Embed Size (px)
DESCRIPTION
Alan Weiner BIOCHEM 530 Wednesday, EE 037 October 8, 2014 RNA structure and function. catalytic (+) strand. template (–) strand. If RNA can be both catalyst and genome, RNA could replicate itself… the first "living" molecule!. precursors. - PowerPoint PPT Presentation
Citation preview

Alan WeinerBIOCHEM 530Wednesday, EE 037October 8, 2014RNA structure and function
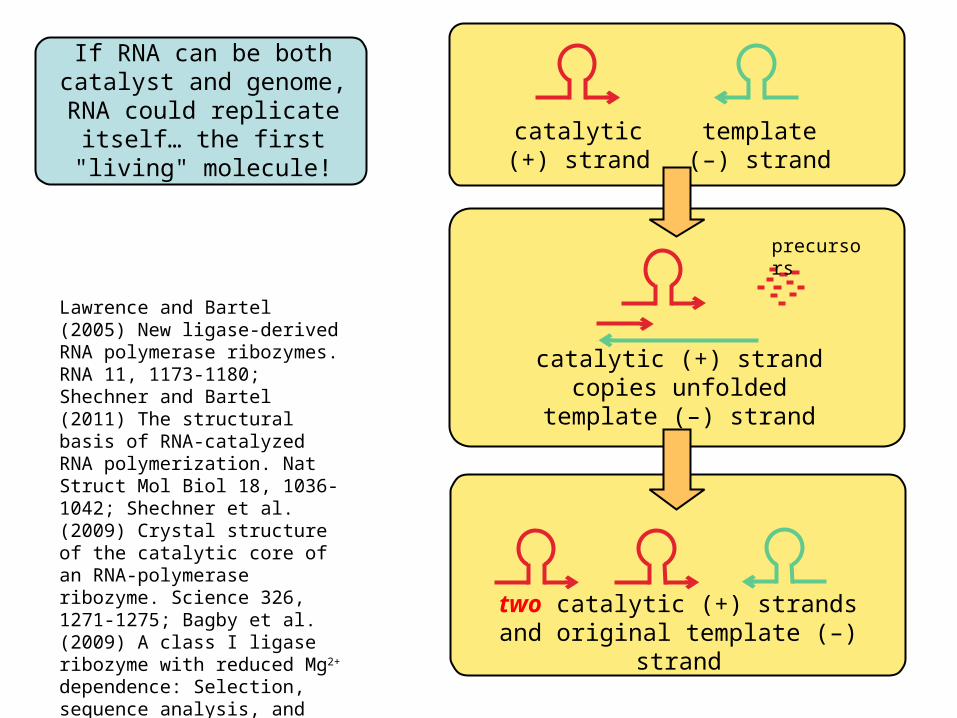
If RNA can be bothcatalyst and genome,
RNA could replicate itself… the first "living" molecule!
catalytic(+) strand
template(–) strand
catalytic (+) strand copies unfolded template (–) strand
two catalytic (+) strandsand original template (–) strand
Lawrence and Bartel (2005) New ligase-derived RNA polymerase ribozymes. RNA 11, 1173-1180; Shechner and Bartel (2011) The structural basis of RNA-catalyzed RNA polymerization. Nat Struct Mol Biol 18, 1036-1042; Shechner et al. (2009) Crystal structure of the catalytic core of an RNA-polymerase ribozyme. Science 326, 1271-1275; Bagby et al. (2009) A class I ligase ribozyme with reduced Mg2+ dependence: Selection, sequence analysis, and identification of functional tertiary interactions. RNA 15, 2129-2146.
precursors
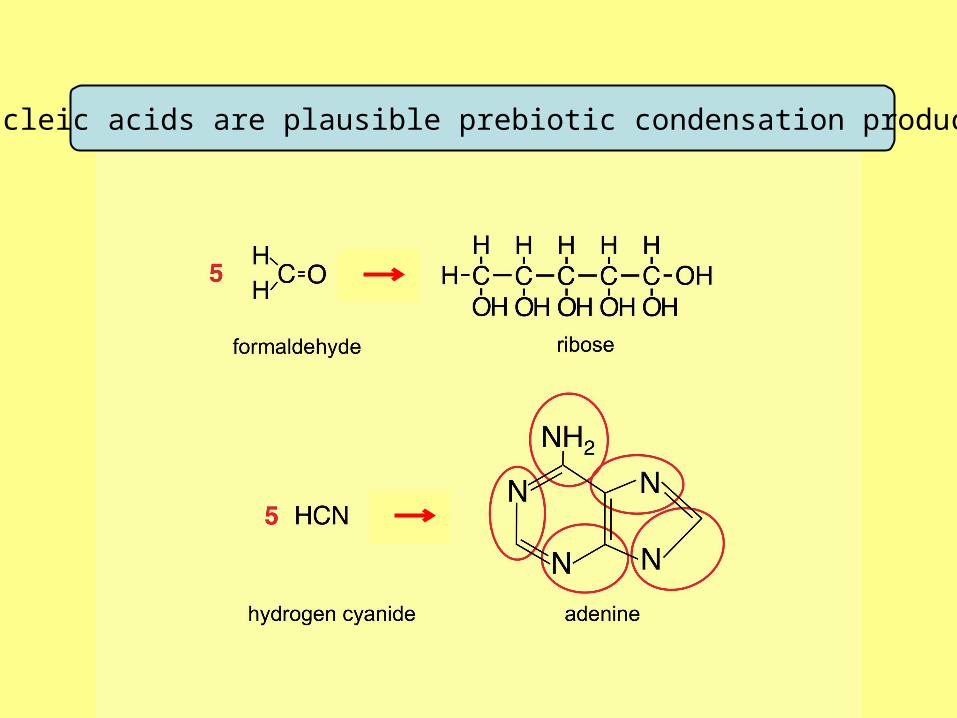
Nucleic acids are plausible prebiotic condensation products
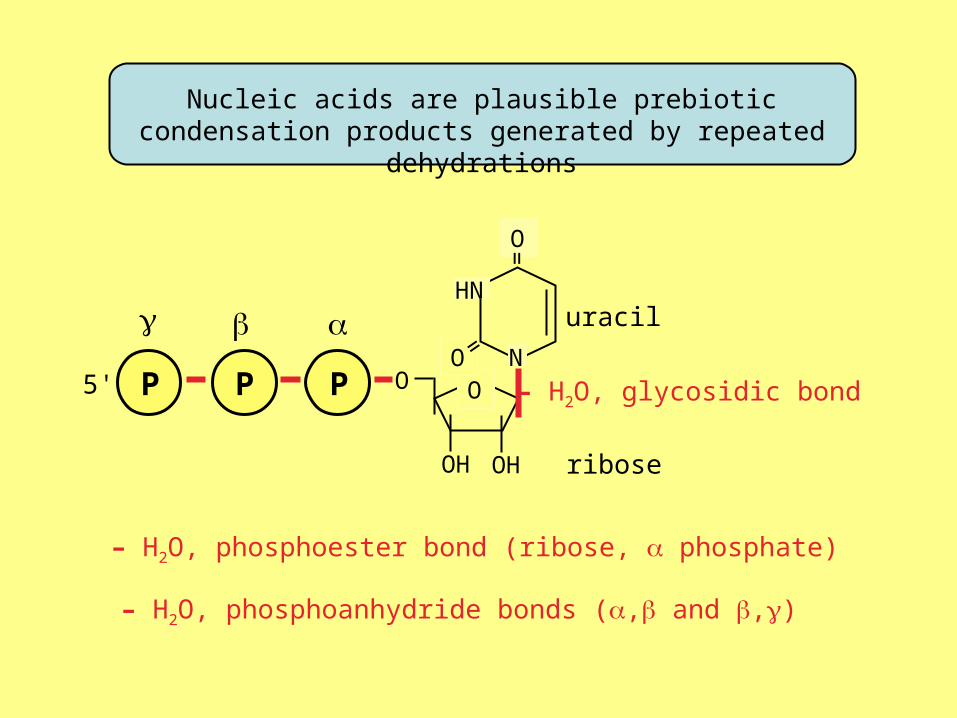
Nucleic acids are plausible prebiotic condensation products generated by repeated dehydrations
uracil
O
OH OH
P O
O
=
O =
5' – H2O, glycosidic bond
– H2O, phosphoester bond (ribose, phosphate)
ribose
PP
– H2O, phosphoanhydride bonds (, and ,)
HN
N
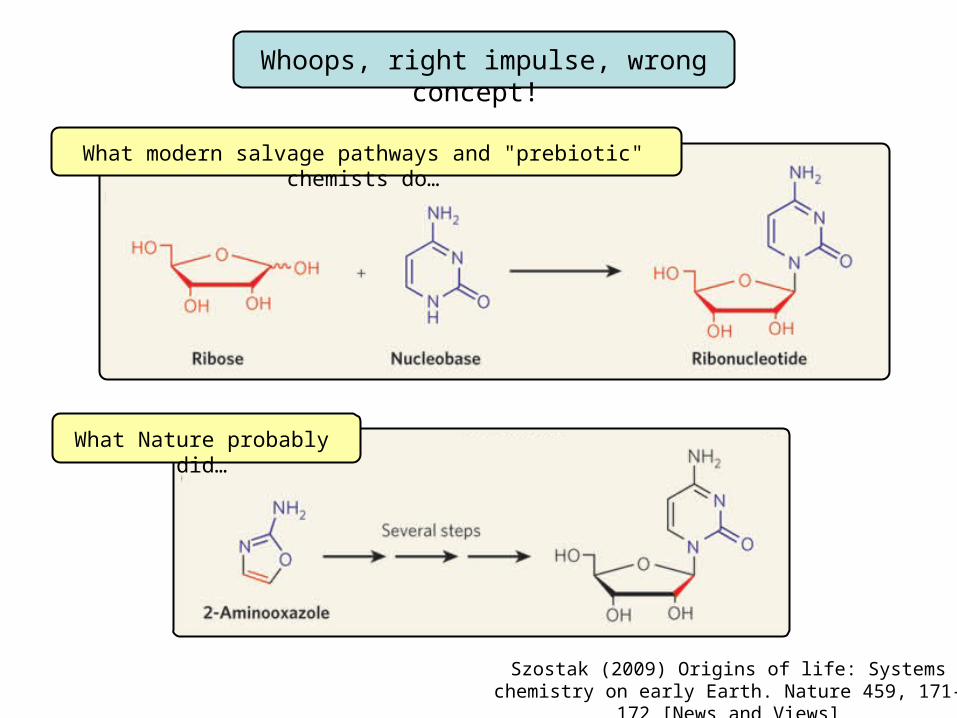
Whoops, right impulse, wrong concept!
Szostak (2009) Origins of life: Systems chemistry on early Earth. Nature 459, 171-172 [News and Views]
What Nature probably did…
What modern salvage pathways and "prebiotic" chemists do…
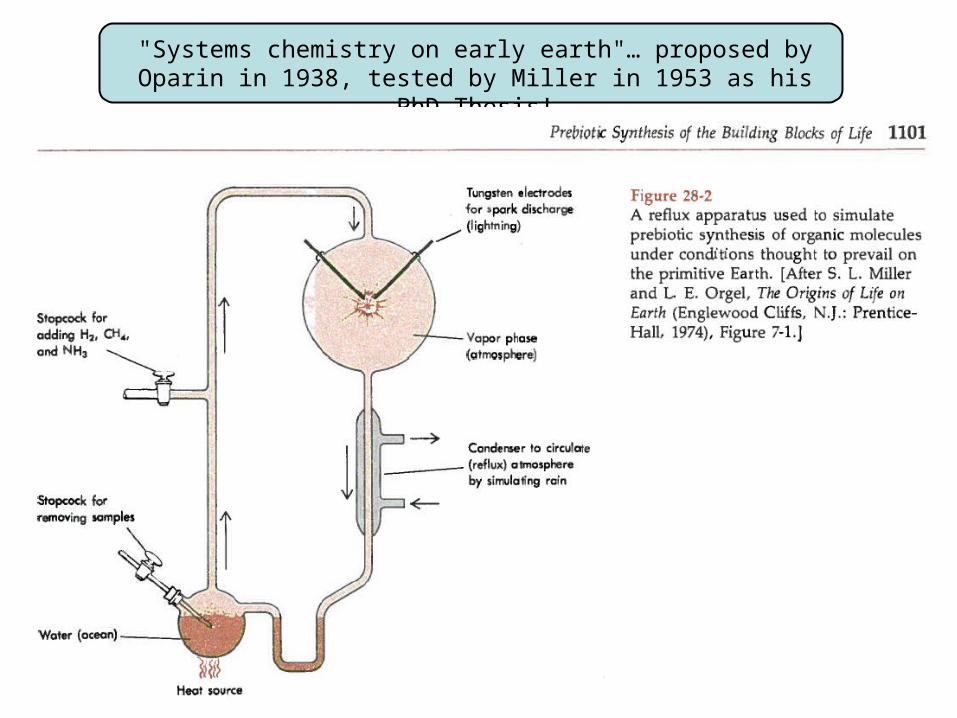
"Systems chemistry on early earth"… proposed by Oparin in 1938, tested by Miller in 1953 as his PhD Thesis!
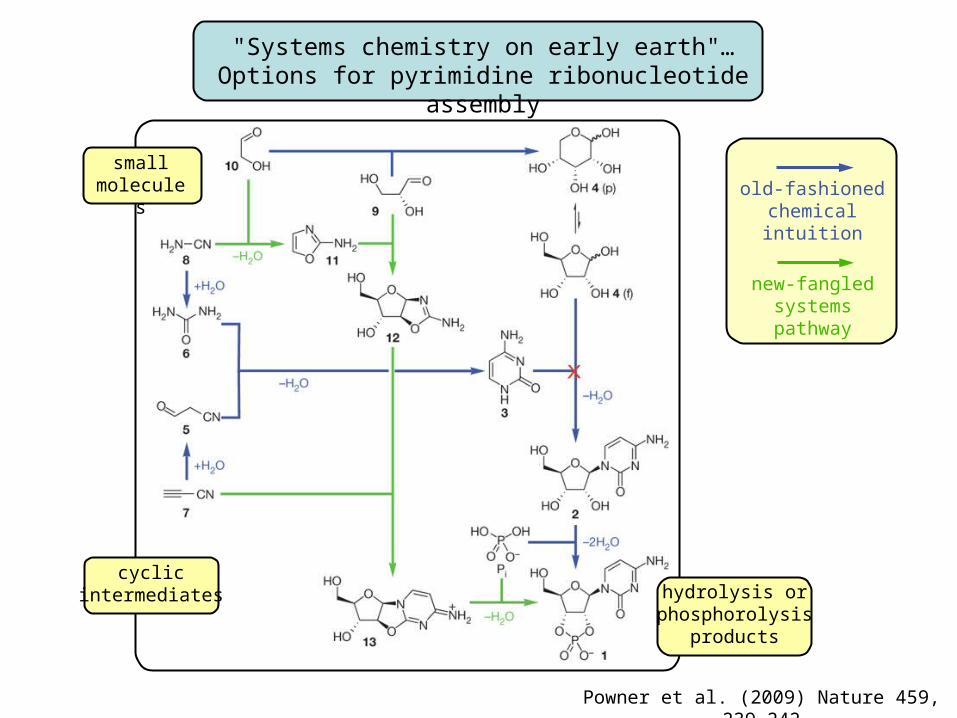
"Systems chemistry on early earth"…Options for pyrimidine ribonucleotide assembly
Powner et al. (2009) Nature 459, 239-242
smallmolecules
cyclic intermediates hydrolysis or
phosphorolysisproducts
old-fashionedchemical intuition
new-fangledsystems pathway
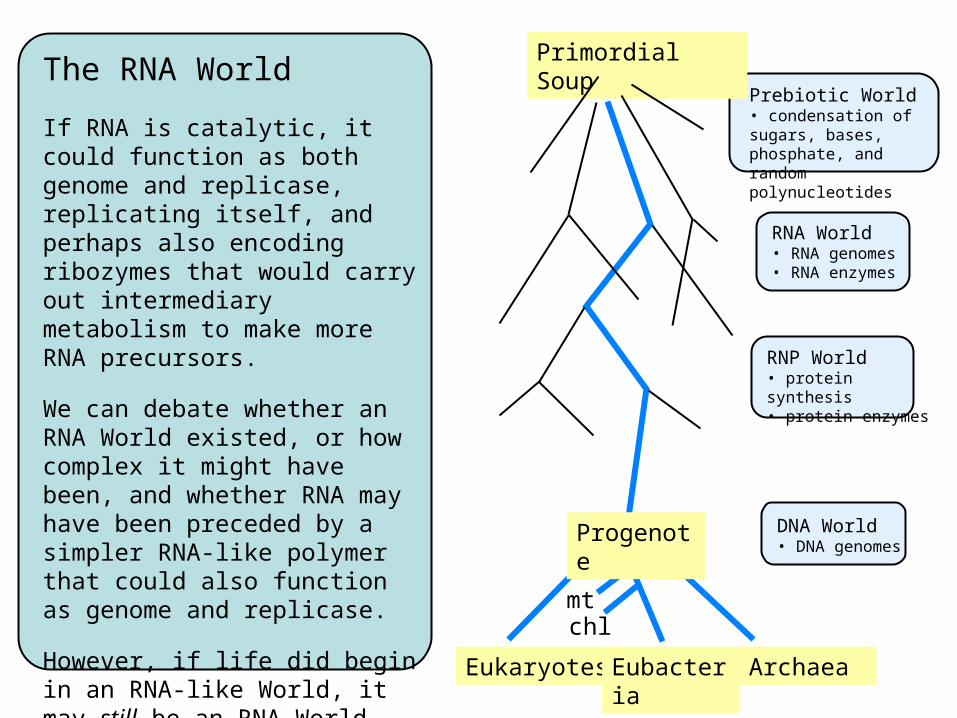
The RNA World
If RNA is catalytic, it could function as both genome and replicase, replicating itself, and perhaps also encoding ribozymes that would carry out intermediary metabolism to make more RNA precursors.
We can debate whether an RNA World existed, or how complex it might have been, and whether RNA may have been preceded by a simpler RNA-like polymer that could also function as genome and replicase.
However, if life did begin in an RNA-like World, it may still be an RNA World today only slightly disguised by a veil of DNA!
Prebiotic World• condensation of sugars, bases, phosphate, and random polynucleotides
Primordial Soup
Progenote
Eukaryotes Eubacteria Archaea
mtchl
DNA World• DNA genomes
RNA World• RNA genomes• RNA enzymes
RNP World• protein synthesis• protein enzymes
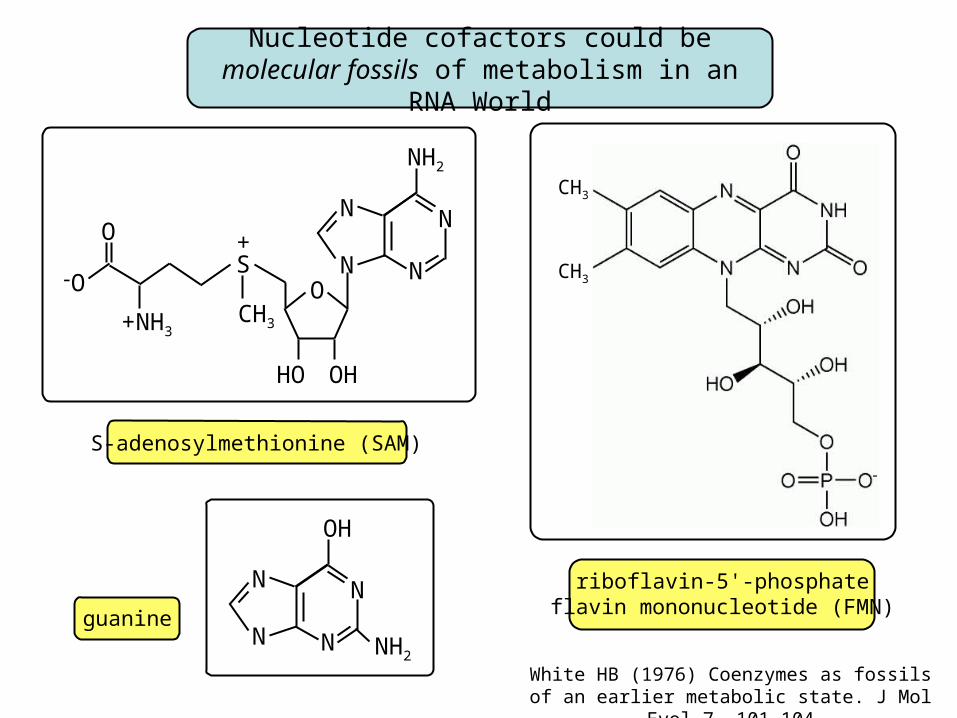
S-adenosylmethionine (SAM)
riboflavin-5'-phosphateflavin mononucleotide (FMN)
Nucleotide cofactors could be molecular fossils of metabolism in an RNA World
guanine
CH3
CH3
NH2
OH
N
NN
N
CH3
NH2
N
NN
NSO
HO OH
+NH3
O
–O
+
White HB (1976) Coenzymes as fossils of an earlier metabolic state. J Mol Evol 7, 101-104
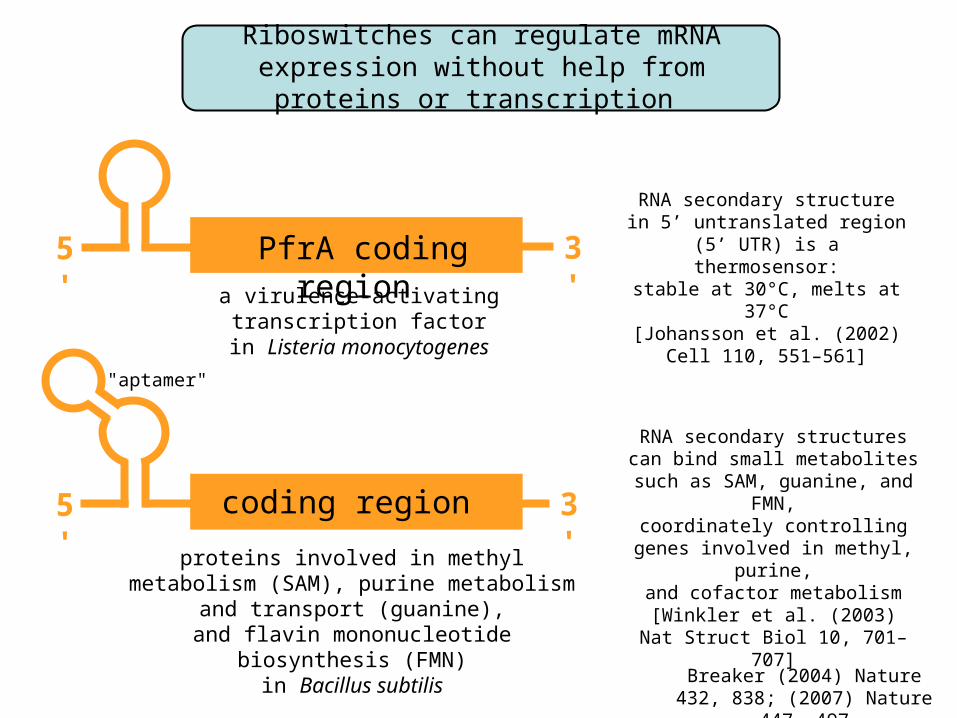
Riboswitches can regulate mRNA expression without help from proteins or transcription
a virulence-activating transcription factor
in Listeria monocytogenes
PfrA coding region
RNA secondary structurein 5’ untranslated region
(5’ UTR) is a thermosensor:stable at 30°C, melts at 37°C
[Johansson et al. (2002)Cell 110, 551–561]
proteins involved in methyl metabolism (SAM), purine metabolism and transport (guanine),
and flavin mononucleotide biosynthesis (FMN)in Bacillus subtilis
coding region
RNA secondary structurescan bind small metabolites
such as SAM, guanine, and FMN,coordinately controlling genes
involved in methyl, purine,and cofactor metabolism
[Winkler et al. (2003)Nat Struct Biol 10, 701–707]
5' 3'
5' 3'
Breaker (2004) Nature 432, 838; (2007) Nature 447, 497
"aptamer"
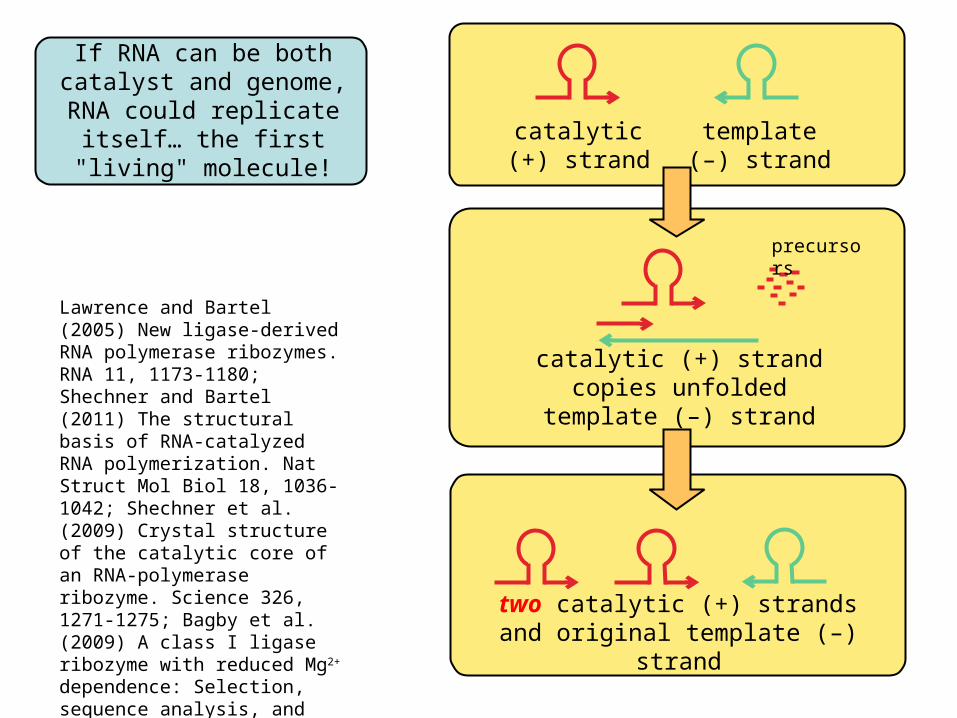
If RNA can be bothcatalyst and genome,
RNA could replicate itself… the first "living" molecule!
catalytic(+) strand
template(–) strand
catalytic (+) strand copies unfolded template (–) strand
two catalytic (+) strandsand original template (–) strand
Lawrence and Bartel (2005) New ligase-derived RNA polymerase ribozymes. RNA 11, 1173-1180; Shechner and Bartel (2011) The structural basis of RNA-catalyzed RNA polymerization. Nat Struct Mol Biol 18, 1036-1042; Shechner et al. (2009) Crystal structure of the catalytic core of an RNA-polymerase ribozyme. Science 326, 1271-1275; Bagby et al. (2009) A class I ligase ribozyme with reduced Mg2+ dependence: Selection, sequence analysis, and identification of functional tertiary interactions. RNA 15, 2129-2146.
precursors
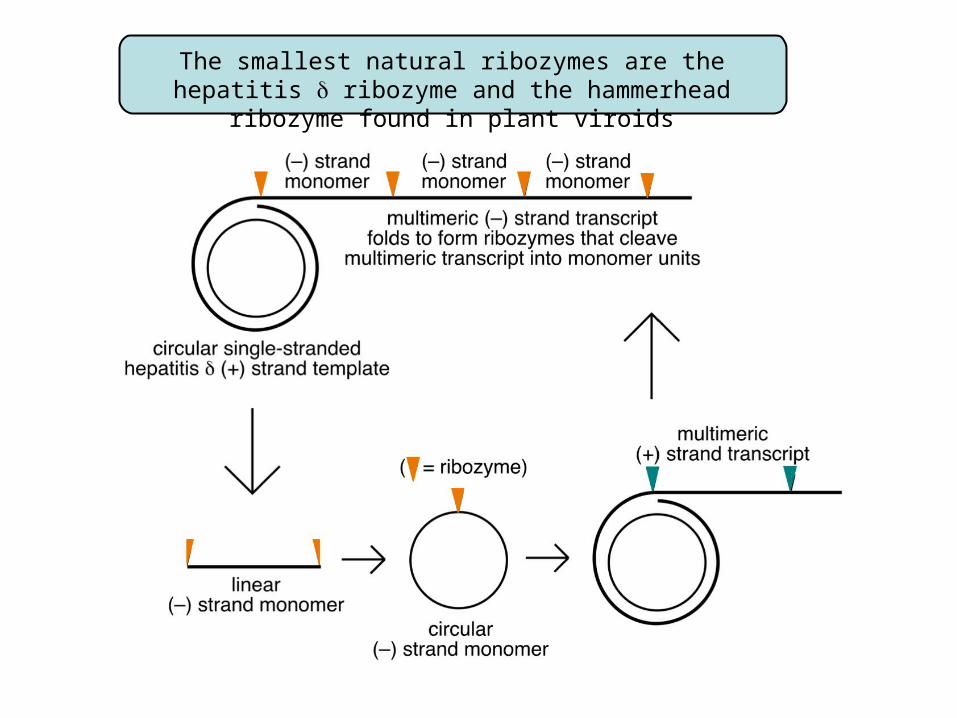
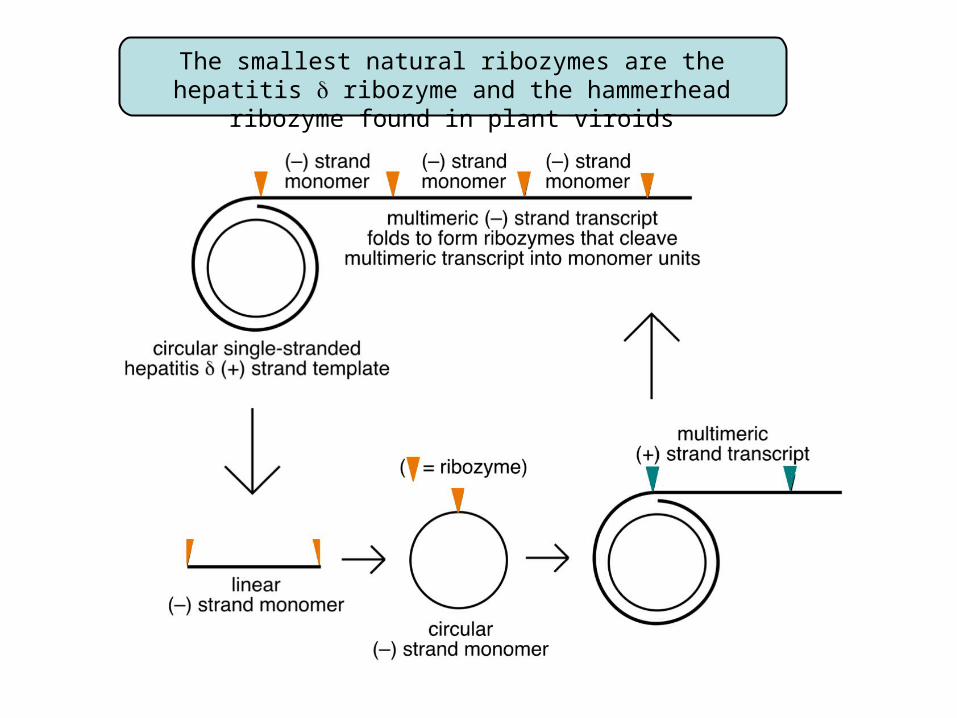
The smallest natural ribozymes are the hepatitis ribozyme and the hammerhead ribozyme found in plant viroids

Australia battled citrus exocortis virus (oranges, lemons, and etrogs on susceptible rootstocks) in the 1940s and
1950s
healthy treestunted by exocortis
http://www.dpi.nsw.gov.au/agriculture/horticulture/citrus/health/diseases/citrus-exocortis
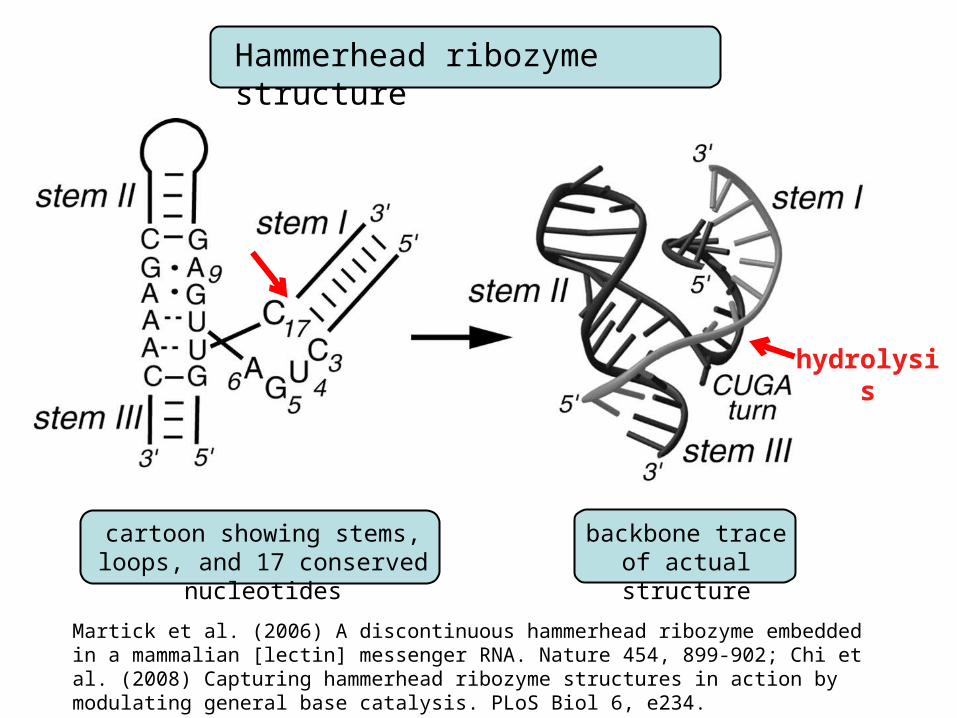
Hammerhead ribozyme structure
cartoon showing stems, loops, and 17 conserved nucleotides
backbone traceof actual structure
hydrolysis
Martick et al. (2006) A discontinuous hammerhead ribozyme embedded in a mammalian [lectin] messenger RNA. Nature 454, 899-902; Chi et al. (2008) Capturing hammerhead ribozyme structures in action by modulating general base catalysis. PLoS Biol 6, e234.
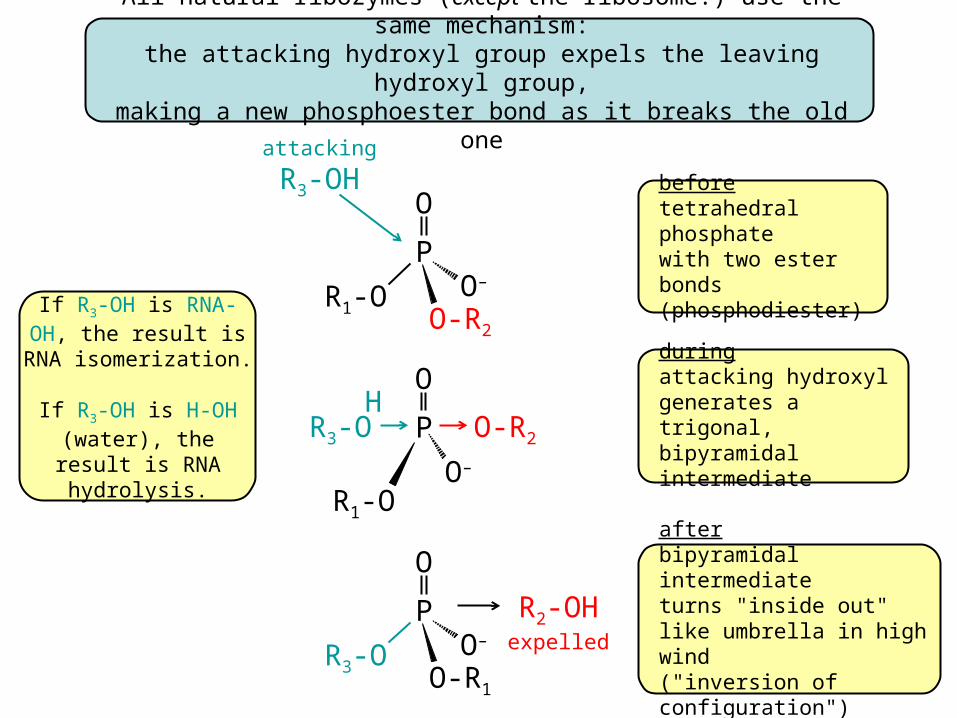
All natural ribozymes (except the ribosome!) use the same mechanism:the attacking hydroxyl group expels the leaving hydroxyl group,
making a new phosphoester bond as it breaks the old one
If R3-OH is RNA-OH, the result is
RNA isomerization.
If R3-OH is H-OH (water), the result is
RNA hydrolysis.
beforetetrahedral phosphatewith two ester bonds(phosphodiester)
duringattacking hydroxylgenerates a trigonal,bipyramidal intermediate
afterbipyramidal intermediateturns "inside out"like umbrella in high wind("inversion of configuration")
P
O-R2
O–
O
R1-O
attacking
R3-OH
P
O-R1
O–
O
R3-O
R2-OHexpelled
P
O–
O
R3-O O-R2
R1-O
H
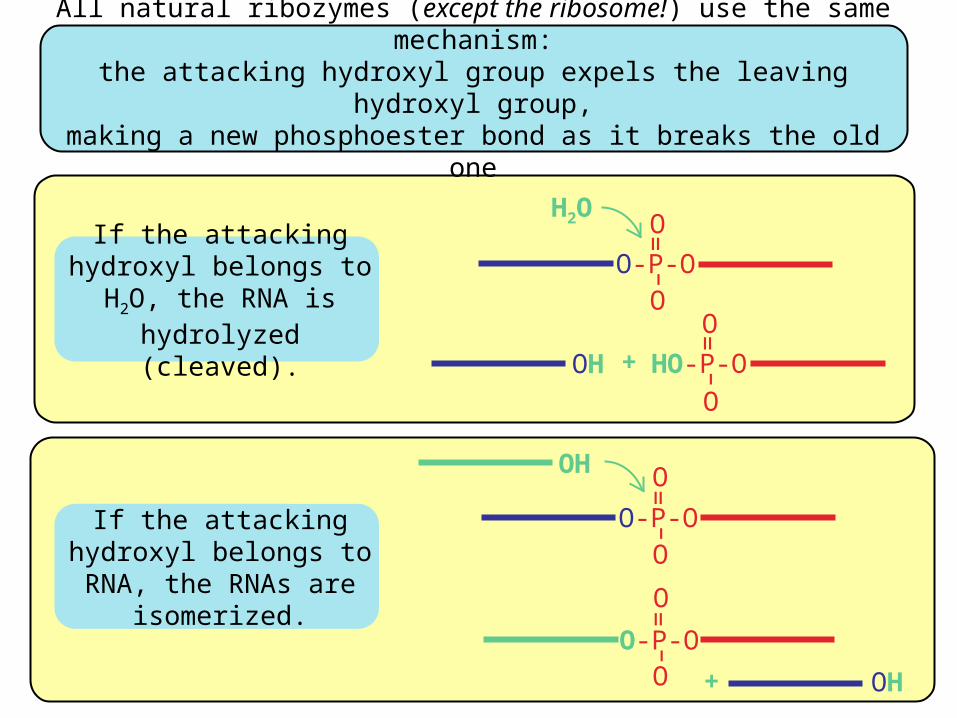
All natural ribozymes (except the ribosome!) use the same mechanism:the attacking hydroxyl group expels the leaving hydroxyl group,
making a new phosphoester bond as it breaks the old one
If the attacking hydroxyl belongs to H2O, the RNA is hydrolyzed (cleaved).
If the attacking hydroxyl belongs to RNA, the
RNAs are isomerized.
H2O
O-P-O
O
O
=–
HO-P-O
O
OH
=–
O
+
OH
O-P-O
O
O
=–
OH+
O-P-O
O
=–
O
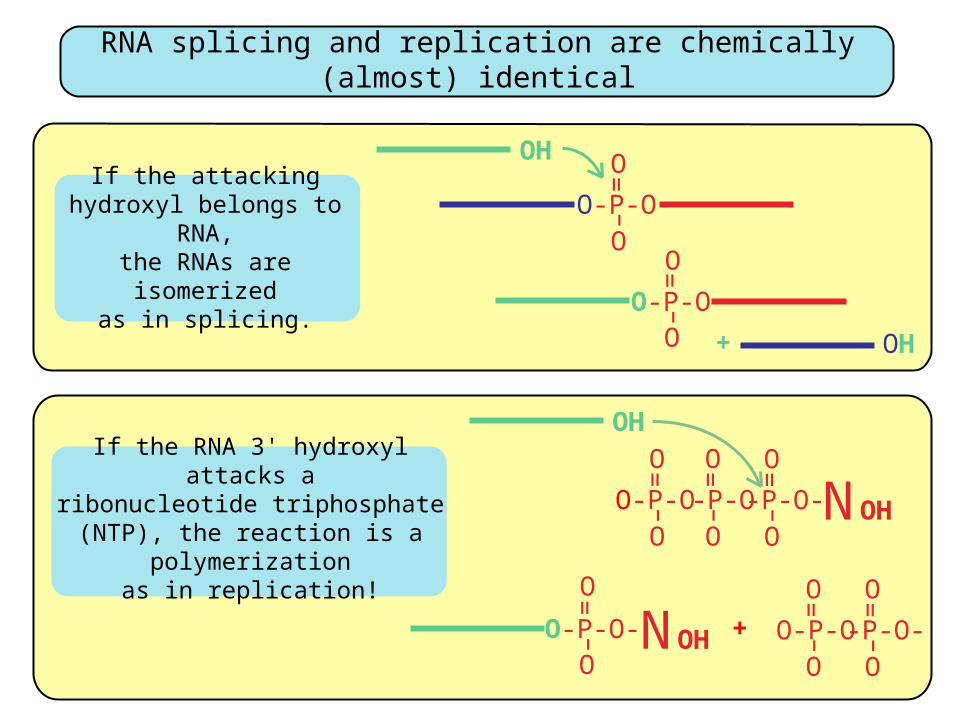
If the attacking hydroxyl belongs to RNA,
the RNAs are isomerizedas in splicing.
OH
O-P-O
O
O
=–
OH+
O-P-O
O
=–
O
If the RNA 3' hydroxyl attacks aribonucleotide triphosphate (NTP),
the reaction is a polymerizationas in replication!
OH
O-P-O
O
O
=–
+O-P-O-
O=
–
O
-P-O
O
O
=– -P-O-
O
O
=– N
N -P-O
O
O
=– -P-O-
O
O
=–O
RNA splicing and replication are chemically (almost) identical
OH
OH
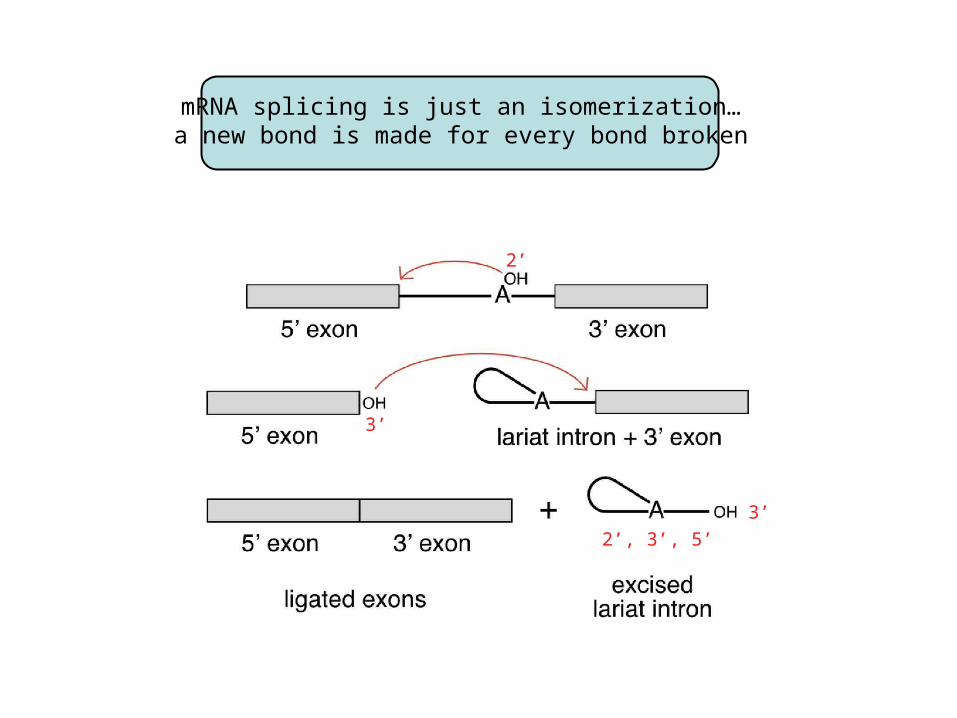
mRNA splicing is just an isomerization…a new bond is made for every bond broken
2’
3’
3’
2’, 3’, 5’
The smallest natural ribozymes are the hepatitis ribozyme and the hammerhead ribozyme found in plant viroids
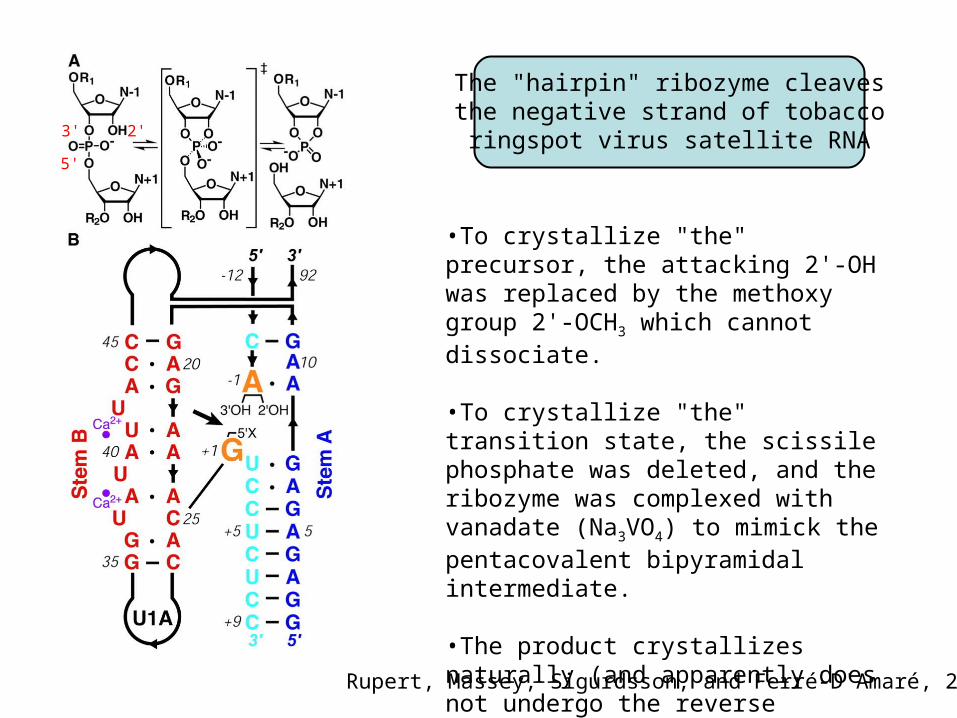
The "hairpin" ribozyme cleavesthe negative strand of tobacco
ringspot virus satellite RNA
•To crystallize "the" precursor, the attacking 2'-OH was replaced by the methoxy group 2'-OCH3 which cannot dissociate.
•To crystallize "the" transition state, the scissile phosphate was deleted, and the ribozyme was complexed with vanadate (Na3VO4) to mimick the pentacovalent bipyramidal intermediate.
•The product crystallizes naturally (and apparently does not undergo the reverse reaction!).
Rupert, Massey, Sigurdsson, and Ferré-D'Amaré, 2002
2'3'
5'
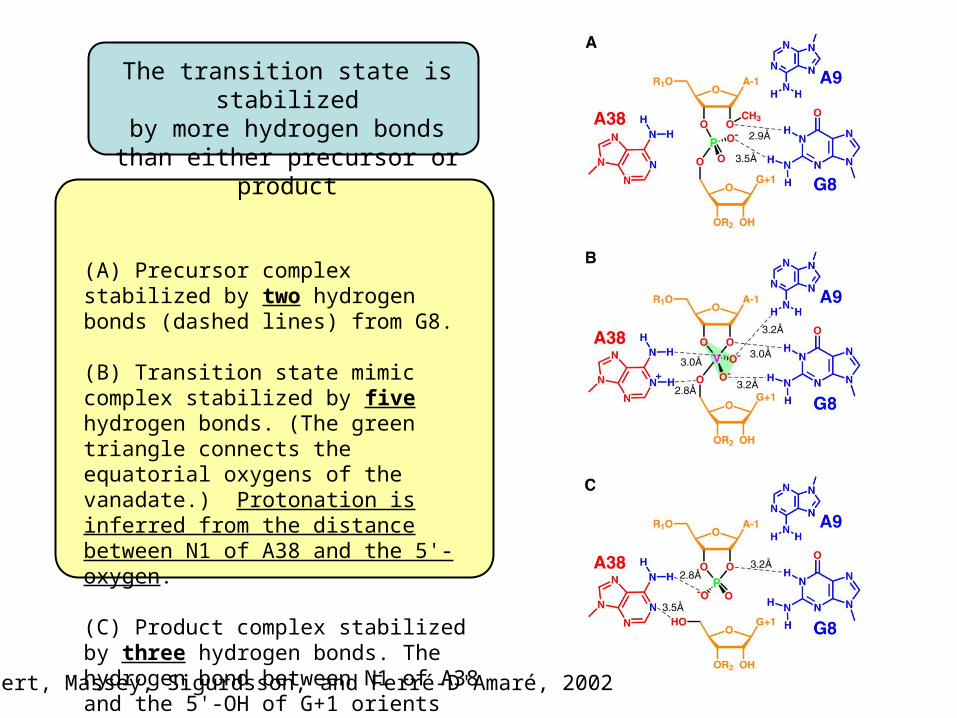
The transition state is stabilizedby more hydrogen bonds
than either precursor or product
(A) Precursor complex stabilized by two hydrogen bonds (dashed lines) from G8.
(B) Transition state mimic complex stabilized by five hydrogen bonds. (The green triangle connects the equatorial oxygens of the vanadate.) Protonation is inferred from the distance between N1 of A38 and the 5'-oxygen.
(C) Product complex stabilized by three hydrogen bonds. The hydrogen bond between N1 of A38 and the 5'-OH of G+1 orients the latter for subsequent ligation.
Rupert, Massey, Sigurdsson, and Ferré-D'Amaré, 2002
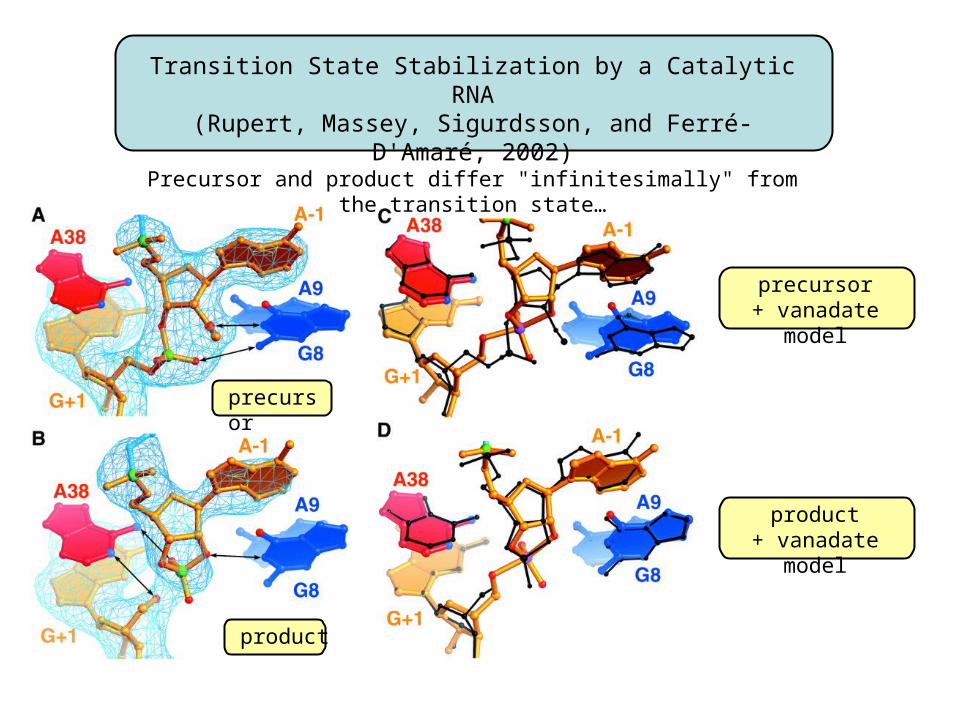
precursor
product
precursor+ vanadate model
product+ vanadate model
Transition State Stabilization by a Catalytic RNA(Rupert, Massey, Sigurdsson, and Ferré-D'Amaré, 2002)
Precursor and product differ "infinitesimally" from the transition state…
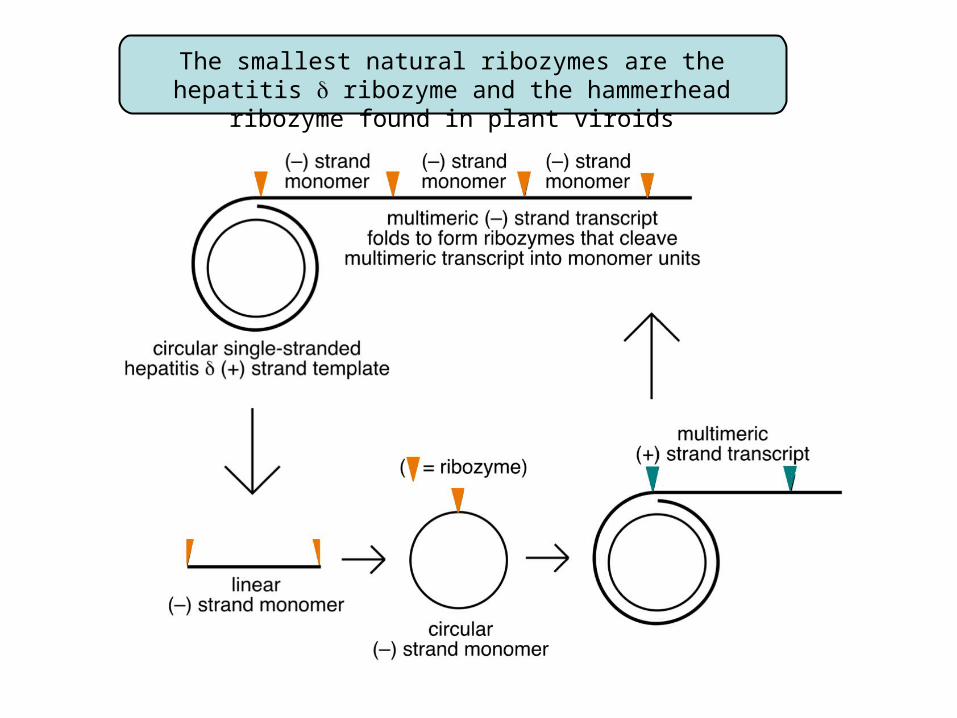
The smallest natural ribozymes are the hepatitis ribozyme and the hammerhead ribozyme found in plant viroids

Doudna and Lorsch (2005)Nat Struct Mol Biol 12, 395-402.

Doudna and Lorsch (2005)Nat Struct Mol Biol 12, 395-402.
Ribozyme-catalyzed reactions: very different,and often far better*
* think of ribosomes and spliceosomes which require large motions of large complexes with many RNA molecules
coming and going!
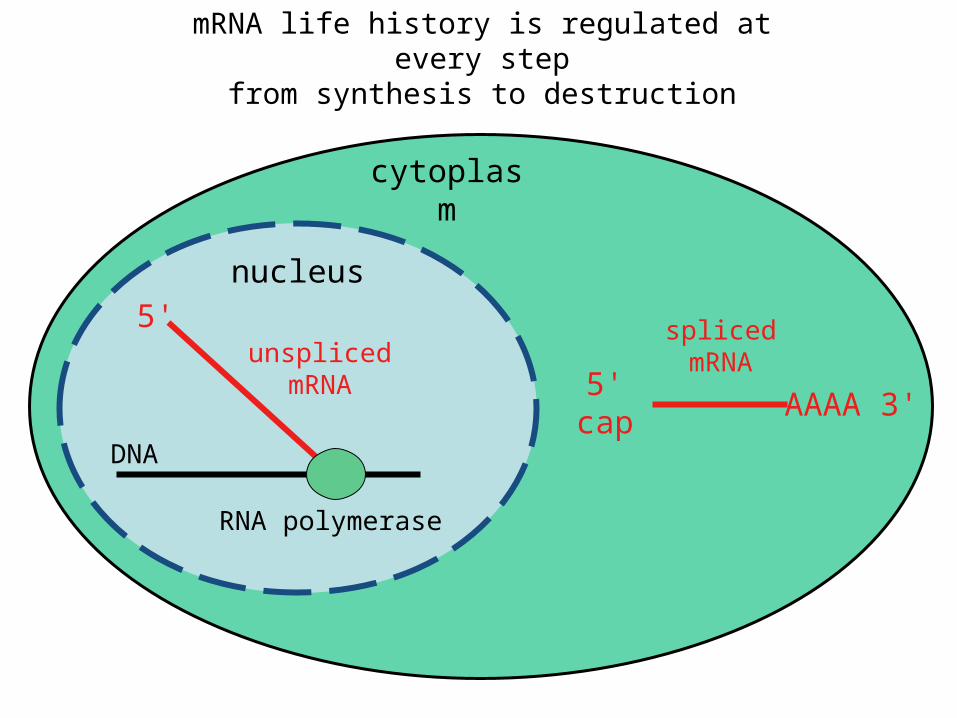
mRNA life history is regulated at every stepfrom synthesis to destruction
5'
RNA polymerase
DNA
unsplicedmRNA
nucleus
cytoplasm
spliced mRNA
5' cap AAAA 3'
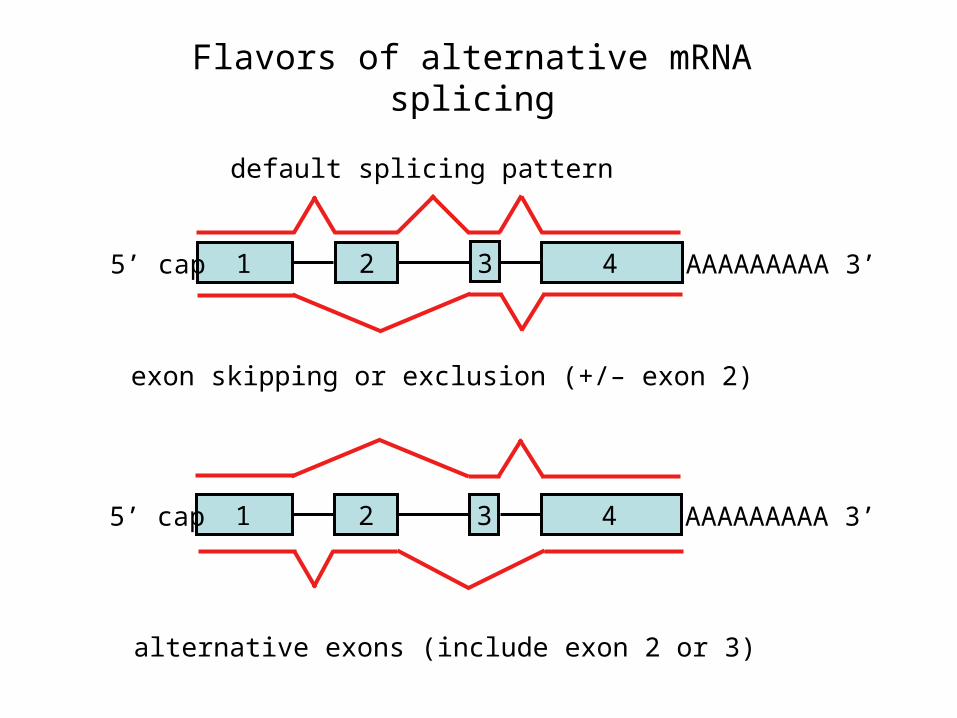
Flavors of alternative mRNA splicing
1 2 3 45’ cap AAAAAAAAA 3’
default splicing pattern
exon skipping or exclusion (+/– exon 2)
1 2 3 45’ cap AAAAAAAAA 3’
alternative exons (include exon 2 or 3)
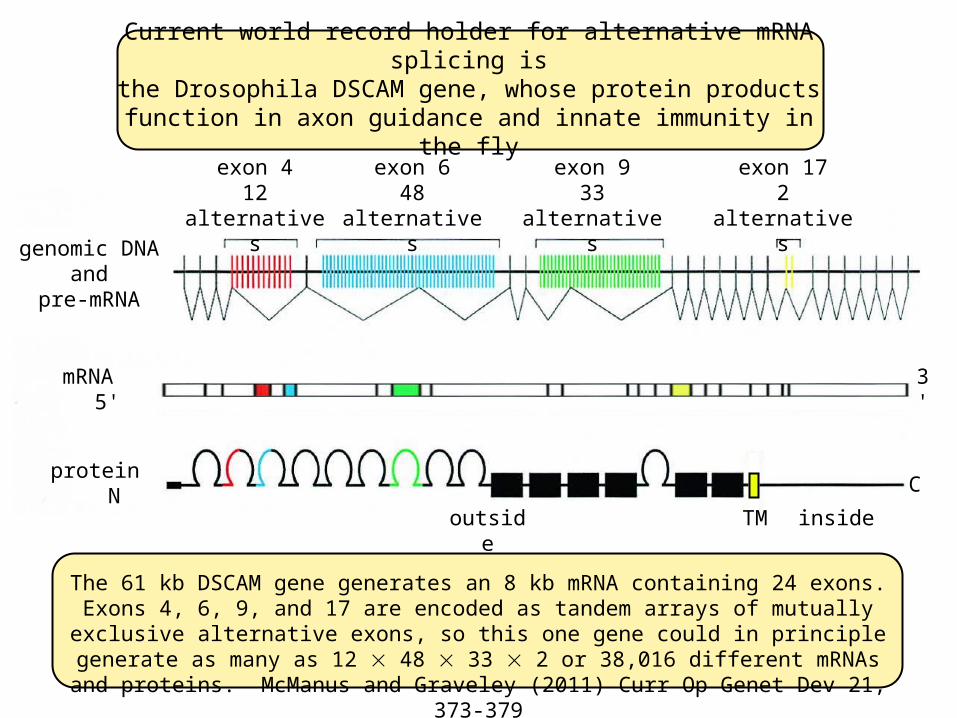
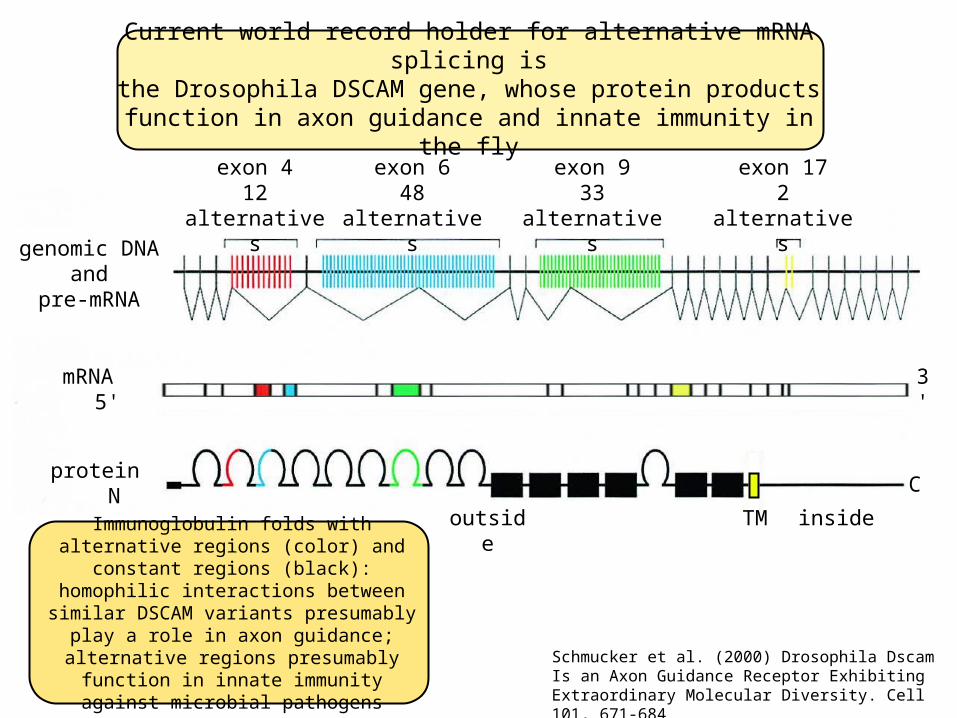
Current world record holder for alternative mRNA splicing isthe Drosophila DSCAM gene, whose protein products
function in axon guidance and innate immunity in the fly
The 61 kb DSCAM gene generates an 8 kb mRNA containing 24 exons. Exons 4, 6, 9, and 17 are encoded as tandem arrays of mutually exclusive alternative exons, so this one gene could in principle generate as many as 12 48 33 2 or 38,016 different
mRNAs and proteins. McManus and Graveley (2011) Curr Op Genet Dev 21, 373-379
exon 412 alternatives
exon 648 alternatives
exon 933 alternatives
exon 172 alternatives
protein N
genomic DNA and
pre-mRNA
TMoutside inside
3'
C
mRNA 5'

New answers pose new questions. We assumed that transcriptional control would be the major determinant of complexity, and we asked What wires transcription? Now it turns out that nerves are wired by alternative mRNA splicing, and so we must now ask What wires alternative splicing? Maybe not an endless hall of mirrors, but certainly no end in sight...
Hattori et al. (2007) Dscam diversity is essential for neuronal wiring and self-recognition. Nature 449, 223-228; Meijers et al. (2007) Structural basis of Dscam isoform specificity. Nature 449, 487-491; Du Pasquier (2005) Insects diversify one molecule to serve two systems. Science 309, 1826-1827..
insideoutside
Current world record holder for alternative mRNA splicing isthe Drosophila DSCAM gene, whose protein products
function in axon guidance and innate immunity in the fly
exon 412 alternatives
exon 648 alternatives
exon 933 alternatives
exon 172 alternatives
protein N
genomic DNA and
pre-mRNA
TMoutside inside
3'
C
mRNA 5'
Immunoglobulin folds with alternative regions (color) and constant regions (black):
homophilic interactions between similar DSCAM variants presumably play a role in
axon guidance; alternative regions presumably function in innate immunity
against microbial pathogensSchmucker et al. (2000) Drosophila Dscam Is an Axon Guidance Receptor Exhibiting Extraordinary Molecular Diversity. Cell 101, 671-684
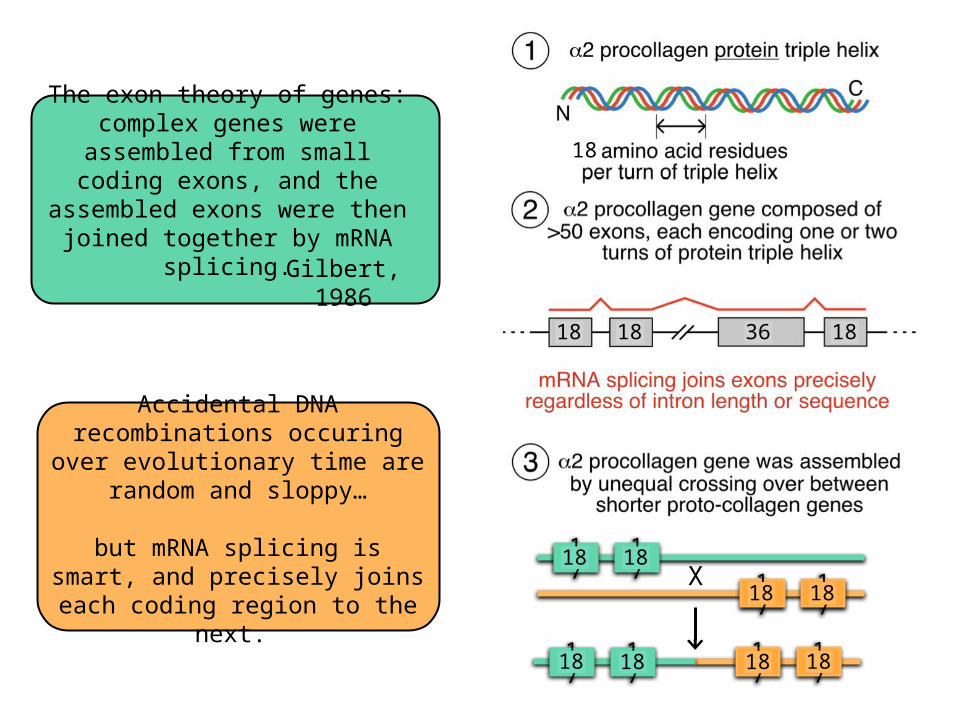
The exon theory of genes: complex genes were assembled from small coding exons, and the
assembled exons were then joined together by mRNA splicing.
Accidental DNA recombinations occuring over evolutionary time are
random and sloppy…
but mRNA splicing is smart, and precisely joins each coding region
to the next.
Gilbert, 1986
17 17
17 17
17 17
17 17
X
18
18 18 1836
18
18 18
18 18
18
18 18
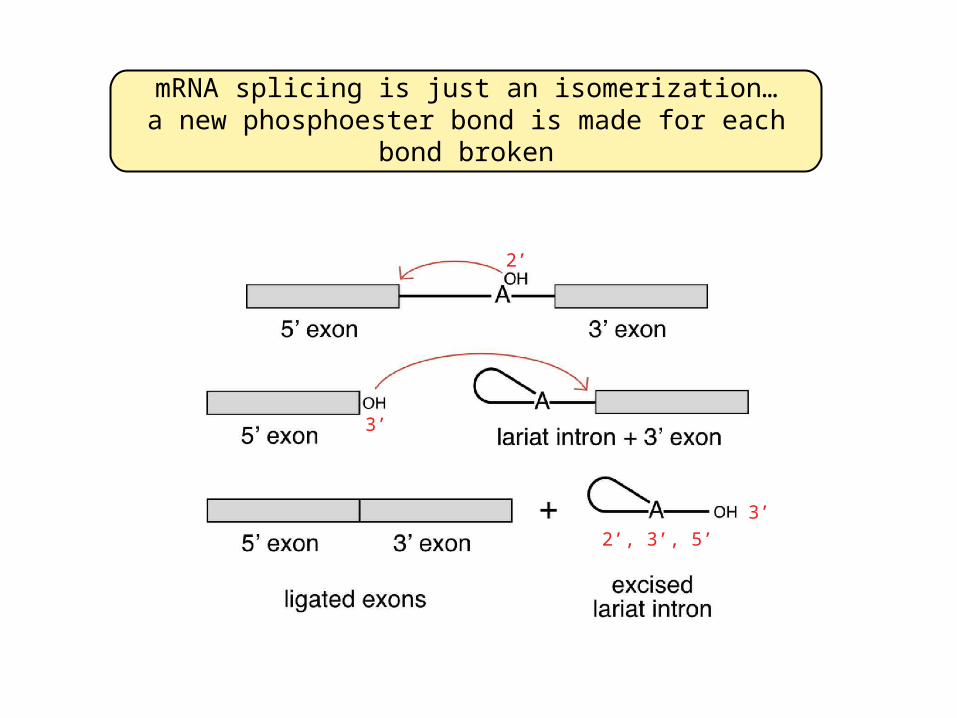
mRNA splicing is just an isomerization…a new phosphoester bond is made for each bond broken
2’
3’
3’
2’, 3’, 5’
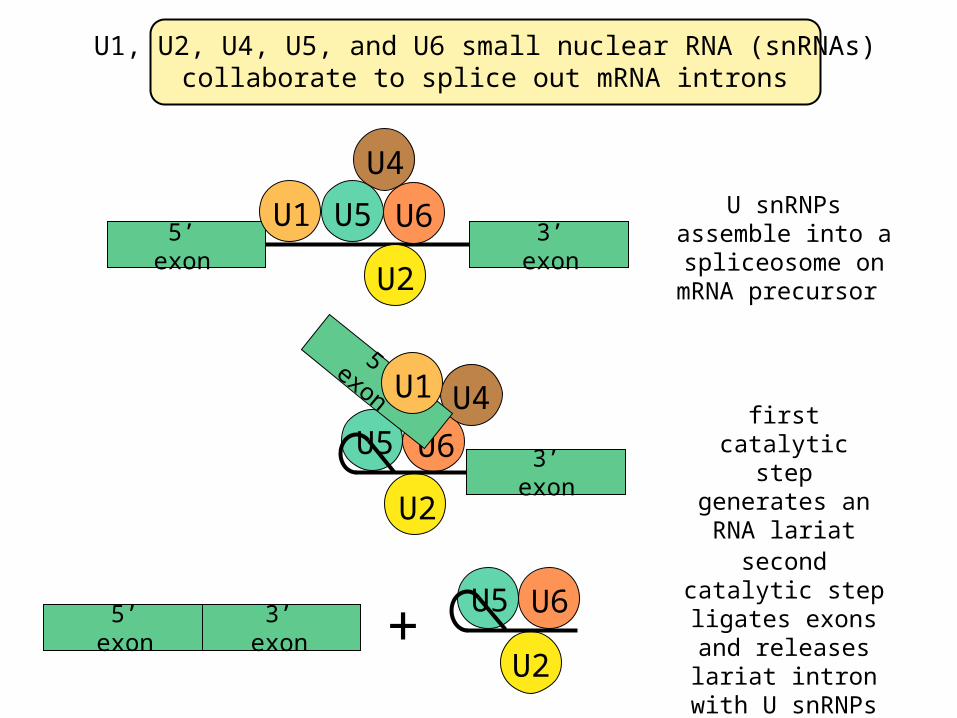
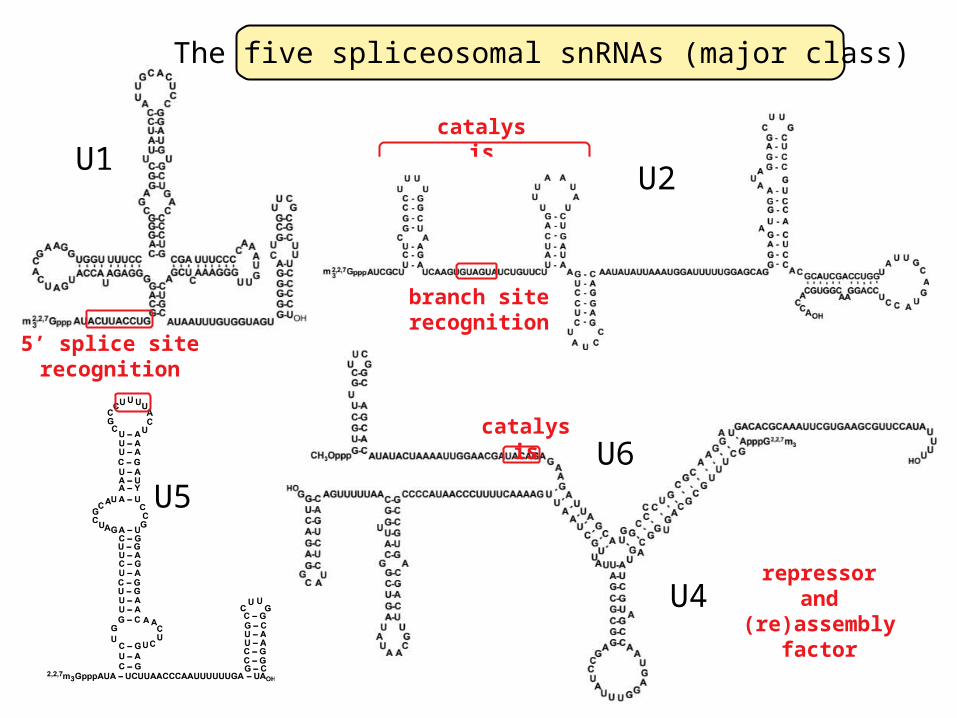
U1, U2, U4, U5, and U6 small nuclear RNA (snRNAs)collaborate to splice out mRNA introns
second catalytic step ligates exons and releases lariat
intron with U snRNPs
U1
U2
U5 U6
U4
U2
U5 U6
U4
U snRNPs assemble into a spliceosome on
mRNA precursor
first catalytic step generates an RNA lariat
+U2
U5 U6
5’ exon 3’ exon
5’ exon
5’ exon
3’ exon
3’ exon
U1
U5
U1
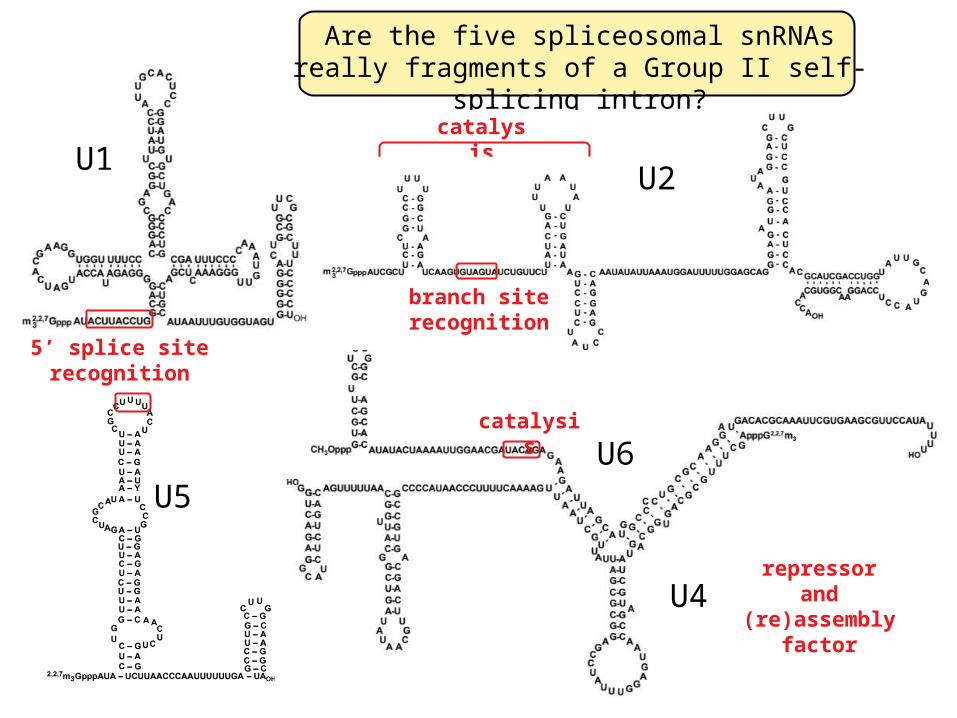
The five spliceosomal snRNAs (major class)
U4
U6catalysi
s
repressorand
(re)assembly factor
branch siterecognition
U2
catalysis
5’ splice siterecognition
U5
U1
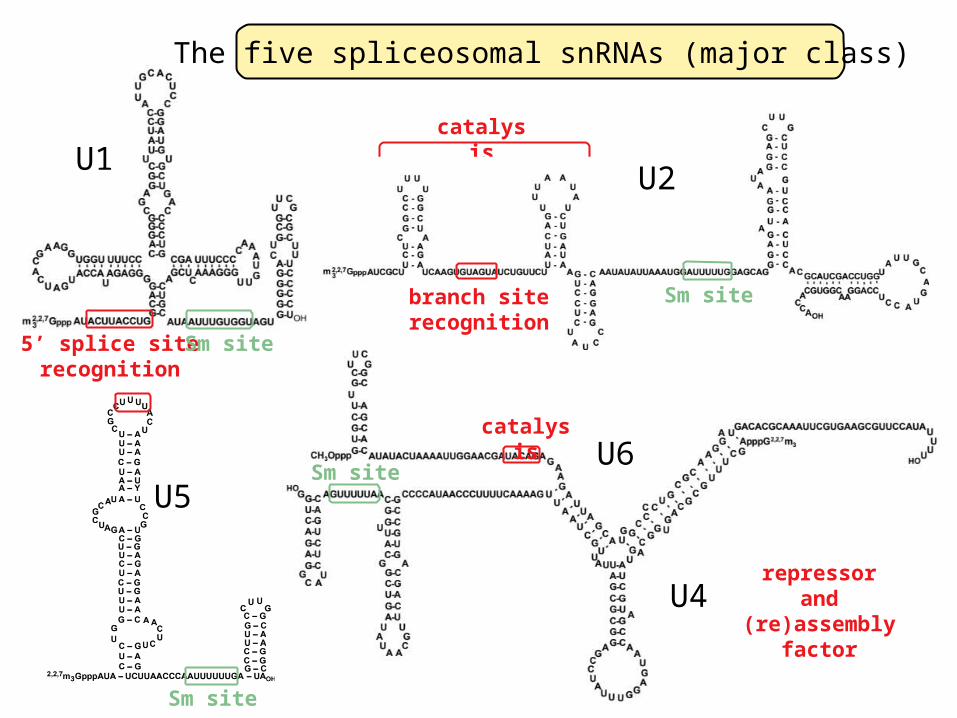
The five spliceosomal snRNAs (major class)
U4
U6
5’ splice siterecognition
catalysis
repressorand
(re)assembly factor
branch siterecognition
U2
catalysis
Sm site
Sm site
Sm site
Sm site
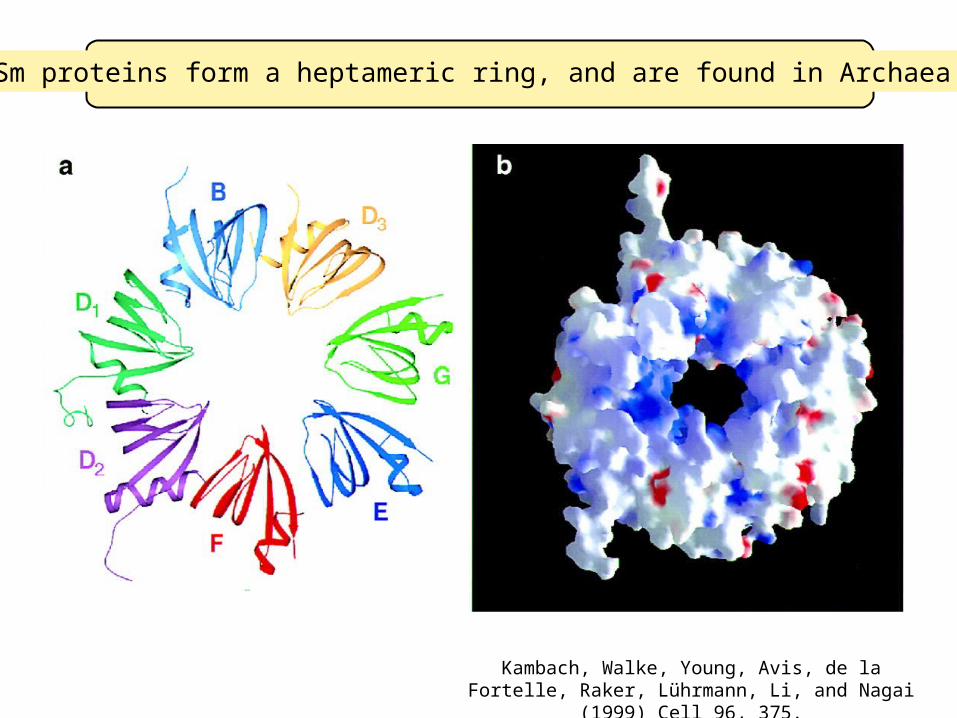
Sm proteins form a heptameric ring, and are found in Archaea!
Kambach, Walke, Young, Avis, de la Fortelle, Raker, Lührmann, Li, and Nagai (1999) Cell 96, 375.
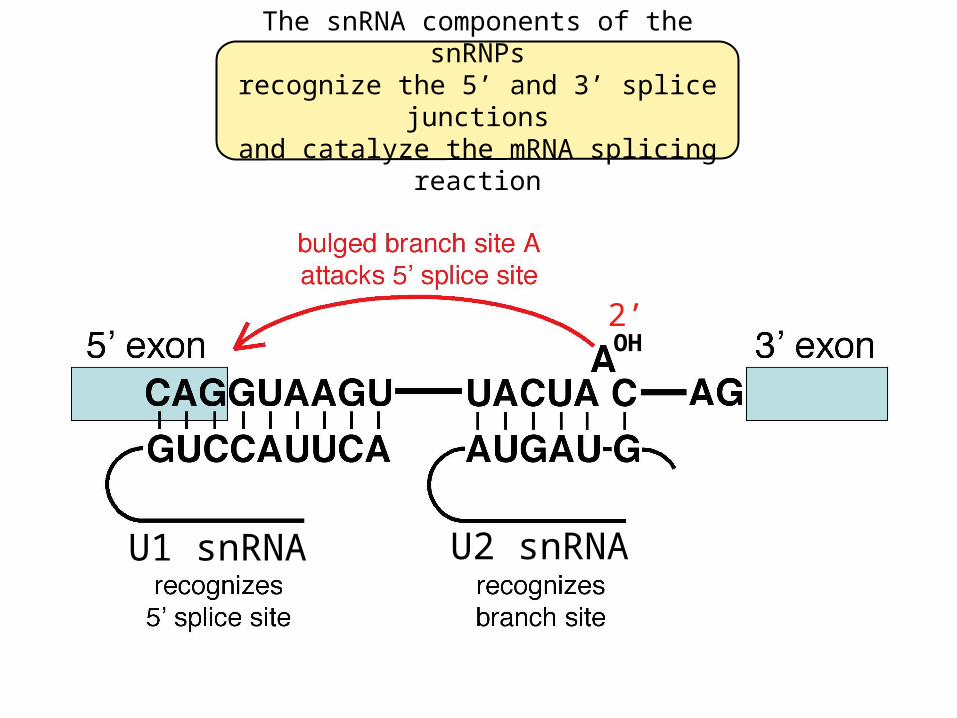
The snRNA components of the snRNPsrecognize the 5’ and 3’ splice junctions
and catalyze the mRNA splicing reaction
OH2’
U1 snRNA U2 snRNA
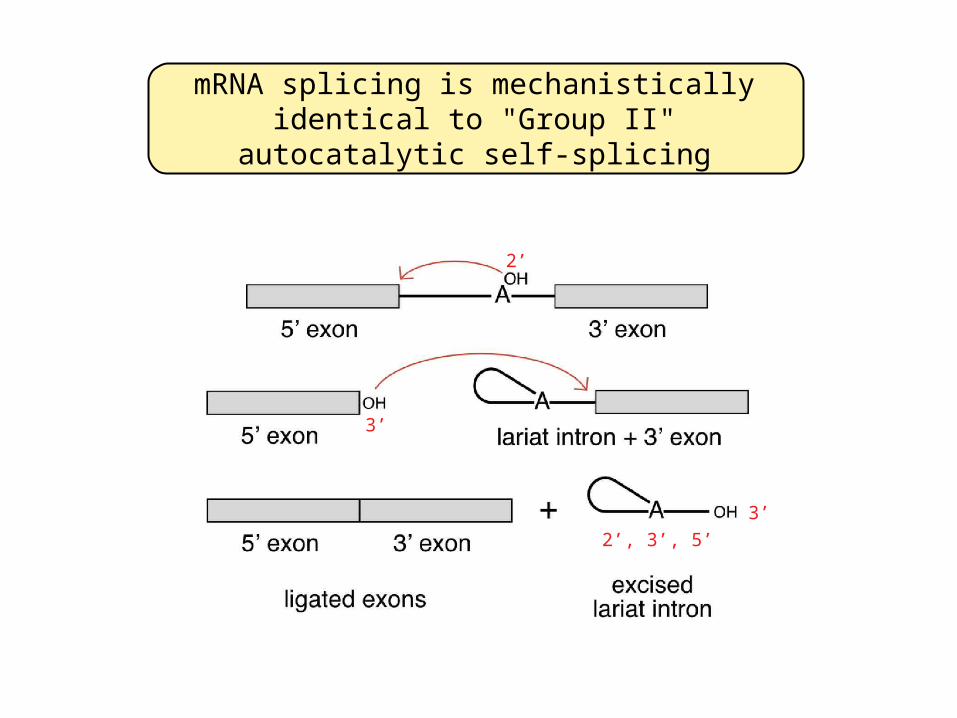
mRNA splicing is mechanistically identical to "Group II" autocatalytic self-splicing
2’
3’
3’
2’, 3’, 5’
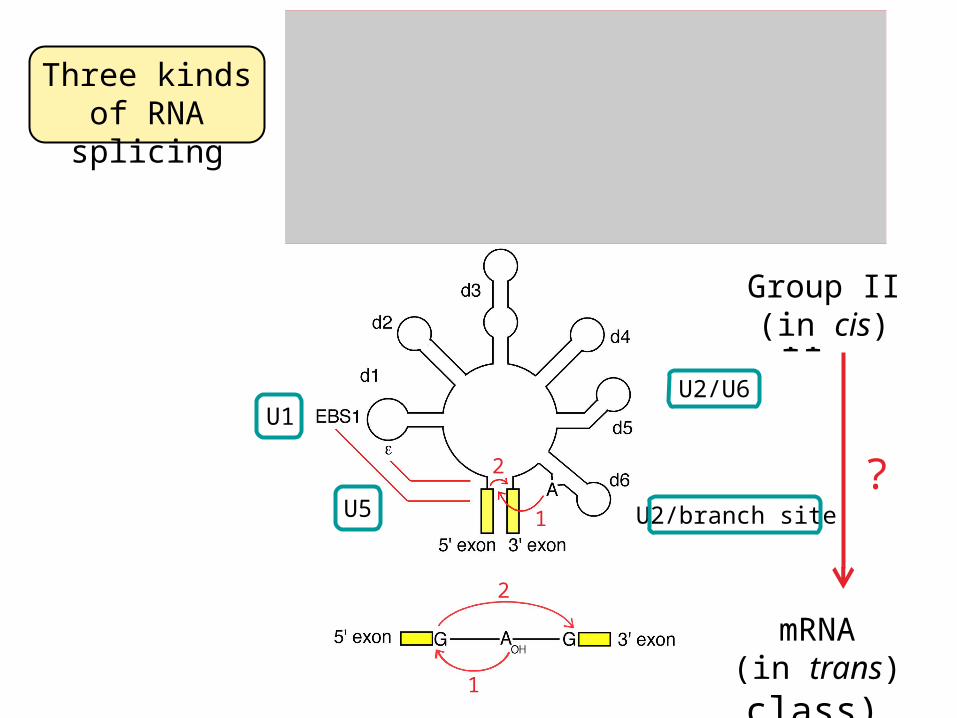
Three kinds of RNA splicing
U2/branch site
U2/U6U1
Group I
Group II
mRNA(major class)
mRNA(in trans)
Group I(in cis)
Group II(in cis)
?
1
2
2
1U5
U5
U1
Are the five spliceosomal snRNAs really fragments of a Group II self-splicing intron?
U4
U6
5’ splice siterecognition
branch siterecognition
catalysis
U2
catalysis
repressorand
(re)assembly factor
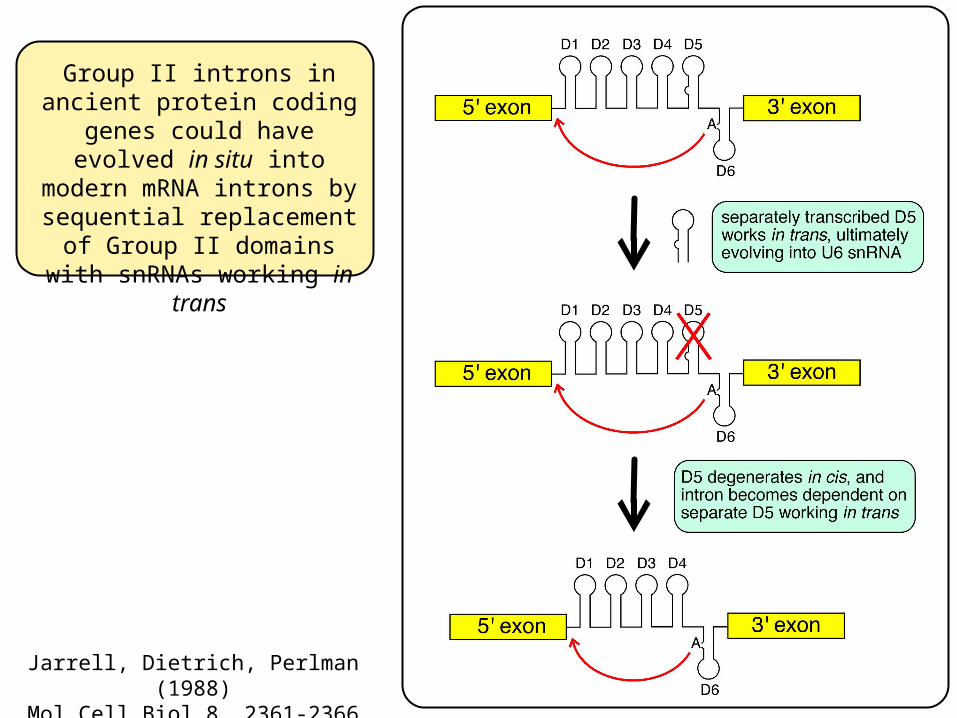
Group II introns in ancient protein coding genes could
have evolved in situ into modern mRNA introns by sequential replacement of
Group II domains with snRNAs working in trans
Jarrell, Dietrich, Perlman (1988)Mol Cell Biol 8, 2361-2366
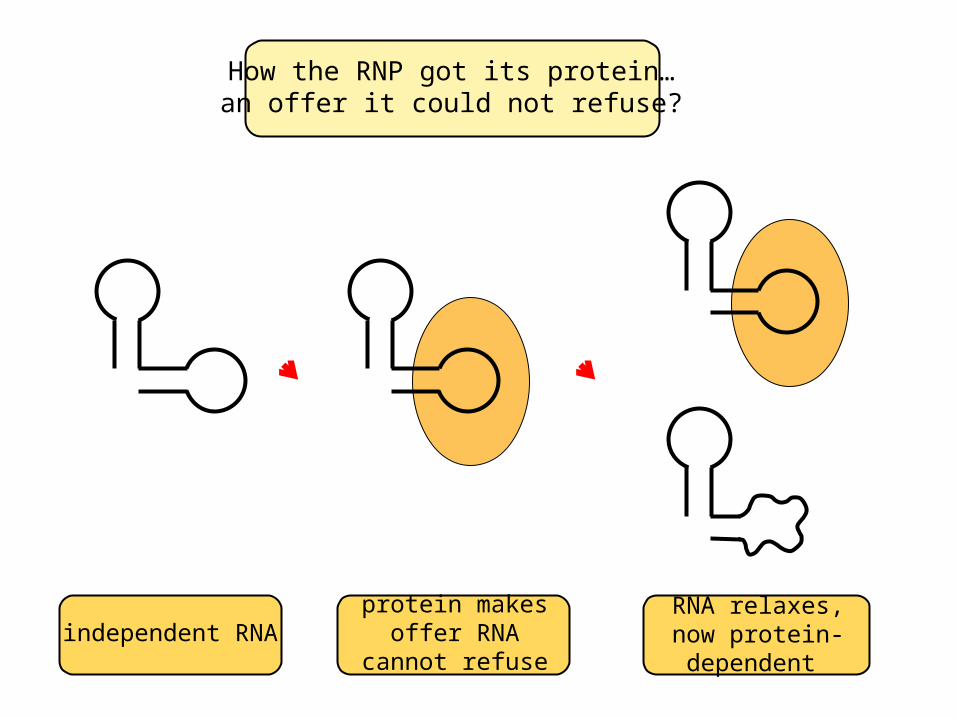
How the RNP got its protein…an offer it could not refuse?
independent RNAprotein makes offer RNA cannot refuse
RNA relaxes, now protein-dependent





























