Embed Size (px)
DESCRIPTION
Aticle about algae
Citation preview

1 3
PlantaDOI 10.1007/s00425-013-1946-5
OrIgInal artIcle
Conversion of membrane lipid acyl groups to triacylglycerol and formation of lipid bodies upon nitrogen starvation in biofuel green algae Chlorella UTEX29
Elton C. Goncalves · Jodie V. Johnson · Bala Rathinasabapathi
received: 11 May 2013 / accepted: 1 august 2013 © Springer-Verlag Berlin Heidelberg 2013
controlling oil accumulation in n-starved biofuel algae and demonstrate membrane recycling during lipid body formation.
Keywords Biofuel · Chlorella · galactolipid · radiolabeling · triacylglycerol · Ultrastructure
Introduction
the development of alternative sources of renewable energy is of utmost importance as world energy consump-tion is projected to grow by 53 % from 2008 to 2035 (Hu et al. 2008). although renewable fuels are the fastest grow-ing form of energy, its contribution to total energy pro-duction is projected to increase a mere 4 % by 2035 (Hu et al. 2008). In addition, the fact that US transportation fuels must contain 36 billion gallons of renewable fuels by 2022 (chisti 2007) highlights the importance of all efforts put into developing renewable fuels to an affordable, sus-tainable and scalable level. algal lipid from microalgae is one of the best sources for biofuels due to the following reasons: impressively high algal growth rates (commonly doubling its biomass within 24 h) (chisti 2007), tolerance to extreme conditions (desert and arid lands) (Hu et al. 2008), their abilities to recycle wastewater (agricultural run-off, industrial and municipal wastewater) and cO2 from flue gases emitted from power plants (Hu et al. 2008; Wilkie et al. 2011), reduced competition with food crops (griffiths and Harrison 2009) and, finally, the production of multiple value-added co-products (e.g., polymers, sur-factants, proteins and pigments) (Foley et al. 2011; Wilkie et al. 2011) in parallel to the accumulation of large quanti-ties of neutral lipids (up to 80 % by weight of dry biomass) (chisti 2007).
Abstract algal lipids are ideal biofuel sources. Our objective was to determine the contributors to triacylglyc-erol (tag) accumulation and lipid body formation in Chlo-rella UteX29 under nitrogen (n) deprivation. a fivefold increase in intracellular lipids following n starvation for 24 h confirmed the oleaginous characteristics of UteX29. Ultrastructural studies revealed increased number of lipid bodies and decreased starch granules in n-starved cells compared to n-replete cells. lipid bodies were observed as early as 3 h after n removal and plastids collapsed after 48 h of stress. Moreover, the identification of intracellu-lar pyrenoids and differences in the expected nutritional requirements for Chlorella protothecoides (as UteX29 is currently classified) led us to conduct a phylogenetic study using 18S and actin cDna sequences. this indicated UteX29 to be more phylogenetically related to Chlorella vulgaris. to investigate the fate of different lipids after n starvation, radiolabeling using 14c-acetate was used. a sig-nificant decrease in 14c-galactolipids and phospholipids matched the increase in 14c-tag starting at 3 h of n star-vation, consistent with acyl groups from structural lipids as sources for tag under n starvation. these results have important implications for the identification of key steps
Electronic supplementary material the online version of this article (doi:10.1007/s00425-013-1946-5) contains supplementary material, which is available to authorized users.
e. c. goncalves · B. rathinasabapathi (*) Plant Molecular and cellular Biology Program, Horticultural Sciences Department, University of Florida, gainesville, Fl 32611-0690, USae-mail: [email protected]
J. V. Johnson chemistry Department, University of Florida, gainesville, Fl 32611-7200, USa

Planta
1 3
green microalgae, especially within the genus Chlorella, are known for their great potential for high-quality biofuel production as they can grow in a variety of environmental conditions, use different carbon sources, achieve high bio-mass and lipid content (up to 55 % of cellular dwt) with adequate fatty acid composition (chen and Walker 2011; guarnieri et al. 2011; Heredia-arroyo et al. 2010; Miao and Wu 2004; O’grady and Morgan 2011; Xu et al. 2006). Several studies have investigated the effect of nitrogen star-vation as an inducer for triacylglycerol (tag) accumula-tion in green algae, with most of the studies conducted in the model green algae Chlamydomonas reinhardtii (good-son et al. 2011; James et al. 2011; Miller et al. 2010; Muj-taba et al. 2012; nguyen et al. 2011; Yeh and chang 2011). However, many aspects of tag accumulation in response to nitrogen starvation remain unresolved. For instance, most studies on tag accumulation examined cells after 24 h or later time points of n starvation, not clarifying the early events of the process. Moreover, it is not known whether what is observed in Chlamydomonas is conserved in other green algae with greater potential for outdoor, large-scale production of biofuels.
a study on Chlamydomonas suggested that the de novo synthesis of fatty acids in the chloroplast is a limit-ing factor for tag production, although the recycling of membrane lipids could account for up to 30 % of the tag produced under n starvation (Fan et al. 2011). Moreover, a tracer experiment with 13c-labeled glucose in Chlorella protothecoides indicated overall stability in global carbon flux during n starvation but did not assess the contribution of recycling of membrane lipids into tag (Xiong et al. 2010). In this study, a series of radiolabeling experiments with 14c-acetate was performed to track membrane lipids’ remodeling into tag during n starvation in a green algal strain with potential for outdoor cultivation, with further identification of lipids through HPlc/UV/(+)eSI–MS2. Molecular phylogeny of cDnas was used to confirm the taxonomic status of the strain. a time-course transmission electron microscopy (teM) study revealed major ultras-tructural changes occurring during the early and late events of lipid droplet formation.
Materials and methods
algal cultures and nitrogen starvation
UteX29 (as of Jan 2012, named as C. protothecoides by the stock center) was purchased from the algae culture collection at the University of texas, austin, and grown in liquid medium under a light:dark regime of 18:6 h (80 μmol photons/m2/s), under continuous agitation at 130 rpm. a nitrogen-replete (+n) mineral basal algal
medium (BaM) (Sorokin and Krauss 1958) was prepared with the following composition (in g/l): KnO3, 1.25; KH2PO4, 1.25; MgSO4·7H20, 1.0; cacl2, 0.0835; H3BO3, 0.1142; FeSO4·7H20, 0.0498; ZnSO4·7H20, 0.0882; Mncl2·4H2O, 0.0144; MoO3, 0.0071; cuSO4·5H2O, 0.0157; co(nO3)2·6H20, 0.0049; eDta, 0.5; and filter-sterilized glucose, 9. the pH was adjusted to 6.8 before autoclaving. For the nitrogen-free (−n) medium, KnO3 was replaced by Kcl for the same final potassium con-centration. to induce nitrogen deficiency, algal cells pre-viously grown in n-replete medium were washed once in −n medium, suspended in fresh −n medium and cultured for 3–48 h. cell counting was done using a hemocytom-eter counting chamber (Fisher Scientific, USa) and turbid-ity measurements were done using a spectrophotometer (DU730, Beckman coulter, Pasadena, ca). the relation-ship between OD600 and cell density was determined as 1 OD600 = 8.3 × 108 cells/ml within a maximum deviation of 14 % of that relationship in both nitrogen-replete and nitrogen-deficient cultures. For calculations, we therefore employed OD600 for normalizing for cell numbers.
lipid staining using nile red
Detection of intracellular lipid bodies was conducted as previously described (chen et al. 2009). corresponding amounts of cells were treated with a solution of the vital flu-orescent stain nile red (greenspan et al. 1985) (50 μg/l) and DMSO (20 % v/v) to increase membrane permeability. after 10 min of incubation, the relative fluorescence (excita-tion at 530 nm and emission at 570 nm) was measured in a Synergy Ht microplate reader (Biotek, USa).
gravimetric method to quantify lipid productivity
cells from nitrogen-replete or nitrogen-depleted cultures were collected by centrifugation at 3,500 rpm at 10 °c for 15 min in a Heraeus labofuge 400r centrifuge in 50-ml tubes. the cell pellets were extracted for total lipids as described elsewhere. the solvent in the lipid extract was evaporated in a nitrogen evaporator (Organomation, Ma) and the total lipid fraction was dried in an oven set at 50 °c until two consecutive weighings were constant. the lipid yield was expressed as g/100 g dry weight of cells.
transmission electron microscopy
algal cells were collected by centrifugation (5 min, 3,500 rpm in swing-out rotors of a Heraeus labofuge 400r centrifuge) fixed in 3 % (v/v) glutaraldehyde in BaM medium overnight and washed three times with 0.1 M sodium cacodylate, 2 mM Mgcl2, 1 mM cacl2, 0.25 % (wt/v) nacl pH 7.23. Fixed cells were processed with the

Planta
1 3
aid of a Pelco BioWave laboratory microwave (ted Pella, redding, ca, USa). the samples were buffer washed, post-fixed with 2 % (wt/v) OsO4, water washed, dehydrated in a graded ethanol series 25, 30, 40, 50, 55, 60, 65, 80, 90, 100 % (v/v), infiltrated with lrWhite acrylic resin (electron Microscopy Sciences, Hatfield, Pa) and cured at 60 °c for 2 days. cured resin blocks were trimmed, thin sectioned and collected on formvar copper 200 mesh grids, post-stained with 2 % aqueous uranyl acetate and rey-nold’s lead citrate. Sections were examined with a Hitachi H-7000 teM (Hitachi High technologies america, Inc., Schaumburg, Il) and digital images were acquired with a Veleta 2k × 2k camera and iteM software (Olympus Soft-Imaging Solutions corp., lakewood, cO). the teM experiments were done in triplicates and repeated twice. three slides were prepared from each replicate and 30 cells were analyzed from each slide.
radiotracer labeling using 14c-acetate
[2-14c]acetic acid (na salt in water) was purchased from american radiolabeled chemicals Inc (St. louis, MO). For labeling studies, the cells were initially grown in 250 ml flasks containing 100 ml of +n medium until the mid-log-arithmic phase, transferred to 4.5 ml glass vials for a 24-h incubation with 3 ml of +n medium containing [14c]-ace-tate (50 nci/μl), followed by centrifugation for 10 min at 3,500 rpm in a swing-out rotor of a Heraeus labofuge 400r centrifuge, removal of supernatant, and finally addition of +n or −n, label-free medium, for a 48-h growth period.
In the pulse-chase experiment, cells were incubated for 1 h in +n medium containing [14c]-acetate (50 nci/μl), centrifuged for 10 min at 3,500 rpm in a swing-out rotor of a Heraeus labofuge 400r centrifuge, supernatant was removed, cell pellets were washed with −n medium and suspended in −n medium containing 0.5 mM unlabeled acetate. the experiments were conducted in triplicate.
extraction of total lipids
equal number of cells were harvested from different treat-ments by centrifugation (10 min, 8,000 rpm, in an eppen-dorf 5418 centrifuge) and frozen in liquid nitrogen. the lipid extraction method reported by Fan et al. (2011) was used with minor modifications. total lipids were extracted with methanol–chloroform–formic acid (2:1:0.1, v:v:v) in the presence of glass beads under maximum agitation for 5 min using a Vortex genie 2 vortex (Fisher Scientific). Maximum extraction efficiency was achieved by repeating this step 7×, followed by phase separation using 1 M Kcl, 0.2 M H3PO4 solution. this procedure was sufficient to extract all lipids detectable by iodine staining in thin layer chromatography (tlc).
analysis of lipids using thin layer chromatography
total lipid extracts were separated in baked ammonium sulfate-impregnated silica plates (J.t. Baker, USa) by thin layer chromatography (tlc) using a double-development solvent system (2/3 in acetone–toluene–water (91:30:3 by volume) followed by full development in hexane–diethyl ether–acetic acid (70:30:1 by volume) (Fan et al. 2011). tlc plates were briefly stained with iodine vapor for quick visualization of lipid bands. Bands were identified based on their relative mobilities (rf) compared to standard lipids. For the quantification of radioactivity, the bands were scrapped off the plate, mixed with 2 ml distilled water and 2 ml ready gel liquid scintillation cocktail, homogenized and radioactivity counted in a lS6500 liquid scintillation counter (Beckman, USa).
HPlc/UV/(+)eSI–mass spectrometry–mass spectrometry
lipid bands isolated from the tlc plates were dissolved in isopropanol and separated using an HPlc (1100 series model g1312a binary pump, agilent, Palo alto, ca) equipped with a c8 column (Hypurity 5 μm; 2.1 × 100 mm with a c8 guard column; thermo Scientific) and gradients utilizing a binary mobile phase system of water and iso-propanol (both were HPlc-grade; Honeywell Burdick & Jackson, Muskegon, MI) with a flow rate of 0.15 ml/min. For the data presented here, the gradient was set to 30 % isopropanol at time 0 and then increased linearly to 60 % isopropanol in 15 min and then to 95 % isopropanol in 50 min and then held at 95 % isopropanol for 20 min. all mass spectrometric data were obtained with a thermoFinni-gan (San Jose, ca) lcQ fitted with the conventional elec-trospray ionization source. the sheath and auxiliary gases were nitrogen (65 and 5, respectively, instrumental unitless parameter) and the heated capillary temperature was set at 250 °c. For (+)eSI–MSn, the spray voltage was set at 3.3 kV, the heated capillary voltage at +12.5 V and the tube lens offset at 0 V. collision-induced dissociation (cID) tan-dem mass spectrometry (MSn) of [M + na]+ ions was per-formed with 3–7 u precursor ion isolation, 30 ms activation time and either 37.5 % normalized cID energy at 0.25 qcID or 40 % normalized cID energy at 0.30 qcID. Ultraviolet/visible (UV) detection at 210 or 220 nm was obtained with an agilent 1100 series g1314a UV/Vis detector.
cloning 18s rrna and actin mrna partial sequences from UteX29
rna extraction was done using nucleoSpin rna Plant Kit (Macherey–nagel, germany). Synthesis of cDna and rt-Pcr were performed using Superscript One-Step rt-Pcr with Platinum taq (Invitrogen, USa) according

Planta
1 3
to the manufacturer’s instructions. the primers for 18s rrna were 5′-Wacctggttgatcctgccagt-3′ and 5′-gatccttcYgcaggttcacctac-3′. the primers for actin were 5′-gtgaccaactgggacgac-3′ and 5′-cgggcagctcgtaIgtctt-3′. the gel-extracted Pcr products were cloned into pcr 2.1-tOPO vector and sequenced (Sambrook 2001).
Molecular phylogeny of UteX29 partial sequences
Database searches were performed using UteX29 18s rrna and actin cDna sequences as queries. Homologous sequences within the taxa “green algae” were obtained by a Basic local alignment Search tool (BlaSt) search at the national center for Biotechnology Information. the sequences with the best e value, coverage and supporting lit-erature were chosen for further analysis (table S1). Multiple sequence alignment was conducted using clustalW software in Mega5 (tamura et al. 2011). Phylogenetic trees were inferred from the aligned sequence data using the neighbor-joining method (Saitou and nei 1987) in Mega5, with the tree being tested by bootstrapping with 1,000 replicates.
Statistical analysis
Quantitative data were analyzed using JMP software, Ver-sion 7 (SaS Institute Inc., cary, nc). Means were com-pared using Student’s t test, α = 0.05. all experiments were conducted in triplicate.
Results
lipid accumulation
Upon arrival of strain 29 from the UteX algae collec-tion, cells were repeatedly streaked in sterile agar plates
containing BaM to guarantee that all experiments were done using axenic cultures. an evaluation of lipid accu-mulation using nile red staining of cells (greenspan et al. 1985) confirmed that Chlorella sp. UteX29 is an oil-accumulating species and demonstrated the effect of its cultivation under photoheterotrophic conditions (under light and addition of glucose into the medium). there was a ~fivefold increase in relative intracellular neutral lipids (as measured by nile red fluorescence) upon 24 h of growth under n-free medium compared to n-replete cultures (Sup-plemental Fig. 1a). total lipid content of n-deplete cul-tures, evaluated by gravimetric method, was 68 % by dry weight (Fig. 1b).
Ultrastructure of nitrogen-starved cells
In order to investigate the event of membrane recycling and the formation of lipid bodies under n-starvation stress, sample cells were fixed for teM after growing them in the presence or absence of n. the microscopic observa-tions supported and confirmed the increased formation of lipid bodies after 48 h of stress, first detected by the nile red assay (Supplemental Fig. 1). the ultrastructure of cells from n-replete medium indicated abundant presence of starch granules and pyrenoid structures in a single large chloroplast, with rare presence of small lBs (Fig. 1a), as previously described in ultrastructural studies in other Chlorella species (Bertagnolli and nadakavukaren 1970; Pyliotis and goodchild 1975). cells in the n-free medium displayed very abundant and large lipid bodies and mor-phological symptoms of programmed cell death (Zuo et al. 2012), with most organelles undergoing fragmenta-tion (Fig. 1b). Of 30 cells randomly chosen for observa-tion from 3 independent microscopic preparations, only 2 of the control cells (under +n condition) had small lBs, whereas 29 of the 30 observed cells under 48 h of n starva-tion contained large lBs. this result led us to investigate
Fig. 1 Ultrastructure of nitrogen-starved cells indicated lipid accumulation and plastid degradation. a cells grown in n-replete medium for 48 h. b cells grown in n-free medium for 48 h. p Pyrenoid, s starch, LB lipid body, Ch chloroplast. See “Materials and methods” for details on processing the cells
Planta
1 3
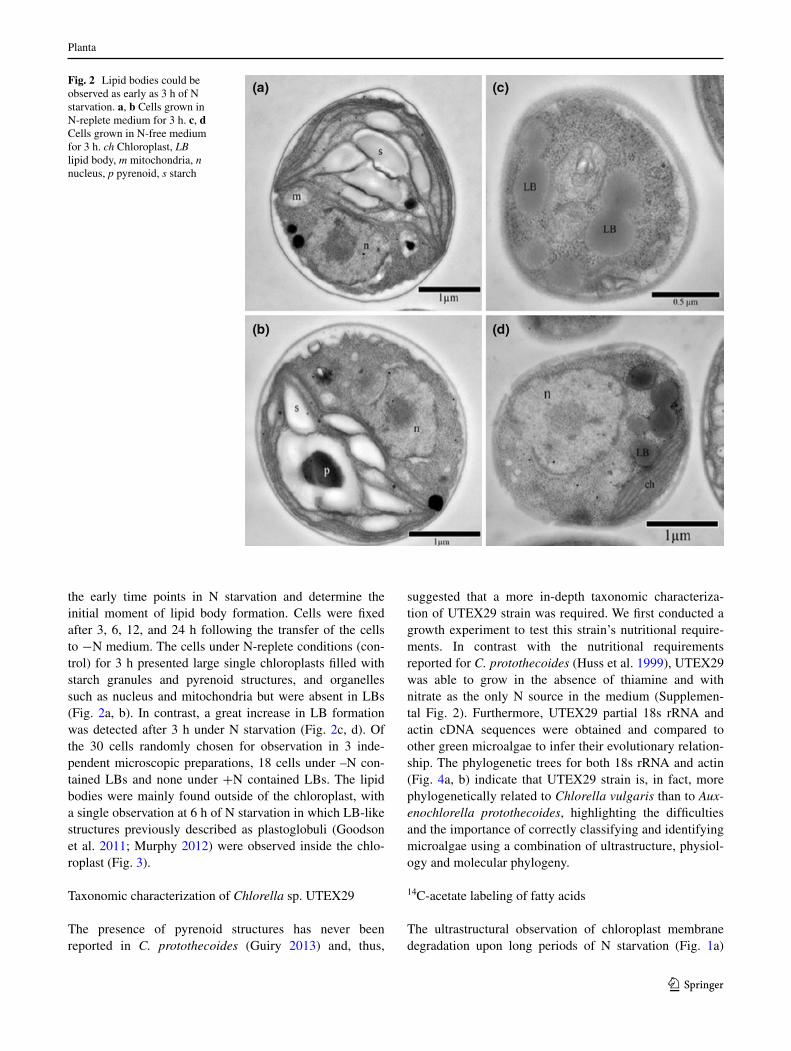
the early time points in n starvation and determine the initial moment of lipid body formation. cells were fixed after 3, 6, 12, and 24 h following the transfer of the cells to −n medium. the cells under n-replete conditions (con-trol) for 3 h presented large single chloroplasts filled with starch granules and pyrenoid structures, and organelles such as nucleus and mitochondria but were absent in lBs (Fig. 2a, b). In contrast, a great increase in lB formation was detected after 3 h under n starvation (Fig. 2c, d). Of the 30 cells randomly chosen for observation in 3 inde-pendent microscopic preparations, 18 cells under –n con-tained lBs and none under +n contained lBs. the lipid bodies were mainly found outside of the chloroplast, with a single observation at 6 h of n starvation in which lB-like structures previously described as plastoglobuli (goodson et al. 2011; Murphy 2012) were observed inside the chlo-roplast (Fig. 3).
taxonomic characterization of Chlorella sp. UteX29
the presence of pyrenoid structures has never been reported in C. protothecoides (guiry 2013) and, thus,
suggested that a more in-depth taxonomic characteriza-tion of UteX29 strain was required. We first conducted a growth experiment to test this strain’s nutritional require-ments. In contrast with the nutritional requirements reported for C. protothecoides (Huss et al. 1999), UteX29 was able to grow in the absence of thiamine and with nitrate as the only n source in the medium (Supplemen-tal Fig. 2). Furthermore, UteX29 partial 18s rrna and actin cDna sequences were obtained and compared to other green microalgae to infer their evolutionary relation-ship. the phylogenetic trees for both 18s rrna and actin (Fig. 4a, b) indicate that UteX29 strain is, in fact, more phylogenetically related to Chlorella vulgaris than to Aux-enochlorella protothecoides, highlighting the difficulties and the importance of correctly classifying and identifying microalgae using a combination of ultrastructure, physiol-ogy and molecular phylogeny.
14c-acetate labeling of fatty acids
the ultrastructural observation of chloroplast membrane degradation upon long periods of n starvation (Fig. 1a)
Fig. 2 lipid bodies could be observed as early as 3 h of n starvation. a, b cells grown in n-replete medium for 3 h. c, d cells grown in n-free medium for 3 h. ch chloroplast, LB lipid body, m mitochondria, n nucleus, p pyrenoid, s starch
Planta
1 3
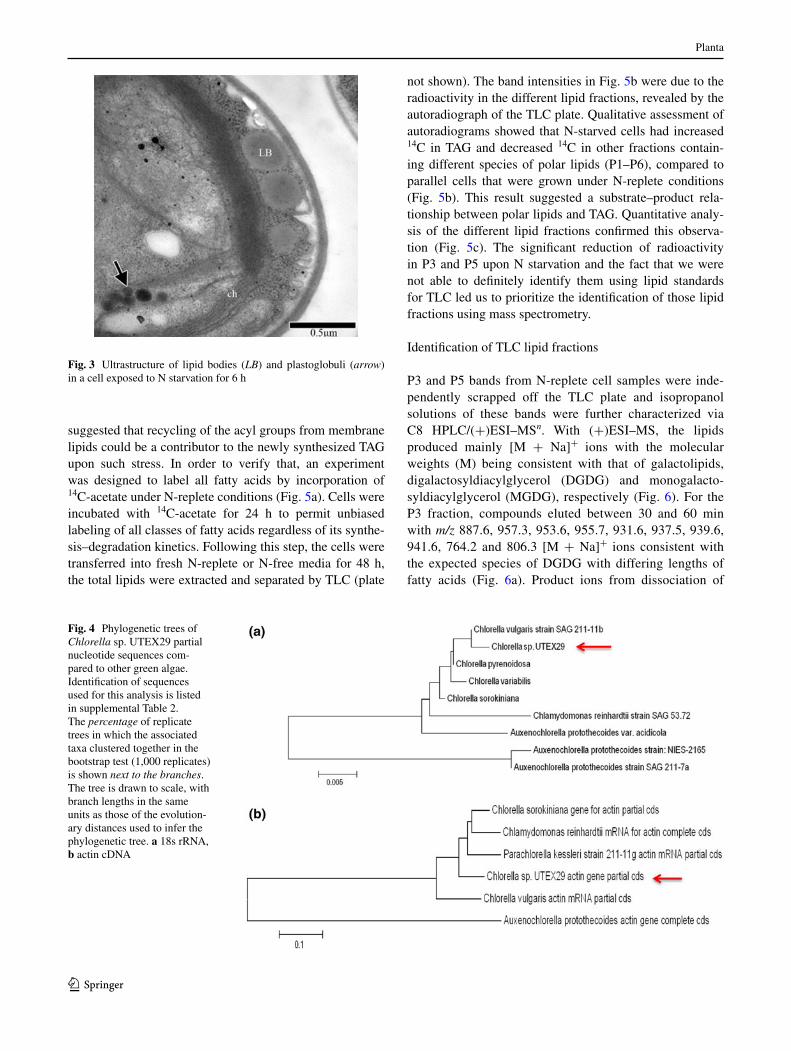
suggested that recycling of the acyl groups from membrane lipids could be a contributor to the newly synthesized tag upon such stress. In order to verify that, an experiment was designed to label all fatty acids by incorporation of 14c-acetate under n-replete conditions (Fig. 5a). cells were incubated with 14c-acetate for 24 h to permit unbiased labeling of all classes of fatty acids regardless of its synthe-sis–degradation kinetics. Following this step, the cells were transferred into fresh n-replete or n-free media for 48 h, the total lipids were extracted and separated by tlc (plate
not shown). the band intensities in Fig. 5b were due to the radioactivity in the different lipid fractions, revealed by the autoradiograph of the tlc plate. Qualitative assessment of autoradiograms showed that n-starved cells had increased 14c in tag and decreased 14c in other fractions contain-ing different species of polar lipids (P1–P6), compared to parallel cells that were grown under n-replete conditions (Fig. 5b). this result suggested a substrate–product rela-tionship between polar lipids and tag. Quantitative analy-sis of the different lipid fractions confirmed this observa-tion (Fig. 5c). the significant reduction of radioactivity in P3 and P5 upon n starvation and the fact that we were not able to definitely identify them using lipid standards for tlc led us to prioritize the identification of those lipid fractions using mass spectrometry.
Identification of tlc lipid fractions
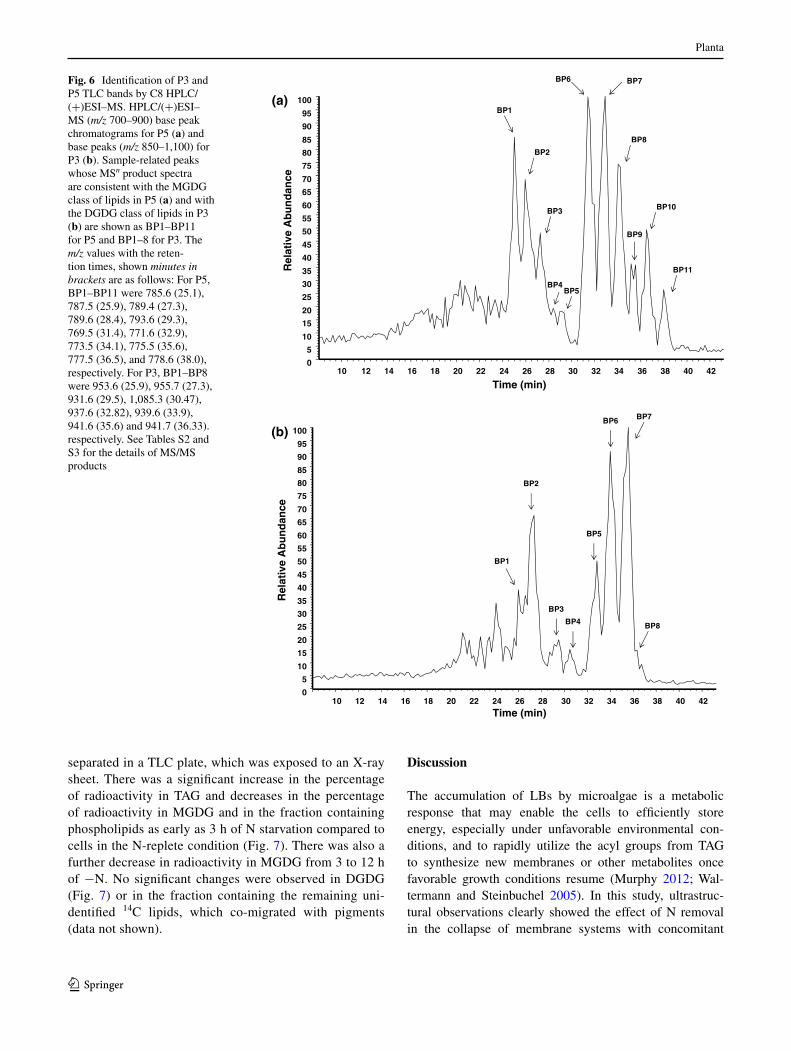
P3 and P5 bands from n-replete cell samples were inde-pendently scrapped off the tlc plate and isopropanol solutions of these bands were further characterized via c8 HPlc/(+)eSI–MSn. With (+)eSI–MS, the lipids produced mainly [M + na]+ ions with the molecular weights (M) being consistent with that of galactolipids, digalactosyldiacylglycerol (DgDg) and monogalacto-syldiacylglycerol (MgDg), respectively (Fig. 6). For the P3 fraction, compounds eluted between 30 and 60 min with m/z 887.6, 957.3, 953.6, 955.7, 931.6, 937.5, 939.6, 941.6, 764.2 and 806.3 [M + na]+ ions consistent with the expected species of DgDg with differing lengths of fatty acids (Fig. 6a). Product ions from dissociation of
Fig. 3 Ultrastructure of lipid bodies (LB) and plastoglobuli (arrow) in a cell exposed to n starvation for 6 h
Fig. 4 Phylogenetic trees of Chlorella sp. UteX29 partial nucleotide sequences com-pared to other green algae. Identification of sequences used for this analysis is listed in supplemental table 2. the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) is shown next to the branches. the tree is drawn to scale, with branch lengths in the same units as those of the evolution-ary distances used to infer the phylogenetic tree. a 18s rrna, b actin cDna
Planta
1 3
the [M + na]+ ions confirmed the parent molecules to be DgDgs (table S2). For the P5 fraction, compounds eluted between 30 and 60 min with m/z 785.4, 787.4, 787.5, 769.6, 771.6, 773.6, 779.5, 619.6 and 806.2 [M + na]+ ions consistent with the expected species of MgDg with differing lengths of fatty acids (Fig. 6b). the patterns of the product ions from parent [M + na]+ ions observed in the P5 fraction were consistent with the pres-ence of MgDg in the fraction P5 (table S2). the regi-ochemical distribution of the acyl chains in MgDg and DgDg (tables S2, S3, respectively), considering no mod-ifications occurred to the molecules while processing the samples, were tentatively deduced as previously described (guella et al. 2003). Using the same strategy, phosphati-dylcholine (Pc) was identified as the major contributor to tlc fraction P1. Phosphatidylethanolamine (Pe) is also expected to be present in P1, by comparing to the previ-ously described rf values in this solvent system (Fan et al. 2011), but its low abundance may have hindered
detection. We tentatively identified P2 as sulfoquinovo-syldiacylglycerol as its mobility in the tlc plate (rf values) matched the previously reported values in Chla-mydomonas (Fan et al. 2011). Fraction P4 co-migrated with P3 and was analyzed as a single sample, reflecting the wide variety of DgDg species detected (table S3). Fraction P6 co-migrated with pigments such as chloro-phylls and was not identified in this study.
Pulse-chase of 14c-acetate-labeled fatty acids
a pulse-chase experiment was done to investigate the moment in which the remodeling of polar lipids into tag began after n starvation. after incubating the cell cultures for 1 h with 14c-acetate, fatty acid labeling was stopped by transferring the cell cultures to fresh n-replete or n-free medium containing excess unlabeled acetate. the cells were harvested after 1, 3 and 12 h following this transfer. total lipids were extracted and fractions were
0
10
20
30
40
50
60
70
80
P1 P2 P3 P4 P5 P6 TAG
Rad
ioac
tivi
ty (
%)
Lipid fraction
Plus N Minus N
+N for 24h
*Centrifuge*Wash (-N)*Resuspend
+N for 48h
-N for 48h
Total Lipids extracted
and separatedby TLC
TAG
+N -N
P:5
P:3 **
* *
*
*
P:6
P:2
P:4
P:1
(a)
(c)
(b)
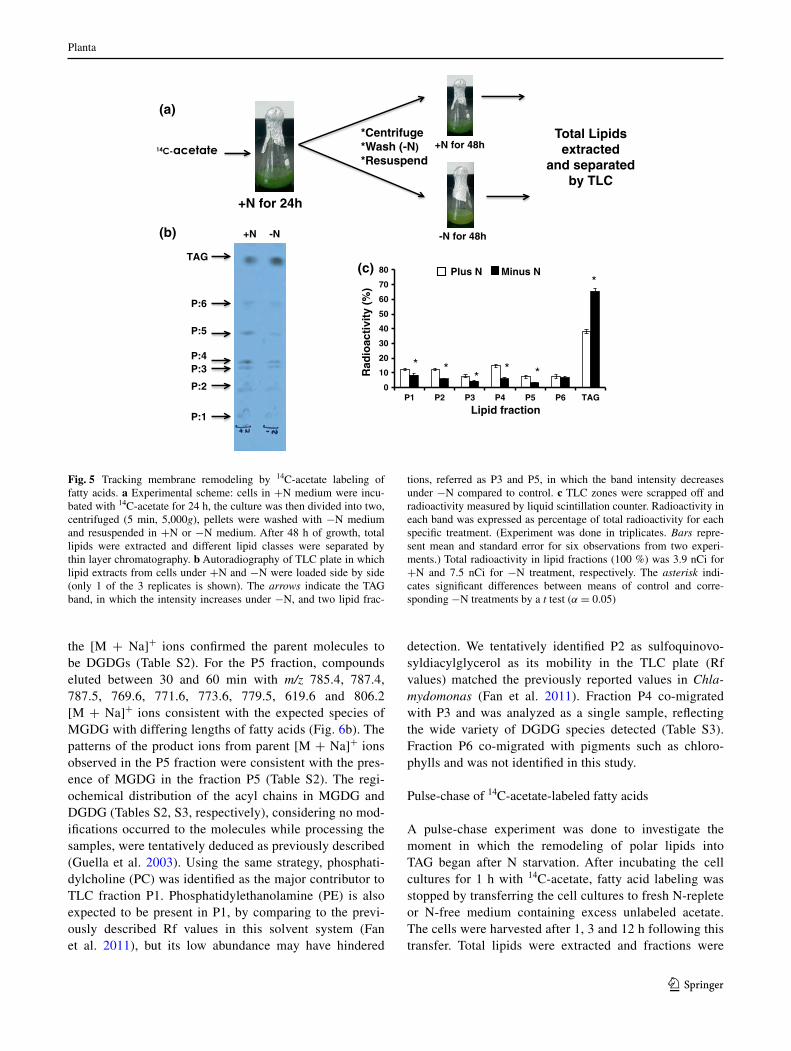
Fig. 5 tracking membrane remodeling by 14c-acetate labeling of fatty acids. a experimental scheme: cells in +n medium were incu-bated with 14c-acetate for 24 h, the culture was then divided into two, centrifuged (5 min, 5,000g), pellets were washed with −n medium and resuspended in +n or −n medium. after 48 h of growth, total lipids were extracted and different lipid classes were separated by thin layer chromatography. b autoradiography of tlc plate in which lipid extracts from cells under +n and −n were loaded side by side (only 1 of the 3 replicates is shown). the arrows indicate the tag band, in which the intensity increases under −n, and two lipid frac-
tions, referred as P3 and P5, in which the band intensity decreases under −n compared to control. c tlc zones were scrapped off and radioactivity measured by liquid scintillation counter. radioactivity in each band was expressed as percentage of total radioactivity for each specific treatment. (experiment was done in triplicates. Bars repre-sent mean and standard error for six observations from two experi-ments.) total radioactivity in lipid fractions (100 %) was 3.9 nci for +n and 7.5 nci for −n treatment, respectively. the asterisk indi-cates significant differences between means of control and corre-sponding −n treatments by a t test (α = 0.05)
Planta
1 3
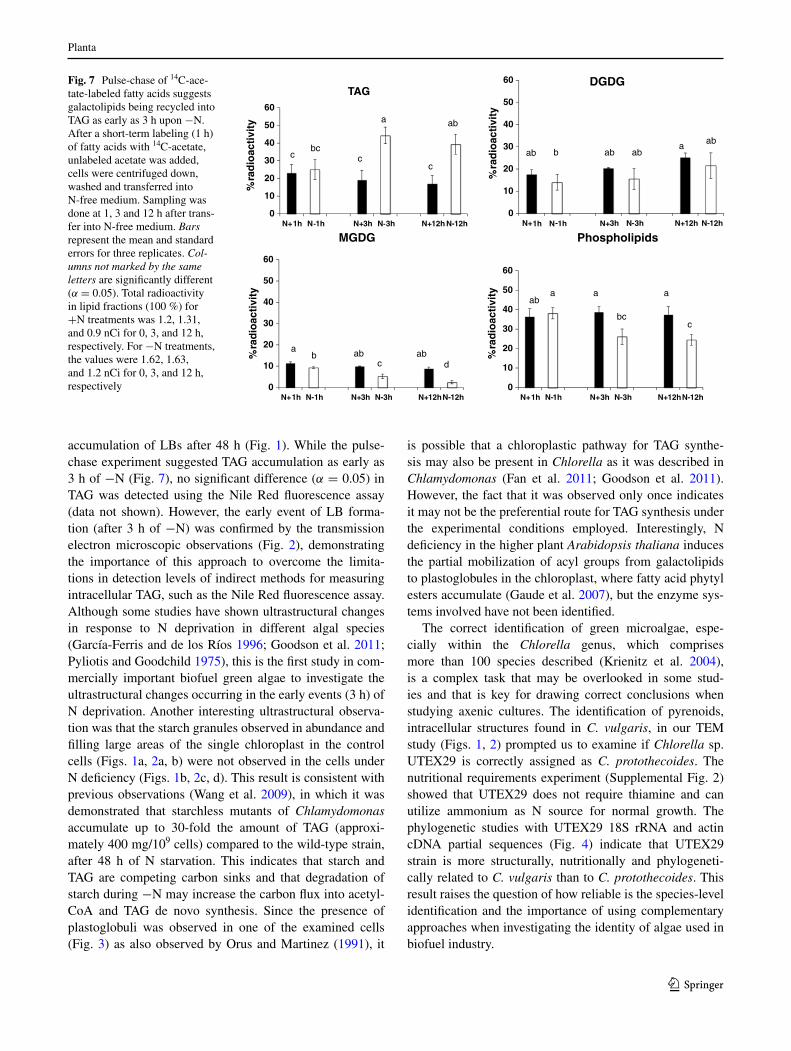
separated in a tlc plate, which was exposed to an X-ray sheet. there was a significant increase in the percentage of radioactivity in tag and decreases in the percentage of radioactivity in MgDg and in the fraction containing phospholipids as early as 3 h of n starvation compared to cells in the n-replete condition (Fig. 7). there was also a further decrease in radioactivity in MgDg from 3 to 12 h of −n. no significant changes were observed in DgDg (Fig. 7) or in the fraction containing the remaining uni-dentified 14c lipids, which co-migrated with pigments (data not shown).
Discussion
the accumulation of lBs by microalgae is a metabolic response that may enable the cells to efficiently store energy, especially under unfavorable environmental con-ditions, and to rapidly utilize the acyl groups from tag to synthesize new membranes or other metabolites once favorable growth conditions resume (Murphy 2012; Wal-termann and Steinbuchel 2005). In this study, ultrastruc-tural observations clearly showed the effect of n removal in the collapse of membrane systems with concomitant
12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Rel
ativ
e A
bu
nd
ance
10
BP1
BP2
BP3
BP4BP5
BP6 BP7
BP8
BP9
BP10
BP11
(a)
10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Rel
ativ
e A
bu
nd
ance
(b)
BP1
BP2
BP3
BP4
BP5
BP6 BP7
BP8
Fig. 6 Identification of P3 and P5 tlc bands by c8 HPlc/(+)eSI–MS. HPlc/(+)eSI–MS (m/z 700–900) base peak chromatograms for P5 (a) and base peaks (m/z 850–1,100) for P3 (b). Sample-related peaks whose MSn product spectra are consistent with the MgDg class of lipids in P5 (a) and with the DgDg class of lipids in P3 (b) are shown as BP1–BP11 for P5 and BP1–8 for P3. the m/z values with the reten-tion times, shown minutes in brackets are as follows: For P5, BP1–BP11 were 785.6 (25.1), 787.5 (25.9), 789.4 (27.3), 789.6 (28.4), 793.6 (29.3), 769.5 (31.4), 771.6 (32.9), 773.5 (34.1), 775.5 (35.6), 777.5 (36.5), and 778.6 (38.0), respectively. For P3, BP1–BP8 were 953.6 (25.9), 955.7 (27.3), 931.6 (29.5), 1,085.3 (30.47), 937.6 (32.82), 939.6 (33.9), 941.6 (35.6) and 941.7 (36.33). respectively. See tables S2 and S3 for the details of MS/MS products
Planta
1 3
accumulation of lBs after 48 h (Fig. 1). While the pulse-chase experiment suggested tag accumulation as early as 3 h of −n (Fig. 7), no significant difference (α = 0.05) in tag was detected using the nile red fluorescence assay (data not shown). However, the early event of lB forma-tion (after 3 h of −n) was confirmed by the transmission electron microscopic observations (Fig. 2), demonstrating the importance of this approach to overcome the limita-tions in detection levels of indirect methods for measuring intracellular tag, such as the nile red fluorescence assay. although some studies have shown ultrastructural changes in response to n deprivation in different algal species (garcía-Ferris and de los ríos 1996; goodson et al. 2011; Pyliotis and goodchild 1975), this is the first study in com-mercially important biofuel green algae to investigate the ultrastructural changes occurring in the early events (3 h) of n deprivation. another interesting ultrastructural observa-tion was that the starch granules observed in abundance and filling large areas of the single chloroplast in the control cells (Figs. 1a, 2a, b) were not observed in the cells under n deficiency (Figs. 1b, 2c, d). this result is consistent with previous observations (Wang et al. 2009), in which it was demonstrated that starchless mutants of Chlamydomonas accumulate up to 30-fold the amount of tag (approxi-mately 400 mg/109 cells) compared to the wild-type strain, after 48 h of n starvation. this indicates that starch and tag are competing carbon sinks and that degradation of starch during −n may increase the carbon flux into acetyl-coa and tag de novo synthesis. Since the presence of plastoglobuli was observed in one of the examined cells (Fig. 3) as also observed by Orus and Martinez (1991), it
is possible that a chloroplastic pathway for tag synthe-sis may also be present in Chlorella as it was described in Chlamydomonas (Fan et al. 2011; goodson et al. 2011). However, the fact that it was observed only once indicates it may not be the preferential route for tag synthesis under the experimental conditions employed. Interestingly, n deficiency in the higher plant Arabidopsis thaliana induces the partial mobilization of acyl groups from galactolipids to plastoglobules in the chloroplast, where fatty acid phytyl esters accumulate (gaude et al. 2007), but the enzyme sys-tems involved have not been identified.
the correct identification of green microalgae, espe-cially within the Chlorella genus, which comprises more than 100 species described (Krienitz et al. 2004), is a complex task that may be overlooked in some stud-ies and that is key for drawing correct conclusions when studying axenic cultures. the identification of pyrenoids, intracellular structures found in C. vulgaris, in our teM study (Figs. 1, 2) prompted us to examine if Chlorella sp. UteX29 is correctly assigned as C. protothecoides. the nutritional requirements experiment (Supplemental Fig. 2) showed that UteX29 does not require thiamine and can utilize ammonium as n source for normal growth. the phylogenetic studies with UteX29 18S rrna and actin cDna partial sequences (Fig. 4) indicate that UteX29 strain is more structurally, nutritionally and phylogeneti-cally related to C. vulgaris than to C. protothecoides. this result raises the question of how reliable is the species-level identification and the importance of using complementary approaches when investigating the identity of algae used in biofuel industry.
0
10
20
30
40
50
60
N+ 1h N- 1h N+ 3h N- 3h N+ 12hN- 12h
% ra
dio
acti
vity
Phospholipids
aa a
bc
ab
c
0
10
20
30
40
50
60
N+ 1h N- 1h N+ 3h N- 3h N+ 12h N- 12h
% r
adio
acti
vity
DGDG
abb abab
aab
0
10
20
30
40
50
60
N+ 1h N- 1h N+ 3h N- 3h N+ 12hN- 12h
% ra
dio
acti
vity
MGDG
abc
abd
0
10
20
30
40
50
60
N+ 1h N- 1h N+ 3h N- 3h N+ 12h N- 12h
% ra
dio
acti
vity
TAG
c
a
c
ab
cbc
ab
Fig. 7 Pulse-chase of 14c-ace-tate-labeled fatty acids suggests galactolipids being recycled into tag as early as 3 h upon −n. after a short-term labeling (1 h) of fatty acids with 14c-acetate, unlabeled acetate was added, cells were centrifuged down, washed and transferred into n-free medium. Sampling was done at 1, 3 and 12 h after trans-fer into n-free medium. Bars represent the mean and standard errors for three replicates. Col-umns not marked by the same letters are significantly different (α = 0.05). total radioactivity in lipid fractions (100 %) for +n treatments was 1.2, 1.31, and 0.9 nci for 0, 3, and 12 h, respectively. For −n treatments, the values were 1.62, 1.63, and 1.2 nci for 0, 3, and 12 h, respectively

Planta
1 3
this study demonstrates that the oil-accumulating characteristic of the green algae Chlorella sp. UteX29 is reproducibly triggered by n removal from the medium, with a ~fivefold increase in detectable lipid content within 24 h of −n (Supplemental Fig. 1). Several studies have investigated the effect of nitrogen starvation as an inducer for tag accumulation in both the genus Chlorella (Hort-ensteiner et al. 2000; Pyliotis and goodchild 1975; Xiong et al. 2010; Yeh and chang 2011) and a related species, the model green algae C. reinhardtii (Fan et al. 2011; goodson et al. 2011; James et al. 2011; Miller et al. 2010; nguyen et al. 2011; Yeh and chang 2011). However, these studies focus at 24 h or later time points of n starvation, without clarifying the early events on lipid body formation. Moreo-ver, information regarding the contribution of membrane lipids toward tag synthesis is also very limited to date. a study on Chlamydomonas indicated that the de novo syn-thesis of fatty acids in the chloroplast is a limiting factor for tag production, and assuming that the decreases in polar lipid amounts during −n are entirely mobilized into tag, Fan et al. (2011) suggested that recycling of membrane lipids could account for up to 30 % of the tag synthesis after 48 h of n deprivation. By labeling the fatty acids with 14c-acetate and tracking the radioactivity shifts after 24 h of n removal (Fig. 5), Fan et al. (2011)’s assumption was confirmed, as a 34 % decrease in polar lipids radioactiv-ity was detected, which accounted for the same percentage increase in tag radioactivity (Fig. 5c). the 14c-acetate-labeling experiment in this study provides evidence in vivo to demonstrate a substrate–product relationship between tag and structural polar lipids upon n deficiency, provid-ing a key support for the hypothesis of membrane recycling into tag. While our study was in progress, li et al. (2012) reported that the activity of a galactoglycerolipid lipase (PgD1) was critical in tag accumulation under n starva-tion in C. reinhardtii (li et al. 2012), consistent with our findings in Chlorella.
the tlc lipid zones P3 and P5, which showed a signifi-cant decrease in radioactivity under −n conditions, were identified by eSI–MS/MS as the galactolipids MgDg and DgDg, which account for up to 80 % of membrane lipids in green plant tissues (guella et al. 2003). this result sug-gests that galactolipids may be a major contributing source of acyl groups for the synthesis of tag under −n. although gc–MS is routinely used to identify fatty acid composition of lipid compounds, the lc(+)eSI–MS/MS approach per-formed in this study has the advantage of generating high-resolution lipid profiles without the fatty acid methyl esteri-fication (FaMe) step, and still obtain information about the acyl chains. the fatty acid composition of the lipids was pos-sible by analyzing the cID products of the [M + na]+ ions (tables S1, S2), and the putative regiochemical distribution was deduced as previously described (guella et al. 2003). the
possible identification of odd-chain fatty acids c19:1, and c19:2 at the sn-1 position of galactolipids needs further study.
the significant increase in radioactivity (α = 0.05) in tag observed as early as 3 h of n starvation with concomi-tant decreased radioactivity in both MgDg and phospholip-ids observed with the pulse-chase experiment (Fig. 7) sug-gests that lipases and acyltransferases may be involved in the response to this nutrient stress at an early time point follow-ing n starvation. In fact, two recent transcriptomic studies in Chlamydomonas under −n detected upregulated genes encoding putative lipases that could play a role in releas-ing fatty acids from membrane lipids (Boyle et al. 2012; Miller et al. 2010). Moreover, a recent proteomics study in Chlamydomonas identified lipase-like oil body associated proteins, which may be localized at contact sites between lBs and chloroplasts, and thus, be involved in the degrada-tion of plastidial membranes (nguyen et al. 2011). In Chla-mydomonas, one of the enzymes involved in this process is phospholipid:diacylglycerol acyltransferase (PDat), which was recently shown to have both acyltransferase and lipase activities in a variety of substrates, such as phospholipids, galactolipids, and tag (Yoon et al. 2012). While PDat and PgD1 homologs would be expected to play a similar role in Chlorella, other enzymes which have not been fully charac-terized may also contribute to this phenomenon of membrane recycling of fatty acids into tag. In fact, the replacement of the membrane lipid phosphatidylcholine present in Chlorella by diacylglycerol-N, N, N-trimethylhomoserine (Fan et al. 2011) in Chlamydomonas, confirmed in this study, illustrates the evolutionary divergences between the two genera.
the present study shows that the recycling of structural membrane lipids significantly contributes to the increased flux of acyl moieties into tag starting as early as 3 h of n starvation in Chlorella. It also provides key ultrastruc-tural information that will serve as a basis for subsequent high-throughput proteomic and transcriptomic studies to investigate the initial and late metabolic changes, leading to the trigger of lB formation and tag accumulation in oleaginous green algae.
Acknowledgments We thank Dr. Byung-Ho Kang, Karen Kel-ley, and Kim Backer-Kelley (University of Florida, Interdisciplinary center for Biotechnology research, electron Microscopy and Bio-Imaging lab) for help with transmission electron microscopy. the HPlc–MS-MS analyses of lipid fractions were done at chemistry Department, University of Florida. eg thanks the Plant Molecular and cellular Biology program, the college of agriculture and life Sciences and the Horticultural Sciences Department, University of Florida for a graduate research assistantship.
References
Bertagnolli Bl, nadakavukaren MJ (1970) an ultrastructural study of pyrenoids from Chlorella pyrenoidosa. J cell Sci 7:623–630

Planta
1 3
Boyle nr, Page MD, liu B, Blaby IK, casero D, Kropat J, cokus SJ, Hong-Hermesdorf a, Shaw J, Karpowicz SJ, gallaher SD, Johnson S, Benning c, Pellegrini M, grossman a, Merchant SS (2012) three acyltransferases and nitrogen-responsive regula-tor are implicated in nitrogen starvation-induced triacylglycerol accumulation in Chlamydomonas. J Biol chem 287:15811–15825
chen YH, Walker tH (2011) Biomass and lipid production of hetero-trophic microalgae Chlorella protothecoides by using biodiesel-derived crude glycerol. Biotechnol lett 33:1973–1983
chen W, Zhang c, Song l, Sommerfeld M, Hu Q (2009) a high throughput nile red method for quantitative measurement of neu-tral lipids in microalgae. J Microbiol Methods 77:41–47
chisti Y (2007) Biodiesel from microalgae. Biotechnol adv 25:294–306
Fan J, andre c, Xu c (2011) a chloroplast pathway for the de novo biosynthesis of triacylglycerol in Chlamydomonas reinhardtii. FeBS lett 585:1985–1991
Foley PM, Beach eS, Zimmerman JB (2011) algae as a source of renewable chemicals: opportunities and challenges. green chem 13:1399–1405
garcía-Ferris c, de los ríos a, ascaso c, Moreno J (1996) corre-lated biochemical and ultrastructural changes in nitrogen-starved Euglena gracilis. J Phycol 32:953–963
gaude n, Brehelin c, tischendorf g, Kessler F, Dormann P (2007) nitrogen deficiency in Arabidopsis affects galactolipid composi-tion and gene expression and results in accumulation of fatty acid phytyl esters. Plant J 49:729–739
goodson c, roth r, Wang Zt, goodenough U (2011) Structural cor-relates of cytoplasmic and chloroplast lipid body synthesis in Chlamydomonas reinhardtii and stimulation of lipid body pro-duction with acetate boost. eukaryot cell 10:1592–1606
greenspan P, Mayer eP, Fowler SD (1985) nile red: a selective fluorescent stain for intracellular lipid droplets. J cell Biol 100:965–973
griffiths M, Harrison S (2009) lipid productivity as a key character-istic for choosing algal species for biodiesel production. J appl Phycol 21:493–507
guarnieri Mt, nag a, Smolinski Sl, Darzins a, Seibert M, Pienkos Pt (2011) examination of triacylglycerol biosynthetic pathways via de novo transcriptomic and proteomic analyses in an unse-quenced microalga. PloS One 6:e25851
guella g, Frassanito r, Mancini I (2003) a new solution for an old problem: the regiochemical distribution of the acyl chains in galactolipids can be established by electrospray ionization tandem mass spectrometry. rapid commun Mass Spectrom 17:1982–1994
guiry MD (2013) algaebase. World-wide electronic publication. national University of Ireland, galway
Heredia-arroyo t, Wei W, Hu B (2010) Oil accumulation via het-erotrophic/mixotrophic Chlorella protothecoides. appl Biochem Biotechnol 162:1978–1995
Hortensteiner S, chinner J, Matile P, thomas H, Donnison IS (2000) chlorophyll breakdown in Chlorella protothecoides: characteri-zation of degreening and cloning of degreening-related genes. Plant Mol Biol 42:439–450
Hu Q, Sommerfeld M, Jarvis e, ghirardi M, Posewitz M, Seibert M, Darzins a (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J 54:621–639
Huss Var, Frank c, Hartmann ec, Hirmer M, Kloboucek a, Seidel BM, Wenzeler P, Kessler e (1999) Biochemical taxonomy and molecular phylogeny of the genus Chlorella sensu lato (chloro-phyta). J Phycol 35:587–598
James gO, Hocart cH, Hillier W, chen H, Kordbacheh F, Price gD, Djordjevic Ma (2011) Fatty acid profiling of Chlamydomonas reinhardtii under nitrogen deprivation. Bioresour technol 102:3343–3351
Krienitz l, Hegewald eH, Hepperle D, Huss Var, rohr t, Wolf M (2004) Phylogenetic relationship of Chlorella and Parachlo-rella gen. nov. (chlorophyta, trebouxiophyceae). Phycologia 43:529–542
li X, Moellering er, liu B, Johnny c, Fedewa M, Sears BB, Kuo MH, Benning c (2012) a galactoglycerolipid lipase is required for triacylglycerol accumulation and survival following nitro-gen deprivation in Chlamydomonas reinhardtii. Plant cell 24:4670–4686
Miao X, Wu Q (2004) High yield bio-oil production from fast pyroly-sis by metabolic controlling of Chlorella protothecoides. J Bio-technol 110:85–93
Miller r, Wu g, Deshpande rr, Vieler a, gartner K, li X, Moeller-ing er, Zauner S, cornish aJ, liu B, Bullard B, Sears BB, Kuo MH, Hegg el, Shachar-Hill Y, Shiu SH, Benning c (2010) changes in transcript abundance in Chlamydomonas reinhardtii following nitrogen deprivation predict diversion of metabolism. Plant Physiol 154:1737–1752
Mujtaba g, choi W, lee cg, lee K (2012) lipid production by Chlo-rella vulgaris after a shift from nutrient-rich to nitrogen starva-tion conditions. Bioresour technol 123:279–283
Murphy DJ (2012) the dynamic roles of intracellular lipid droplets: from archaea to mammals. Protoplasma 249:541–585
nguyen HM, Baudet M, cuine S, adriano JM, Barthe D, Billon e, Bruley c, Beisson F, Peltier g, Ferro M, li-Beisson Y (2011) Proteomic profiling of oil bodies isolated from the unicellular green microalga Chlamydomonas reinhardtii: with focus on pro-teins involved in lipid metabolism. Proteomics 11:4266–4273
O’grady J, Morgan Ja (2011) Heterotrophic growth and lipid produc-tion of Chlorella protothecoides on glycerol. Bioprocess Biosyst eng 34:121–125
Orus M, Martinez F (1991) chlorophyll a/b ratio and thylakoid stack-ing modification in response to glucose in Chlorella vulgaris UaM 101. Biochem Physiol Pflanzen 187:197–202
Pyliotis na, goodchild DJ (1975) the regreening of nitrogen-defi-cient Chlorella fusca II. Structural changes during synchronous regreening. arch Microbiol 103:259–270
Saitou n, nei M (1987) the neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol evol 4:406–425
Sambrook J (2001) Molecular cloning: a laboratory manual. cold spring harbor laboratory Press, cold Spring Harbor
Sorokin c, Krauss rW (1958) the effects of light intensity on the growth rates of green algae. Plant Physiol 33:109–113
tamura K, Peterson D, Peterson n, Stecher g, nei M, Kumar S (2011) Mega5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsi-mony methods. Mol Biol evol 28:2731–2739
Waltermann M, Steinbuchel a (2005) neutral lipid bodies in prokary-otes: recent insights into structure, formation, and relationship to eukaryotic lipid depots. J Bacteriol 187:3607–3619
Wang Zt, Ullrich n, Joo S, Waffenschmidt S, goodenough U (2009) algal lipid bodies: stress induction, purification, and biochemi-cal characterization in wild-type and starchless Chlamydomonas reinhardtii. eukaryot cell 8:1856–1868
Wilkie ac, edmundson SJ, Duncan Jg (2011) Indigenous algae for local bioresource production: phycoprospecting. energy Sustain Dev 15:365–371
Xiong W, liu l, Wu c, Yang c, Wu Q (2010) 13c-tracer and gas chromatography-mass spectrometry analyses reveal metabolic flux distribution in the oleaginous microalga Chlorella protothe-coides. Plant Physiol 154:1001–1011
Xu H, Miao X, Wu Q (2006) High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J Biotechnol 126:499–507
Yeh Kl, chang JS (2011) nitrogen starvation strategies and photo-bioreactor design for enhancing lipid content and lipid production

Planta
1 3
of a newly isolated microalga Chlorella vulgaris eSP-31: impli-cations for biofuels. Biotechnol J 6:1358–1366
Yoon K, Han D, li Y, Sommerfeld M, Hu Q (2012) Phospholipid:diacylglycerol acyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation
while synthesizing triacylglycerol in the unicellular green micro-alga Chlamydomonas reinhardtii. Plant cell
Zuo Z, Zhu Y, Bai Y, Wang Y (2012) acetic acid-induced programmed cell death and release of volatile organic compounds in Chla-mydomonas reinhardtii. Plant Physiol Biochem 51:175–184