Embed Size (px)
DESCRIPTION
Chronic cigarette smoke exposure induces microbial and inflammatory shifts and mucin changes in the murine gut
Citation preview

Chronic cigarette smoke exposure induces microbialand inflammatory shifts and mucin changes in themurine gut
Liesbeth Allais,1* Frederiek-Maarten Kerckhof,2
Stephanie Verschuere,3 Ken R. Bracke,4
Rebecca De Smet,1 Debby Laukens,5
Pieter Van den Abbeele,2 Martine De Vos,5
Nico Boon,2 Guy G. Brusselle,4 Claude A. Cuvelier1
and Tom Van de Wiele2
1Department of Medical and Forensic Pathology,2Laboratory of Microbial Ecology and Technology(LabMET), Faculty of Bioscience Engineering, GhentUniversity, Ghent, Belgium.3Department of Pathology, AZ Sint-Jan, Brugge,Belgium.4Laboratory for Translational Research in ObstructivePulmonary Diseases, Department of RespiratoryMedicine, Ghent University Hospital, Ghent, Belgium.5Department of Gastroenterology, Ghent University,Ghent, Belgium.
Summary
Inflammatory bowel diseases (IBD) are complex mul-tifactorial diseases characterized by an inappropriatehost response to an altered commensal microbiomeand dysfunctional mucus barrier. Cigarette smoking isthe best known environmental risk factor in IBD. Here,we studied the influence of chronic smoke exposureon the gut microbiome, mucus layer composition andimmune factors in conventional mice. We comparedsmoke-exposed with air-exposed mice (n = 12) after asmoke exposure of 24 weeks. Both Illumina sequenc-ing (n = 6) and denaturing gradient gel electrophoresis(n = 12) showed that bacterial activity and communitystructure were significantly altered in the colon dueto smoke exposure. Interestingly, an increase ofLachnospiraceae sp. activity in the colon wasobserved. Also, the mRNA expression of Muc2 andMuc3 increased in the ileum, whereas Muc4 increasedin the distal colon of smoke-exposed mice (n = 6).Furthermore, we observed increased Cxcl2 anddecreased Ifn-γ in the ileum, and increased Il-6 and
decreased Tgf-β in the proximal colon. Tight junctiongene expression remained unchanged. We infer thatthe modulating role of chronic smoke exposure as alatently present risk factor in the gut may be driven bythe altered epithelial mucus profiles and changes inmicrobiome composition and immune factors.
Introduction
Inflammatory bowel diseases (IBD), comprising Crohn’sdisease (CD) and ulcerative colitis (UC), result from acomplex interplay between environmental factors, genet-ics and intestinal microbiota, which combine to initiate andperpetuate chronic inflammation in the gastrointestinaltract. CD is associated with a transmural and discontinu-ous inflammation that most frequently involves the ileumand colon, but can affect the entire gastrointestinal tract,while in UC, a superficial inflammation occurs which islimited to the colon (Khan and Collins, 2006; Jin et al.,2012). The prevalence of IBD ranges from 174 to 210 forCD and 79 to 122 for UC per 100 000 inhabitants inWestern countries and is still increasing (Loftus, 2004;Lakatos, 2006; Hovde and Moum, 2012). Disrupted intes-tinal homeostasis and abnormal immune responses tohost intestinal resident microbiota play a major role in thepathogenesis, although the exact mechanisms have notyet been elucidated (Strober et al., 2007; Xavier andPodolsky, 2007).
Disturbance of the microbial equilibrium, termeddysbiosis, is characterized by quantitative and qualitativechanges in the microbiota. This is marked by an increase ofcommon bacterial inhabitants of the gut that becomepathogenic under permissive conditions (Stecher et al.,2013). Dysbiosis provokes dysregulation of adaptiveimmune responses in the gut and is increasingly recog-nized as a contributing factor in the pathogenesis of IBD(Carbonnel et al., 2009; Round and Mazmanian, 2009).Previous studies showed that bacterial diversity isdecreased in stool samples of IBD patients (Manichanhet al., 2006; Erickson et al., 2012). Alterations in theabundance of species of all prominent intestinalphyla Firmicutes, Bacteroidetes, Verrucomicrobia, Actino-bacteria and Proteobacteria are associated with the devel-opment of IBD (Frank et al., 2007; Png et al., 2010b).
Received 9 February, 2015; accepted 28 May, 2015. *For corre-spondence. E-mail [email protected] or [email protected]; Tel. +32 9 332 49 53; Fax +32-93324965.
bs_bs_banner
Environmental Microbiology (2015) doi:10.1111/1462-2920.12934
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd

Cigarette smoke (CS) is the most prominent environ-mental risk factor for developing CD; however, itexerts a protective role in UC (Persson et al., 1990;Ananthakrishnan, 2013). In addition, smoke exposuremay have an important impact on the composition anddynamics of the gut microbiome. For instance, theBifidobacterium population increased in the caecum ofrats after 4 weeks of CS exposure (Tomoda et al., 2011).In mouse, side-stream smoking increased Clostridiumsp., but decreased the Firmicutes phylum (Lactococcusand Ruminococcus sp.), the Enterobacteriaceae familyand the segmented filamentous bacteria in the caecum(Wang et al., 2012). In human studies, smoking hasbeen associated with a higher rate of Clostridium difficileinfection, in which current smokers (actively smokingduring the study) have the highest risk compared withformer (quitted smoking before the start of the study)and never smokers (never smoked in their whole life)(Rogers et al., 2012). Moreover, CS is a known riskdeterminant for both bacterial and viral infectionsthrough alterations in cell- and humoral-mediatedimmune responses in the respiratory tract (Arcavi andBenowitz, 2004). We recently demonstrated that CS trig-gers the gut immune system through the recruitment ofimmune cells to the Peyer’s patches (PP), the main lym-phoid organs in the gut, and through the inductionof autophagy and apoptosis in the follicle-associatedepithelium (FAE) overlying the PP (Verschuere et al.,2011; 2012).
Many studies have investigated the impact of specificintestinal bacteria on host gene and protein expression inthe intestine and their role in IBD development (Rolli et al.,2010). Nevertheless, the role of smoking in the emergenceof dysbiosis, which might modulate the risk for thedevelopment of IBD, still needs further investigation. CSalters host–microorganism interaction dynamics in theairways, causing respiratory tract infections and contribut-ing to chronic obstructive pulmonary disease (COPD)(Garmendia et al., 2012). This study addresses the effectof chronic CS exposure on the gut mucosa and its micro-bial environment. We examined the diversity of the bacte-rial community in the ileum and colon of mice exposed toCS or air for 24 weeks. Additionally, we explored whichspecies were sensitive to CS exposure and to what extentthe composition of the mucus layer and inflammatory geneexpression is affected.
Results
The gut bacterial community shifts in response tocigarette smoke
To determine the response of the host microbiome to CS,the taxonomical community structure of the microbiome inileal, caecal and distal colonic samples of 12 smoke-
exposed and 12 air-exposed mice was analysed by dena-turing gradient gel electrophoresis (DGGE) as a profilingtechnique. Dendrograms applying the abundance-basedJaccard index (Fig. 1) and Yue and Clayton’s theta index(Fig. S1) were used to visualize the clustering of thesamples. Considering the distinct intestinal regions, thedendrograms showed clear clusters for air-exposed mice(49 ± 11% and 58 ± 9% within-group similarity, respec-tively), separated from smoke-exposed mice in caecumand distal colon, suggesting a bacterial shift (P = 0.001 forboth using multivariate analysis of variance (MANOVA)).This kind of separated clusters was not observed in thedendrogram of the ileal samples, with the air-exposedsamples only showing 42 ± 11% within-group similarity.
The activity of specific bacterial species is influenced bysmoke exposure
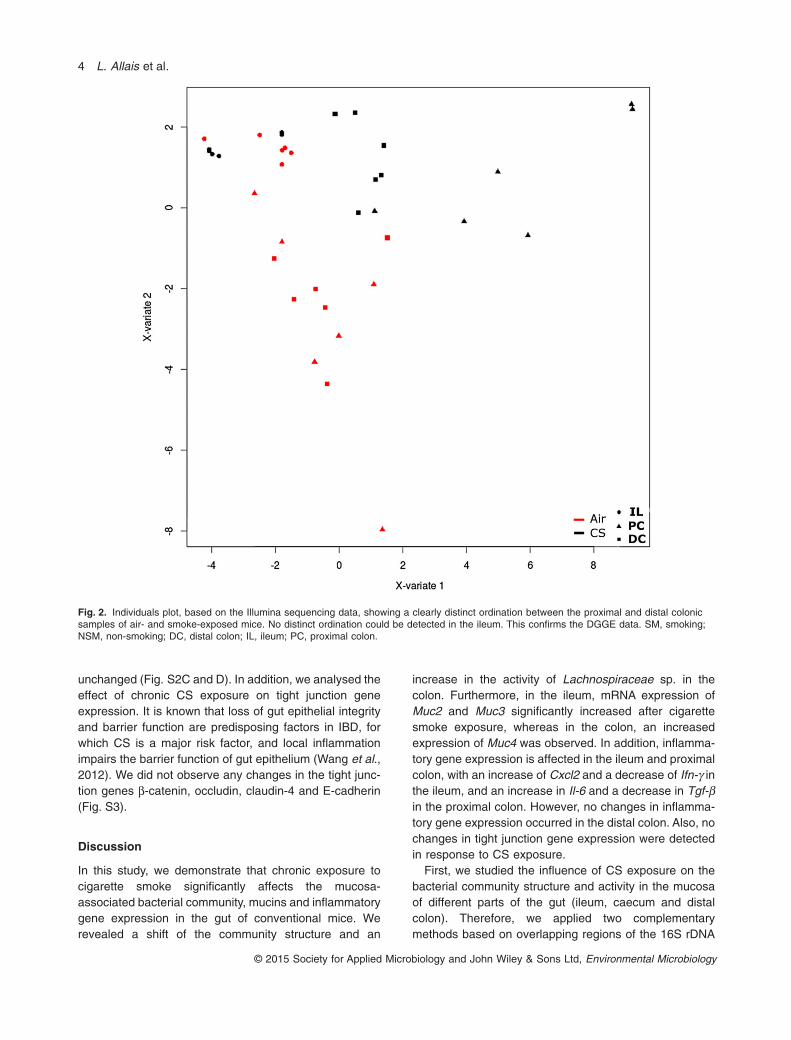
A shift in the microbial composition may potentiallyimply a shift in microbial activity. Using cDNA samples,we evaluated the activity of the microbiome with Illuminasequencing as a more high-resolution method. We ran-domly selected 12 mice (six smoke- and six air-exposedmice, originating from four distinct cages) of whichileal, caecal and distal colonic samples were included inthe analysis. Changes in 16S rRNA levels were evalu-ated by the sparse partial least square discriminantanalysis (sPLS-DA). This clearly showed a distinct ordi-nation for proximal and distal colon between the smoke-and air-exposed murine microbiome activity (Fig. 2),but not for ileum, which parallels the shift in microbialcomposition as shown by the DGGE-based clustering(Fig. 1).
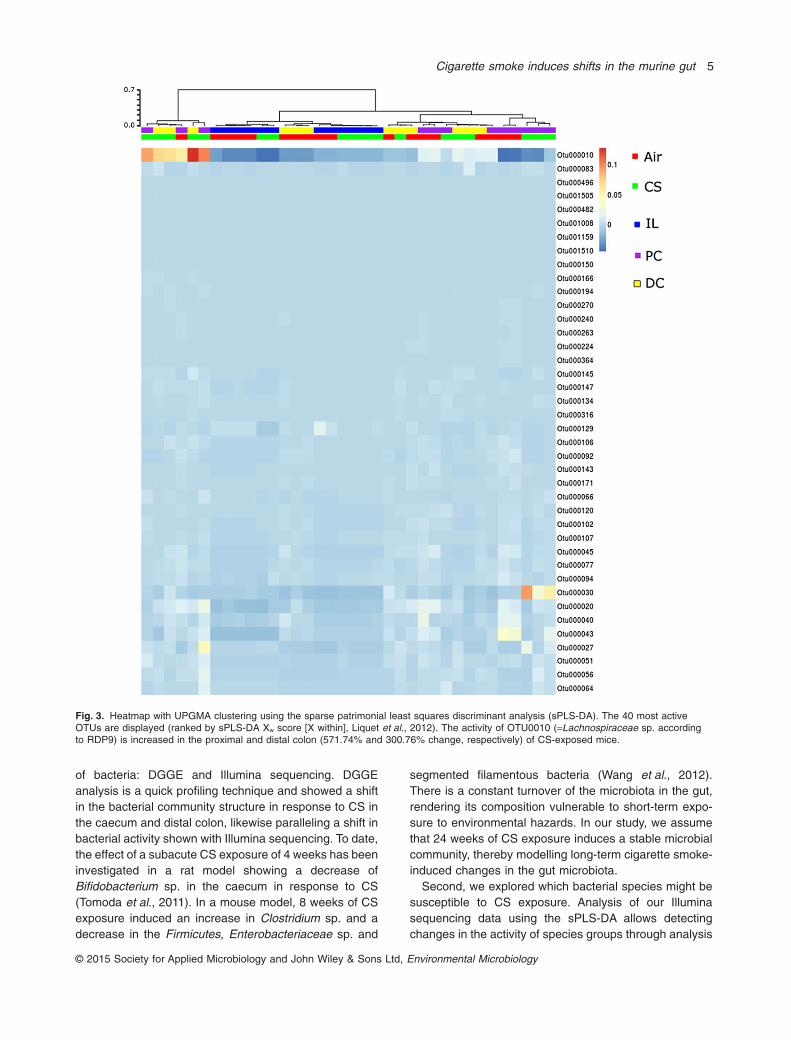
To further investigate the effect of smoke exposure onbacterial activity in the murine gut, we used Illuminasequencing to identify the operating taxonomic units(OTUs) showing changes in activity in the different gutregions of six smoke- and six air-exposed mice. Weapplied the sPLS-DA method to identify the specific OTUsin each separate intestinal region. All relevant OTUs wereclustered using the unweighted pair group method witharithmetic mean (UPGMA) algorithm and displayed in aheatmap (Fig. 3), which demonstrated that the expressed16S rRNA level and therefore the activity of OTU010,representing Lachnospiraceae sp., strongly increases inproximal and distal colon (571.74% and 300.76% change,respectively) in response to smoke exposure.
Cigarette smoke exposure affects the specific mucinexpression pattern, but not the main mucin classes inileum and distal colon
The detected bacterial changes prompted us to investigatewhether the mucin composition of the mucus layer is
2 L. Allais et al.
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology

altered upon CS exposure. Mucins in the gut can besubdivided in two main classes: secretable mucins and cellsurface mucins (Hoebler et al., 2006). We examined theamount of goblet cells, which secrete mucins into the gutlumen, inAlcian Blue (AB)/periodic acid Schiff (PAS)-staineileum and distal colon sections of 10 smoke-exposed of 10air-exposed mice. For each group, ileum and distal colonsections were scored. No significant differences weredetected. Additionally, tissue sections of the same gutsamples were stained with high iron diamine (HID) and ABto differentiate between sulfo- and sialomucins. Again, nosignificant differences were detected.
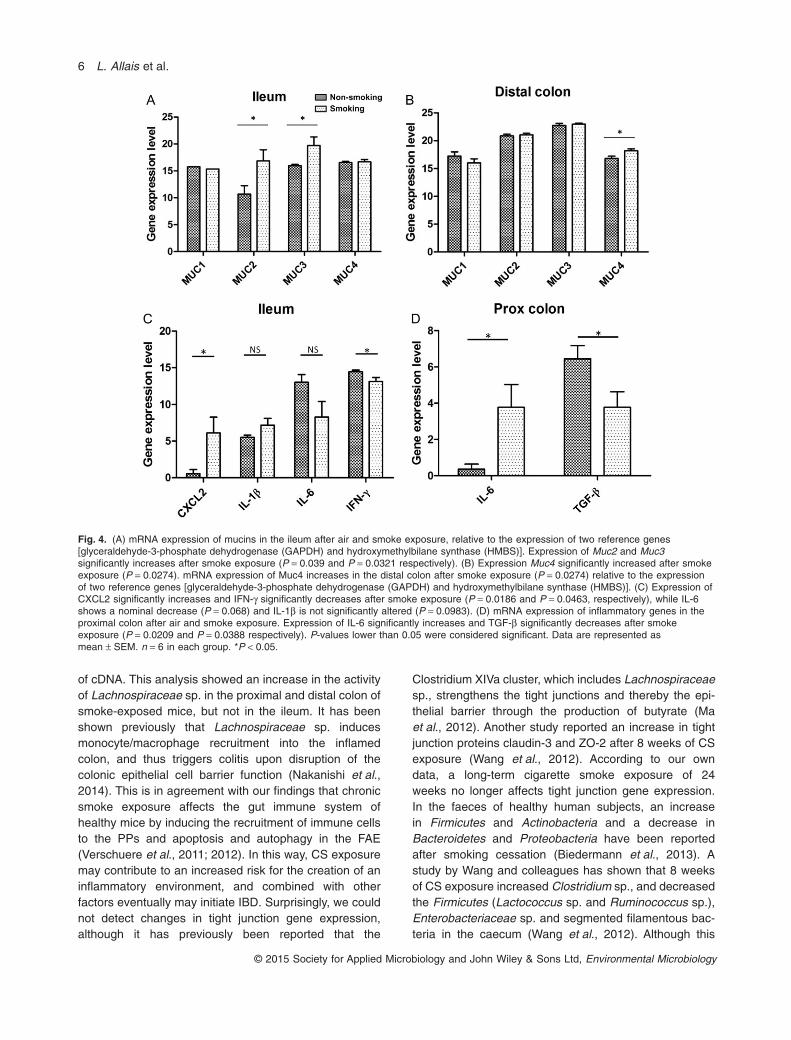
Furthermore, we examined the mRNA expressionpattern of MUC1, MUC2, MUC3 and MUC4. MUC2 isstored in goblet cells and typically secreted, whereasMUC1, MUC3 and MUC4 are membrane-bound cellsurface mucins, which are anchored in the glycocalyx,located at the apical surface of the enterocyte (Hoebleret al., 2006). Expression analysis in both the smoke-and air-exposed group (n = 6 in each group) showed ashift in the mucin expression pattern in response to CSin ileum and distal colon (Fig. 4A and B). In the ileum,Muc2 and Muc3 expression was upregulated inresponse to CS, while Muc1 and Muc4 expression wasnot significantly altered. Muc2 and Muc3 expressionin air-exposed ileum was much lower than in air-exposed distal colon, which is not surprising as bacterialload and therefore bacteria-induced mucus in the ileumis much lower (Johansson, 2014). Interestingly, beinginduced by CS exposure, Muc2 and Muc3 expression inthe ileum resembles a more colon-like expression sig-nature. In the distal colon, only Muc4 expression isincreased in smoke-exposed compared with air-exposedanimals.
Cigarette smoke exposure affects inflammatory geneexpression, but not the tight junction gene expression
The gut microbiota play an important role in the immuno-logical response of the gut (Round et al., 2010). Interest-ingly, we have previously shown that 24 weeks of CSexposure affects the ileal immunological response by trig-gering the attraction of immune cells to the PP, theimmune inductive sites of the gut (Verschuere et al.,2011). Therefore, we investigated the effect of CS expo-sure on inflammatory gene expression in the immuneeffector regions of the gut (n = 6 in each group). Weobserved a significant increase of Cxcl2, a decrease ofIfn-γ and a nominal decrease of Il-6 in the ileum (Fig. 4C),whereas Tnf-α, Il-10, Nfκb and Tgf-β remained unaltered(Fig. S2A). In the proximal colon, Il-6 was increased andTgf-β was decreased (Fig. 4D), while Il-10, Nfκb, Ifn-γ,Cxcl2 and Il-1β did not change (Fig. S2B). However, in thedistal colon, inflammatory gene expression remained
Fig. 1. Agglomerative nesting clustering dendrograms using theabundance-based Jaccard distance measure based on the DGGEdata with average linkage (UPGMA), normalized in BIONUMERICS
5.10 software. (A) Clustering dendrogram for ileum, (B) clusteringdendrogram for caecum and (C) clustering dendrogram for distalcolon. SM, smoking; NSM, non-smoking; DC, distal colon; IL,ileum; CC, caecum.
Cigarette smoke induces shifts in the murine gut 3
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology
unchanged (Fig. S2C and D). In addition, we analysed theeffect of chronic CS exposure on tight junction geneexpression. It is known that loss of gut epithelial integrityand barrier function are predisposing factors in IBD, forwhich CS is a major risk factor, and local inflammationimpairs the barrier function of gut epithelium (Wang et al.,2012). We did not observe any changes in the tight junc-tion genes β-catenin, occludin, claudin-4 and E-cadherin(Fig. S3).
Discussion
In this study, we demonstrate that chronic exposure tocigarette smoke significantly affects the mucosa-associated bacterial community, mucins and inflammatorygene expression in the gut of conventional mice. Werevealed a shift of the community structure and an
increase in the activity of Lachnospiraceae sp. in thecolon. Furthermore, in the ileum, mRNA expression ofMuc2 and Muc3 significantly increased after cigarettesmoke exposure, whereas in the colon, an increasedexpression of Muc4 was observed. In addition, inflamma-tory gene expression is affected in the ileum and proximalcolon, with an increase of Cxcl2 and a decrease of Ifn-γ inthe ileum, and an increase in Il-6 and a decrease in Tgf-βin the proximal colon. However, no changes in inflamma-tory gene expression occurred in the distal colon. Also, nochanges in tight junction gene expression were detectedin response to CS exposure.
First, we studied the influence of CS exposure on thebacterial community structure and activity in the mucosaof different parts of the gut (ileum, caecum and distalcolon). Therefore, we applied two complementarymethods based on overlapping regions of the 16S rDNA
Fig. 2. Individuals plot, based on the Illumina sequencing data, showing a clearly distinct ordination between the proximal and distal colonicsamples of air- and smoke-exposed mice. No distinct ordination could be detected in the ileum. This confirms the DGGE data. SM, smoking;NSM, non-smoking; DC, distal colon; IL, ileum; PC, proximal colon.
4 L. Allais et al.
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology
of bacteria: DGGE and Illumina sequencing. DGGEanalysis is a quick profiling technique and showed a shiftin the bacterial community structure in response to CS inthe caecum and distal colon, likewise paralleling a shift inbacterial activity shown with Illumina sequencing. To date,the effect of a subacute CS exposure of 4 weeks has beeninvestigated in a rat model showing a decrease ofBifidobacterium sp. in the caecum in response to CS(Tomoda et al., 2011). In a mouse model, 8 weeks of CSexposure induced an increase in Clostridium sp. and adecrease in the Firmicutes, Enterobacteriaceae sp. and
segmented filamentous bacteria (Wang et al., 2012).There is a constant turnover of the microbiota in the gut,rendering its composition vulnerable to short-term expo-sure to environmental hazards. In our study, we assumethat 24 weeks of CS exposure induces a stable microbialcommunity, thereby modelling long-term cigarette smoke-induced changes in the gut microbiota.
Second, we explored which bacterial species might besusceptible to CS exposure. Analysis of our Illuminasequencing data using the sPLS-DA allows detectingchanges in the activity of species groups through analysis
Fig. 3. Heatmap with UPGMA clustering using the sparse patrimonial least squares discriminant analysis (sPLS-DA). The 40 most activeOTUs are displayed (ranked by sPLS-DA Xw score [X within], Liquet et al., 2012). The activity of OTU0010 (=Lachnospiraceae sp. accordingto RDP9) is increased in the proximal and distal colon (571.74% and 300.76% change, respectively) of CS-exposed mice.
Cigarette smoke induces shifts in the murine gut 5
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology
of cDNA. This analysis showed an increase in the activityof Lachnospiraceae sp. in the proximal and distal colon ofsmoke-exposed mice, but not in the ileum. It has beenshown previously that Lachnospiraceae sp. inducesmonocyte/macrophage recruitment into the inflamedcolon, and thus triggers colitis upon disruption of thecolonic epithelial cell barrier function (Nakanishi et al.,2014). This is in agreement with our findings that chronicsmoke exposure affects the gut immune system ofhealthy mice by inducing the recruitment of immune cellsto the PPs and apoptosis and autophagy in the FAE(Verschuere et al., 2011; 2012). In this way, CS exposuremay contribute to an increased risk for the creation of aninflammatory environment, and combined with otherfactors eventually may initiate IBD. Surprisingly, we couldnot detect changes in tight junction gene expression,although it has previously been reported that the
Clostridium XIVa cluster, which includes Lachnospiraceaesp., strengthens the tight junctions and thereby the epi-thelial barrier through the production of butyrate (Maet al., 2012). Another study reported an increase in tightjunction proteins claudin-3 and ZO-2 after 8 weeks of CSexposure (Wang et al., 2012). According to our owndata, a long-term cigarette smoke exposure of 24weeks no longer affects tight junction gene expression.In the faeces of healthy human subjects, an increasein Firmicutes and Actinobacteria and a decrease inBacteroidetes and Proteobacteria have been reportedafter smoking cessation (Biedermann et al., 2013). Astudy by Wang and colleagues has shown that 8 weeksof CS exposure increased Clostridium sp., and decreasedthe Firmicutes (Lactococcus sp. and Ruminococcus sp.),Enterobacteriaceae sp. and segmented filamentous bac-teria in the caecum (Wang et al., 2012). Although this
Fig. 4. (A) mRNA expression of mucins in the ileum after air and smoke exposure, relative to the expression of two reference genes[glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and hydroxymethylbilane synthase (HMBS)]. Expression of Muc2 and Muc3significantly increases after smoke exposure (P = 0.039 and P = 0.0321 respectively). (B) Expression Muc4 significantly increased after smokeexposure (P = 0.0274). mRNA expression of Muc4 increases in the distal colon after smoke exposure (P = 0.0274) relative to the expressionof two reference genes [glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and hydroxymethylbilane synthase (HMBS)]. (C) Expression ofCXCL2 significantly increases and IFN-γ significantly decreases after smoke exposure (P = 0.0186 and P = 0.0463, respectively), while IL-6shows a nominal decrease (P = 0.068) and IL-1β is not significantly altered (P = 0.0983). (D) mRNA expression of inflammatory genes in theproximal colon after air and smoke exposure. Expression of IL-6 significantly increases and TGF-β significantly decreases after smokeexposure (P = 0.0209 and P = 0.0388 respectively). P-values lower than 0.05 were considered significant. Data are represented asmean ± SEM. n = 6 in each group. *P < 0.05.
6 L. Allais et al.
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology

study suggests that Firmicutes, to which theLachnospiraceae sp. belongs, decreases due to smoking,many other species groups determine changes in thephylum besides the Lachnospiraceae sp. Also, weexposed the mice to 24 weeks of CS, which is a long-termexposure and sheds light on long-term changes in themicrobiota. In future experiments, it might be of interest tomonitor microbiota changes over time in response to CSexposure.
A third important finding in this study was that smokingis able to induce changes in mucin expression. Previousstudies have shown that smoking causes mucinhypersecretion in the lung of patients suffering fromCOPD (Di et al., 2012; Kim, 2012; Yu et al., 2012). Themajor mucins in the intestine are MUC2, a secretablemucin mainly produced and delivered to the lumen bygoblet cells, and MUC3, a cell surface mucin (Shiraziet al., 2000; Hoebler et al., 2006; Linden et al., 2008).MUC4 is a cell surface mucin that acts purely as ananti-adhesive (Hattrup and Gendler, 2008). Here, weshowed that ileal expression of Muc2 and Muc3 increasedsignificantly upon CS exposure. In the distal colon, CScaused an increase of Muc4 mRNA. Interestingly, quan-titative changes in mucin secretion also occur in humanIBD (Boltin et al., 2013). The composition of the protectivemucin layer plays an important role in inhibiting directcontact between the host and potentially offending bacte-ria, and reducing the exposure time by increasing bacte-rial transit. The changes in mucin expression that weobserved might either be a direct effect of the exposure tothe chemical components of CS or induced as a protec-tion mechanism to counteract local physiological changesin the gut. In addition, a shifting composition of mucolyticorganisms from A. muciniphila to other mucolytic species,such as Ruminococcus gnavus, occurs in pathologicalconditions, such as IBD, and may therefore be a suitablebiomarker for mucosal integrity (Png et al., 2010a; Berryand Reinisch, 2013). However, to date, the mucin degra-dation specificity of different Akkermansia speciesremains unclear.
In addition, we found that inflammatory gene expres-sion is altered in the ileum and proximal colon in responseto CS exposure, but not in the distal colon. We demon-strated that ileal Cxcl2, a critical effector for neutrophiltrafficking, is increased by chronic CS exposure. In con-trast, ileal Ifn-ɣ, involved in macrophage activation, isdecreased. Furthermore, we showed that proximalcolonic Il-6 was increased. Proximal colonic Tgf-β, beinginvolved in tempering the immune response, wasdecreased. Surprisingly, expression of inflammatoryfactors was not affected in the distal colon, althoughsmoke-induced changes in the microbiota occur in thecolon. The ileum contains important immune inductivesites, such as the PPs, taking up antigens and generating
immune responses. This kind of immune inductive sitesare much less frequent in the colon, which makes it mainlyan effector site (lamina propria and surface epithelia) ofileal-generated immune responses (Brandtzaeg andPabst, 2004). Wang et al. showed an inhibition of theNFκB pathway, but no changes in the expression ofcolonic Tnf-α and Il-6 after 8 weeks of CS exposure(Wang et al., 2012). In our study, we did not observe anychanges in Nfκb expression; however, we showed thatcolonic Il-6 was increased. Again, this might be due to thelong-term (24 weeks) effect of CS exposure in our ownexperimental set-up.
Our data show that chronic CS exposure has a pro-found effect on the gut microbiota, the associated mucincomposition and cytokine/chemokine production in mice,which is likely to occur in human as well. Although the gutmicrobial similarity between mice and humans at thephylum level is remarkable, many differences exist at thespecies level (Dethlefsen et al., 2007). Both in human andC57BL6/J mice, the two most abundant phyla are theFirmicutes and the Bacteroidetes; however, 85% of themurine sequences represent species that have not beendetected in humans (Ley et al., 2005). Therefore, extrapo-lation from mouse to man might be challenging due to theheterogeneity of the human population. Nevertheless, theuse of a murine model offers the advantage of housingin standardized conditions, minimizing environmentalinfluences.
Interestingly, our findings show that chronic CS expo-sure affects the immune system in the ileum while it exertsa more pronounced effect on the microbiota in the colon.This may be partly explained by the much lower bacterialload in the ileum compared with the colon. It is known thatsmoking worsens CD, while UC is rather a disease ofex-smokers (Ananthakrishnan, 2015). Inflammation in CDmainly originates in the terminal ileum, whereas UC isinitiated in the colon. Taking all these facts together, itmight be that CS affects the ileum and colon viadistinct mechanisms, resulting in an even more distinctoutcome for inflammatory diseases as CD and UC. Givenour data, we speculate that cigarette smoke exposuretends to affect the ileal immune system, which may giverise to CD-related inflammation. In contrast, cigarettesmoke exposure targets the colonic microbiome ratherthan its immune system. Moreover, the activity ofLachnospiraceae sp., a butyrate producer, is increased inresponse to CS exposure, which may contribute to thesmoke-induced protective effect on UC-related colonicinflammation.
Today, the link between host (patho)physiology andthe gut microbiome is being increasingly recognized(Arumugam et al., 2011; Elinav et al., 2011). In-depthknowledge of the human bacterial ecosystem in relation todisease might pave the way for the generation of
Cigarette smoke induces shifts in the murine gut 7
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology

customized therapeutics and probiotics. We hypothesizethat the induction of dysbiosis and the alteration of epi-thelial mucus and cytokine/chemokine profiles by chroniccigarette smoke exposure modulate the risk for the devel-opment of inflammation-related disorders, such as IBD.Further work in this field may contribute to the adaptationof treatment strategies for IBD depending on smokingbehaviour. Future research exposing colitic mice tochronic smoke exposure will be necessary to be able toattribute a specific role of dysbiosis and mucus changes inthe development of disease.
Experimental procedures
Animals
Male C57BL/6 wild-type mice were purchased from CharlesRiver Laboratories and were co-housed in the same animalfacility before the start of the experiment to homogenize gutmicrobiota between groups. All mice were 8–9 weeks old atthe start of the smoke exposure. Mice were housed in atemperature-controlled room with a 12 h light and 12 h dark-ness cycle and were given standardized food (Carfil,Turnhout, Belgium) and water ad libitum. The mice weredivided in two cages per experimental group (n = 12) with sixmice per cage. The local ethics committee for animal experi-mentation of the Faculty of Medicine and Health Sciences(Ghent, Belgium) approved all experiments (ECD 27/07).
Cigarette smoke exposure
Mice were exposed to mainstream cigarette smoke, asdescribed previously (D’Hulst et al., 2005). Briefly, groups of12 mice were exposed to the tobacco smoke of 20 cigarettes(Reference Cigarette 3R4F without filter; University of Ken-tucky, Lexington, KY, USA). The exposure to five cigaretteswas applied for 7 min, with 30 min smoke-free intervals,which was repeated four times a day, 5 days per week for 24weeks (chronic smoke exposure). An optimal smoke : airratio of 1:6 was obtained. The control groups were exposed toair. Carboxyhaemoglobin in serum of smoke-exposed micereached a non-toxic level of 8.7 ± 0.31% (compared with0.65 ± 0.25% in air-exposed mice), which is similar tocarboxyhaemoglobin blood concentrations of humansmokers (Macdonald et al., 2004).
Staining methods
Paraffin-embedded tissue sections of 4 μm taken fromileum and distal colon were de-waxed and re-hydrated. Thesections were stained with either AB/PAS, in which ABstains blue for secretable mucins and PAS stains purple-pink for cell surface mucins, or with HID/AB, in whichsulfated mucins are stained dark brown by HID andsialylated mucins are stained blue by AB. In case of theAB/PAS staining method, slides were stained using the PASStaining Kit and AB for PAS Staining Kit on the automatedVentana system (Roche Diagnostics, Vilvoorde, Belgium).Slides were immersed in the AB solution for 5 min before
incubation in 1% aqueous periodic acid for 5 min. Slideswere then washed in distilled water and immersed inSchiff’s reagent for 15 min. In case of the HID/ABstaining method, slides were treated with diamine solu-tion (N,N-dimethyl-meta-phenylenediamine-dihydrochloride;N,N-dimethyl-para-phenylene-diamine-dihydrochloride; ferricchloride 60% solution and distilled water) for 24 h at roomtemperature. After incubation, slides were washed andcounterstained in 1% AB solution for 5 min. Finally, sectionsfrom both staining methods were dehydrated in absolutealcohol, cleared in three changes of xylene and mounted.
Scoring of microscopic analysis
Histological and goblet cell assessment was performed usinglight microscopy. For 10 AB/PAS-stained sections per group,positively stained goblet cells were counted in two regions of10 aligning longitudinal crypts. For 10 HID/AB-stained sec-tions per group, sulfomucin-positive stained goblet cells werecounted in two regions of 10 aligning longitudinal crypts.Histological assessments were carried out in a blindedfashion.
Quantitative real-time polymerase chain reaction(qRT-PCR)
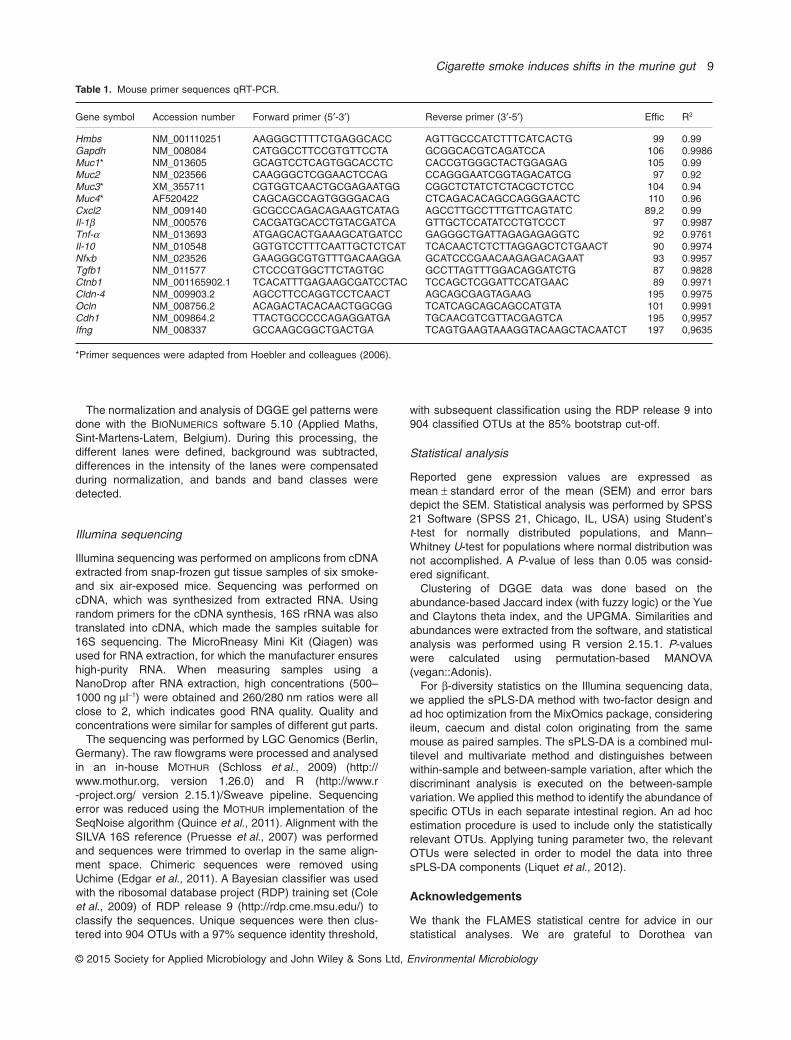
RNA from ileum and distal colon (both were taken from 10smoke- and 10 air-exposed mice) was extracted using theQiagen miRNeasy Mini Kit (Qiagen, Hilden, Germany). Sub-sequently, cDNA was synthesized by reverse transcriptionusing the iScript™ cDNA synthesis kit (Bio-Rad Laboratories,Nazareth, Belgium) following the manufacturer’s instructions.Expression of target genes Muc1, Muc2, Muc3, Muc4, Cxcl2,I1-1β, I1-6, Ifn-γ, Tnf-α, Il-10, Nfκb, Tgfb1, Ctnb1, Cldb-4,Ocln and Cdh1, and reference genes glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and hydroxymethyl-bilane synthase (HMBS) (Table 1), was analysed byqRT-PCR using the SensiMix™ SYBR No-ROX Kit (Bioline,London, UK). The qRT-PCR was performed on aLightCycler480 detection system (Roche Diagnostics) withthe following cycling conditions: 10 min incubation at 95°C,45 cycles of 95°C for 10 s and 60°C for 1 min. Melting curveanalysis confirmed primer specificity. The PCR efficiency ofeach primer pair was calculated using a standard curve fromreference cDNA. The amplification efficiency was determinedusing the formula 10−1/SLOPE − 1.
DGGE
The ileal and colonic samples of 12 smoke- and 12 air-exposed mice were obtained snap-frozen and stored at−80°C. The 16S rRNA genes for all bacteria were amplified byPCR adding a GC-clamp of 40 bp. The DGGE was performedusing the INGENYPHORU System (Ingeny International BV,The Netherlands), after which PCR fragments were loadedonto 8% (w/v) polyacrylamide gels containing 40–60% dena-turing gradients (Muyzer et al., 1993; Ovreas et al., 1997). Toprocess and compare the different gels, an in-house devel-oped marker of different PCR fragments was loaded on eachgel (Boon et al., 2002).
8 L. Allais et al.
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology
The normalization and analysis of DGGE gel patterns weredone with the BIONUMERICS software 5.10 (Applied Maths,Sint-Martens-Latem, Belgium). During this processing, thedifferent lanes were defined, background was subtracted,differences in the intensity of the lanes were compensatedduring normalization, and bands and band classes weredetected.
Illumina sequencing
Illumina sequencing was performed on amplicons from cDNAextracted from snap-frozen gut tissue samples of six smoke-and six air-exposed mice. Sequencing was performed oncDNA, which was synthesized from extracted RNA. Usingrandom primers for the cDNA synthesis, 16S rRNA was alsotranslated into cDNA, which made the samples suitable for16S sequencing. The MicroRneasy Mini Kit (Qiagen) wasused for RNA extraction, for which the manufacturer ensureshigh-purity RNA. When measuring samples using aNanoDrop after RNA extraction, high concentrations (500–1000 ng μl−1) were obtained and 260/280 nm ratios were allclose to 2, which indicates good RNA quality. Quality andconcentrations were similar for samples of different gut parts.
The sequencing was performed by LGC Genomics (Berlin,Germany). The raw flowgrams were processed and analysedin an in-house MOTHUR (Schloss et al., 2009) (http://www.mothur.org, version 1.26.0) and R (http://www.r-project.org/ version 2.15.1)/Sweave pipeline. Sequencingerror was reduced using the MOTHUR implementation of theSeqNoise algorithm (Quince et al., 2011). Alignment with theSILVA 16S reference (Pruesse et al., 2007) was performedand sequences were trimmed to overlap in the same align-ment space. Chimeric sequences were removed usingUchime (Edgar et al., 2011). A Bayesian classifier was usedwith the ribosomal database project (RDP) training set (Coleet al., 2009) of RDP release 9 (http://rdp.cme.msu.edu/) toclassify the sequences. Unique sequences were then clus-tered into 904 OTUs with a 97% sequence identity threshold,
with subsequent classification using the RDP release 9 into904 classified OTUs at the 85% bootstrap cut-off.
Statistical analysis
Reported gene expression values are expressed asmean ± standard error of the mean (SEM) and error barsdepict the SEM. Statistical analysis was performed by SPSS21 Software (SPSS 21, Chicago, IL, USA) using Student’st-test for normally distributed populations, and Mann–Whitney U-test for populations where normal distribution wasnot accomplished. A P-value of less than 0.05 was consid-ered significant.
Clustering of DGGE data was done based on theabundance-based Jaccard index (with fuzzy logic) or the Yueand Claytons theta index, and the UPGMA. Similarities andabundances were extracted from the software, and statisticalanalysis was performed using R version 2.15.1. P-valueswere calculated using permutation-based MANOVA(vegan::Adonis).
For β-diversity statistics on the Illumina sequencing data,we applied the sPLS-DA method with two-factor design andad hoc optimization from the MixOmics package, consideringileum, caecum and distal colon originating from the samemouse as paired samples. The sPLS-DA is a combined mul-tilevel and multivariate method and distinguishes betweenwithin-sample and between-sample variation, after which thediscriminant analysis is executed on the between-samplevariation. We applied this method to identify the abundance ofspecific OTUs in each separate intestinal region. An ad hocestimation procedure is used to include only the statisticallyrelevant OTUs. Applying tuning parameter two, the relevantOTUs were selected in order to model the data into threesPLS-DA components (Liquet et al., 2012).
Acknowledgements
We thank the FLAMES statistical centre for advice in ourstatistical analyses. We are grateful to Dorothea van
Table 1. Mouse primer sequences qRT-PCR.
Gene symbol Accession number Forward primer (5′-3′) Reverse primer (3′-5′) Effic R2
Hmbs NM_001110251 AAGGGCTTTTCTGAGGCACC AGTTGCCCATCTTTCATCACTG 99 0.99Gapdh NM_008084 CATGGCCTTCCGTGTTCCTA GCGGCACGTCAGATCCA 106 0.9986Muc1* NM_013605 GCAGTCCTCAGTGGCACCTC CACCGTGGGCTACTGGAGAG 105 0.99Muc2 NM_023566 CAAGGGCTCGGAACTCCAG CCAGGGAATCGGTAGACATCG 97 0.92Muc3* XM_355711 CGTGGTCAACTGCGAGAATGG CGGCTCTATCTCTACGCTCTCC 104 0.94Muc4* AF520422 CAGCAGCCAGTGGGGACAG CTCAGACACAGCCAGGGAACTC 110 0.96Cxcl2 NM_009140 GCGCCCAGACAGAAGTCATAG AGCCTTGCCTTTGTTCAGTATC 89,2 0.99Il-1β NM_000576 CACGATGCACCTGTACGATCA GTTGCTCCATATCCTGTCCCT 97 0.9987Tnf-α NM_013693 ATGAGCACTGAAAGCATGATCC GAGGGCTGATTAGAGAGAGGTC 92 0.9761Il-10 NM_010548 GGTGTCCTTTCAATTGCTCTCAT TCACAACTCTCTTAGGAGCTCTGAACT 90 0.9974Nfκb NM_023526 GAAGGGCGTGTTTGACAAGGA GCATCCCGAACAAGAGACAGAAT 93 0.9957Tgfb1 NM_011577 CTCCCGTGGCTTCTAGTGC GCCTTAGTTTGGACAGGATCTG 87 0.9828Ctnb1 NM_001165902.1 TCACATTTGAGAAGCGATCCTAC TCCAGCTCGGATTCCATGAAC 89 0.9971Cldn-4 NM_009903.2 AGCCTTCCAGGTCCTCAACT AGCAGCGAGTAGAAG 195 0.9975Ocln NM_008756.2 ACAGACTACACAACTGGCGG TCATCAGCAGCAGCCATGTA 101 0.9991Cdh1 NM_009864.2 TTACTGCCCCCAGAGGATGA TGCAACGTCGTTACGAGTCA 195 0,9957Ifng NM_008337 GCCAAGCGGCTGACTGA TCAGTGAAGTAAAGGTACAAGCTACAATCT 197 0,9635
*Primer sequences were adapted from Hoebler and colleagues (2006).
Cigarette smoke induces shifts in the murine gut 9
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology

Limbergen, Ran Rumes and Lynn Supply for the support withthe animal experiments and the processing of the samples,and Eliane Castrique, Christelle Snauwaert, Marie-RoseMouton, Katleen de Saedeleer, Anouk Goethals, AnnNeesen, Indra de Borle, Evelyn Spruyt and Greet Barbier forthe excellent technical support with the animal experiments.We thank Tim Lacoere, Siska Maertens and Lois Maignienfrom LabMET for the support with the DGGE and Illuminasequencing. This work was supported by the SpecialResearch Fund of Ghent University (01D41012), the Con-certed Research Actions of Ghent University (BOF09/GOA/005, BOF10/GOA/021 and BOF12/GOA/008) and theInteruniversity Attraction Poles Program (IUAP, P7/30).Liesbeth Allais is supported by a doctoral grant from theSpecial Research Fund of Ghent University (01D41012).Frederiek-Maarten Kerckhof is supported by a doctoral grantfrom the Concerted Research Actions of Ghent University(BOF09/GOA/005). Ken R. Bracke is a postdoctoralresearcher of the Fund for Scientific Research Flanders(FWO Vlaanderen). No author has an ethical or financialconflict of interest.
References
Ananthakrishnan, A.N. (2013) Environmental risk factors forinflammatory bowel disease. Gastroenterol Hepatol (N Y)9: 367–374.
Ananthakrishnan, A.N. (2015) Environmental risk factors forinflammatory bowel diseases: a review. Dig Dis Sci 60:290–298.
Arcavi, L., and Benowitz, N.L. (2004) Cigarette smoking andinfection. Arch Intern Med 164: 2206–2216.
Arumugam, M., Raes, J., Pelletier, E., Le Paslier, D.,Yamada, T., Mende, D.R., et al. (2011) Enterotypes of thehuman gut microbiome. Nature 473: 174–180.
Berry, D., and Reinisch, W. (2013) Intestinal microbiota: asource of novel biomarkers in inflammatory bowel dis-eases? Best Pract Res Clin Gastroenterol 27: 47–58.
Biedermann, L., Zeitz, J., Mwinyi, J., Sutter-Minder, E.,Rehman, A., Ott, S.J., et al. (2013) Smoking cessationinduces profound changes in the composition of the intes-tinal microbiota in humans. PLoS ONE 8: e59260.
Boltin, D., Perets, T.T., Vilkin, A., and Niv, Y. (2013) Mucinfunction in inflammatory bowel disease: an update. J ClinGastroenterol 47: 106–111.
Boon, N., De Windt, W., Verstraete, W., and Top, E.M. (2002)Evaluation of nested PCR-DGGE (denaturing gradient gelelectrophoresis) with group-specific 16S rRNA primers forthe analysis of bacterial communities from different waste-water treatment plants. FEMS Microbiol Ecol 39: 101–112.
Brandtzaeg, P., and Pabst, R. (2004) Let’s go mucosal: com-munication on slippery ground. Trends Immunol 25: 570–577.
Carbonnel, F., Jantchou, P., Monnet, E., and Cosnes, J.(2009) Environmental risk factors in Crohn’s disease andulcerative colitis: an update. Gastroenterol Clin Biol 33(Suppl. 3): S145–S157.
Cole, J.R., Wang, Q., Cardenas, E., Fish, J., Chai, B., Farris,R.J., et al. (2009) The Ribosomal Database Project:improved alignments and new tools for rRNA analysis.Nucleic Acids Res 37: D141–D145.
Dethlefsen, L., McFall-Ngai, M., and Relman, D.A. (2007) Anecological and evolutionary perspective on human-microbemutualism and disease. Nature 449: 811–818.
D’Hulst, A.I., Vermaelen, K.Y., Brusselle, G.G., Joos, G.F.,and Pauwels, R.A. (2005) Time course of cigarette smoke-induced pulmonary inflammation in mice. Eur Respir J 26:204–213.
Di, Y.P., Zhao, J., and Harper, R. (2012) Cigarette smokeinduces MUC5AC protein expression through the activa-tion of Sp1. J Biol Chem 287: 27948–27958.
Edgar, R.C., Haas, B.J., Clemente, J.C., Quince, C., andKnight, R. (2011) UCHIME improves sensitivity and speedof chimera detection. Bioinformatics 27: 2194–2200.
Elinav, E., Strowig, T., Kau, A.L., Henao-Mejia, J., Thaiss,C.A., Booth, C.J., et al. (2011) NLRP6 inflammasome regu-lates colonic microbial ecology and risk for colitis. Cell 145:745–757.
Erickson, A.R., Cantarel, B.L., Lamendella, R., Darzi, Y.,Mongodin, E.F., Pan, C., et al. (2012) Integratedmetagenomics/metaproteomics reveals human host-microbiota signatures of Crohn’s disease. PLoS ONE 7:e49138.
Frank, D.N., St Amand, A.L., Feldman, R.A., Boedeker, E.C.,Harpaz, N., and Pace, N.R. (2007) Molecular-phylogeneticcharacterization of microbial community imbalances inhuman inflammatory bowel diseases. Proc Natl Acad SciUSA 104: 13780–13785.
Garmendia, J., Morey, P., and Bengoechea, J.A. (2012)Impact of cigarette smoke exposure on host-bacterialpathogen interactions. Eur Respir J 39: 467–477.
Hattrup, C.L., and Gendler, S.J. (2008) Structure and functionof the cell surface (tethered) mucins. Annu Rev Physiol 70:431–457.
Hoebler, C., Gaudier, E., De Coppet, P., Rival, M., andCherbut, C. (2006) MUC genes are differently expressedduring onset and maintenance of inflammation in dextransodium sulfate-treated mice. Dig Dis Sci 51: 381–389.
Hovde, O., and Moum, B.A. (2012) Epidemiology and clinicalcourse of Crohn’s disease: results from observationalstudies. World J Gastroenterol 18: 1723–1731.
Jin, Y., Lin, Y., Lin, L., and Zheng, C. (2012) IL-17/IFN-gammainteractions regulate intestinal inflammation in TNBS-induced acute colitis. J Interferon Cytokine Res 32: 548–556.
Johansson, M.E. (2014) Mucus layers in inflammatory boweldisease. Inflamm Bowel Dis 20: 2124–2131.
Khan, W.I., and Collins, S.M. (2006) Gut motor function:immunological control in enteric infection and inflamma-tion. Clin Exp Immunol 143: 389–397.
Kim, K.C. (2012) Role of epithelial mucins during airwayinfection. Pulm Pharmacol Ther 25: 415–419.
Lakatos, P.L. (2006) Recent trends in the epidemiology ofinflammatory bowel diseases: up or down? World JGastroenterol 12: 6102–6108.
Ley, R.E., Backhed, F., Turnbaugh, P., Lozupone, C.A.,Knight, R.D., and Gordon, J.I. (2005) Obesity alters gutmicrobial ecology. Proc Natl Acad Sci USA 102: 11070–11075.
Linden, S.K., Florin, T.H., and McGuckin, M.A. (2008) Mucindynamics in intestinal bacterial infection. PLoS ONE 3:e3952.
10 L. Allais et al.
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology

Liquet, B., Le Cao, K.A., Hocini, H., and Thiebaut, R. (2012)A novel approach for biomarker selection and the integra-tion of repeated measures experiments from two assays.BMC Bioinformatics 13: 325.
Loftus, E.V. (2004) Clinical epidemiology of inflammatorybowel disease: incidence, prevalence, and environmentalinfluences. Gastroenterology 126: 1504–1517.
Ma, X., Fan, P.X., Li, L.S., Qiao, S.Y., Zhang, G.L., and Li,D.F. (2012) Butyrate promotes the recovering of intestinalwound healing through its positive effect on the tight junc-tions. J Anim Sci 90 (Suppl. 4): 266–268.
Macdonald, G., Kondor, N., Yousefi, V., Green, A., Wong, F.,and Aquino-Parsons, C. (2004) Reduction ofcarboxyhaemoglobin levels in the venous blood of ciga-rette smokers following the administration of carbogen.Radiother Oncol 73: 367–371.
Manichanh, C., Rigottier-Gois, L., Bonnaud, E., Gloux, K.,Pelletier, E., Frangeul, L., et al. (2006) Reduced diversity offaecal microbiota in Crohn’s disease revealed by ametagenomic approach. Gut 55: 205–211.
Muyzer, G., Dewaal, E.C., and Uitterlinden, A.G. (1993) Pro-filing of complex microbial-populations by denaturing gra-dient gel-electrophoresis analysis of polymerase chainreaction-amplified genes-coding for 16S ribosomal-RNA.Appl Environ Microbiol 59: 695–700.
Nakanishi, Y., Sato, T., and Ohteki, T. (2014) CommensalGram-positive bacteria initiates colitis by inducingmonocyte/macrophage mobilization. Mucosal Immunol 8:152–160.
Ovreas, L., Forney, L., Daae, F.L., and Torsvik, V. (1997)Distribution of bacterioplankton in meromictic LakeSaelenvannet, as determined by denaturing gradient gelelectrophoresis of PCR-amplified gene fragments codingfor 16S rRNA. Appl Environ Microbiol 63: 3367–3373.
Persson, P.G., Ahlbom, A., and Hellers, G. (1990) Inflamma-tory bowel disease and tobacco smoke–a case-controlstudy. Gut 31: 1377–1381.
Png, C.W., Linden, S.K., Gilshenan, K.S., Zoetendal, E.G.,McSweeney, C.S., Sly, L.I., et al. (2010a) Mucolytic bacte-ria with increased prevalence in IBD mucosa augment invitro utilization of mucin by other bacteria. Am JGastroenterol 105: 2420–2428.
Png, C.W., Linden, S.K., Gilshenan, K.S., Zoetendal, E.G.,McSweeney, C.S., Sly, L.I., et al. (2010b) Mucolytic bacte-ria with increased prevalence in IBD mucosa augment invitro utilization of mucin by other bacteria. Am JGastroenterol 105: 2420–2428.
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig,W.G., Peplies, J., and Glockner, F.O. (2007) SILVA: a com-prehensive online resource for quality checked and alignedribosomal RNA sequence data compatible with ARB.Nucleic Acids Res 35: 7188–7196.
Quince, C., Lanzen, A., Davenport, R.J., and Turnbaugh, P.J.(2011) Removing noise from pyrosequenced amplicons.BMC Bioinformatics 12: 38.
Rogers, M.A., Greene, M.T., Saint, S., Chenoweth, C.E.,Malani, P.N., Trivedi, I., and Aronoff, D.M. (2012) Higherrates of Clostridium difficile infection among smokers.PLoS ONE 7: e42091.
Rolli, J., Loukili, N., Levrand, S., Rosenblatt-Velin, N.,Rignault-Clerc, S., Waeber, B., et al. (2010) Bacterial
flagellin elicits widespread innate immune defense mecha-nisms, apoptotic signaling, and a sepsis-like systemicinflammatory response in mice. Crit Care 14: R160.
Round, J.L., and Mazmanian, S.K. (2009) The gut microbiotashapes intestinal immune responses during health anddisease. Nat Rev Immunol 9: 313–323.
Round, J.L., O’Connell, R.M., and Mazmanian, S.K. (2010)Coordination of tolerogenic immune responses by the com-mensal microbiota. J Autoimmun 34: J220–J225.
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R.,Hartmann, M., Hollister, E.B., et al. (2009) IntroducingMothur: open-source, platform-independent, community-supported software for describing and comparing microbialcommunities. Appl Environ Microbiol 75: 7537–7541.
Shirazi, T., Longman, R.J., Corfield, A.P., and Probert, C.S.(2000) Mucins and inflammatory bowel disease. PostgradMed J 76: 473–478.
Stecher, B., Maier, L., and Hardt, W.D. (2013) ‘Blooming’ inthe gut: how dysbiosis might contribute to pathogen evo-lution. Nat Rev Microbiol 11: 277–284.
Strober, W., Fuss, I., and Mannon, P. (2007) The fundamentalbasis of inflammatory bowel disease. J Clin Invest 117:514–521.
Tomoda, K., Kubo, K., Asahara, T., Andoh, A., Nomoto, K.,Nishii, Y., et al. (2011) Cigarette smoke decreases organicacids levels and population of bifidobacterium in thecaecum of rats. J Toxicol Sci 36: 261–266.
Verschuere, S., Bracke, K.R., Demoor, T., Plantinga, M.,Verbrugghe, P., Ferdinande, L., et al. (2011) Cigarettesmoking alters epithelial apoptosis and immune composi-tion in murine GALT. Lab Invest 91: 1056–1067.
Verschuere, S., Allais, L., Bracke, K.R., Lippens, S.,De Smet, R., Vandenabeele, P., et al. (2012) Cigarettesmoke and the terminal ileum: increased autophagy inmurine follicle-associated epithelium and Peyer’s patches.Histochem Cell Biol 137: 293–301.
Wang, H., Zhao, J.X., Hu, N., Ren, J., Du, M., and Zhu, M.J.(2012) Side-stream smoking reduces intestinal inflamma-tion and increases expression of tight junction proteins.World J Gastroenterol 18: 2180–2187.
Xavier, R.J., and Podolsky, D.K. (2007) Unravelling thepathogenesis of inflammatory bowel disease. Nature 448:427–434.
Yu, H., Li, Q., Kolosov, V.P., Perelman, J.M., and Zhou, X.(2012) Regulation of cigarette smoke-mediated mucinexpression by hypoxia-inducible factor-1alpha via epider-mal growth factor receptor-mediated signaling pathways.J Appl Toxicol 32: 282–292.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article at the publisher’s web-site:
Fig. S1. Agglomerative nesting clustering dendrogramsusing the abundance-based Yue and Clayton’s θ based onthe DGGE data with average linkage (UPGMA), normalizedin BIONUMERICS 5.10 software. (A) clustering dendrogram forileum, (B) clustering dendrogram for caecum and (C) cluster-ing dendrograms for distal colon. SM, smoking; NSM, non-smoking; DC, distal colon; IL, ileum; CC, caecum.
Cigarette smoke induces shifts in the murine gut 11
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology

Fig. S2. (A) mRNA expression of TNF-α, IL-10, NFκB andTGF-β in the ileum after air and smoke exposure, relative tothe expression of two reference genes [glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and hydroxymethyl-bilane synthase (HMBS)]. (B) mRNA expression of IL-10,NFκB, IFN-γ, CXCL2, IL-1β and TNF-α in the proximal colonafter air and smoke exposure. (C) mRNA expression ofCXCL2, IL-1β, IL-6 and IFN-γ in the distal colon after air andsmoke exposure. (D) mRNA expression of TNF-α, IL-10,NFκB and IFN-γ in the distal colon after air and smoke
exposure. Data are represented as mean ± SEM; n = 6 ineach group.Fig. S3. mRNA expression of tight junction genes in theileum, proximal and distal colon after air and smoke expo-sure, relative to the expression of two reference genes[glyceraldehyde-3-phosphate dehydrogenase (GAPDH) andhydroxymethylbilane synthase (HMBS)]. Data are repre-sented as mean ± SEM; n = 6 in each group.Appendix. Raw Illumina sequencing data, containing OTUtable and LefSe data.
12 L. Allais et al.
© 2015 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology
![ABOUT ALLAIS EFFECT AND EARTH´S ELECTROCONVERGENCE · Allais = sin[2 (x – φ)], (1) is referred to as Allais effect term in this text. M.Allais does not define parameter k. Parameter](https://img.pdfslide.net/doc/110x75/605ee10fcc849009d665e9e3/about-allais-effect-and-earths-electroconvergence-allais-sin2-x-a-.jpg)




























