Embed Size (px)
Citation preview

67Medit. Mar. Sci., 8/1, 2007, 67-89
Introduction
The Mediterranean is one of the mostimportant enclosed seas in the world witha large shipping circulation but narrowcommunication with two oceans by theGibraltar strait (Atlantic Ocean) and bythe man-made Suez Canal (Indian Oceanthrough the Red Sea). It also has a freewater exchange with its annexed Black Seathrough the Dardanelles, the Sea of Mar-mara and the Bosporus, all of which, thatin spite of marked particularities are inreality annexed parts of the Mediter-
ranean. This whole frame of marine andbrackish waters conveys a large variety ofbiota, which enter the Mediterranean con-tinuously and include organisms frequent-ly described as ‘allochthonous’, ‘exotic’,‘alien’, ‘introduced’ by man or ‘vagrantvisitors’. According to dependable ecolog-ical parameters they have settled in theMediterranean for a few or a large num-ber of years, temporarily or permanently,with large or scarce populations, followingtheir ecological requirements and theshort or long term environmental oscilla-tions. Therefore, the Mediterranean is by
Mediterranean Marine ScienceVolume 8/1, 2007, 67-89
Allochthonous and vagrant ichthyofauna in Hellenic marine and estuarine waters
M. CORSINI-FOKA1 and P. S. ECONOMIDIS2
1 Hellenic Center for Marine Research,Hydrobiological Station of Rhodes, Cos Street, 85100 Rhodes, Hellas
2 Karakasi Street, 79, 54453 Thessaloniki, Hellas
e-mail: [email protected]
Abstract
A review of the non-indigenous ichthyofauna occurring in the Hellenic marine and estuarine watersis presented, including Atlantic origin colonizers, aquaculture introduced and Lessepsian alien species. 34non-native species have been registered. The majority of allochthonous fish are Lessepsian immigrants,which represent approximately 80% of the ascertained non-native ichthyofauna in Hellenic waters; theirestablishment, spread, habitat, abundance and interaction with indigenous fish are discussed.
Keywords: Allochthonous fish species; Alien fish species; Greece; Ionian; Aegean; Mediterranean;Lessepsian migration.
Review Article

far the major recipient of exotic speciesand vagrant visitors like macrophytes,invertebrates and fish. Apart from therather free energetic movement throughthe above passages, ships’ ballast, fouling,clinging of ship hulls and aquaculture arealso important vectors of non-indigenousmarine species. Since the last glacial peri-od, it seems the Mediterranean has pre-sented a continuous exchange of faunawith the Atlantic and the Black Sea. Dur-ing the last 150 years since the opening ofthe Suez Canal, it has been presentingmany Indo-Pacific species, which migratethrough it. The rate of this so-calledLessepsian migration has increased in thelast decades, with ecological and economicimpacts, mainly in the Levantine basin.
The Mediterranean Ichthyofauna
In a rather simplified scheme, whichcould also be adapted and used for theHellenic seas, recent Mediterraneanichthyofauna is composed of four maingroups of species: (1) endemic(QUIGNARD & TOMASINI, 2000), (2)Atlantic, Boreal and Tropical (sensuTORTONESE, 1960, 1964, 1967, 1970;WHITEHEAD et al., 1984-86). TheseAtlanto-Mediterranean species occur ornot in the Mediterranean according toenvironmental factors, many of whichoscillate significantly depending on longor short term climatic changes. Altogetherthese species are called ‘autochthonous’.Several such species, with rare or periodicappearance, could be described as‘vagrant’ or ‘visitors’ but not ‘alien’ or‘exotic’, even if they are only present inlimited concentration and many of themare, in fact, detected with difficulty. Otherscould also be called ‘new colonizers’, par-ticularly when their spread is favoured by
special circumstances such as the tendencytowards a change in temperature and cur-rent regime in the NE Atlantic. (3) Intro-duced by man (sensu HUNTER &GIBBS, 2006) mainly for aquaculture oraquarium purposes, and which escape intothe wild or are carelessly released. This is arather obscure phenomenon and actuallythere is no or only very little elementaryknowledge of its impact on the naturalenvironment and genetic compatibilityafter mating with closely-related wildspecies. In this category should also beincluded the release with the ballast waterof various taxa, many of which maybe‘exotic’, and (4) the Lessepsian or Erythre-an, of Red Sea origin immigrants, whichare real ‘exotic’ or ‘alien’ species. In sev-eral cases, when a population explosionhas occurred, these last could be charac-terized as really ‘invasive’ (HUNTER &GIBBS, 2006), as in the case of the recentcolonizer Fistularia commersonii in manyareas of the Eastern Mediterranean.
Greek Allochthonous fish species
Among a number of about 500 nativemarine fish species in the Hellenic waters,34 allochthonous have been registered,while seven others are not yet confirmedfor various reasons (Table 1). Among thelast, six species belong to the category ofintroduced for aquaculture and their pres-ence in nature is based on unverifiedobservation of free swimming specimensor unidentified samples, strongly suggest-ing careless release or escape from fishfarms. From the remainder, the majority(27 species plus one uncertain, i.e. 79.4%)originate from the Indo-Pacific Ocean andthe Red Sea having penetrated into theMediterranean via the Suez Canal. Amongthe other seven species, four are usually
68 Medit. Mar. Sci., 8/1, 2007, 67-89
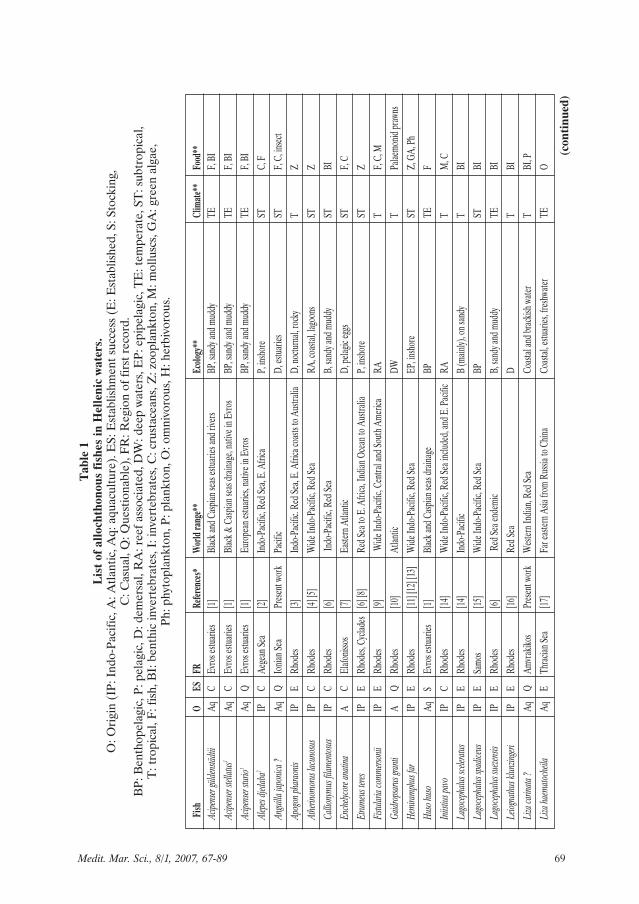
69Medit. Mar. Sci., 8/1, 2007, 67-89
Tab
le 1
Lis
t of
all
ocht
hono
us f
ishe
s in
Hel
leni
c w
ater
s.
O: O
rigi
n (I
P: I
ndo-
Pac
ific,
A: A
tlant
ic, A
q: a
quac
ultu
re).
ES:
Est
ablis
hmen
t suc
cess
(E
: Est
ablis
hed,
S: S
tock
ing,
C
: Cas
ual,
Q: Q
uest
iona
ble)
, FR
: Reg
ion
of fi
rst r
ecor
d.B
P: B
enth
opel
agic
, P: p
elag
ic, D
: dem
ersa
l, R
A: r
eef a
ssoc
iate
d, D
W: d
eep
wat
ers,
EP
: epi
pela
gic,
TE
: tem
pera
te, S
T: s
ubtr
opic
al,
T: t
ropi
cal,
F: f
ish,
BI:
ben
thic
inve
rteb
rate
s, I
: inv
erte
brat
es, C
: cru
stac
eans
, Z: z
oopl
ankt
on, M
: mol
lusc
s, G
A: g
reen
alg
ae,
Ph:
phy
topl
ankt
on, P
: pla
nkto
n, O
: om
nivo
rous
, H: h
erbi
voro
us.
Fish
OES
FRRe
feren
ces*
Wor
ld ra
nge*
*Ec
ology
**Cl
imat
e**
Food
**Ac
ipens
er gü
ldens
tädtii
AqC
Evro
s estu
aries
[1]
Blac
k and
Cas
pian s
eas e
stuar
ies an
d rive
rsBP
, san
dy an
d mud
dyTE
F, B
I
Acipe
nser
stella
tus1
AqC
Evro
s estu
aries
[1]
Blac
k & C
aspia
n sea
s dra
inage
, nati
ve in
Evro
sBP
, san
dy an
d mud
dyTE
F, B
I
Acipe
nser
sturio
1Aq
QEv
ros e
stuar
ies[1]
Eu
rope
an es
tuar
ies, n
ative
in E
vros
BP, s
andy
and m
uddy
TEF,
BI
Alep
es dje
daba
2IP
CAe
gean
Sea
[2]
Indo
-Pac
ific,
Red S
ea, E
. Afri
caP,
insh
ore
STC,
F
Angu
illa ja
ponic
a ?Aq
QIo
nian S
eaPr
esen
t wor
kPa
cific
D, es
tuar
iesST
F, C
, inse
ct
Apog
on ph
arao
nisIP
ERh
odes
[3]
Indo
-Pac
ific.
Red S
ea, E
. Afri
ca co
asts
to A
ustra
liaD,
noctu
rnal,
rock
yT
Z
Athe
rinom
orus
lacu
nosu
sIP
CRh
odes
[4] [5
] W
ide In
do-P
acifi
c, Re
d Sea
RA, c
oasta
l, lag
oons
STZ
Callio
nymu
s fila
mento
sus
IPC
Rhod
es[6]
In
do-P
acifi
c, Re
d Sea
B, sa
ndy a
nd m
uddy
STBI
Ench
elyco
re an
atina
AC
Elafo
nisso
s[7]
Ea
stern
Atla
ntic
D, pe
lagic
eggs
STF,
C
Etru
meus
teres
IPE
Rhod
es, C
yclad
es[6]
[8]
Red S
ea to
E. A
frica
, Ind
ian O
cean
to A
ustra
liaP,
insh
ore
STZ
Fistu
laria
comm
erson
iiIP
ERh
odes
[9]
Wide
Indo
-Pac
ific,
Cent
ral a
nd So
uth A
meric
aRA
TF,
C, M
Gaidr
opsa
rus g
ranti
AQ
Rhod
es[10
] At
lantic
DWT
Palae
monid
praw
ns
Hemi
ramp
hus f
arIP
ERh
odes
[11] [
12] [
13]
Wide
Indo
-Pac
ific,
Red S
eaEP
, insh
ore
STZ,
GA,
Ph
Huso
huso
AqS
Evro
s estu
aries
[1]
Blac
k and
Cas
pian s
eas d
raina
geBP
TEF
Iniis
tius p
avo
IPC
Rhod
es[14
] W
ide In
do-P
acifi
c, Re
d Sea
inclu
ded,
and E
. Pac
ific
RAT
M, C
Lago
ceph
alus s
celer
atus
IPE
Rhod
es[14
]In
do-P
acifi
cB
(main
ly), o
n san
dy
TBI
Lago
ceph
alus s
padic
eus
IPE
Samo
s[15
] W
ide In
do-P
acifi
c, Re
d Sea
BPST
BI
Lago
ceph
alus s
uezen
sisIP
ERh
odes
[6]
Red S
ea en
demi
cB,
sand
y and
mud
dyTE
BI
Leiog
nathu
s klun
zinge
riIP
ERh
odes
[16]
Red S
eaD
TBI
Liza
carin
ata ?
AqQ
Amvra
kikos
Pres
ent w
ork
Wes
tern I
ndian
, Red
Sea
Coas
tal an
d bra
ckish
wate
rT
BI, P
Liza
haem
atoch
eila
AqE
Thra
cian S
ea[17
] Fa
r eas
tern A
sia fr
om R
ussia
to C
hina
Coas
tal, e
stuar
ies, f
resh
water
TEO
(con
tinu
ed)
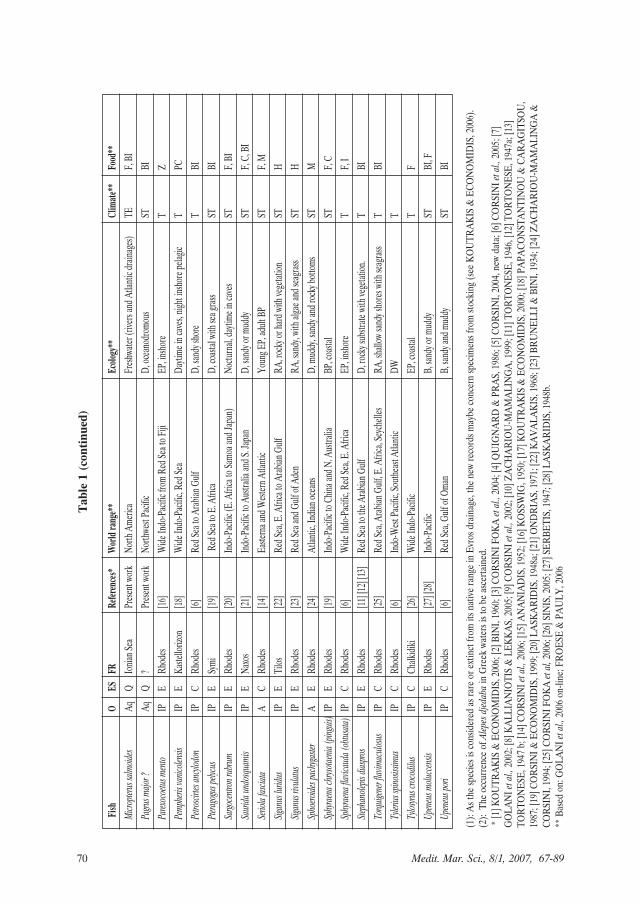
70 Medit. Mar. Sci., 8/1, 2007, 67-89
Fish
OES
FRRe
feren
ces*
Wor
ld ra
nge*
*Ec
ology
**Cl
imat
e**
Food
**
Micr
opter
us sa
lmoid
esAq
QIo
nian S
eaPr
esen
t wor
kNo
rth A
meric
aFr
eshw
ater (
river
s and
Atla
ntic
drain
ages
)TE
F, B
I
Pagru
s majo
r ?Aq
Q?
Pres
ent w
ork
North
west
Pacif
icD,
ocea
nodr
omou
sST
BI
Parex
ocoe
tus m
ento
IPE
Rhod
es[16
] W
ide In
do-P
acifi
c fro
m Re
d Sea
to F
ijiEP
, insh
ore
TZ
Pemp
heris
vanic
olens
isIP
EKa
stello
rizon
[18]
Wide
Indo
-Pac
ific,
Red S
eaDa
ytime
in ca
ves,
night
insh
ore p
elagic
TPC
Petro
scirte
s anc
ylodo
nIP
CRh
odes
[6]
Red S
ea to
Ara
bian G
ulfD,
sand
y sho
reT
BI
Ptera
gogu
s pely
cus
IPE
Symi
[19]
Red S
ea to
E. A
frica
D, co
astal
with
sea g
rass
STBI
Sargo
centr
on ru
brum
IPE
Rhod
es[20
] In
do-P
acifi
c (E.
Afri
ca to
Samo
a and
Japa
n)No
cturn
al, da
ytime
in ca
ves
STF,
BI
Saur
ida un
dosq
uami
sIP
ENa
xos
[21]
Indo
-Pac
ific t
o Aus
tralia
and S
. Jap
anD,
sand
y or m
uddy
ST
F, C
, BI
Serio
la fas
ciata
AC
Rhod
es[14
] Ea
stern
a and
Wes
tern A
tlant
icYo
ung E
P, ad
ult B
PST
F, M
Sigan
us lu
ridus
IPE
Tilos
[22]
Red S
ea, E
. Afri
ca to
Ara
bian G
ulfRA
, roc
ky or
hard
with
vege
tation
STH
Sigan
us ri
vulat
usIP
ERh
odes
[23]
Red S
ea an
d Gulf
of A
den
RA, s
andy
, with
alga
e and
seag
rass
STH
Spho
eroide
s pac
hyga
ster
AE
Rhod
es[24
] At
lantic
, Ind
ian oc
eans
D, m
uddy
, san
dy an
d roc
ky bo
ttoms
STM
Sphy
raen
a chr
ysotae
nia (p
inguis
)IP
ERh
odes
[19]
Indo
-Pac
ific t
o Chin
a and
N. A
ustra
liaBP
, coa
stal
STF,
C
Sphy
raen
a flav
icaud
a (ob
tusata
)IP
CRh
odes
[6]
Wide
Indo
-Pac
ific,
Red S
ea, E
. Afri
caEP
, insh
ore
TF,
I
Steph
anole
pis di
aspr
osIP
ERh
odes
[11] [
12] [
13]
Red S
ea to
the A
rabia
n Gulf
D, ro
cky s
ubstr
ate w
ith ve
getat
ion.
TBI
Torqu
igene
r flav
imac
ulosu
sIP
CRh
odes
[25]
Red S
ea, A
rabia
n Gulf
, E. A
frica
, Sey
chell
esRA
, sha
llow
sand
y sho
res w
ith se
agra
ssT
BI
Tyler
ius sp
inosis
simus
IPC
Rhod
es[6]
In
do-W
est P
acifi
c, So
uthe
ast A
tlant
icDW
T
Tylos
yrus c
roco
dilus
IPC
Chalk
idiki
[26]
Wide
Indo
-Pac
ific
EP, c
oasta
lT
F
Upen
eus m
olucc
ensis
IPE
Rhod
es[27
] [28
] In
do-P
acifi
cB,
sand
y or m
uddy
ST
BI, F
Upen
eus p
ori
IPC
Rhod
es[6]
Re
d Sea
, Gulf
of O
man
B, sa
ndy a
nd m
uddy
STBI
(1):
As t
he sp
ecie
s is c
onsid
ered
as r
are
or e
xtin
ct fr
om it
s nat
ive
rang
e in
Evr
os d
rain
age,
the
new
reco
rds m
aybe
con
cern
spec
imen
s fro
m st
ocki
ng (s
ee K
OU
TRA
KIS
& E
CO
NO
MID
IS, 2
006)
.(2
): T
he o
ccur
renc
e of
Alep
es d
jedab
ain
Gre
ek w
ater
s is t
o be
asc
erta
ined
.*
[1] K
OU
TRA
KIS
& E
CO
NO
MID
IS, 2
006;
[2] B
INI,
1960
; [3]
CO
RSI
NI F
OK
A e
t al.,
200
4; [4
] QU
IGN
AR
D &
PR
AS,
198
6; [5
] CO
RSI
NI,
2004
, new
dat
a; [6
] CO
RSI
NI e
t al.,
2005
; [7]
GO
LAN
I et a
l.,20
02; [
8] K
ALL
IAN
IOTI
S &
LEK
KA
S, 2
005;
[9] C
OR
SIN
I et a
l.,20
02; [
10] Z
AC
HA
RIO
U-M
AM
ALI
NG
A, 1
999;
[11]
TO
RTO
NES
E, 1
946,
[12]
TO
RTO
NES
E, 1
947a
; [13
]TO
RTO
NES
E, 1
947
b; [1
4] C
OR
SIN
I et a
l.,20
06; [
15] A
NA
NIA
DIS
, 195
2; [1
6] K
OSS
WIG
, 195
0; [1
7] K
OU
TRA
KIS
& E
CO
NO
MID
IS, 2
000;
[18]
PA
PAC
ON
STA
NTI
NO
U &
CA
RA
GIT
SOU
,19
87; [
19] C
OR
SIN
I & E
CO
NO
MID
IS, 1
999;
[20]
LA
SKA
RID
IS, 1
948a
; [21
] ON
DR
IAS,
197
1; [2
2] K
AV
ALA
KIS
, 196
8; [2
3] B
RU
NEL
LI &
BIN
I, 19
34; [
24] Z
AC
HA
RIO
U-M
AM
ALI
NG
A &
CO
RSI
NI,
1994
; [25
] CO
RSI
NI F
OK
A e
t al,
2006
; [26
] SIN
IS, 2
005;
[27]
SER
BETI
S, 1
947;
[28]
LA
SKA
RID
IS, 1
948b
.**
Bas
ed o
n: G
OLA
NI e
t al.,
200
6 on
-line
; FR
OES
E &
PA
ULY
, 200
6
Tab
le 1
(co
ntin
ued)

considered to originate from the AtlanticOcean and three, two sturgeons and onemullet, come from aquaculture eitherfrom stocking or escape.
All these allochthonous species can beclassified in three main groups accordingto their origin: (a) Atlantic, (b) Aquacul-ture and (c) Indo-Pacific. The first groupof species could eventually be includedmore easily in the indigenous Mediter-ranean species, according to the aboveexpressed points. The list of species com-ing from aquaculture is expected to grad-ually increase, as the number of aquacul-ture installations is increasing in the Hel-lenic seas and in the neighbouring areas aswell. The Indo-Pacific group includes themajority of allochthonous species, whichare real alien species in the Mediter-ranean as they have penetrated an areaout of their original distribution. It isworth noting that the majority of thesespecies have quite a slow process ofspread and settlement, underlining theirthermophilous and stenohaline, tropicalor sub-tropical, character. In the Mediter-ranean such environmental conditions arecurrent in the eastern basin, mainly alongthe Asian coasts. In the Aegean Sea aclosely related situation occurs in the SEcorner around the Dodecanese continen-tal plateau. There is evidence that the con-tribution of the Indo-Pacific alien speciesto the Aegean Sea fish species composi-tion and ecosystem structure, is graduallyaccelerating, as many populations of thesespecies have already constructed commer-cial stocks, as did long ago the Upeneusand Siganus species in the Dodecaneseislands and more recently Fistularia com-mersonii (CORSINI et al., 2002;KALOGIROU et al., 2007) and Etrumeusteres in the Cyclades (KALLIANIOTIS &LEKKAS, 2005) and in the Dodecanese
as well (CORSINI-FOKA, unpubl. data).
Atlantic vagrant and new colonizer
As aforementioned, the vagrant or vis-itor and the new colonizer fish species ofAtlantic origin are not easy to enumeratebecause their flux has been continuing formore than 10,000 years. Various attempts,however, have been made at standardiza-tion of their origin and at the calculation ofthe number of Mediterranean fish species(TORTONESE, 1938, 1963, 1970, 1975,1982; QUIGNARD & TOMASINI,2000). Furthermore, as research in variouspart of the Mediterranean becomes moreintensive, many new species are registeredeach year, mainly in the western part of thesea, which is closest to the Atlantic. Thiscould also be attributed to other reasons,such as the warming trend of the Mediter-ranean, the expansion of maritime traffic,etc. (GOLANI et al., 2002) or combinationof all these reasons. There is widespreaddiscussion regarding the exact number ofAtlantic vagrant and new colonizer fishspecies (QUIGNARD & TOMASINI,2000). Recent estimations (up to 2006)have considered those species enteringthrough the Gibraltar straits and crossinginto the Mediterranean to be over 30(GOLANI et al., 2006 on-line; REINA-HERVA’ S et al., 2004), also reaching theAegean (ERYILMAZ et al., 2003;CORSINI et al., 2006) and the AdriaticSea as with the recently recordedCyclopterus lumpus (DUL
v
CIv
C &GOLANI, 2006). Some of them, particu-larly Sphoeroides pachygaster, Seriola fasci-ata and Seriola carpenteri have achievedsignificant biomass (ANDALORO, 2001;ANDALORO et al., 2005).
Three such species have been record-ed in the Aegean: Sphoeroides pachygaster,
71Medit. Mar. Sci., 8/1, 2007, 67-89

a species well established throughout theMediterranean, Enchelycore anatina andSeriola fasciata, while the consideration ofGaidropsarus granti as a non-indigenousAtlantic colonizer is questionable (Table 1).
The regular or rare appearance ofAtlantic species in the Mediterranean is amore complicated phenomenon, depend-ing mainly on hydro-climatic long-termchanges and short-term trophic relationsamong species. In a very preliminary andsimplified approach, when North Atlanticwaters become warmer, there is an accessof temperate, even sub-tropical, speciesinto the Mediterranean. On contrary,when Atlantic waters become colder,northern species enter and colonize theMediterranean. This play seems to havetaken place repeatedly during the glacierperiod dominating in the northern hemi-sphere of the planet during the last onemillion years. The inter-glacier periodfavours the first while the glacier the sec-ond movement (see also QUIGNARD &TOMASINI, 2000). Actually, there is arather clear range of these warm water or‘thermophilous’ and cold water or ‘psy-chrophilous’ fish fauna in the Mediter-ranean, as the first is distributed in thesouth mainly along the African and Asiancoasts, while the second is restricted to theEuropean coasts in areas where cold waterdominates (North Adriatic, NorthAegean, Black Sea, etc.), following theactual temperature and current regime.
Active movement according to trophicavailability concerns the errant largepelagic species such as sharks, variousscombrids, tetrapterids, swordfish, etc.Several such species are season regular orrare vagrant visitors. In some other cases,such as the sea lamprey (Petromyzon mari-nus), a parasitic mode of life forces it tomove far away from its normal distribu-
tion. Therefore, the recording of this lastspecies in the North Aegean Sea(ECONOMIDIS et al., 1999), where thereare no spawning grounds for permanentsettlement, could be qualified as acciden-tal and the species as ‘vagrant’ and not asexotic or as alien, because the area is with-in its large range. A similar example wouldbe any species of remoras, accompanyinglarge marine animals, such as the Remile-gia australis (= Remora australis), noticedlong ago in Hellenic waters(TORTONESE, 1946, 1947a), and which,according to GOLANI et al. (2002),although very rare could be considered aMediterranean resident.
In several other cases, such as theAtlantic muraenid Enchelycore anatina,spread from the Atlantic to the Mediter-ranean, where there are only two recordsone of which being from Elafonissos(Peloponnesus coasts) (GOLANI et al.,2002), could be explained by passive dis-semination of the leptocephali larvaeand/or via transport in ship ballast.
Aquaculture introduced
Uncontrolled and/or collapsed aqua-cultures are responsible for the releasingof specimens into nature from their rearedstocks. Some few cases have already beenreported in the Greek seas by picturedand/or preserved specimen, but informa-tion has not been yet published. One ofthese may concern some mullet specimens,apparently Liza carinata, captured in west-ern Greek estuaries (Table 1). Such aspecies was introduced long ago as fryfrom Egypt to a fish farm near Arta (riverArachthos estuary in the AmvrakikosGulf), which subsequently closed down(unpublished data). However, Liza carina-ta is listed among Indo-Pacific aliens
72 Medit. Mar. Sci., 8/1, 2007, 67-89

(GOLANI et al., 2002), since it has alsobeen recorded in the Levantine basin,including the Turkish Mediterranean andAegean coasts (TORCU & MATER,2000; ZAITSEV & ÖZTÜRK, 2001;BILECENOGLU et al., 2002). Conse-quently, its occurrence could be also dueto the expansion of this lessepsian immi-grant distribution from the EasternMediterranean to the Ionian Greek waterspreviously mentioned.
It is known that around Greek andother European estuaries, among cap-tured growing European eel (Anguillaanguilla) there also are some specimens ofthe elvers of the Japanese eel (Anguillajaponica), species largely used in fishfarms, which have escaped from eel farms(unpublished data). Another case con-cerns the largemouth bass (Micropterussalmoides) that was observed in 2003 andphotographed near the estuaries of thestream Lessini, flowing directly into theIonian Sea (unpublished data) (Table 1).Recent records concerning several speciesof sturgeons such as Acipenser güldenstän-di, Huso huso or various rearing hybrids, aswell as Acipenser sturio, captured in theThracian Sea, near the Evros estuaries orin the river itself, strongly suggest that theyoriginate from the release of fry orescapees from hatcheries lying in theEvros river catchment of Bulgaria and/orof Turkey, as there are no such installa-tions in the Hellenic part of the river(KOUTRAKIS & ECONOMIDIS, 2006).Therefore, some previous records ofAcipenser stellatus (ECONOMIDIS et al.,2000) maybe have had the same origin.Furthermore, there is unverified informa-tion that certain alien fish species (forinstance Pagrus major) or even fry of nativeMediterranean fish, such as Dicentrarchuslabrax and Sparus auratus, originating from
distant areas (i.e. Spain), are used widelyin marine Greek fish farms. Consequently,very often these species of different genet-ic stocks are released or escape as fry oradults into the wild. But there is a con-firmed record of the Indo-Pacific nativemullet, Liza haematocheilus (Mugil soiuy),which was reared in Black Sea cage fishfarms. Many of these farms collapsed inthe nineties and specimens were carelesslyreleased in the Azov and Black Sea areas(STARUSHENKO & KAZANSKY,1996). Soon after, the species was observedin the Turkish Aegean Sea (Gulf of Smyr-na: KAYA et al., 1998) and in the ThracianSea (KOUTRAKIS & ECONOMIDIS,2000) (see also HARRISON, 2004). Actu-ally, it forms fished population in variousThracian and Macedonia lagoons and it iscaptured and appears frequently in the fishmarkets of North Greece. (After MINOSet al., 2007 and in press).
Lessepsian Aliens
The movement of various marineorganisms from the Red Sea to theMediterranean via the Suez Canal wasnamed Lessepsian migration by POR(1969), a term widely used in literaturethereafter; it is very often also called Ery-threan migration. According to GALIL &ZENETOS (2002), the Eastern Mediter-ranean, open to Atlantic, Pontic and Ery-threan biota, is particularly prone to inva-sion. Concerning fish, the Lessepsianspecies in the Mediterranean are real alienspecies because they belong to anotherecological status (thermophilous) and to adifferent bio-geographical zone (Indo-Pacific), and the Mediterranean is out oftheir native distribution.
In recent years intensive research hasbrought to light more alien species which
73Medit. Mar. Sci., 8/1, 2007, 67-89

might have been present in low densitiesand thus escaped our attention(CORSINI & ECONOMIDIS, 1999).The immigration of such alien species isfar from ceasing after approaching a stan-dard level, since in practice the flux ofinvaders is continuous (STREFTARIS etal., 2005). The dynamics of invasions andextension of the distribution of alienspecies obviously need a continuousupdate and a monitoring procedure(GOLANI et al., 2004; ZENETOS et al.,2005; PANCUCCI-PAPADOPOULOUet al., 2005a, b).
Establishment. According to the morerecent inventories the Lessepsian fishmigrants in the Mediterranean are oneElasmobranchii species and more than 60Teleostei species (GOLANI et al., 2006on-line; GOKOGLU et al., 2003;CORSINI et al., 2005; AKYOL et al., 2005;CORSINI et al., 2006; BILECENOGLU& KAYA, 2006; CINAR et al., 2006;GOLANI, 2006; ERYLMAZ &DALYAN, 2006; GOLANI & SONIN,2006). Once arrived in the MediterraneanSea through the Suez Canal, these speciesmove either westwards to the African
74
Fig. 1: The Dodecanese islands and the Hellenic Seas.
Medit. Mar. Sci., 8/1, 2007, 67-89

coasts or usually eastwards first and thennorthwards, following mainly the Asiaticcontinental shelf. They normally establish,fast or slow, dense or scarce populations,in the Levantine Sea where they influenceor even modify essentially the compositionand the structure of the local marineecosystems. The common way for furthercolonization of the Mediterranean is main-ly by using the South coast of Turkey andits narrow continental shelf. Besides, thewater masses around the DodecaneseIslands form the most important gates forentering and establishing in the AegeanSea and the main pathway of spread with-in the Mediterranean (PANCUCCI-PAPADOPOULOU et al., 2005a, b) (Fig.1). A number of 28 Lessepsian immigrantfish species has entered Greek waters(Table 1). Three additional ones havebeen reported from the SE Aegean Sea,along the Turkish coasts, Scomberomoruscommerson (BUHAN et al., 1997) andOxyurichthys petersi (AKYOL et al., 2006)in Gokova Bay and Sillago sihama in Datçapeninsula (BILECENOGLU, 2004).Older records of Lessepsian fish inHellenic waters amounted tothirteen species until 1990(PAPACONSTANTINOU, 1987, 1990;also References in Table 1). An evidentincrease in records has been observed inthe last decade, when 15 other such specieswere added. A similar picture is presentedby the Erythrean crustaceans, which sud-denly occurred in the SE Aegean in thepast decade and, according to GALIL &KEVREKIDIS (2002), this may be due tothe more extensive inflow of the AsiaMinor Current in the area.
CORSINI & ECONOMIDIS (1999)mention that several Lessepsian specieswere reported in the Dodecanese almostsimultaneously as along the coasts of
Israel. Such are the cases of Siganus rivula-tus, Upeneus moluccensis, Sargocentronrubrum, Pteragogus pelycus and Fistulariacommersonii. On the contrary, as for exam-ple the cases of Sphyraena chrysotaenia,Apogon pharaonis, Etrumeus teres andsome others, the advance along the Anato-lian coasts was gradual and the immigrantsreached this area after a relatively longtime, depending on biotic and abiotic fac-tors (GOLANI, 1998a; CORSINI &ECONOMIDIS, 1999; CORSINI et al.,2002, 2005; CORSINI FOKA et al., 2004)(Table 1). There is a direct relationbetween the environmental conditions andthe speed of spread and adaptation of thenew settler. In fact, species have to face acombination of unfavourable ecologicalconditions (water temperature, substra-tum, currents, trophic conditions etc.)when following the narrow continentalshelf of the South Anatolian coast as themain way for westward spread, as themajority of species are coastal. Conse-quently, once they reach the continentalshelf of the Dodecanese islands, they donot usually advance any further; some ofthem are also commercially exploited.
On the other hand, a large number ofLessepsian immigrants, being quite com-mon in the Levantine, have not beenrecorded in the SE Aegean Sea. This dif-ferent trend could be owing to many bioticand/or abiotic factors as, for example, thetemperature regime and the thermal toler-ance of the colonizing species, food avail-ability, competition with indigenousspecies, local pathogens and the suitablesubstrate (GOLANI, 1998a). The exten-sion of the spawning season, the aggressivebehaviour of the new species, their foodpreferences, could also be added. Coloniz-ers may also occupy free niches alreadyexisting or created by over-fishing of local
75Medit. Mar. Sci., 8/1, 2007, 67-89

species. All these, like many other factors,need a more accurate survey to obtain acomplete view of the real situation.
In comparison, along the Turkish terri-torial waters of the Aegean and the EastMediterranean, 38 species of teleosts ofRed Sea origin have been recorded(GOLANI et al., 2006 on-line; BILECE-NOGLU et al., 2002; CINAR et al., 2005,2006; BILECENOGLU & KAYA, 2006).
A number of 25 species is common inthe Hellenic and the Turkish waters.
It is obvious that several fish speciesalready detected and established along theLevantine Turkish coasts can be expectedalso in Hellenic waters, especially Dussum-ieria elopsoides, Herklotsichthys punctatus,Pelates quadrilineatus, Cynoglossussinusarabici, Liza carinata.
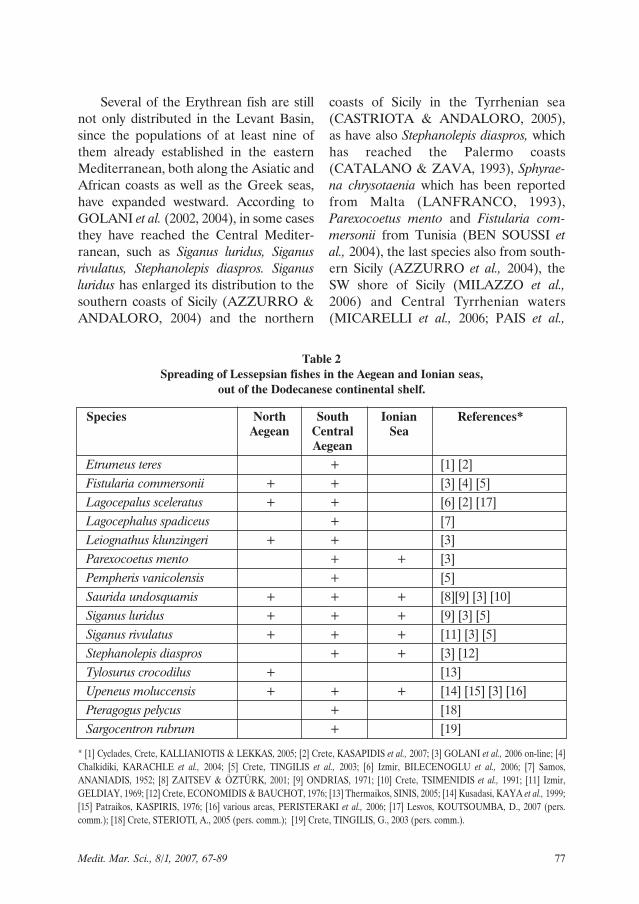
Spreading. Lessepsian migrant speciesonce having reached the Dodecanese con-tinental shelf seem to have difficulties infurther spread to the rest of the AegeanSea, mainly to the north, or in continuingtheir expansion to all the Mediterranean.Among the above 28 Lessepsian fishspecies (Table 1), only nine have beenrecorded further north than the Dode-canese plateau, five before the last decadeand the other four during the last sevenyears (Table 2). The recording of the Tylo-surus crocodilus in the Chalkidiki peninsu-la, based on a dead fish found on Gerakinibeach (SINIS, 2005), needs a special men-tion as it is a recording which merits fur-ther investigation. This is a first recordingin the Mediterranean and its derivation asa rejected specimen from undelivered for-eign stock should also be considered.
Spreading to the south, central or west-ern Greek waters of the Aegean and theIonian seas seems to have the same low suc-cess, as only fourteen and six species respec-tively followed these routes (Table 2).
This statement shows that some abiot-ic (water temperature, salinity, currentregime, size of continental shelf, etc.)and/or biotic factors (food quality andquantity, spawning facilities, etc.) areimpeding the spread of Erythrean fish.
In general, the species which havespread out of the Dodecanese ‘refuge’(Table 2) have a wide native distributionarea (Table 1). Although tropical or sub-tropical fish, they are able to colonize tem-perate regions quite easily, the water tem-perature not prohibiting their spread.
Although the Cretan continental shelflies in the south Aegean Sea in a ratherwarm water zone, colonization by Lessep-sian immigrants presents some interestingparticularities. Since these species areregarded as thermophilous, it would beexpected that they would settle more easi-ly in the coastal waters of Crete. However,unfavourable factors obviously seem to bean interdiction for their large scale settle-ment there as only eleven fish species wererecorded, two before 2002, Stephanolepisdiaspros and Saurida undosquamis, andanother nine in the last four years: Siganusluridus, Fistularia commersonii, Siganusrivulatus, Pempheris vanicolensis, Sargocen-tron rubrum and recently, Lagocephalussceleratus, Etrumeus teres, Upeneus moluc-censis and Pteragogus pelycus (Table 2).These new records are the result of recentmore intensive local investigations, but stillthe number of the species remains low.This phenomenon could be attributed tothe fact that although the continental shelfof Crete is accessible to Lessepsian immi-grants through the Kasos-Karpathos corri-dor, its colonization shows a rather slowprogress mainly because it is very narrowand delimited by exceptionally deepwaters, thus isolating the coastal zone ofCrete.
76 Medit. Mar. Sci., 8/1, 2007, 67-89
Several of the Erythrean fish are stillnot only distributed in the Levant Basin,since the populations of at least nine ofthem already established in the easternMediterranean, both along the Asiatic andAfrican coasts as well as the Greek seas,have expanded westward. According toGOLANI et al. (2002, 2004), in some casesthey have reached the Central Mediter-ranean, such as Siganus luridus, Siganusrivulatus, Stephanolepis diaspros. Siganusluridus has enlarged its distribution to thesouthern coasts of Sicily (AZZURRO &ANDALORO, 2004) and the northern
coasts of Sicily in the Tyrrhenian sea(CASTRIOTA & ANDALORO, 2005),as have also Stephanolepis diaspros, whichhas reached the Palermo coasts(CATALANO & ZAVA, 1993), Sphyrae-na chrysotaenia which has been reportedfrom Malta (LANFRANCO, 1993),Parexocoetus mento and Fistularia com-mersonii from Tunisia (BEN SOUSSI etal., 2004), the last species also from south-ern Sicily (AZZURRO et al., 2004), theSW shore of Sicily (MILAZZO et al.,2006) and Central Tyrrhenian waters(MICARELLI et al., 2006; PAIS et al.,
77Medit. Mar. Sci., 8/1, 2007, 67-89
Species North South Ionian References*Aegean Central Sea
AegeanEtrumeus teres + [1] [2] Fistularia commersonii + + [3] [4] [5] Lagocepalus sceleratus + + [6] [2] [17]Lagocephalus spadiceus + [7] Leiognathus klunzingeri + + [3] Parexocoetus mento + + [3] Pempheris vanicolensis + [5] Saurida undosquamis + + + [8][9] [3] [10] Siganus luridus + + + [9] [3] [5] Siganus rivulatus + + + [11] [3] [5] Stephanolepis diaspros + + [3] [12]Tylosurus crocodilus + [13] Upeneus moluccensis + + + [14] [15] [3] [16]Pteragogus pelycus + [18]Sargocentron rubrum + [19]
Table 2Spreading of Lessepsian fishes in the Aegean and Ionian seas,
out of the Dodecanese continental shelf.
* [1] Cyclades, Crete, KALLIANIOTIS & LEKKAS, 2005; [2] Crete, KASAPIDIS et al., 2007; [3] GOLANI et al., 2006 on-line; [4]Chalkidiki, KARACHLE et al., 2004; [5] Crete, TINGILIS et al., 2003; [6] Izmir, BILECENOGLU et al., 2006; [7] Samos,ANANIADIS, 1952; [8] ZAITSEV & ÖZTÜRK, 2001; [9] ONDRIAS, 1971; [10] Crete, TSIMENIDIS et al., 1991; [11] Izmir,GELDIAY, 1969; [12] Crete, ECONOMIDIS & BAUCHOT, 1976; [13] Thermaikos, SINIS, 2005; [14] Kusadasi, KAYA et al., 1999;[15] Patraikos, KASPIRIS, 1976; [16] various areas, PERISTERAKI et al., 2006; [17] Lesvos, KOUTSOUMBA, D., 2007 (pers.comm.); [18] Crete, STERIOTI, A., 2005 (pers. comm.); [19] Crete, TINGILIS, G., 2003 (pers. comm.).

2007). Some Lessepsian fish arefurthrmore colonizing the coasts of theAdriatic Sea, like Siganus rivulatus,Stephanolepis diaspros, Sphyraena chryso-taenia, Saurida undosquamis, Parexocoetusmento, Leiognathus klunzingeri and Hemi-ramphus far (DUL
v
CIv
C et al., 2003).In conclusion the westward spread of
Lessepsian species will proceed slowly, butmay accelerate if the increase in watertemperature continues. A rearrangementof the ecosystem structure is alreadyappearing in the eastern basin and thiscould be an important factor for fisherymanagement.
Habitat. At present most ErythreanIndo-Pacific alien invertebrates occupythe Mediterranean littoral and infralit-toral zones to a depth of approximately50m, and are hardly ever found in deeperwaters (GALIL & ZENETOS, 2002).This is also true for the Lessepsian fishspecies, which are in the majority coastalspecies dwelling in rather shallow, sandyor muddy habitats (GOLANI, 1993) orrocky shores, often covered by sea-grass.
In Rhodes and other Dodecanesecoastal areas the majority of Erythreanfish are collected by trawlers at depths ofup to 50m, more frequently on sandy-muddy bottoms covered by well-devel-oped Chlorophyceae beds (Caulerpa pro-lifera and Caulerpa racemosa) andPhanerogames prairies, mainly Posidoniaoceanica and Halophila stipulacea(CORSINI & ECONOMIDIS, 1999;CORSINI et al., 2005, 2006). Otherspecies are encountered on sandy-rockybottoms, such as Pempheris vanicolensis,Sargocentron rubrum and schools of juve-nile Shyraena chrysotaenia or in very shal-low sandy areas, as for Iniistius pavo. Onthe other hand, as assessed by CORSINI
et al. (2005), Tylerius spinosissimus wascollected at a lower depth than those it isfound in in its original tropical waters, butdeeper than that usual for Lessepsianmigrant fish in the SE Aegean Sea. Thisprobably shows that there may be severalunexplored niches suitable for the newcolonizers. The first finding of this youngpufferfish in Rhodes, far from the SuezCanal, indicates that a certain populationof the species is already established in theeastern Mediterranean, especially closerto the Suez Canal areas; its spread followsquite unusual paths, as observed in thecase of Tetrosomus gibbosus (SPANIER& GOREN, 1988). However, the possibil-ity that other vectors (ship ballast, aqua-culture or aquaria purposes transport),different from the natural pathway, maybe responsible for the introduction ofsuch Indo-Pacific species as the spinyblaasop Tylerius spinosissimus should notbe undervalued, in common with othertaxa also introduced in this way into Hel-lenic waters (PANCUCCI-PAPADOPOULOU et al., 2006).
Abundance and interactions. The abun-dance of some Lessepsian immigrant fishhas assumed economic importance in thesouth-eastern Levantine and Anatolianfisheries (GUCU et al., 1994; TORCU &MATER, 2000; GOLANI et al., 2002;CICEK & ASVAR, 2003; HARMELIN-VIVIEN et al., 2005), while other Erythtre-an invaders are considered as economicburden by GOREN & GALIL (2005).
Populations of Lessepsian fishthroughout the coastal waters of theDodecanese continental shelf generallycorrespond to species that have been pre-viously established there and they havebeen large enough to be detectable(CORSINI et al., 2005). This is confirmed
78 Medit. Mar. Sci., 8/1, 2007, 67-89

by the fact that such various species havenormally been present in fishery activitiesfrom their first recording in the regionuntil today (CORSINI FOKA et al., 2004).Among the established species listed inTable 1, the majority are caught regularly,but these species cannot be consideredabundant. They are: Hemiramphus far,Stephanolepis diaspros, Upeneus moluccen-sis, Sargocentron rubrum, Sauridaundosquamis, Atherinomorus lacunosus,Pempheris vanicolensis, Pteragogus pelycusand Apogon pharaonis. On the contrary,some other successfully established speciesare common and have acquired a commer-cial importance, i.e. Siganus luridus, S. rivu-latus and Sphyraena chrysotaenia; the latteris normally confused with the two indige-nous species S. sphyhraena and S. viridensiscoexisting in the same coastal area(CORSINI & ECONOMIDIS, 1999), andalso with the new Erythrean colonizer S.flavicauda. The quickly increasing popula-tion of the recent colonizer Etrumeus teresis already contributing to fishery in theDodecanese (CORSINI-FOKA, unpubl.data) and in the Cyclades Islands(KALLIANIOTIS & LEKKAS, 2005), asalso has been observed in Israel, Cyprusand Turkey (GOLANI et al., 2002, CICEK& AVSAR, 2003).
Local indigenous fish, especiallysmall-sized and fry, are subjected to aremarkable predation pressure. This factpresents the other side of the coin, per-haps suggesting that several invaders andnative species are in competition. Conse-quently, the impact on the local exploitedpopulations by Lessepsian immigrantsseems to be serious and accelerated insome cases. Thus a more detailedapproach on this matter is needed.
The permanent establishment of aLessepsian species in the area, and conse-
quently how harmful this would be for thenative fish, is related to its reproductivesuccess. In many cases such a successfulbreeding, in combination with food avail-ability and other favourable environmen-tal factors, leads to a population explo-sion. Such a case of population explosionwas observed in the area during the1940's for Upeneus moluccensis (LASKA-RIDIS, 1948a), followed by a dramaticcrash shortly after, so that nowadays thisspecies is rarely found in the area(CORSINI & ECONOMIDIS, 1999).
Several Erythrean invaders may havefound vacant ecological niches and obvi-ously they do not compete with localnative species. Thus, Siganus luridus andS. rivulatus were well established in theentire Levantine basin initially and theninvaded the Central Mediterranean, dueto the presence of few herbivorous com-petitors and/or abundant available food.Furthermore, Upeneus moluccensis ispresent but it does not dominate Mullusbarbatus. Apogon pharaonis is regularlycaught in trawl nets, as well as its indige-nous counterpart Apogon imberbis, whilePteragogus pelycus occurs with otherlabrid species of similar size (for exampleSymphodus sp., Coris julis, Thalassomapavo). Schools of Sphyraenidae are alsonoted, both belonging to the indigenousspecies Sphyraena sphyraena and S. viri-densis but also to the Lessepsian S. chryso-taenia and S. flavicauda (see above). Sau-rida undosquamis is very rare comparedto Synodus saurus. On the other hand, nolocal fish species disappeared in theRhodes marine area as assessed for therest of the Eastern Mediterranean(GOLANI, 1998b).
Fistularia invasion. In terms ofimpact, the recent invader, blue cornet-
79Medit. Mar. Sci., 8/1, 2007, 67-89

fish, Fistularia commersonii, caught nor-mally by trawl-nets up to 50-60m, hasdeveloped an important population and isconsidered one of the worst invasive fishspecies in the Mediterranean Sea(PANCUCCI-PAPADOPOULOU et al.,2005b; STREFTARIS & ZENETOU,2006). This species presents a very fastexpansion along the coasts of the Levan-tine basin (GOLANI, 2000; CORSINI etal., 2002) and recently up to the northAegean Sea (Chalkidiki peninsula), to thenorthern coasts of Crete and westward tothe Central Mediterranean and Tyrrhen-ian Sea (GOLANI et al., 2006 on-line;Table 2; ICES, 2006; PAIS et al., 2007).The phenomenon is alarming becausethis fish reproduces and grows very rapid-ly, reaching a remarkably large size. Sinceits first appearance in the Rhodes marinearea in 2001, the blue cornetfish occurs ata number of 5 to 20 specimens occasion-ally in a catch of any trawl net operation,mainly at a depth of 20-25 m, but also inthe very shallow waters of beaches andharbours (KALOGIROU et al., 2007).
Additionally, the species presents anextreme feeding activity and a clearaggressive behaviour when in schools. Asa free-swimming carnivore, it is merelydwelling over reefs and sea-grass beds. Asit has no or little commercial value, it issubjected to a very low fishing pressure,being more a ‘by catch’ fish than a target.Consequently, it is free to form large pop-ulations, which seriously affect economi-cally and ecologically important nativespecies not only in the limited area of theSE Aegean Sea, but in all occupied newhabitats, as in the South Aegean Sea(unpublished data). It mainly feeds onfish fry, gobiids and several exploitedpopulations of economically valuablenative fish, namely Spicara smaris, Mullus
spp. and Boops boops (CORSINI et al.,2002; KALOGIROU et al., 2007).Anatomical features of mouth and feed-ing behaviour observed in nature andfood items found in stomach contentreveal that Fistularia commersonii shouldnot be ranked in the highest trophic lev-els. Various sized individuals of thespecies could be seen rather as an easyprey-fish for any energetic predator, if itaccepts and/or appreciates them.
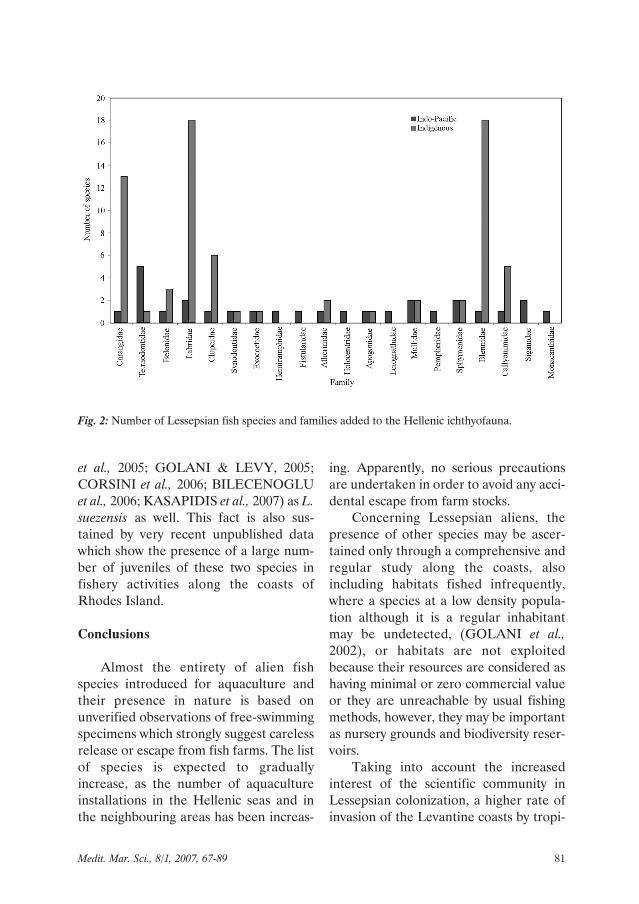
Inventorying Lessepsian migrants.Lessepsian alien fish species occurring inGreek waters belong to 20 families. Thenon-indigenous species have added sevenfamilies to the Hellenic ichthyofauna:Siganidae, Hemiramphidae, Fistularidae,Holocentridae, Leiognathidae, Pem-pheridae, Monacanthidae, while otheraliens increased the species number offive already well-represented families(ECONOMIDIS, 1973; WHITEHEADet al., 1984-1986; BAUCHOT, 1987;PAPAKONSTANTINOU, 1988) (Fig. 2).The remaining aliens increased thespecies number in families represented byone or two species, with the exception ofthe Tetraodontidae, which needs particu-lar attention. Until 1994, only theautochthonous Lagocephalus lago-cephalus, the Lessepsian Lagocephalusspadiceus and Sphoeroides pachygasterfrom the Atlantic were known in the area.A rapid increase of records in the last twoyears added four more Indo-Pacifictetraodontid species: Tylerius spinosis-simus, Lagocephalus suezensis, Lago-cephalus sceleratus and Torquigener flavi-maculosus (Table 1). Among these lastalien tetraodontid colonizers, the popula-tion of the large sized L. sceleratus, whichis not marketable since it may cause foodpoisioning, is increasing rapidly (AKYOL
80 Medit. Mar. Sci., 8/1, 2007, 67-89
et al., 2005; GOLANI & LEVY, 2005;CORSINI et al., 2006; BILECENOGLUet al., 2006; KASAPIDIS et al., 2007) as L.suezensis as well. This fact is also sus-tained by very recent unpublished datawhich show the presence of a large num-ber of juveniles of these two species infishery activities along the coasts ofRhodes Island.
Conclusions
Almost the entirety of alien fishspecies introduced for aquaculture andtheir presence in nature is based onunverified observations of free-swimmingspecimens which strongly suggest carelessrelease or escape from fish farms. The listof species is expected to graduallyincrease, as the number of aquacultureinstallations in the Hellenic seas and inthe neighbouring areas has been increas-
ing. Apparently, no serious precautionsare undertaken in order to avoid any acci-dental escape from farm stocks.
Concerning Lessepsian aliens, thepresence of other species may be ascer-tained only through a comprehensive andregular study along the coasts, alsoincluding habitats fished infrequently,where a species at a low density popula-tion although it is a regular inhabitantmay be undetected, (GOLANI et al.,2002), or habitats are not exploitedbecause their resources are considered ashaving minimal or zero commercial valueor they are unreachable by usual fishingmethods, however, they may be importantas nursery grounds and biodiversity reser-voirs.
Taking into account the increasedinterest of the scientific community inLessepsian colonization, a higher rate ofinvasion of the Levantine coasts by tropi-
81Medit. Mar. Sci., 8/1, 2007, 67-89
Fig. 2: Number of Lessepsian fish species and families added to the Hellenic ichthyofauna.

cal fish species and their successfulacclimatization is evident, confirming that‘the littoral and infralittoral biota of theLevantine Sea is undergoing a profoundchange due to the influx of Erythreaninvaders’ (GALIL & ZENETOS, 2002),and furthermore giving support to theindication of a tropicalization of the area(BIANCHI & MORRI, 2003). In thelong run, this phenomenon will little bylittle increase the complexity of the entireeastern Mediterranean ecosystem andwill most probably lead to a new equilib-rium, which should be characterized by ahigher productivity and stability.
Ackowledgements
The authors are grateful to two anony-mous referees for their constructive com-ments and advice on the manuscript.
References
AKYOL, O., ÜNAL, V., CEYHAN, T. &BILECENOGLU, M., 2005. First con-firmed record of Lagocephalus scelera-tus (Gmelin, 1789) in the Mediter-ranean Sea. Journal of Fish Biology, 66:1183-1186.
AKYOL, O., ÜNAL, V. & CEYHAN, T.,2006. Occurrence of two Lessepsianmigrant fish, Oxyurichthys petersi(Gobiidae) and Upeneus pori (Mulli-dae), from the Aegean Sea. Cybium,30 (4): 389-390.
ANANIADIS, C., 1952. On the appearanceof the fish Tetrodon spadiceus (Rich.) inGreek seas. Praktika of Hellenic Hydro-biological Institute, 6 (1): 73-74.
ANDALORO, F., 2001. La tropicaliz-zazione del Mediterraneo. p. 52-54. In:La biodiversità nella regione bio-geografica mediterranea. Versione inte-
grata del contributo dell’ANPA al rap-porto dell’EEA sulla biodiversità inEuropa. Rome, A.N.P.A., Stato del-l’Ambiente 4/2001.
ANDALORO, F., FALAUTANO, M.,SINOPOLI, M., PASSARELLI, F.M., PIPITONE, C., ADDIS, P., CAU,A. & CASTRIOTA, L., 2005. Thelesser amberjack Seriola fasciata (Per-ciformes: Carangidae) in the Mediter-ranean: A recent colonist? Cybium, 29(2): 141-145.
AZZURRO, E. & ANDALORO, F.,2004. A new settled population of thelessepsian migrant Siganus luridus(Pisces: Siganidae) in Linosa Island-Sicily Strait. Journal of the Marine Bio-logical Association United Kingdom, 84:819-821.
AZZURRO, E., PIZZICORI, F. &ANDALORO, F., 2004. First recordof Fistularia commersonii (Fistulari-dae) from the Central Mediterranean.Cybium, 28: 72-74.
BAUCHOT, M.-L., 1987. Poissons osseux.p. 891-1422. In: Fiches FAO d’identifi-cation des espèces pour les besoins de lapêches. (Révision 1). Méditerranée et MerNoire. Zone de pêche 37. Vol. 2: Vertébrés,edited by W. Fischer, M.-L. Bauchot.and M. Schneider, Rome, FAO.
BEN SOUSSI, J., ZAOUALI, J.,BRADAI, M. N. & QUIGNARD, J.P., 2004. Lessepsian migrant fishes offthe coast of Tunisia. First record ofFistularia commersonii (Osteichthyes,Fistularidae) and Parexocoetus mento(Osteichthyes, Exocoetidae). Vie etMilieu, 54 (4): 247-248.
BEN SOUSSI, J., GOLANI, D., MEJRI,H. & CAPAPE, C., 2005. On theoccurrence of Cheilopogon furcatus inthe Mediterranean Sea. Journal FishBiology, 67: 1144-1149.
82 Medit. Mar. Sci., 8/1, 2007, 67-89

BIANCHI, C. N. & MORRI, C., 2003.Global Sea warming and tropicaliza-tion of the Mediterranean Sea: bio-geographic and ecological aspects.Biogeographia, 24: 319-328.
BILECENOGLU, M., 2004. Occurrenceof the lessepsian migrant fish, Sillagosihama (ForsskaÆl, 17775) (Oste-ichthyes: Sillaginidae) from theAegean Sea. Israel Journal of Zoology,50: 420-421.
BILECENOGLU, M., & KAYA, M.,2006. A new alien fish in the Mediter-ranean Sea – Platax teira (ForsskaÆl,1775) (Osteichthyes: Ephippidae).Aquatic Invasions, 1 (2): 80-83.
BILECENOGLU, M., TASKAVAK, E.,MATER, S. & KAYA, M., 2002.Checklist of the marine fishes ofTurkey. Zootaxa, 113: 1-194.
BILECENOGLU, M., KAYA, M. &AKALIN, S., 2006. Range expansionof silverstripe blaasop, Lagocephalussceleratus (Gmelin, 1789), to thenorthern Aegean Sea. Aquatic Inva-sions, 1 (4): 289-291.
BINI, G., 1960. Catalogue of the names offishes of commercial importance in theMediterranean. FAO, Rome, 250 p.
BRUNELLI, G. & BINI, G., 1934. Sullaimmigrazione di una specie di‘Teuthis‘ del Mar Rosso al Mar Egeo.Atti della Reale Accademia dei Lincei,19: 255-260.
BUHAN, E., YILMAZ, H., MORKAN,Y., BÜKE, Y. & YÜKSEK, A., 1997.A new catch potential for Güllük bayand Gökova bay Scomberomorus com-merson (Lacépède, 1980) (Pisces-Teleostei). Mediterranean FisheriesCongress, Izmir, 9-11 April 1997, 7 p.(Turkish with English abstract).
CASTRIOTA, L. & ANDALORO, F.2005. First record of the lessepsian fish
Siganus luridus (Osteichthyes:Siganidae) in the Tyrrhenian Sea.Journal of the Marine Biological Associ-ation 2 – Biodiversity Records, Pub-lished online.
CATALANO, E. & ZAVA, B., 1993. Sullapresenza di Stephanolepis diasprosFraser Brunner. Doriana, supplementodegli Annali del Museo Civico di StoriaNaturale "G. Doria", 4 (181): 1-4.
CORSINI, M. & ECONOMIDIS, P. S.,1999. Distribution extension of twoLessepsian migrants found in themarine area of the island of Rhodes(Aegean Sea, Greece). Cybium, 23 (2):195-199.
CORSINI, M., KONDILATOS, G. &ECONOMIDIS, P.S., 2002. Lessep-sian migrant Fistularia commersoniifrom the Rhodes marine area. Journalof Fish Biology, 60 (4): 1061-1062.
CORSINI M., MARGIES, P., KONDI-LATOS, G. & ECONOMIDIS, P. S.,2005. Lessepsian migration in AegeanSea: Tylerius spinosissimus (Pisces,Tetraodontidae) new for the Mediter-ranean, and six more fish records fromRhodes. Cybium, 29 (4): 347-354.
CORSINI M., MARGIES, P., KONDI-LATOS, G. & ECONOMIDIS, P. S.,2006. Three new exotic fish recordsfrom the SE Aegean Greek waters.Scientia Marina, 70 (2): 325-329.
CORSINI FOKA, M., KONDYLATOS,G. & ECONOMIDIS, P. S., 2004.Occurrence of the lessepsian speciesPortunus pelagicus (Crustacea) andApogon pharaonis (Pisces) in themarine area of Rhodes Island. Mediter-ranean Marine Science, 5 (1): 5-17.
CORSINI FOKA, M., MARGIES, P.,KONDILATOS, G. & ECONO-MIDIS, P. S., 2006. Torquigener flavi-maculosus Hardy and Randall, 1983
83Medit. Mar. Sci., 8/1, 2007, 67-89

(Pisces: Tetraodontidae) at the RhodesIsland marine area: a new alien fish inthe Hellenic waters. MediterraneanMarine Science, 7/2: 73-76.
CICEK E. & AVSAR, D., 2003. Monthlydistribution and abundance of Lessep-sian fishes off Karatas (NortheasternMediterranean) throughout 2002-2003 fishing season. International Sym-posium of Fisheries and Zoology.Abstract. www.istanbul.edu.tr
CINAR, M. E., BILECENOGLU, M.,ÖZTÜRK, B., KATAGAN, T. &AYSEL, V., 2005. Alien species on thecoasts of Turkey. MediterraneanMarine Science, 6/2: 119-146.
CINAR, M. E., BILECENOGLU, M.,ÖZTÜRK, B. & CAN, A., 2006. Newrecords of alien species on the Levan-tine coast of Turkey. Aquatic Inva-sions, 1 (2): 84-90.
DULv
CIC’ , J. & GOLANI, D., 2006. Firstrecord of Cyclopterus lumpus L., 1758(Osteichthyes: Cyclopteridae) in theMediterranean Sea. Journal of FishBiology, 69: 300-303.
DULv
CIC’ , J., PALLAORO, A., & LIPEJ,L., 2003: Lessepsian fish migrantsreported in the eastern Adriatic Sea:an annotated list. Annals for Istrianand Mediterranean Studies Series Histo-ria Naturalis, 13: 137–144.
ECONOMIDIS, P. S., 1973. Cataloguedes poissons de la Grèce. HellenicOceanology and Limnology, 11: 421-598 (in Greek).
ECONOMIDIS, P. S. & BAUCHOT, M.-L., 1976. Sur une collection de pois-sons des mers helléniques (mers Egéeet Ionienne) déposée au MuséumNational d’Histoire Naturelle. Bulletindu Muséum National d’HistoireNaturelle, Paris, 3e sér., no 392, Zoolo-gie, 274: 871-903.
ECONOMIDIS, P. S., KALLIANIOTIS,A. & PSALTOPOULOU, H., 1999.Sea Lamprey, Petromyzon marinus(Cyclostomata, Petromyzontidae), inthe eastern Mediterranean-tworecords from the Aegean Sea(Greece). Journal of Fish Biology, 55:1114-1118.
ECONOMIDIS, P. S., KOUTRAKIS, E.TH. & BOBORI, D. C., 2000. Distrib-ution and conservation of Acipensersturio and related species in the Greekwaters. Boletin Instituto Espanol deOceanografia, 16(1-4): 81-88.
ERYLMAZ, L. & DALYAN, C., 2006.First record of Apogon quekettiGilchrist (Osteichthyes: Apogonidae)in the Mediterranean Sea. Journal ofFish Biology, 69: 1251-1254.
ERYILMAZ, L., ÖZULUG, M. &MERIC, N., 2003. The Smooth Puf-ferfish, Sphoeroides pachygaster(Müller and Troschel, 1848)(Teleostei: Tetraodontidae), new tothe Northern Aegean Sea. Zoology inthe Middle East, 28: 125-126.
FROESE, R. AND PAULY, D. (Eds.),2006. FishBase. World Wide Webelectronic publication. www.fishbase.org,version (12/2006).
GALIL, B. S. & KEVREKIDIS, K., 2002.Exotic decapods and a stomatopod offRhodes Island (Greece) and the east-ern Mediterranean Transient. Crus-taceana, 75: 925-930.
GALIL, B. S. & ZENETOS, A., 2002. ASea change-Exotics in the EasternMediterranean Sea. p. 325-336.In: Invasive Aquatic Species ofEurope. Distribution, Impacts andManagement, E. Leppäkoski, GollaschS. & S. Olenin (Eds). Netherlands:Kluwer Academic publishers.www.biomareweb.org
84 Medit. Mar. Sci., 8/1, 2007, 67-89

GELDIAY, R., 1969. The fishes of theIzmir Bay and the possibility of theirinvasions. Ege Universitesi fe Fak.Genel. Zooloji Kursusu, Izmir, p. 135.
GOKOGLU, M., BODUR, T. & KAYA,Y., 2003. First record of the bonner-fish (Heniochus intermedius Stein-dachner, 1893) from the Mediter-ranean Sea. Israel Journal fo Zoology,49: 324-325.
GOLANI, D., 1993. The sandy shore of theRed Sea-launching pad for Lessepsian(Suez Canal) migrant fish colonizers ofthe eastern Mediterranean. Journal ofBiogeography, 20: 579-585.
GOLANI, D., 1998a. Distribution ofLessepsian migrant fish in theMediterranean. Italian Journal ofZoology, 65, Suppl.: 95-99.
GOLANI, D., 1998b. Impact of Red SeaFish Migrants through the Suez Canalon the Aquatic Environment of theEastern Mediterranean. Bulletin YaleUniversity School of Forestry and Envi-ronmental Studies, 103: 375-387.
GOLANI, D., 2000. First record of theBluespotted cornetfish from theMediterranean Sea. Journal of FishBiology, 56: 1545-1547.
GOLANI, D., 2006. The Indian scad(Decapterus russelli), (Osteichthyes:Carangidae), a new Indo-Pacific fishinvader of the eastern Mediterranean.Scientia Marina, 70 (4): 603-605.
GOLANI, D. & LEVY, Y., 2005. New re-cords and rare occurrences of fish speciesfrom the Mediterranean coast of Israel.Zoology in the Middle East, 36: 27-32.
GOLANI, D. & SONIN, O., 2006. TheJapanese threadfin bream Nemipterusjaponicus, a new Indo-Pacific fish inthe Mediterranean Sea. Journal of FishBiology, 68: 940-943.
GOLANI, D., ORSI-RELINI, D., L.,
MASSUTI, E. & QUIGNARD, J.- P.,2002. CIESM Atlas of Exotic speciesin the Mediterranean Volume 1: Fish-es. F. Briand (Ed.), Monaco. CIESMPublishers, 256 pp.
GOLANI, D., ORSI-RELINI, L.,MASSUTI, E. & QUIGNARD, J.- P.,2004. Dynamics of fish invasions inthe Mediterranean: update of theCIESM Fish Atlas. Rapport Commis-sion International Mer Méditerranée,37: 367.
GOLANI, D., ORSI-RELINI, L.,MASSUTI, E. & QUIGNARD, J.- P.,2006 on-line. CIESM Atlas of Exoticfishes in the Mediterranean.http://www.ciesm.org/atlas/, visited inDecember 2006.
GOREN, M. & GALIL, B. S., 2005. Areview of changes in the fish assem-blages of Levantine inland and marineecosystems following the introductionof non-native fishes. Journal ofApplied Ichthyology, 21: 364-370.
GUCU, A. C., BINGEL, F., AVSAR, D.& UYSAL, N., 1994. Distribution andoccurrence of Red Sea fish at theTurkish Mediterranean coast-north-ern Cilician Basin. Acta Adriatica, 34:103-113.
HARMELIN-VIVIEN, M., BITAR, G.,HARMELIN, J.-G. & MONESTIEZ,P., 2005. The littoral fish communityof the Lebanese rocky coast (easternMediterranean Sea) with emphasis onRed Sea immigrants. Biological Inva-sions, 7: 625-637.
HARRISON, I., 2004. Liza haematocheila(Temminck and Schlegel, 1845).Addentum to vol. 8, part I, Mugilidae.In: The Freshwater Fishes of Europe, P.J. Miller (Eds), vol. 8/II. Gobiidae 2,pp. 469-471.
HUNTER, M. L. & GIBBS, P., 2006. Fun-
85Medit. Mar. Sci., 8/1, 2007, 67-89

damentals of Conservation Biology.Blackwell publishing, p. 516.
ICES, 2006. Working Group on Introduc-tions and Transfers of Marine Organ-isms (WGITMO), 16–17 March 2006,Oostende, Belgium. ICES CM2006/ACME:05. p.334.
KALLIANIOTIS, A. & LEKKAS, V.,2005. First documented report on theLessepsian migrant Etrumeus teres DeKay, 1842 (Pisces: Clupeidae) inGreek Seas. Journal of BiologicalResearch, 4: 225-229.
KALOGIROU S., CORSINI, M.,KONDILATOS, G. &WENNHAGE, H., 2007. Diet of theinvasive piscivorous fish Fistulariacommersonii in a recently colonizedarea of the eastern Mediterranean.Biological Invasions, 9: 887-896.
KARACHLÉ, P. K., TRIANTA-PHYLLIDIS, C. & STERGIOU, K. I.,2004. Fistularia commersonii Rüppell,1838: a lessepsian sprinter. Acta Ichthy-ologica et Piscatoria, 34 (1): 103-108.
KASAPIDIS, P., PERISTERAKI, P.,TSERPES, G. & MAGOULAS, A.,2007. First record of the Lessepsianmigrant Lagocephalus sceleratus(Gmelin, 1789) (Osteichthyes:Tetraodontidae) in the Cretan Sea(Aegean, Greece). Aquatic Invasions,(2) 1: 71-73.
KASPIRIS, P., 1976. New fish recordsfrom the Greek part of the Ionian Sea.Revue des Travaux de l’ Institut desPêches Maritimes, 40: 627-628.
KAVALAKIS, G., 1968. Siganus luridus andSiganus rivulatus in the DodecaneseIslands. Aliia, 248: 307-308 (In Greek).
KAYA, M., MATER, S. & KORKUT,A.Y., 1998. A new grey mullet species‘Mugil so-iuy Basilewsky’ (Teleostei:Mugilidae) from the Aegean coast of
Turkey. Turkish Journal of Zoology, 22(4): 303-306.
KAYA, M., BENLI, H. A., KATAGAN,T. & OZAYDIN, O., 1999. Age,growth, sex-ratio, spawning season andfood of golden banded goatfish,Upeneus moluccensis Bleeker (1855)from the Mediterranean and southAegean coasts of Turkey. FisheriesResearch, 41: 317-328.
KOSSWIG, C., 1950. Erythräische Fischeim Mittelmeer und an der Grenze derAgäis. Syllegomena Biologica.Festschrift Kleinschmidt, p. 203-212.Leipzig Akademie Verlag.
KOUTRAKIS, E. T. & ECONOMIDIS,P. S., 2000. First record in the Mediter-ranean (north Aegean Sea, Greece) ofthe Pacific mullet Mugil soiuyBasilewsky, 1855 (Mugilidae). Cybium,24 (3): 299-302.
KOUTRAKIS E. T. & ECONOMIDIS, P.S., 2006. Did sturgeons return to theRiver Evros? Alieftika Nea, 306: 68-83(in Greek).
LANFRANCO, G. G., 1993. The fishesaround Malta (Central Mediterranean).Progress Press Co., Malta, p. 132.
LASKARIDIS, K., 1948a. Holocentrumrubrum (Forsk) and Lagocephalus lago-cephalus (L.), two newly reported mem-bers of the Greek fish fauna (Dode-canesian islands). Praktika HellenicHydrobiological Institute, 2 (1): 127-129.
LASKARIDIS, K., 1948b. Contribution tothe biology of the fish Mulloides auri-flamma (Forsk.). Praktika HellenicHydrobiological Institute, 2 (1): 103-118.
MICARELLI, P., BARLETTANI, M. &CECCARELLI, R., 2006. First recordof Fistularia commersonii (Fistulari-idae, Pisces) in the North TyrrhenianSea. Biologia Marina Mediterranea,13(1):887-889.
86 Medit. Mar. Sci., 8/1, 2007, 67-89

MILAZZO, A., BARRACO, B.,BELTRAME, A.M., CANNIZZARO,L., PULEO, M., RIZZO, P., SALVO,G. & VITALE, S., 2006. Capture ofFistularia commersonii, Rüppel, 1839(Pisces, Fistularidae) in the coastalwaters of Selinunte (S-W Sicily).Biologia Marina Mediterranea, 13(1):890-891.
MINOS, G., IMSIRIDOU, A &ECONOMIDIS, P. S., in press. Lizahaematocheilus (Pisces, Mugilidae) inNorthern Aegean Sea. Fish Invasionsof the Mediterrranean Sea.
MINOS, G., A. IMSIRIDOU & P. S.ECONOMIDIS, 2007. Liza haema-tochilus (Pisces, Mugilidae) in North-ern Aegean Sea. New records andgonad structure. XII EuropeanCongress of Ichthyology in Cavtat(Dubrovnik), Croatia 9–13 September2007 Book of Abstacts, p.243.
ONDRIAS, I. C., 1971. A list of the freshand sea water fishes of Greece. HellenicOceanology and Limnology, 10: 23-96.
PAIS, A., MERELLA, P., FOLLESA, M.C. & GARIPPA, G., 2007. Westwardrange expansion of the Lessepsianmigrant Fistularia commersonii (Fistu-lariidae) in the Mediterranean Sea,with notes on its parasites Journal ofFish Biology, 70 (1): 269-277.
PANCUCCI-PAPADOPOULOU, M. A.,KEVREKIDIS, K., CORSINI-FOKA, M. & SIMBOURA, N., 2005a.Changes in species: invasion of exoticspecies. p. 336-342. In: State of the Hel-lenic Marine Environment, E. Pap-athanassiou & A. Zenetos (Eds),Athens, HCMR Publications.
PANCUCCI-PAPADOPOULOU, M. A.,ZENETOS, A., CORSINI-FOKA,M., & POLITOU, CH., 2005b.Update of marine alien species in Hel-
lenic waters. Mediterranean MarineScience, 6/2: 147-158
PANCUCCI-PAPADOPOULOU, M. A.,ZENETOS, A. & CORSINI-FOKA,M., 2006. Aliens in Hellenic Seas:emphasis on introductions in ports.Communication presented atICES/IOC/IMO Working Group onballast and other ship vectors. Oos-tende, Belgium, 13-15 March, 2006.
PAPACONSTANTINOU, C., 1987.Distribution of the Lessepsian fishmigrants in the Aegean Sea. BiologiaGallo-Hellenica, 13: 15-20.
PAPACONSTANTINOU, C., 1990. Thespreading of Lessepsian fish migrantsinto the Aegean Sea (Greece). ScientiaMarina, 54 (4): 313-316.
PAPACONSTANTINOU, C. &CARAGITSOU, E., 1987. The firstoccurrence of the lessepsian fish immi-grant Pempheris vanicolensis in Greekterritorial waters. Thalassographica, 9(2): 78-79.
PAPAKONSTANTINOU K., 1988.Check-List of Marine Fishes of Greece.FAUNA GRAECIAE IV, NationalCenter for Marine Research, HellenicZoological Society, Athens, p. 257.
PERISTERAKI, P., LAZARAKIS, G.,SKARVELIS, C., GEORGIADIS, M.& TSERPES, G., 2006. Additionalrecords on the occurrence of alien fishspecies in the eastern MediterraneanSea. Mediterranean Marine Science,7/2: 61-66.
POR, F. D., 1969. The Canuellidae (Cope-poda, Harpacticoida) in the watersaround the Sinai Peninsula and theproblem of ‘Lessepsian’ migration ofthis family. Israel Journal of Zoology,18: 169-178.
QUIGNARD, J. P. & PRAS, A., 1986.Atherinidae. p. 1207-1210. In: Fishes
87Medit. Mar. Sci., 8/1, 2007, 67-89

of the North-eastern Atlantic and theMediterranean, P. J. P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielsen &E. Tortonese (Eds). Vol. 3. Paris,UNESCO.
QUIGNARD, J. P. & TOMASINI, J. A.,2000. Mediterranean fish biodiversity.Biologia Marina Mediterranea, 7(3): 1-66.
REINA-HERVA’ S, J. A., GARCIARASO, J. E. & MANJO’ N-CABEZA,M. E., 2004. First record of Spho-eroides spengleri (Osteichthyes:Tetraodontidae) in the Mediter-ranean Sea. Journal of the Marine Bio-logical Association United Kingdom,84: 1089-1090.
SERBETIS, CH., 1947. The appearance ofa new fish (Mulloides flavolineatus) inthe Mediterranean Sea in theDodecanese area. Praktika AcademyAthens, 22: 209.
SINIS, A. I., 2005. First record of Tylosu-rus crocodilus (Péron & Lesueur1821) (Pisces: Belonidae) in theMediterranean (North Aegean Sea,Greece). Journal of BiologicalResearch, 4: 221-224.
SPANIER, E. & GOREN, M., 1988. AnIndo-Pacific trunkfish Tetrosomus gibbo-sus (Linnaeus): first record of the familyOstracionidae in the Mediterranean.Journal of Fish Biology, 32: 797-798.
STARUSHENKO L. I. & KAZANSKY,A. B., 1996. Introduction of mullethaarder (Mugil so-iuy Basilewsky) intothe Black Sea and the sea of Azov.Studies and Reviews, General FisheriesCouncil for the Mediterranean. No 67.Rome, FAO. p. 29.
STREFTARIS, N., ZENETOS, A. &PAPATHANASSIOU, E., 2005.Globalisation in marine ecosystems:the story of non-indigenous marinespecies across European seas.
Oceanography and Marine Biology: AnAnnual Review, 43: 419-453.
STREFTARIS, N. & ZENETOU, A.,2006. Alien marine species in theMediterranean – the 100 ‘Worst inva-sives’ and their impact. MediterraneanMarine Science, 7/1: 87-118.
TINGILIS, G., ANEZAKI, A., VALLA,E., LYMBERAKIS, P.,KAPATANGAKIS, A., BIYIAKIS, S.& KAOURI, S., 2003. First records ofLessepsian fishfauna migrants fromthe Crete marine area. Proceedings ofthe 11th Hellenic Ichthyologists Sympo-sium, Preveza, 10-13 April 2003, 47-50.
TORCU, H. & MATER, S., 2000. Lessep-sian fishes spreading along the coastsof the Mediterranean and the South-ern Aegean Sea of Turkey. TurkishJournal of Zoology, 24: 139-148.
TORTONESE, E., 1938. L’ ittiofaunamediterranea in rapporto alla zoo-geografia. Bollettino del Museo diZoologia e Anatomia Comparata diTorino, 46: 1-35.
TORTONESE, E., 1946. On some fishesfrom the Eastern Mediterranean(Island of Rhodes). Annals and Ma-gazine of Natural History, 13: 710-715.
TORTONESE, E., 1947a. Biologicalinvestigations in the Aegean Sea.Nature, 159: 887-888.
TORTONESE, E., 1947b. Ricerche zoo-logiche nell’isola di Rodi (Mar Egeo).Pesci. Bollettino di Pesca Piscicoltura eIdrobiologia, 23 (2): 143-192.
TORTONESE, E., 1960. General charac-ters of the Mediterranean fish fauna.Idrobiologi BV (1-2): 43-50.
TORTONESE, E., 1963. Elenco rivedutodei Leptocardi, Ciclostomi, Pesci Car-tilaginei e Ossei del Mar Mediterra-neo. Annali del Museo di Storia Natu-rale di Genova, 74: 156-185.
88 Medit. Mar. Sci., 8/1, 2007, 67-89

89
TORTONESE, E., 1964. The main bio-geographical features and problems ofthe Mediterranean fish fauna. Copeia,1964: 98-107.
TORTONESE, E., 1967. Citeva note com-parative privind pestii din Mediter-rana ci din Marea Neagra. BuletinulInstitutului de Cercetari Si ProiectariPiscicole, 26(4): 37-55.
TORTONESE, E., 1970. Osteichthyes(Pesci Ossei). Parte prima. In: Faunad’Italia. Bologna, Ed. Calderini, p. 565.
TORTONESE, E., 1975. Osteichthyes(Parte seconda). In: Fauna d’Italia.Bologna, Ed. Calderini. p. 636.
TORTONESE, E., 1982. I pesci a dis-tribuzione circumtropicale presentinel Mediterraneo. Memorie BiologiaMarina e Oceanografia, 12: 191-203.
TSIMENIDIS, N., TSERPES, G.,MACHIAS A. & KALLIANIOTIS,A., 1991. Distribution of fishes on theCretan shelf. Journal of Fish Biology,39: 661-672.
WHITEHEAD, P.J.P., BAUCHOT, M.-L.,HUREAU, J.-C., NIELSEN, J. ANDTORTONESE, E. (Eds.), 1984-1986.Fishes of the North-eastern Atlanticand the Mediterranean (FNAM) Vol. I,II, III, UNESCO, 1473 pp.
ZACHARIOU-MAMALINGA, ∂., 1999.The occurrence of the Atlantic fishGaidropsarus granti in the Mediter-ranean Sea (South-eastern AegeanSea, Dodecanese, Greece, EasternMediterranean). Annales MuseiGoulandris, 10: 261-266.
ZACHARIOU-MAMALINGA, H. &CORSINI, M., 1994. The occurrenceof the fish Sphoeroides pachygaster inthe south-eastern Aegean Sea(Greece). Annales Musei Goulandris, 9:479-483.
ZAITSEV, Y. & ÖZTÜRK, B. (Eds.),2001. Exotic species in the Aegean, Mar-mara, Black, Azov and Caspian Seas.Published by Turkish MarineResearch Foundation, Istanbul,Turkey, 265 pp.
ZENETOS, A., CINAR, M.E.,PANCUCCI- PAPDOPOULOU, M.A., HARMELIN, J. G., FURNARI,G., ANDALORO, F., BELLOU, N.,STREFTARIS, N. & ZIBROWIUS,H., 2005. Annotated list of marinealien species in the Mediterraneanwith records of the worst invasivespecies. Mediterranean Marine Science,6/2: 63-118.
Medit. Mar. Sci., 8/1, 2007, 67-89
Accepted in 2007





























