Embed Size (px)
Citation preview

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 1/52

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 2/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Animal Learning &Cognition

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 3/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 4/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Animal Learning &Cognition,An Introduction
Third Edition
John M. Pearce
Cardiff University

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 5/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Published in 2008by Psychology Press27 Church Road, Hove, East Sussex, BN3 2FA
Simultaneously published in the USA and Canadaby Psychology Press270 Madison Ave, New York, NY 10016
www.psypress.com
Psychology Press is an imprint of the Taylor & Francis Group,an informa business
© 2008 Psychology Press
All rights reserved. No part of this book may be reprinted or reproduced or utilized inany form or by any electronic, mechanical, or other means, now known or hereafterinvented, including photocopying and recording, or in any information storage orretrieval system, without permission in writing from the publishers.
The publisher makes no representation, express or implied, with regard to theaccuracy of the information contained in this book and cannot accept any legalresponsibility or liability for any errors or omissions that may be made.
British Library Cataloguing in Publication DataA catalogue record for this book is available from the British Library
Library of Congress Cataloging-in-Publication DataPearce, John M.
Animal learning and cognition: an introduction / John M. Pearce.
p. cm.Includes bibliographical references and index.ISBN 978–1–84169–655–3—ISBN 978–1–84169–656–01. Animal intelligence. I. Title.
QL785.P32 2008591.5'13—dc22 2007034019
ISBN: 978–1–84169–655–3 (hbk)ISBN: 978–1–84169–656–0 (pbk)
Typeset by Newgen Imaging Systems (P) Ltd, Chennai, IndiaPrinted and bound in Slovenia

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 6/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
For Victoria

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 7/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 8/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Contents
Preface ix
1 The study of animal
intelligence 2
The distribution of intelligence 4
Defining animal intelligence 12
Why study animal intelligence? 16
Methods for studying animal
intelligence 20
Historical background 22
2 Associative learning 34
Conditioning techniques 36
The nature of associative learning 42
Stimulus–stimulus learning 49
The nature of US representations 52
The conditioned response 55
Concluding comment: the reflexive
nature of the conditioned
response 60
3 The conditions for learning:
Surprise and attention 62
Part 1: Surprise and conditioning 64
Conditioning with a single CS 64
Conditioning with a compound CS 68
Evaluation of the Rescorla–Wagner
model 72
Part 2: Attention and conditioning 74
Wagner’s theory 76
Stimulus significance 80
The Pearce–Hall theory 86
Concluding comments 91
4 Instrumental conditioning 92
The nature of instrumental
learning 93
The conditions of learning 97
The performance of instrumental
behavior 106The Law of Effect and
problem solving 111
5 Extinction 122
Extinction as generalization
decrement 123
The conditions for extinction 125
Associative changes during
extinction 134
Are trials important for Pavlovian
extinction? 142
6 Discrimination learning 148
Theories of discrimination learning 149
Connectionist models of
discrimination learning 161
Metacognition and discrimination
learning 166
7 Category formation 170
Examples of categorization 171
Theories of categorization 173
Abstract categories 179
Relationships as categories 180
The representation of knowledge 188
8 Short-term retention 190
Methods of study 191
Forgetting 199
Theoretical interpretation 202
Serial position effects 206
Metamemory 207
9 Long-term retention 212
Capacity 214
Durability 215

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 9/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
viii Contents
Theoretical interpretation 218
Episodic memory 225
10 Time, number, and
serial order 232
Time 233Number 243
Serial order 253
Transitive inference 259
Concluding comments 262
11 Navigation 264
Part 1: Short-distance travel 265
Methods of navigation 265
Part 2: Long-distance travel 283
Navigational cues 284
Homing 286
Migration 289
Concluding comments 293
12 Social learning 296
Diet selection and foraging 298
Choosing a mate 301
Fear of predators 301
Copying behavior: mimicry 302
Copying behavior: imitation 304
Theory of mind 312
Self-recognition 319
Concluding comments 324
13 Animal communication andlanguage 326
Animal communication 327
Communication and language 336
Can an ape create
a sentence? 339
Language training with other
species 350
The requirements for learning
a language 356
14 The distribution of
intelligence 360
Intelligence and brain size 361
The null hypothesis 364
Intelligence and evolution 369
References 373
Author index 403
Subject index 411

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 10/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Preface
In preparing the third edition of this book, my aim, as it was for the previous editions,
has been to provide an overview of what has been learned by pursuing one particular
approach to the study of animal intelligence. It is my belief that the intelligence of
animals is the product of a number of mental processes. I think the best way of
understanding these processes is by studying the behavior of animals in an
experimental setting. This book, therefore, presents what is known about animal
intelligence by considering experimental findings from the laboratory and from more
naturalistic settings.
I do not attach any great importance to the distinction between animal learning
and animal cognition. Research in both areas has the common goal of elucidating the
mechanisms of animal intelligence and, very often, this research is conducted using
similar procedures. If there is any significance to the distinction, then it is that thefields of animal learning and animal cognition are concerned with different aspects
of intelligence. Chapters 2 to 6 are concerned predominantly with issues that fall
under the traditional heading of animal learning theory. My main concern in these
chapters is to show how it is possible with a few simple principles of associative
learning to explain a surprisingly wide range of experimental findings. Readers
familiar with the previous edition will notice that apart from a new chapter devoted
to extinction, there are relatively few changes to this part of the book. This lack of
change does not mean that researchers are no longer actively investigating the basic
learning processes in animals. Rather, it means that the fundamental principles of

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 11/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
x Preface
learning are now reasonably well established and that current research is directed
towards issues that are too advanced to be considered in an introductory text book.
The second half of the book covers material that is generally treated under the
heading of animal cognition. My overall aim in these chapters is to examine what has
been learned from studying animal behavior about such topics as memory, the
representation of knowledge, navigation, social learning, communication, and
language. I also hope to show that the principles developed in the earlier chapters areof relevance to understanding research that is reviewed in the later chapters. It is in
this part of the book that the most changes have been made. Research on animal
cognition during the last 10 years has headed in many new directions. I have tried to
present a clear summary of this research, as well as a balanced evaluation of its
theoretical implications.
Those who wish to study the intelligence of animals face a daunting task. Not
only are there numerous different species to study, but there is also an array of
intellectual skills to be explored, each posing a unique set of challenging theoretical
problems. As a result, many of the topics that I discuss are still in their infancy. Some
readers may therefore be disappointed to discover that we are still trying to answer
many of the interesting questions that can be asked about the intelligence of animals.On the other hand, it is just this lack of knowledge that makes the study of animal
learning and cognition so exciting. Many fascinating discoveries remain to be made
once the appropriate experiments have been conducted.
One of the rewards for writing a book is the opportunity it provides to thank the
many friends and colleagues who have been so generous with the help they have
given me. The way in which this book is organized and much of the material it
contains have been greatly influenced by numerous discussions with A. Dickinson,
G. Hall, N. J. Mackintosh, and E. M. Macphail. Different chapters have benefited
greatly from the critical comments on earlier versions by A. Aydin, N. Clayton,
M. Haselgrove, C. Heyes, V. LoLordo, A. McGregor, E. Redhead, and P. Wilson.
A special word of thanks is due to Dave Lieberman, whose thoughtful comments onan earlier draft of the present edition identified numerous errors and helped to clarify
the manner in which much of the material is presented. The present edition has
also greatly benefited from the detailed comments on the two previous editions by
N. J. Mackintosh.
I should also like to express my gratitude to the staff at Psychology Press.
Without the cajoling and encouragement of the Assistant Editor, Tara Stebnicky, it is
unlikely that I would have embarked on this revision. I am particularly grateful to the
Production Editor, Veronica Lyons, who, with generous amounts of enthusiasm and
imagination, has done a wonderful job in trying to transform a sow’s ear into a silk
purse. Thanks are also due to the colleagues who were kind enough to send me
photographs of their subjects while they were being tested. Finally, there is the
pleasure of expressing gratitude to Victoria, my wife, who once again patiently
tolerated the demands made on her while this edition was being prepared. In previous
editions I offered similar thanks to my children, but there is no need on this occasion
now that they have left home. Even so, Jess, Alex, and Tim would never forgive me
if I neglected to mention their names.
While preparing for this revision I read a little about Darwin’s visit to the
Galapagos Islands. I was so intrigued by the influence they had on him that I felt
compelled to visit the islands myself. During the final stages of preparing this
edition, Veronica and Tania, somewhat reluctantly, allowed me a two-week break to
travel to the Galapagos Islands. The holiday was one of the highlights of my life.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 12/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Preface xi
The sheer number of animals, and their absolute indifference to the presence of
humans, was overwhelming. The picture on the previous page shows me trying
unsuccessfully to engage a giant tortoise in conversation. This, and many other
photographs, were taken without any elaborate equipment and thus reveal how the
animals allowed me to approach as close as I wished in order to photograph them.
I came away from the islands having discovered little that is new about the
intelligence of animals, but with a deeper appreciation of how the environmentshapes not only their form, but also their behavior.
John M. Pearce
October, 2007

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 13/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
C
H A
P T E
R
4C O N T E N T S
The nature of instrumental learning 93
The conditions of learning 97
The performance of instrumental behavior 106
The Law of Effect and problem solving 111

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 14/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4Instrumentalconditioning
Behavior is affected by its consequences. Responses that lead to reward are repeated,
whereas those that lead to punishment are withheld. Instrumental conditioning refers
to the method of using reward and punishment in order to modify an animal’s
behavior. The first laboratory demonstration of instrumental conditioning was
provided by Thorndike (1898) who, as we saw in Chapter l, trained cats to make a
response in order to escape from a puzzle box and earn a small amount of fish. Since
this pioneering work, there have been many thousands of successful demonstrations
of instrumental conditioning, employing a wide range of species, and a variety of
experimental designs. Skinner, for example, taught two pigeons, by means of
instrumental conditioning, to play ping-pong with each other.From the point of view of understanding the mechanisms of animal intelligence,
three important issues are raised by a successful demonstration of instrumental
conditioning. We need to know what information an animal acquires as a result of its
training. Pavlovian conditioning was shown to promote the growth of
stimulus–stimulus associations, but what sort of associations develop when a response
is followed by a reward or punishment? Once the nature of the associations formed
during instrumental conditioning has been identified, we then need to specify the
conditions that promote their growth. Surprise, for example, is important for successful
Pavlovian conditioning, but what are the necessary ingredients to ensure the success of
instrumental conditioning? Finally, we need to understand the factors that determine
when, and how vigorously, an instrumental response will be performed.Before turning to a detailed discussion of these issues, we must be clear what is
meant by the term reinforcer. This term refers to the events that result in the
strengthening of an instrumental response. The events are classified as either positive
reinforcers, when they consist of the delivery of a stimulus, or negative reinforcers,
when it involves the removal of a stimulus.
THE NATURE OF INSTRUMENTAL LEARN ING
Historical background
Thorndike (1898) was the first to propose that instrumental conditioning is based on
learning about responses. According to his Law of Effect, when a response is
followed by a reinforcer, then a stimulus–response (S–R) connection is strengthened.
In the case of a rat that must press a lever for food, the stimulus might be the lever
itself and the response would be the action of pressing the lever. Each successful
lever press would thus serve to strengthen a connection between the sight of the lever
and the response of pressing it. As a result, whenever the rat came across the lever in
the future, it would be likely to press it and thus gain reward. This analysis of
instrumental conditioning has formed the basis of a number of extremely influential
theories of learning (e.g. Hull, 1943).
K E Y T E R M
Reinforcer
An event thatincreases theprobability of aresponse when
presented after it.If the event is theoccurrence of astimulus, such asfood, it is referredto as a positivereinforcer; but if theevent is the removalof a stimulus, such asshock, it is referredto as a negativereinforcer.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 15/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
94 A N I M A L L E A R N I N G & C O G N I T I O N
A feature of the Law of Effect that has proved unacceptable to the intuitions of
many psychologists is that it fails to allow the animal to anticipate the goal for which
it is responding. The only knowledge that an S–R connection permits an animal to
possess is the knowledge that it must make a particular response in the presence of a
given stimulus. The delivery of food after the response will, according to the Law of
Effect, effectively come as a complete surprise to the animal. In addition to sounding
implausible, this proposal has for many years conflicted with a variety of experimental findings.
One early finding is reported by Tinkelpaugh (1928), who required monkeys to
select one of two food wells to obtain reward. On some trials the reward was a
banana, which was greatly preferred to the other reward, a lettuce leaf. Once the
animals had been trained they were occasionally presented with a lettuce leaf when
they should have received a banana. The following quote, which is cited in
Mackintosh (1974), provides a clear indication that the monkey expected a more
attractive reward for making the correct response (Tinkelpaugh, 1928, p. 224):
She extends her hand to seize the food. But her hand drops to the floor without
touching it. She looks at the lettuce but (unless very hungry) does not touch it.
She looks around the cup and behind the board. She stands up and looks under
and around her. She picks the cup up and examines it thoroughly inside and out.
She had on occasion turned toward the observers present in the room and
shrieked at them in apparent anger.
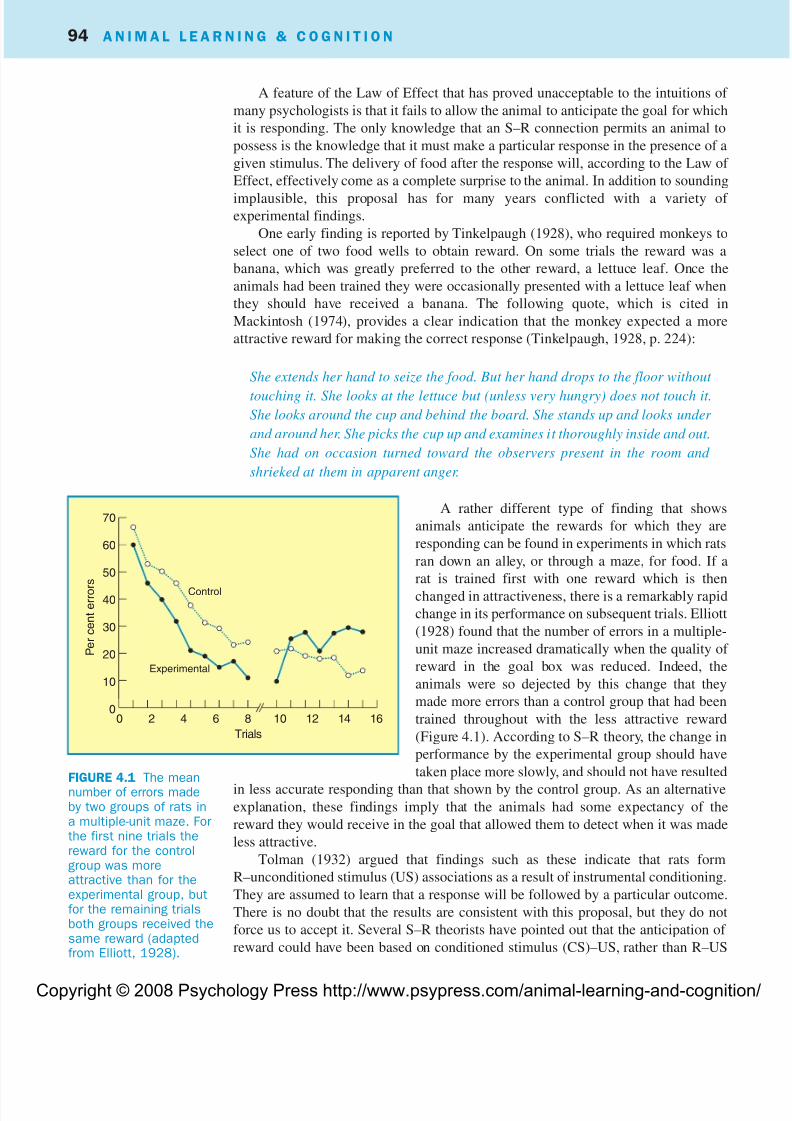
A rather different type of finding that shows
animals anticipate the rewards for which they are
responding can be found in experiments in which rats
ran down an alley, or through a maze, for food. If a
rat is trained first with one reward which is then
changed in attractiveness, there is a remarkably rapid
change in its performance on subsequent trials. Elliott
(1928) found that the number of errors in a multiple-
unit maze increased dramatically when the quality of
reward in the goal box was reduced. Indeed, the
animals were so dejected by this change that they
made more errors than a control group that had been
trained throughout with the less attractive reward
(Figure 4.1). According to S–R theory, the change in
performance by the experimental group should have
taken place more slowly, and should not have resulted
in less accurate responding than that shown by the control group. As an alternativeexplanation, these findings imply that the animals had some expectancy of the
reward they would receive in the goal that allowed them to detect when it was made
less attractive.
Tolman (1932) argued that findings such as these indicate that rats form
R–unconditioned stimulus (US) associations as a result of instrumental conditioning.
They are assumed to learn that a response will be followed by a particular outcome.
There is no doubt that the results are consistent with this proposal, but they do not
force us to accept it. Several S–R theorists have pointed out that the anticipation of
reward could have been based on conditioned stimulus (CS)–US, rather than R–US
FIGURE 4.1 The meannumber of errors madeby two groups of rats ina multiple-unit maze. Forthe first nine trials thereward for the controlgroup was moreattractive than for theexperimental group, butfor the remaining trialsboth groups received thesame reward (adaptedfrom Elliott, 1928).
70
60
50
40
30
20
10
00 2 4 6 8 10 12 14 16
Trials
P e r c e n t e r r o r s
Experimental
Control

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 16/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 95
associations. In Elliott’s (1928) experiment, for example, the animal consumed the
reward in the goal box. It is possible that the stimuli created by this part of the
apparatus served as a CS that became associated with food. After a number of
training trials, therefore, the sight of the goal box would activate a representation of
the reward and thereby permit the animal to detect when its value was changed. Both
Hull (1943) and Spence (1956) seized on this possibility and proposed that the
strength of instrumental responding is influenced by the Pavlovian properties of thecontext in which the response is performed.
The debate between S–R theorists and what might be called the expectancy
(R–US) theorists continued until the 1970s (see for example, Bolles, 1972). In the
last 20 years or so, however, experiments have provided new insights into the
nature of the associations that are formed during instrumental conditioning. To
anticipate the following discussion, these experiments show that both the S–R and
the expectancy theorists were correct. The experiments also show that these theorists
underestimated the complexity of the information that animals can acquire in even
quite simple instrumental conditioning tasks.
Evidence for R–US associations
To demonstrate support for an expectancy theory of instrumental conditioning,
Colwill and Rescorla (1985) adopted a reinforcer devaluation design (see also
Adams & Dickinson, 1981). A single group of rats was trained in the manner
summarized in Table 4.1. In the first (training) stage of the experiment subjects were
able to make one response (R1) to earn one reinforcer (US1) and another response
(R2) to earn a different reinforcer (US2). The two responses were lever pressing or
pulling a small chain that was suspended from the ceiling, and the two reinforcers
were food pellets or sucrose solution. After a number of sessions of this training, an
aversion was formed to US1 by allowing subjects free access to it and then injecting
them with a mild poison (lithium chloride; LiCl). This treatment was so effective thatsubjects completely rejected US1 when it was subsequently presented to them. For
the test trials subjects were again allowed to make either of the two responses, but
this time neither response led to the delivery of a reinforcer. The results from the
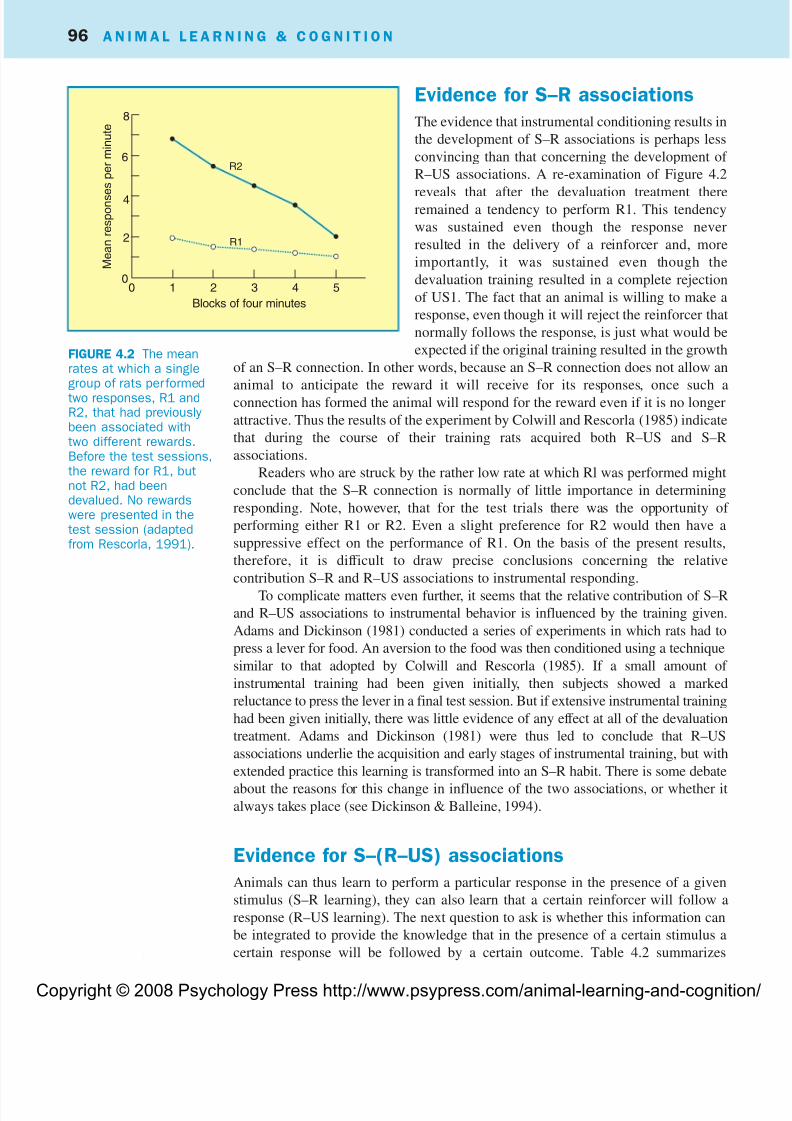
experiment are shown in Figure 4.2, which indicates that R2 was performed more
vigorously than R1. The figure also shows a gradual decline in the strength of R2,
which reflects the fact that neither response was followed by reward. This pattern of
results can be most readily explained by assuming that during their training rats
formed Rl–US1 and R2–US2 associations. They would then be reluctant to perform
R1 in the test phase because of their knowledge that this response produced a
reinforcer that was no longer attractive.
Training Devaluation Test
R1 → US1 US1 → LiCl R1 versus R2
R2 → US2
LiCl, lithium chloride; R, response; US, unconditioned stimulus.
TABLE 4.1 Summary of the training given to a single group of rats in an
experiment by Colwill and Rescorla (1985)
K E Y T E R M
Reinforcer devaluation
A technique in whichthe positive reinforcerfor an instrumentalresponse issubsequentlydevalued, normallyby pairing itsconsumption withillness.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 17/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
96 A N I M A L L E A R N I N G & C O G N I T I O N
Evidence for S–R associations
The evidence that instrumental conditioning results in
the development of S–R associations is perhaps less
convincing than that concerning the development of
R–US associations. A re-examination of Figure 4.2
reveals that after the devaluation treatment thereremained a tendency to perform R1. This tendency
was sustained even though the response never
resulted in the delivery of a reinforcer and, more
importantly, it was sustained even though the
devaluation training resulted in a complete rejection
of US1. The fact that an animal is willing to make a
response, even though it will reject the reinforcer that
normally follows the response, is just what would be
expected if the original training resulted in the growth
of an S–R connection. In other words, because an S–R connection does not allow an
animal to anticipate the reward it will receive for its responses, once such a
connection has formed the animal will respond for the reward even if it is no longer
attractive. Thus the results of the experiment by Colwill and Rescorla (1985) indicate
that during the course of their training rats acquired both R–US and S–R
associations.
Readers who are struck by the rather low rate at which Rl was performed might
conclude that the S–R connection is normally of little importance in determining
responding. Note, however, that for the test trials there was the opportunity of
performing either R1 or R2. Even a slight preference for R2 would then have a
suppressive effect on the performance of R1. On the basis of the present results,
therefore, it is difficult to draw precise conclusions concerning the relative
contribution S–R and R–US associations to instrumental responding.
To complicate matters even further, it seems that the relative contribution of S–R
and R–US associations to instrumental behavior is influenced by the training given.
Adams and Dickinson (1981) conducted a series of experiments in which rats had to
press a lever for food. An aversion to the food was then conditioned using a technique
similar to that adopted by Colwill and Rescorla (1985). If a small amount of
instrumental training had been given initially, then subjects showed a marked
reluctance to press the lever in a final test session. But if extensive instrumental training
had been given initially, there was little evidence of any effect at all of the devaluation
treatment. Adams and Dickinson (1981) were thus led to conclude that R–US
associations underlie the acquisition and early stages of instrumental training, but with
extended practice this learning is transformed into an S–R habit. There is some debate
about the reasons for this change in influence of the two associations, or whether italways takes place (see Dickinson & Balleine, 1994).
Evidence for S–(R–US) associations
Animals can thus learn to perform a particular response in the presence of a given
stimulus (S–R learning), they can also learn that a certain reinforcer will follow a
response (R–US learning). The next question to ask is whether this information can
be integrated to provide the knowledge that in the presence of a certain stimulus a
certain response will be followed by a certain outcome. Table 4.2 summarizes
FIGURE 4.2 The meanrates at which a singlegroup of rats performedtwo responses, R1 andR2, that had previouslybeen associated withtwo different rewards.Before the test sessions,the reward for R1, butnot R2, had beendevalued. No rewardswere presented in thetest session (adaptedfrom Rescorla, 1991).
8
6
4
2
00 1 2 3 4 5
Blocks of four minutes
M e a n r e s p o n s e s p e r m i n u t e
R2
R1

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 18/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 97
the design of an experiment by Rescorla (1991) that was conducted to test this
possibility.
A group of rats first received discrimination training in which a light or a noise
(S1 or S2) was presented for 30 seconds at a time. During each stimulus the rats were
trained to perform two responses (pulling a chain or pressing a lever), which each
resulted in a different reinforcer (food pellets or sucrose solution). The design of
conditioning experiments is rarely simple and, in this case, it was made moredifficult by reversing the response–reinforce relationships for the two stimuli. Thus
in S1, R1 led to US1 and R2 led to US2; but in S2, R1 led to US2 and R2 led to US1.
For the second stage of the experiment, the reinforcer devaluation technique was
used to condition an aversion to US2. Finally, test trials were conducted in extinction
in which subjects were provided with the opportunity of performing the two
responses in the presence of each stimulus. The result from these test trials was quite
clear. There was a marked preference to perform R1, rather than R2, in the presence
of S1; but in the presence of S2 there was a preference to perform R2 rather than R1.
These findings cannot be explained by assuming that the only associations acquired
during the first stage were S–R, otherwise the devaluation technique would have
been ineffective. Nor can the results be explained by assuming that only R–USassociations developed, otherwise devaluation treatment should have weakened R1
and R2 to the same extent in both stimuli. Instead, the results can be most readily
explained by assuming that the subjects were sensitive to the fact that the devalued
reinforcer followed R2 in S1, and followed R1 in S2. Rescorla (1991) has argued that
this conclusion indicates the development of a hierarchical associative structure that
he characterizes as S–(R–US). Animals are first believed to acquire an R–US
association, and this association in its entirety is then assumed to enter into a new
association with S. Whether it is useful to propose that an association can itself enter
into an association remains to be seen. There are certainly problems with this type of
suggestion (see, for example, Holland, 1992). In addition, as Dickinson (1994) points
out, there are alternative ways of explaining the findings of Rescorla (1991). Despite
these words of caution, the experiment demonstrates clearly that animals are able to
anticipate the reward they will receive for making a certain response in the presence
of a given stimulus.
THE COND IT IONS OF LEARN ING
There is, therefore, abundant evidence to show that animals are capable of learning
about the consequences of their actions. We turn now to consider the conditions that
enable this learning to take place.
Discrimination training Devaluation Test
Sl: R1 → US1 and R2 → US2 S1: R1 R2
US2 → LiClS2: R1 → US2 and R2 → US1 S2: R2 Rl
LiCl, lithium chloride; R, response; S, stimulus; US, unconditioned stimulus.
TABLE 4.2 Summary of the training given to a single group of rats in an
experiment by Rescorla (1991)
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 19/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
98 A N I M A L L E A R N I N G & C O G N I T I O N
Contiguity
A fundamental principle of the early theories of
learning was that instrumental conditioning is most
effective when the response is contiguous with or, in
other words, followed immediately by the reinforcer.
An early demonstration of this influence of contiguity on instrumental conditioning was made by
Logan (1960), who trained rats to run down an alley
for food. He found that the speed of running was
substantially faster if the rats received food as soon as
they reached the goal box, as opposed to waiting in
the goal box before food was made available. This
disruptive effect of waiting was found with delays
from as little as 3 seconds. Moreover, the speed of
running down the alley was directly related to the
duration of the delay in the goal box. This
effect, which is referred to as the gradient of
delay, has been reported on numerous occasions
(e.g. Dickinson, Watt, & Griffiths, 1992).
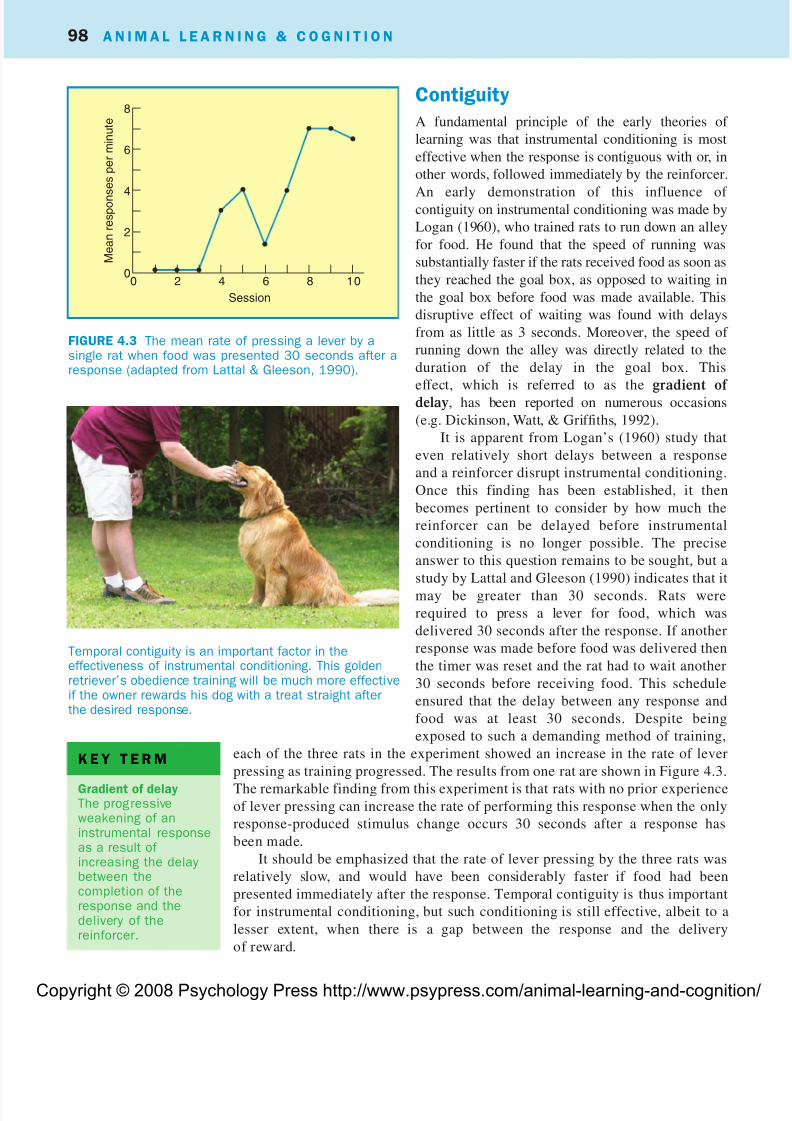
It is apparent from Logan’s (1960) study that
even relatively short delays between a response
and a reinforcer disrupt instrumental conditioning.
Once this finding has been established, it then
becomes pertinent to consider by how much the
reinforcer can be delayed before instrumental
conditioning is no longer possible. The precise
answer to this question remains to be sought, but a
study by Lattal and Gleeson (1990) indicates that it
may be greater than 30 seconds. Rats were
required to press a lever for food, which was
delivered 30 seconds after the response. If another
response was made before food was delivered then
the timer was reset and the rat had to wait another
30 seconds before receiving food. This schedule
ensured that the delay between any response and
food was at least 30 seconds. Despite being
exposed to such a demanding method of training,
each of the three rats in the experiment showed an increase in the rate of lever
pressing as training progressed. The results from one rat are shown in Figure 4.3.
The remarkable finding from this experiment is that rats with no prior experienceof lever pressing can increase the rate of performing this response when the only
response-produced stimulus change occurs 30 seconds after a response has
been made.
It should be emphasized that the rate of lever pressing by the three rats was
relatively slow, and would have been considerably faster if food had been
presented immediately after the response. Temporal contiguity is thus important
for instrumental conditioning, but such conditioning is still effective, albeit to a
lesser extent, when there is a gap between the response and the delivery
of reward.
FIGURE 4.3 The mean rate of pressing a lever by asingle rat when food was presented 30 seconds after aresponse (adapted from Lattal & Gleeson, 1990).
8
6
4
2
00
M e a n r e s p o n s e s p e r m i n u t e
2 4 6 8 10
Session
Temporal contiguity is an important factor in theeffectiveness of instrumental conditioning. This goldenretriever’s obedience training will be much more effectiveif the owner rewards his dog with a treat straight afterthe desired response.
K E Y T E R M
Gradient of delayThe progressiveweakening of aninstrumental responseas a result of increasing the delaybetween thecompletion of theresponse and thedelivery of thereinforcer.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 20/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 99
Response–reinforcer contingency
We saw in Chapter 3 that the CS–US contingency is
important for Pavlovian conditioning because learning
is more effective when the US occurs only in the
presence of the CS than when the US also occurs both
in the presence and absence of the CS. An experimentby Hammond (1980) makes a similar point for
instrumental behavior, by demonstrating the
importance of the response–reinforcer contingency
for effective conditioning. The training schedule was
quite complex and required that the experimental
session was divided into 1-second intervals. If a
response occurred in any interval then, for three groups
of thirsty rats, water was delivered at the end of the
interval with a probability of 0.12. The results from a
group that received only this training, and no water in
the absence of lever pressing (Group 0), are shown in
the left-hand histogram of Figure 4.4. By the end of
training this group was responding at more than 50
responses a minute. For the remaining two groups,
water was delivered after some of the 1-second
intervals in which a response did not occur. For Group
0.08, the probability of one of these intervals being followed by water was 0.08, whereas
for Group 0.12 this probability was 0.12. The remaining two histograms show the final
response rates for these two groups. Both groups responded more slowly than Group 0,
but responding was weakest in the group for which water was just as likely to be
delivered whether or not a response had been made. The contingency between response
and reinforcer thus influences the rate at which the response will be performed. We now
need to ask why this should be the case. In fact, there are two answers to this question.
One answer is based on a quite different view of instrumental conditioning to that
considered thus far. According to this account, instrumental conditioning will be
effective whenever a response results in an increase in the rate of reinforcement
(e.g. Baum, 1973). Thus there is no need for a response to be followed closely by
reward for successful conditioning, all that is necessary is for the overall probability of
reward being delivered to increase. In other words, the contingency between a response
and reward is regarded as the critical determinant for the outcome of instrumental
conditioning. This position is referred to as a molar theory of reinforcement because
animals are assumed to compute the rate at which they make a response over a
substantial period of time and, at the same time, compute the rate at which reward is
delivered over the same period. If they should detect that an increase in the rate of responding is correlated with an increase in the rate of reward delivery, then the
response will be performed more vigorously in the future. Moreover, the closer the
correlation between the two rates, the more rapidly will the response be performed.
Group 0 of Hammond’s (1980) experiment demonstrated a high correlation between
the rate at which the lever was pressed and the rate at which reward was delivered, and
this molar analysis correctly predicts that rats will learn to respond rapidly on the lever.
In the case of Group 0.12, however, the rate of lever pressing had some influence on
the rate at which reward was delivered, but this influence was slight because the reward
would be delivered even if a rat refused to press the lever. In these circumstances,
FIGURE 4.4 The mean rates of lever pressing for waterby three groups of thirsty rats in their final session of training. The groups differed in the probability withwhich free water was delivered during the intervals
between responses. Group 0 received no water duringthese intervals, Group 0.08 and Group 0.12 receivedwater with a probability of 0.08 and 0.12, respectively, atthe end of each period of 1 second in which a responsedid not occur (adapted from Hammond, 1980).
60
40
20
00 0.08 0.12
Group
M e a n r e s p o n s e s p e r m i n u t e
K E Y T E R M S
Response–reinforcer
contingency
The degree to whichthe occurrence of the
reinforcer dependson the instrumentalresponse. Positivecontingency: thefrequency of thereinforcer is increasedby making the response.Negative contingency:the frequency of thereinforcer is reduced bymaking the response.Zero contingency: thefrequency of thereinforcer is unaffectedby making the response.
Molar theory of
reinforcement
The assumption thatthe rate of instrumentalresponding isdetermined by theresponse–reinforcercontingency.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 21/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
100 A N I M A L L E A R N I N G & C O G N I T I O N
responding is predicted to be slow and again the theory
is supported by the findings.
The molar analysis of instrumental behavior has
received a considerable amount of attention and
generated a considerable body of experimental
research, but there are good reasons for believing that it
may be incorrect. In an experiment by Thomas (1981),rats in a test chamber containing a lever were given a
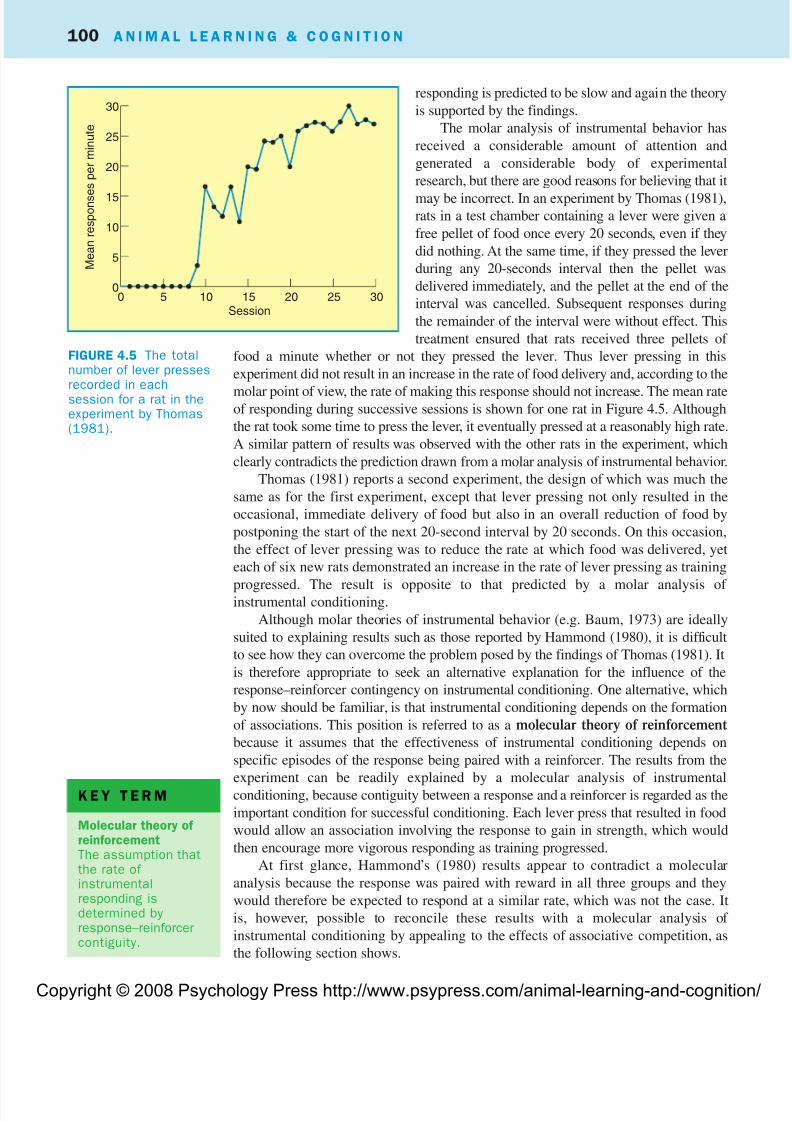
free pellet of food once every 20 seconds, even if they
did nothing. At the same time, if they pressed the lever
during any 20-seconds interval then the pellet was
delivered immediately, and the pellet at the end of the
interval was cancelled. Subsequent responses during
the remainder of the interval were without effect. This
treatment ensured that rats received three pellets of
food a minute whether or not they pressed the lever. Thus lever pressing in this
experiment did not result in an increase in the rate of food delivery and, according to the
molar point of view, the rate of making this response should not increase. The mean rateof responding during successive sessions is shown for one rat in Figure 4.5. Although
the rat took some time to press the lever, it eventually pressed at a reasonably high rate.
A similar pattern of results was observed with the other rats in the experiment, which
clearly contradicts the prediction drawn from a molar analysis of instrumental behavior.
Thomas (1981) reports a second experiment, the design of which was much the
same as for the first experiment, except that lever pressing not only resulted in the
occasional, immediate delivery of food but also in an overall reduction of food by
postponing the start of the next 20-second interval by 20 seconds. On this occasion,
the effect of lever pressing was to reduce the rate at which food was delivered, yet
each of six new rats demonstrated an increase in the rate of lever pressing as training
progressed. The result is opposite to that predicted by a molar analysis of instrumental conditioning.
Although molar theories of instrumental behavior (e.g. Baum, 1973) are ideally
suited to explaining results such as those reported by Hammond (1980), it is difficult
to see how they can overcome the problem posed by the findings of Thomas (1981). It
is therefore appropriate to seek an alternative explanation for the influence of the
response–reinforcer contingency on instrumental conditioning. One alternative, which
by now should be familiar, is that instrumental conditioning depends on the formation
of associations. This position is referred to as a molecular theory of reinforcement
because it assumes that the effectiveness of instrumental conditioning depends on
specific episodes of the response being paired with a reinforcer. The results from the
experiment can be readily explained by a molecular analysis of instrumental
conditioning, because contiguity between a response and a reinforcer is regarded as the
important condition for successful conditioning. Each lever press that resulted in food
would allow an association involving the response to gain in strength, which would
then encourage more vigorous responding as training progressed.
At first glance, Hammond’s (1980) results appear to contradict a molecular
analysis because the response was paired with reward in all three groups and they
would therefore be expected to respond at a similar rate, which was not the case. It
is, however, possible to reconcile these results with a molecular analysis of
instrumental conditioning by appealing to the effects of associative competition, as
the following section shows.
FIGURE 4.5 The totalnumber of lever pressesrecorded in eachsession for a rat in theexperiment by Thomas(1981).
30
25
20
15
10
5
00 5 10 15 20 25 30
Session
M e a n r e s p o n s e s p e r m i n u t e
K E Y T E R M
Molecular theory of
reinforcement
The assumption thatthe rate of instrumentalresponding isdetermined byresponse–reinforcercontiguity.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 22/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 101
Associative competition
If two stimuli are presented together for Pavlovian conditioning, the strength of the
conditioned response (CR) that each elicits when tested individually is often weaker
than if they are presented for conditioning separately. This overshadowing effect is
explained by assuming the two stimuli are in competition for associative strength so
that the more strength acquired by one the less is available for the other (Rescorla &Wagner, 1972). Overshadowing is normally assumed to take place between stimuli
but, if it is accepted that overshadowing can also occur between stimuli and
responses that signal the same reinforcer, then it is possible for molecular theories of
instrumental behavior to explain the contingency effects reported by Hammond
(1980). In Group 0 of his study, each delivery of water would strengthen a
lever-press–water association and result eventually in rapid lever pressing. In the
other groups, however, the delivery of free water would allow the context to enter
into an association with this reinforcer. The delivery of water after a response will
then mean that it is signaled by both the context and the response, and theories of
associative learning predict that the context–water association will restrict, through
overshadowing, the growth of the response–water association. As the strength of the
response–water association determines the rate at which the response is performed,
responding will be slower when some free reinforcers accompany the instrumental
training than when all the reinforcers are earned. Furthermore, the more often that
water is delivered free, the stronger will be the
context–water association and the weaker will be the
response–water association. Thus the pattern of
results shown in Figure 4.4 can be explained by a
molecular analysis of instrumental conditioning,
providing it is assumed that responses and stimuli
compete with each other for their associative
strength. The results from two different experiments
lend support to this assumption.
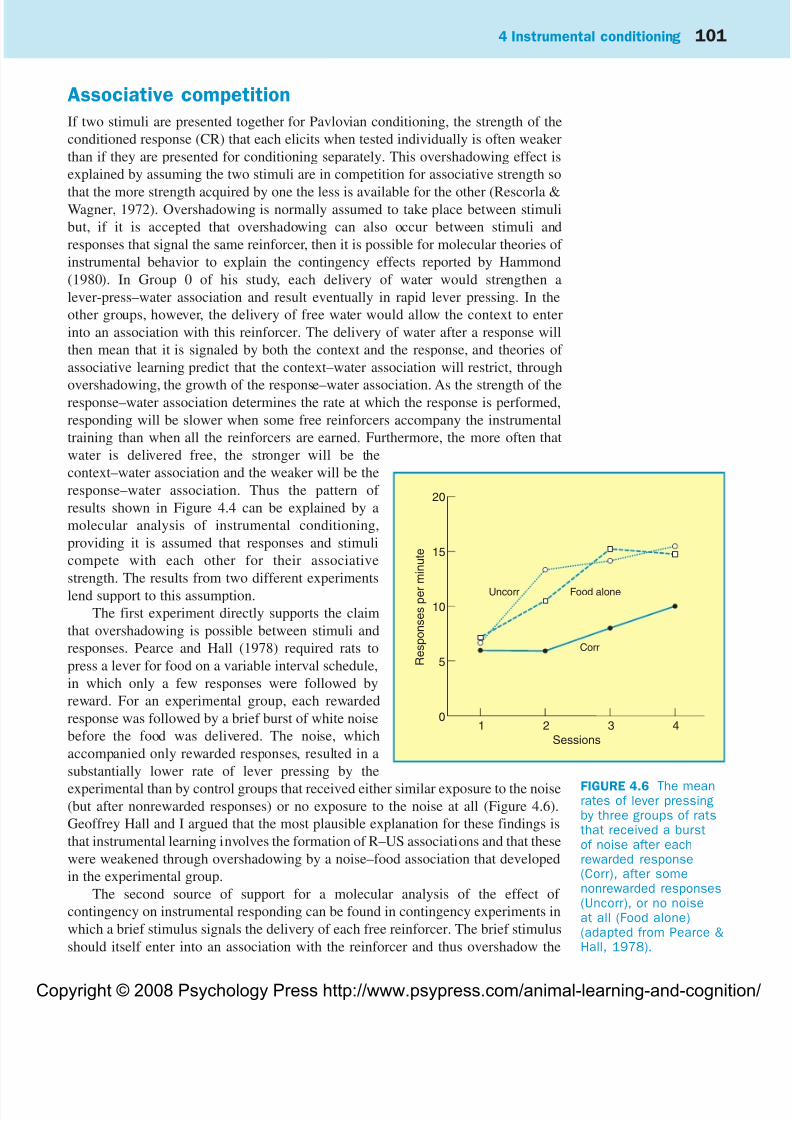
The first experiment directly supports the claim
that overshadowing is possible between stimuli and
responses. Pearce and Hall (1978) required rats to
press a lever for food on a variable interval schedule,
in which only a few responses were followed by
reward. For an experimental group, each rewarded
response was followed by a brief burst of white noise
before the food was delivered. The noise, which
accompanied only rewarded responses, resulted in a
substantially lower rate of lever pressing by the
experimental than by control groups that received either similar exposure to the noise(but after nonrewarded responses) or no exposure to the noise at all (Figure 4.6).
Geoffrey Hall and I argued that the most plausible explanation for these findings is
that instrumental learning involves the formation of R–US associations and that these
were weakened through overshadowing by a noise–food association that developed
in the experimental group.
The second source of support for a molecular analysis of the effect of
contingency on instrumental responding can be found in contingency experiments in
which a brief stimulus signals the delivery of each free reinforcer. The brief stimulus
should itself enter into an association with the reinforcer and thus overshadow the
FIGURE 4.6 The meanrates of lever pressingby three groups of ratsthat received a burstof noise after eachrewarded response(Corr), after somenonrewarded responses(Uncorr), or no noiseat all (Food alone)(adapted from Pearce &Hall, 1978).
20
15
10
5
01 2 3 4
Sessions
R e s p o n s e s p e r
m i n u t e
Corr
Uncorr Food alone

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 23/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
102 A N I M A L L E A R N I N G & C O G N I T I O N
development of an association between the context and the reinforcer. Whenever a
response is followed by the reinforcer it will now be able to enter a normal R–US
association, because of the lack of competition from the context. Responding in these
conditions should thus be more vigorous than if the free US is not signaled. In
support of this argument, both Hammond and Weinberg (1984) and Dickinson and
Charnock (1985) have shown that free reinforcers disrupt instrumental responding to
a greater extent when they are unsignaled than when they are signaled. Thesefindings make a particularly convincing case for the belief that competition for
associative strength is an important influence on the strength of an instrumental
response. They also indicate that this competition is responsible for the influence of
the response–reinforcer contingency on the rate of instrumental responding.
The nature of the reinforcer
Perhaps the most important requirement for successful instrumental conditioning is
that the response is followed by a reinforcer. But what makes a reinforcer? In nearly
all the experiments that have been described thus far, the reinforcer has been food fora hungry animal, or water for a thirsty animal. As these stimuli are of obvious
biological importance, it is hardly surprising to discover that animals are prepared to
engage in an activity such as lever pressing in order to earn them. However, this does
not mean that a reinforcer is necessarily a stimulus that is of biological significance
to the animal. As Schwartz (1989) notes, animals will press a lever to turn on a light,
and it is difficult to imagine the biological need that is satisfied on these occasions.
Thorndike (1911) was the first to appreciate the need to identify the defining
characteristics of a reinforcer, and his solution was contained within the Law of
Effect. He maintained that a reinforcer was a stimulus that resulted in a satisfying
state of affairs. A satisfying state of affairs was then defined as “ . . . one which the
animal does nothing to avoid, often doing things which maintain or renew it”
(Thorndike, 1913, p. 2). In other words, Thorndike effectively proposed that a
stimulus would serve as a reinforcer (increase the likelihood of a response) if animals
were willing to respond in order to receive that stimulus. The circularity in this
definition should be obvious and has served as a valid source of criticism of the Law
of Effect on more than one occasion (e.g. Meehl, 1950). Thorndike was not alone in
providing a circular definition of a reinforcer. Skinner has been perhaps the most
blatant in this respect, as the following quotation reveals (Skinner, 1953, pp. 72–73):
The only way to tell whether or not a given event is reinforcing to a given
organism under given conditions is to make a direct test. We observe the
frequency of a selected response, then make an event contingent upon it and
observe any change in frequency. If there is a change, we classify the event as
reinforcing.
To be fair, for practical purposes this definition is quite adequate. It provides a
useful and unambiguous terminology. At the same time, once we have decided that
a stimulus, such as food, is a positive reinforcer, then we can turn to a study of a
number of issues that are important to the analysis of instrumental learning. For
instance, we have been able to study the role of the reinforcer in the associations that
are formed during instrumental learning, without worrying unduly about what it is
that makes a stimulus a reinforcer. But the definitions offered by Thorndike and
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 24/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 103
Skinner are not very helpful if a general statement is
being sought about the characteristics of a stimulus
that dictate whether or not it will function as a
reinforcer. And the absence of such a general
statement makes our understanding of the conditions
that promote instrumental learning incomplete.
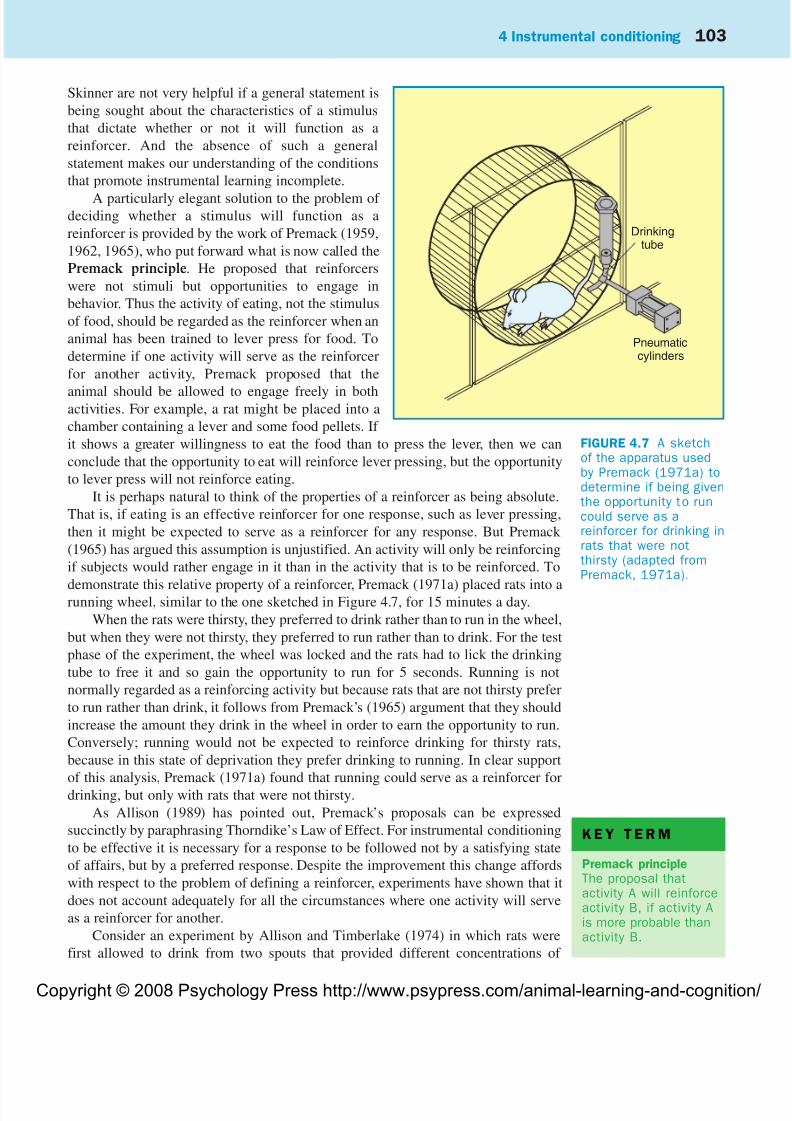
A particularly elegant solution to the problem of deciding whether a stimulus will function as a
reinforcer is provided by the work of Premack (1959,
1962, 1965), who put forward what is now called the
Premack principle. He proposed that reinforcers
were not stimuli but opportunities to engage in
behavior. Thus the activity of eating, not the stimulus
of food, should be regarded as the reinforcer when an
animal has been trained to lever press for food. To
determine if one activity will serve as the reinforcer
for another activity, Premack proposed that the
animal should be allowed to engage freely in bothactivities. For example, a rat might be placed into a
chamber containing a lever and some food pellets. If
it shows a greater willingness to eat the food than to press the lever, then we can
conclude that the opportunity to eat will reinforce lever pressing, but the opportunity
to lever press will not reinforce eating.
It is perhaps natural to think of the properties of a reinforcer as being absolute.
That is, if eating is an effective reinforcer for one response, such as lever pressing,
then it might be expected to serve as a reinforcer for any response. But Premack
(1965) has argued this assumption is unjustified. An activity will only be reinforcing
if subjects would rather engage in it than in the activity that is to be reinforced. To
demonstrate this relative property of a reinforcer, Premack (1971a) placed rats into arunning wheel, similar to the one sketched in Figure 4.7, for 15 minutes a day.
When the rats were thirsty, they preferred to drink rather than to run in the wheel,
but when they were not thirsty, they preferred to run rather than to drink. For the test
phase of the experiment, the wheel was locked and the rats had to lick the drinking
tube to free it and so gain the opportunity to run for 5 seconds. Running is not
normally regarded as a reinforcing activity but because rats that are not thirsty prefer
to run rather than drink, it follows from Premack’s (1965) argument that they should
increase the amount they drink in the wheel in order to earn the opportunity to run.
Conversely; running would not be expected to reinforce drinking for thirsty rats,
because in this state of deprivation they prefer drinking to running. In clear support
of this analysis, Premack (1971a) found that running could serve as a reinforcer for
drinking, but only with rats that were not thirsty.
As Allison (1989) has pointed out, Premack’s proposals can be expressed
succinctly by paraphrasing Thorndike’s Law of Effect. For instrumental conditioning
to be effective it is necessary for a response to be followed not by a satisfying state
of affairs, but by a preferred response. Despite the improvement this change affords
with respect to the problem of defining a reinforcer, experiments have shown that it
does not account adequately for all the circumstances where one activity will serve
as a reinforcer for another.
Consider an experiment by Allison and Timberlake (1974) in which rats were
first allowed to drink from two spouts that provided different concentrations of
FIGURE 4.7 A sketchof the apparatus usedby Premack (1971a) todetermine if being giventhe opportunity to runcould serve as areinforcer for drinking inrats that were notthirsty (adapted fromPremack, 1971a).
Drinkingtube
Pneumaticcylinders
K E Y T E R M
Premack principle
The proposal thatactivity A will reinforceactivity B, if activity Ais more probable thanactivity B.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 25/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
104 A N I M A L L E A R N I N G & C O G N I T I O N
saccharin solution. This baseline test session revealed a preference for the sweeter
solution. According to Premack’s proposals, therefore, rats should be willing to
increase their consumption of the weaker solution if drinking it is the only means
by which they can gain access to the sweeter solution. By contrast, rats should not
be willing to increase their consumption of the sweeter solution to gain access to the
weaker one. To test this second prediction, rats were allowed to drink from the spout
supplying the sweeter solution and, after every 10 licks, they were permitted onelick at the spout offering the less-sweet solution. This 10 : 1 ratio meant that,
relative to the amount of sweet solution consumed, the rats received less of the
weaker solution than they chose to consume in the baseline test session. As a
consequence of this constraint imposed by the experiment, Allison and Timberlake
(1974) found that rats increased their consumption of the stronger solution. It is
important to emphasize that this increase occurred in order to allow the rats to gain
access to the less preferred solution, which, according to Premack’s theory, should
not have taken place.
Timberlake and Allison (1974) explained their results in terms of an equilibrium
theory of behavior. They argued that when an animal is able to engage in a variety of
activities, it will have a natural tendency to allocate more time to some than others.The ideal amount of time that would be devoted to an activity is referred to as its bliss
point, and each activity is assumed to have its own bliss point. By preventing an
animal from engaging in even its least preferred activity, it will be displaced from the
bliss point and do its best to restore responding to this point.
In the experiment by Allison and Timberlake (1974), therefore, forcing the
subjects to drink much more of the strong than the weak solution meant that they
were effectively deprived of the weak solution. As the only way to overcome this
deficit was to drink more of the sweet solution, this is what they did. Of course, as
the rats approached their bliss point for the consumption of the weak solution, they
would go beyond their bliss point for the consumption of the sweet solution. To cope
with this type of conflict, animals are believed to seek a compromise, or state of equilibrium, in which the amount of each activity they perform will lead them as
close as possible to the bliss points for all activities. Thus the rats completed the
experiment by drinking rather more than they would prefer of the strong solution,
and rather less than they would prefer of the weak solution.
By referring to bliss points, we can thus predict when the opportunity to engage
in one activity will serve as a reinforcer for another activity. But this does not mean
that we have now identified completely the circumstances in which the delivery of a
particular event will function as a reinforcer. Some reinforcers do not elicit responses
that can be analyzed usefully by equilibrium theory. Rats will press a lever to receive
stimulation to certain regions of the brain, or to turn on a light, or to turn off an
electric shock to the feet. I find it difficult to envisage how any measure of baseline
activity in the presence of these events would reveal that they will serve as
reinforcers for lever pressing. In the next section we will find that a stimulus that has
been paired with food can reinforce lever pressing in hungry rats. Again, simply by
observing an animal’s behavior in the presence of the stimulus, it is hard to imagine
how one could predict that the stimulus will function as a reinforcer. Our
understanding of the nature of a reinforcer has advanced considerably since
Thorndike proposed the Law of Effect. However, if we wish to determine with
confidence if a certain event will act as a reinforcer for a particular response, at times
there will be no better alternative than to adopt Skinner’s suggestion of testing for
this property directly.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 26/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 105
Conditioned reinforcement
The discussion has been concerned thus far with primary reinforcers, that is, with
stimuli that do not need to be paired with another stimulus to function as reinforcers
for instrumental conditioning. There are, in addition, numerous studies that have
shown that even a neutral stimulus may serve as an instrumental reinforcer by virtue
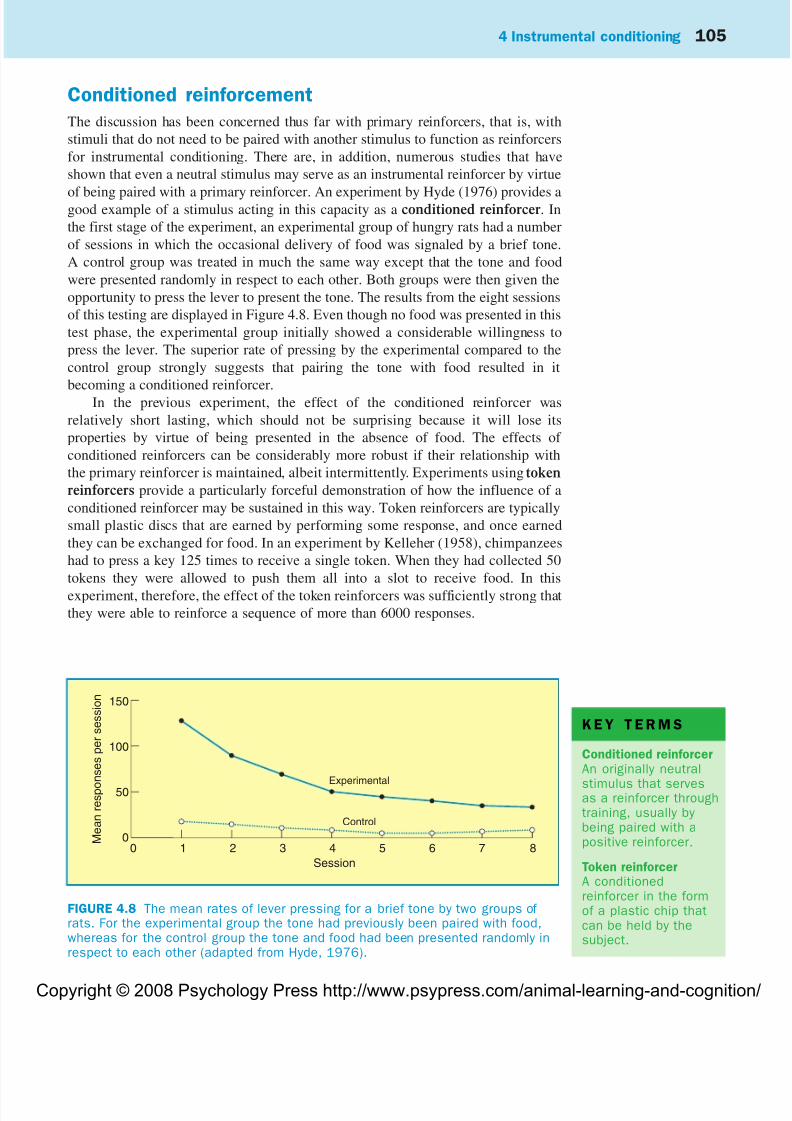
of being paired with a primary reinforcer. An experiment by Hyde (1976) provides agood example of a stimulus acting in this capacity as a conditioned reinforcer. In
the first stage of the experiment, an experimental group of hungry rats had a number
of sessions in which the occasional delivery of food was signaled by a brief tone.
A control group was treated in much the same way except that the tone and food
were presented randomly in respect to each other. Both groups were then given the
opportunity to press the lever to present the tone. The results from the eight sessions
of this testing are displayed in Figure 4.8. Even though no food was presented in this
test phase, the experimental group initially showed a considerable willingness to
press the lever. The superior rate of pressing by the experimental compared to the
control group strongly suggests that pairing the tone with food resulted in it
becoming a conditioned reinforcer.
In the previous experiment, the effect of the conditioned reinforcer was
relatively short lasting, which should not be surprising because it will lose its
properties by virtue of being presented in the absence of food. The effects of
conditioned reinforcers can be considerably more robust if their relationship with
the primary reinforcer is maintained, albeit intermittently. Experiments using token
reinforcers provide a particularly forceful demonstration of how the influence of a
conditioned reinforcer may be sustained in this way. Token reinforcers are typically
small plastic discs that are earned by performing some response, and once earned
they can be exchanged for food. In an experiment by Kelleher (1958), chimpanzees
had to press a key 125 times to receive a single token. When they had collected 50
tokens they were allowed to push them all into a slot to receive food. In this
experiment, therefore, the effect of the token reinforcers was sufficiently strong that
they were able to reinforce a sequence of more than 6000 responses.
FIGURE 4.8 The mean rates of lever pressing for a brief tone by two groups of rats. For the experimental group the tone had previously been paired with food,whereas for the control group the tone and food had been presented randomly inrespect to each other (adapted from Hyde, 1976).
150
100
50
00 1 2 3 4 5 6 7 8
Session
M e a n r e s p o
n s e s p e r s e s s i o n
Experimental
Control
K E Y T E R M S
Conditioned reinforcer
An originally neutral
stimulus that servesas a reinforcer throughtraining, usually bybeing paired with apositive reinforcer.
Token reinforcer
A conditionedreinforcer in the formof a plastic chip thatcan be held by thesubject.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 27/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
106 A N I M A L L E A R N I N G & C O G N I T I O N
A straightforward explanation for the results of the experiment by Hyde (1976)
is that the tone became an appetitive Pavlovian CS and thus effectively served as a
substitute for food. The results from experiments such as that by Kelleher (1958)
have led Schwartz (1989) to argue that there are additional ways in which
conditioned reinforcers can be effective (see also Golub, 1977):
• They provide feedback that the correct response has been made. Delivering a tokenafter the completion of 125 responses would provide a useful signal that the subject
is engaged in the correct activity.
• Conditioned reinforcers might act as a cue for the next response to be performed.
Kelleher (1958) observed that his chimpanzees often waited for several hours
before making their first response in a session. This delay was virtually eliminated
by giving the subject some tokens at the start of the session, thus indicating that the
tokens acted as a cue for key pressing.
• Conditioned reinforcers may be effective because they help to counteract the
disruptive effects of imposing a long delay between a response and the delivery of
a primary reinforcer. Interestingly, as far as tokens are concerned, this property of the token is seen only when the chimpanzee is allowed to hold it during the delay.
Taken together, these proposals imply that the properties of a conditioned
reinforcer are considerably more complex than would be expected if they were based
solely on its Pavlovian properties.
THE PERFORMANCE OF INSTRUMENTAL
BEHAV IOR
The experiments considered so far have been concerned with revealing theknowledge that is acquired during the course of instrumental conditioning. They
have also indicated some of the factors that influence the acquisition of this
knowledge. We turn our attention now to examining the factors that determine the
vigor with which an animal will perform an instrumental response. We have already
seen that certain devaluation treatments can influence instrumental responding, and
so too can manipulations designed to modify the strength of the instrumental
association. But there remain a number of other factors that influence instrumental
behavior. In the discussion that follows we shall consider two of these influences in
some detail: deprivation state and the presence of Pavlovian CSs.
Deprivation
The level of food deprivation has been shown, up to a point, to be directly related to
the vigor with which an animal responds for food. This is true when the response is
running down an alley (Cotton, 1953) or pressing a lever (Clark, 1958). To explain this
relationship, Hull (1943) suggested that motivational effects are mediated by activity
in a drive center. Drive is a central state that is excited by needs and energizes
behavior. It was proposed that the greater the level of drive, the more vigorous will be
the response that the animal is currently performing. Thus, if a rat is pressing a lever
for food, then hunger will excite drive, which, in turn, will invigorate this activity.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 28/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 107
A serious shortcoming of Hull’s (1943) account is the claim that drive is
nonspecific, so that it can be enhanced by an increase in any need of the animal.
A number of curious predictions follow from this basic aspect of his theorizing. For
example, the pain produced by electric shock is assumed to increase drive, so that if
animals are given shocks while lever pressing for food, they should respond more
rapidly than in the absence of shock. By far the most frequent finding is that this
manipulation has the opposite effect of decreasing appetitive instrumentalresponding (e.g. Boe & Church, 1967). Conversely, the theory predicts that
enhancing drive by making animals hungrier should facilitate the rate at which they
press a lever to escape or avoid shock. Again, it should not be surprising to discover
that generally this prediction is not confirmed. Increases in deprivation have been
found, in this respect, to be either without effect (Misanin & Campbell, 1969) or to
reduce the rate of such behavior (Meyer, Adams, & Worthen, 1969; Leander, 1973).
In response to this problem, more recent theorists have proposed that animals
possess two drive centers: One is concerned with energizing behavior that leads to
reward, the other is responsible for invigorating activity that minimizes contact
with aversive stimuli. These can be referred to, respectively, as the positive and
negative motivational systems. A number of such dual-system theories of motivation have been proposed (Konorski, 1967; Rescorla & Solomon, 1967;
Estes, 1969).
The assumption that there are two motivational systems rather than a single drive
center allows these theories to overcome many of the problems encountered by
Hull’s (1943) theory. For example, it is believed that deprivation states like hunger
and thirst will increase activity only in the positive system, so that a change in
deprivation should not influence the vigor of behavior that minimizes contact with
aversive stimuli such as shock. Conversely, electric shock should not invigorate
responding for food as it will excite only the negative system.
But even this characterization of the way in which deprivation states influence
behavior may be too simple. Suppose that an animal that has been trained to leverpress for food when it is hungry is satiated by being granted unrestricted access to
food before it is returned to the conditioning chamber. The account that has just been
developed predicts that satiating the animal will reduce the motivational support for
lever pressing by lowering the activity in the positive system. The animal would thus
be expected to respond less vigorously than one that was still hungry. There is some
evidence to support this prediction (e.g. Balleine, Garner, Gonzalez & Dickinson,
1995), but additional findings by Balleine (1992) demonstrate that dual-system
theories of motivation are in need of elaboration if they are to provide a complete
account of the way in which deprivation states influence responding.
In one experiment by Balleine (1992), two groups of rats were trained to press a
bar for food while they were hungry (H). For reasons that will be made evident
shortly, it is important to note that the food pellets used as the instrumental reinforcer
were different to the food that was presented at all other times in this experiment.
Group H–S was then satiated (S) by being allowed unrestricted access to their normal
food for 24 hours, whereas Group H–H remained on the deprivation schedule.
Finally, both groups were again given the opportunity to press the bar, but responding
never resulted in the delivery of the reinforcer. Because of their different deprivation
states, dual-system theories of motivation, as well as our intuitions, predict that
Group H–H should respond more vigorously than Group H–S in this test session. But
it seems that our intuitions are wrong on this occasion. The mean number of
responses made by each group in the test session are shown in the two gray
K E Y T E R M
Dual-system theories
of motivation
Theories that assumethat behavior ismotivated by activityin a positive system,which energizesapproach to an object,and a negativesystem, whichenergizes withdrawalfrom an object.
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 29/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
108 A N I M A L L E A R N I N G & C O G N I T I O N
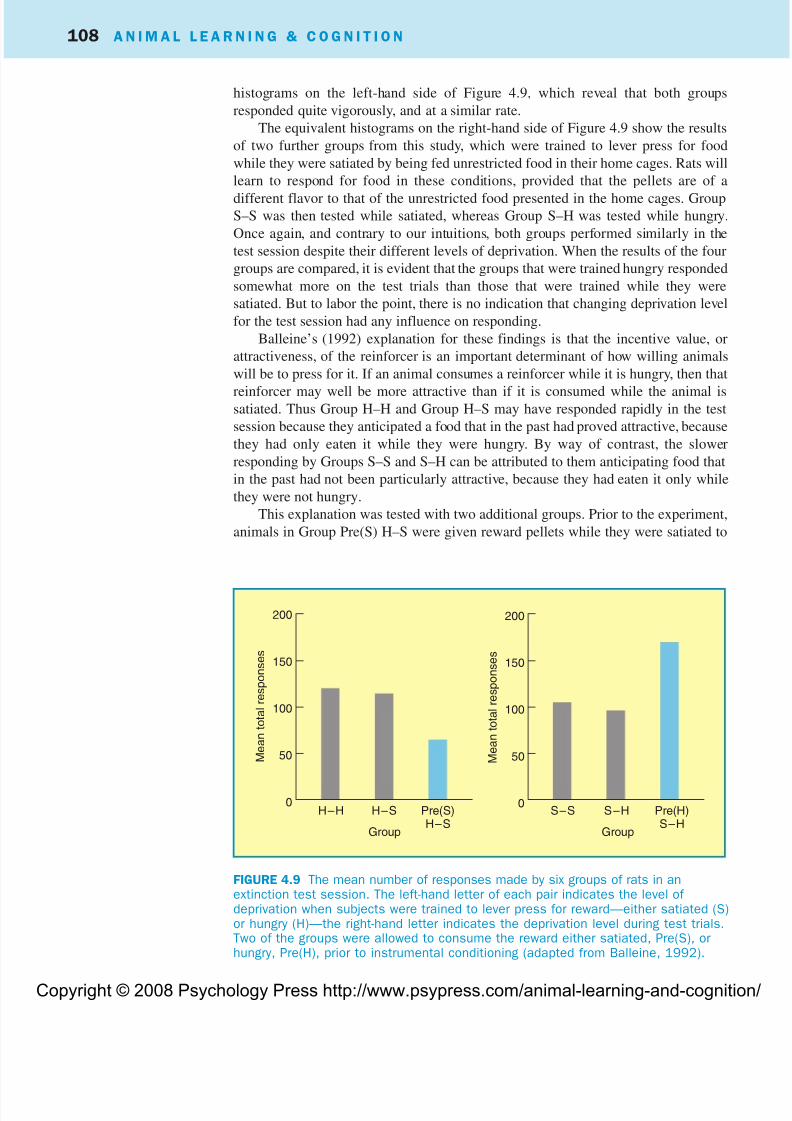
histograms on the left-hand side of Figure 4.9, which reveal that both groups
responded quite vigorously, and at a similar rate.
The equivalent histograms on the right-hand side of Figure 4.9 show the results
of two further groups from this study, which were trained to lever press for food
while they were satiated by being fed unrestricted food in their home cages. Rats will
learn to respond for food in these conditions, provided that the pellets are of a
different flavor to that of the unrestricted food presented in the home cages. GroupS–S was then tested while satiated, whereas Group S–H was tested while hungry.
Once again, and contrary to our intuitions, both groups performed similarly in the
test session despite their different levels of deprivation. When the results of the four
groups are compared, it is evident that the groups that were trained hungry responded
somewhat more on the test trials than those that were trained while they were
satiated. But to labor the point, there is no indication that changing deprivation level
for the test session had any influence on responding.
Balleine’s (1992) explanation for these findings is that the incentive value, or
attractiveness, of the reinforcer is an important determinant of how willing animals
will be to press for it. If an animal consumes a reinforcer while it is hungry, then that
reinforcer may well be more attractive than if it is consumed while the animal issatiated. Thus Group H–H and Group H–S may have responded rapidly in the test
session because they anticipated a food that in the past had proved attractive, because
they had only eaten it while they were hungry. By way of contrast, the slower
responding by Groups S–S and S–H can be attributed to them anticipating food that
in the past had not been particularly attractive, because they had eaten it only while
they were not hungry.
This explanation was tested with two additional groups. Prior to the experiment,
animals in Group Pre(S) H–S were given reward pellets while they were satiated to
FIGURE 4.9 The mean number of responses made by six groups of rats in anextinction test session. The left-hand letter of each pair indicates the level of deprivation when subjects were trained to lever press for reward—either satiated (S)or hungry (H)—the right-hand letter indicates the deprivation level during test trials.Two of the groups were allowed to consume the reward either satiated, Pre(S), orhungry, Pre(H), prior to instrumental conditioning (adapted from Balleine, 1992).
200
150
100
50
0H–H H–S Pre(S)
H–SS–S S–H Pre(H)
S–HGroup Group
M e a n t o t a l r e s p o n s e s
200
150
100
50
0
M e a n t o t a l r e s p o n s e s

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 30/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 109
demonstrate that the pellets are not particularly attractive in this deprivation state.
The group was then trained to lever press while hungry and received test trials while
satiated. On the test trials, the subjects should know that because, of their low level
of deprivation, the reward pellets are no longer attractive and they should be reluctant
to press the lever. The results, which are shown in the blue histogram in the left-hand
side of Figure 4.9, confirmed this prediction. The final group to be considered, Group
Pre(H) S–H, was first allowed to eat reward pellets in the home cage while hungry,instrumental conditioning was then conducted while the group was satiated and the
test trials were conducted while the group was hungry. In contrast to Group S–H and
Group S–S, this group should appreciate that the reward pellets are attractive while
hungry and respond more rapidly than the other two groups during the test trials.
Once again, the results confirmed this prediction—see the blue histogram on the
right-hand side of Figure 4.9
By now it should be evident that no simple conclusion can be drawn concerning
the way in which deprivation states influence the vigor of instrumental responding.
On some occasions a change in deprivation state is able to modify directly the rate
of responding, as dual-systems theories of motivation predict. On other occasions,
this influence is more indirect by modifying the attractiveness of the reinforcer. Aninformative account of the way in which these findings may be integrated can be
found in Balleine et al. (1995).
Pavlovian–instrumental interactions
For a long time, theorists have been interested in the way in which Pavlovian CSs
influence the strength of instrumental responses that are performed in their
presence. One reason for this interest is that Pavlovian and instrumental
conditioning are regarded as two fundamental learning processes, and it is
important to appreciate the way in which they work together to determine how an
animal behaves. A second reason was mentioned at the end of Chapter 2, where wesaw that Pavlovian CSs tend to elicit reflexive responses that may not always be in
the best interests of the animal. If a Pavlovian CS was also able to modulate the
vigor of instrumental responding, then this would allow it to have a more general,
and more flexible, influence on behavior than has so far been implied. For example,
if a CS for food were to invigorate instrumental responses that normally lead to
food, then such responses would be strongest at a time when they are most needed,
that is, in a context where food is likely to occur. The experiments described in this
section show that Pavlovian stimuli can modulate the strength of instrumental
responding. They also show that there are at least two ways in which this influence
takes place.
Motivational influences
Konorski (1967), it should be recalled from Chapter 2, believed that a CS can excite
an affective representation of the US that was responsible for arousing a preparatory
CR. He further believed that a component of this CR consists of a change in the level
of activity in a motivational system. A CS for food, say, was said to increase activity
in the positive motivational system, whereas a CS for shock should excite the
negative system. If these proposals are correct, then it should be possible to alter the
strength of instrumental responding by presenting the appropriate Pavlovian CS
(see also Rescorla & Solomon, 1967).

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 31/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
110 A N I M A L L E A R N I N G & C O G N I T I O N
An experiment by Lovibond (1983), using Pavlovian–instrumental transfer
design, provides good support for this prediction. Hungry rabbits were first trained
to operate a lever with their snouts to receive a squirt of sucrose into the mouth. The
levers were then withdrawn for a number of sessions of Pavlovian conditioning in
which a clicker that lasted for 10 seconds signaled the delivery of sucrose. In a final
test stage, subjects were again able to press the lever and, as they were doing so, the
clicker was occasionally operated. The effect of this appetitive CS was to increasethe rate of lever pressing both during its presence and for a short while after it was
turned off. A similar effect has also been reported in a study using an aversive US.
Rescorla and LoLordo (1965) found that the presentation of a CS previously paired
with shock enhanced the rate at which dogs responded to avoid shock.
In addition to explaining the findings that have just been described, a further
advantage of dual-system theories of motivation is that they are able to account for
many of the effects of exposing animals simultaneously to both appetitive and aversive
stimuli. For example, an animal may be exposed to one stimulus that signals reward
and another indicating danger. In these circumstances, instead of the two systems
working independently, they are assumed to be connected by mutually inhibitory links,
so that activity in one will inhibit the other (Dickinson & Pearce, 1977).To understand this relationship, consider the effect of presenting a signal for
shock to a rat while it is lever pressing for food. Prior to the signal, the level of
activity in the positive system will be solely responsible for the rate of pressing.
When the aversive CS is presented, it will arouse the negative system. The existence
of the inhibitory link will then allow the negative system to suppress activity in the
positive system and weaken instrumental responding. As soon as the aversive CS is
turned off, the inhibition will be removed and the original response rate restored. By
assuming the existence of inhibitory links, dual-system theories can provide a very
simple explanation for conditioned suppression. It occurs because the aversive CS
reduces the positive motivational support for the instrumental response.
Response-cueing properties of Pavlovian CRs
In addition to modulating activity in motivational systems, Pavlovian stimuli can
influence instrumental responding through a response-cueing process (Trapold &
Overmier, 1972). To demonstrate this point we shall consider an experiment by
Colwill and Rescorla (1988), which is very similar in design to an earlier study by
Kruse, Overmier, Konz, and Rokke (1983).
In the first stage of the experiment, hungry rats received Pavlovian conditioning
in which US1 was occasionally delivered during a 30-second CS. Training was then
given, in separate sessions, in which R1 produced US1 and R2 produced US2. The
two responses were chain pulling and lever pressing, and the two reinforcers were
food pellets and sucrose solution. For the test stage, animals had the opportunity forthe first time to perform R1 and R2 in the presence of the CS, but neither response led
to a reinforcer. As Figure 4.10 shows, R1 was performed more vigorously than R2.
The first point to note is that it is not possible to explain these findings by
appealing to the motivational properties of the CS. The CS should, of course,
enhance the level of activity in the positive system. But this increase in activity
should then invigorate R1 to exactly the same extent as R2 because the motivational
support for both responses will be provided by the same, positive, system.
In developing an alternative explanation for the findings by Colwill and Rescorla
(1988), note that instrumental conditioning with the two responses was conducted in
K E Y T E R M
Pavlovian–instrumental
transfer
Training in which a CSis paired with a USand then the CS ispresented while thesubject is performingan instrumentalresponse.
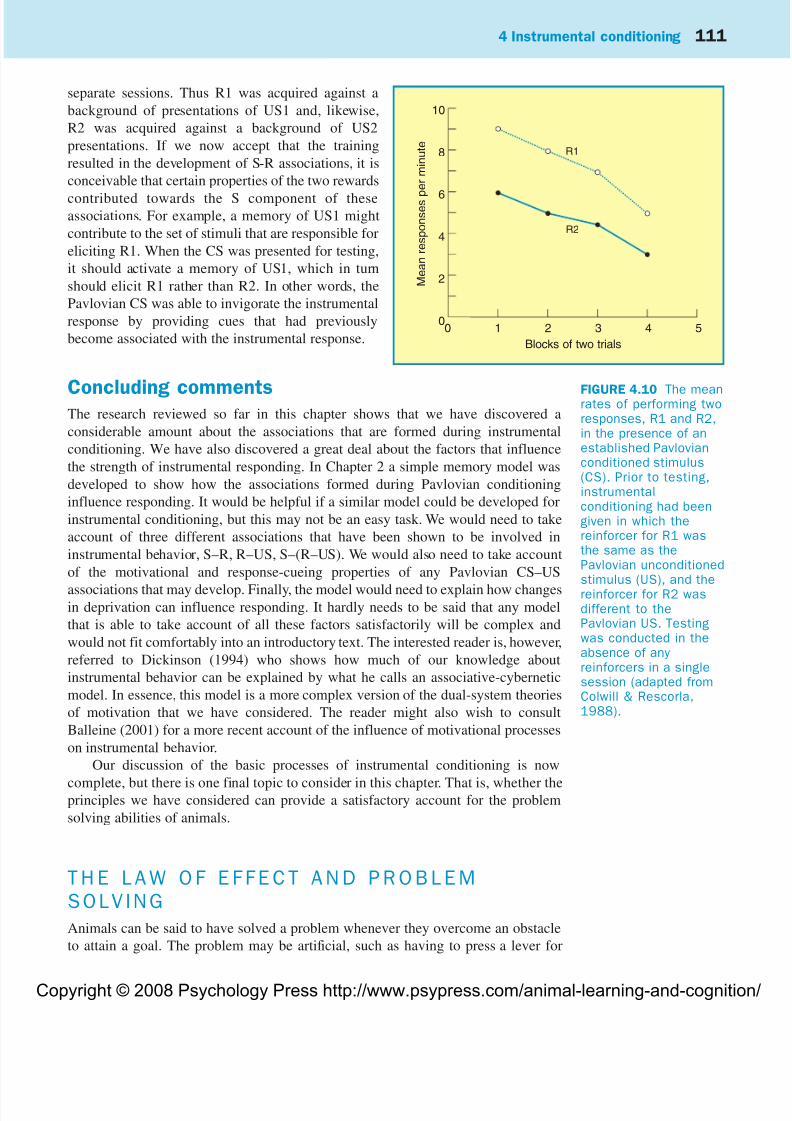
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 32/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 111
separate sessions. Thus R1 was acquired against a
background of presentations of US1 and, likewise,
R2 was acquired against a background of US2
presentations. If we now accept that the training
resulted in the development of S-R associations, it is
conceivable that certain properties of the two rewards
contributed towards the S component of theseassociations. For example, a memory of US1 might
contribute to the set of stimuli that are responsible for
eliciting R1. When the CS was presented for testing,
it should activate a memory of US1, which in turn
should elicit R1 rather than R2. In other words, the
Pavlovian CS was able to invigorate the instrumental
response by providing cues that had previously
become associated with the instrumental response.
Concluding commentsThe research reviewed so far in this chapter shows that we have discovered a
considerable amount about the associations that are formed during instrumental
conditioning. We have also discovered a great deal about the factors that influence
the strength of instrumental responding. In Chapter 2 a simple memory model was
developed to show how the associations formed during Pavlovian conditioning
influence responding. It would be helpful if a similar model could be developed for
instrumental conditioning, but this may not be an easy task. We would need to take
account of three different associations that have been shown to be involved in
instrumental behavior, S–R, R–US, S–(R–US). We would also need to take account
of the motivational and response-cueing properties of any Pavlovian CS–US
associations that may develop. Finally, the model would need to explain how changesin deprivation can influence responding. It hardly needs to be said that any model
that is able to take account of all these factors satisfactorily will be complex and
would not fit comfortably into an introductory text. The interested reader is, however,
referred to Dickinson (1994) who shows how much of our knowledge about
instrumental behavior can be explained by what he calls an associative-cybernetic
model. In essence, this model is a more complex version of the dual-system theories
of motivation that we have considered. The reader might also wish to consult
Balleine (2001) for a more recent account of the influence of motivational processes
on instrumental behavior.
Our discussion of the basic processes of instrumental conditioning is now
complete, but there is one final topic to consider in this chapter. That is, whether theprinciples we have considered can provide a satisfactory account for the problem
solving abilities of animals.
THE LAW OF EF FECT AND PROBLEM
SOLVING
Animals can be said to have solved a problem whenever they overcome an obstacle
to attain a goal. The problem may be artificial, such as having to press a lever for
FIGURE 4.10 The meanrates of performing tworesponses, R1 and R2,in the presence of anestablished Pavlovianconditioned stimulus(CS). Prior to testing,instrumentalconditioning had beengiven in which thereinforcer for R1 wasthe same as thePavlovian unconditionedstimulus (US), and thereinforcer for R2 wasdifferent to thePavlovian US. Testingwas conducted in theabsence of anyreinforcers in a singlesession (adapted fromColwill & Rescorla,1988).
10
8
6
4
2
0
Blocks of two trials
M e a n r e s p o n s e s p
e r m i n u t e
R2
R1
0 1 2 3 4 5

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 33/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
112 A N I M A L L E A R N I N G & C O G N I T I O N
reward, or it might be one that occurs naturally, such as having to locate a new source
of food. Early studies of problem solving in animals were conducted by means of
collecting anecdotes, but this unsatisfactory method was soon replaced by
experimental tests in the laboratory (see Chapter 1). As a result of his experiments,
Thorndike (1911) argued that despite the range of potential problems that can
confront an animal, they are all solved in the same manner. Animals are assumed to
behave randomly until by trial and error the correct response is made and reward isforthcoming. To capture this idea, Thorndike (1911) proposed the Law of Effect,
which stipulates that one effect of reward is to strengthen the accidentally occurring
response and to make its occurrence more likely in the future. This account may
explain adequately the way cats learn to escape from puzzle boxes, but is it suitable
for all aspects of problem solving? A number of researchers have argued that animals
are more sophisticated at solving problems than is implied by the Law of Effect. It has
been suggested that they are able to solve problems through insight. It has also been
suggested that animals can solve problems because they have an understanding of the
causal properties of the objects in their environment or, as it is sometimes described,
an understanding of folk physics. We shall consider each of these possibilities.
Insight
An early objector to Thorndike’s (1911) account of problem solving was Kohler
(1925). Thorndike’s experiments were so restrictive, he argued, that they prevented
animals from revealing their capacity to solve problems by any means other than the
most simple. Kohler spent the First World War on the
Canary Islands, where he conducted a number of
studies that were meant to reveal sophisticated
intellectual processes in animals. He is best known
for experiments that, he claimed, demonstrate the
importance of insight in problem solving. Many of
his findings are described in his book The mentality
of apes, which documents some remarkable feats of
problem solving by chimpanzees and other animals.
Two examples should be sufficient to give an
indication of his methodology. These example
involve Sultan (Figure 4.11), whom Kohler (1925)
regarded as the brightest of his chimpanzees. On one
occasion Sultan, was in a cage in which there was
also a small stick. Outside the cage was a longer
stick, which was beyond Sultan’s reach, and even
further away was a reward of fruit (p. 151):
Sultan tries to reach the fruit with the smaller of
the sticks. Not succeeding, he tries a piece of
wire that projects from the netting in his cage,
but that, too, is in vain. Then he gazes about him
(there are always in the course of these tests
some long pauses, during which the animal
scrutinizes the whole visible area). He suddenly
picks up the little stick once more,
K E Y T E R M
Insight
An abrupt change in
behavior that leads toa problem beingsolved. The change inbehavior is sometimesattributed to a periodof thought followed bya flash of inspiration.
FIGURE 4.11 Sultan stacking boxes in an attempt toreach a banana (drawing based on Kohler, 1956).

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 34/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 113
goes to the bars directly opposite to the long stick, scratches it towards him
with the auxiliary, seizes it and goes with it to the point opposite the objective
which he secures. From the moment that his eyes fell upon the long stick, his
procedure forms one consecutive whole.
In the other study, Kohler (1925) hung a piece of fruit from the ceiling of a cage
housing six apes, including Sultan. There was a wooden box in the cage (p. 41):
All six apes vainly endeavored to reach the fruit by leaping up from the ground.
Sultan soon relinquished this attempt, paced restlessly up and down, suddenly
stood still in front of the box, seized it, tipped it hastily straight towards the
objective, but began to climb upon it at a (horizontal) distance of 1 ⁄ 2 meter and
springing upwards with all his force, tore down the banana.
In both examples there is a period when the animal responds incorrectly; this is
then followed by activity that, as it is reported, suggests that the solution to the
problem has suddenly occurred to the subject. There is certainly no hint in thesereports that the problem was solved by trial and error. Does this mean, then, that
Kohler (1925) was correct in his criticism of Thorndike’s (1911) theorizing?
A problem with interpreting Kohler’s (1925) findings is that all of the apes had
played with boxes and sticks prior to the studies just described. The absence of
trial-and-error responding may thus have been due to the previous experience of the
animals. Sultan may, by accident, have learned about the consequences of jumping
from boxes in earlier sessions, and he was perhaps doing no more than acting on the
basis of his previous trial-and-error learning. This criticism of Kohler’s (1925) work
is by no means original. Birch (1945) and Schiller (1952) have both suggested that
without prior experience with sticks and so forth, there is very little reason for
believing that apes can solve Kohler’s problems in the manner just described.
The absence of trial-and-error responsesin Kohler’s (1925)findings might havebeen due to the factthat most apes wouldhave had priorexperience of playingwith sticks.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 35/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
114 A N I M A L L E A R N I N G & C O G N I T I O N
An amusing experiment by Epstein, Kirshnit, Lanza, and Rubin (1984) also
shows the importance of past experience in problem solving and, at the same time,
raises some important issues concerning the intellectual abilities of animals. Pigeons
were given two different types of training. They were rewarded with food for pushing
a box towards a spot randomly located at the base of a wall of the test chamber.
Pushing in the absence of the spot was never rewarded. They were also trained to
stand on the box when it was fixed to the floor and peck for food at a plastic bananasuspended from the ceiling. Attempts to peck the banana when not standing on the
box were never rewarded. Finally, on a test session they were confronted with a novel
situation in which the banana was suspended from the ceiling and the box was placed
some distance from beneath it. Epstein et al. (1984) report that (p. 61):
At first each pigeon appeared to be “confused”; it stretched and turned beneath
the banana, looked back and forth from banana to box, and so on. Then each
subject began rather suddenly to push the box in what was clearly the direction
of the banana. Each subject sighted the banana as it pushed and readjusted the
box as necessary to move it towards the banana. Each subject stopped pushing
it in the appropriate place, climbed and pecked the banana. This quite
remarkable performance was achieved by one bird in 49 sec, which compares
very favorably with the 5 min it took Sultan to solve his similar problem.
There can be no doubt from this study that the prior training of the pigeon played
an important role in helping it solve the problem. Even so, the study clearly reveals
that the pigeons performed on the test session in a manner that extends beyond
trial-and-error responding. The act of pecking the banana might have been acquired
by trial-and-error learning, and so, too, might the act of moving the box around. But
the way in which the box was moved to below the banana does not seem to be
compatible with this analysis.
The description by Epstein et al. (1984) of the pigeons’behavior bears a striking
similarity to Kohler’s (1925) account of Sultan’s reaction to the similar problem. It
might be thought, therefore, that it would be appropriate to account for the pigeons’
success in terms of insight. In truth, this would not be a particularly useful approach
as it really does not offer an account of the way in which the problem was solved.
Other than indicating that the problem was solved suddenly, and not by trial and
error, the term “insight” adds little else to our understanding of these results.
I regret that I find it impossible to offer, with confidence, any explanation for the
findings by Epstein et al. (1984). But one possibility is that during their training with the
blue spot, pigeons learned that certain responses moved the box towards the spot, and
that the box by the spot was a signal for food. The combination of these associations
would then result in them pushing the box towards the spot. During their training with
the banana, one of the things the pigeons may have learned is that the banana is
associated with food. Then, for the test session, although they would be unable to push
the box towards the blue spot, generalization from their previous training might result in
them pushing the box in the direction of another signal for food, the banana.
Causal inference and folk physics
The term “insight” is now rarely used in discussions of problem solving by animals.
As an alternative, it has been proposed that animals have some understanding of

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 36/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 115
causal relationships and that they can draw inferences based on this understanding to
solve problems. When a problem is encountered, therefore, animals are believed to
solve it through reasoning based on their understanding of the physical and causal
properties of the objects at their disposal. To take the example of Sultan joining two
sticks together to reach food, if he understood that this action would create a longer
stick that would allow him to reach further, he would then be able to solve the
problem in a manner that is considerably more sophisticated than relying on trial anderror. Of course, we have just seen that the studies by Kohler (1925) do not provide
evidence that animals can solve problems in this way, but the results from other
experiments have been taken as evidence that animals are capable of making causal
inferences. The following discussion will focus separately on this work with
primates and birds.
Primates
Premack (1976, pp. 249–261) describes an experiment with chimpanzees in which
a single subject would be shown an array of objects similar to the one in Figure 4.12.
To gain reward, the chimpanzee was required to replace the strange shape in
the upper row with the knife from the lower row. The choice of the knife was
intended to reveal that the ape understood this object causes an apple to be cut in
half. Two of the four subjects that were tested performed consistently well on this
task. They received a novel problem on each trial, thus their success could not
depend on them solving the problem by associating a given choice with a particular
array of objects. An alternative explanation for the problem shown in Figure 4.12 is
that the apes had repeatedly seen an apple being cut with a knife and they may have
learned to select the object from the lower row that was most strongly associated
with the one from the upper row. Although this explanation will work for many of
the test trials, Premack (1976) argues there were certain occasions where it provides
an implausible explanation for the successful choice. For instance, one trial was
similar to that shown in Figure 4.12 except the apple was replaced by a whole ball
and a ball cut into pieces. Even though the subjects had rarely seen a knife and a
ball together, they still made the correct choice (see also Premack & Premack, 1994,
pp. 354–357).
FIGURE 4.12 Sketchof an array of objectsused by Premack(1976) to test for
causal inference inchimpanzees (adaptedfrom Premack, 1976).

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 37/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
116 A N I M A L L E A R N I N G & C O G N I T I O N
The findings from the experiment are encouraging, but there is some doubt about
how they should be interpreted. The two apes who performed well on the task had
received extensive training on other tasks, and one of them, Sarah, had even been taught
an artificial language (see Chapter 13). Perhaps the extensive training given to the
chimpanzees resulted in them acquiring a rich array of associations that helped them
perform correctly on the tests, without the need to understand the relevant causal
relationships. It is also possible that because of the similarity between the shapes of anapple and a ball, stimulus generalization rather than a causal inference was responsible
for the selection of the knife during the test with the ball. To properly evaluate the results
from the experiment it would be necessary to have a complete account of the training
that was given before it started. It would also be important to have a full description of
the method and results from the experiment itself. Unfortunately, the information that is
available is rather brief and the reader is left in some doubt as to how Premack’s findings
should be interpreted. Another possibility is that because of their extensive training, the
two chimpanzees were able to appreciate causal relationships and to draw inferences
from them in a way that is not open to relatively naïve chimpanzees. Once again, there
is insufficient information available for this possibility to be assessed. There is no
denying that the experiment by Premack has revealed some intriguing findings but,before its full significance can be appreciated, additional experiments are needed in
order to pursue the issues that have just been raised.
Rather than study causal inference, some researchers have investigated what they
call “folk physics”, which refers to a common-sense appreciation of the causal
properties of objects in the environment. Problems could then be solved by drawing
inferences from the understanding about these properties. Povinelli (2000) has
conducted a thorough series of experiments to explore whether chimpanzees make
use of folk physics in problem solving, and they point to rather different conclusions
to those drawn by Premack (1976).
In one of Povinelli’s experiments, chimpanzees were confronted with a clear
tube that contained a peanut. To retrieve the food, they were required to push it outof the tube with a stick. Once they had mastered this skill they were given the same
task but this time there was a trap in the tube. Pushing the stick in one direction
caused the peanut to fall out of the tube, pushing it in the other direction caused the
peanut to fall in the trap where it was inaccessible. A sketch of this simple apparatus
can be seen in Figure 4.13. Three of the four chimpanzees that were given this task
never mastered it, and the fourth chimpanzee came to terms with it only after many
practice trials. The conclusion to be drawn from this study is that chimpanzees did
not have any appreciation of the problem created by the presence of the trap, that is,
they lacked an understanding provided by folk physics concerning the properties of
traps. Instead, the eventual success of the single chimpanzee can be explained by
assuming that she learned through trial and error how to avoid pushing the food into
the trap by inserting the stick into the end of the tube that was furthest from the
peanut. A similar failure to find evidence of successful performance on the same
problem has been found with capuchin monkeys (Visalberghi & Limongelli, 1994).
Povinelli (2000) cites a total of 27 experiments, using a variety of tests, all of which
show that chimpanzees have a complete lack of understanding of the physical
properties of the problems that confront them. The interested reader might also refer
to an article by Nissani (2006), which reports a failure by elephants to display causal
reasoning in a tool-use task.
The negative results that have just been cited make it all the more important to
determine whether Premack (1976) was correct in claiming that chimpanzees are

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 38/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 117
capable of causal inference. For the present, it is perhaps wisest to keep an open mind
about the capacity of primates to refer to folk physics when solving problems, but
what about other species? Clayton and Dickinson (2006) have suggested that the
most compelling evidence that animals have an appreciation of folk physics can be
found in certain species of birds.
Birds
In one study, Seed, Tebbich, Emery, and Clayton (2006) presented a group of naïve
rooks a version of the trap problem used by Povinelli (2000; described above). When
first confronted with this problem, the direction in which the birds pushed the foodwas determined by chance but, as training progressed, they showed a marked
improvement in avoiding the trap. To test whether this improvement reflected
anything more than learning through trial and error, the birds were given a new
problem where performance was not expected to be influenced by the effects of any
prior trial-and-error learning. Six out of seven birds performed poorly on the
new problem, but one bird performed extremely well from the outset. As the authors
point out, it is hard to know what conclusions to draw when one bird passes a test
that six others have failed, but the performance of the one successful bird encourages
the view that future research with rooks might reveal promising results.
Another species of bird that might possess an understanding of folk physics is
the raven. Heinrich (2000; see also Heinrich and Bugnyar, 2005) describes a seriesof experiments with hand-reared ravens in which the birds were presented with a
piece of meat hanging from a perch (see the left-hand side of Figure 4.14). The meat
could not be obtained by flying towards it and clasping it in the beak. Instead, to
reach the meat some birds settled on the perch where the string was attached and
grasped the string below the perch with their beak and pulled it upwards. To stop the
meat falling back they placed a foot on the string and then let it drop from their beak
whereupon they bent down to grasp again the string below the perch. This operation
was repeated until the meat was near enough to be grasped directly with the beak. In
another test, ravens were confronted with the arrangement shown in the right-hand
side of Figure 4.14. On this occasion, meat could be retrieved by standing on the
Failure
Success
FIGURE 4.13 Right: Diagram of the apparatus used by Povinelli (2000) and by Visalberghi and Limongelli(1994) to test whether an animal will push a peanut in the direction that ensures it does not fall into a trap.From Visalberghi and Limongelli, 1994. Copyright © 1994 American Psychological Association. Reproducedwith permission. Left: A monkey about to attempt to retrieve a nut from the apparatus. Photograph byElisabetta Visalberghi.
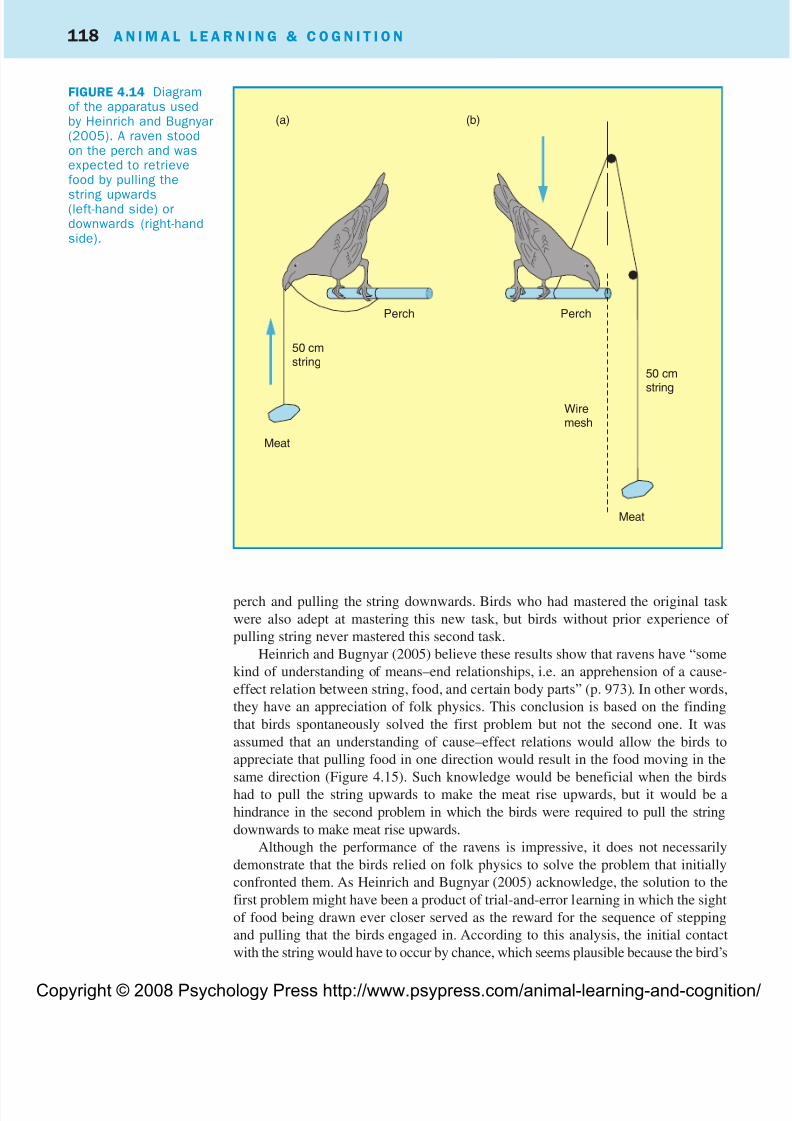
8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 39/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
118 A N I M A L L E A R N I N G & C O G N I T I O N
perch and pulling the string downwards. Birds who had mastered the original task
were also adept at mastering this new task, but birds without prior experience of
pulling string never mastered this second task.
Heinrich and Bugnyar (2005) believe these results show that ravens have “some
kind of understanding of means–end relationships, i.e. an apprehension of a cause-
effect relation between string, food, and certain body parts” (p. 973). In other words,
they have an appreciation of folk physics. This conclusion is based on the finding
that birds spontaneously solved the first problem but not the second one. It was
assumed that an understanding of cause–effect relations would allow the birds to
appreciate that pulling food in one direction would result in the food moving in the
same direction (Figure 4.15). Such knowledge would be beneficial when the birds
had to pull the string upwards to make the meat rise upwards, but it would be a
hindrance in the second problem in which the birds were required to pull the string
downwards to make meat rise upwards.
Although the performance of the ravens is impressive, it does not necessarily
demonstrate that the birds relied on folk physics to solve the problem that initially
confronted them. As Heinrich and Bugnyar (2005) acknowledge, the solution to the
first problem might have been a product of trial-and-error learning in which the sight
of food being drawn ever closer served as the reward for the sequence of stepping
and pulling that the birds engaged in. According to this analysis, the initial contact
with the string would have to occur by chance, which seems plausible because the bird’s
FIGURE 4.14 Diagramof the apparatus usedby Heinrich and Bugnyar(2005). A raven stoodon the perch and wasexpected to retrievefood by pulling the
string upwards(left-hand side) ordownwards (right-handside).
(a) (b)
PerchPerch
Meat
Meat
50 cm
stringWiremesh
50 cmstring

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 40/52

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 41/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
120 A N I M A L L E A R N I N G & C O G N I T I O N
They use long thin strips of leaf with barbs running down one edge to draw prey from
cracks and holes in trees. It is not known if this ability is learned or inherited, but if
the conclusions drawn from the study by Tebbich et al. (2001) have any generality,
then it will be a mixture of the two.
In the experiment by Weir et al. (2002), a male and a female crow were expected
to retrieve a piece of food from a bucket with a handle that was placed in a clear,
vertical tube (Figure 4.16). The tube was so deep that it was impossible for the birdsto reach the handle of the bucket with their beaks. A piece of straight wire and a piece
of wire with a hook at one end were placed near the tube and the birds were expected
to use the hooked wire to lift the bucket out of the tube by its handle. On one
occasion, the male crow selected the hooked wire, which left the female with the
straight wire. She picked up one end in her beak inserted the other end in a small
opening and then bent the wire to create a hook which was of a suitable size to enable
her to lift the bucket from the tube. A video clip of this sequence can be seen by
going to the following web address http://www.sciencemag.org/cgi/content/full/
297/5583/981/DC1. As one watches the female crow bend the wire it is hard not to
agree with the authors that she was deliberately modifying the wire to create a tool,
and that this modification relied on an understanding of “folk physics” and causality.However, appearances can be deceptive, and it would be a mistake to ignore the
possibility that the bird’s behavior was a consequence of less sophisticated processes.
As with the woodpecker finches, it is possible that the skill displayed by the female
crow was a consequence of the interaction between inherited tendencies and learning
based on prior experience with sticks, twigs, and so on. Before this explanation can
be rejected with complete confidence, more needs to be known about the
development of tool use in New Caledonian crows in their natural habitat. It is also
a pity that rather little is known about the prior experiences of the bird in question,
which was captured in the wild.
The results from these tests for an understanding of folk physics in birds can
perhaps most fairly be described as ambiguous in their theoretical significance.On the one hand, it is possible to explain most, if not all, of them in terms of
the trial-and-error principles advocated by Thorndike (1911) almost a century ago.
FIGURE 4.16 A NewCaledonian crow liftinga bucket out of a tubein order to retrieve foodin an experiment byWeir, et al., (2002).Photograph by AlexWeir © Behavioural
Ecology ResearchGroup, University of Oxford.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 42/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
4 Instrumental conditioning 121
However, a critic of this type of explanation would argue that it is so versatile that it
can explain almost any result that is obtained. Moreover, although the trial-and-error
explanations we have considered may be plausible, there is no evidence to confirm
that they are necessarily correct. On the other hand, some of the behavior that has
been discovered with birds is so impressive to watch that many researchers find it
hard to believe they lack any understanding of the problem that confronts them.
For myself, my sympathies rest with an analysis of problem solving in terms of trial-and-error learning. The great advantage of this explanation is that it is based on
firmly established principles of associative learning. Problem solving relies on the
capacity for rewards to strengthen associations involving responses, and the transfer
of the solution from one problem to another is explained through stimulus
generalization. By contrast, much less is known about the mental processes that
would permit animals to make causal inferences, or to reason using folk physics.
Seed et al. (2006) note briefly that these processes may involve the capacity for
acquiring abstract rules about simple, physical properties of the environment. Given
such a proposal two questions arise: first, how is such knowledge about the
environment acquired, and second, are animals capable of abstract thought? As far as
I am aware, no-one has offered an answer to the first question and, as for the secondquestion, we shall see in later chapters that whether or not animals are capable of
abstract thought is a contentious issue that has yet to be fully resolved.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 43/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Subject index
abstract categories 179–80, 189, 358abstract mental codes 325, 359,
368, 371abstract thought 121acquired distinctiveness 157–8,
157 , 158acquired equivalence 157–8, 157 , 158active memory 220, 221adaptability 12–13addition 249–50, 250African claw-toed frog 216, 216 African grey parrot
and category formation 172,182, 182
and communication 351–3and mimicry 304and mirror-use 322, 323number skills of 248–50, 248–9,
251, 252aggression, displays of 337–8, 337 AI see artificial intelligenceair pressure 285Akeakamai (dolphin) 353–5alarm calls 331, 332–5, 336, 337albatross 286Alex (parrot) 248–50, 248–9, 251,
252, 351–3, 368alley running 94, 98, 106
and extinction 131–3, 132and navigation 267–8and number skills 245–6, 246
alpha conditioning 44American goldfinch 331American sign language 341–2,
349–50, 349amnesia, drug-induced 224–5amodal representation 240–1, 241amygdala 21, 224analogical reasoning 187, 188, 368, 371
anecdotes 23, 112animal intelligence 2–33
and brain size 10–12, 10, 361–4definition 12–16, 362–3distribution 4–12, 4, 360–71ecological view 228, 229, 231and evolution 369–71general process view 230–1historical background 22–33null hypothesis of 261, 364–9reasons to study 16–20research methods 20–2
animal kingdom 4–5animal welfare 19–20anisomycin 224–5anthropomorphism 23–4, 168–9,
262–3ants 265–8, 267 apes 16
and category formation 182and communication 329–50, 335–6,
340, 356–7and language 116, 329–50, 356–7and self-recognition 369
vocalizations 339–40, 340see also chimpanzee; gorilla;
orang-utan Aplysia californica (marine snail) 43,
43–5, 46, 47, 149, 263Arctic tern 289artificial intelligence (AI) 19associability/conditionability 67–8,
74–5, 91associative competition 101–2, 101associative learning 35–61, 123, 143
and attention 365–6conditioning techniques 26–42CRs 55–61and deception studies 313, 315definition 35and evolution 370–1memory model of 46–9, 46–9, 59nature of 42–9and the null hypothesis 364–6and problem solving 121and the reflexive nature of the CR
60–1and stimulus-stimulus learning
49–52and surprise 365–6and time 233
and US representations 52–5see also instrumental (operant)
conditioning; Pavlovian(classical) conditioning
associative strength 64–71, 66 , 68, 83,88–9, 91, 101, 102
and category formation 175–7, 179and discrimination learning 152–3equations 65–70, 83, 87, 88, 89and extinction 125–31, 134, 142negative 127–8, 130, 176and stimulus significance 83–4
associative-cybernetic model 111asymptote 36attention 365–6, 371
automatic 86and conditioning 63, 74–91, 86controlled/deliberate 86and the CS 63, 76, 77, 78–80dual 86multiple modes of 86
audience effects 335–6auditory templates 332Austin (chimpanzee) 345, 346, 357
autoshaping 37, 37 , 38, 60, 126, 127 omission schedules 60
baboonand deception 314, 314, 315predatory nature 333, 336
Bach, J.S. 172–3back-propagation networks 164–5
two-layer 164bees
light detection 285navigation 269–71, 271, 280see also bumble-bee; honey-bee
behaviorists 28bicoordinate navigation 288bidirectional control 308–9, 308–9,
310biological significance 80–1,
84–5, 102birds
evolution 6and imitation 304–5, 305and navigation 275and problem solving 117–21,
118–20short-term memory of 228,
229–30
song birds 331–2, 363see also specific species
black and white images 173–4black-capped chickadee 305, 363, 363blackbird 302bliss points 104blocking 53, 63–4, 63, 69–70, 78,
365–6LePelley on 91and Mackintosh’s theory 83–4and navigation 293, 294and the Pearce–Hall model 88–9
Note: Page numbers in italic refer to information contained in tables and diagrams.

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 44/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
412 Subject index
blocking (Continued )and the Rescorla–Wagner model
72–3, 73blue tit 230, 304–5body concept 323–4bombykol 265bonobo, and communication 343–5,
344, 346, 347–8, 348, 349, 357bottle-nosed dolphin
and communication 338, 353–6,354–5
and learning set studies 362brain 20–1
human 19size 10–12, 10, 11, 361–4
brain size-body ratio 10–11, 11, 361brown bear 266budgerigar 309–10, 367bumble-bee 298Burmese jungle fowl 298
cache 226canary 363capuchin monkey
and category formation 184and diet selection 300and problem solving 116, 117 see also cebus monkey
carp 173cat
and communication 328and the puzzle box escape task
26–7, 26–7 , 28category formation 170–89, 358
abstract categories 179–80, 189,358
categories as concepts 179–80examples of 171–3exemplar theories of 177–8, 178feature theory of 173–9and knowledge representation
188–9prototype theory of 178–9relationships as categories 180–7theories of 173–9
causal inference 114–21, 115cebus monkey
and imitation 312, 312and serial order 256–7, 256–7 see also capuchin monkey
cephalization index (K) 11–12, 12,13, 361
chaffinch 331chain pulling tasks 95, 97, 110, 141, 253cheremes 341chick, and discrimination learning
159–61, 159–61chicken
and communication 331and discrimination learning 150
chimpanzee 4, 368, 371and abstract representation 189and category formation 179–81,
180, 186–7, 188, 189and communication 24, 116, 327,
336, 339–50, 342–4, 357, 359dominant 317–18
and emulation learning 310and imitation 306, 307, 310,
311, 367and instrumental conditioning
105, 106lexigram use 343–5, 343–4, 346and mimicry 303and number skills 245, 245, 248,
250, 250, 252and plastic token use 342–3, 343and problem solving 112–17,
112–13, 115, 117 and self-recognition 319–24,
320, 322short-term memory of 228and social learning 367subordinate 317–18and theory of mind 312–13,
314–18, 316 , 317 , 324and transitive inference
259–60, 262vocal tracts 340, 340welfare issues 19see also bonobo
chinchilla 172, 172chunking 255–6circadian rhythms 233–4
circannual rhythms (internalcalendars) 291
Clark’s nutcracker 214, 228, 230,272–3, 294, 370
Clever Hans 16, 243, 243clock-shift experiment 288coal tit 230cockroach 233–4, 262–3cognition, and language 358–9cognitive maps 276–80, 330, 369–70color perception 173–4communication 326–59, 369
definition 327
development of 330–2honey-bees and 328–30, 328–30,331, 338–9
as innate process 330–3, 336, 356and intention 335–6and interpretation of the signal
330–1as learnt process 330–3, 336, 337and representation 334–5and the significance of signals
334–5vervet monkeys and 332–7, 333–4see also language
compass bearings 269–70, 271, 272compound stimuli 68–73, 83–4, 88–9,
126–9, 151–3, 160, 176–7concepts, categories as 179–80concrete codes 188, 371conditioned emotional response (CER)
see conditioned suppression
conditioned inhibition see inhibitoryconditioning
conditioned reinforcement 105–6, 105conditioned response (CR) 29, 35,
39–40, 44, 55–60, 285air pressure as 285and autoshaping 37and blocking 63compensatory 56–8, 58consummatory 55–6and discrimination learning 149,
155, 156, 164, 165and eye-blink conditioning 36–7and fear of predators 302influence of the CS on 58–9and inhibitory conditioning 42and the memory model of
conditioning 47preparatory 56reflexive nature of 60–1and representation of the US 109response-cueing properties
110–11, 111and stimulus-stimulus
conditioning 50–2strength of 64, 65, 70, 71, 73
conditioned stimulus (CS) 29, 30,
32–3, 123associability/conditionability of
67–8, 75associative strength 125–31,
134, 142and attention 63, 76, 77, 78–80aversive 110and compensatory CRs 57, 58compound 68–73, 83–4, 88–9,
126–9, 151–3, 160and conditioned suppression 38and discrimination learning 162and excitatory conditioning 35,
36–7, 38, 39, 40and extinction 124, 125–30, 134–5,136–46
and eye-blink conditioning 36–7and fear of predators 302and imitation 305influence on the CR 58–9inhibitory 40, 41–2, 217and instrumental conditioning 106,
109–11intensity 67–8, 68and long-term memory
studies 216, 217

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 45/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Subject index 413
memory model of 46–8, 59and the nature of US representations
52–4neural mechanisms of 43–5protection from extinction 126–8and the reflexive nature of the CR
60–1
and the renewal effect 136–8single 64–8spontaneous recovery 134–6and stimulus-stimulus conditioning
50–2and taste aversion conditioning 39and timing 242and trace conditioning 194see also CS–no–US association;
CS–R association; CS–USassociation/contingency
conditioned suppression (conditionedemotional response) 37–8, 38,41, 42, 63, 67, 67–8, 71–2, 72,85, 85, 89–90, 89
conditioningalpha 44and attention 63, 74–91, 86compound 68–73, 83–4, 88–9,
126–9, 151–3, 160excitatory 35–40, 42–5, 44–5,
52–4, 72–3eye-blink 36–7, 36–7 , 53–4, 64,
64, 68–9, 69, 124, 125,135, 135
information-processing model of 46, 46
instrumental 92–121observational 302, 308, 309second-order (higher-order) 50–2,
51, 185–7, 186 serial 49–50single CS 64–8and surprise 62–74trace 194–6, 195–6 see also inhibitory conditioning;
instrumental (operant)conditioning; Pavlovian(classical) conditioning; tasteaversion conditioning
conditioning techniques 26–42autoshaping 37, 37 , 38conditioned suppression 37–8, 38control groups 39–40, 40excitatory conditioning 35, 36–40inhibitory conditioning 35, 40–2
configural cues 153configural theory of discrimination
learning 155–7, 156 connectionist models
of discrimination learning 161–6,162–6
exemplar-based networks 165–6, 165
hidden layers 165multi-layer networks 164–5, 165single-layer networks 162–4, 162–4of time 241
conscious thought 210–11consolidation theory (rehearsal theory)
of long-term memory 218–20,
218, 225conspecifics 300, 324, 335context 72, 138, 229context-stimulus associations
78–80, 90–1contextual variables 15contiguity 31, 63, 98, 98continuous reinforcement 130–1,
131–2, 134, 134, 145–6, 145control groups 39–40, 40
truly random control 40cooperation, and language 338–9copying behavior 297, 298, 301,
302–12, 324mimicry 303–4, 312, 324see also imitation
coriolis force 288corvids 181, 182, 368counting 233, 243, 245–9, 251–2
cardinal principle 252one–one principle 252stable-ordering principle 252
cow 229coyote 58–9CS–no–US association 135–6CS–R (response) association 364–5
inhibitory 140, 142
CS–US association/contingency 71–2,88–90, 91, 94–5, 99, 111,346, 364
associative strength 64–71, 83, 88and discrimination learning 162and extinction 123, 136–8, 138–40and inhibitory connections
136–8, 142and learned irrelevance 85–6, 90negative 71–2positive 71and stimulus significance 83–4, 85–7and the strength of the CR 64
zero 71
dark-eyed juncos 363dead reckoning/path integration 266–9,
269–70, 278, 280, 293, 367decay theory 202–3deception 313–15, 314delay, gradient of 98, 98delay conditioning 194delayed matching to sample (DMTS)
197–9, 228and decay theory 202–3and forgetting 200, 201–3, 204, 205
density discrimination 167–9,167 , 168
deprivation 106–109, 108, 111detours 276–8, 277 diana monkeys 335diet selection 298–301, 299
see also food
discrimination learning 148–69,171, 178, 368
configural theory of 155–7, 156 connectionist models of 161–6,
162–6 elemental theories of 155, 156and learning sets 361–2, 362and metacognition 166–9,
167–8and relational learning 149–50Rescorla–Wagner theory of
152–5, 153–4Spence on 150–2, 155, 156stimulus preexposure 157–61theories of 149–61and theory of mind studies
317, 318and transposition 149–51
dishabituation 79, 193–4displacement (language) 337, 345,
354–6distance effect 257, 259distractors 193–4, 201–2, 203–4
surprising 203DMTS see delayed matching to
sample“Do as I do” test 307–8, 310,
311, 367dog 22–3
and classical conditioning29–30, 29
and communication 337, 337 ,350–1
and trial and error learning 25–6dolphin 16, 16 , 124
and category formation 181and communication 338–9, 353–6,
354–5and learning set studies 362and self-recognition 322, 324,
369short-term retention of 198,201, 206, 206 , 207 , 228
see also bottle-nosed dolphindrive centers 106, 107drives 30–1, 106–7drug tolerance 56–8, 58
eagle 80, 333, 334, 335ecological niches 369–70ecological view of animal intelligence
228, 229, 231EDS see extradimensional shift

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 46/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
414 Subject index
electric shocks 14, 31–2, 43–4, 50, 53,63–4, 67, 69, 71–3, 84–6, 89–90,107, 124–5, 129–30, 135
and conditioned suppression 38and eye-blink conditioning 36and inhibitory conditioning 41, 42and long-term memory studies 216,
216 , 217, 219–21, 219and navigation experiments 285and preparatory CRs 56and stimulus-stimulus learning
51–2electroconvulsive shock (ECS) 218,
218–19, 220, 224–5, 228elemental theories of discrimination
learning 155, 156elephant 36
long-term memory of 213, 213and problem solving 116and self-recognition 322, 324, 369and theory of mind 317
emulation learning 310endogenous control, of migration
290–2, 290English language, spoken 339–40,
344–5environment 7–8episodic memory 225–31episodic-like memories 226–8equilibrium theories of behavior 104European robin 291–2evolutionary development 3, 4–10,
22, 23and intelligence 369–71
tree of evolution 6, 7 excitatory conditioning 35–40, 72–3
and the nature of US representations52–4
of neural processes 42–5, 44–5exemplar effect 177exemplars 175, 177–9, 178expectancy (R–US) theorists 94–5
see also R–US associationsexperience 113–14extinction 36, 51–2, 57–8, 65–6, 66,
108, 109, 122–47, 224associative changes during 134–42
of conditioned inhibition 129–30conditions for 125–42enhanced 128–9, 128–9as generalization decrement 123–5,
125, 128, 132–4, 144, 146and an inhibitory S–R connection
140–2, 141not affecting CS–US associations
138–40, 139partial reinforcement 130–4, 131,
132, 134, 145–7, 145and Pavlovian conditioning 142–7,
143–6
protection from 126–8, 126–7 and the renewal effect 136–8,
136 , 137 and surprise 125–30trial-by-trial basis 142–7, 143–6 as unlearning 123with a US 129
extradimensional shift (EDS) 81–3, 82eye-blink conditioning 36–7, 36–7 ,
53–4, 64, 64, 68–9, 69extinction 124, 125, 135, 135
face recognition 173–4, 176, 176 , 178family trees 6, 8–9fear, of predators 301–2fear conditioning see electric shocks;
electroconvulsive shock feature theory of category formation
173–9feature-positive discrimination 151–2,
155–6Fellow (dog) 350–1fish
and category formation 173and learning 13, 15and navigation 274short-term memory of 228see also specific species
folk physics 116–21food 8, 9
food-pulling tasks 117–19, 118–19hiding/storing behavior 191, 214,
226–7, 229–30, 363, 370rewards 13–14, 14, 15, 27–8
see also diet selectionforaging behavior 300–1, 301forgetting 216–25, 228
decay theory of 202–3deliberate 204–5and proactive interference 200–1,
201, 203, 204and retroactive interference 163,
200, 201–2, 203and short-term retention 199–205
fossil record 6fruit-fly 327frustration theory 132–3, 135
Galapagos Islands 5, 6 , 119geese 268–9, 268–9, 270general process view of animal
intelligence 230–1generalization
and category formation 174, 177,178
gradients 150, 150, 156mediated 180numerosity 244, 244temporal 236–8, 237 , 240–1see also stimulus generalization
generalization decrement 37as extinction 123–5, 125, 128,
132–4, 144, 146geometric modules 274–6geometric relations 272–4,
273–4, 367geraniol 234
gerbils 268, 269–72, 270, 272ghost control 310goal directed learning 32goldfish 364–5gorilla
and communication 339and self-recognition 319, 321, 322
gradient of delay 98, 98grain selection 80–1, 81grammar 337–8, 346–50, 353–6,
355, 358of American sign language 341and lexigrams 343using plastic tokens 342
gravity 288great spotted woodpecker 191great tit 304–5, 305, 363green turtle 283, 284, 291grief 341guppy 301
habituation 75, 77, 79, 80, 91definition 192and retroactive interference 201,
203and short-term retention 192–4,
193, 201, 203
see also dishabituationhamster
and navigation 268, 277–8, 277 social behavior of 59–60
harbor seal 303heading vectors 272hedgehog 229higher vocal centre 363hippocampal place cells 280–3hippocampus 21, 363–4homing 284, 286–9
and bicoordinate navigation 288and landmark use 286
map and compass hypothesis of 287–8and olfaction 289by retracing the outward route
287, 287 Homo sapiens 5
see also human beingshoney-bee
communication (waggle dance) of 328–30, 328–30, 331, 338–9
and navigation 268short-term memory of 228sounds made by 329–30

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 47/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Subject index 415
and time perception 234, 235, 236,262
Hooker, John Lee 173 Horizon (TV programme) 338horse 16, 243, 243human beings 371
brain 19
and category formation 177, 178episodic memory of 225and the exemplar effect 177and language 327, 338, 339–40,
344–5, 356and learning 74, 86long-term memory of 213metamemory of 207–8navigation skills 266, 275and number 248and serial position effects 207see also Homo sapiens
hunger 106–9, 110“hunting by search image”
80–1, 80–1hyena 370
IDS see intradimensional shiftiguana 5, 6 imaginal codes 188imitation 303, 304–12, 324, 367–8
and bidirectional control 308–9,308–9, 310
chimpanzees and 306, 307, 310,311, 367
“Do as I do’’ test 307–8, 310, 311,367
and emulation learning 310laboratory studies of 306–10mechanisms of 311–12monkeys and 311–12, 312, 367,
368naturalistic evidence 304–6and two-action control 309–10,
309inactive memory 220indigo bunting 292information-processing 15–16, 21information-processing model
of conditioning 46, 46
of time 237–8, 237 , 239, 239,240, 241infrasound 285, 288inheritance, of problem solving
abilities 119–20inhibitory conditioning 35, 40–2,
41–3, 70–1, 73, 127–8detection 41–2extinction 129–30and latent inhibition 76and long-term memory 217and the nature of US representations
54–5, 55
innate processescommunication 330–3, 336, 356fear 302
insight 112–14instrumental (operant) conditioning
28, 30, 32–3, 92–121conditions of 97–106
and discriminatory learning 153extinction 123–8, 131–4, 141historical background 93–5and long-term memory studies
216–17and the memory model 111and mimicry 303nature of 93–7and the null hypothesis 364, 365–6Pavlovian interactions 106, 109–11and problem solving 111–21and serial order 253and vigor of performance 106–11
intellectual curiosity 16intelligence see animal intelligenceintention, and communication 335–6internal clocks 233–6, 262,
287–8, 292alternatives to 241–2
interneurons 45intertrial interval 197interval scales 250interval timing 233, 236–41, 237–41,
242, 263midpoints 238, 239–40pacemaker 241, 263
intradimensional shift (IDS) 81–3, 82
Japanese macaque monkey 300, 306Japanese quail
and imitation 309, 309and mate choice 301
jaw movement 39–40, 40
Kanzi (bonobo) 344–5, 344, 346,347–8, 348, 349, 357
kinesthetic self concept 323–4knowledge attribution 315–19knowledge representation 188–9,
358–9, 368–9
abstract mental code 325, 359,368, 371concrete mental code 188, 371self-recognition 319–24, 320–2,
368–9see also number; serial order; time
Koko (gorilla) 319, 321
Lana (chimpanzee) 343, 346, 352landmarks 293–4
and cognitive maps 278–80and compass bearings 269–70,
271, 272
in distinctively shaped environments274–6, 274–5
and geometric relations 272–4,273–4, 367
and heading vectors 272and hippocampal place
cells 281–3
and homing 286piloting with multiple 271–2,
271–2, 367piloting with single 269–71, 367retinal snapshots of 270, 271
language 327, 336–59, 371American sign language 341–2,
349–50, 349apes and 116, 329–50, 356–7arbitrariness of units 337, 338and cognition 358–9and cooperation 338–9definition 336–8discreteness of 337, 338, 345and displacement 337, 345, 354–6grammar 337–8, 341–3, 346–50,
353–6, 355, 358human 327, 338, 339–40, 344–5,
356as innate process 356and lexigrams 343–5, 343–4, 346and motivation 357and the null hypothesis 369and plastic tokens 342–3and problem solving 19productivity of 337–8, 346–7requirements for learning 356–79
and semanticity 337, 346sentences 342–3, 346–50, 352–4,
355, 356, 358spoken English 339–40, 344–5spontaneous 357training assessment (apes) 345–50training methods (apes) 339–40training methods (non-primates)
350–6language acquisition device 356–7latent inhibition 76, 79–80, 88–91,
365–6Law of Effect 27–8, 30–1, 93–4,
102–4and problem solving 111–21learned irrelevance 84, 85–6, 85, 90learning 8–9, 13–15
communication and 330–3,336, 337
definition 13emulation learning 310goal directed 32human beings and 74, 86and migration 292–3perceptual 158–61, 159–61relational 149–50

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 48/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
416 Subject index
learning (Continued )speed of 13–15, 14stimulus-stimulus 49–52and surprise 62–74trial and error 25–7, 112–14,
116–17, 119–21, 169, 211,297, 323, 336
see also associative learning;discrimination learning; sociallearning
learning curve 64, 64, 65learning sets 361–2, 362leopard 335lever pressing 93, 95–6, 98–102,
105–8, 110and conditioned suppression 37–8and CS–US contingency 71and discrimination learning
151–2, 153and extinction 124, 141and imitation studies 306–7, 307 and interval timing 236–8, 237 ,
239–41, 240–1and number skills 245, 246, 251and preparatory CRs 56and serial order 253
lexigrams 343–5, 343–4, 346light
polarized 285ultraviolet 285
light-dark cycle 287–8, 292artificial 288, 291
limited capacity theory 202, 203–4loggerhead turtle 290–1, 290
long-distance travel 265, 283–93and air pressure 285homing 284, 286–9and magnetic fields 284, 288, 291migration 284, 289–93and navigational cues 284–5, 293–5and polarized light 285and ultraviolet light 285
long-term memory 191, 212–31capacity 214–15, 214, 215comparative studies of 228–31consolidation theory of 218–20,
218, 225
durability 215–17, 216 , 217 episodic 225–31neural circuitry 218, 224–5retrieval theory of 218, 220–5,
222, 223loris 266Loulis (chimpanzee) 341–2, 342, 350
magnetic fields 269–70, 284,288, 291
magnitude effect 256, 259mammalian evolution 6Manx shearwater 286
map and compass hypothesis 287–8maps, cognitive 276–80, 330, 369–70marmoset monkey 83
and imitation 310, 367short-term memory of 366–7, 366
marsh tit 363marsh warbler 332
Matata (bonobo) 344matching to sample 180–3, 181, 184,
186–7, 186 , 189, 215, 368see also delayed matching to sample
matingand deception 314–15mate choice 301
maze running 94, 94and foraging behaviour 300–1, 301and long-term memory 222–4, 223and navigation skills 276–7, 277 ,
281–3, 281–2, 294and trace conditioning 195–6,
195, 196 see also radial maze; T-mazes
mean length of utterances (mlu) 349, 349meaning 337, 346Melton (baboon) 314, 315memory 188–9
active 220, 221episodic 225–31episodic-like 226–8and evolution 370human 371inactive 220and the null hypothesis 366–7, 366 reference 237–8
retrieval 21spatial 363–4, 366 , 366–7 and standard operating procedures
76–9and stimulus significance 81visual 304see also long-term memory;
short-term retentionmemory model of associative learning
46–9, 46–9, 59, 111memory traces 202, 211, 241–2mental states 28, 312, 324–5metacognition
definition 166and discrimination learning 166–9,167–8
metamemorydefinition 208and short-term retention 207–11, 209
metamphetamine 241Mexican jay 230migration 284, 289–93
definition 284, 289endogenous control of 290–2, 290and learning 292–3and the null hypothesis 367
mimicry 303–4, 312, 324vocal 303–4
mind 12mirror neurons 311–12, 324mirrors 319–24, 320, 321, 368–9Missouri cave bat 286mlu see mean length of utterances
molar theory of reinforcement 99–100molecular theory of reinforcement
100, 101–2monkeys 368
and category formation 179, 187and discrimination learning 166–9,
167 , 168episodic-like memory of 227fear of predators 301–2and imitation 311–12, 312, 367, 368and instrumental conditioning 93and metacognition 166–9, 167 , 168mirror-use 322, 323number skills of 246–8, 247 ,
251, 252and self-recognition 322and serial order 253, 256–9,
256–8, 358short-term retention of 198, 201–2,
204, 228and trace conditioning 195and transitive inference 260–1,
260, 262see also specific species
morphine 56–8, 58motivation
dual-system theories of 107–8,
109–10and language 357
motor neurons 44–5, 45, 46mouse 233music 172–3mynah bird 303
natural selection 5navigation 21, 264–95
bicoordinate 288and communication in honey-bees
328–30, 328–30, 331long-distance travel 265, 283–93
methods of 265–83and the null hypothesis 367and the Rescorla-Wagner theory 293short-distance travel 265–83, 293and the sun 267
navigational cues 284–5, 293–5need 30, 106–7negative patterning 153, 156, 162neocortex 364neophobia 298–9, 300neural circuitry
of associative learning 42–5, 44, 45of long-term memory 218, 224–5

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 49/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Subject index 417
neural net theories see connectionistmodels
New Caledonian crow 119–20, 120Nim (Neam Chimpsky) (chimpanzee)
341, 346, 349–50, 357no–US center 134–6, 135no–US representation 55
nominal scales 247nous (mind) 12null hypothesis of intelligence 12,
261, 364–9and associative learning 364–6as impossible to refute 369and language 369and memory 366–7, 366 and navigation 367and the representation of knowledge
368–9and social learning 367–8
number 233, 243–52, 263, 368absolute number 245–7, 245–7 addition 249–50, 250interval scales 250nominal scales 247numerical symbols 248–50, 248–9,
252ordinal scales 247–8and perceptual matching 251–2relative numerosity 243–4, 244representation 247–8subitizing 251
number-identification task 86numerons 252numerosity generalizations 244, 244
oak leaf silhouette experiment 171,172, 173
observational conditioning 302, 308, 309octopus 298odour cues 309olfaction 289omission schedules 60operant conditioning see instrumental
(operant) conditioningorang-utan
and communication 339–40, 340and mimicry 303
and self-recognition 323–4ordinal scales 247–8orienting response (OR) 74–6, 75,
87–8, 87 oscillators 236, 263overshadowing 70, 101–2, 293
Pacific sea anemone 192paramecium 149, 149, 192, 193parrot
and communication 351–3and mimicry 303see also African grey parrot
partial reinforcement 130–4, 131, 132,134, 145–7, 145
partial reinforcement effect (PRE)130, 132, 133, 145–7
path integration see deadreckoning/path integration
Pavlovian (classical) conditioning 20,
28–30, 29, 32–3, 35–61,93, 99
using air pressure 285and autoshaping 37and the conditioned response 55–61and conditioned suppression 38and diet selection 299and discrimination learning
149–50, 162and dual attention 86and evolution 370–1and extinction 123–6, 128–31,
133–4, 138–47eye-blink conditioning 36–7and fear of predators 302and imitation 305using infrasound 285instrumental interactions 106,
109–11and long-term memory studies 217using magnetic fields 284memory model of 46–9, 46 neural mechanisms of 43–5,
44, 45and the null hypothesis 364–6and overshadowing 101and stimulus-stimulus learning
49–52taste aversion conditioning 39and timing 242using ultraviolet and polarized
light 285Pavlovian-instrumental transfer
design 110peak procedure 239, 239peak shift 151, 153, 178Pearce–Hall theory 86–91pedometers 267penguin 24, 24–5pentobarbitol 224
perception, colour 173–4perceptual learning 158–61, 159–61perceptual processing 21periodic timing 233–6, 263
oscillators 236, 263pheromone trails 265–6Phoenix (dolphin) 353phonemes 173, 341phylogenetic scale 5physiological techniques 20–1pigeon 16, 17–18, 55–6, 365–6, 368
and autoshaping 37, 37 , 38, 60and category formation 171–2, 172,
173–5, 174–5, 176, 177,182–5, 182, 183–5, 358
and communication 339and discrimination learning 151,
151, 154episodic-like memory of 227–8and extinction 124, 126–8
and homing 286–9, 293and imitation 310, 311and inhibitory conditioning 40and long-term memory 214–17,
214, 215, 227–8metamemory of 211navigation skills of 264, 272,
284–9, 293number skills of 243–4, 244, 251and problem solving 114and selective association 84and self-recognition 322and serial order 253–6, 254–5, 259short-term retention of 197–8, 200,
201–2, 204, 205, 206, 211, 228and stimulus significance 80–3, 84and stimulus-stimulus
conditioning 50and time 241–2and transitive inference 261–2, 261
pilotingwith multiple landmarks 271–2,
271–2with single landmarks 269–71
pinyon jay 363place cells 280–3plastic tokens 342–3, 343
population-specific behavioraltraditions 305–6
PRE see partial reinforcement effectpredators
and alarm calls 331, 332–5, 334,337
fear of 301–2prefrontal lobe 21Premack principle 103–4primacy effect 206–7primates 6, 8–9
and imitation 305–6and navigation 275
and social learning 367see also specific speciesproactive interference 200–1, 201,
203, 204problem solving 26–7
and causal inference 114–21, 115and folk physics 116–21and insight 112–14and language 19and the Law of Effect 111–21
prospective code 198“protection from extinction” 126–8,
126–7

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 50/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
418 Subject index
prototype theory 178–9protozoan 327punishment 14, 31–2, 93
see also electric shocks;electroconvulsive shock
puzzle boxes 26–7, 26–7 , 28
quail 367
R–US associations 94–5, 95, 97,101–2, 110–11, 311, 365
rabbit 64, 64, 68–9, 69, 77–80, 110and extinction 124–5, 135and habituation 192–4, 193and Pavlovian conditioning 36–7,
39–40, 53–4raccoon 60–1, 191radial maze 228–9, 366
and forgetting 200–1, 201, 202,203, 204–5
and navigation 294and serial position effects 206–7,
208and short-term retention 196–7,
196–7 , 198, 199rat 14, 31–2, 32, 59
and attention and conditioning75–6, 75, 78, 83–8, 86 , 89–90
and compensatory CRs 56–8and conditioned suppression 37–8and CS–US contingency 71and diet selection 298–301,
299, 301and discrimination learning 151–2,
153, 153, 157–8, 163episodic-like memory of 227, 228and extinction 128, 128, 130–4,
139, 140–2, 146and imitation 306–7, 307 , 308–9,
308, 367–8and inhibitory conditioning 42and instrumental conditioning
93–101, 94–101, 103–8, 103,105, 108, 110, 365
and knowledge representation 188and learned irrelevance 85–6, 86 long-term memory of 216–25,
218–19, 222, 227–8and the memory model of conditioning 47–9, 47
and method of learning 14–15navigation skills of 268, 273–9,
273–5, 277–9, 281–3, 281,293, 294
number skills of 245–6, 246 , 251–2and orienting responses 87–8and preparatory CRs 56and selective association 84, 85and serial order 253short-term retention of 194–7,
195–7 , 199–207, 201, 204,208, 228–9
and stimulus significance 83, 84, 85and stimulus-response theorists
31–2, 32and stimulus-stimulus learning
49–50
and time 235–8, 237 , 239–41,240, 241
and trace conditioning 194–6,195–6
raven 117–19, 118–19reactivation effects 221–5, 222, 223reasoning, analogical 187, 188,
368, 371recency effect 206–7reference memory 237–8reinforcement
conditioned 105–6, 105continuous 130–1, 131–2, 134,
134, 145–6, 145molar theory of 99–100molecular theory of 100, 101–2
reinforcer devaluation design 95–6,95, 97
reinforcers 93and associative competition 101–2conditioned 105–6definition 102–4incentive value of the 108, 109nature of the 102–4negative 93positive 93response contiguity 98
token 105–6relational learning 149–50relationships between objects 180–7,
188–9second-order 185–7, 186
renewal effect 136–8, 136 , 137 representation 21–2
A1 76–8, 76 , 81A2 76–80, 76 , 81amodal 240–1, 241and communication 334–5inactive 76, 76 of number 247–8
retrieval-generated A2representations 78–9self-generated A2 representations
77–8, 79spatial 258–9, 262standard operating procedures
model 76, 76 US 109see also knowledge representation
reptiles 6Rescorla–Wagner model 64–5, 68–71,
72–4, 83, 84, 91and category formation 175, 176
and discrimination learning 152–5,153–4, 156, 162, 163, 164–5
and extinction 123, 125–6,127–31, 142
and navigation 293research methods 20–2response-chaining 253–7, 254–7
response-cueing 110–11, 111response-reinforcer contingency
99–100, 99, 100, 101–2retardation test 41–2retention interval 197–8, 216retinal snapshots 270, 271retrieval theory 218, 220–5,
222, 223retroactive interference 163, 200,
201–2, 203retrospective code 198reward 93
anticipation of 94–5, 96, 108and category formation 175–6and gradient of delay 98, 98and the Law of Effect 112and response-reinforcer contingency
99–100and short-term memory
studies 199and stimulus-response theorists
31, 32rhesus monkey
and category formation 181, 184and metamemory 208–11, 210and serial order 257–8, 257–8and short-term retention 206,
208–11Rico (dog) 351rook 117running wheels 103, 103
S–R associations 365and imitation 311and inhibition in extinction 140–2,
141and instrumental conditioning 93–4,
96and serial order 253and theory of mind studies 318
S–R theorists 46, 48–9, 52, 94–5, 96 ,111S–(R–US) associations 96–7, 111sage grouse 301sameness 180–5, 181–5, 188–9Sarah (chimpanzee) 116, 187, 188,
260, 342–3, 346, 347, 349, 359,368, 371
satiety 107, 108–9satisfaction 28, 30scala naturae 4–5scrub jay 318–19, 324sea lion 181, 181, 215, 216

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 51/52
Copyright © 2008 Psychology Press http://www.psypress.com/animal-learning-and-cognition
Subject index 419
Second World War 266second-order (higher-order)
conditioning 50–2, 51,185–7, 186
selective association 84–6self-awareness 323self-recognition 319–24, 320–2,
368–9semanticity 337, 346sensory neurons 44, 45, 46sensory preconditioning 50sentences 342–3, 346–50, 352–4, 355,
356, 358comprehension 347–8, 352,
353–4, 355production 348–50, 352–3,
356, 358sequential stimuli 245–6, 245sequential theory 132, 133serial conditioning 49–50serial delayed alternation 203serial order 233, 253–9, 254–8, 263,
358, 368and chunking 255–6and the distance effect 257, 259and the magnitude effect 256, 259spatial representation of 258–9and transitive inference 259–61
serial position effects 206–7, 208serial recognition tasks 253–7, 254–7 serotonin 45Sheba (chimpanzee) 248, 250,
252, 368Sherman (chimpanzee) 345, 346, 357
short-distance travel 265–83, 293and cognitive maps 276–80and dead reckoning/path integration
266–9, 269, 270, 278, 280,293, 367
and detours 276–8, 277 and distinctively shaped
environments 274–6, 274–5and geometric modules 274–6and geometric relations 272–4,
273–4and hippocampal place cells 280–3methods of navigation 265–83
and novel release sites 278–80, 279and pheromone trails 265–6piloting with multiple landmarks
271–2, 271–2piloting with single landmarks
269–71short-term retention 190–211,
366, 366 comparative studies of 228–31and decay theory 202–3forgetting 199–205and habituation 192–4, 193limited capacity theory of 202
and metamemory 207–11, 209methods of study 191–9and serial position effects 206–7and trace conditioning 194–6
shuttle-boxes 220–1Siamese fighting fish 229sign language 24, 341–2, 349–50, 349
signals 334–5silkworm moth 265simultaneous chaining 253–7, 254–7 simultaneous stimuli 246–7, 247 Slocum (sailor) 266snakes 301–2, 302, 333, 334social behavior 59–60social enhancement 305social groups 367social learning 296–325
and copying behavior 302–12and diet selection 298–301, 299and foraging behavior 300–1, 301and mate choice 301and the null hypothesis 367–8and predator fear 301–2and self-recognition 319–24, 320–2and theory of mind 312–19, 324–5
song birds 331–2, 363SOP model see standard operating
procedures (SOP) modelspatial memory 363–4, 366 , 366–7 spatial representation 258–9, 262spontaneous recovery 134–6squirrel monkey 300
short-term retention of 206and transitive inference 260, 260
standard operating procedures (SOP)model 76–80, 76 , 81, 83, 192
starling 292–3stars 292Stellar’s jay 328, 328, 337stickleback 192, 228, 328, 334stimulus 20
context-stimulus associations78–80, 90–1
and habituation 192–4preexposure 157–61significance 80–6see also conditioned stimulus; S–R
associations; S–(R–US)associationsstimulus enhancement 298, 306, 307,
308, 309stimulus generalization
definition 37and discrimination learning 150,
153, 155–6, 157–8neural basis of 45and problem solving 116, 121
stimulus-response theorists 30–3, 32see also S–R associations
stimulus-stimulus learning 49–52
Stravinsky, Igor 172–3strawberry finch 332subitizing 251Sultan (chimpanzee) 112–13, 112,
114, 115summation test 42, 43sun 267, 287–8
sun compass 287–8surprise
and distractors 203and extinction 125–30and learning 62–74, 84,
86–7, 90symbolic distance effect 260–1
T-mazes 195, 203–4, 204tamarin monkey 229, 300
and self-recognition 322and short-term memory 366–7, 366
taste aversion conditioning 14, 39,47–9, 58–9, 78, 78, 95–6, 300
and discrimination learning 158,159–61
and trace conditioning 194–5, 195temporal generalization 236–8, 237 ,
240–1theory of mind 312–19, 324–5, 371
and communication 336and deception 313–15, 314and knowledge attribution 315–19
three-spined stickleback 192tiger 297 time 233–42, 262–3, 368
circadian rhythms 233–4
connectionist model of 241information-processing model of
237–8, 237 , 239, 239, 240,241
interval timing 233, 236–41,237–41, 242, 263
and memory traces 241–2periodic timing 233–6, 263scalar timing theory 238, 238,
239–40, 240tool use 116–17, 117 , 119–20trace conditioning 194–6, 195–6 trace intervals 194
transitive inference 259–62, 260–1and spatial representation 262and the symbolic distance effect
260–1and value transfer 261–2
transposition 149–51transposition test 150trial and error learning 25–7, 112–14,
116–17, 119–21, 169, 211, 297,323, 336
trial-based theories 142truly random control 40two-action control 309–10, 309

8/3/2019 Amimal Learning
http://slidepdf.com/reader/full/amimal-learning 52/52
420 Subject index
ultraviolet (UV) light 285unconditioned stimulus (US) 29, 30,
32–3, 35, 40, 123and attention 78–9, 80aversive 110and compensatory CRs 58and diet selection 299
and discrimination learning 155,156, 162, 165
and extinction 125–6, 128–9, 135,139–43, 145–6
and eye-blink conditioning 36and fear of predators 302and imitation 305and inhibitory conditioning 40,
41, 42intensity 66–7and long-term memory studies 217memory model of 46–8, 59and the nature of representations
52–5, 54and the reflexive nature of the
CR 60
representation 109and stimulus-stimulus conditioning
49–50surprising 63–4, 69, 365–6and trace conditioning 194see also CS–US
association/contingency;
no–US center; no–USrepresentation; R–USassociations; S–(R–US)associations
unobservable processes 21–2urine 266utterances, mean length of
(mlu) 349, 349
value transfer 261–2vasoconstriction 77–8,
192–3, 193vasopressin 241vervet monkey 332–7, 333–4vestibular system 268Vicki (chimpanzee) 307, 340
visual category formation 171–2,173–6
visual memory 304vocal mimicry 303–4vocal tract 340, 340
waggle dance (bees) 328–30, 328–30,
331, 338–9Washoe (chimpanzee) 24, 341–2,
342, 344, 346, 348–9,350, 357
Western scrub jay 226–7, 226 white-crowned sparrow 331–2, 370wind direction 289wolf 58–9, 59Woodpecker finch 119, 120words 337working memory 237–8wrasse 228
Yerkish 343
Zugunruhe 291, 292





























