Embed Size (px)
Citation preview

An analysis of cyanobacterial bloom occurrence in Missisquoi Bay (Québec, Canada) between 2000 and 2008, and possible environmental factors underlying them

Published by the NSW Department of Primary Industries, Office of Water
An analysis of cyanobacterial bloom occurrence in Missisquoi Bay (Québec, Canada) between 2000 and 2008, and
possible environmental factors underlying them
First published July 2014
ISBN 978 1 74256 652 8
More information
Lee Bowling, Office of Water, NSW Department of Primary Industries, Menangle, New South Wales, Australia
Sylvie Blais, Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Québec City,
Québec, Canada
Marc Sinotte, Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Québec City,
Québec, Canada
www.dpi.nsw.gov.au
© State of New South Wales through the Department of Trade and Investment, Regional Infrastructure and Services, 2014. You may copy, distribute
and otherwise freely deal with this publication for any purpose, provided that you attribute the NSW Department of Primary Industries as the owner.
Disclaimer: The information contained in this publication is based on knowledge and understanding at the time of writing (July 2014). However, because
of advances in knowledge, users are reminded of the need to ensure that information upon which they rely is up to date and to check currency of the
information with the appropriate officer of the Department of Primary Industries or the user’s independent adviser.
JT 12990
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
Acknowledgments
This study was supported by a travel grant to Lee Bowling from the Australian Academy of Science under their
Scientific Visits to North America program. We thank the Ministère du Développement durable, de l’Environnement,
de la Faune et des Parcs (MDDEFP) of Québec, Canada, for hosting this visit. Office of Water, New South Wales
Department of Primary Industries, Australia, also supported this visit and the completion of the study.
The authors want to thank all the members of the MDDEFP production team (listed below) for contributing their
expertise to one or more of the following stages of this study:
Scientific editing Sylvie Blais1, Marc Sinotte
1, Martin Mimeault
2
Field sampling, measurement and
comments
Daniel Blanchard3, Patrick Chevrette
3, Mathieu Choinière
3, Guy
Desgranges3
, Martin Mimeault2, Sylvain Primeau
2, Richard Smith
3,
Alain Tanguay3, Sylvie Legendre
1,6, Yves Laporte
1,6
Cyanotoxin laboratory analysis Christian Deblois4 (senior analyst), Annie Laverdière
4, Ginette
Côté4, Mireille Brunet
4, Valérie Gauthier
4, Martin Duchesneau
4,
François Houde4, Nathalie Dassylva
4, Danielle Thomassin
4, Marie-
Claire Grenon4 and Benoît Sarrasin
4 as well as some part-time
employees4
Cyanobacteria and eukaryotic algae
(except in 2000), chlorophyll and
phycocyanin laboratory analysis
Christian Bastien4
(senior analyst), Éloïse Veilleux4, Richard
Cardin4, Karine Ménard-Cloutier
4, Nancy Desrosiers
4, Cathy
Tremblay4, Nicole Cadoret
4
Cyanobacteria and eukaryotic algae
laboratory analysis (2000)
Serge Paquet5
Laboratory inorganic chemistry
parameter analyses
Nathalie Dassylva4
, François Houde4, Danielle Thomassin
4,
Andrée Gendron4
(senior analysts), Gilles Labbé4, Jean-Guy
Samson4, Hélène Ferland
4, Alberte Lauzier
4, Louise Simoneau
4,
Diane Veillette4, Serge Morissette
4 and Steeve Roberge
4
Data capture and verification Nathalie Bourbonnais1, Yves Laporte
1, Sylvie Legendre
1,
Guillaume Desrosiers1, Marie-Ève Thériault
1
Maps Mona Frenette1
Computer graphics France Gauthier1
1. Direction du suivi de l’état de l’environnement (DSEE), Ministère du Développement durable, de l’Environnement,
de la Faune et des Parcs (MDDEFP)
2. Direction régionale de l’analyse et de l’expertise de l’Estrie et de la Montérégie, (DRAE), MDDEFP
3. Centre de contrôle environnemental du Québec de l'Estrie et de la Montérégie (CCEQ), MDDEFP
4. Centre d’expertise en analyse environnementale du Québec (CEAEQ) au MDDEFP.
5. Université du Québec à Montréal
6. DSEE participated in fieldwork only in the first annual study.
Special thanks go to Martin Mimeault, coordinator of Lake Champlain integrated management at the MDDEFP’s
Direction régionale de l’analyse et de l’expertise de l’Estrie et de la Montérégie (DRAE). In addition to providing the
data, he also was the advocate at DRAE for ensuring that it contributed regional technical resources for fieldwork and
laboratory time for sample analysis. He contributed also by reviewing the draft report and advising on nutrients
sources within Missisquoi Bay.
We also thank Mme France Gauthier of MDDEFP for the production of figures within the report.
We thank David Ryan of the NSW Office of Water for assistance with the statistical analyses.
Cover photo: Cyanobacteria bloom in Missisquoi Bay, north of the Saint-Armand public dock, August 26, 2008.
(Source: Patrick Chevrette, Centre de contrôle environnemental du Québec de l'Estrie et de la Montérégie, MDDEFP).

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
i NSW Office of Water, July 2014
Contents Executive summary ..................................................................................................................... v
1 Introduction .............................................................................................................................. 1
2 Methods ................................................................................................................................... 3
2.1 Field samping and data collection ...................................................................................... 3
2.1.1 Sampling sites and strategies ...................................................................................... 3
2.1.2 Data collection............................................................................................................. 4
2.2 Laboratory analysis............................................................................................................ 5
2.3 Classification of potentially toxic cyanobacteria ................................................................. 6
2.4 Statistical analysis ............................................................................................................. 6
3 Results ..................................................................................................................................... 8
3.1 Annual variation in cyanobacterial presence at sites a, b, d2 and d8 ................................. 8
3.2 Contribution of Cyanobacteria to the Total Phytoplankton Biomass ................................. 12
3.3 Interactions between phytoplankton and cyanobacterial taxa .......................................... 16
3.4 Cyanobacteria and toxin presence ................................................................................... 18
3.5 Eukaryotic and cyanobacterial presence and water quality indicators .............................. 25
3.5.1 Analysis of the combined data for sites a, b, d2 and d8 ............................................. 25
3.5.2 Site by site analysis ................................................................................................... 27
3.6 Meteorological factors and cyanobacterial and eukaryotic algal presence ....................... 28
3.7 Multivariate analysis of the cyanobacterial and environmental data ................................. 32
3.7.1 Preliminary analysis of the complete cyanobacterial data set .................................... 32
3.7.2 Analysis of the reduced cyanobacterial data set ........................................................ 33
3.7.3 Analysis of the environmental data set ...................................................................... 34
3.7.4 Comparison of the reduced cyanobacterial data set and the environmental data set 35
3.7.5 Comparisons on a site by site basis .......................................................................... 36
4 Discussion.............................................................................................................................. 36
5 Conclusions ........................................................................................................................... 43
6 References ............................................................................................................................. 45
Appendix A List of sampling sites in the Québec portion of Missisquoi Bay, their location, the
number of samples collected at each (between 2000 and 20081), and the period of record ...... 49
Appendix B Analytical methodology used by CEAEQ ................................................................ 51

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
ii NSW Office of Water, July 2014
Appendix C Cyanobacterial taxa that were present at sites a, b, d2 and d8 for the entire
sampling campaigns between 2000 and 2008, and their dominance ......................................... 52
Appendix D Spearman Rank Order Correlation coefficients (Rho) for interactions between
various cyanobacterial and eukaryotic algal groupings and taxa ............................................... 58
Appendix E Scatter diagrams of total microcystin concentrations against various water quality
attributes (sites a, b, d2 and d8 combined), and Spearman Rank Order Correlation coefficients
(Rho) ......................................................................................................................................... 62
Appendix F Spearman Rank Order Correlation coefficients (Rho) for interactions between
various cyanobacterial and eukaryotic algal groupings and taxa against physico-chemical
variables ................................................................................................................................... 66
Appendix G Spearman Rank Order Correlation coefficients (Rho) and probabilities (p) for
interactions between the biomass of various cyanobacterial and eukaryotic algal taxa (log(x)
transformed) and meteorological variables measured at Philipsburg manual weather station ... 75
Appendix H Output from the multivariate analyses .................................................................... 99
Figure 1 Location of sampling sites and main public usage areas in Missisquoi Bay. “Patrolled
public beach” indicates that at least one lifeguard is on duty. For this report, only data from sites
a, b, d2 and d8 were used ........................................................................................................... 4
Figure 2 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site a in the
Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 3, 4 and 5 .... 9
Figure 3 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site b in the
Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 2, 4 and 5 . 10
Figure 4 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site d2 in
the Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 2, 3 and 5
.................................................................................................................................................. 11
Figure 5 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site d8 in
the Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 2, 3 and 4
.................................................................................................................................................. 13
Figure 6 Contribution provided by various phytoplankton divisions (both cyanobacteria and
eukaryotes) to total phytoplankton biomass at site a, from 2000 to 2008 inclusive. Note the
difference in Y axis scale from Figures 7, 8 and 9 ..................................................................... 14
Figure 7 Contribution provided by various phytoplankton divisions (both cyanobacteria and
eukaryotes) to total phytoplankton biomass at site b, from 2000 to 2008 inclusive. Note the
difference in Y axis scale from Figures 6, 8 and 9 ..................................................................... 15
Figure 8 Contribution provided by various phytoplankton divisions (both cyanobacteria and
eukaryotes) to total phytoplankton biomass at site d2, from 2000 to 2008 inclusive. Note the
difference in Y axis scale from Figures 6, 7 and 9 ..................................................................... 15
Figure 9 Contribution provided by various phytoplankton divisions (both cyanobacteria and
eukaryotes) to total phytoplankton biomass at site d8, from 2000 to 2008 inclusive. Note the
difference in Y axis scale from Figures 6, 7 and 8. .................................................................... 16
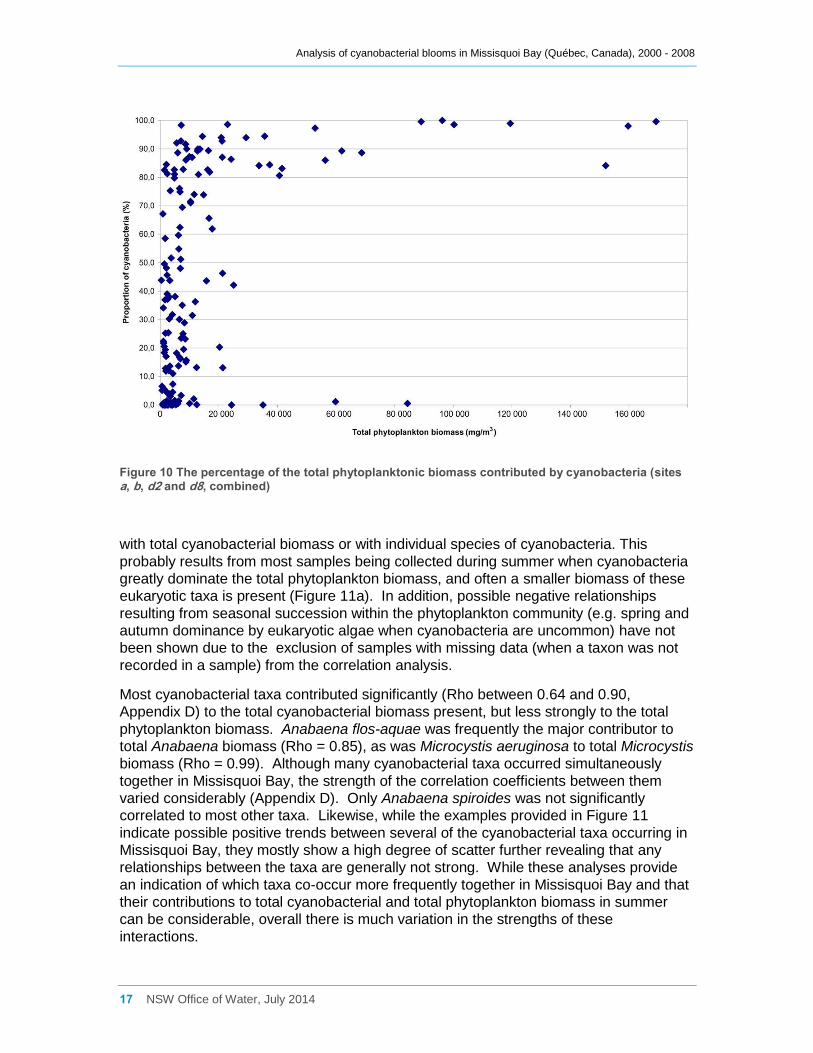
Figure 10 The percentage of the total phytoplanktonic biomass contributed by cyanobacteria
(sites a, b, d2 and d8, combined) .............................................................................................. 17

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
iii NSW Office of Water, July 2014
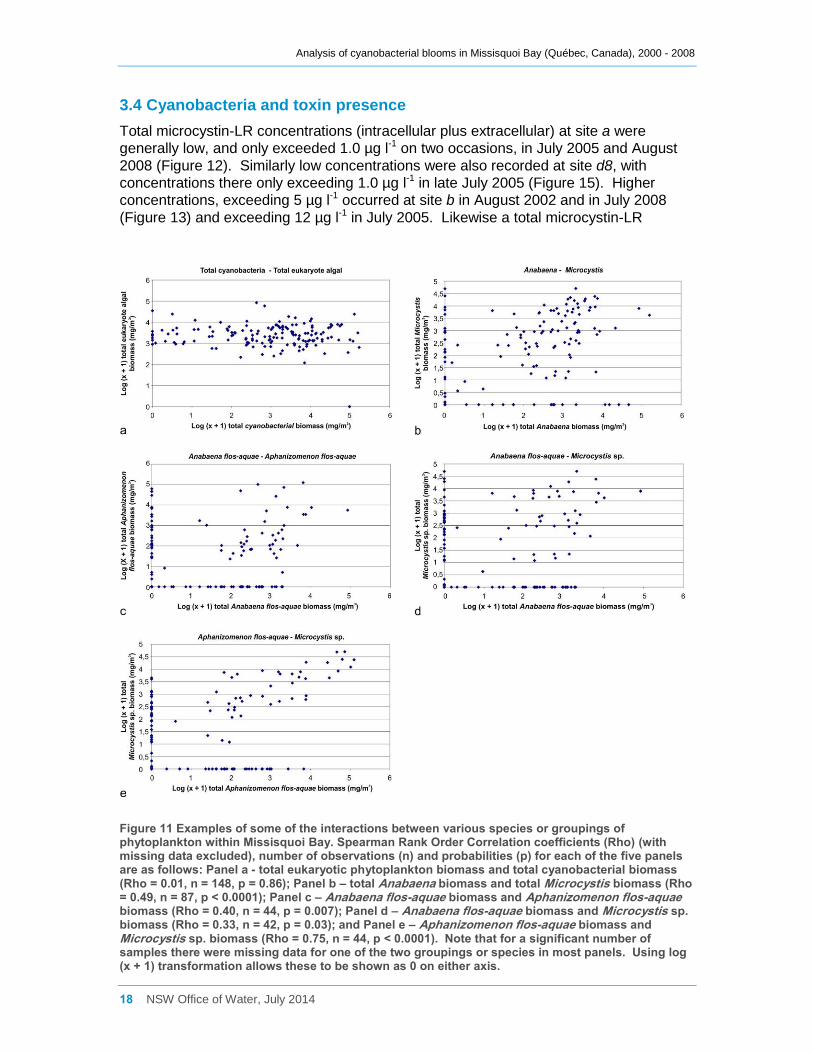
Figure 11 Examples of some of the interactions between various species or groupings of
phytoplankton within Missisquoi Bay. Spearman Rank Order Correlation coefficients (Rho) (with
missing data excluded), number of observations (n) and probabilities (p) for each of the five
panels are as follows: Panel a - total eukaryotic phytoplankton biomass and total cyanobacterial
biomass (Rho = 0.01, n = 148, p = 0.86); Panel b – total Anabaena biomass and total
Microcystis biomass (Rho = 0.49, n = 87, p < 0.0001); Panel c – Anabaena flos-aquae biomass
and Aphanizomenon flos-aquae biomass (Rho = 0.40, n = 44, p = 0.007); Panel d – Anabaena
flos-aquae biomass and Microcystis sp. biomass (Rho = 0.33, n = 42, p = 0.03); and Panel e –
Aphanizomenon flos-aquae biomass and Microcystis sp. biomass (Rho = 0.75, n = 44, p <
0.0001). Note that for a significant number of samples there were missing data for one of the
two groupings or species in most panels. Using log (x + 1) transformation allows these to be
shown as 0 on either axis. ......................................................................................................... 18
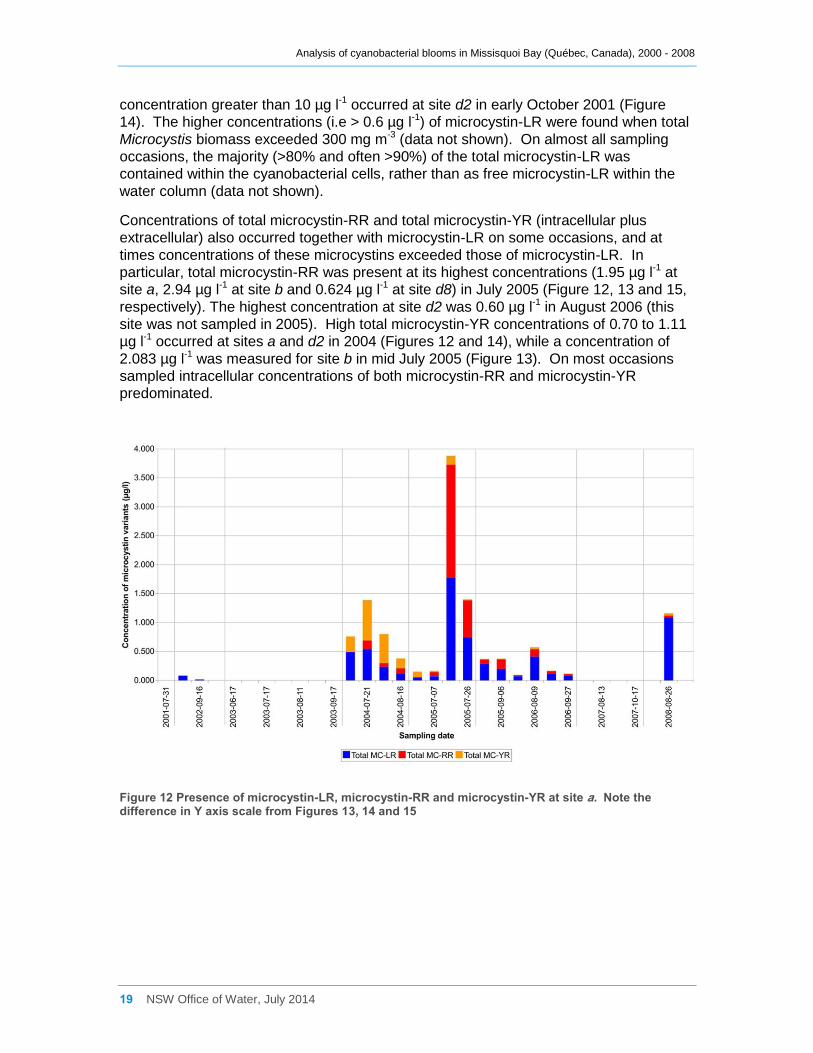
Figure 12 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site a. Note the
difference in Y axis scale from Figures 13, 14 and 15 ............................................................... 19
Figure 13 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site b. Note the
difference in Y axis scale from Figures 12, 14 and 15 ............................................................... 20
Figure 14 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site d2. Note the
difference in Y axis scale from Figures 12, 13 and 15 ............................................................... 20
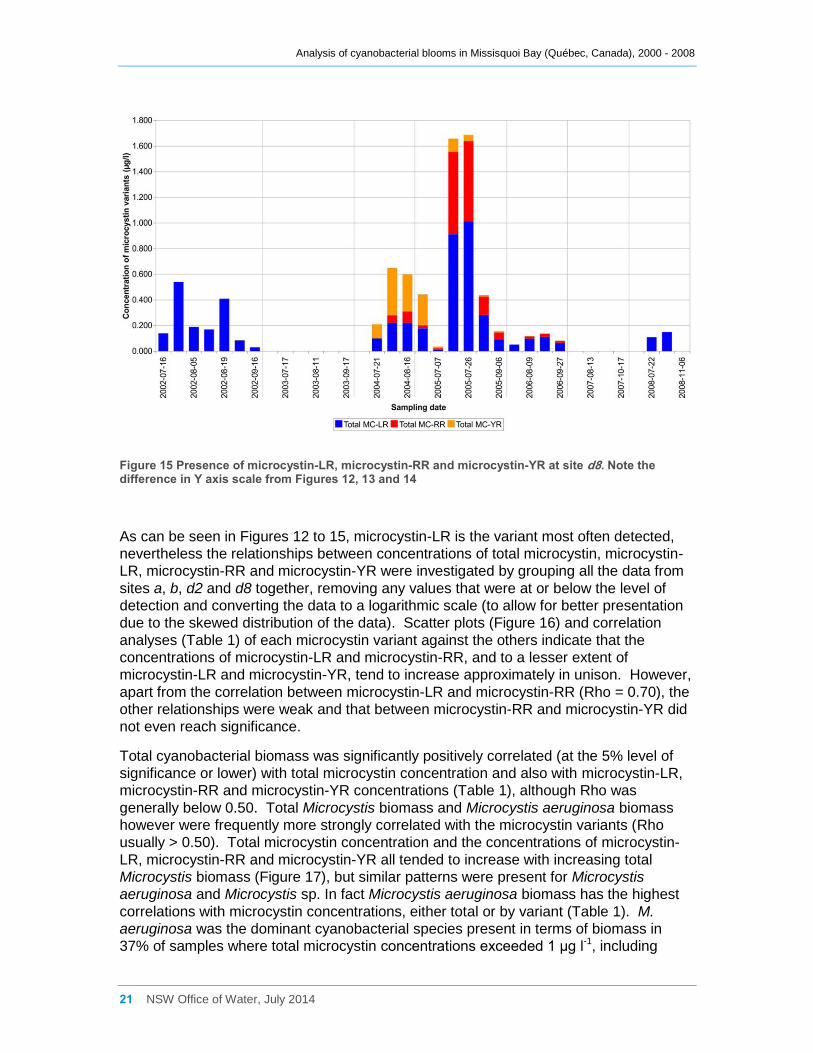
Figure 15 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site d8. Note the
difference in Y axis scale from Figures 12, 13 and 14 ............................................................... 21
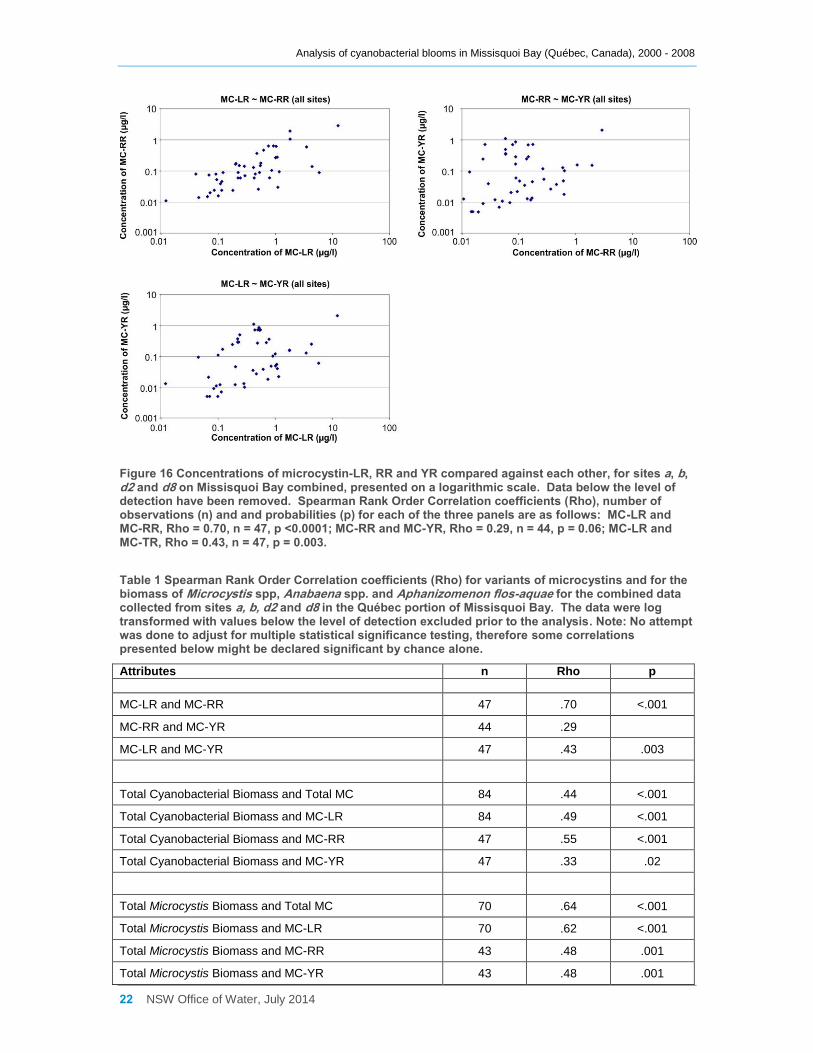
Figure 16 Concentrations of microcystin-LR, RR and YR compared against each other, for sites
a, b, d2 and d8 on Missisquoi Bay combined, presented on a logarithmic scale. Data below the
level of detection have been removed. Spearman Rank Order Correlation coefficients (Rho),
number of observations (n) and and probabilities (p) for each of the three panels are as follows:
MC-LR and MC-RR, Rho = 0.70, n = 47, p <0.0001; MC-RR and MC-YR, Rho = 0.29, n = 44, p
= 0.06; MC-LR and MC-TR, Rho = 0.43, n = 47, p = 0.003. ...................................................... 22
Figure 17 Concentrations of microcystin-LR, RR and YR and the total of all three variants
compared against total Microcystis biomass for sites a, b, d2 and d8 in Missisquoi Bay
combined, presented on a log scale. Data below the level of detection have been removed.
Spearman Rank Order Correlation coefficients (Rho), number of observations (n) and and
probabilities (p) for each of the four panels are as follows: Total Microcystis biomass and total
MC, Rho = 0.64, n = 70, p <0.0001; Total Microcystis biomass and MC-LR, Rho = 0.62, n = 70, p
<0.0001; Total Microcystis biomass and MC-RR, Rho = 0.48, n = 43, p = 0.001; Total
Microcystis biomass and MC-YR, Rho = 0.48, n = 43, p = 0.001. .............................................. 24
Figure 18 Variations of concentrations of total microcystin, MC-LR, MC-RR and MC-YR at sites
a, b, d2 and d8 in Missisquoi Bay combined in relation to water temperature. Microcystin data
are presented on a log scale, and data below the level of detection have been removed.
Spearman Rank Order Correlation coefficients (Rho), number of observations (n) and and
probabilities (p) for each of the four panels are as follows: Total microcystin and surface water
temperature, Rho = 0.20, n = 82, p = 0.08; MC-LR and surface water temperature, Rho = 0.08, n
= 82, p = 0.47; MC-RR and surface water temperature, Rho = 0.43, n = 45, p = 0.003; MC-YR
and surface water temperature, Rho = 0.47, n = 45, p = 0.001. ................................................. 26
Figure 19. Examples of scatterplots where the strongest correlations between cyanobacterial
biomass at sites a, b, d2 and d8 combined and meteorological conditions were usually obtained,
except wind speed (panel f). Note that in most panels there are a number of samples where
there is missing cyanobacterial biomass data. Using log (x + 1) transformation allows these to
be shown as 0 on the Y axis. Spearman Rank Order Correlation coefficients (Rho), number of

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
iv NSW Office of Water, July 2014
observations (n) and probabilities (p) for each of the six panels are as follows: Total Anabaena
biomass and antecedent 10 day maximum temperature, Rho = 0.22, n = 94, p = 0.03; A
spiroides biomass and antecedent 10 day minimum temperature, Rho = 0.36, n = 38, p = 0.02;
Total cyanobacterial biomass and antecedent 10 day rainfall, Rho = -0.26, n = 132, p = 0.003;
Total cyanobacterial biomass and antecedent 10 day cloud cover, Rho = -0.25, n = 132, p =
0.004; M. aeruginosa biomass and antecedent 10 day wind direction, Rho = -0.62, n = 22, p =
0.002; M. aeruginosa biomass and antecedent 10 day wind velocity, Rho = -0.14, n = 22, p =
0.52. .......................................................................................................................................... 30
Figure 20 Total cyanobacterial presence for all four Missiquoi Bay sites combined (blue
diamonds) compared against various meteorological variables from Philipsburg manual weather
station (histogram bars) ............................................................................................................ 31
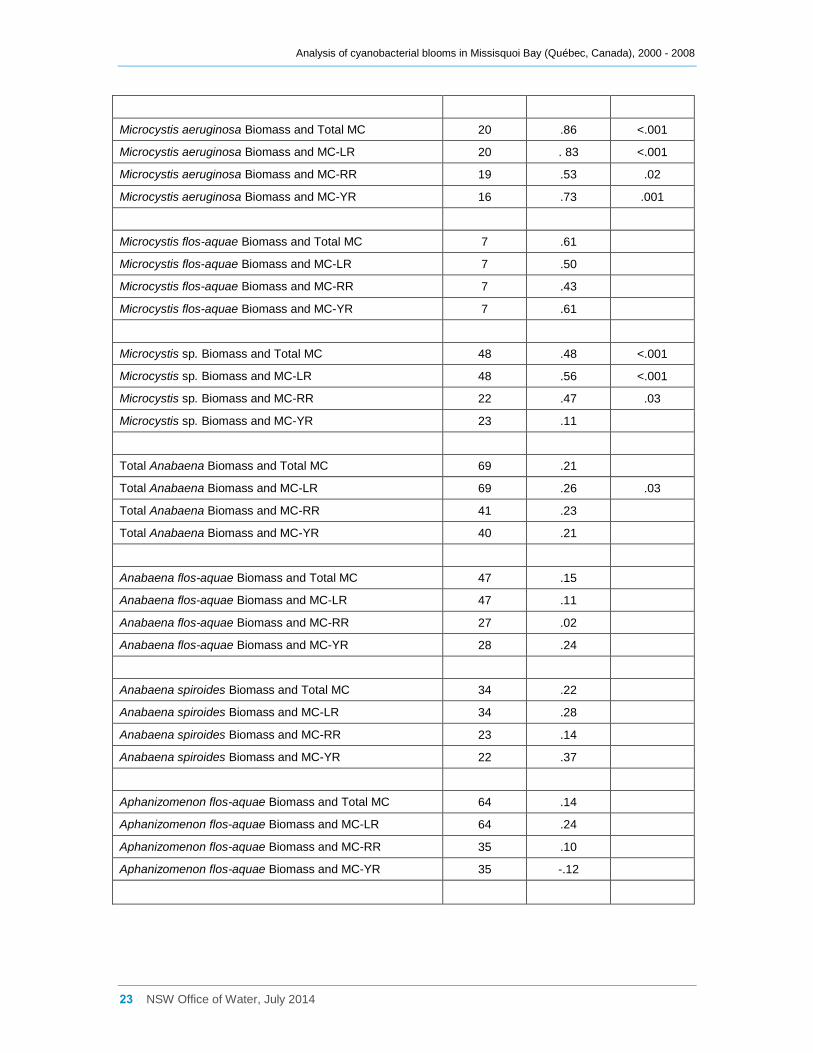
Table 1 Spearman Rank Order Correlation coefficients (Rho) for variants of microcystins and for
the biomass of Microcystis spp, Anabaena spp. and Aphanizomenon flos-aquae for the
combined data collected from sites a, b, d2 and d8 in the Québec portion of Missisquoi Bay.
The data were log transformed with values below the level of detection excluded prior to the
analysis. Note: No attempt was done to adjust for multiple statistical significance testing,
therefore some correlations presented below might be declared significant by chance alone. .. 22
Table 2 90th percentile, median (in bold) and 10th percentile values for various water quality
variables at different sites on Missisquoi Bay. Number of observations (n) in brackets ............ 28

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
v NSW Office of Water, July 2014
Executive summary
Cyanobacterial (blue-green algal) and water quality data has been collected for the
northern (Québec, Canada) portion of Missisquoi Bay by the Ministère du
Développement durable, de l’Environnement, de la Faune et des Parcs (MDDEFP). The
sampling protocol was designed primarily to check for compliance against local
guidelines, mainly for recreational water use on the sampling dates when major blooms
or scums occurred. Samples were also collected to document biological, physical and
chemical conditions within the bay. This report focuses on the biological data and how
environmental factors may be driving cyanobacterial blooms within the bay. Compliance
with management guidelines are reported elsewhere (Blais, 2014).
Data were collected at four sites, including two along the eastern shoreline near Saint-
Armand (sites b, d2), one near the centre of the bay at the point of maximum depth (site
a) and one in the western part of the bay, near Venise-en-Québec (site d8). Sampling
occurred each summer period from 2000 to 2008, however not all sites were sampled
every year or at the same frequency each year, with greatest data collection occurring in
2001 and 2002. There were also slight changes in sampling method over the duration of
the monitoring.
Despite these temporal and spatial variations in sampling effort, the results highlight a
very dynamic and variable cyanobacterial bloom occurrence within Missisquoi Bay.
Generally sites a and d8 had lower cyanobacterial abundance than the two eastern
sites, but cyanobacterial blooms were particularly abundant at most sites from 2001 to
2006 and in 2008. In contrast, 2007 was a low bloom year. Microcystis was the
dominant genus present at most sites between 2001 and 2005. Blooms in subsequent
years have been more mixed, with dominance by potentially toxic species such as
Aphanizomenon flos-aquae, Anabaena flos-aquae and Anabaena spiroides at certain
times. Non-toxic species including Aphanothece clathrata and Chroococcus dispersus
were also sometimes dominant.
Cyanobacterial blooms in Missisquoi Bay are generally preceded by spring blooms of
diatoms and cryptophytes, but exponential growth of cyanobacteria in summer means
that these often come to contribute greater than 80% of the phytoplanktonic biomass at
that time of year. The cyanobacterial blooms decline in autumn, and are replaced again
by diatoms and cryptophytes, although generally at a lower biomass. In 2007 when
cyanobacterial blooms did not develop, green algae (Chlorophyceae) and dinoflagellates
(Peridinium) were dominant components of the phytoplankton.
The cyanotoxin microcystin was recorded at most sites in most years, apart from in
2007. Microcystin-LR was the most common variant present, but microcystin-YR and
microcystin-RR were also frequently present. The microcystin concentrations were
significantly positively correlated with Microcystis biomass but were rarely present when
other cyanobacterial genera were dominant. Microcystin-YR and microcystin-RR were
significantly positively correlated with surface water temperature, although microcystin-
LR and total microcystin were not; even if this relationship is not very strong it might
suggest that some taxa or genetic strains producing these microcystin variants are more
frequently present in warmer waters. Other factors that may have affected microcystin
concentrations include nutrients, pH and turbidity. Other cyanotoxins were usually below
detectable concentrations.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
vi NSW Office of Water, July 2014
Multivariate analysis revealed that water quality varies considerably both spatially and
temporally. In particular the eastern sites tended to have higher total phosphorus
concentrations, turbidity and pH, and lower Secchi disc depths. The spatial variability in
water quality across the bay may be one factor leading to the variable composition of the
cyanobacterial communities. The bay is eutrophic, and a significant positive correlation
was found between cyanobacterial biomass and total phosphorus concentrations. In
comparison, total nitrogen concentrations were low and were only weakly correlated with
total cyanobacterial biomass. However oxidised nitrogen (NOx) was often significantly
negatively correlated with the biomass of various cyanobacterial taxa, indicating possible
uptake of this nutrient by the cyanobacteria. Cyanobacteria also frequently occurred at
TN:TP ratios of less than 7, indicating possible nitrogen limitation regardless of whether
heterocystous species were dominant or not. Significant positive correlations were also
shown between cyanobacterial biomass and pH and turbidity, and a significant negative
correlation occurred with Secchi disc depth. A number of taxa, especially of Anabaena,
were significantly positively correlated with dissolved organic carbon and several species
with water temperature.
Few significant correlations occurred between cyanobacterial biomass and the
meteorological conditions measured at the Philipsburg weather station, apart from the 7
and 10 day mean antecedent maximum air temperature, indicating that a week or longer
of hotter weather may benefit cyanobacteria in Missisquoi Bay in summer. Wind
conditions, periods of rainfall and prolonged cloud cover showed little effect.
Multivariate analysis also demonstrated the considerable temporal and spatial
heterogeneity in cyanobacterial bloom distribution and community composition in
Missisquoi Bay. Multivariate comparison of the cyanobacterial and environmental data
suggested that total nitrogen concentrations, pH and surface water temperature were the
environmental attributes most likely affecting cyanobacterial composition, but these
relationships were weak and there was a lot of unexplained variance in these data.
Correlation analysis indicated that chlorophyll-a and phycocyanin may both be good
indicators of cyanobacterial presence in Missisquoi Bay. There were few significant
correlations between eukaryotic algal biomass and the environmental attributes apart
from pH and surface water temperature.
The large amounts of unexplained variance suggest other unmeasured factors may be
causing major perturbations in the cyanobacterial communities of Missisquoi Bay.
Factors such as herbicide concentrations, the presence of zebra mussels and also of a
zooplanktivorous fish, the alewife, have been suggested by other authors. Unfortunately
there are no data to investigate these potential drivers of cyanobacterial community
composition and abundance in Missisquoi Bay.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
1 NSW Office of Water, July 2014
1 Introduction
Missisquoi Bay comprises the north-eastern end of Lake Champlain. It is a shallow
(mean depth 2.8 metres) embayment, with a surface area of 77.5 km2 (McQuaid et al.,
2011; Wheeler et al., 2011). The international boundary between Canada and the
United States of America divides the bay in two, with the northern half being part of the
Canadian Province of Québec, and the southern half part of the State of Vermont in the
USA.
Problems of eutrophication were first documented for the Québec portion of Missisquoi
Bay during the late 1960s (Fontaine et al., 1968). Cyanobacterial blooms were first
recorded in the bay in the early 1980s (Bérubé, 1981; Germain et al., 1987). However
cyanobacterial blooms have become a prominent feature of Missisquoi Bay since 2000
(Watzin et al., 2006; Boyer, 2008; Zananski et al., 2010; Wheeler et al., 2012). Although
no single factor has been identified as the primary cause of cyanobacterial blooms in
Missisquoi Bay (Zananski et al., 2010), the bay is eutrophic, suffering excess
phosphorus inputs from agricultural and urban runoff (Wheeler et al., 2012). Medalie et
al. (2012) have shown that the Pike and Missisquoi Rivers which both discharge into
Missisquoi Bay provide high annual phosphorus and nitrogen loads, although annual
variation is considerable. Total phosphorus concentrations increased by 72% between
1979 and 2009 (Smeltzer et al., 2012), with concentrations averaging 45 μg l-1 between
1999 and 2003 (Zananski et al., 2010). McCarthy et al. (2013) have also found that
water column nitrogen regeneration processes play a primary role in supporting
productivity in Missisquoi Bay during the growing season, although this and external
nitrogen inputs from tributaries combined were still insufficient to fully support nitrogen
demand in the bay.
The Province of Québec has undertaken regular monitoring of cyanobacterial presence
in its half of Missisquoi Bay each summer since 2000. Between 2000 to 2008, public
health advisories due to cyanobacteria were issued for the bay each summer except for
2000 (Brault, 2012), with microcystin concentrations in scum regularly exceeding
Québec’s guidelines, therefore triggering partial or total closures of the bay for
recreational uses (including beach closures). Occasionally no-drinking advisories were
also issued for the drinking water treatment plant in Saint-Armand.
A number of previous studies have reported on cyanobacteria in Missisquoi Bay.
Smeltzer et al. (2012) compared phytoplankton data collected for mainly Vermont
monitoring programs on Missisquoi Bay for 1970-74, 1991-92, and 2006-09 and found a
shift in dominance from diatoms in the early 1970s and 1990s to cyanobacteria by 2006-
09. Palaeolimnological studies (Levine et al., 2012) using cores from the Québec part of
Missisquoi Bay have indicated that eutrophication of the bay began during the 1970s
when agricultural activities, including dairying and corn production intensified in the
catchment, with cyanobacterial blooms commencing as a result in the 1990s. Using
data from a New York monitoring program, Mihac et al. (2005) reported a spring
dominance of diatoms in 2002 in Lake Champlain, with a late summer shift to Microcystis
being most notable in the shallower northern parts of the lake, including Missisquoi Bay.
These authors also considered that there had been a recent change in cyanobacterial
dominance from Anabaena to Microcystis in northern parts of the lake. Watzin et al.
(2006), while reporting on cyanobacteria in Lake Champlain as a whole between 2002
and 2004 inclusive, reported that the southern (Vermont) portion of Missisquoi Bay had
the most consistent and severe cyanobacterial problems compared with other American

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
2 NSW Office of Water, July 2014
parts of the lake, exceeding World Health Organisation Alert Level 1 guidelines (Bartram
et al., 1999) each year. These blooms were dominated by large Microcystis colonies,
with the hepatotoxin microcystin also being detected. Mihac et al. (2006) reported that
the Missisquoi Bay phytoplankton community during the 2004 summer was comprised
primarily of cyanobacteria, with Microcystis as the dominant taxon but with a large
proportion of Aphanizomenon also present. Mihac et al. (2008) also mapped
cyanobacterial distributions throughout the whole of Missisquoi Bay during three cruises
conducted during the summer of 2006, which illustrated a complex spatial distribution of
blooms within the bay. They found extensive spatial variability in the size, extent and
location of cyanobacterial blooms around the bay, and that different taxa were dominant
at different locations. Finally Wheeler et al. (2012) have used satellite data to map the
distribution of cyanobacteria across the entire Missisquoi Bay in 2004, finding marked
spatial and temporal differences in its distribution.
Fortin et al. (2010) used a quantitative Polymerase Chain Reaction (qPCR) method to
track cyanobacterial blooms at Saint-Armand (Philipsburg sector) in the north-east of the
bay in 2006 and 2007. A bloom throughout summer and early autumn in 2006 was
numerically dominated by Microcystis aeruginosa, with Anabaena species occurring later
in autumn. Cyanobacterial presence at the site was much reduced in 2007, with some
Anabaena spiroides appearing in September. They concluded that Microcystis was the
principal microcystin producing genus in Missisquoi Bay during this study. Microcystis
was also the dominant cyanobacterium at a number of locations in the northern portion
of the bay in 2009, with toxigenic strains comprising 2 to 23% of the total Microcystis
population (Ngwa et al., 2014).
McQuaid et al. (2011) also reported on cyanobacterial presence in the north-east of
Missisquoi Bay close to the inlet of a water treatment plant in 2007 and 2008, reporting
low total cyanobacterial biovolume present in 2007, although high cell counts of the tiny
picoplanktonic species Aphanothece minutissima were encountered. In 2008 however
cyanobacteria once again dominated the phytoplankton community at this site, with both
Microcystis sp. and Anabaena sp. dominating. Zamyadi et al. (2012) further studied the
amount of cyanobacteria entering this drinking water treatment plant in untreated water
drawn from Missisquoi Bay, and reported that Microcystis aeruginosa dominated in the
inflow water in 2008 and 2009. In 2010 they found lower cyanobacterial concentrations
but high concentrations of cyanotoxins, especially microcystin-LR but also some
cylindrospermopsin.
Davis et al. (2009) included sites in Missisquoi Bay (in the US but also sometimes in
Québec) in their study of toxic and non-toxic strains of Microcystis undertaken in 2006.
The bay was devoid of cyanobacteria until July and then never numerically dominated by
Microcystis, but by Anabaena instead. Nutrients at the time were elevated. Toxic
Microcystis cells, determined by PCR, comprised approximately 6% of the total
Microcystis cells present. Pearce et al. (2013) utilised a self-organising map (SOM), a
non-linear and non-parametric clustering method, to investigate associations between
cyanobacterial blooms in Missisquoi Bay and in-lake environmental conditions from 2007
to 2009. They found the method separated the data into bloom and non-bloom periods
based on similarities in water and sediment chemistry and non-cyanobacterial
phytoplankton abundance. They also found that the dissolved nitrogen to soluble
reactive phosphorus ratios had a strong influence on determining Microcystis presence,
which was generally favoured by low ratios. However other undetermined environmental
factors may trigger blooms. Zananski et al. (2010) studied the effects of the herbicide
atrazine on cyanobacteria in water samples collected from the south-west (Vermont)

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
3 NSW Office of Water, July 2014
corner of the bay during the 2005 summer. Eukaryotic phytoplankton, in particular
Heterokontophyceae and Dinophyceae, were consistently major contributors to
chlorophyll-a, with cyanobacteria only dominating in June. These authors found some
seasonal variation in the responses of different divisions of phytoplankton to atrazine,
but were unable to conclude if any were more resistant to the herbicide than others.
Atrazine may have acted as a positive selective factor for cyanobacteria in one month of
their experiments, but at concentrations an order of magnitude higher than
concentrations found in Missisquoi Bay.
Most of the past studies of the cyanobacteria in Missisquoi Bay have been of short
duration (1 to 3 years), of limited spatial coverage, or have been part of broader studies
covering the entire Lake Champlain. The objective of this report is to provide an
integrated analysis of the blooms in the northern (Québec) portion of Missisquoi Bay
from 2000 to 2008 using data from the Québec monitoring program. Compliance with
management guidelines are reported elsewhere (Blais, 2014).
2 Methods
2.1 Field samping and data collection
2.1.1 Sampling sites and strategies
The largest cyanobacterial data sets for the Québec portion of Missisquoi Bay over the
longest periods of record are for sites a, b, d2 and d8. Data analysis for this report has
therefore concentrated on these 4 sites. Their locations are shown in Figure 1. A brief
description of all sites and the number of samples taken at each between 2000 and 2008
is given in Appendix A. The data used in this report are all for samples collected during
both regular sampling and bloom sampling occasions at these four sites between June
and October (summer and autumn in Québec, mostly for the years 2000 to 2008
inclusive). Some data from 2009 was also included in analyses that did not require inter-
year comparisons, but was otherwise not used elsewhere as 2009 data were only
available from late August to mid October and excluded the worst blooms of that
summer.
Site a is located in the deepest sector of the bay and at its centre. The justification for
sites b and d8 is firstly that they are located in important recreational use areas where
major body contact with the water occurs and secondly that there have been frequent
major blooms in these areas in recent years. Site b is also located facing the dominant
winds. Whenever the water level was too low, site b would be relocated further offshore
and as a consequence is considered a sector rather than a specific location. Site d2 is
frequently monitored because it is close to the intake point for the drinking water
treatment plant at Saint-Armand.
From one bloom study to another, site locations either remained unchanged or new ones
were set up. The goal was sampling at accessible locations where blooms and scum
seemed to be the most intense in both the eastern and western (i.e. Baie de Venise)
parts of Missisquoi Bay (access permission obtained from owners of private lands
however decreased over time). If permanent sites a or b were part of a major bloom
sector, they would also be used in the bloom studies. Otherwise, new “d” sites would be
set up or existing “d” sites would be resampled.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
4 NSW Office of Water, July 2014
Figure 1 Location of sampling sites and main public usage areas in Missisquoi Bay. “Patrolled public beach” indicates that at least one lifeguard is on duty. For this report, only data from sites a, b, d2 and d8 were used
2.1.2 Data collection
The cyanobacterial monitoring reported in this report originated from two types of data
collection:
Regular studies that aim at providing a picture of the situation prior to visible blooms.
Generally speaking, these studies take place two or three times a year, at the
beginning of the season, with the last samples generally collected around mid-July.
For 2000, only regular studies took place, since no blooms were observed during that
year.
Bloom studies deal with situations where blooms are present, but representative
water samples were collected rather than concentrated surface scum samples.
Regular studies include the following laboratory-analyzed attributes: cyanobacteria,
eukaryotic algae, pigments such as chlorophyll-a and physicochemical attributes such as
phosphorus and nitrogen. During regular studies because of the absent of any blooms,
cyanotoxins are not usually measured, given the very low probability of detection and/or
measurement in appreciable concentrations. Bloom studies included all the above
attributes plus cyanotoxins.
Regular and bloom studies involved the collection of integrated samples from the
offshore water column from a boat. From 2000 to 2007 inclusive, integrated water

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
5 NSW Office of Water, July 2014
column sampling extended throughout the depth of the photic zone, but in 2008 and
2009 the sampling was restricted to the surface 1 metre (0-1 m). Between 2000 and
2003, samples were collected using a vertical Kemmerer sampling bottle. In 2000,
photic zone sampling was discontinuous, with sub-samples taken at 2-metre intervals,
starting at 30 cm below the water surface. From 2001 to 2003, the sub-samples
incorporated the entire depth of the photic zone on a continuous basis, starting from the
surface. From 2004 to 2007, the depth of the photic zone was sampled a single time
using a flexible plastic tube that was sufficiently long enough to extend through the entire
depth of the water column, independent of wherever the sampling occurred. In 2008 and
2009, 0-1 m samples were collected using a rigid PVC tube. The change from photic
zone to 0-1 m sampling was justified by the fact that the top metre of water is the depth
most frequently used during recreational activities that include water contact.
The depth of the photic zone was calculated by multiplying water transparency
measured with a Secchi disc by a factor of 2.7. This value is based on Cole (1975) and
Holdren et al. (2001), who recommend multiplying Secchi disk transparency by factors of
2.7 to 3 or 2 to 3 respectively. However, when the calculated photic zone reached or
exceeded the water column depth, the entire depth of the water column was sampled
apart from the bottom 30 to 50 cm, the goal being to avoid collection of water
contaminated with bottom sediments. Samples were well mixed before being divided into
attrribute-specific bottles that were subsequently analyzed in the laboratory. Surface
water temperature was measured by thermometer.
Over the entire monitoring period, sampling depth and instrumentation differences (three
different types of instruments were used) may have led to some disparity in results.
However, since the bay is quite large but not very deep and is highly wind-sensitive, as
such the water column is often stirred and is quite probably relatively homogenous.
Moreover, the number of studies and sampling sites per study varied from one year to
the next according to the available human and funding resources.
Meteorological data were obtained for the Philipsburg manual weather station for 2002
to 2008 inclusive. There were two records per day – one for 08:00 and one for 18:00
Québec local time. These were averaged to provide a single daily reading for the days
when phytoplankton samples had been collected. The data for the antecedent 4 days, 7
days and 10 days prior to phytoplankton sample collection were also averaged to obtain
a mean value for each of these antecedent periods. Some data within these antecedent
periods was sometimes missing and only the data available could be used to obtain the
mean value for that period.
2.2 Laboratory analysis
Water samples were analyzed by the MDDEFP’s Centre d’expertise en analyse
environnementale du Québec (CEAEQ). The methods used have been tabulated in
Appendix B. The 2000 cyanobacteria and eukaryotic algae samples were analyzed by
Université du Québec à Montréal (UQAM) which had developed the methodology earlier.
For the subsequent years, cyanobacteria and eukaryotic algae samples were analyzed
by CEAEQ using the expertise obtained by UQAM. Microcystin (MC) variants analyzed
during the entire monitoring period were limited to microcystin-LR (MC-LR), microcystin-
RR (MC-RR) and microcystin-YR (MC-YR).

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
6 NSW Office of Water, July 2014
2.3 Classification of potentially toxic cyanobacteria
These cyanobacteria are those likely to produce hepatotoxins such as microcystin (MC)
or cylindrospermopsin or neurotoxins such as anatoxin-a, saxitoxin or neosaxitoxin, etc.
Cyanobacterial species or types that according to the scientific literature (reviewed by
Blais and Sinotte, 2008) are known to produce at least these cyanotoxins in various
parts of the world were listed to define those potentially toxic species. Using a preventive
approach, we considered that these cyanobacteria are also likely to produce similar
toxins in Québec water bodies.
The taxonomy of a number of cyanobacterial species has been revised recently,
resulting in name changes to some species. In particular, gas-vacuolate planktonic
species from the genus Anabaena have now been assigned to a revised genus,
Dolichospermum (Waklin et al., 2009). A number of these species occur commonly in
Missisquoi Bay, including Anabaena flos-aquae (now Dolichospermum flos-aquae), A.
planctonica (now D. planctonicum), A. spiroides (now D. spiroides) and A. crassa (or A.
spiroides var crassa) (now D. crassum). In addition, Aphanothece clathrata has been
renamed Anathece clathrata (Komárek et al., 2011). However for consistency with
previous reporting from Missisquoi Bay, the previous taxonomic names have been
retained for this report.
2.4 Statistical analysis
The data number (n) often differs between various phytoplankton and water quality
attributes, as not all attributes were analysed for every sampling occasion. This missing
data explains why n often differs from one set of data analyses to another, especially
with the correlation analyses.
Initial exploratory analysis to visualise the data as histograms or scatter plots was
undertaken using Microsoft Excel 2007. STATISTICA v9 was used to undertake
Spearman Rank Order Correlation (Rho – the symbol for Rho, ρ, has not been used in
this report to avoid confusion with p, for probability) analysis between individual
cyanobacterial, eukaryotic algal, toxin and environmental variables. Spearman Rank
Order Correlation, a nonparametric correlation method, was chosen because the data
were not normally distributed, having a large number of low observations for many taxa
of cyanobacteria within the data set and occasional very high values; these would
otherwise lead to the violation of parametric assumptions. This correlation method also
does not simply calculate a linear correlation, but any correlation shape. Where taxa
were not observed in a sample, these were not assigned a zero value for that sample,
but excluded from the analysis. The correlation analyses were undertaken both on the
combined data from sites a, b, d2 and d8, and separately for each site after logarithmic
(log(x)) transformation of the cyanobacterial data and other environmental data where
appropriate, to account for the occasional high values that were skewing the
distributions.
In contrast, the data used for the scatter plots and other figures included zeros, and was
log (x + 1) transformed to avoid negative logarithmic values and to allow zero values to
be shown on either axis.
Despite the multiple comparisons that were made using correlation analysis, all results
having a level of significance of ≤ 0.05 are presented in this report. The exploratory
nature of this study justifies the absence of adjustment for multiple statistical significance
tests. We therefore acknowledge that type I error (i.e. probability of accepting a

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
7 NSW Office of Water, July 2014
correlation coefficient as different from zero when it is not) could be much higher than
5%. On the other hand applying any correction method, such as Bonferroni adjustment,
would in turn increase type II error (i.e. probability of rejecting a correlation coefficient as
non significant when it is actually different from zero), thereby reducing the power of the
test. The reader must keep in mind that some of the correlations presented might have
been declared statistically significant due to chance alone.
Multivariate analysis of the combined data for sites a, b, d2 and d8 was undertaken
using PRIMER v6 (Clarke and Gorley, 2006) and PERMANOVA+ (Anderson et al.,
2008). Separate site-specific analyses were also undertaken for sites a, b and d8, but
not for site d2 as there were insufficient data for this location. Rare species and those
with very low abundance (contributing very little to the total cyanobacterial biomass)
were either excluded from the cyanobacterial data set or rolled up to a genus level of
taxonomy, as appropriate. Samples where no cyanobacteria had been detected were
also excluded. Bray-Curtis similarity coefficients were then calculated using logarithmic
transformed (x + 1) cyanobacterial biomass data which were then used for non-metric
multidimensional scaling (nMDS) ordination and “Similarity, percentages” analysis
(SIMPER) to identify differences between sites, months and years and which
cyanobacterial taxa were contributing to this. The differences in community composition
between sampling sites, months and years were also tested using the Permutational
Multivariate Analysis of Variance (PERMANOVA) technique in the PERMANOVA+
program.
Water quality data from the 4 sites collected over the 2001 to 2008 sampling periods
were combined into an environmental data set containing total phosphorus, total
nitrogen, TN:TP (by concentration), pH, turbidity, surface water temperature, and Secchi
disc depth. Because some water quality attributes were not measured on all sampling
occasions, samples with missing data had to be deleted from the data set prior to
multivariate analysis. Meteorological data from Philipsburg manual weather station
(7026040) was not included as the same data would have been used for all 4 lake
sampling sites (i.e. there should be no differences between sites based on
meteorological data).
The environmental data set was normalised and then subjected to Principal Components
Analysis (PCA) and SIMPER to examine any variation between sites, years and months
using PRIMER v6 (Clarke and Gorley, 2006). The normalised data was also used to
create a resemblance matrix based on Euclidean distance, and PERMANOVA then used
to test for significant differences in the multivariate environmental data between sites,
years and months.
Because the environmental data set contained fewer samples than the cyanobacterial
data base, a second cyanobacterial data set of reduced size was created using only
samples matching those in the environmental data base. This reduced cyanobacterial
data set was then subjected to the same analyses as the original larger data set, so that
these new analytical outcomes could be directly compared with those of the PCA
analyses of the environmental data. This was undertaken to determine which of the
measured environmental attributes, if any, may be driving cyanobacterial community
composition and abundance in Missisquoi Bay. This was done in PRIMER v6 using
RELATE, an analysis which provides a measure of any overall correlation between the
two data sets. RELATE is also a rank correlation test similar to the Spearman Rank
Order Correlation test used previously, thereby providing analytical consistency over all
analyses. When RELATE showed a significant correlation between the environmental

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
8 NSW Office of Water, July 2014
and cyanobacterial data, BEST was used to determine which subset of environmental
variables was most strongly correlated with cyanobacterial community composition.
The reduced cyanobacterial data set and the environmental data set were then split into
data sets for sampling sites a, b and d8, and the 7 day average antecedent
meteorological data for Philipsburg added to each site-specific environmental data set.
There were insufficient data for site d2. Preliminary analysis indicated that surface water
temperature, minimum air temperature and maximum air temperature were all strongly
inter-corelated, containing effectively the same information and therefore 2 of these 3
variables were redundant for the purposes of the multivariate analysis. Likewise cloud
cover and rainfall were also strongly inter-correlated. Because of this, minimum air
temperature, maximum air temperature and rainfall were excluded from the
environmental data sets for further analysis. PCA, PERMANOVA and SIMPER analyses
were then performed on the environmental data for each site, and nMDS, PERMANOVA
and SIMPER on the cyanobacterial data for each site. RELATE and BEST were then
used to determine if there were any site-specific environmental drivers of the
cyanobacterial communities at each site.
3 Results
3.1 Annual variation in cyanobacterial presence at sites a, b, d2 and d8
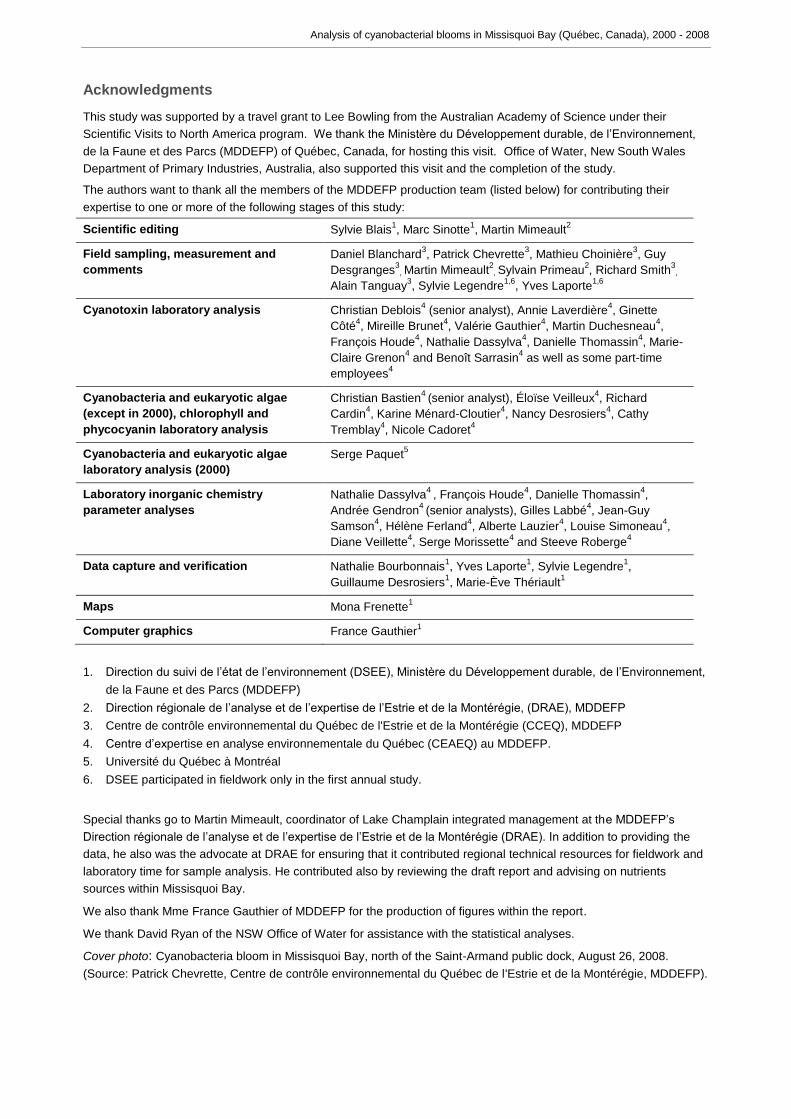
Total cyanobacterial abundance at site a, as cell density (cells ml-1), is shown in Figure
2a, while abundance in terms of biomass is shown in Figure 2b. A summary of all
species occurring each year and their contributions according to biomass is provided in
Appendix C. Cyanobacterial abundance never exceeded 200,400 cells ml-1 (or 11,500
mg m-3 of biomass). Blooms in excess of 50,000 cells ml-1 were present on many
occasions when samples were collected at site a. Low cyanobacterial presence was
recorded here in 2001, but this may result from the site only being sampled in early
summer (June and July) and not throughout the entire bloom season. Cyanobacterial
presence was also low on the five occasions sampled during 2007 (mid monthly from
June to October inclusive). Cyanobacterial presence was also low on most occasions
sampled in 2008, apart from a brief bloom in late August. In terms of biomass however
cyanobacterial presence was low in 2000, 2001 and 2007, when in never exceeded
1000 mg m-3, and also in 2008 when the maximum biomass on the dates sampled never
exceeded 3500 mg m-3. Species composition varied annually, and although there were
high counts of small non-toxic species such as Aphanocapsa spp., Aphanothece spp.,
Chroococcus spp. and Merismopedia spp. present between 2000 and 2004, these often
contributed little to the total cyanobacterial biomass present. Increased counts of
potentially toxic species of Microcystis occurred between 2002 and 2006, and also in
2008, and these tended to dominate the cyanobacterial biomass present during these
years, with a smaller contribution from potentially toxic species of Anabaena. However
Aphanizomenon flos-aquae had become dominant by late September 2006.
Sites b and d2 were sampled more intensively than site a in 2001, and a major bloom in
excess of 1,000,000 cells ml-1 (Figures 3a, 4a) and 120,000 mg m-3 of cyanobacterial
biomass (Figures 3b, 4b) was recorded at both, especially in September. This bloom
was dominated by Aphanizomenon flos-aquae and Microcystis sp. at both sites, in terms
of both cell counts and biomass. Smaller blooms with lower cell counts and biomass,
but generally in excess of 200,000 cells ml-1 and 20,000 mg m-3 biomass occurred at site
b from 2002 through to 2005, and again in 2008 (Figure 3a, b, Appendix C). These
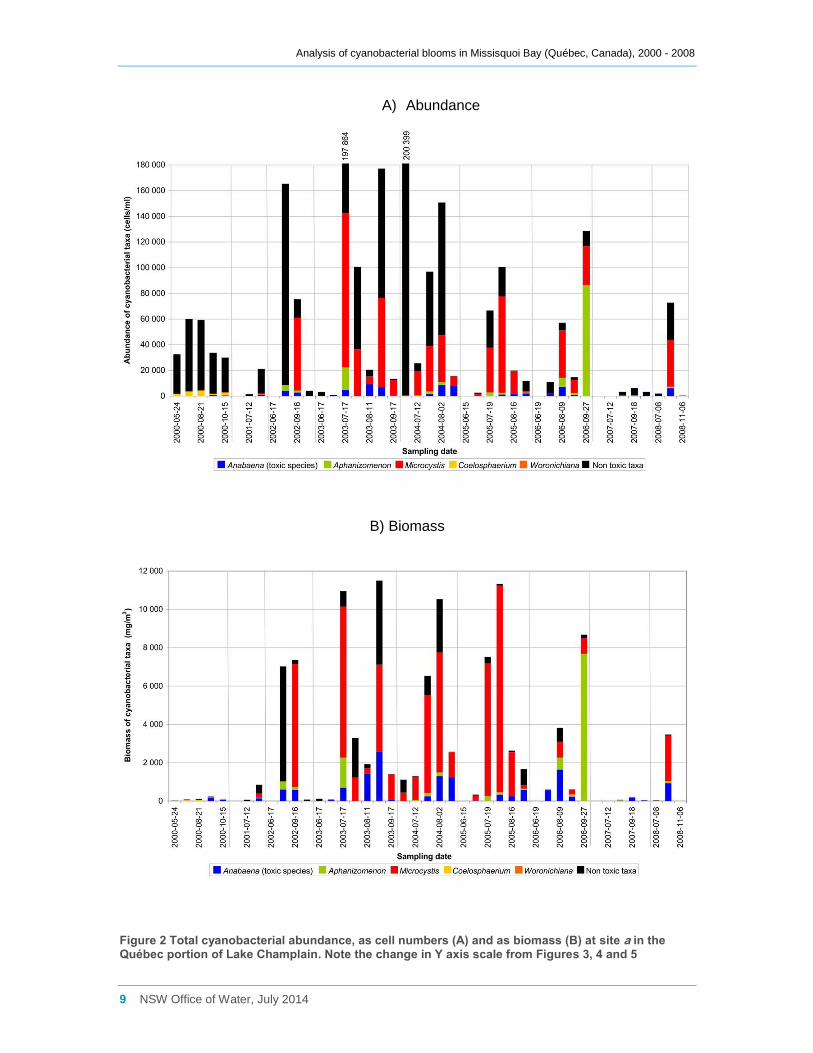
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
9 NSW Office of Water, July 2014
A) Abundance
B) Biomass
Figure 2 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site a in the Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 3, 4 and 5
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
10 NSW Office of Water, July 2014
A) Abundance
B) Biomass
Figure 3 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site b in the Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 2, 4 and 5
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
11 NSW Office of Water, July 2014
A) Abundance
B) Biomass
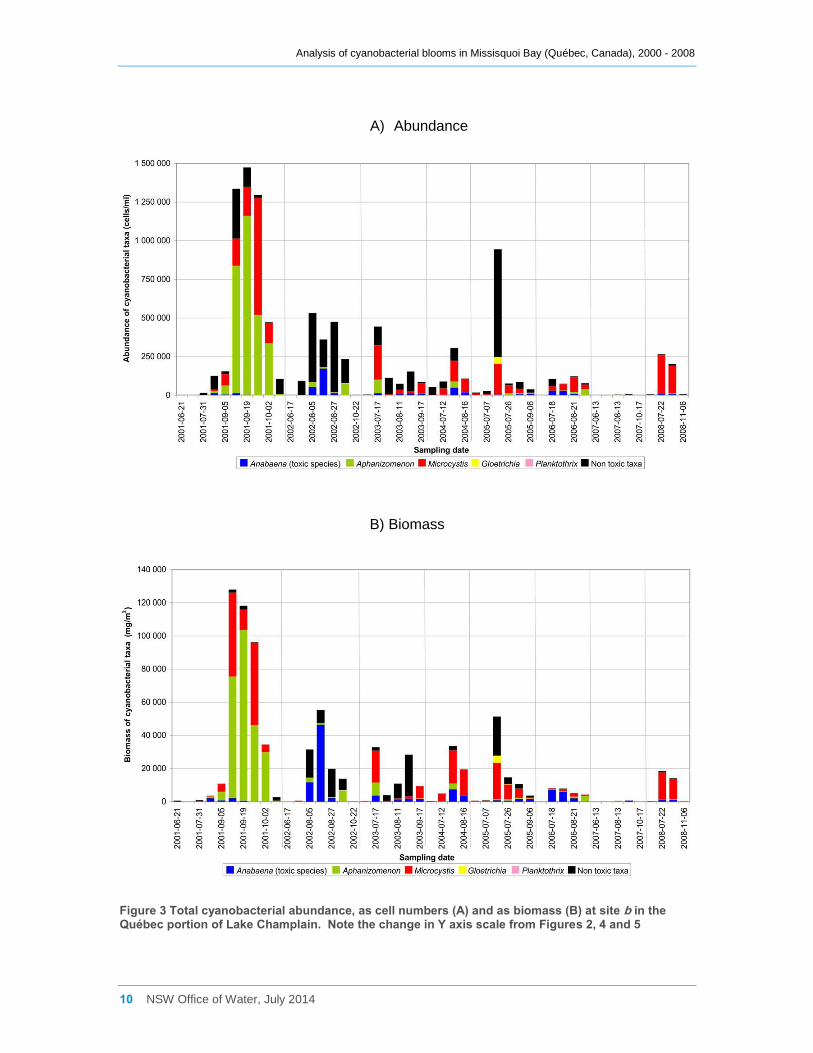
Figure 4 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site d2 in the Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 2, 3 and 5

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
12 NSW Office of Water, July 2014
blooms often displayed a considerable mix of cyanobacterial species present, with
Anabaena spiroides being more common in 2002 and several Microcystis species being
dominant in subsequent years. Chroococcus dispersus and Phormidium mucicola were
the more common non-toxic taxa present during these years. Cyanobacterial presence
at site b during 2006 and especially during 2007 was often low.
Apart from 2001, site d2 was much less frequently sampled than the other sites. A small
bloom of mixed cyanobacterial species was recorded there in early August 2004, and a
larger bloom dominated by Anabaena spiroides was measured in early August 2006
(Figure 4a, b, Appendix C).
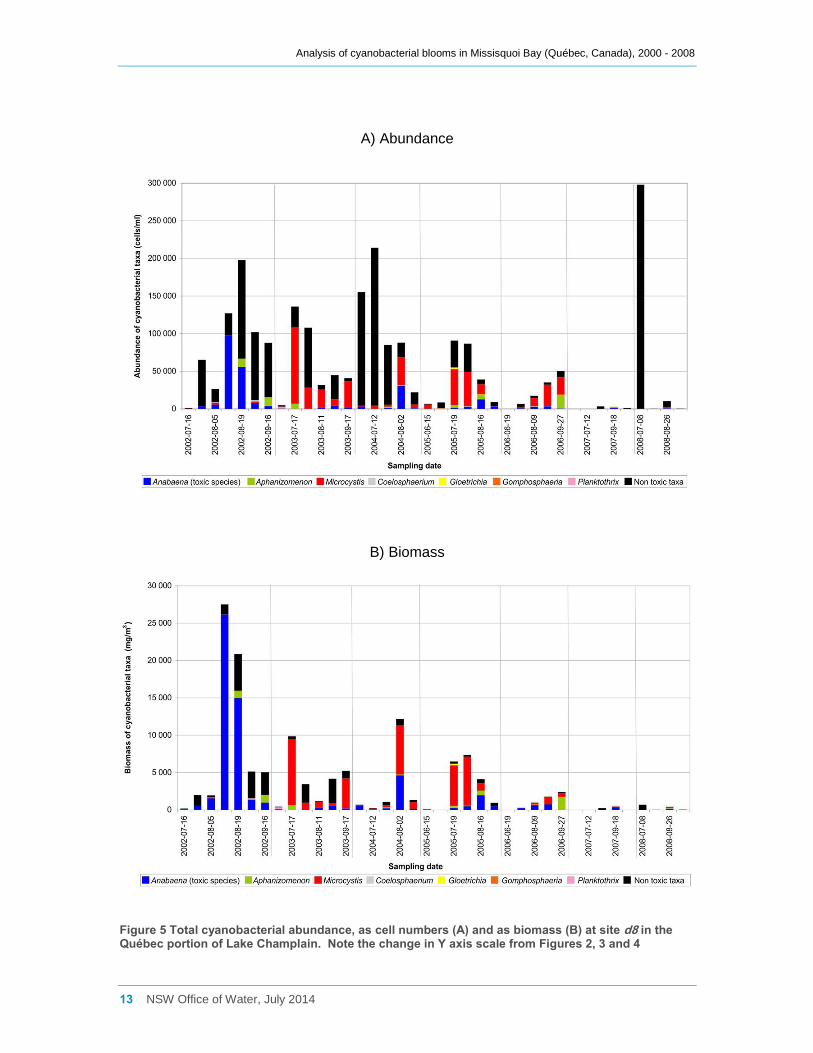
Blooms sampled at site d8 in the north-west corner of Missisquoi Bay near Venise-en-
Québec (in Baie de Venise) generally had a lower cyanobacterial abundance both as
cell counts and total cyanobacterial biomass than sites b and d2 on the eastern side of
the bay. Blooms rarely exceeded 100,000 cells ml-1 or had a total cyanobacterial
biomass greater than 5000 mg m-3 (Figure 5a, b). Anabaena spiroides was dominant in
2002, with a smaller contribution from non-toxic Chroococcus dispersus. In 2004 the cell
counts at site d8 were dominated by non-toxic Aphanocapsa delicatissima, Aphanothece
clathrata and Chroococcus dispersus, although Microcystis species were the dominant
contributors to the total cyanobacterial biomass. Various Microcystis species were also
major contributors to total cyanobacterial biomass in 2003, 2004, 2005 and 2006. Only
a small cyanobacterial biomass was recorded at site d8 during 2007 and 2008, although
Aphanothece clathrata contributed to very high cell counts of non-toxic cyanobacteria in
early August 2008 (Figs 5a, b, Appendix C).
The contribution of potentially toxic species varied considerably both within and between
sites, and is dependent in part on how abundance is measured, whether as cell counts
or as biomass. For example at site a in 2000, potentially toxic cyanobacteria contributed
less the 10% of the total cyanobacterial cell count, but over 50% of the total biomass
because the cell sizes of many of the toxic species present are considerably larger than
the cell sizes of the non-toxic species present. At other times when there are large
numbers of potentially toxic cells present, they can dominate both cell count and
biomass measures, such as at most sites in 2003, 2004, 2005 and 2006.
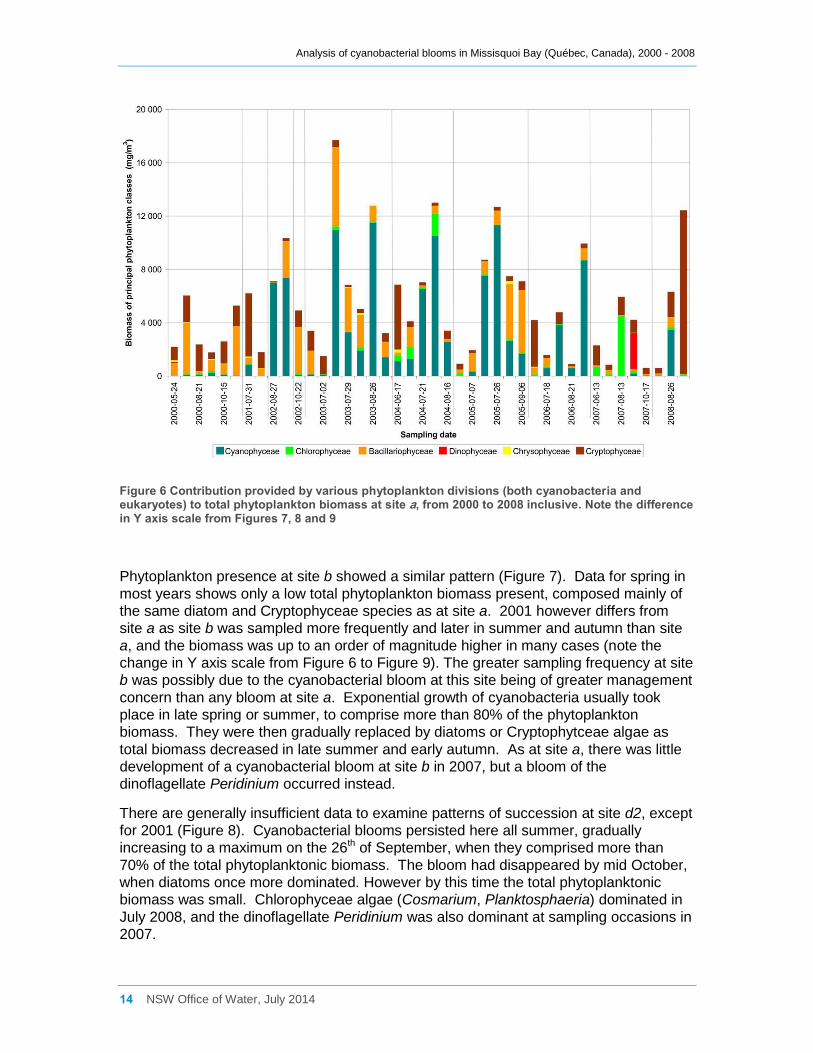
3.2 Contribution of Cyanobacteria to the Total Phytoplankton Biomass
The contributions provided by cyanobacteria to total phytoplankton biomass are shown
for site a in Figure 6. Cyanobacteria comprised less than 15% of the total phytoplankton
biomass in all samples collected at site a in 2000, 2001 and in 2007. The phytoplankton
community in these years was usually dominated by diatoms (also called
Bacillariophyceae) (Cyclotella, Stephanodiscus, Fragellaria and Melosira) and
Cryptophyceae (Cryptomonas, Rhodomonas). The Chlorophyceae (green algae) genus
Pandorina was dominant in August 2007 and the dinoflagellate Peridinium was dominant
in September that year.
Cyanobacterial blooms at site a in other years (2002 – 2006, and 2008) were generally
preceded by a mixed community comprised mainly of diatom and cryptophyte species.
During the blooms, cyanobacteria at times contributed more than 80% of the total
phytoplankton biomass, but the cyanobacteria were usually progressively replaced by
diatoms (and some Cryptophyceae) as the bloom declined. Various Chrysophyceae
species were also frequently present, but usually at low biomass.
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
13 NSW Office of Water, July 2014
A) Abundance
B) Biomass
Figure 5 Total cyanobacterial abundance, as cell numbers (A) and as biomass (B) at site d8 in the Québec portion of Lake Champlain. Note the change in Y axis scale from Figures 2, 3 and 4
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
14 NSW Office of Water, July 2014
Figure 6 Contribution provided by various phytoplankton divisions (both cyanobacteria and eukaryotes) to total phytoplankton biomass at site a, from 2000 to 2008 inclusive. Note the difference in Y axis scale from Figures 7, 8 and 9
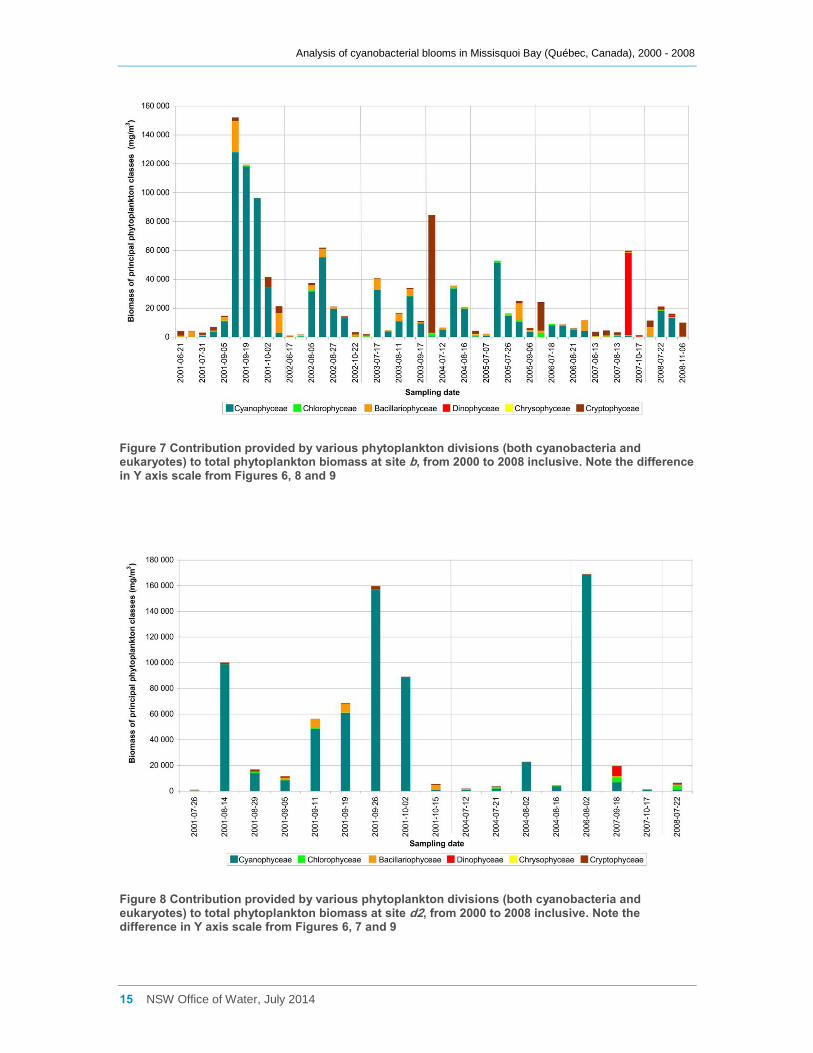
Phytoplankton presence at site b showed a similar pattern (Figure 7). Data for spring in
most years shows only a low total phytoplankton biomass present, composed mainly of
the same diatom and Cryptophyceae species as at site a. 2001 however differs from
site a as site b was sampled more frequently and later in summer and autumn than site
a, and the biomass was up to an order of magnitude higher in many cases (note the
change in Y axis scale from Figure 6 to Figure 9). The greater sampling frequency at site
b was possibly due to the cyanobacterial bloom at this site being of greater management
concern than any bloom at site a. Exponential growth of cyanobacteria usually took
place in late spring or summer, to comprise more than 80% of the phytoplankton
biomass. They were then gradually replaced by diatoms or Cryptophytceae algae as
total biomass decreased in late summer and early autumn. As at site a, there was little
development of a cyanobacterial bloom at site b in 2007, but a bloom of the
dinoflagellate Peridinium occurred instead.
There are generally insufficient data to examine patterns of succession at site d2, except
for 2001 (Figure 8). Cyanobacterial blooms persisted here all summer, gradually
increasing to a maximum on the 26th of September, when they comprised more than
70% of the total phytoplanktonic biomass. The bloom had disappeared by mid October,
when diatoms once more dominated. However by this time the total phytoplanktonic
biomass was small. Chlorophyceae algae (Cosmarium, Planktosphaeria) dominated in
July 2008, and the dinoflagellate Peridinium was also dominant at sampling occasions in
2007.
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
15 NSW Office of Water, July 2014
Figure 7 Contribution provided by various phytoplankton divisions (both cyanobacteria and eukaryotes) to total phytoplankton biomass at site b, from 2000 to 2008 inclusive. Note the difference in Y axis scale from Figures 6, 8 and 9
Figure 8 Contribution provided by various phytoplankton divisions (both cyanobacteria and eukaryotes) to total phytoplankton biomass at site d2, from 2000 to 2008 inclusive. Note the difference in Y axis scale from Figures 6, 7 and 9
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
16 NSW Office of Water, July 2014
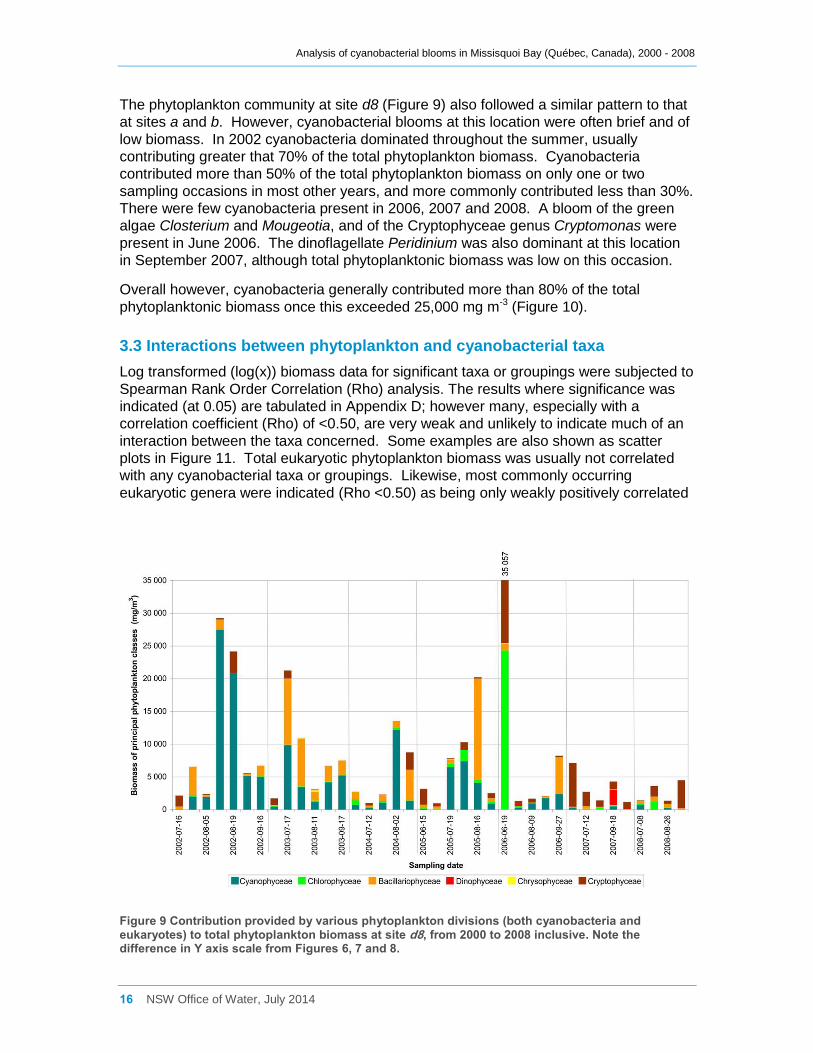
The phytoplankton community at site d8 (Figure 9) also followed a similar pattern to that
at sites a and b. However, cyanobacterial blooms at this location were often brief and of
low biomass. In 2002 cyanobacteria dominated throughout the summer, usually
contributing greater that 70% of the total phytoplankton biomass. Cyanobacteria
contributed more than 50% of the total phytoplankton biomass on only one or two
sampling occasions in most other years, and more commonly contributed less than 30%.
There were few cyanobacteria present in 2006, 2007 and 2008. A bloom of the green
algae Closterium and Mougeotia, and of the Cryptophyceae genus Cryptomonas were
present in June 2006. The dinoflagellate Peridinium was also dominant at this location
in September 2007, although total phytoplanktonic biomass was low on this occasion.
Overall however, cyanobacteria generally contributed more than 80% of the total
phytoplanktonic biomass once this exceeded 25,000 mg m-3 (Figure 10).
3.3 Interactions between phytoplankton and cyanobacterial taxa
Log transformed (log(x)) biomass data for significant taxa or groupings were subjected to
Spearman Rank Order Correlation (Rho) analysis. The results where significance was
indicated (at 0.05) are tabulated in Appendix D; however many, especially with a
correlation coefficient (Rho) of <0.50, are very weak and unlikely to indicate much of an
interaction between the taxa concerned. Some examples are also shown as scatter
plots in Figure 11. Total eukaryotic phytoplankton biomass was usually not correlated
with any cyanobacterial taxa or groupings. Likewise, most commonly occurring
eukaryotic genera were indicated (Rho <0.50) as being only weakly positively correlated
Figure 9 Contribution provided by various phytoplankton divisions (both cyanobacteria and eukaryotes) to total phytoplankton biomass at site d8, from 2000 to 2008 inclusive. Note the difference in Y axis scale from Figures 6, 7 and 8.
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
17 NSW Office of Water, July 2014
Figure 10 The percentage of the total phytoplanktonic biomass contributed by cyanobacteria (sites a, b, d2 and d8, combined)
with total cyanobacterial biomass or with individual species of cyanobacteria. This
probably results from most samples being collected during summer when cyanobacteria
greatly dominate the total phytoplankton biomass, and often a smaller biomass of these
eukaryotic taxa is present (Figure 11a). In addition, possible negative relationships
resulting from seasonal succession within the phytoplankton community (e.g. spring and
autumn dominance by eukaryotic algae when cyanobacteria are uncommon) have not
been shown due to the exclusion of samples with missing data (when a taxon was not
recorded in a sample) from the correlation analysis.
Most cyanobacterial taxa contributed significantly (Rho between 0.64 and 0.90,
Appendix D) to the total cyanobacterial biomass present, but less strongly to the total
phytoplankton biomass. Anabaena flos-aquae was frequently the major contributor to
total Anabaena biomass (Rho = 0.85), as was Microcystis aeruginosa to total Microcystis
biomass (Rho = 0.99). Although many cyanobacterial taxa occurred simultaneously
together in Missisquoi Bay, the strength of the correlation coefficients between them
varied considerably (Appendix D). Only Anabaena spiroides was not significantly
correlated to most other taxa. Likewise, while the examples provided in Figure 11
indicate possible positive trends between several of the cyanobacterial taxa occurring in
Missisquoi Bay, they mostly show a high degree of scatter further revealing that any
relationships between the taxa are generally not strong. While these analyses provide
an indication of which taxa co-occur more frequently together in Missisquoi Bay and that
their contributions to total cyanobacterial and total phytoplankton biomass in summer
can be considerable, overall there is much variation in the strengths of these
interactions.
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
18 NSW Office of Water, July 2014
3.4 Cyanobacteria and toxin presence
Total microcystin-LR concentrations (intracellular plus extracellular) at site a were
generally low, and only exceeded 1.0 µg l-1 on two occasions, in July 2005 and August
2008 (Figure 12). Similarly low concentrations were also recorded at site d8, with
concentrations there only exceeding 1.0 µg l-1 in late July 2005 (Figure 15). Higher
concentrations, exceeding 5 µg l-1 occurred at site b in August 2002 and in July 2008
(Figure 13) and exceeding 12 µg l-1 in July 2005. Likewise a total microcystin-LR
Figure 11 Examples of some of the interactions between various species or groupings of phytoplankton within Missisquoi Bay. Spearman Rank Order Correlation coefficients (Rho) (with missing data excluded), number of observations (n) and probabilities (p) for each of the five panels are as follows: Panel a - total eukaryotic phytoplankton biomass and total cyanobacterial biomass (Rho = 0.01, n = 148, p = 0.86); Panel b – total Anabaena biomass and total Microcystis biomass (Rho = 0.49, n = 87, p < 0.0001); Panel c – Anabaena flos-aquae biomass and Aphanizomenon flos-aquae biomass (Rho = 0.40, n = 44, p = 0.007); Panel d – Anabaena flos-aquae biomass and Microcystis sp. biomass (Rho = 0.33, n = 42, p = 0.03); and Panel e – Aphanizomenon flos-aquae biomass and Microcystis sp. biomass (Rho = 0.75, n = 44, p < 0.0001). Note that for a significant number of samples there were missing data for one of the two groupings or species in most panels. Using log (x + 1) transformation allows these to be shown as 0 on either axis.
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
19 NSW Office of Water, July 2014
concentration greater than 10 µg l-1 occurred at site d2 in early October 2001 (Figure
14). The higher concentrations (i.e > 0.6 µg l-1) of microcystin-LR were found when total
Microcystis biomass exceeded 300 mg m-3 (data not shown). On almost all sampling
occasions, the majority (>80% and often >90%) of the total microcystin-LR was
contained within the cyanobacterial cells, rather than as free microcystin-LR within the
water column (data not shown).
Concentrations of total microcystin-RR and total microcystin-YR (intracellular plus
extracellular) also occurred together with microcystin-LR on some occasions, and at
times concentrations of these microcystins exceeded those of microcystin-LR. In
particular, total microcystin-RR was present at its highest concentrations (1.95 µg l-1 at
site a, 2.94 µg l-1 at site b and 0.624 µg l-1 at site d8) in July 2005 (Figure 12, 13 and 15,
respectively). The highest concentration at site d2 was 0.60 µg l-1 in August 2006 (this
site was not sampled in 2005). High total microcystin-YR concentrations of 0.70 to 1.11
µg l-1 occurred at sites a and d2 in 2004 (Figures 12 and 14), while a concentration of
2.083 µg l-1 was measured for site b in mid July 2005 (Figure 13). On most occasions
sampled intracellular concentrations of both microcystin-RR and microcystin-YR
predominated.
Figure 12 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site a. Note the difference in Y axis scale from Figures 13, 14 and 15

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
20 NSW Office of Water, July 2014
Figure 13 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site b. Note the difference in Y axis scale from Figures 12, 14 and 15
Figure 14 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site d2. Note the difference in Y axis scale from Figures 12, 13 and 15
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
21 NSW Office of Water, July 2014
Figure 15 Presence of microcystin-LR, microcystin-RR and microcystin-YR at site d8. Note the difference in Y axis scale from Figures 12, 13 and 14
As can be seen in Figures 12 to 15, microcystin-LR is the variant most often detected,
nevertheless the relationships between concentrations of total microcystin, microcystin-
LR, microcystin-RR and microcystin-YR were investigated by grouping all the data from
sites a, b, d2 and d8 together, removing any values that were at or below the level of
detection and converting the data to a logarithmic scale (to allow for better presentation
due to the skewed distribution of the data). Scatter plots (Figure 16) and correlation
analyses (Table 1) of each microcystin variant against the others indicate that the
concentrations of microcystin-LR and microcystin-RR, and to a lesser extent of
microcystin-LR and microcystin-YR, tend to increase approximately in unison. However,
apart from the correlation between microcystin-LR and microcystin-RR (Rho = 0.70), the
other relationships were weak and that between microcystin-RR and microcystin-YR did
not even reach significance.
Total cyanobacterial biomass was significantly positively correlated (at the 5% level of
significance or lower) with total microcystin concentration and also with microcystin-LR,
microcystin-RR and microcystin-YR concentrations (Table 1), although Rho was
generally below 0.50. Total Microcystis biomass and Microcystis aeruginosa biomass
however were frequently more strongly correlated with the microcystin variants (Rho
usually > 0.50). Total microcystin concentration and the concentrations of microcystin-
LR, microcystin-RR and microcystin-YR all tended to increase with increasing total
Microcystis biomass (Figure 17), but similar patterns were present for Microcystis
aeruginosa and Microcystis sp. In fact Microcystis aeruginosa biomass has the highest
correlations with microcystin concentrations, either total or by variant (Table 1). M.
aeruginosa was the dominant cyanobacterial species present in terms of biomass in
37% of samples where total microcystin concentrations exceeded 1 μg l-1, including
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
22 NSW Office of Water, July 2014
Figure 16 Concentrations of microcystin-LR, RR and YR compared against each other, for sites a, b, d2 and d8 on Missisquoi Bay combined, presented on a logarithmic scale. Data below the level of detection have been removed. Spearman Rank Order Correlation coefficients (Rho), number of observations (n) and and probabilities (p) for each of the three panels are as follows: MC-LR and MC-RR, Rho = 0.70, n = 47, p <0.0001; MC-RR and MC-YR, Rho = 0.29, n = 44, p = 0.06; MC-LR and MC-TR, Rho = 0.43, n = 47, p = 0.003.
Table 1 Spearman Rank Order Correlation coefficients (Rho) for variants of microcystins and for the biomass of Microcystis spp, Anabaena spp. and Aphanizomenon flos-aquae for the combined data collected from sites a, b, d2 and d8 in the Québec portion of Missisquoi Bay. The data were log transformed with values below the level of detection excluded prior to the analysis. Note: No attempt was done to adjust for multiple statistical significance testing, therefore some correlations presented below might be declared significant by chance alone.
Attributes n Rho p
MC-LR and MC-RR 47 .70 <.001
MC-RR and MC-YR 44 .29
MC-LR and MC-YR 47 .43 .003
Total Cyanobacterial Biomass and Total MC 84 .44 <.001
Total Cyanobacterial Biomass and MC-LR 84 .49 <.001
Total Cyanobacterial Biomass and MC-RR 47 .55 <.001
Total Cyanobacterial Biomass and MC-YR 47 .33 .02
Total Microcystis Biomass and Total MC 70 .64 <.001
Total Microcystis Biomass and MC-LR 70 .62 <.001
Total Microcystis Biomass and MC-RR 43 .48 .001
Total Microcystis Biomass and MC-YR 43 .48 .001
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
23 NSW Office of Water, July 2014
Microcystis aeruginosa Biomass and Total MC 20 .86 <.001
Microcystis aeruginosa Biomass and MC-LR 20 . 83 <.001
Microcystis aeruginosa Biomass and MC-RR 19 .53 .02
Microcystis aeruginosa Biomass and MC-YR 16 .73 .001
Microcystis flos-aquae Biomass and Total MC 7 .61
Microcystis flos-aquae Biomass and MC-LR 7 .50
Microcystis flos-aquae Biomass and MC-RR 7 .43
Microcystis flos-aquae Biomass and MC-YR 7 .61
Microcystis sp. Biomass and Total MC 48 .48 <.001
Microcystis sp. Biomass and MC-LR 48 .56 <.001
Microcystis sp. Biomass and MC-RR 22 .47 .03
Microcystis sp. Biomass and MC-YR 23 .11
Total Anabaena Biomass and Total MC 69 .21
Total Anabaena Biomass and MC-LR 69 .26 .03
Total Anabaena Biomass and MC-RR 41 .23
Total Anabaena Biomass and MC-YR 40 .21
Anabaena flos-aquae Biomass and Total MC 47 .15
Anabaena flos-aquae Biomass and MC-LR 47 .11
Anabaena flos-aquae Biomass and MC-RR 27 .02
Anabaena flos-aquae Biomass and MC-YR 28 .24
Anabaena spiroides Biomass and Total MC 34 .22
Anabaena spiroides Biomass and MC-LR 34 .28
Anabaena spiroides Biomass and MC-RR 23 .14
Anabaena spiroides Biomass and MC-YR 22 .37
Aphanizomenon flos-aquae Biomass and Total MC 64 .14
Aphanizomenon flos-aquae Biomass and MC-LR 64 .24
Aphanizomenon flos-aquae Biomass and MC-RR 35 .10
Aphanizomenon flos-aquae Biomass and MC-YR 35 -.12
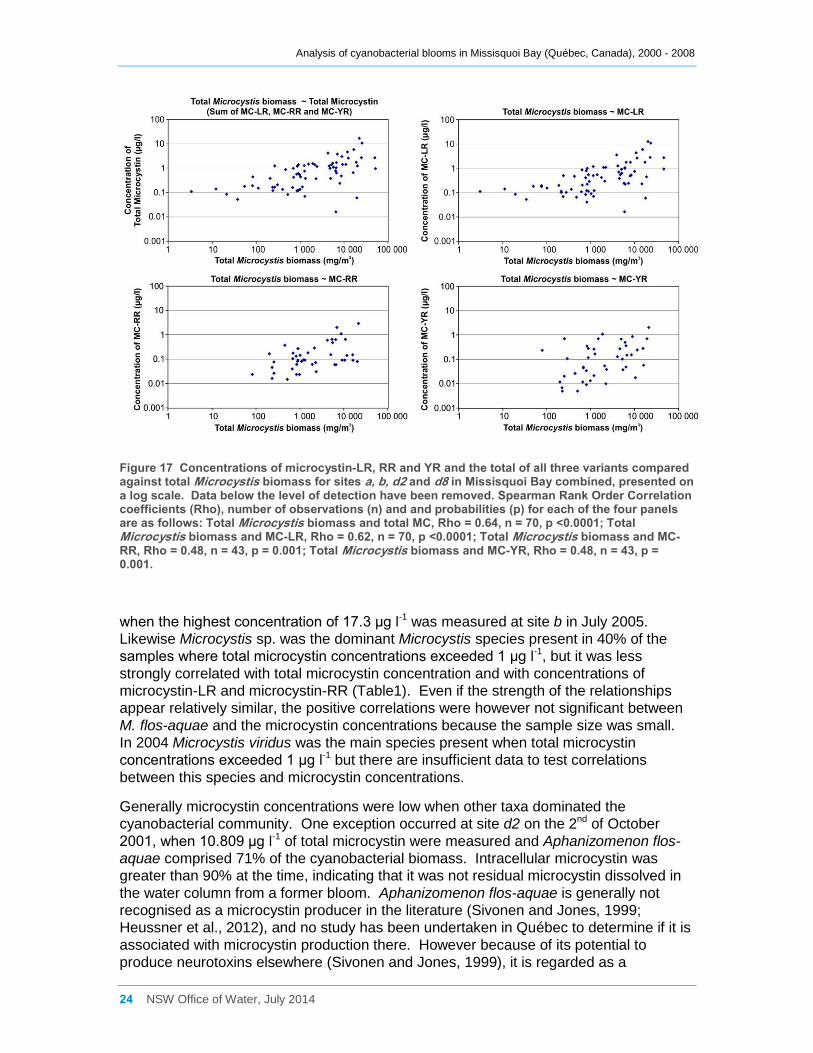
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
24 NSW Office of Water, July 2014
Figure 17 Concentrations of microcystin-LR, RR and YR and the total of all three variants compared against total Microcystis biomass for sites a, b, d2 and d8 in Missisquoi Bay combined, presented on a log scale. Data below the level of detection have been removed. Spearman Rank Order Correlation coefficients (Rho), number of observations (n) and and probabilities (p) for each of the four panels are as follows: Total Microcystis biomass and total MC, Rho = 0.64, n = 70, p <0.0001; Total Microcystis biomass and MC-LR, Rho = 0.62, n = 70, p <0.0001; Total Microcystis biomass and MC-RR, Rho = 0.48, n = 43, p = 0.001; Total Microcystis biomass and MC-YR, Rho = 0.48, n = 43, p = 0.001.
when the highest concentration of 17.3 μg l-1 was measured at site b in July 2005.
Likewise Microcystis sp. was the dominant Microcystis species present in 40% of the
samples where total microcystin concentrations exceeded 1 μg l-1, but it was less
strongly correlated with total microcystin concentration and with concentrations of
microcystin-LR and microcystin-RR (Table1). Even if the strength of the relationships
appear relatively similar, the positive correlations were however not significant between
M. flos-aquae and the microcystin concentrations because the sample size was small.
In 2004 Microcystis viridus was the main species present when total microcystin
concentrations exceeded 1 μg l-1 but there are insufficient data to test correlations
between this species and microcystin concentrations.
Generally microcystin concentrations were low when other taxa dominated the
cyanobacterial community. One exception occurred at site d2 on the 2nd of October
2001, when 10.809 μg l-1 of total microcystin were measured and Aphanizomenon flos-
aquae comprised 71% of the cyanobacterial biomass. Intracellular microcystin was
greater than 90% at the time, indicating that it was not residual microcystin dissolved in
the water column from a former bloom. Aphanizomenon flos-aquae is generally not
recognised as a microcystin producer in the literature (Sivonen and Jones, 1999;
Heussner et al., 2012), and no study has been undertaken in Québec to determine if it is
associated with microcystin production there. However because of its potential to
produce neurotoxins elsewhere (Sivonen and Jones, 1999), it is regarded as a

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
25 NSW Office of Water, July 2014
potentially toxic species in Québec (Blais and Sinotte, 2008). In early October 2001
most of the remaining 29% of potentially toxic cyanobacteria at site d2 was Microcystis
sp., which was nevertheless present at high biomass (25,282 mg m-3) and most likely
produced the microcystin on this occasion. Higher microcystin concentrations also
occurred at site b in August 2002 and at site d2 in August 2006 when Anabaena
spiroides was dominant in terms of biomass. Despite this, correlation analysis found no
significant correlations between Anabaena spirodes and any microcystin concentration,
or between Aphanizomenon flos-aquae and any microcystin concentration (Table 1).
Relationships between microcystin presence and a range of physico-chemical attributes
of the lake water were also examined. Microcystin concentrations may possibly co-vary
with environmental factors in the same manner as cyanobacterial biomass, especially
total Microcystis biomass. For example, microcystin concentrations (microcystin-LR,
microcystin-RR, microcystin-YR, and total microcystin) were significantly positively
correlated with increasing chlorophyll-a concentration and also with pH and turbidity
(Table E1 of Appendix E). The majority of the correlation coefficients (Rho) were
however weak, with a value of under 0.50. The correlations with chlorophyll-a and
turbidity may simply be due to increasing concentrations of toxin-producing
cyanobacteria within the water column, which would cause increases in both chlorophyll-
a concentrations and turbidity. Increased photosynthesis by increased cyanobacterial
biomass also causes pH to increase. Total microcystin, microcystin-LR, microcystin-RR
and microcystin-YR were also weakly (Rho < 0.50) negatively correlated with euphotic
depth, possibly resulting from increasing cyanobacterial biomass (with associated
increased microcystin production) causing a reduction in light penetration through the
water column. Scatter diagrams of total microcystin concentrations against various other
water quality attributes can be also found in Appendix E.
Microcystin-LR, microcystin-YR and total microcystin concentrations were also positively
correlated with total phosphorus concentrations, while total microcystin (seemingly
driven by microcystin-YR) was weakly correlated with the TN:TP ratio, and microcystin-
LR appeared negatively correlated (n = 18) with oxidised nitrogen concentrations. Most
of these correlations were however of borderline significance (Table E1 of Appendix E),
as Rho was frequently less than 0.50. Although microcystin-RR and microcystin-YR
concentrations both showed a significant positive correlation with water temperature,
being present mainly when the temperature was above 20°C (Figure 18), this was weak
(Rho again < 0.50), and water temperature appears to have little influence on
microcystin-LR concentrations.
Concentrations of saxitoxin, neosaxitoxin and cylindrospermopsin were always below the
detection limit of the analytical method. Anatoxin-a was detected only once in 180
samples that were analysed for this toxin.
3.5 Eukaryotic and cyanobacterial presence and water quality indicators
3.5.1 Analysis of the combined data for sites a, b, d2 and d8
Cyanobacterial and other commonly occurring eukaryotic phytoplankton (Chlorella,
Closterium, Cosmarium, Cryptomonas, Cyclotella, Fragilaria, Melosira, Oocystis,
Peridinium, Planktosphaeria, Rhodomonas, Scenedesmus and Stephanodiscus) have
been examined against a range of water quality attributes. Chlorella, Oocystis and
Scenedesmus were chosen as they occurred in >40% of samples, whereas the other
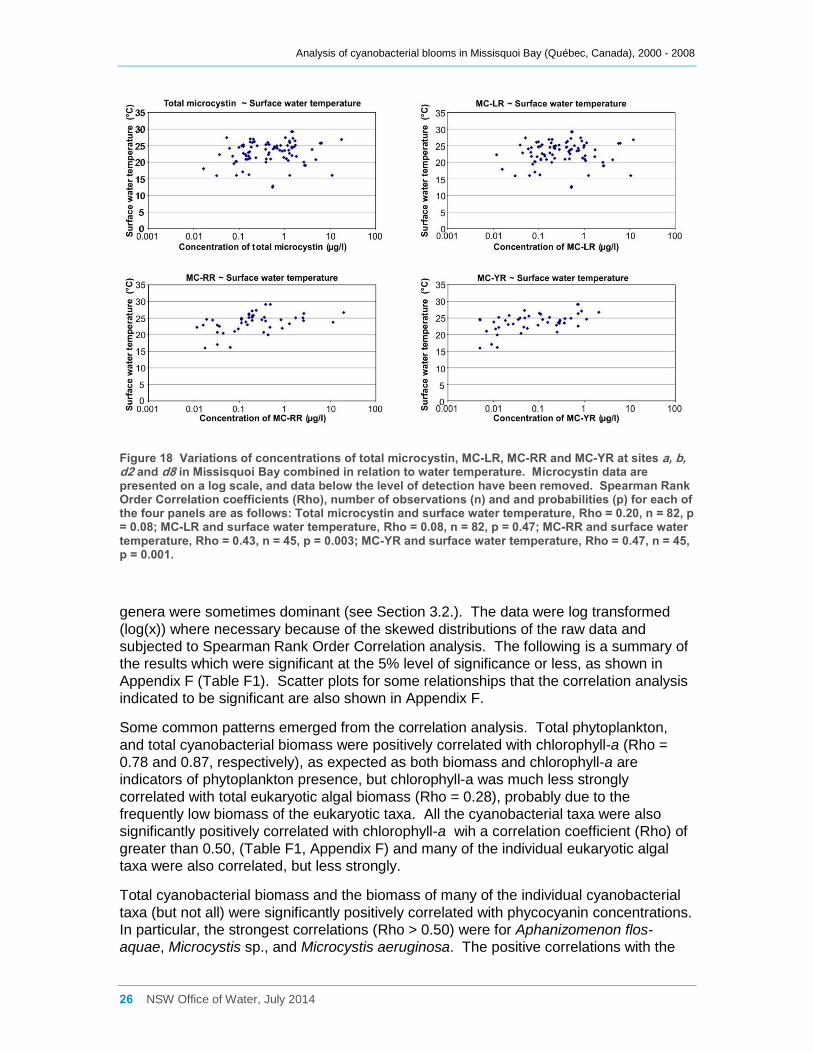
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
26 NSW Office of Water, July 2014
Figure 18 Variations of concentrations of total microcystin, MC-LR, MC-RR and MC-YR at sites a, b, d2 and d8 in Missisquoi Bay combined in relation to water temperature. Microcystin data are presented on a log scale, and data below the level of detection have been removed. Spearman Rank Order Correlation coefficients (Rho), number of observations (n) and and probabilities (p) for each of the four panels are as follows: Total microcystin and surface water temperature, Rho = 0.20, n = 82, p = 0.08; MC-LR and surface water temperature, Rho = 0.08, n = 82, p = 0.47; MC-RR and surface water temperature, Rho = 0.43, n = 45, p = 0.003; MC-YR and surface water temperature, Rho = 0.47, n = 45, p = 0.001.
genera were sometimes dominant (see Section 3.2.). The data were log transformed
(log(x)) where necessary because of the skewed distributions of the raw data and
subjected to Spearman Rank Order Correlation analysis. The following is a summary of
the results which were significant at the 5% level of significance or less, as shown in
Appendix F (Table F1). Scatter plots for some relationships that the correlation analysis
indicated to be significant are also shown in Appendix F.
Some common patterns emerged from the correlation analysis. Total phytoplankton,
and total cyanobacterial biomass were positively correlated with chlorophyll-a (Rho =
0.78 and 0.87, respectively), as expected as both biomass and chlorophyll-a are
indicators of phytoplankton presence, but chlorophyll-a was much less strongly
correlated with total eukaryotic algal biomass (Rho = 0.28), probably due to the
frequently low biomass of the eukaryotic taxa. All the cyanobacterial taxa were also
significantly positively correlated with chlorophyll-a wih a correlation coefficient (Rho) of
greater than 0.50, (Table F1, Appendix F) and many of the individual eukaryotic algal
taxa were also correlated, but less strongly.
Total cyanobacterial biomass and the biomass of many of the individual cyanobacterial
taxa (but not all) were significantly positively correlated with phycocyanin concentrations.
In particular, the strongest correlations (Rho > 0.50) were for Aphanizomenon flos-
aquae, Microcystis sp., and Microcystis aeruginosa. The positive correlations with the

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
27 NSW Office of Water, July 2014
two eukaryotic algal taxa are probably due to chance or their occurrence at the same
time as cyanobacterial, as these taxa should not contain phycocyanin.
Most cyanobacterial biomass indicators were significantly positively correlated (Rho
often > 0.5) with total phosphorus concentrations, with higher biomass associated with
higher concentrations of this major nutrient. Although total eukaryotic algal biomass was
also positively (but weakly) correlated with increasing total phosphorus, there were few
significant correlations between total phosphorus and the eukaryotic taxa tested (Table
F1, Appendix F), and those that were present are weak. There were few significant
correlations between either cyanobacteria or eukaryotic algal taxa and total nitrogen.
Many of the cyanobacterial biomass indicators were however significantly negatively
correlated with oxidised nitrogen (NOx) concentrations with Rho values between -0.63
and -0.85, suggesting that oxidised nitrogen is not a main source of nitrogenous nutrient
for the cyanobacteria in Missisquoi Bay, or that NOx is rapidly utilised. The biomass of
many cyanobacterial taxa was also significantly negatively correlated with the TN:TP
ratio (with Rho values between -0.32 and -0.56), apparently reflecting the positive
correlations with TP in the univariate analysis described above (Table F1, Appendix F).
Few eukaryotic algal indicators were significantly correlated with total nitrogen, NOx and
with the TN:TP ratio.
Increasing cyanobacterial biomass and eukaryotic algal biomass were frequently
significantly and positively correlated with pH, often with correlation coefficients (Rho)
above 0.50, and likewise cyanobacterial biomass with turbidity. Higher pH can be
attributed to the effects of photosynthesis, although cyanobacteria are also advantaged
by a pH of 8.0 or more (Shapiro, 1990). Increased turbidity could be due in part to the
increased number of cyanobacterial cells and colonies entrained within the water
column. Increasing water temperature may also favour increased biomass of the more
common cyanobacterial and eukaryotic algal taxa present in Missisquoi Bay, although
correlations were weaker (Rho often <0.50). Several groupings (Anabaena taxa in
particular) were positively correlated with dissolved organic carbon. Euphotic depth was
also negatively correlated with almost all taxa and groupings of cyanobacteria and with
three eukaryotic algal taxa, although the strength of these correlations varied
considerably (Table F1, Appendix F). This may be due to increased phytoplankton
presence within the water column reducing water column transparency and light
penetration (turbidity and euphotic depth were also significantly negatively correlated).
Electrical conductivity and dissolved oxygen had little effect on either cyanobacterial or
eukaryotic algal biomass.
3.5.2 Site by site analysis
Although there were often few oxidised nitrogen and ammonia data available for some
sites (Table 2), Spearman Rank Order Correlation analysis were also undertaken on a
site by site basis to determine if there were different responses between various
cyanobacterial and eukaryotic algal taxa and the water quality variables at different sites.
This was because some water quality variables vary spatially across the four sites on
Missisquoi Bay (Table 2). In particular, sites b and d2 tended to have higher TP
concentrations than sites a and d8, as well as slightly higher pH values, higher turbidity,
lower TN:TP ratios and shallower Secchi disc depths.
The results of the site by site correlation analysis are provided in Table F2, Appendix F.
Note that Table F2 provides only a summary of those correlations found to be significant
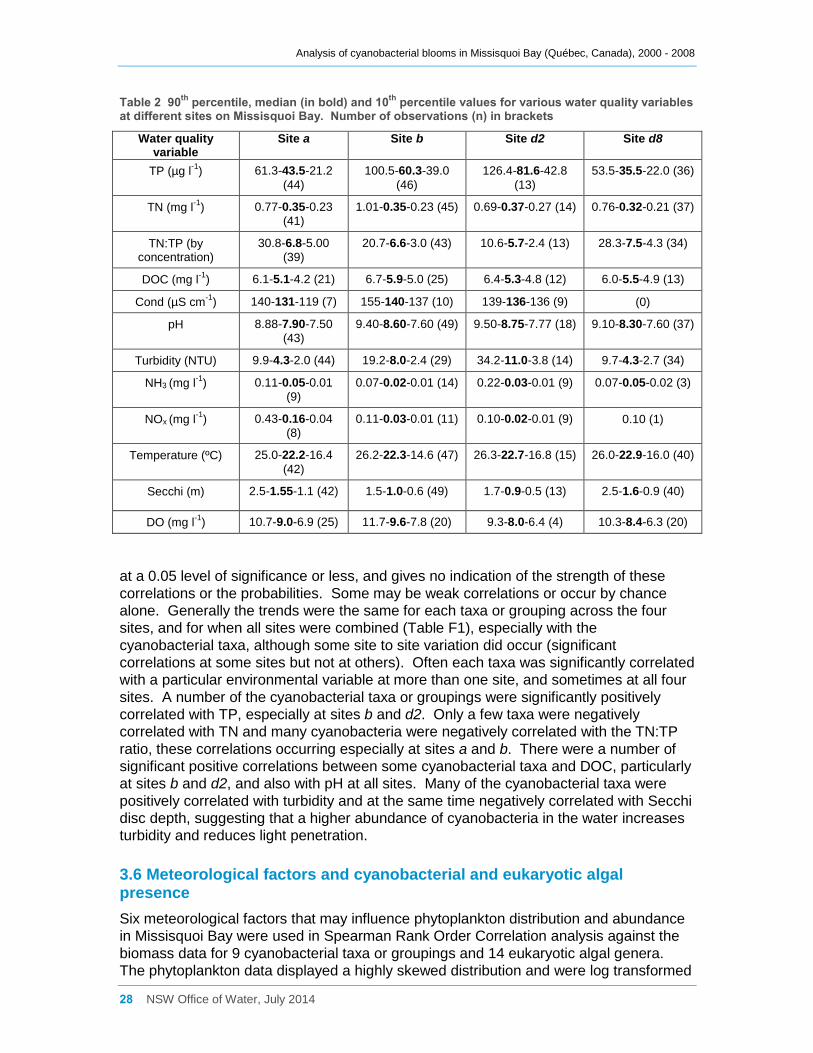
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
28 NSW Office of Water, July 2014
Table 2 90th
percentile, median (in bold) and 10th
percentile values for various water quality variables at different sites on Missisquoi Bay. Number of observations (n) in brackets
Water quality variable
Site a Site b Site d2 Site d8
TP (µg l-1
) 61.3-43.5-21.2
(44)
100.5-60.3-39.0
(46)
126.4-81.6-42.8
(13)
53.5-35.5-22.0 (36)
TN (mg l-1
) 0.77-0.35-0.23
(41)
1.01-0.35-0.23 (45) 0.69-0.37-0.27 (14) 0.76-0.32-0.21 (37)
TN:TP (by concentration)
30.8-6.8-5.00
(39)
20.7-6.6-3.0 (43) 10.6-5.7-2.4 (13) 28.3-7.5-4.3 (34)
DOC (mg l-1
) 6.1-5.1-4.2 (21) 6.7-5.9-5.0 (25) 6.4-5.3-4.8 (12) 6.0-5.5-4.9 (13)
Cond (µS cm-1
) 140-131-119 (7) 155-140-137 (10) 139-136-136 (9) (0)
pH 8.88-7.90-7.50
(43)
9.40-8.60-7.60 (49) 9.50-8.75-7.77 (18) 9.10-8.30-7.60 (37)
Turbidity (NTU) 9.9-4.3-2.0 (44) 19.2-8.0-2.4 (29) 34.2-11.0-3.8 (14) 9.7-4.3-2.7 (34)
NH3 (mg l-1
) 0.11-0.05-0.01
(9)
0.07-0.02-0.01 (14) 0.22-0.03-0.01 (9) 0.07-0.05-0.02 (3)
NOx (mg l-1
) 0.43-0.16-0.04
(8)
0.11-0.03-0.01 (11) 0.10-0.02-0.01 (9) 0.10 (1)
Temperature (ºC) 25.0-22.2-16.4
(42)
26.2-22.3-14.6 (47) 26.3-22.7-16.8 (15) 26.0-22.9-16.0 (40)
Secchi (m) 2.5-1.55-1.1 (42) 1.5-1.0-0.6 (49) 1.7-0.9-0.5 (13) 2.5-1.6-0.9 (40)
DO (mg l-1
) 10.7-9.0-6.9 (25) 11.7-9.6-7.8 (20) 9.3-8.0-6.4 (4) 10.3-8.4-6.3 (20)
at a 0.05 level of significance or less, and gives no indication of the strength of these
correlations or the probabilities. Some may be weak correlations or occur by chance
alone. Generally the trends were the same for each taxa or grouping across the four
sites, and for when all sites were combined (Table F1), especially with the
cyanobacterial taxa, although some site to site variation did occur (significant
correlations at some sites but not at others). Often each taxa was significantly correlated
with a particular environmental variable at more than one site, and sometimes at all four
sites. A number of the cyanobacterial taxa or groupings were significantly positively
correlated with TP, especially at sites b and d2. Only a few taxa were negatively
correlated with TN and many cyanobacteria were negatively correlated with the TN:TP
ratio, these correlations occurring especially at sites a and b. There were a number of
significant positive correlations between some cyanobacterial taxa and DOC, particularly
at sites b and d2, and also with pH at all sites. Many of the cyanobacterial taxa were
positively correlated with turbidity and at the same time negatively correlated with Secchi
disc depth, suggesting that a higher abundance of cyanobacteria in the water increases
turbidity and reduces light penetration.
3.6 Meteorological factors and cyanobacterial and eukaryotic algal presence
Six meteorological factors that may influence phytoplankton distribution and abundance
in Missisquoi Bay were used in Spearman Rank Order Correlation analysis against the
biomass data for 9 cyanobacterial taxa or groupings and 14 eukaryotic algal genera.
The phytoplankton data displayed a highly skewed distribution and were log transformed

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
29 NSW Office of Water, July 2014
(log(x)) prior to use. Four timeframes (day of sampling, 4 day antecedent, 7 day
antecedent, 10 day antecedent) were available for the meteorological data. Analyses
were performed for each site (a, b, d2 and d8) and for the 4 sites combined
Only the correlations found to be significant at a probability of ≤0.05 are shown in the
tables in Appendix G. Most correlations between the meteorological variables and the
phytoplankton taxonomic groupings were not significant. Of the cyanobacteria that were
significant, most were weakly correlated (Rho < 0.50), and sometimes the significance
possibly results from the high number of observations involved (the higher the n, the
lower the coefficient value that becomes significant at the 5% level of significance),
especially when all sites were combined. Moreover, due to the multiple correlations
tested, some significance by chance alone cannot be excluded. Examples of
scatterplots where the strongest correlations were obtained between cyanobacteria
(from all four sites combined) and meteorological conditions are shown in Figure 19.
With the exception of Aphanizomenon flos-aquae, maximum and minimum air
temperatures when significantly correlated to cyanobacterial presence were positively
correlated. Significant correlations for minimum air temperature and for maximum air
temperature frequently coincided. The greater number of significant correlations
between the 7 day mean antecedent maximum temperature and cyanobacterial taxa
suggest that a week or more of hotter weather prior to sampling may lead to increased
cyanobacterial presence. Maximum air temperature on the same day of sampling was
only occasionally correlated with most cyanobacterial presence. Significant correlations
were found on occasions for all sites seperately, although the data are generally
insufficient to make a confident assessment of the effect of air temperature on
cyanobacterial distributions across the bay.
There were few significant correlations between cyanobacteria and wind direction.
Cyanobacterial presence tended to be negatively correlated with wind speed. Longer
antecedent conditions (10 days) of stronger winds are more likely to reduce
cyanobacterial presence than shorter periods, especially with the Anabaena species.
The results of the correlation analysis are insufficient to be conclusive about spatial
effects of either wind direction or wind velocity on the distribution of cyanobacteria in
Missisquoi Bay.
Both rainfall and cloud cover may provide measures of the amount of sunshine received
by phytoplankton within Missisquoi Bay, as possibly fewer cyanobacteria may be present
on cloudy and rainy days when solar radiation is reduced. Rainfall showed few
correlations with cyanobacterial presence, although total cyanobacterial biomass may
have been reduced by rainfall on the same day of sampling, and if there was higher
rainfall across the preceeding 10 days. However the correlation coefficients (Rho) are
weak, all being below -0.39. In comparison, Anabaena spiroides and Microcystis
aeruginosa showed a slight increase in response to rain, especially on the day of
sampling. Increased cloud cover may reduce both total cyanobacterial biomass plus that
of a number of common species, especially if these conditions persist for a week or
more. However again the significant correlations are few and mostly represent weak
relationships between the attributes.
Several genera of eukaryotic algae also showed responses to meteorological conditions,
in particular to the maximum temperature. Several taxa appear to be favoured by hotter
air temperatures whereas Rhodomonas and to a lesser extent Cryptomonas were
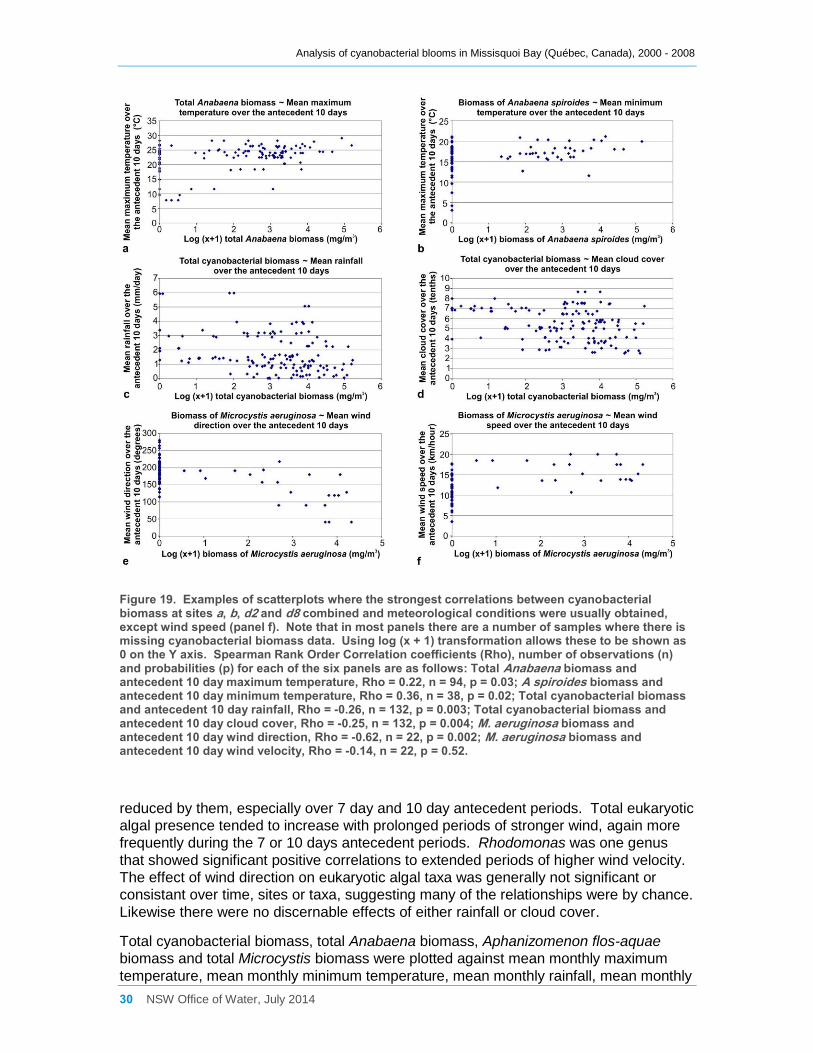
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
30 NSW Office of Water, July 2014
Figure 19. Examples of scatterplots where the strongest correlations between cyanobacterial biomass at sites a, b, d2 and d8 combined and meteorological conditions were usually obtained, except wind speed (panel f). Note that in most panels there are a number of samples where there is missing cyanobacterial biomass data. Using log (x + 1) transformation allows these to be shown as 0 on the Y axis. Spearman Rank Order Correlation coefficients (Rho), number of observations (n) and probabilities (p) for each of the six panels are as follows: Total Anabaena biomass and antecedent 10 day maximum temperature, Rho = 0.22, n = 94, p = 0.03; A spiroides biomass and antecedent 10 day minimum temperature, Rho = 0.36, n = 38, p = 0.02; Total cyanobacterial biomass and antecedent 10 day rainfall, Rho = -0.26, n = 132, p = 0.003; Total cyanobacterial biomass and antecedent 10 day cloud cover, Rho = -0.25, n = 132, p = 0.004; M. aeruginosa biomass and antecedent 10 day wind direction, Rho = -0.62, n = 22, p = 0.002; M. aeruginosa biomass and antecedent 10 day wind velocity, Rho = -0.14, n = 22, p = 0.52.
reduced by them, especially over 7 day and 10 day antecedent periods. Total eukaryotic
algal presence tended to increase with prolonged periods of stronger wind, again more
frequently during the 7 or 10 days antecedent periods. Rhodomonas was one genus
that showed significant positive correlations to extended periods of higher wind velocity.
The effect of wind direction on eukaryotic algal taxa was generally not significant or
consistant over time, sites or taxa, suggesting many of the relationships were by chance.
Likewise there were no discernable effects of either rainfall or cloud cover.
Total cyanobacterial biomass, total Anabaena biomass, Aphanizomenon flos-aquae
biomass and total Microcystis biomass were plotted against mean monthly maximum
temperature, mean monthly minimum temperature, mean monthly rainfall, mean monthly
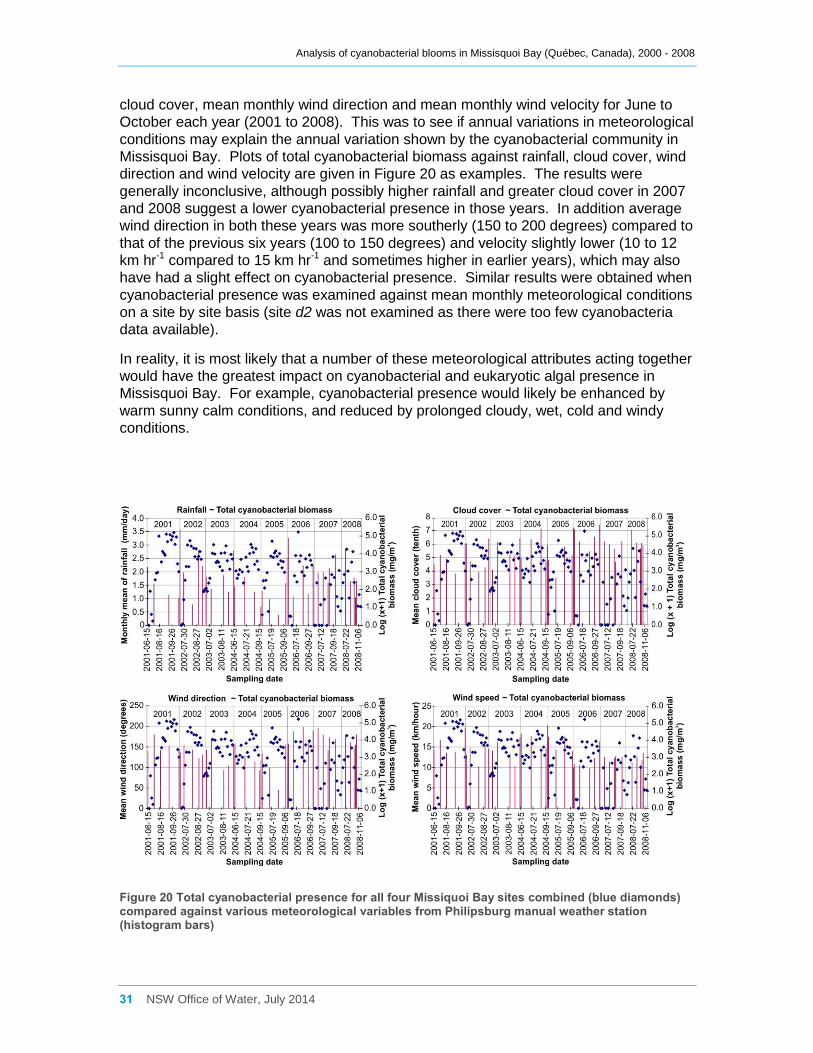
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
31 NSW Office of Water, July 2014
cloud cover, mean monthly wind direction and mean monthly wind velocity for June to
October each year (2001 to 2008). This was to see if annual variations in meteorological
conditions may explain the annual variation shown by the cyanobacterial community in
Missisquoi Bay. Plots of total cyanobacterial biomass against rainfall, cloud cover, wind
direction and wind velocity are given in Figure 20 as examples. The results were
generally inconclusive, although possibly higher rainfall and greater cloud cover in 2007
and 2008 suggest a lower cyanobacterial presence in those years. In addition average
wind direction in both these years was more southerly (150 to 200 degrees) compared to
that of the previous six years (100 to 150 degrees) and velocity slightly lower (10 to 12
km hr-1 compared to 15 km hr-1 and sometimes higher in earlier years), which may also
have had a slight effect on cyanobacterial presence. Similar results were obtained when
cyanobacterial presence was examined against mean monthly meteorological conditions
on a site by site basis (site d2 was not examined as there were too few cyanobacteria
data available).
In reality, it is most likely that a number of these meteorological attributes acting together
would have the greatest impact on cyanobacterial and eukaryotic algal presence in
Missisquoi Bay. For example, cyanobacterial presence would likely be enhanced by
warm sunny calm conditions, and reduced by prolonged cloudy, wet, cold and windy
conditions.
Figure 20 Total cyanobacterial presence for all four Missiquoi Bay sites combined (blue diamonds) compared against various meteorological variables from Philipsburg manual weather station (histogram bars)

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
32 NSW Office of Water, July 2014
3.7 Multivariate analysis of the cyanobacterial and environmental data
3.7.1 Preliminary analysis of the complete cyanobacterial data set
Preliminary analysis was undertaken on the complete set of cyanobacterial data from
sites a, b, d2 and d8 combined, for the years 2000 to 2009 inclusive. Unfortunately the
nMDS ordinations had a high stress value (0.23), meaning that the ordination provides
only a limited representation of the multidimensional data distribution when this is
reduced to 2 dimensional space. There was a large amount of overlap in the ordinations
for sites, months and location (east/west), and any differences were not well shown.
However some differences are evident in the ordination for years, with the years 2001 to
2004 appearing more towards the top left of the ordination, and the years 2005, 2007,
2008 and 2009 appearing more towards the bottom and right side of the ordination. The
outputs of the nMDS are provided in Appendix H (Figure H1).
PERMANOVA analysis revealed significant differences in community composition
between sites, years and months. The overall PERMANOVA results were as follows:-
For sites, Pseudo-F value = 1.95, permutation P (P(perm)) = 0.005, unique
permutations = 9905 out of a total of 9999. Pairwise analysis indicated that
differences in community composition between sites were not always significant.
Sites a and d8 were not significantly different from each, as were sites b and d2 and
sites b and d8. However significant differences were found between sites a and b (t
= 1.43, P(perm) = 0.046, unique permutations = 9934), sites a and d2 (t = 1.77,
P(perm) = 0.005, unique permutations = 9936), and sites d2 and d8 (t = 1.70,
P(perm) = 0.005, unique permutations = 9929).
Location on the eastern or western side of the Bay (sites a and d8 compared to sites
b and d2) was significantly different (Pseudo-F value = 3.24, permutation P (P(perm))
= 0.002, unique permutations = 9943). Globally, the sites from the western and
centre of the bay (sites a and d8) and those from the eastern side (sites b and d2)
were internally more similar while both sides of the bay were different. This may
reflect the effect of winds and other meteorological conditions that may push
cyanobacteria towards the eastern shoreline, although this was not indicated by the
analysis of the meteorological data (Section 3.6, above). It may also reflect nutrient
inputs from major tributaries which may affect community composition.
For years, Pseudo-F value = 9.95, permutation P (P(perm)) = 0.0001, unique
permutations = 9859 out of a total of 9999. Pairwise comparisons between years
indicated significant differences between all years except for 2008 and 2009
(pairwaise PERMANOVA results not shown).
For months, Pseudo-F value = 3.84, permutation P (P(perm)) = 0.0001, unique
permutations = 9708 out of a total of 9999. Pairwise comparisons indicated
significant differences between all months sampled except June and July and June
and October (analysis results not shown). This tends to confirm the relative
unpredictability of cyanobacterial proliferation in Missisquoi Bay and suggests the
complexity of underlying causual factors, at least with the sampling protocols used for
this study.
SIMPER analysis indicated only a low within-sites percent similarity in cyanobacterial
communities (Table H1, Appendix H). Anabaena flos-aquae, Aphanizomenon flos-
aquae, Microcystis sp., Phormidium mucicola and Chroococcus dispersus were the most
common species, but their contributions to the cyanobacterial communities varied from

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
33 NSW Office of Water, July 2014
site to site. Between-site similarities were therefore also generally low, ranging from
23.9% for sites a and d8 to 31.0% for sites b and d2 (data not otherwise shown in
Appendix H).
Part of the within-site variation in cyanobacterial community composition would be due to
the annual variations occurring at each site, as shown by SIMPER (Table H2, Appendix
H). Cyanobacterial communities were most similar throughout 2000 (68.7%) and in
2006 (61.3%). Communities were least similar during 2007 and 2008 (24.1% and
27.0%, respectively). Some of this variation would be due to year-to-year variations in
sampling effort; for example in 2000 only site a was sampled, whereas in other years all
sites were sampled. The cyanobacterial community recorded for 2000 also displayed
the least similarity to the communities of most other years as 2000 was dominated by
small Chroococcales species, whereas different species occurred in most other years
except 2007 when Aphanothece spp. were also present in sufficient biomass. The years
most similar to each other were 2003-04 (41.7% similar), 2002-03 (38.6% similar) and
2003 and 2007 (37.3% similar).
SIMPER analysis also indicated low within-month similarities in most instances (Table
F3, Appendix F). Within-month similarity was greatest in September, at 37.0%. May
and November have been deleted due to only 1 and 3 samples respectively for these
months. Chroococcales (Microcystis sp., Aphanocapsa spp., Chroococcus spp.) were
the taxa contributing most to similarities in June, July and October, whereas it was
Nostocales (Anabaena spp., Aphanizomenon flos-aquae) in August and September.
However a range of other taxa were commonly present most months. The two months
most similar were for August and September at 33.6% similarity, and those least similar
were August and October, at 15.1% similarity.
3.7.2 Analysis of the reduced cyanobacterial data set
nMDS ordinations undertaken on the reduced cyanobacterial data set yielded similar
results to those done on the complete data set – the stress value was also high (0.22)
and the ordinations did not reveal much separation between the samples, especially
based on site and month. Some groupings were more apparent for the ordination based
on years (2001 to 2004 samples being more to the right of the ordination, 2008 and 2009
more to the left) and on location (eastern or western side of Missisquoi Bay) (Figure H2,
Appendix H). These results were consistent with those obtained with the larger data set.
PERMANOVA analysis revealed a significant difference in community composition
between sites, years and months. The overall PERMANOVA results were as follows:-
For sites, Pseudo-F value = 1.63, permutation P (P(perm)) = 0.03, unique
permutations = 9906 out of a total of 9999. However pairwise analysis only indicated
significant differences in community composition between sites a and d2 (t value =
1.61, permutation P (P(perm)) = 0.01, unique permutations = 9952) and between
sites d2 and d8 (t value = 1.79, permutation P (P(perm)) = 0.003, unique
permutations = 9940). There was also a significant difference indicated between the
sites on the eastern side of Missisquoi Bay (sites b, d2) compared to those on the
western side (sites a and d8) (Pseudo-F value = 2.86, permutation P (P(perm)) =
0.006, unique permutations = 9946).
For years, Pseudo-F value = 7.92, permutation P (P(perm)) = 0.0001, unique
permutations = 9851 out of a total of 9999. Pairwise comparisons between years

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
34 NSW Office of Water, July 2014
indicated significant differences between all years except 2008 and 2009 (t value =
0.98, permutation P (P(perm)) = 0.44, unique permutations = 9660).
For months, Pseudo-F value = 3.15, permutation P (P(perm)) = 0.0001, unique
permutations = 9880 out of a total of 9999. Pairwise comparisons indicated
significant differences between all months, although the comparison between July
and October (t value = 1.37, permutation P (P(perm)) = 0.0503, unique permutations
= 9927) was on the significance threshold.
Again, the results of the PERMANOVA analysis of the reduced cyanobacterial data set
were very similar to those undertaken on the complete cyanobacterial data set (see
Section 3.7.1).
SIMPER analysis for the reduced cyanobacterial data set was also similar to that of the
complete cyanobacterial data set. Average within-site similarities were below 30%
except for site d2 where there were fewer samples (39.4% similarity), with the same
species being the major contributors at each site as in the complete data set (Table H1,
Appendix H). Between-site similarities were also generally less than 30%, and the
cyanobacterial community composition at the eastern sites (b and d2) was only 26.8%
similar to the community composition at the western sites (a and d8). Within-year
similarities were also mostly the same as in the larger data set, although with fewer
samples within-year similarities in 2004 and 2005 were greater at 70.2% (7 samples)
and 52.7% (6 samples) respectively (compare with Table H2, Appendix H). Between-
year similarities were also mostly low, at less then 30% similarity (range 45.6% to
10.7%). The most similar cyanobacterial communities occurred in 2003 and 2004,
followed by 2002 and 2003, 2002 and 2004, and 2003 and 2006, while the least similar
years were 2005 and 2007. The reduced data set also showed similarities and species
composition little different to the complete data set. Between month similarities were
also low, with the closest months (35.2% similarity) being August and September.
3.7.3 Analysis of the environmental data set
Principal components analysis (PCA) was undertaken on the environmental data to
determine if differences in environmental conditions occurred in Missisquoi Bay between
sites, years and months. Ordinations showing the first and second principal components
of the PCA are shown in Figure H3, Appendix H. These two principal components
represent 68.4% of the total variation within the data set. The main variables
represented by the first principal component (47.1% of the variation) and their
coefficients were total phosphorus (-0.44), TN:TP (0.36), pH (-0.45), turbidity (-0.46) and
Secchi disc depth (0.44). The main variables represented by the second principal
component (21.3% of the variation) were total nitrogen (-0.72), and the TN:TP ratio (-
0.54).
The ordinations show slight separation between the sites, with samples from sites a and
d8 plotting more to the right side of the ordination, and samples from sites b and d2
located more towards the left of the ordination. This is also better illustrated in the
ordination based on location (east or west side of Missisquoi Bay). Basically sites a and
d8 are distinguished as having lower nutrient concentrations, pH and better water clarity
(increasing Secchi disc depth) compared to sites b and d2. The ordinations show little
variation between years, although possibly 2006 and 2009 may have had better water
clarity. There were few differences in environmental variables indicated by month,
although July would have had slightly lower surface water temperature, but surface
water temperature has little effect on the distribution of samples based on these first two

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
35 NSW Office of Water, July 2014
principal components. September may often have had lower TN concentrations and
TN:TP ratios, but so too may have August and October. This observation suggests that
nitrogen is reduced during late summer and early autumn.
PERMANOVA analysis of the environmental data revealed significant differences in
environmental variables between sites, years and months. The overall PERMANOVA
results were as follows:-
For sites, Pseudo-F value = 5.45, permutation P (P(perm)) = 0.0001, unique
permutations = 9935 out of a total of 9999. However pairwise analysis indicated that
there were no significant differences in environmental variables between sites a and
d8 (t value = 0.99, permutation P (P(perm)) = 0.40, unique permutations = 9946) and
between sites b and d2 (t value = 1.19, permutation P (P(perm)) = 0.22, unique
permutations = 9940). There was also a significant difference between the sites on
the eastern side of Missisquoi Bay (sites b, d2) and on the western side (sites a and
d8) (Pseudo-F value = 13.81, permutation P (P(perm)) = 0.0001, unique
permutations = 9952).
For years, Pseudo-F value = 4.99, permutation P (P(perm)) = 0.0001, unique
permutations = 9874 out of a total of 9999. Pairwise comparisons between years
indicated significant differences between most years, exceptions being 2001 and
2005, 2001 and 2007, 2002 and 2005, 2002 and 2007, 2003 and 2005, and 2003
and 2007. Low sample numbers in some years (and thus a low number of unique
permutations) may have led to the lack of significance in the latter two comparisons.
For months, Pseudo-F value = 4.49, permutation P (P(perm)) = 0.0001, unique
permutations = 9911 out of a total of 9999. Pairwise comparisons between some
months are not reliable due to the low number of samples in these months (June and
November in particular), although significant differences were recorded between July
and August, September and October; between August and October; and between
September and October. Environmental variables were not significantly different
between August and September (t value = 1.63, permutation P (P(perm)) = 0.07,
unique permutations = 9949).
SIMPER analysis indicated highly variable within-site and between-site similarities within
the environmental data set. Different water quality factors appeared to have different
contributions to the environmental conditions at different sites. The main contributing
factors to similarity between samples at each site were surface water temperature at site
a (24%), total nitrogen at site b (27%), turbidity at site d2 (32%) and the TN:TP ratio (by
concentration) at site d8 (34%). In summary, surface water temperature was a factor
contributing to sample similarity at sites a, b and d8; Secchi disc depth and the TN:TP
ratio at sites a and d8; pH at sites a and b; turbidity at sites a, b and d2; TN at sites b, d2
and d8, and TP at sites b and d2. Likewise there was considerable variability in the
contributions of the various environmental factors to within and between year and month
similarity – each year and month varied considerably from the others.
3.7.4 Comparison of the reduced cyanobacterial data set and the environmental data set
The resemblance matrices generated for the cyanobacterial data and the corresponding
environmental data in PRIMER v6 were compared to each other using RELATE. A small
but statistically significant correlation was found between the multivariate data sets
(global statistic Rho = 0.28, P(perm) = 0.001, total permutations = 999). This correlation

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
36 NSW Office of Water, July 2014
is not strong enough to identify water quality as a major determinant of cyanobacterial
community composition. Further analysis using BEST indicated that TN, surface water
temperature and pH were the environmental factors most correlated with the changes in
cyanobacterial community composition. However the data also indicated that there is
considerable other unexplained variance that may be driving cyanobacterial community
composition in Missisquoi Bay.
3.7.5 Comparisons on a site by site basis
A site by site analysis was also undertaken for sites a, b and d8 (there were insufficient
data for site d2), but the results did not differ greatly from those obtained for the
combined sites (a, b, d2, d8) described above, so the analyses are not reported in any
detail here. The environmental data for these analyses also included the meteorological
data, but this was found to have little influence on cyanobacterial community composition
at any of the sites. In brief, the analyses found that both the environmental data and the
cyanobacterial community composition data at each of the three sites displayed
considerable variability in terms of both years and months, as also shown by the
previous analysis of the combined data for all sites. Comparisons of the environmental
data set with the cyanobacterial data set using RELATE for each site demonstrated a
very poor correlation between the two.
4 Discussion
The planktonic cyanobacteria in the northern (Québec) half of Missisquoi Bay have
displayed considerable temporal and spatial diversity during the period 2000 until 2008,
illustrated in particular by Figures 2 to 5 and confirmed by the PERMANOVA and
SIMPER analyses. Generally sites a and d8 on the western side of Missisquoi Bay had
a lower cyanobacterial presence than sites b and d2 on the eastern side of the bay.
Likewise, although some variation in the data may be due to differences in sampling
effort between years, there were nevertheless considerable annual differences.
Cyanobacterial were present at particularly high cell counts and biomass at most sites in
2001, 2002, 2003, 2004 and 2005, while cell counts and biomass were low at all 4 sites
sampled in 2007. Only site a was sampled in 2000, and although this site had high cell
counts, these were predominantly contributed by small Aphanothece clathrata so
biomass that year at that site was low. In 2006 sites a and d2 had considerable
cyanobacterial presence, whereas sites b and d8 had only a low to moderate
cyanobacterial presence. Similarly in 2008, site b recorded a considerably higher
cyanobacterial presence compared to sites a and d8. In most years the peak blooms did
not develop until later in summer, occurring in late July, August and September.
Microcystis was the dominant genus present at most sites in the major bloom years of
2001 to 2005, although the species did vary from year to year. Microcystis sp. was often
recorded as the major taxa present in 2001 and 2003, whereas M. viridus was dominant
at all four sites in 2004, and M. aeruginosa in 2005 and also in 2008. Microcystis has
also been recorded as the major cyanobacterial genus occurring in Missisquoi Bay prior
to 2006 by a number of other authors (Watzin et al., 2006; Mihuc et al., 2006; Boyer,
2008; Wheeler et al., 2012) who sampled mainly the American portion of the bay, but
who may have sometimes also sampled in Québec. Taxa comprising the cyanobacterial
communities in 2006 and 2007 have been more mixed. A later summer bloom of
Aphanizomenon flos-aquae occurred in 2006, whereas Anabaena flos-aquae and A.
spiroides occurred in 2006 and 2007. Correlation analysis indicated that a number of

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
37 NSW Office of Water, July 2014
cyanobacterial species were significantly positively correlated, forming mixed blooms in
Missisquoi Bay together.
Non-toxic species such as Aphanothece clathrata and Chroococcus dispersus were
sometimes dominant at some sites. Fortin et al. (2010) and McQuaid et al. (2011)
reported considerable variation in the cyanobacterial community in the north-east portion
of the bay near Saint-Armand occurring in 2006, 2007, and 2008 and they also reported
low cyanobacterial abundance in 2007. Likewise Davis et al. (2009) found that
Microcystis never numerically dominated at their sampling site in Missisquoi Bay in 2006,
with Anabaena dominating in late summer. Zamyadi et al. (2012) found Microcystis
aeruginosa dominant in the north-east of the bay in 2008, with other cyanobacterial taxa
as subdominants. Levine et al. (2012) reported that there were varying combinations of
Aphanizomenon, Anabaena, Gloeotrichia, Microcystis and Woronichinia occurring in
Missisquoi Bay since 2006, with Aphanizomenon dominating. In addition, Pearce et al.
(2013) have reported that Microcystis has become less dominant in summer blooms in
Missisquoi Bay in recent years, and suggested that the phytoplankton community may
be changing due to processes such as global warming or invasive species introductions.
This spatial and temporal variation in the cyanobacterial communities of Missisquoi Bay
was also highlighted by the multivariate analysis of the community data, with
PERMANOVA in particular showing significant differences between sites, between years
and between months. SIMPER analysis also showed considerable within-site, within-
year and within-month heterogeneity within the cyanobacterial community. Satellite
images of blooms in Missisquoi Bay from 2004 (Wheeler et al., 2012), and cruise data
from 2006 (Mihac et al., 2008) have also illustrated the large temporal and spatial
variability of cyanobacterial blooms and their community composition across the bay.
Cyanobacterial blooms in Missisquoi Bay are generally preceded in spring by eukaryotic
communities dominated by diatoms and cryptophytes, although the biomass of these
communities is usually low. Exponential growth in summer can lead to cyanobacteria at
times contributing to more than 80% of the total phytoplankton biomass present. As the
cyanobacterial blooms decline in late autumn, they are replaced again by diatoms and
cryptophytes. In 2007 when major cyanobacterial blooms did not develop, green algae
(Chlorophyceae) and the dinoflagellate Peridinium became dominant components of the
phytoplankton community. Correlation analysis indicated no significant correlation
between total cyanobacterial biomass and total eukaryotic biomass. Mihuc et al. (2005)
have reported a spring dominance of diatoms in Lake Champlain that were followed by
later summer shifts to Microcystis in the shallow waters of Missisquoi Bay. McQuaid et
al. (2011) reported that diatoms became dominant at end of September 2008.
The cyanotoxin microcystin was recorded in Missisquoi Bay at most sites each year from
2001 when toxin analysis commenced, apart from 2007 when all measurements were
below the level of detection and total cyanobacterial biomass was low. Microcystin-LR
was the most commonly occurring cyanotoxin, but microcystin-YR and microcystin-RR
also frequently co-occurred and increased in unison with microcystin-LR, and
occasionally concentrations of these two microcystin variants even exceeded those of
microcystin-LR. Watzin et al. (2006) and Boyer (2008) also reported that microcystin
was commonly detected in samples from the American side of Missisquoi Bay between
2000 and 2004. Fortin et al. (2010) reported very low microcystin concentrations in
2007, and that microcystin-LA was the predominant microcystin encountered in 2006.
Davis et al. (2009) also reported microcystins present in 2006.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
38 NSW Office of Water, July 2014
Spearman Rank Order Correlation analyses indicated that total microcystin
concentrations in Missisquoi Bay were significantly positively correlated with total
Microcystis biomass, as well as with Microcystis aeruginosa and Microcystis sp.
biomass, as were the separate variants (with the exception of microcystin-YR and
Microcystis sp.). With some exceptions, microcystin concentrations were low when
species other than Microcystis dominated the cyanobacterial community. Fortin et al.
(2010) also found that Microcystis was the principal cyanobacterium producing
microcystin in Missisquoi Bay during their study, although Ngwa et al. (2014) reported a
substantial spatial variability in the proportion of toxic to non-toxic genotypes (2 to 23%)
of Microcystis at Québec sites in Missisquoi Bay in 2009, with non-toxic strains
dominating. Davis et al. (2009) found a significant positive correlation between
microcystin concentrations and densities of toxic Microcystis cells determined by
Polymerase Chain Reaction (PCR). Elsewhere, Rinta-Kanto et al. (2009) have also
found significant positive correlations between microcystin concentrations and
Microcystis abundance determined by PCR in Lake Erie, and Wood et al. (2012) have
found that high cell densities of Microcystis may enhance microcystin production. In
comparison, Watzin et al. (2006) reported that microcystin concentrations in other parts
of Lake Champlain in the USA were not correlated with cell densities, especially late in
the bloom period when cells may die and lyse, releasing the microcystin in soluble form
into the water column. In addition, Microcystis also appears to be able to switch
microcystin production on and off leading to large and rapid (within hours) increases and
decreases in microcystin concentrations in blooms (Wood et al., 2010). Such short-term
fluctuations would also affect correlations found between Microcystis abundance and
microcystin concentrations. Sinang et al. (2013) reported that the temporal variability of
total microcystin concentrations was up to four times greater than the temporal variability
of total cyanobacterial biomass in Western Australian lakes, and that the relationship
between the two was weak and site specific.
For the Québec portion of Missisquoi Bay, anatoxin-a was detected only once in this
study. However Boyer (2008) has reported animal fatalities due to anatoxin-a from
“northern regions of Lake Champlain” (not necessarily Missisquoi Bay), and that peaks in
anatoxin did not coincide with large microcystin-producing cyanobacterial blooms, but
occurred when significant concentrations of Anabaena spp. were present. This species
could be an anatoxin producer. In regards to other cyanotoxins, to our knowledge
saxitoxins and neosaxitoxins have never been measured in the Québec portion of
Missisquoi Bay, although this may perhaps be because appropriate standards to analyse
them have not been available. Zamyadi et al. (2012) did however detect traces of
cylindrospermopsin once in samples collected from near Saint-Armand in 2010.
Microcystin concentrations are likely to vary depending on environmental factors in much
the same way as cyanobacterial blooms vary with the environment, although Sinang et
al. (2013) consider that different environmental factors may influence each. In
Missisquoi Bay, total microcystin concentrations and concentrations of microcystin-LR,
microcystin-RR and microcystin-YR were significantly correlated with a number of
environmental variables (Appendix E). The correlation with chlorophyll-a may be due to
increased toxin concentrations with increased cyanobacterial biomass. Rinta-Kanto et
al. (2009) also found a significant positive correlation between total microcystin
(determined by Protein Phosphatase Inhibition analysis) and chlorophyll-a
concentrations in Lake Erie. Microcystin-RR and microcystin-YR in Missisquoi Bay were
positively correlated with water temperature, although total microcystin concentrations
were not.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
39 NSW Office of Water, July 2014
Davis et al. (2009) found that the maximum cellular toxin content of Microcystis was
achieved at water temperatures of 25 to 30°C, their optimal growth temperature range.
Increasing turbidity and decreasing euphotic depth would result in lower light intensities
experienced by cyanobacteria entrained within the water column, while pH can influence
CO2 availability. Neilan et al. (2013) suggest that differences in light conditions, iron
availability, and CO2 concentrations may influence cyanobacterial blooms at a
community level as variations in the ratio of toxic to non-toxic genotypes may vary
according to these factors. This in turn would affect microcystin production by that
community.
Total microcystin and microcystin-LR and microcystin-YR concentrations in Missisquoi
Bay were also significantly positively correlated with total phosphorus concentrations,
and significantly negatively correlated with the TN:TP ratio. This probably results from
the significant positive correlations between many of the cyanobacterial taxa and total
phosphorus and the significant negative correlations with the TN:TP ratio, including
microcystin producing taxa; that is, increasing cyanobacterial biomass due to these
nutrient conditions was also reflected by higher microcystin concentrations. Rinta-Kanto
et al. (2009) reported similar results for Lake Erie, while Wilhelm et al. (2011) reported
only weak relationships between microcystin concentrations and nutrients in Taihu, a
large lake in eastern China, and no other strong relationships between toxin
concentrations and other environmental variables. Sinang et al. (2013) found that
relationships betwewen microcystin concentrations and total phosphorus varied
considerably from lake to lake and may be site specific. However Neilan et al. (2013)
point out that while eutrophication may stimulate toxic bloom development, it is currently
not known if phosphorus and nitrogen have direct effects on toxin production or if they
regulate the presence toxic and non-toxic genotypes, although they may stimulate
overall bloom community composition.
The very heterogeneous composition of the cyanobacterial community within Missisquoi
Bay, both spatially and temporally, may result partly from the heterogeneous nature of
water quality across the bay (Table 2). Multivariate analysis of the environmental data
using PCA and PERMANOVA also indicated significant spatial and temporal differences
in water quality. In particular sites b and d2 on the eastern side of Missisquoi Bay tend
to have generally higher TP concentrations, pH and turbidity and lower Secchi disc
depths compared to sites a and d8 in the centre and west of the bay. Both sites b and
d2, and site b in particular, are close to where the Pike River (Rivière aux Brochets)
discharges into Missisquoi Bay. The Pike River catchment is intensely agricultural with
considerable increases in the area of land cultivated for corn over the past two decades,
and it subsequently has high annual phosphorus and nitrogen yields and P and N
concentrations (Medalie et al., 2012). Moreover, the major source of nutrients in
Missisquoi Bay is the Missisquoi River (Smeltzer and Simoneau, 2008). A hydraulic
simulation confirmed by the results of a sampling program designed as a follow-up study
of the use of a lampricide in this river have shown that it discharges from its outlet in the
south of the bay towards Saint-Armand (Crowley et al., 2009). The nutrients contributed
by these rivers may partly account for the higher abundance of cyanobacteria measured
at these eastern sites (b and d2) during much of the 2001 – 2009 monitoring period. The
heterogeneous distribution of cyanobacterial blooms, including their community
composition, has been noted previously by Mihuc et al. (2008), and by Wheeler et al.
(2012).
Overall, Missisquoi Bay is eutrophic in terms of total phosphorus concentrations (Table
2), as has been described by several other authors (McQuaid et al., 2011; Zamyadi et

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
40 NSW Office of Water, July 2014
al., 2012; Levine et al., 2012; Smeltzer et al., 2012). McCarthy et al. (2013) consider
that nutrients probably do not limit primary production in the bay, although Davis et al.
(2009) reported that increased nitrogen stimulated increased growth rate of toxic
Microcystis at their sampling locations. Spearman Rank Order Correlation analysis
indicated that the biomass of many taxa of cyanobacteria, as well as total cyanobacterial
biomass were significantly positively correlated with total phosphorus concentrations,
both overall across all four sites (a, b, d2, d8) and often on a site by site basis, especially
at sites b and d2.
In comparison, total nitrogen (TN) concentrations recorded for the four sites were low,
even at site b (Table 3). The general trend is for TN and oxidised nitrogen (NOx)
concentrations to decrease over the summer period (June to August), then increase
once more in September, while TP concentrations show an opposite trend (data not
shown). McCarthy et al. (2013) also reported this trend with nitrogen. Spearman Rank
Order Correlation analysis showed few significant correlations between cyanobacterial
taxa and total nitrogen concentrations when data from all sites were combined, although
a number of taxa were significantly negatively correlated with NOx (Table F1, Appendix
F). On a site to site basis, there were significant negative correlations between the
biomass of several cyanobacterial taxa and NOx at sites b and d2 (Table F2, Appendix
F). This indicates the possible uptake of NOx by the cyanobacteria. The TN:TP ratios
(by concentration) were frequently less than 7.0 and are also indicative of possible
nitrogen limitation (or phosphorus excess). Decreasing nitrogen concentrations occurred
when heterocystis cyanobacterial species such as Anabaena spp. and Aphanizomenon
flos-aquae, which can overcome nitrogen limitation by fixing atmospheric nitrogen, were
present. However non-nitrogen fixing species such as Microcystis spp. were at times
also present when nitrogen concentrations were low, especially at site b. McCarthy et al.
(2013) reported few heterocytes being present in heterocystis species of Nostocales in
Missisquoi Bay, suggesting that cyanobacterial fixation of atmospheric nitrogen does not
contribute significantly to the overall nitrogen budget of the bay.
Total nitrogen concentrations in the Pike River (and in the Missisquoi River) appear to be
much higher (Medalie et al., 2012) than those in Missisquoi Bay (Table 2), even at sites
close to their inflow points, and McCarthy et al. (2013) have reported a similar spatial
variation. This suggests a rapid loss of nitrogen once inflows enter Missisquoi Bay in
summer, either by sedimentation or by denitrification. Denitrification is microbially
mediated and occurs more rapidly at warmer summer temperatures McCarthy et al.
(2013) also found that although inputs from tributaries contributed to the nitrogen
available for primary production within Missisquoi Bay, regeneration of autochthonous
organic material within the lake was a more important source of nitrogen for primary
productivity. Nevertheless, McCarthy et al. (2013) considered that neither tributary
inputs nor within-lake regeneration were sufficient to account for all the nitrogen demand
and therefore primary producers must supplement their nitrogen requirements from other
sources, including from the sediments. Pearce et al. (2013) report brief periods of
stratification and anoxic layers overlying the sediment-water interface that would allow
the diffusion of ammonium from the sediments that could be utilised by cyanobacteria.
Spearman Rank Order Correlation analysis also indicated some relationships between
cyanobacterial biomass and several other environmental attributes, such as dissolved
organic carbon (DOC), pH, turbidity and Secchi disc depth. Increasing DOC with
increasing cyanobacterial presence could be due to the production of soluble
metabolites by the cyanobacteria that are then released into the water. For example,
microcystins may be released into the water column from cells, especially if they die and

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
41 NSW Office of Water, July 2014
the cells lyse (Watzin et al., 2006). Correlations with pH, turbidity and Secchi disc depth
may be due to effects of the cyanobacteria themselves. Photosynthesis by
cyanobacteria during bloom periods removes most soluble CO2 from the water column
during daylight hours and results in an increase in pH (Paerl and Paul, 2012). The
presence of large amounts of cyanobacterial biomass can substantially increase the
turbidity of the water column, and at the same time reduce Secchi disc depth. These
interactions would lead to significant correlations between cyanobacteria and these
environmental attributes – positive in the case of pH and turbidity, and negative in the
case of Secch disc depth. These interactions also occurred at all four sites across the
bay, regardless of between site differences in these water quality variables.
Several taxa of cyanobacteria, especially Anabaena spp., were also positively correlated
with surface water temperature. Cyanobacteria are generally considered to grow better
at higher temperatures, especially above 25ºC (Elliott, 2012), and Davis et al. (2009)
have noted that blooms at their study sites on Missisquoi Bay corresponded to a
maximum water temperature of 26ºC. Likewise Mihuc et al. (2008) considered that high
abundance of cyanobacteria would only occur in Missisquoi Bay once temperature
thresholds of 25.5ºC were exceeded in late July, or thresholds of 21.5ºC were exceeded
in late August. Higher summer water temperatures (>25ºC) may support dominance by
Microcystis species as these have higher optimal growth rate temperatures (Carey et al.,
2012) than Anabaena or Aphanizomenon species, which may be an underlying factor for
some of the seasonal changes in cyanobacterial community composition observed in
this study and by others (Davis et al., 2009; Fortin et al., 2010).
There were few significant correlations between cyanobacterial presence and the
meteorological data from Philipsburg, although warm calm sunny conditions are
expected to enhance cyanobacterial presence, while cloudy wet cold and windy
conditions may reduce cyanobacterial presence. Higher maximum and minimum air
temperatures were sometimes correlated with a number of cyanobacterial taxa , and
especially when the 4, 7 or 10 day antecedent period was considered, indicating that a
week or more of hotter weather may benefit cyanobacteria in Missisquoi Bay in summer.
Wind direction was rarely correlated with cyanobacterial presence, and wind conditions
on the day of sampling had little effect on cyanobacterial distribution. Longer periods of
strong winds (10 days) may reduce Anabaena presence. Prolonged rain and cloud
cover may also reduce cyanobacterial presence, but this was not demonstrated
conclusively in this study. Investigations of the effects of annual variations in
meteorological conditions on cyanobacterial presence using site by site multivariate
analysis were also generally inconclusive, although possibly higher rainfall, increased
cloud cover and more southerly winds may have reduced cyanobacterial presence in
2007 and 2008. Fortin et al. (2010) considered that the predominant south-west winds
may lead to onshore accumulations of cyanobacterial scum near Saint-Armand, and also
suggested that rainfall may play a role in cyanobacterial bloom dynamics in the bay.
McQuaid et al. (2011) reported maximum cyanobacterial abundance during calm
conditions, while Wheeler et al. (2012) suggested that scums may form along the
eastern shoreline of the bay, but cyanobacteria would be mixed and dispersed down the
water column by wind on the western side. Mihuc et al. (2008) also suggested that wind
patterns and water currents would influence cyanobacterial bloom distribution around
Missisquoi Bay.
The multivariate analysis of the data tended to differ somewhat from the Spearman Rank
Order Correlation analysis. The multivariate analyses once again illustrate the
considerable heterogeneity in cyanobacterial bloom distribution and community

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
42 NSW Office of Water, July 2014
composition within Missisquoi Bay, both on a spatial and annual basis, and also on a
seasonal basis. The multivariate analysis of all four sites combined found a significant
positive correlation between the cyanobacterial community data and the environmental
data. Although total nitrogen concentrations, pH and surface water temperature were
suggested as the water quality attributes having the most effect on cyanobacterial
community composition, the relationships were not strong enough to identify these as
being the major drivers. Likewise total phosphorus was not indicated as a major driver
of cyanobacetrial presence, and there was a lot of unexplained variance as well. Site by
site analysis also displayed the heterogeneous temporal nature of much of the data, with
no significant correlations between the environmental data and the cyanobacterial
community data indicated for sites a and d8. Significant correlations between the
environmental and cyanobacterial data were only found for site b, where total nitrogen
concentrations, the TN:TP ratio (by concentration) and surface water temperature were
the environmental variables most correlated with the cyanobacterial community data.
Any relationships between cyanobacterial biomass and pigment concentrations in
Missisquoi Bay were also examined using Spearman Rank Order Correlation analysis.
Total cyanobacterial biomass and the biomass of most of the individual taxa or
groupings of cyanobacteria were significantly positively correlated with chlorophyll-a, and
also many with phycocyanin. This indicates that these two pigments are good indicators
of cyanobacterial presence within Missisquoi Bay in summer when cyanobacteria
dominate the phytoplankton, and especially of total cyanobacterial biomass. McQuaid et
al. (2011) has also found a significant positive relationship between phycocyanin
fluorescence measured in-situ in Missisquoi Bay and cyanobacterial biovolume, as have
Brient et al. (2008) and Bowling et al. (2013) elsewhere in the world.
Cyanobacterial blooms in Missisquoi Bay tend to occur in summer, and dominate the
phytoplankton community, contributing more than 70% of the total phytoplankton
biomass at times in July, August and September most years, except in 2000 and 2007.
In contrast, eukaryotic algae, especially diatoms and Cryptophyceae, tend to dominate
the phytoplankton community prior to the onset of the cyanobacterial blooms in spring,
and following the decline of the blooms in autumn, albeit at much lower total biomass.
Chlorophyceae and dinoflagellates also occur, but less frequently than the diatoms and
Cryptophyceae. Similar successional patterns have been reported in Missisquoi Bay by
Mihuc et al. (2005, 2008) and Levine et al. (2012), especially spring diatom abundance.
Spearman Rank Order Correlation analysis of major eukaryotic algal species with
environmental data indicated some significant correlations, but generally fewer than the
corresponding analyses with the cyanobacterial taxa. Significant correlations occurred
between several eukaryotic algal species and total phosphorus and especially with pH,
and surface water temperature, with the eukaryotic algae and cyanobacteria generally
showing similar responses to environmental cues in Missisquoi Bay. Carey et al. (2012)
have described the eco-physiological adaptations that cyanobacteria possess that may
give them a competative advantage over eukaryotic phytoplankton at warmer water
temperatures, and possibly such factors are supporting cyanobacterial abundance and
dominance during summer in Missisquoi Bay. There was usually a marked seasonal
succession pattern within the phytoplankton community each year with a spring
dominance of diatoms and a summer dominance of the cyanobacteria.
The multivariate analysis of the environmental and cyanobacterial data revealed only
weak linkages between the two data sets, and suggested that there were many

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
43 NSW Office of Water, July 2014
unexplained factors causing the heterogeneity shown by the cyanobacterial communities
in Missisquoi Bay. Several other environmental and biological factors not measured in
our study could interact to generate further variation in the cyanobacterial communities.
For example, increased concentrations of agricultural herbicides such as atrazine have
been proposed as one possible cause by Zananski et al. (2010), as this may kill off some
diatom species, but these authors were unable to conclude that cyanobacteria were
more resistant to herbicides than eukaryotic groups of phytoplankton and thus have a
competitive advantage. One biotic factor that has been proposed to have had major
impacts on the phytoplankton community and cyanobacterial presence is the invasion of
exotic species into Lake Champlain such as the zebra mussel (Dreissena polymorpha),
first reported from the lake in 1996 (Mihuc et al., 2005; 2006; 2012). Selective filtering
by zebra mussels is thought to remove many species of phytoplankton, while the larger
colonial forming Microcystis are not ingested, leading to their dominance (Mihuc et al.
2005; 2006). Smeltzer et al. (2012) however point out that Missisquoi Bay hosts a
relatively small population of zebra mussels, so their impacts on the phytoplankton
community may be less severe here than in other parts of Lake Champlain. An exotic
zooplanktovorious fish, the alewife (Alosa pseudoharengus) was also first discovered in
Missisquoi Bay in 2003 (Mihuc et al., 2012). Alewifes are size selective predators of
zooplankton in Lake Champlain, removing the larger sized species. Loss of the larger
zooplankton would reduce grazing pressure on phytoplankton, including cyanobacteria,
and could potentially lead to enhanced blooms (Mihuc et al., 2012), including in
Missisquoi Bay where nutrient concentrations and other environmental factors are
already favouring cyanobacterial blooms.
5 Conclusions
Analysis of the long term (2000 to 2008) phytoplankton and environmental data for
Missisquoi Bay collected by the MDDEFP has highlighted the following:-
Cyanobacterial blooms within the bay are diverse and highly variable in terms of both
abundance and composition, both between sites, within sites and between years and
seasons.
This heterogeneity in cyanobacterial bloom occurrence is apparent despite some
limitations encountered with the data, in particular the different sampling effort
maintained at different sampling sites and in different years.
Major cyanobacterial blooms occurred in Missisquoi Bay from 2001 until 2006, and
again in 2008. Microcystis was the dominant genus present in the bay prior to 2006,
with more mixed cyanobacterial communities occurring from 2006 onwards that
included both Aphanizomenon flos-aquae and Anabaena flos-aquae.
A general successional pattern was apparent in the phytoplankton community, with
diatoms and cryptophytes occurring in cooler periods in spring and autumn, and
cyanobacterial blooms confined to the warmer summer periods.
The cyanotoxin microcystin was recorded most years, with microcystin-LR as the
most common variant present compared to the two other variants analysed
(microcystin-RR and microcystin-YR). Because only three of the many variants of
microcystin were analysed, the data may underestimate total microcystin
concentrations as demonstrated by results for other variants measured in the bay
since 2009 (data not shown). Nevertheless microcystin concentrations were highly

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
44 NSW Office of Water, July 2014
correlated with Microcystis biomass, and were only present occasionally when other
cyanobacterial genera dominated. Factors that have been associated with increased
microcystin production include higher surface water temperature, and increased total
phosphorus, pH and turbidity (reduced light).
Principal Components Analysis (PCA) of the environmental data indicates that water
quality varies considerably at different locations across the bay, especially total
phosphorus, turbidity and pH which are higher on the eastern side compared to the
western side, and shallower Secchi disc depths also occur in the east. This
heterogeneous water quality distribution may be one cause of the high variability in
the cyanobacterial community composition and abundance.
Overall, Missisquoi Bay is eutrophic, and total phosphorus concentrations are a
major factor behind the cyanobacterial blooms.
Total nitrogen concentrations are often not high, and tend to decrease during
summer. Loss by sedimentation or bacteriologically mediated denitrification could be
possible causes. Uptake of soluble oxidized nitrogen by cyanobacteria could also be
occurring.
Cyanobacterial blooms also tend to occur at low TN:TP ratios, suggesting nitrogen
limitation, or excess phosphorus, during bloom periods. Input of nitrogen from
catchment sources would contribute to bloom development, despite low ambient
concentrations in the lake. In-situ nitrogen regeneration and inputs from other
sources have also been suggested in the literature as important sources of nitrogen.
The cyanobacteria themselves change some environmental conditions within the
lake, including increasing pH through use of carbon dioxide during photosynthesis.
Likewise the presence of their cells and colonies increases turbidity and reduces
Secchi disc depth. Their release of metabolites such as extracellular toxins may
increase DOC concentrations, although DOC increases may also be due to other
bacterial activity.
Spearman Rank Order Correlation analysis found few significant correlations with
meteorological factors, although 7 to 10 day periods of hotter weather may benefit
cyanobacteria in Missisquoi Bay in summer.
Spearman Rank Order Correlation analysis found cyanobacterial biomass was
strongly positively correlated with pigment concentrations (both chlorophyll-a and
phycocyanin), and these may be good indicators of cyanobacterial abundance in
summer. Phycocyanin in particular is asssociated with cyanobacteria, while high
chlorophyll-a concentrations can indicate blooms of both cyanobacteria and
eukaryotic algae.
Multivariate analysis of the cyanobacterial and environmental data (RELATE, BEST)
indicated no strong relationships between the cyanobacterial community composition
and environmental attributes. It showed that total nitrogen, pH and surface water
temperature were the factors most associated with cyanobacterial abundance and
community composition. The analysis also indicated a lot of unexplained variance in
these data.
Other environmental and biological factors for which there were no data for analysis
for this report, including herbicides, zebra mussels and the alewife, have been
suggested by other authors as possible factors influencing cyanobacterial community
composition in Missisquoi Bay and elsewhere in Lake Champlain.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
45 NSW Office of Water, July 2014
6 References
Anderson, M.J., Gorley, R.N., and Clarke, K.R. (2008). PERMANOVA+ for PRIMER: Guide to
software and statistical methods. (PRIMER-E, Plymouth, UK.)
Bartram, J., Burch, M., Falconer, I.R., Jones, G. and Kuiper-Goodman, T. (1999). Situation
assessment, planning and management. pp 179–209 in Chorus, I. and Bartram, J.
(eds) Toxic Cyanobacteria in water. A guide to their public health consequences,
monitoring and management. (E & FN Spon, London).
Bastien, C. (2012). 17 May. Validation d'informations méthodologiques - Rapport baie Missisquoi
2000-2009, ([email protected]) [Email]
Bérubé, G. (1981). Aucun moyen de s’en débarrasser à court terme. Des algues microscopiques
dans la baie Missisquoi! Le Canada Français, Saint-Jean. (26 août).
Blais, S. (2014). État de situation sur les cyanobactéries et les cyanotoxines à l’égard de la
protection des usages à la baie Missisquoi de 2000 à 2008. Québec. Ministère du
Développement durable, de l’Environnement et des Parcs.
Blais, S. and Sinotte, M. (2008). Fiche: Cyanobactéries à potentiel toxique. (Document non
publié). Québec. Ministère du Développement durable, de l’Environnement et des
Parcs. 14 pp.
Brault, N. (2012) 27 and 28 September and 7 November. Liste des avis de santé publique (ou de
fermeture de plage) à la baie Missisquoi de 2000 à 2009?
([email protected]) [Emails] ([email protected]).
Brient, L., Lengronne, M., Bertrand, E., Rolland, D., Sipel, A., Steinmann, D., Baudin, I., Legeas,
M., Le Rouzic, B. and Bormans, M. (2008). A phycocyanin probe as a tool for
monitoring cyanobacteria in freshwater bodies. Journal of Environmental Monitoring
10, 248–255.
Bowling, L., Ryan, D., Holliday, J. and Honeyman, G. (2013). Evaluation of in-situ fluorometry to
determine cyanobacterial abundance in the Murray and Darling Rivers, Australia. River
Research and Applications 29, 1059–1071.
Boyer, G.L. (2008). Cyanobacterial toxins in New York and the lower Great Lakes ecosystems. In
Hudnell, H.K. (ed.) Proceedings of the Interagency International Symposium on
cyanobacterial harmful algal blooms. Advances in Experimental Medicine and Biology
619, 153–165.
Carey, C.C., Ibelings, B.W., Hoffmann, E.P., Hamilton, D.P. and Brookes, J.D. (2012). Eco-
physiological adaptations that favour freshwater cyanobacteria in a changing climate.
Water Research 46, 1394–1407.
Clarke, K.R. and Gorley, R.N. (2006). PRIMER v6: user manual/tutorial. (PRIMER-E Ltd.
Plymouth Marine Laboratory, Plymouth, United Kingdom).
Cole, G.A. (1975). Textbook of limnology. (The C.V. Mosby Company, Saint-Louis)..
Crowley, D., Mendelsohn, D. and Swanson, C. (2009). Lampricide hindcast study in Missisquoi
Bay. Report to Vermont Fish and Wildlife Department, October 2009. 63 pp.
Dassylva, N. (2012). 31 May and 5 June. Validation d'informations méthodologiques - Rapport
baie Missisquoi 2000-2009, ([email protected] ) [Email]
Davis, T.W., Berry, D.L., Boyer, G.L. and Gobler, C.J. (2009). The effects of temperature and
nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis
during cyanobacterial blooms. Harmful Algae 8, 715–725.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
46 NSW Office of Water, July 2014
Deblois, C. (2012,a). 28 May. Validation d'informations méthodologiques - Rapport baie
Missisquoi 2000-2009, ([email protected] ) [Email]
Deblois, C. (2012,b). 11 November. Comment vous citer dans l'équipe de réalisation? / rapport
Baie Missisquoi, ([email protected] ) [Email]
Elliott, J.A. (2012). Is the future blue-green? A review of the current model predictions of how
climate change could affect pelagic freshwater cyanobacteria. Water Research 46,
1364–1371.
Fontaine, L., Jolicoeur, G., Desjardins, R., Paquette, P.-É. and Dubé, M. H. (1968). Étude de
l’état des eaux de la baie Missisquoi (Lac Champlain) – Comté de Missisquoi. Régie
des eaux du Québec. 60 pp.
Fortin, N., Aranda-Rodriguez, R., Jing, H., Pick, F., Bird, D. and Greer, C.W. (2010). Detection of
microcystin-producing cyanobacteria in Missisquoi Bay, Quebec, Canada, using
quantitative PCR. Applied and Environmental Microbiology 76, 5105–5112.
Germain, A., Lavoie, F., and Janson, M. (1987). Étude sur l’eutrophisation de la baie Missisquoi.
Québec. Environnement Canada, Conservation et Protection, Direction des eaux
intérieures et des terres, Région de Québec. 97 pp.
Heussner, A.H., Mazija, L., Fastner, J. and Dietrich, D.R. (2012). Toxin content and cytotoxicity of
algal dietary supplements. Toxicology and Applied Pharmacology 265, 263–271.
Holdren, C., Jones, W. and Taggart, J. (2001). Managing Lakes and Reservoirs. Madison, North
American Lake Management Society and Terrene Institute in cooperation with Office of
Water, Assessment and Watershed Protection Division, U.S. Environmental Protection
Agency. 382 pp.
Komárek, J., Kaštovský, J. and Jezberová, J. (2011). Phylogenetic and taxonomic delimitation of
the cyanobacterial genus Aphanothece and description of Anathece gen. nov.
European Journal of Phycology 46, 315–326.
Levine, S.N., Lini, A., Ostrofsky, M.L., Bunting, L., Burgess, H., Leavitt, P.R., Reuter, D., Lami, A.,
Guilizzoni, P., and Gilles, E. (2012). The eutrophication of Lake Champlain’s
northeastern arm: Insights from paleolimnological analyses. Journal of Great Lakes
Research 38, 35–48.
McCarthy, M.J., Gardner, W.S., Lehmann, M.F. and Bird, D.F. (2013). Implications of water
column ammonia uptake and regeneration for the nitrogen budget in temperate,
eutrophic Missisquoi Bay, Lake Champlain (Canada/USA). Hydrobiologia 718, 173-
188.
McQuaid, N., Zamyadi, A., Prévost, M., Bird, D.F. and Dorner, S. (2011). Use of in-vivo
phycocyanin fluorescence to monitor potential microcystin-producing cyanobacterial
biovolume in a drinking water source. Journal of Environmental Monitoring 13, 455–
463.
Medalie, L., Hirsch, R.M. and Archfield, S.A. (2012). Use of flow-normalisation to evaluate
nutrient concentration and flux changes in Lake Champlain tributaries, 1990-2009.
Journal of Great Lakes Research 38, 58–67.
Mihuc, T.B., Boyer, G., Satchwell, M.F., Pellam, M., Jones, J., Vasile, J., Bouchard, A. and
Bonham, R. (2005). 2002 phytoplankton community composition and cyanobacterial
toxins in Lake Champlain, USA. Verhandlungen der Internationale Vereinigung fur
Theoretische und Angewandte Limnologie 29, 328–333.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
47 NSW Office of Water, July 2014
Mihuc, T.B., Boyer, G.L., Jones, J., Satchwell, M.F, and Watzin, M.C. (2006). Lake Champlain
phytoplankton and algal toxins: past and present. Great Lakes Research Review 7, 18–
21.
Mihuc, T.B., Pershyn, C., Thomas, S., Boyer, G., Satchwell, M., Jones, J., Allen, E. and Greene,
M. (2008). Cyanobacteria in the sixth Great Lake: geospatial pigment mapping and
phytoplankton community composition in Missisquoi Bay, Lake Champlain.
Verhandlungen der Internationale Vereinigung fur Theoretische und Angewandte
Limnologie 30, 312–317.
Mihuc, T.B., Dunlap, F., Binggeli, C., Myers, L., Pershyn, C., Groves, A. and Waring, A. (2012).
Long-term patterns in Lake Champlain’s zooplankton: 1992-2010. Journal of Great
Lakes Research 38, 49–57.
Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs (2012).
CLIMATOLOGIE, Direction du suivi de l’état de l’environnement, Québec.
Neilan, B.A., Pearson, L.A., Muenchhoff, J., Moffitt, M.C. and Dittmann, E. (2013). Environmental
conditions that influence toxin biosynthesis in cyanobacteria. Environmental
Microbiology 15, 1239-1253.
Ngwa, F., Madramootoo, C. and Jabaji, S. (2014). Monitoring toxigenic Microcystis strains in the
Missisquoi Bay, Quebec, by PCR targeting multiple gene loci. Environmental
Toxicology 29, 440-451.
Paerl, H.W. and Paul, V.J. (2012). Climate change: Links to global expansion of harmful
cyanobacteria. Water Research 46, 1349–1363.
Pearce, A.R., Rizzo, D.M., Watzin, M.C. and Druschel, G.K. (2013). Unraveling associations
between cyanobacteria blooms and in-lake environmental conditions in Missisquoi Bay,
Lake Champlain, USA, using a modified self-organizing map. Environmental Science
and Technology 47, 14267-14274.
Rinta-Kanto, J.M., Konopko, E.A., DeBruyn, J.M., Bourbonniere, R.A., Boyer, G.L. and Wilhem,
S.W. (2009). Lake Erie Microcystis: Relationship between microcystin production,
dynamics of genotypes and environmental parameters in a large lake. Harmful Algae 8,
665–673.
Shapiro, J. (1990). Current beliefs regarding dominance by blue-greens: the case for the
importance of CO2 and pH. Verhandlungen Internationale Vereinigung für
Theoretische und Angewandte Limnologie 24, 38–54.
Sinang, S.C., Reichwaldt, E.S. and Ghadouani, A. (2013). Spatial and temporal variability in the
relationship between cyanobacterial biomass and microcystins. Environmental
Monitoring and Assessment 185, 6379–6395.
Sivonen, K. and Jones, G. (1999). Cyanobacterial toxins. pp 41–111 in Chorus, I. and Bartram, J.
(eds) Toxic Cyanobacteria in water. A guide to their public health consequences,
monitoring and management. (E & FN Spon, London).
Smeltzer, E., Shambaugh, A.D. and Stangel, P. (2012). Environmental change in Lake
Champlain revealed by long-term monitoring. Journal of Great Lakes Research 38, 6–
18.
Smeltzer, E. and Simoneau, M. (2008). Phosphorus loading to Missisquoi Bay from sub-basins in
Vermont and Québec, 2002-2005. Report submitted to the Lake Champlain Steering
Committee, November 25, 2008. 22 pp.
Wacklin, P., Hoffmann, L. and Komárek, J. (2009). Nomenclatural validation of the genetically
revised cyanobacterial genus Dolichospermum (Ralfs ex Bornet et Flahault) comb.
Nova. Fottea 9, 59–64.

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
48 NSW Office of Water, July 2014
Watzin, M.C., Miller, E.B., Shambaugh, A.D. and Kreider, M.A. (2006). Application of the WHO
Alert Level Framework to cyanobacterial monitoring of Lake Champlain, Vermont.
Environmental Toxicology 21, 278–288.
Wheeler, S.M., Morrissey, L.A., Levine, S.N., Livingston, G.P. and Vincent, W.F. (2012). Mapping
cyanobacterial blooms in Lake Champlain’s Missisquoi Bay using Quickbird and
MERIS satellite data. Journal of Great Lakes Research 38, 68–75.
Wilhelm, S.W., Farnsley, S.E., LeCleir, G.R., Layton, A.C., Satchwell, M.F., DeBruyn, J.M.,
Boyer, G.L., Zhu, G. and Paerl, H.W. (2011). The relationships between nutrients,
cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China. Harmful
Algae 10, 207-215.
Wood, S.A., Rueckert, A., Hamilton, D.P., Cary, S.C. and Dietrich, D.R. (2010). Switching toxin
production on and off: intermittent microcystin synthesis in a Microcystis bloom.
Environmental Microbiology Reports 3, 118-124.
Wood, S.A., Dietrich, D.R., Cary, S.C. and Hamilton, D.P. (2012). Increasing Microcystis cell
density enhances microcystin synthesis: a mesocosm study. Inland Water 2, 17-22.
Zamyadi, A., MacLeod, S.L., Fan, Y., McQuaid, N., Dorner, S., Sauvé, S. and Prévost, M. (2012).
Toxic cyanobacterial breakthrough and accumulation in a drinking water plant: A
monitoring and treatment challenge. Water Research 46, 1511–1523.
Zananski, T.J., Twiss, M.R. and Mihuc, T.B. (2010). Use of fluorimetry to evaluate atrazine
toxicity to phytoplankton communities. Aquatic Ecosystem Health and Management 13,
56–65.
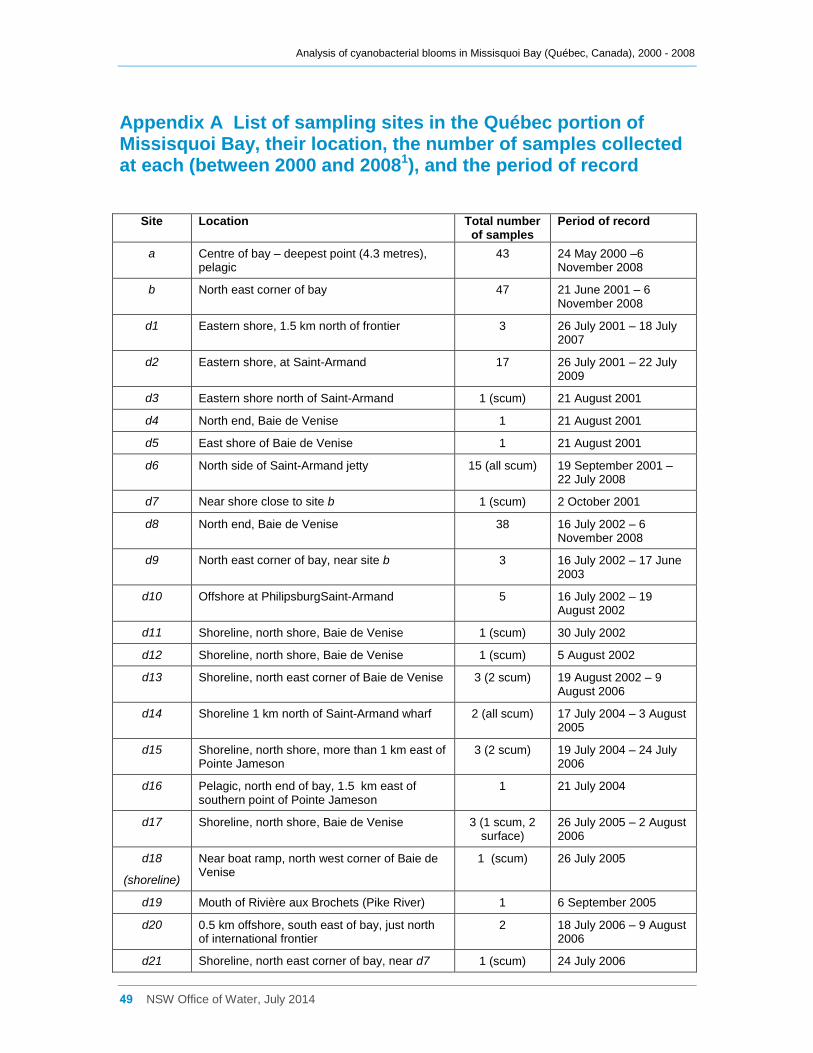
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
49 NSW Office of Water, July 2014
Appendix A List of sampling sites in the Québec portion of Missisquoi Bay, their location, the number of samples collected at each (between 2000 and 20081), and the period of record
Site Location Total number of samples
Period of record
a Centre of bay – deepest point (4.3 metres), pelagic
43 24 May 2000 –6 November 2008
b North east corner of bay 47 21 June 2001 – 6 November 2008
d1 Eastern shore, 1.5 km north of frontier 3 26 July 2001 – 18 July 2007
d2 Eastern shore, at Saint-Armand 17 26 July 2001 – 22 July 2009
d3 Eastern shore north of Saint-Armand 1 (scum) 21 August 2001
d4 North end, Baie de Venise 1 21 August 2001
d5 East shore of Baie de Venise 1 21 August 2001
d6 North side of Saint-Armand jetty 15 (all scum) 19 September 2001 – 22 July 2008
d7 Near shore close to site b 1 (scum) 2 October 2001
d8 North end, Baie de Venise 38 16 July 2002 – 6 November 2008
d9 North east corner of bay, near site b 3 16 July 2002 – 17 June 2003
d10 Offshore at PhilipsburgSaint-Armand 5 16 July 2002 – 19 August 2002
d11 Shoreline, north shore, Baie de Venise 1 (scum) 30 July 2002
d12 Shoreline, north shore, Baie de Venise 1 (scum) 5 August 2002
d13 Shoreline, north east corner of Baie de Venise 3 (2 scum) 19 August 2002 – 9 August 2006
d14 Shoreline 1 km north of Saint-Armand wharf 2 (all scum) 17 July 2004 – 3 August 2005
d15 Shoreline, north shore, more than 1 km east of Pointe Jameson
3 (2 scum) 19 July 2004 – 24 July 2006
d16 Pelagic, north end of bay, 1.5 km east of southern point of Pointe Jameson
1 21 July 2004
d17 Shoreline, north shore, Baie de Venise 3 (1 scum, 2 surface)
26 July 2005 – 2 August 2006
d18
(shoreline)
Near boat ramp, north west corner of Baie de Venise
1 (scum) 26 July 2005
d19 Mouth of Rivière aux Brochets (Pike River) 1 6 September 2005
d20 0.5 km offshore, south east of bay, just north of international frontier
2 18 July 2006 – 9 August 2006
d21 Shoreline, north east corner of bay, near d7 1 (scum) 24 July 2006

Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
50 NSW Office of Water, July 2014
d22 North west shoreline (south west corner of Baie de Venise)
1 (surface) 24 July 2006
d23 Shoreline, north west corner of Baie de Venise 1 (scum) 27 September 2006
d24 Shoreline, north west corner of Baie de Venise 1 (scum) 27 September 2006
d25 Pelagic, north end of bay, 0.5 km east of southern point of Pointe Jameson
1 (surface) 12 July 2007
1 4 samples from site a, 4 samples from site b, 1 sample from site d2, and 3 samples from site d8 collected
in 2009 were also included in some of the statistical analyses. This is indicated in the text where
appropriate.
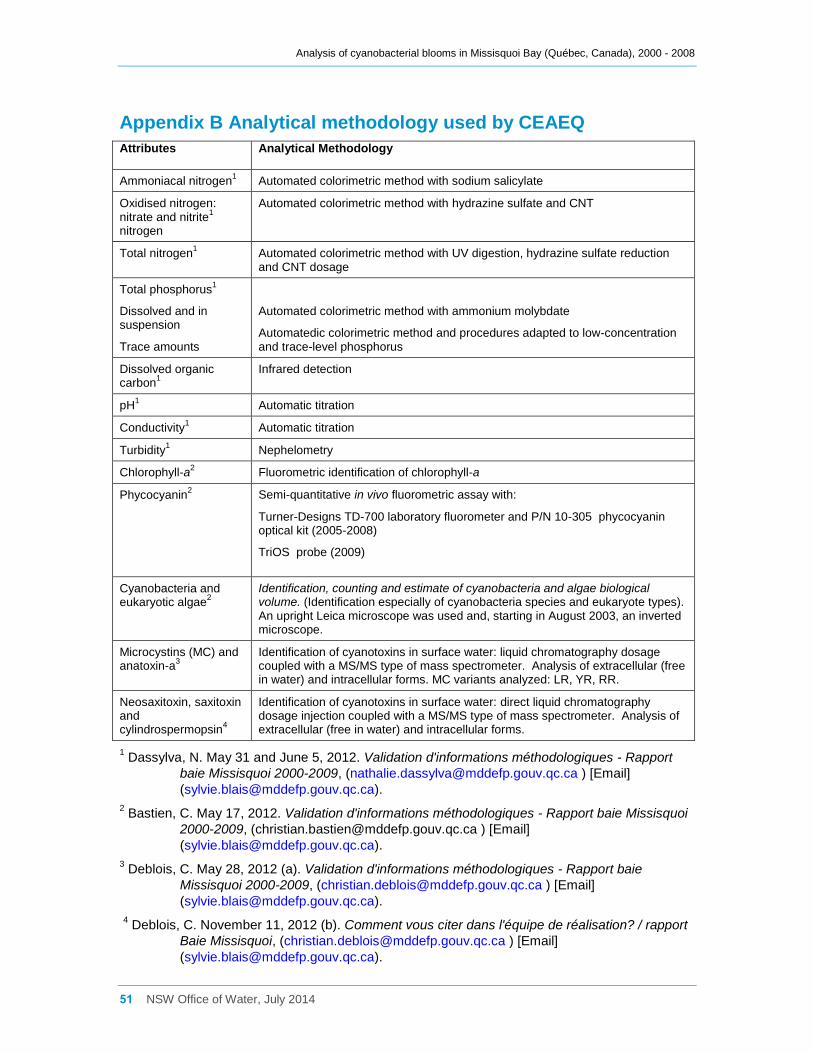
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
51 NSW Office of Water, July 2014
Appendix B Analytical methodology used by CEAEQ
Attributes
Analytical Methodology
Ammoniacal nitrogen1 Automated colorimetric method with sodium salicylate
Oxidised nitrogen: nitrate and nitrite
1
nitrogen
Automated colorimetric method with hydrazine sulfate and CNT
Total nitrogen1 Automated colorimetric method with UV digestion, hydrazine sulfate reduction
and CNT dosage
Total phosphorus1
Dissolved and in suspension
Trace amounts
Automated colorimetric method with ammonium molybdate
Automatedic colorimetric method and procedures adapted to low-concentration and trace-level phosphorus
Dissolved organic carbon
1
Infrared detection
pH1 Automatic titration
Conductivity1 Automatic titration
Turbidity1 Nephelometry
Chlorophyll-a2 Fluorometric identification of chlorophyll-a
Phycocyanin2 Semi-quantitative in vivo fluorometric assay with:
Turner-Designs TD-700 laboratory fluorometer and P/N 10-305 phycocyanin optical kit (2005-2008)
TriOS probe (2009)
Cyanobacteria and eukaryotic algae
2
Identification, counting and estimate of cyanobacteria and algae biological volume. (Identification especially of cyanobacteria species and eukaryote types). An upright Leica microscope was used and, starting in August 2003, an inverted microscope.
Microcystins (MC) and anatoxin-a
3
Identification of cyanotoxins in surface water: liquid chromatography dosage coupled with a MS/MS type of mass spectrometer. Analysis of extracellular (free in water) and intracellular forms. MC variants analyzed: LR, YR, RR.
Neosaxitoxin, saxitoxin and cylindrospermopsin
4
Identification of cyanotoxins in surface water: direct liquid chromatography dosage injection coupled with a MS/MS type of mass spectrometer. Analysis of extracellular (free in water) and intracellular forms.
1 Dassylva, N. May 31 and June 5, 2012. Validation d'informations méthodologiques - Rapport
baie Missisquoi 2000-2009, ([email protected] ) [Email]
2 Bastien, C. May 17, 2012. Validation d'informations méthodologiques - Rapport baie Missisquoi
2000-2009, ([email protected] ) [Email]
3 Deblois, C. May 28, 2012 (a). Validation d'informations méthodologiques - Rapport baie
Missisquoi 2000-2009, ([email protected] ) [Email]
4 Deblois, C. November 11, 2012 (b). Comment vous citer dans l'équipe de réalisation? / rapport
Baie Missisquoi, ([email protected] ) [Email]
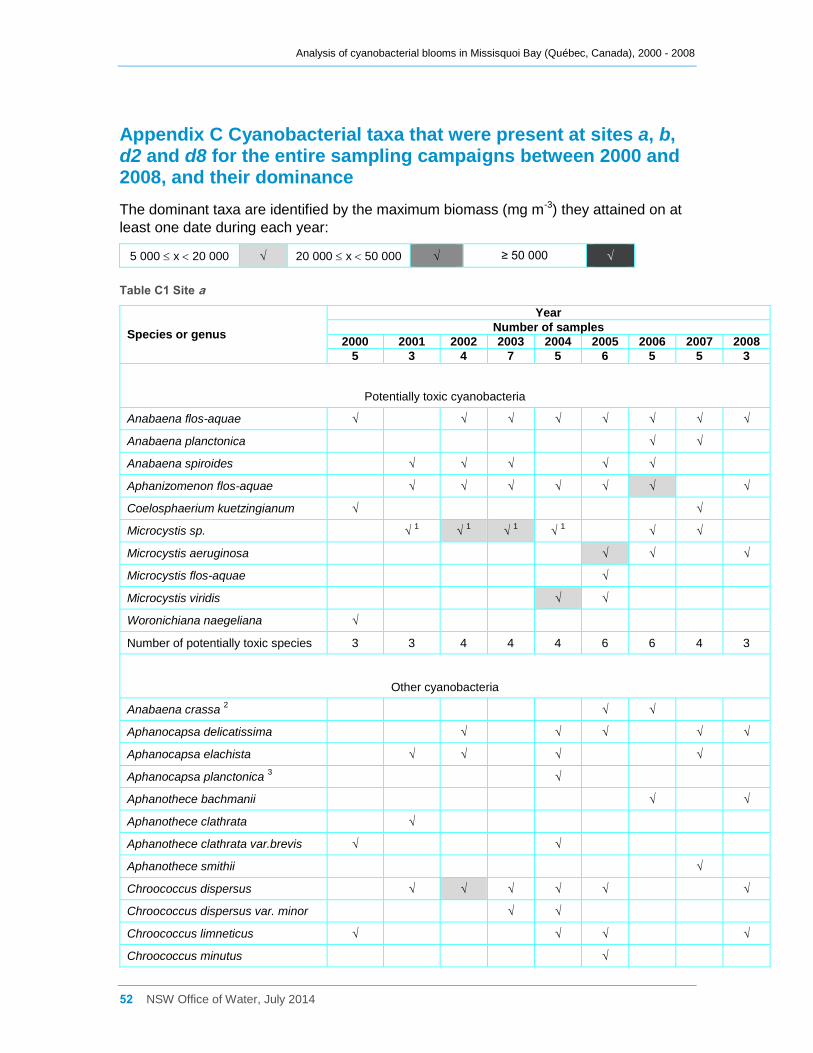
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
52 NSW Office of Water, July 2014
Appendix C Cyanobacterial taxa that were present at sites a, b, d2 and d8 for the entire sampling campaigns between 2000 and 2008, and their dominance
The dominant taxa are identified by the maximum biomass (mg m-3) they attained on at
least one date during each year:
5 000 x 20 000 20 000 x 50 000 ≥ 50 000
Table C1 Site a
Species or genus
Year
Number of samples
2000 2001 2002 2003 2004 2005 2006 2007 2008
5 3 4 7 5 6 5 5 3
Potentially toxic cyanobacteria
Anabaena flos-aquae √ √ √ √ √ √ √ √
Anabaena planctonica √ √
Anabaena spiroides √ √ √ √ √
Aphanizomenon flos-aquae √ √ √ √ √ √ √
Coelosphaerium kuetzingianum √ √
Microcystis sp. √ 1 √
1 √
1 √
1 √ √
Microcystis aeruginosa √ √ √
Microcystis flos-aquae √
Microcystis viridis √ √
Woronichiana naegeliana √
Number of potentially toxic species 3 3 4 4 4 6 6 4 3
Other cyanobacteria
Anabaena crassa 2 √ √
Aphanocapsa delicatissima √ √ √ √ √
Aphanocapsa elachista √ √ √ √
Aphanocapsa planctonica 3 √
Aphanothece bachmanii √ √
Aphanothece clathrata √
Aphanothece clathrata var.brevis √ √
Aphanothece smithii √
Chroococcus dispersus √ √ √ √ √ √
Chroococcus dispersus var. minor √ √
Chroococcus limneticus √ √ √ √
Chroococcus minutus √
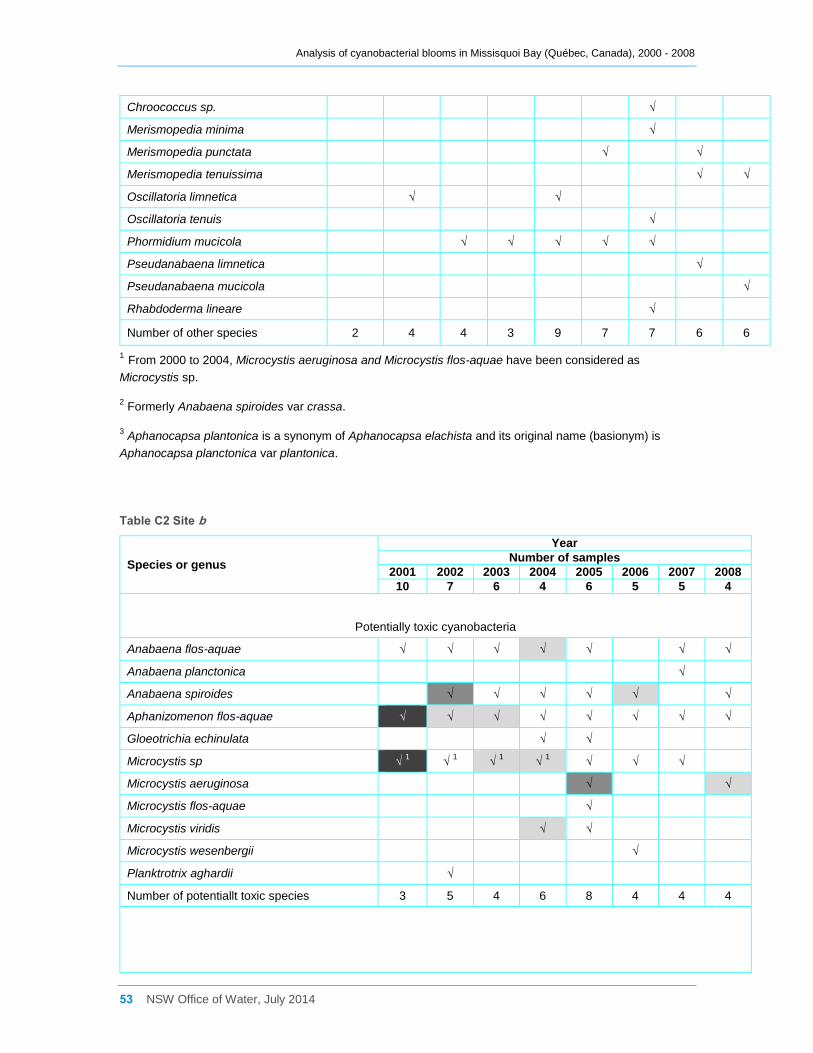
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
53 NSW Office of Water, July 2014
Chroococcus sp. √
Merismopedia minima √
Merismopedia punctata √ √
Merismopedia tenuissima √ √
Oscillatoria limnetica √ √
Oscillatoria tenuis √
Phormidium mucicola √ √ √ √ √
Pseudanabaena limnetica √
Pseudanabaena mucicola √
Rhabdoderma lineare √
Number of other species 2 4 4 3 9 7 7 6 6
1 From 2000 to 2004, Microcystis aeruginosa and Microcystis flos-aquae have been considered as
Microcystis sp.
2 Formerly Anabaena spiroides var crassa.
3 Aphanocapsa plantonica is a synonym of Aphanocapsa elachista and its original name (basionym) is
Aphanocapsa planctonica var plantonica.
Table C2 Site b
Species or genus
Year
Number of samples
2001 2002 2003 2004 2005 2006 2007 2008
10 7 6 4 6 5 5 4
Potentially toxic cyanobacteria
Anabaena flos-aquae √ √ √ √ √ √ √
Anabaena planctonica √
Anabaena spiroides √ √ √ √ √ √
Aphanizomenon flos-aquae √ √ √ √ √ √ √ √
Gloeotrichia echinulata √ √
Microcystis sp √ 1 √
1 √
1 √
1 √ √ √
Microcystis aeruginosa √ √
Microcystis flos-aquae √
Microcystis viridis √ √
Microcystis wesenbergii √
Planktrotrix aghardii √
Number of potentiallt toxic species 3 5 4 6 8 4 4 4
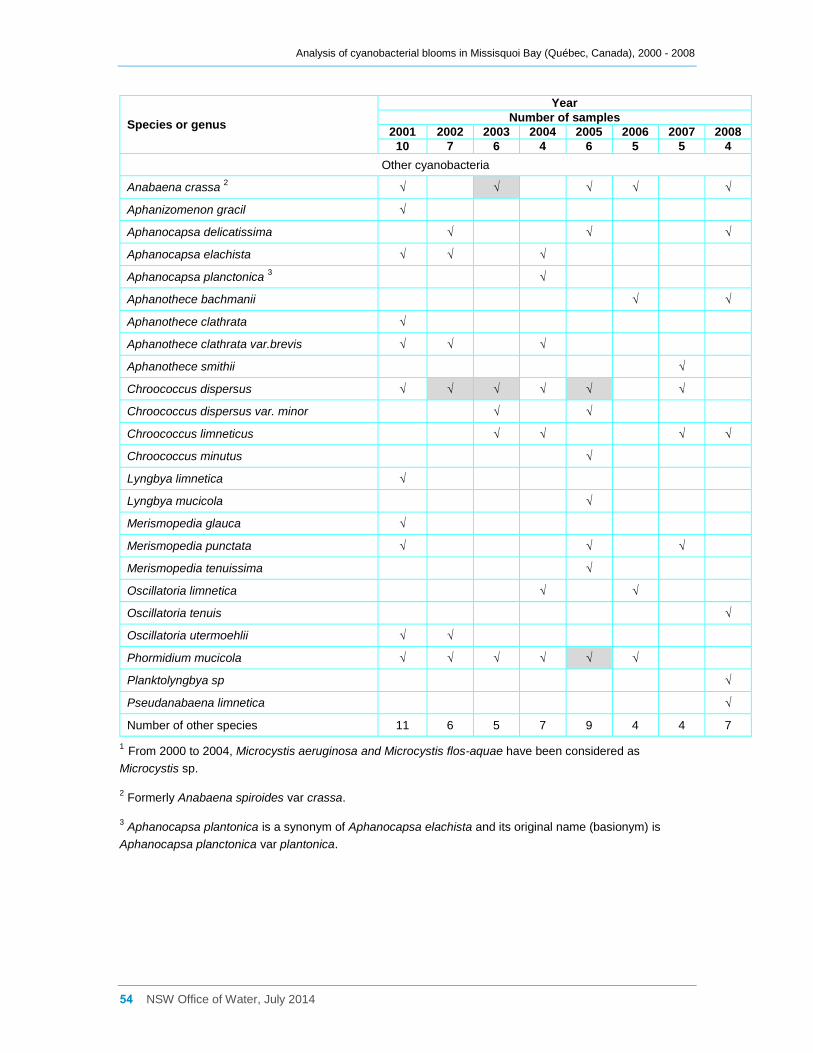
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
54 NSW Office of Water, July 2014
Species or genus
Year
Number of samples
2001 2002 2003 2004 2005 2006 2007 2008
10 7 6 4 6 5 5 4
Other cyanobacteria
Anabaena crassa 2 √ √ √ √ √
Aphanizomenon gracil √
Aphanocapsa delicatissima √ √ √
Aphanocapsa elachista √ √ √
Aphanocapsa planctonica 3 √
Aphanothece bachmanii √ √
Aphanothece clathrata √
Aphanothece clathrata var.brevis √ √ √
Aphanothece smithii √
Chroococcus dispersus √ √ √ √ √ √
Chroococcus dispersus var. minor √ √
Chroococcus limneticus √ √ √ √
Chroococcus minutus √
Lyngbya limnetica √
Lyngbya mucicola √
Merismopedia glauca √
Merismopedia punctata √ √ √
Merismopedia tenuissima √
Oscillatoria limnetica √ √
Oscillatoria tenuis √
Oscillatoria utermoehlii √ √
Phormidium mucicola √ √ √ √ √ √
Planktolyngbya sp √
Pseudanabaena limnetica √
Number of other species 11 6 5 7 9 4 4 7
1 From 2000 to 2004, Microcystis aeruginosa and Microcystis flos-aquae have been considered as
Microcystis sp.
2 Formerly Anabaena spiroides var crassa.
3 Aphanocapsa plantonica is a synonym of Aphanocapsa elachista and its original name (basionym) is
Aphanocapsa planctonica var plantonica.
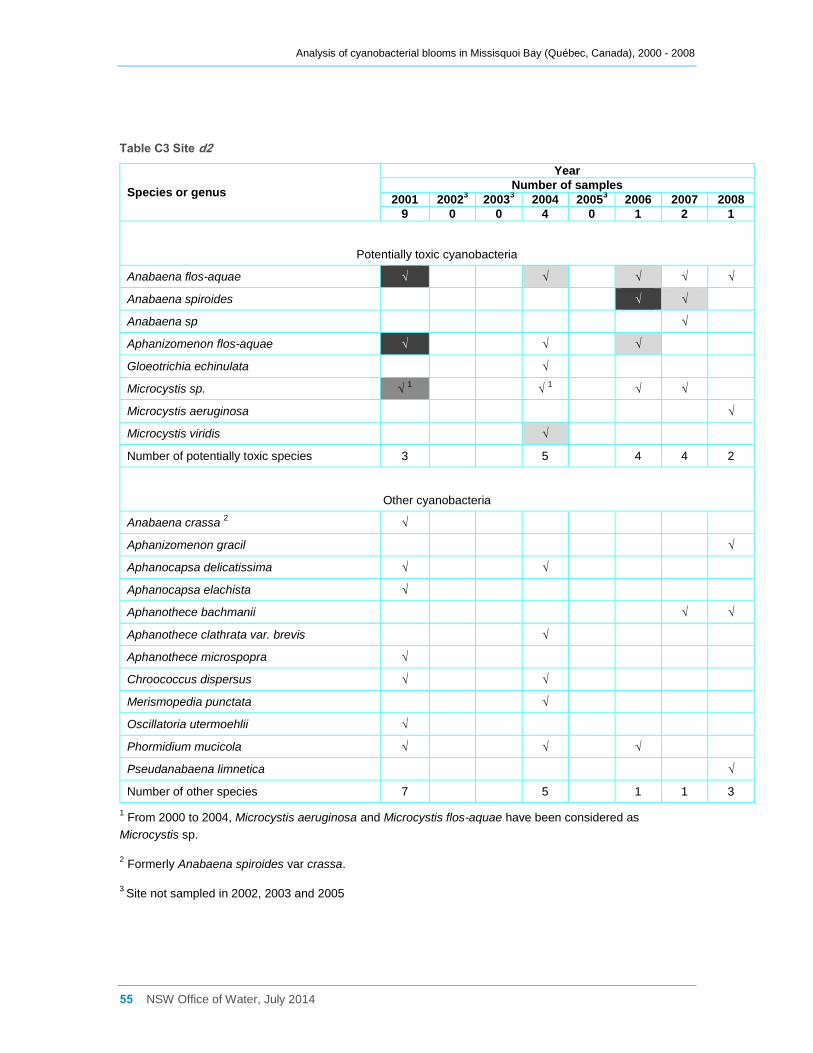
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
55 NSW Office of Water, July 2014
Table C3 Site d2
Species or genus
Year
Number of samples
2001 20023 2003
3 2004 2005
3 2006 2007 2008
9 0 0 4 0 1 2 1
Potentially toxic cyanobacteria
Anabaena flos-aquae √ √ √ √ √
Anabaena spiroides √ √
Anabaena sp √
Aphanizomenon flos-aquae √ √ √
Gloeotrichia echinulata √
Microcystis sp. √ 1 √
1 √ √
Microcystis aeruginosa √
Microcystis viridis √
Number of potentially toxic species 3 5 4 4 2
Other cyanobacteria
Anabaena crassa 2 √
Aphanizomenon gracil √
Aphanocapsa delicatissima √ √
Aphanocapsa elachista √
Aphanothece bachmanii √ √
Aphanothece clathrata var. brevis √
Aphanothece microspopra √
Chroococcus dispersus √ √
Merismopedia punctata √
Oscillatoria utermoehlii √
Phormidium mucicola √ √ √
Pseudanabaena limnetica √
Number of other species 7 5 1 1 3
1 From 2000 to 2004, Microcystis aeruginosa and Microcystis flos-aquae have been considered as
Microcystis sp.
2 Formerly Anabaena spiroides var crassa.
3 Site not sampled in 2002, 2003 and 2005
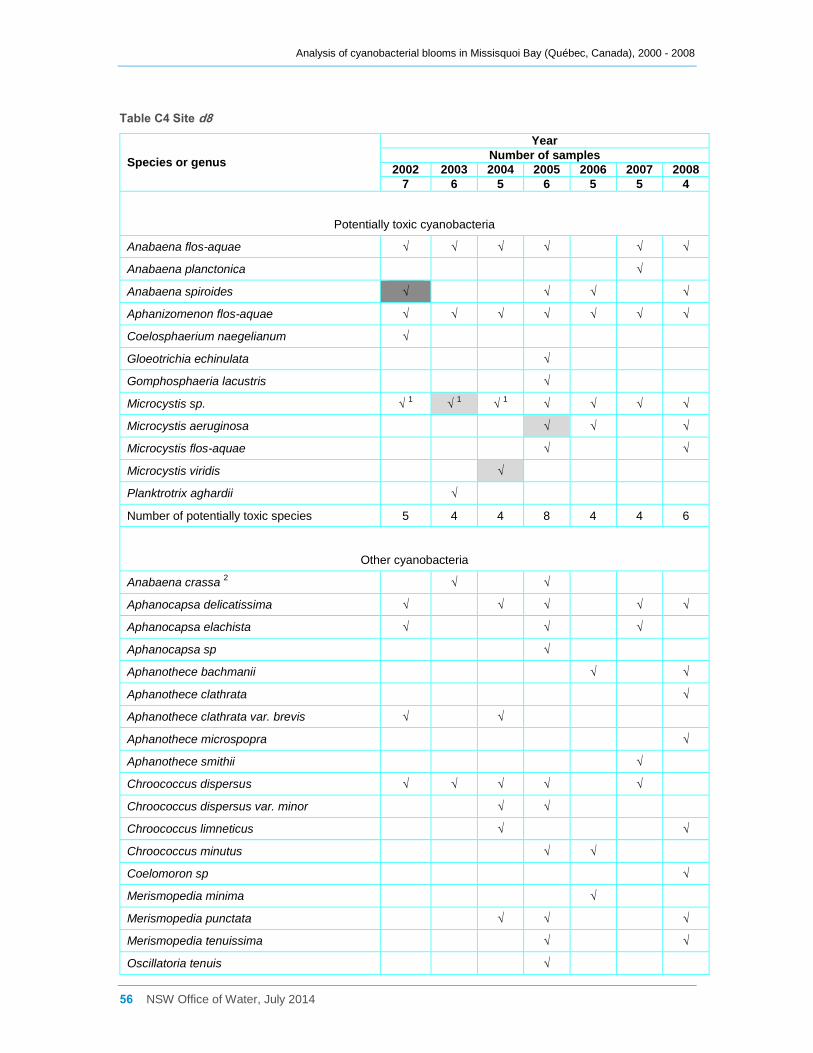
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
56 NSW Office of Water, July 2014
Table C4 Site d8
Species or genus
Year
Number of samples
2002 2003 2004 2005 2006 2007 2008
7 6 5 6 5 5 4
Potentially toxic cyanobacteria
Anabaena flos-aquae √ √ √ √ √ √
Anabaena planctonica √
Anabaena spiroides √ √ √ √
Aphanizomenon flos-aquae √ √ √ √ √ √ √
Coelosphaerium naegelianum √
Gloeotrichia echinulata √
Gomphosphaeria lacustris √
Microcystis sp. √ 1 √
1 √
1 √ √ √ √
Microcystis aeruginosa √ √ √
Microcystis flos-aquae √ √
Microcystis viridis √
Planktrotrix aghardii √
Number of potentially toxic species 5 4 4 8 4 4 6
Other cyanobacteria
Anabaena crassa 2 √ √
Aphanocapsa delicatissima √ √ √ √ √
Aphanocapsa elachista √ √ √
Aphanocapsa sp √
Aphanothece bachmanii √ √
Aphanothece clathrata √
Aphanothece clathrata var. brevis √ √
Aphanothece microspopra √
Aphanothece smithii √
Chroococcus dispersus √ √ √ √ √
Chroococcus dispersus var. minor √ √
Chroococcus limneticus √ √
Chroococcus minutus √ √
Coelomoron sp √
Merismopedia minima √
Merismopedia punctata √ √ √
Merismopedia tenuissima √ √
Oscillatoria tenuis √
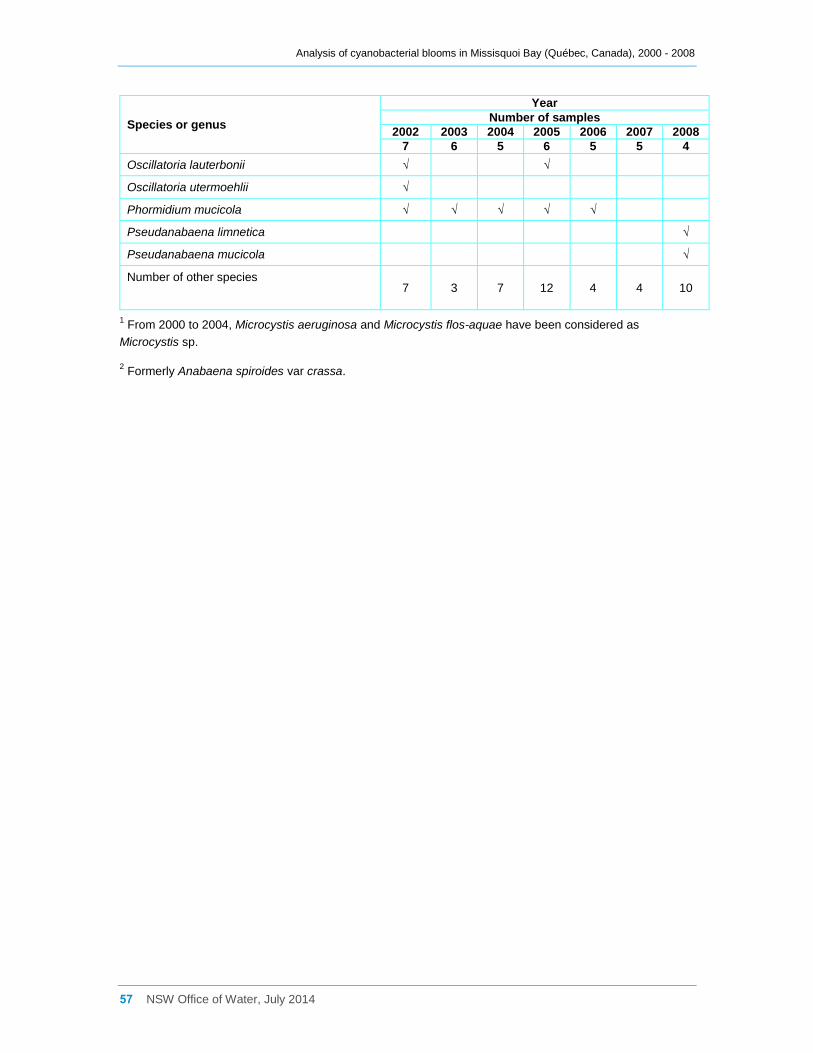
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
57 NSW Office of Water, July 2014
Species or genus
Year
Number of samples
2002 2003 2004 2005 2006 2007 2008
7 6 5 6 5 5 4
Oscillatoria lauterbonii √ √
Oscillatoria utermoehlii √
Phormidium mucicola √ √ √ √ √
Pseudanabaena limnetica √
Pseudanabaena mucicola √
Number of other species
7 3 7 12 4 4 10
1 From 2000 to 2004, Microcystis aeruginosa and Microcystis flos-aquae have been considered as
Microcystis sp.
2 Formerly Anabaena spiroides var crassa.
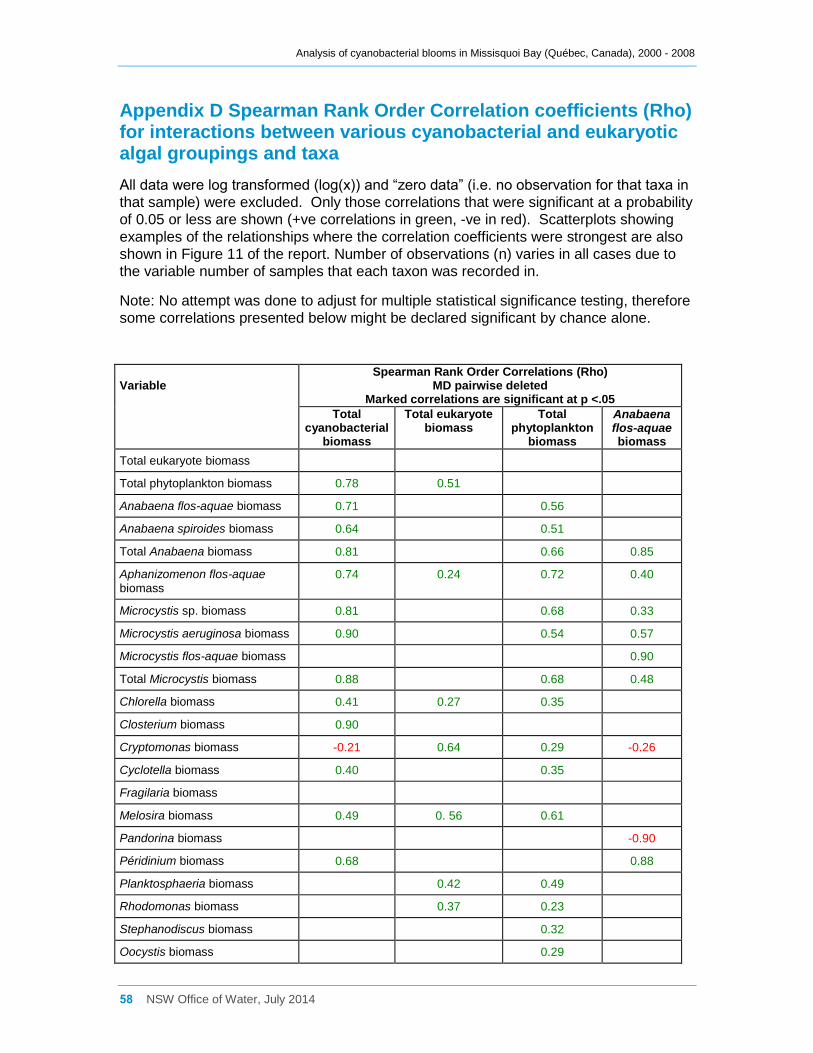
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
58 NSW Office of Water, July 2014
Appendix D Spearman Rank Order Correlation coefficients (Rho) for interactions between various cyanobacterial and eukaryotic algal groupings and taxa
All data were log transformed (log(x)) and “zero data” (i.e. no observation for that taxa in
that sample) were excluded. Only those correlations that were significant at a probability
of 0.05 or less are shown (+ve correlations in green, -ve in red). Scatterplots showing
examples of the relationships where the correlation coefficients were strongest are also
shown in Figure 11 of the report. Number of observations (n) varies in all cases due to
the variable number of samples that each taxon was recorded in.
Note: No attempt was done to adjust for multiple statistical significance testing, therefore
some correlations presented below might be declared significant by chance alone.
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Total cyanobacterial
biomass
Total eukaryote biomass
Total phytoplankton
biomass
Anabaena flos-aquae biomass
Total eukaryote biomass
Total phytoplankton biomass 0.78 0.51
Anabaena flos-aquae biomass 0.71 0.56
Anabaena spiroides biomass 0.64 0.51
Total Anabaena biomass 0.81 0.66 0.85
Aphanizomenon flos-aquae
biomass 0.74 0.24 0.72 0.40
Microcystis sp. biomass 0.81 0.68 0.33
Microcystis aeruginosa biomass 0.90 0.54 0.57
Microcystis flos-aquae biomass 0.90
Total Microcystis biomass 0.88 0.68 0.48
Chlorella biomass 0.41 0.27 0.35
Closterium biomass 0.90
Cryptomonas biomass -0.21 0.64 0.29 -0.26
Cyclotella biomass 0.40 0.35
Fragilaria biomass
Melosira biomass 0.49 0. 56 0.61
Pandorina biomass -0.90
Péridinium biomass 0.68 0.88
Planktosphaeria biomass 0.42 0.49
Rhodomonas biomass 0.37 0.23
Stephanodiscus biomass 0.32
Oocystis biomass 0.29
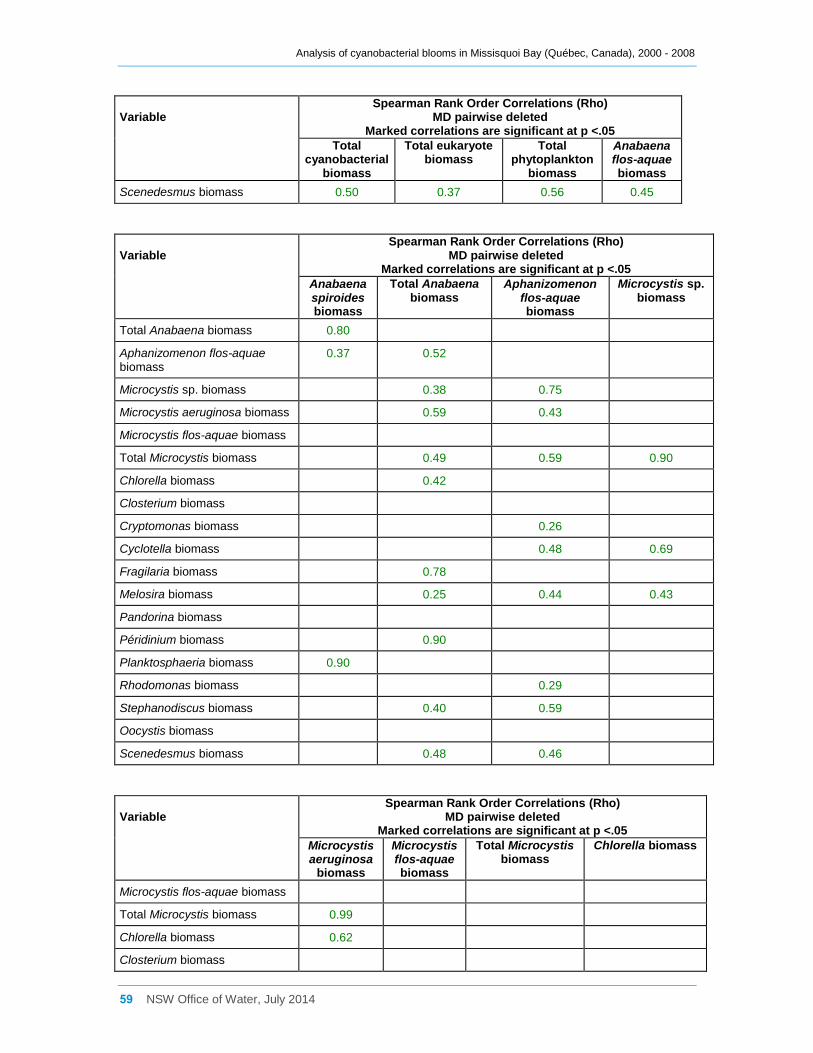
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
59 NSW Office of Water, July 2014
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Total cyanobacterial
biomass
Total eukaryote biomass
Total phytoplankton
biomass
Anabaena flos-aquae biomass
Scenedesmus biomass 0.50 0.37 0.56 0.45
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Anabaena spiroides biomass
Total Anabaena biomass
Aphanizomenon flos-aquae biomass
Microcystis sp. biomass
Total Anabaena biomass 0.80
Aphanizomenon flos-aquae biomass
0.37 0.52
Microcystis sp. biomass 0.38 0.75
Microcystis aeruginosa biomass 0.59 0.43
Microcystis flos-aquae biomass
Total Microcystis biomass 0.49 0.59 0.90
Chlorella biomass 0.42
Closterium biomass
Cryptomonas biomass 0.26
Cyclotella biomass 0.48 0.69
Fragilaria biomass 0.78
Melosira biomass 0.25 0.44 0.43
Pandorina biomass
Péridinium biomass 0.90
Planktosphaeria biomass 0.90
Rhodomonas biomass 0.29
Stephanodiscus biomass 0.40 0.59
Oocystis biomass
Scenedesmus biomass 0.48 0.46
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Microcystis aeruginosa
biomass
Microcystis flos-aquae biomass
Total Microcystis biomass
Chlorella biomass
Microcystis flos-aquae biomass
Total Microcystis biomass 0.99
Chlorella biomass 0.62
Closterium biomass
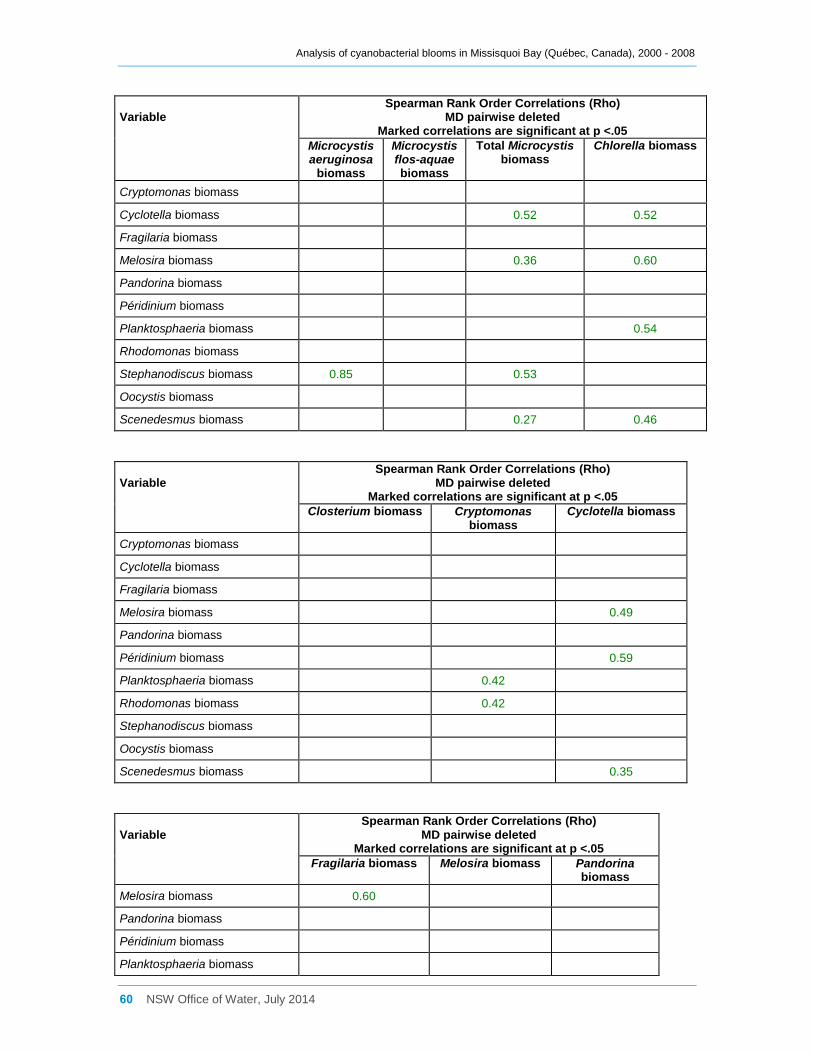
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
60 NSW Office of Water, July 2014
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Microcystis aeruginosa
biomass
Microcystis flos-aquae biomass
Total Microcystis biomass
Chlorella biomass
Cryptomonas biomass
Cyclotella biomass 0.52 0.52
Fragilaria biomass
Melosira biomass 0.36 0.60
Pandorina biomass
Péridinium biomass
Planktosphaeria biomass 0.54
Rhodomonas biomass
Stephanodiscus biomass 0.85 0.53
Oocystis biomass
Scenedesmus biomass 0.27 0.46
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Closterium biomass Cryptomonas biomass
Cyclotella biomass
Cryptomonas biomass
Cyclotella biomass
Fragilaria biomass
Melosira biomass 0.49
Pandorina biomass
Péridinium biomass 0.59
Planktosphaeria biomass 0.42
Rhodomonas biomass 0.42
Stephanodiscus biomass
Oocystis biomass
Scenedesmus biomass 0.35
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Fragilaria biomass Melosira biomass Pandorina biomass
Melosira biomass 0.60
Pandorina biomass
Péridinium biomass
Planktosphaeria biomass
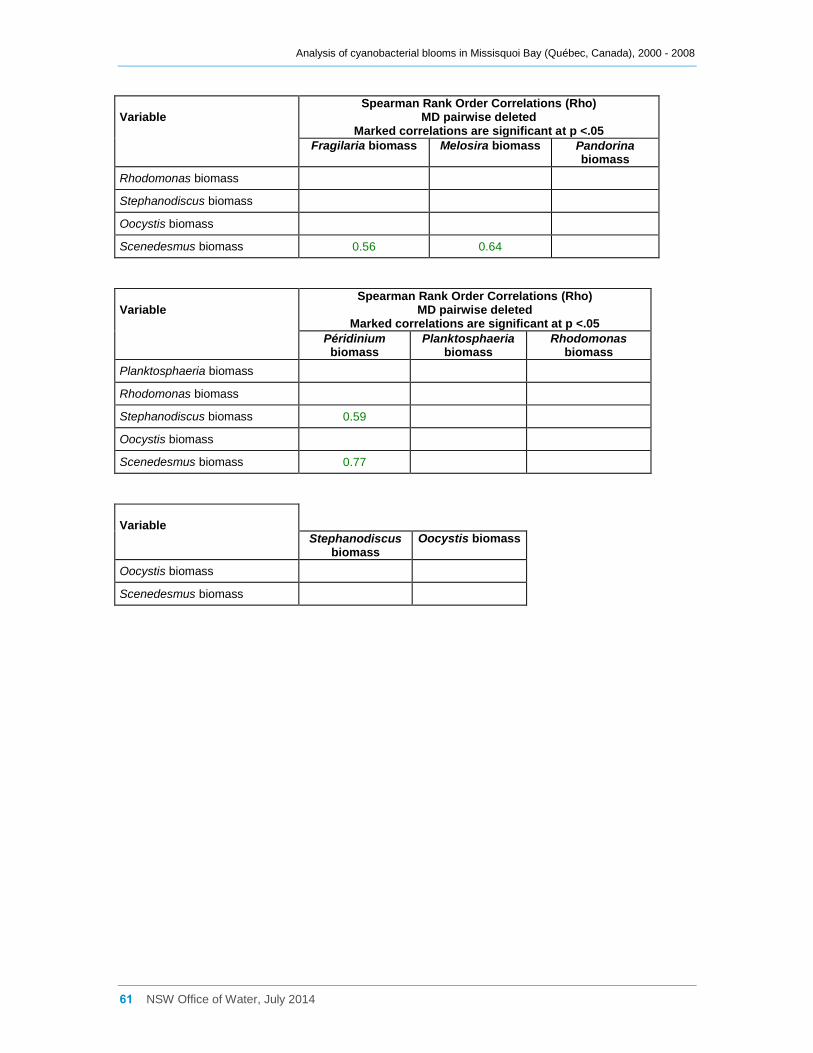
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
61 NSW Office of Water, July 2014
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Fragilaria biomass Melosira biomass Pandorina biomass
Rhodomonas biomass
Stephanodiscus biomass
Oocystis biomass
Scenedesmus biomass 0.56 0.64
Variable
Spearman Rank Order Correlations (Rho) MD pairwise deleted
Marked correlations are significant at p <.05
Péridinium biomass
Planktosphaeria biomass
Rhodomonas biomass
Planktosphaeria biomass
Rhodomonas biomass
Stephanodiscus biomass 0.59
Oocystis biomass
Scenedesmus biomass 0.77
Variable
Stephanodiscus biomass
Oocystis biomass
Oocystis biomass
Scenedesmus biomass
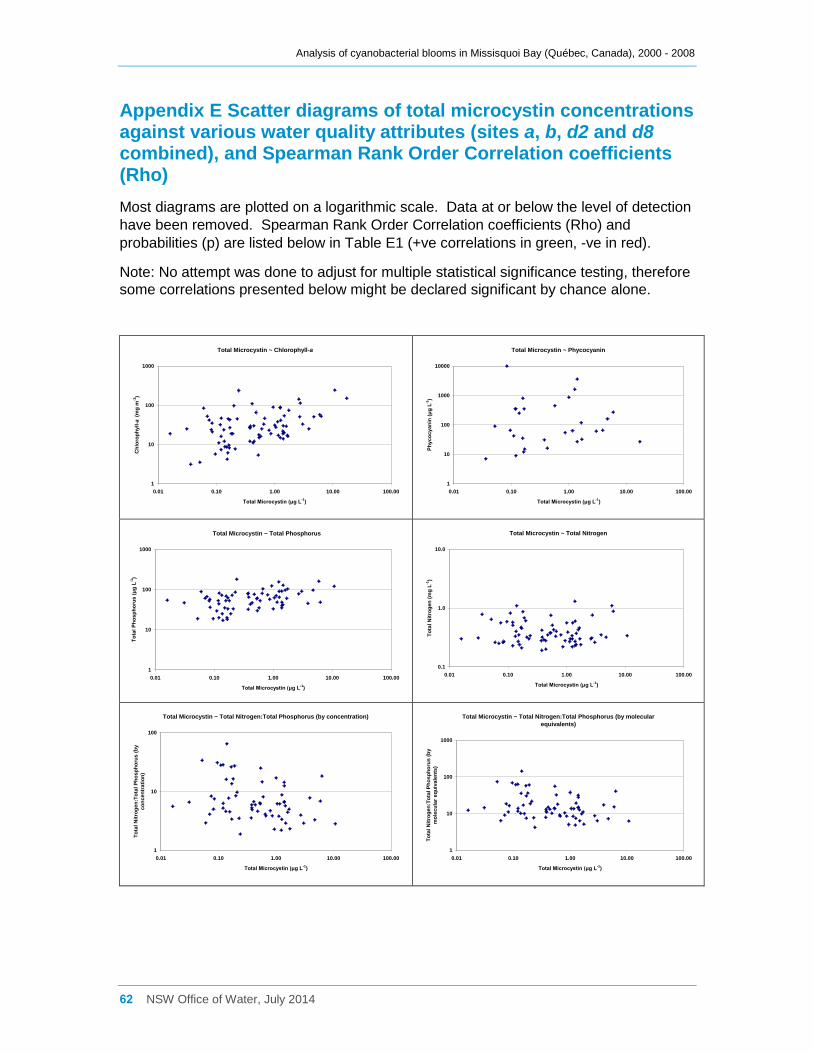
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
62 NSW Office of Water, July 2014
Appendix E Scatter diagrams of total microcystin concentrations against various water quality attributes (sites a, b, d2 and d8 combined), and Spearman Rank Order Correlation coefficients (Rho)
Most diagrams are plotted on a logarithmic scale. Data at or below the level of detection
have been removed. Spearman Rank Order Correlation coefficients (Rho) and
probabilities (p) are listed below in Table E1 (+ve correlations in green, -ve in red).
Note: No attempt was done to adjust for multiple statistical significance testing, therefore
some correlations presented below might be declared significant by chance alone.
Total Microcystin ~ Chlorophyll-a
1
10
100
1000
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Ch
loro
ph
yll
- a (
mg
m-3
)
Total Microcystin ~ Phycocyanin
1
10
100
1000
10000
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Ph
yc
oc
ya
nin
(μ
g L
-1)
Total Microcystin ~ Total Phosphorus
1
10
100
1000
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
To
tal
Ph
os
ph
oru
s (
μg
L-1
)
Total Microcystin ~ Total Nitrogen
0.1
1.0
10.0
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
To
tal
Nit
rog
en
(m
g L
-1)
Total Microcystin ~ Total Nitrogen:Total Phosphorus (by concentration)
1
10
100
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
To
tal
Nit
rog
en
:To
tal
Ph
os
ph
oru
s (
by
co
nc
en
tra
tio
n)
Total Microcystin ~ Total Nitrogen:Total Phosphorus (by molecular
equivalents)
1
10
100
1000
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
To
tal
Nit
rog
en
:To
tal
Ph
os
ph
oru
s (
by
mo
lec
ula
r e
qu
iva
len
ts)
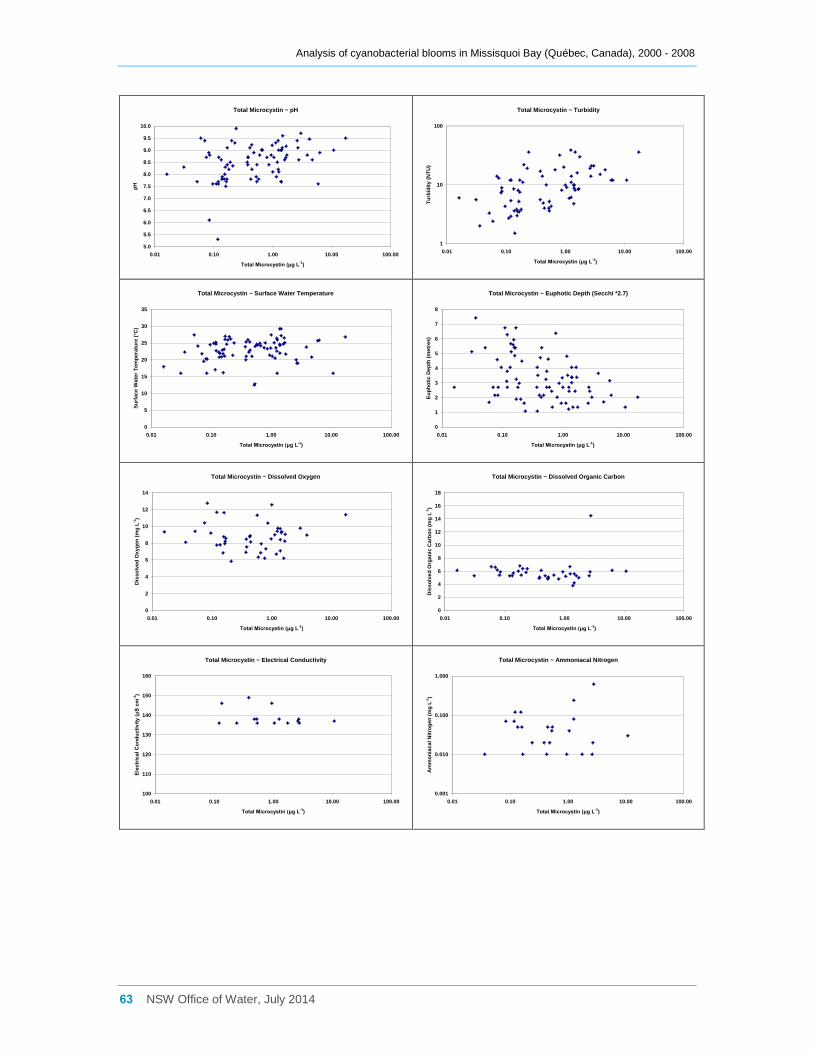
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
63 NSW Office of Water, July 2014
Total Microcystin ~ pH
5.0
5.5
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
10.0
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
pH
Total Microcystin ~ Turbidity
1
10
100
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Tu
rbid
ity
(N
TU
)
Total Microcystin ~ Surface Water Temperature
0
5
10
15
20
25
30
35
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Su
rfa
ce
Wa
ter
Te
mp
era
ture
(°C
)
Total Microcystin ~ Euphotic Depth (Secchi *2.7)
0
1
2
3
4
5
6
7
8
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Eu
ph
oti
c D
ep
th (
me
tre
s)
Total Microcystin ~ Dissolved Oxygen
0
2
4
6
8
10
12
14
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Dis
so
lve
d O
xy
ge
n (
mg
L-1
)
Total Microcystin ~ Dissolved Organic Carbon
0
2
4
6
8
10
12
14
16
18
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Dis
so
lve
d O
rga
nic
Ca
rbo
n (
mg
L-1
)
Total Microcystin ~ Electrical Conductivity
100
110
120
130
140
150
160
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Ele
ctr
ical
Co
nd
ucti
vit
y (
μS
cm
-1)
Total Microcystin ~ Ammoniacal Nitrogen
0.001
0.010
0.100
1.000
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Am
mo
nia
ca
l N
itro
ge
n (
mg
L-1
)
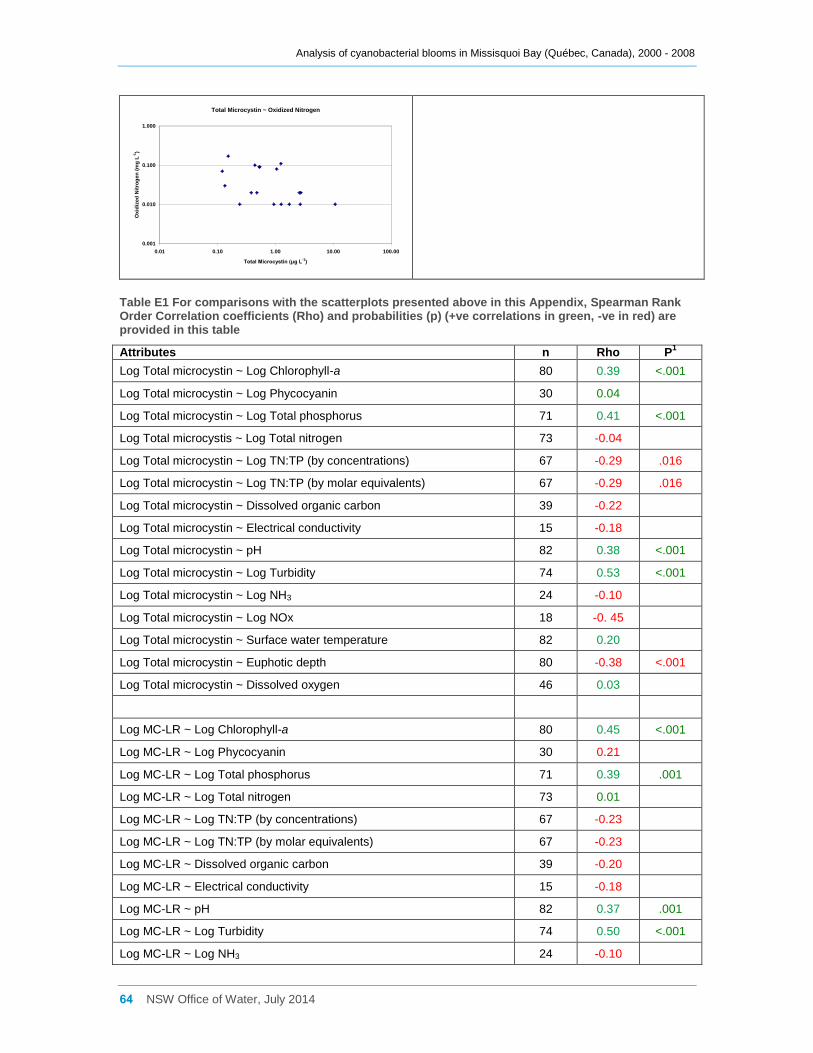
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
64 NSW Office of Water, July 2014
Total Microcystin ~ Oxidized Nitrogen
0.001
0.010
0.100
1.000
0.01 0.10 1.00 10.00 100.00
Total Microcystin (μg L-1
)
Ox
idiz
ed
Nit
rog
en
(m
g L
-1)
Table E1 For comparisons with the scatterplots presented above in this Appendix, Spearman Rank Order Correlation coefficients (Rho) and probabilities (p) (+ve correlations in green, -ve in red) are provided in this table
Attributes n Rho P1
Log Total microcystin ~ Log Chlorophyll-a 80 0.39 <.001
Log Total microcystin ~ Log Phycocyanin 30 0.04
Log Total microcystin ~ Log Total phosphorus 71 0.41 <.001
Log Total microcystis ~ Log Total nitrogen 73 -0.04
Log Total microcystin ~ Log TN:TP (by concentrations) 67 -0.29 .016
Log Total microcystin ~ Log TN:TP (by molar equivalents) 67 -0.29 .016
Log Total microcystin ~ Dissolved organic carbon 39 -0.22
Log Total microcystin ~ Electrical conductivity 15 -0.18
Log Total microcystin ~ pH 82 0.38 <.001
Log Total microcystin ~ Log Turbidity 74 0.53 <.001
Log Total microcystin ~ Log NH3 24 -0.10
Log Total microcystin ~ Log NOx 18 -0. 45
Log Total microcystin ~ Surface water temperature 82 0.20
Log Total microcystin ~ Euphotic depth 80 -0.38 <.001
Log Total microcystin ~ Dissolved oxygen 46 0.03
Log MC-LR ~ Log Chlorophyll-a 80 0.45 <.001
Log MC-LR ~ Log Phycocyanin 30 0.21
Log MC-LR ~ Log Total phosphorus 71 0.39 .001
Log MC-LR ~ Log Total nitrogen 73 0.01
Log MC-LR ~ Log TN:TP (by concentrations) 67 -0.23
Log MC-LR ~ Log TN:TP (by molar equivalents) 67 -0.23
Log MC-LR ~ Dissolved organic carbon 39 -0.20
Log MC-LR ~ Electrical conductivity 15 -0.18
Log MC-LR ~ pH 82 0.37 .001
Log MC-LR ~ Log Turbidity 74 0.50 <.001
Log MC-LR ~ Log NH3 24 -0.10
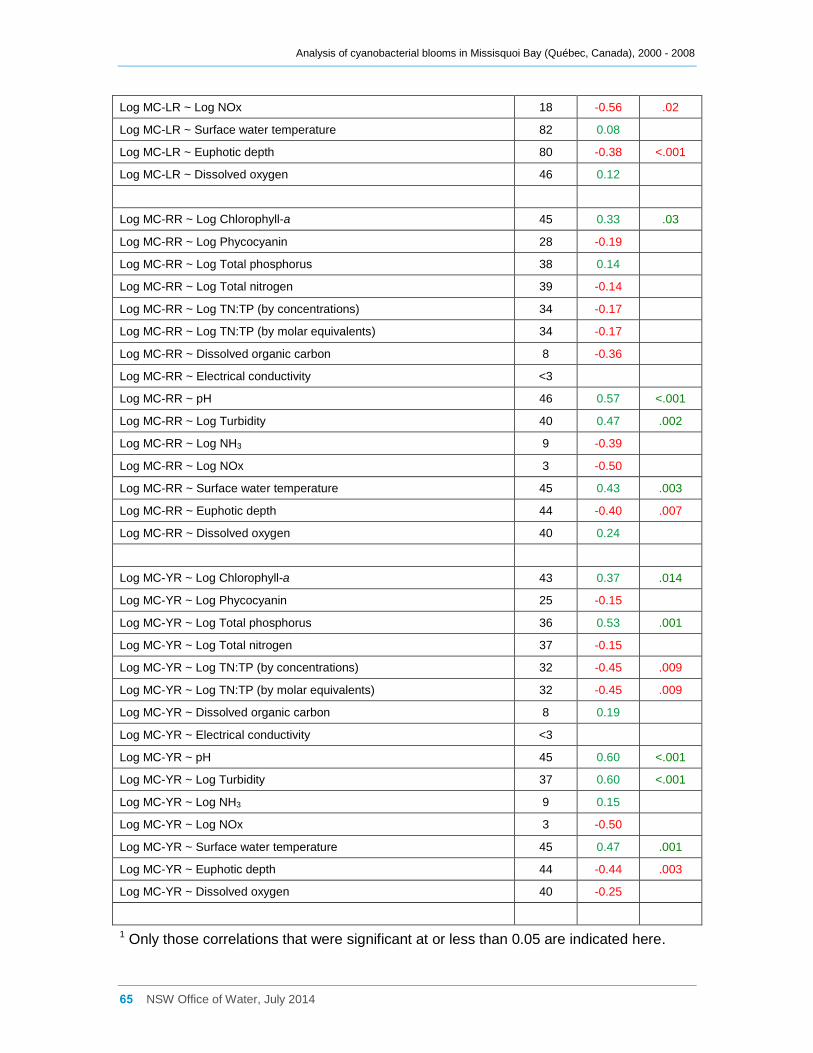
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
65 NSW Office of Water, July 2014
Log MC-LR ~ Log NOx 18 -0.56 .02
Log MC-LR ~ Surface water temperature 82 0.08
Log MC-LR ~ Euphotic depth 80 -0.38 <.001
Log MC-LR ~ Dissolved oxygen 46 0.12
Log MC-RR ~ Log Chlorophyll-a 45 0.33 .03
Log MC-RR ~ Log Phycocyanin 28 -0.19
Log MC-RR ~ Log Total phosphorus 38 0.14
Log MC-RR ~ Log Total nitrogen 39 -0.14
Log MC-RR ~ Log TN:TP (by concentrations) 34 -0.17
Log MC-RR ~ Log TN:TP (by molar equivalents) 34 -0.17
Log MC-RR ~ Dissolved organic carbon 8 -0.36
Log MC-RR ~ Electrical conductivity <3
Log MC-RR ~ pH 46 0.57 <.001
Log MC-RR ~ Log Turbidity 40 0.47 .002
Log MC-RR ~ Log NH3 9 -0.39
Log MC-RR ~ Log NOx 3 -0.50
Log MC-RR ~ Surface water temperature 45 0.43 .003
Log MC-RR ~ Euphotic depth 44 -0.40 .007
Log MC-RR ~ Dissolved oxygen 40 0.24
Log MC-YR ~ Log Chlorophyll-a 43 0.37 .014
Log MC-YR ~ Log Phycocyanin 25 -0.15
Log MC-YR ~ Log Total phosphorus 36 0.53 .001
Log MC-YR ~ Log Total nitrogen 37 -0.15
Log MC-YR ~ Log TN:TP (by concentrations) 32 -0.45 .009
Log MC-YR ~ Log TN:TP (by molar equivalents) 32 -0.45 .009
Log MC-YR ~ Dissolved organic carbon 8 0.19
Log MC-YR ~ Electrical conductivity <3
Log MC-YR ~ pH 45 0.60 <.001
Log MC-YR ~ Log Turbidity 37 0.60 <.001
Log MC-YR ~ Log NH3 9 0.15
Log MC-YR ~ Log NOx 3 -0.50
Log MC-YR ~ Surface water temperature 45 0.47 .001
Log MC-YR ~ Euphotic depth 44 -0.44 .003
Log MC-YR ~ Dissolved oxygen 40 -0.25
1 Only those correlations that were significant at or less than 0.05 are indicated here.
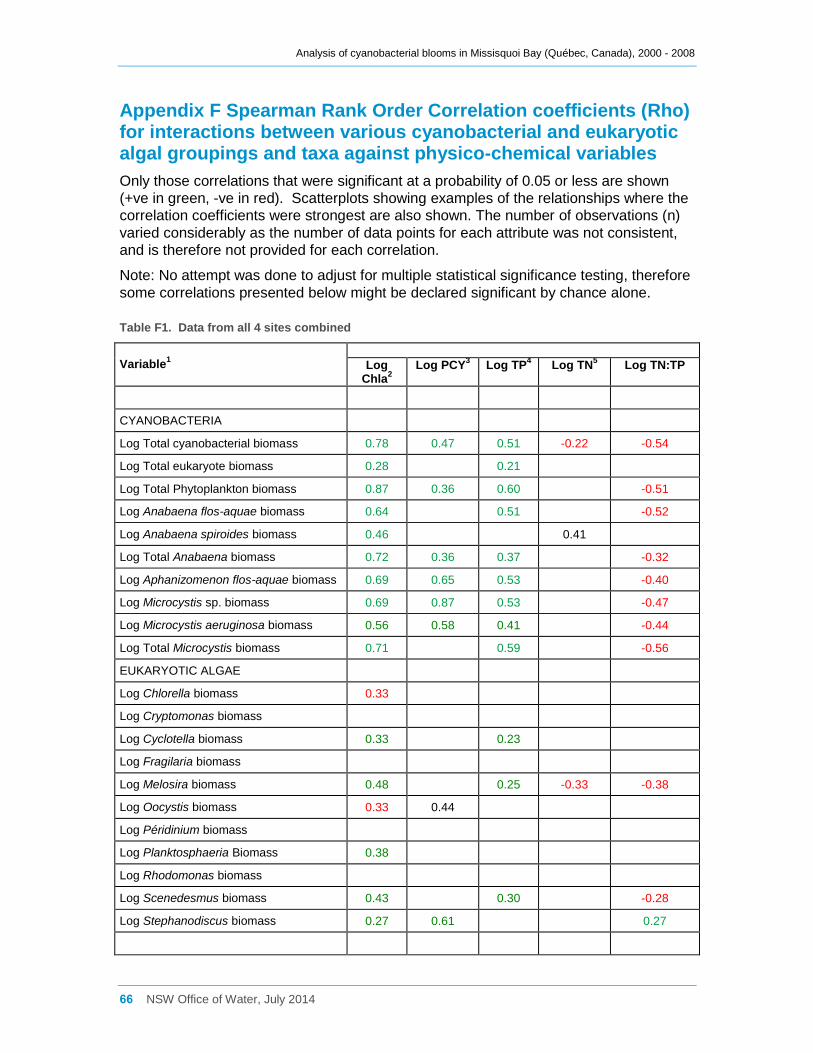
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
66 NSW Office of Water, July 2014
Appendix F Spearman Rank Order Correlation coefficients (Rho) for interactions between various cyanobacterial and eukaryotic algal groupings and taxa against physico-chemical variables
Only those correlations that were significant at a probability of 0.05 or less are shown
(+ve in green, -ve in red). Scatterplots showing examples of the relationships where the
correlation coefficients were strongest are also shown. The number of observations (n)
varied considerably as the number of data points for each attribute was not consistent,
and is therefore not provided for each correlation.
Note: No attempt was done to adjust for multiple statistical significance testing, therefore
some correlations presented below might be declared significant by chance alone.
Table F1. Data from all 4 sites combined
Variable
1
Log Chla
2
Log PCY3 Log TP
4 Log TN
5 Log TN:TP
CYANOBACTERIA
Log Total cyanobacterial biomass 0.78 0.47 0.51 -0.22 -0.54
Log Total eukaryote biomass 0.28 0.21
Log Total Phytoplankton biomass 0.87 0.36 0.60 -0.51
Log Anabaena flos-aquae biomass 0.64 0.51 -0.52
Log Anabaena spiroides biomass 0.46 0.41
Log Total Anabaena biomass 0.72 0.36 0.37 -0.32
Log Aphanizomenon flos-aquae biomass 0.69 0.65 0.53 -0.40
Log Microcystis sp. biomass 0.69 0.87 0.53 -0.47
Log Microcystis aeruginosa biomass 0.56 0.58 0.41 -0.44
Log Total Microcystis biomass 0.71 0.59 -0.56
EUKARYOTIC ALGAE
Log Chlorella biomass 0.33
Log Cryptomonas biomass
Log Cyclotella biomass 0.33 0.23
Log Fragilaria biomass
Log Melosira biomass 0.48 0.25 -0.33 -0.38
Log Oocystis biomass 0.33 0.44
Log Péridinium biomass
Log Planktosphaeria Biomass 0.38
Log Rhodomonas biomass
Log Scenedesmus biomass 0.43 0.30 -0.28
Log Stephanodiscus biomass 0.27 0.61 0.27
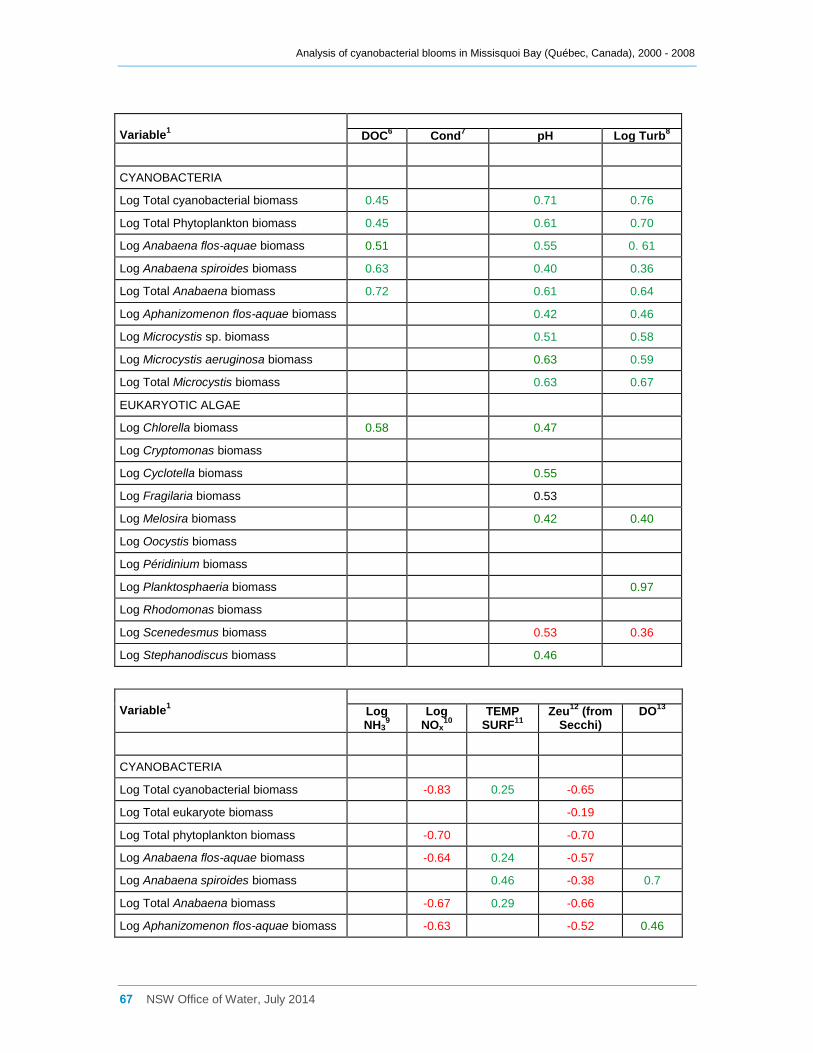
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
67 NSW Office of Water, July 2014
Variable
1
DOC6 Cond
7 pH Log Turb
8
CYANOBACTERIA
Log Total cyanobacterial biomass 0.45 0.71 0.76
Log Total Phytoplankton biomass 0.45 0.61 0.70
Log Anabaena flos-aquae biomass 0.51 0.55 0. 61
Log Anabaena spiroides biomass 0.63 0.40 0.36
Log Total Anabaena biomass 0.72 0.61 0.64
Log Aphanizomenon flos-aquae biomass 0.42 0.46
Log Microcystis sp. biomass 0.51 0.58
Log Microcystis aeruginosa biomass 0.63 0.59
Log Total Microcystis biomass 0.63 0.67
EUKARYOTIC ALGAE
Log Chlorella biomass 0.58 0.47
Log Cryptomonas biomass
Log Cyclotella biomass 0.55
Log Fragilaria biomass 0.53
Log Melosira biomass 0.42 0.40
Log Oocystis biomass
Log Péridinium biomass
Log Planktosphaeria biomass 0.97
Log Rhodomonas biomass
Log Scenedesmus biomass 0.53 0.36
Log Stephanodiscus biomass 0.46
Variable
1
Log NH3
9
Log NOx
10
TEMP SURF
11
Zeu12
(from Secchi)
DO13
CYANOBACTERIA
Log Total cyanobacterial biomass -0.83 0.25 -0.65
Log Total eukaryote biomass -0.19
Log Total phytoplankton biomass -0.70 -0.70
Log Anabaena flos-aquae biomass -0.64 0.24 -0.57
Log Anabaena spiroides biomass 0.46 -0.38 0.7
Log Total Anabaena biomass -0.67 0.29 -0.66
Log Aphanizomenon flos-aquae biomass -0.63 -0.52 0.46
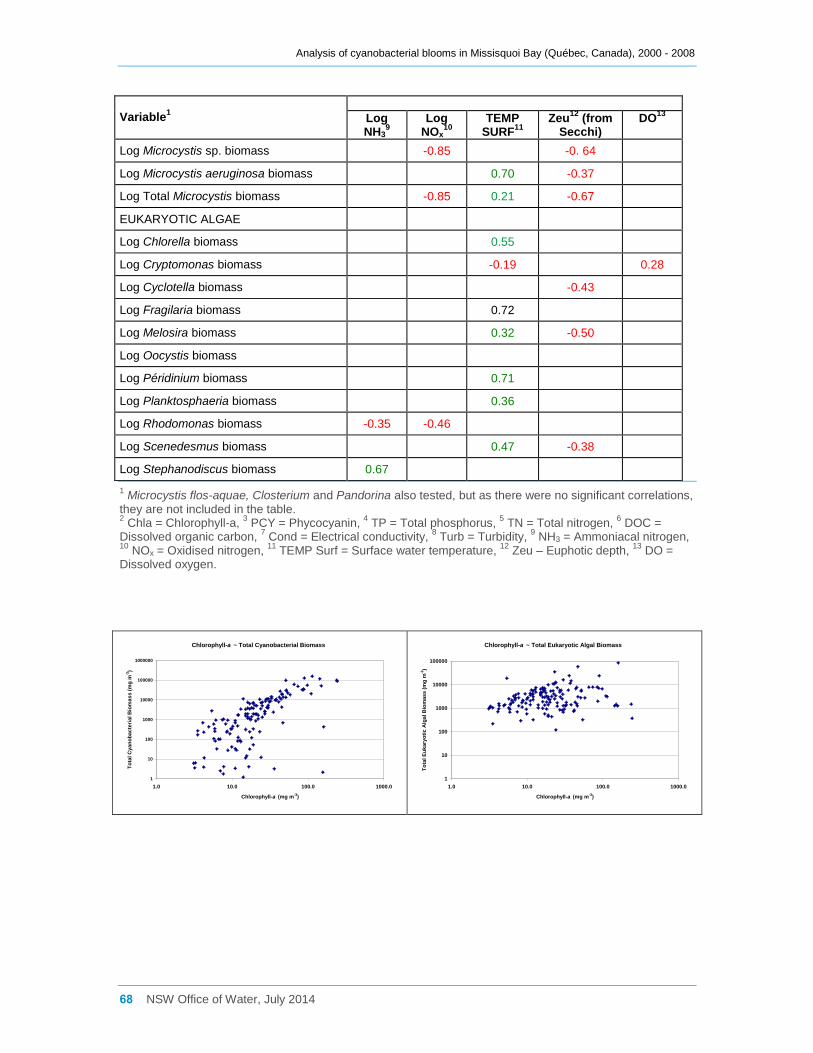
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
68 NSW Office of Water, July 2014
Variable
1
Log NH3
9
Log NOx
10
TEMP SURF
11
Zeu12
(from Secchi)
DO13
Log Microcystis sp. biomass -0.85 -0. 64
Log Microcystis aeruginosa biomass 0.70 -0.37
Log Total Microcystis biomass -0.85 0.21 -0.67
EUKARYOTIC ALGAE
Log Chlorella biomass 0.55
Log Cryptomonas biomass -0.19 0.28
Log Cyclotella biomass -0.43
Log Fragilaria biomass 0.72
Log Melosira biomass 0.32 -0.50
Log Oocystis biomass
Log Péridinium biomass 0.71
Log Planktosphaeria biomass 0.36
Log Rhodomonas biomass -0.35 -0.46
Log Scenedesmus biomass 0.47 -0.38
Log Stephanodiscus biomass 0.67
1 Microcystis flos-aquae, Closterium and Pandorina also tested, but as there were no significant correlations,
they are not included in the table. 2 Chla = Chlorophyll-a,
3 PCY = Phycocyanin,
4 TP = Total phosphorus,
5 TN = Total nitrogen,
6 DOC =
Dissolved organic carbon, 7 Cond = Electrical conductivity,
8 Turb = Turbidity,
9 NH3 = Ammoniacal nitrogen,
10 NOx = Oxidised nitrogen,
11 TEMP Surf = Surface water temperature,
12 Zeu – Euphotic depth,
13 DO =
Dissolved oxygen.
Chlorophyll-a ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
1.0 10.0 100.0 1000.0
Chlorophyll-a (mg m-3
)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
Chlorophyll-a ~ Total Eukaryotic Algal Biomass
1
10
100
1000
10000
100000
1.0 10.0 100.0 1000.0
Chlorophyll-a (mg m-3
)
To
tal
Eu
ka
ryo
tic
Alg
al
Bio
ma
ss
(m
g m
-3)
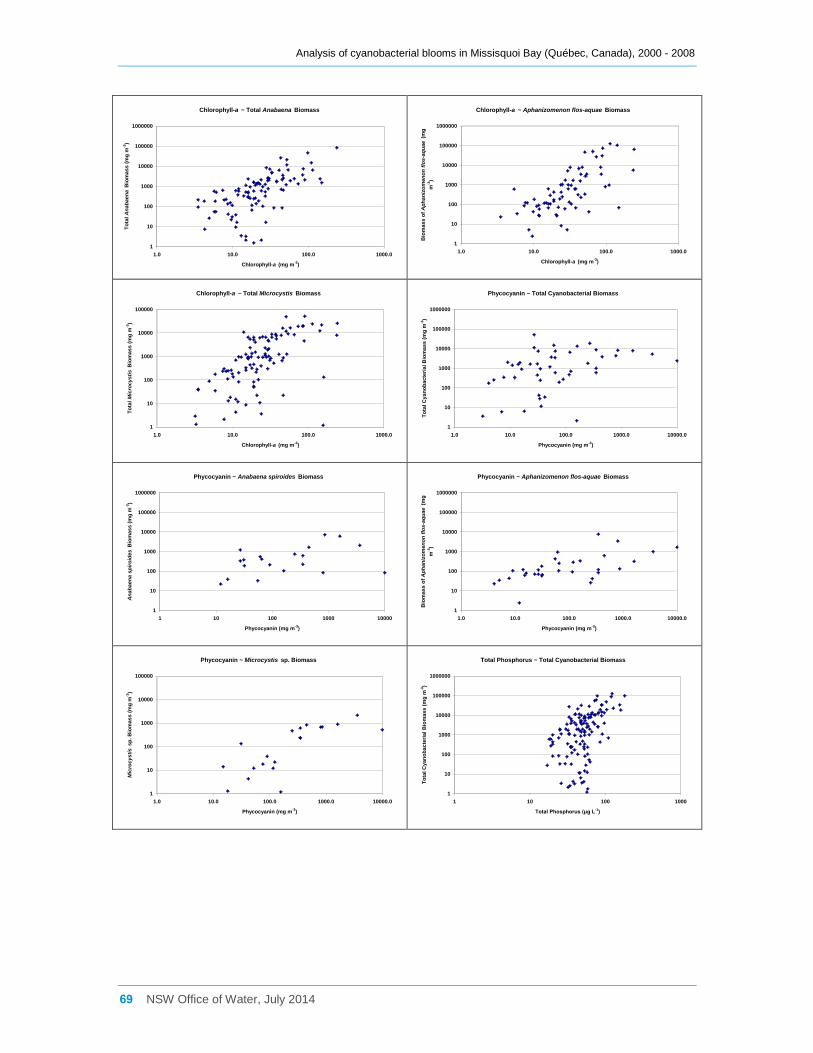
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
69 NSW Office of Water, July 2014
Chlorophyll-a ~ Total Anabaena Biomass
1
10
100
1000
10000
100000
1000000
1.0 10.0 100.0 1000.0
Chlorophyll-a (mg m-3
)
To
tal
An
ab
ae
na
Bio
ma
ss
(m
g m
-3)
Chlorophyll-a ~ Aphanizomenon flos-aquae Biomass
1
10
100
1000
10000
100000
1000000
1.0 10.0 100.0 1000.0
Chlorophyll-a (mg m-3
)
Bio
ma
ss
of
Ap
ha
niz
om
en
on
flo
s-a
qu
ae
(m
g
m-3
)
Chlorophyll-a ~ Total Microcystis Biomass
1
10
100
1000
10000
100000
1.0 10.0 100.0 1000.0
Chlorophyll-a (mg m-3
)
To
tal
Mic
roc
ys
tis
Bio
ma
ss
(m
g m
-3)
Phycocyanin ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
1.0 10.0 100.0 1000.0 10000.0
Phycocyanin (mg m-3
)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
Phycocyanin ~ Anabaena spiroides Biomass
1
10
100
1000
10000
100000
1000000
1 10 100 1000 10000
Phycocyanin (mg m-3
)
An
ab
ae
na
sp
iro
ide
s B
iom
as
s (
mg
m-3
)
Phycocyanin ~ Aphanizomenon flos-aquae Biomass
1
10
100
1000
10000
100000
1000000
1.0 10.0 100.0 1000.0 10000.0
Phycocyanin (mg m-3
)
Bio
ma
ss
of
Ap
ha
niz
om
en
on
flo
s-a
qu
ae
(m
g
m-3
)
Phycocyanin ~ Microcystis sp. Biomass
1
10
100
1000
10000
100000
1.0 10.0 100.0 1000.0 10000.0
Phycocyanin (mg m-3
)
Mic
roc
ys
tis
sp
. B
iom
as
s (
mg
m-3
)
Total Phosphorus ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
1 10 100 1000
Total Phosphorus (μg L-1
)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
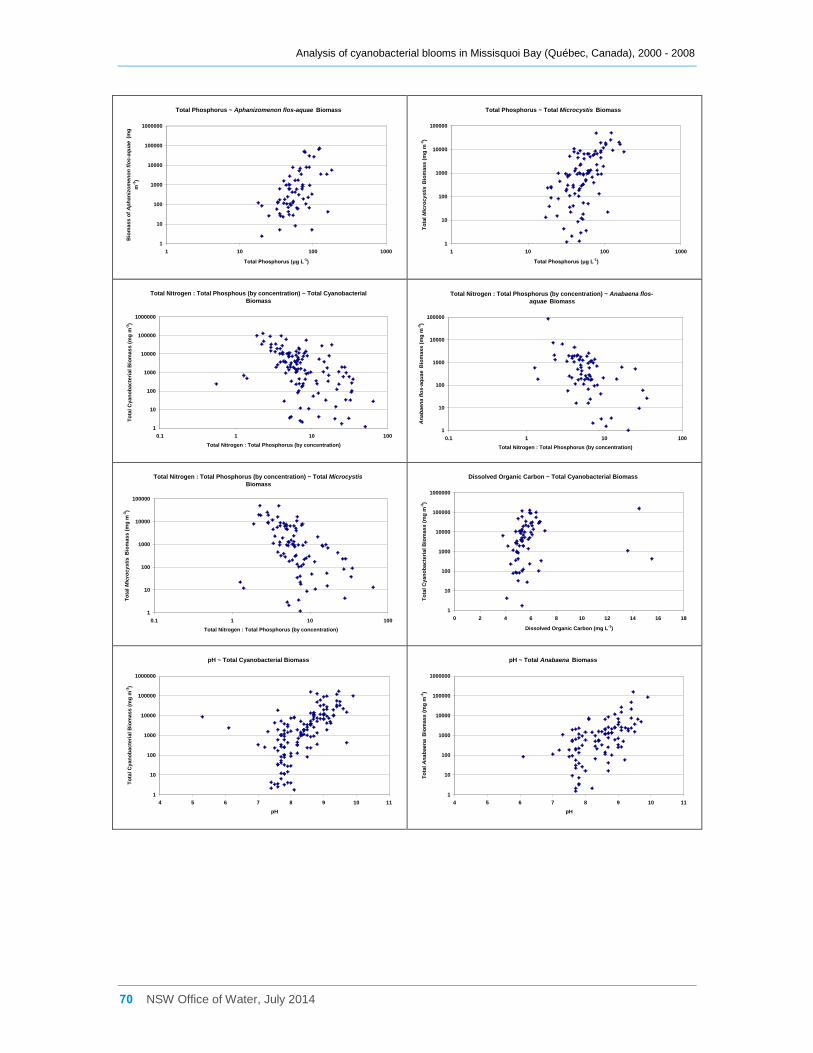
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
70 NSW Office of Water, July 2014
Total Phosphorus ~ Aphanizomenon flos-aquae Biomass
1
10
100
1000
10000
100000
1000000
1 10 100 1000
Total Phosphorus (μg L-1
)
Bio
ma
ss
of
Ap
ha
niz
om
en
on
flo
s-a
qu
ae
(m
g
m-3
)
Total Phosphorus ~ Total Microcystis Biomass
1
10
100
1000
10000
100000
1 10 100 1000
Total Phosphorus (μg L-1
)
To
tal
Mic
rocysti
s B
iom
ass (
mg
m-3
)
Total Nitrogen : Total Phosphous (by concentration) ~ Total Cyanobacterial
Biomass
1
10
100
1000
10000
100000
1000000
0.1 1 10 100
Total Nitrogen : Total Phosphorus (by concentration)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
Total Nitrogen : Total Phosphorus (by concentration) ~ Anabaena flos-
aquae Biomass
1
10
100
1000
10000
100000
0.1 1 10 100
Total Nitrogen : Total Phosphorus (by concentration)
An
ab
ae
na
flo
s-a
qu
ae
Bio
ma
ss
(m
g m
-3)
Total Nitrogen : Total Phosphorus (by concentration) ~ Total Microcystis
Biomass
1
10
100
1000
10000
100000
0.1 1 10 100
Total Nitrogen : Total Phosphorus (by concentration)
To
tal
Mic
roc
ys
tis
Bio
ma
ss
(m
g m
-3)
Dissolved Organic Carbon ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
0 2 4 6 8 10 12 14 16 18
Dissolved Organic Carbon (mg L-1
)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
pH ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
4 5 6 7 8 9 10 11
pH
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
pH ~ Total Anabaena Biomass
1
10
100
1000
10000
100000
1000000
4 5 6 7 8 9 10 11
pH
To
tal
An
ab
ae
na
Bio
ma
ss
(m
g m
-3)
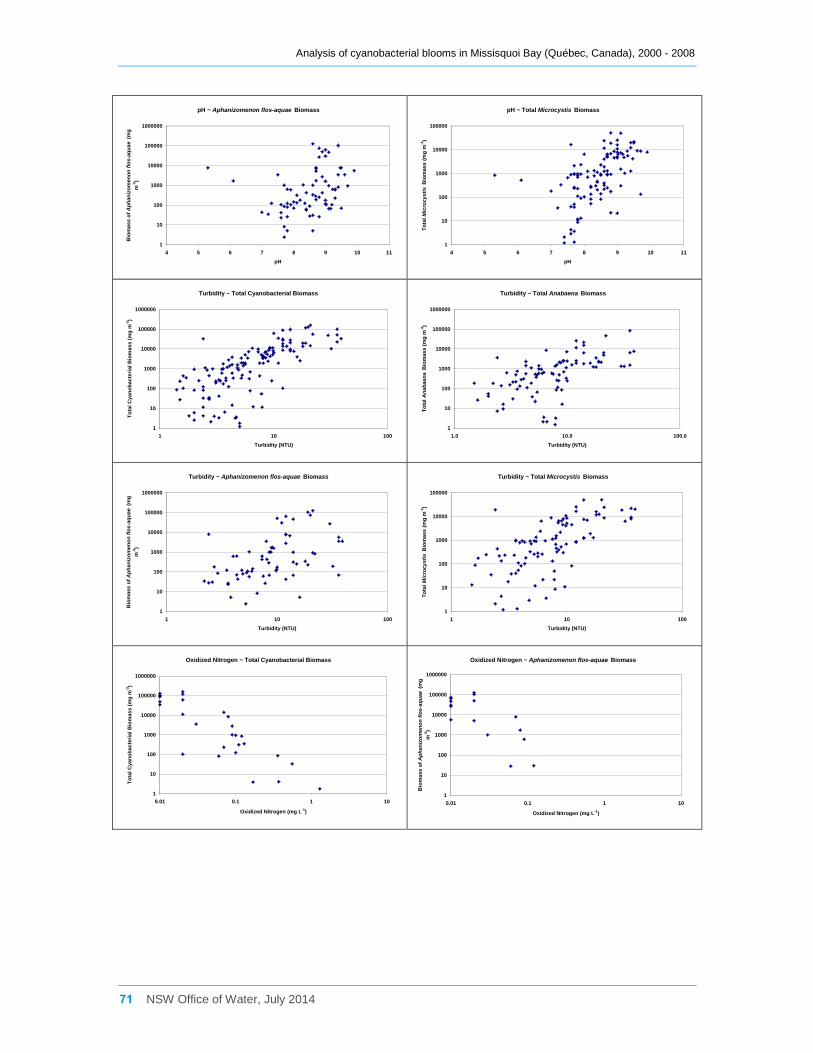
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
71 NSW Office of Water, July 2014
pH ~ Aphanizomenon flos-aquae Biomass
1
10
100
1000
10000
100000
1000000
4 5 6 7 8 9 10 11
pH
Bio
ma
ss
of
Ap
ha
niz
om
en
on
flo
s-a
qu
ae
(m
g
m-3
)
pH ~ Total Microcystis Biomass
1
10
100
1000
10000
100000
4 5 6 7 8 9 10 11
pH
To
tal
Mic
roc
ys
tis
Bio
ma
ss
(m
g m
-3)
Turbidity ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
1 10 100
Turbidity (NTU)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
Turbidity ~ Total Anabaena Biomass
1
10
100
1000
10000
100000
1000000
1.0 10.0 100.0
Turbidity (NTU)
To
tal
An
ab
ae
na
Bio
ma
ss
(m
g m
-3)
Turbidity ~ Aphanizomenon flos-aquae Biomass
1
10
100
1000
10000
100000
1000000
1 10 100
Turbidity (NTU)
Bio
ma
ss
of
Ap
ha
niz
om
en
on
flo
s-a
qu
ae
(m
g
m-3
)
Turbidity ~ Total Microcystis Biomass
1
10
100
1000
10000
100000
1 10 100
Turbidity (NTU)
To
tal
Mic
roc
ys
tis
Bio
ma
ss
(m
g m
-3)
Oxidized Nitrogen ~ Total Cyanobacterial Biomass
1
10
100
1000
10000
100000
1000000
0.01 0.1 1 10
Oxidized Nitrogen (mg L-1
)
To
tal
Cy
an
ob
ac
teri
al
Bio
ma
ss
(m
g m
-3)
Oxidized Nitrogen ~ Aphanizomenon flos-aquae Biomass
1
10
100
1000
10000
100000
1000000
0.01 0.1 1 10
Oxidized Nitrogen (mg L-1
)
Bio
ma
ss
of
Ap
ha
niz
om
en
on
flo
s-a
qu
ae
(m
g
m-3
)
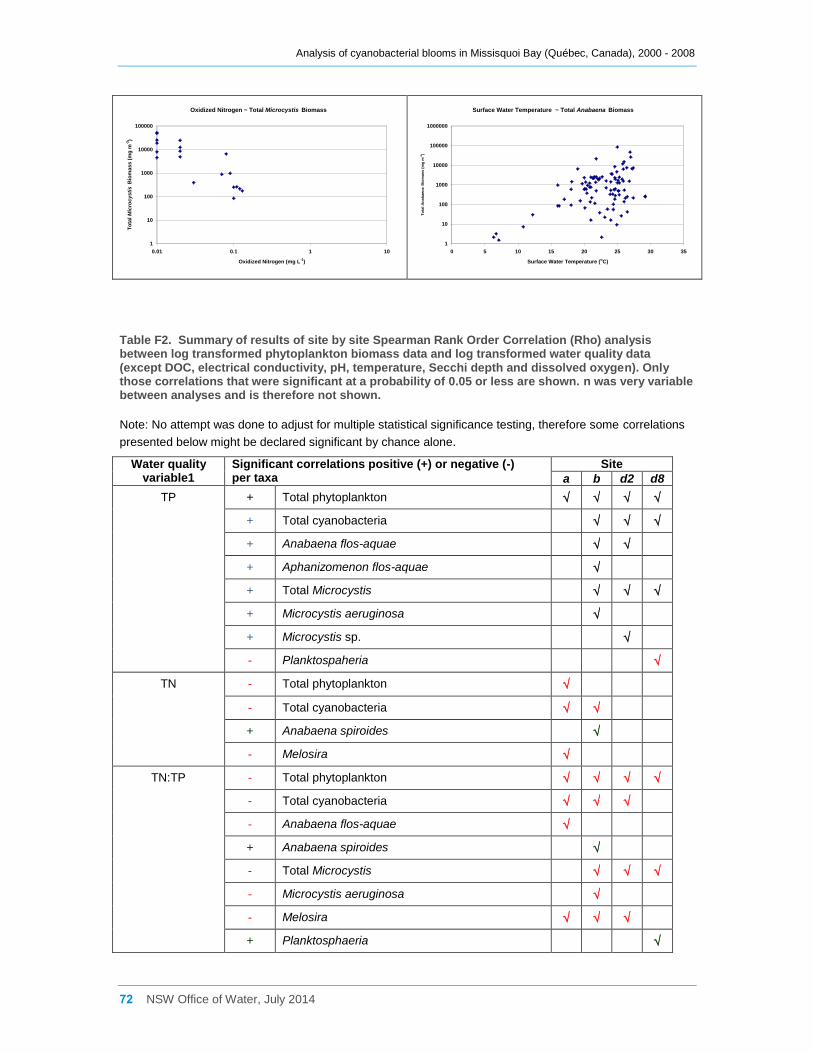
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
72 NSW Office of Water, July 2014
Oxidized Nitrogen ~ Total Microcystis Biomass
1
10
100
1000
10000
100000
0.01 0.1 1 10
Oxidized Nitrogen (mg L-1
)
To
tal
Mic
roc
ys
tis
Bio
ma
ss
(m
g m
-3)
Surface Water Temperature ~ Total Anabaena Biomass
1
10
100
1000
10000
100000
1000000
0 5 10 15 20 25 30 35
Surface Water Temperature (oC)
To
tal
An
ab
aen
a B
iom
ass (
mg
m-3
)
Table F2. Summary of results of site by site Spearman Rank Order Correlation (Rho) analysis between log transformed phytoplankton biomass data and log transformed water quality data (except DOC, electrical conductivity, pH, temperature, Secchi depth and dissolved oxygen). Only those correlations that were significant at a probability of 0.05 or less are shown. n was very variable between analyses and is therefore not shown.
Note: No attempt was done to adjust for multiple statistical significance testing, therefore some correlations
presented below might be declared significant by chance alone.
Water quality variable1
Significant correlations positive (+) or negative (-) per taxa
Site
a b d2 d8
TP + Total phytoplankton
+ Total cyanobacteria
+ Anabaena flos-aquae
+ Aphanizomenon flos-aquae
+ Total Microcystis
+ Microcystis aeruginosa
+ Microcystis sp.
- Planktospaheria
TN - Total phytoplankton
- Total cyanobacteria
+ Anabaena spiroides
- Melosira
TN:TP - Total phytoplankton
- Total cyanobacteria
- Anabaena flos-aquae
+ Anabaena spiroides
- Total Microcystis
- Microcystis aeruginosa
- Melosira
+ Planktosphaeria
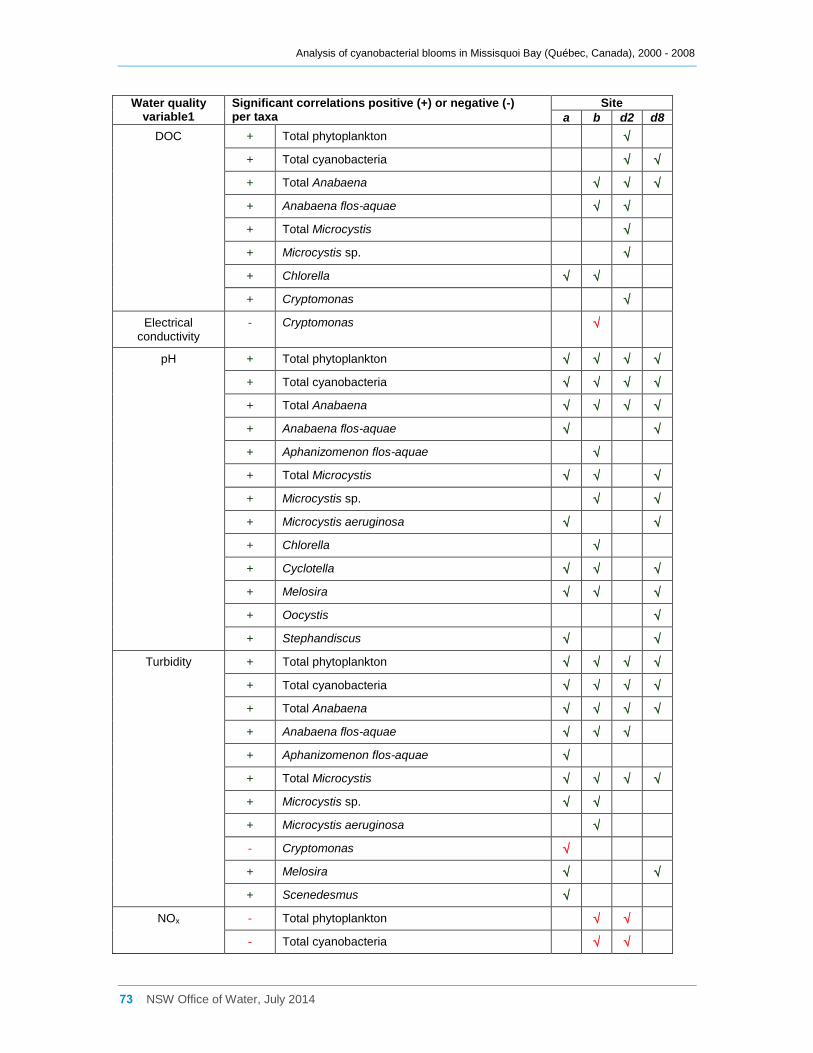
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
73 NSW Office of Water, July 2014
Water quality variable1
Significant correlations positive (+) or negative (-) per taxa
Site
a b d2 d8
DOC + Total phytoplankton
+ Total cyanobacteria
+ Total Anabaena
+ Anabaena flos-aquae
+ Total Microcystis
+ Microcystis sp.
+ Chlorella
+ Cryptomonas
Electrical conductivity
- Cryptomonas
pH + Total phytoplankton
+ Total cyanobacteria
+ Total Anabaena
+ Anabaena flos-aquae
+ Aphanizomenon flos-aquae
+ Total Microcystis
+ Microcystis sp.
+ Microcystis aeruginosa
+ Chlorella
+ Cyclotella
+ Melosira
+ Oocystis
+ Stephandiscus
Turbidity + Total phytoplankton
+ Total cyanobacteria
+ Total Anabaena
+ Anabaena flos-aquae
+ Aphanizomenon flos-aquae
+ Total Microcystis
+ Microcystis sp.
+ Microcystis aeruginosa
- Cryptomonas
+ Melosira
+ Scenedesmus
NOx - Total phytoplankton
- Total cyanobacteria
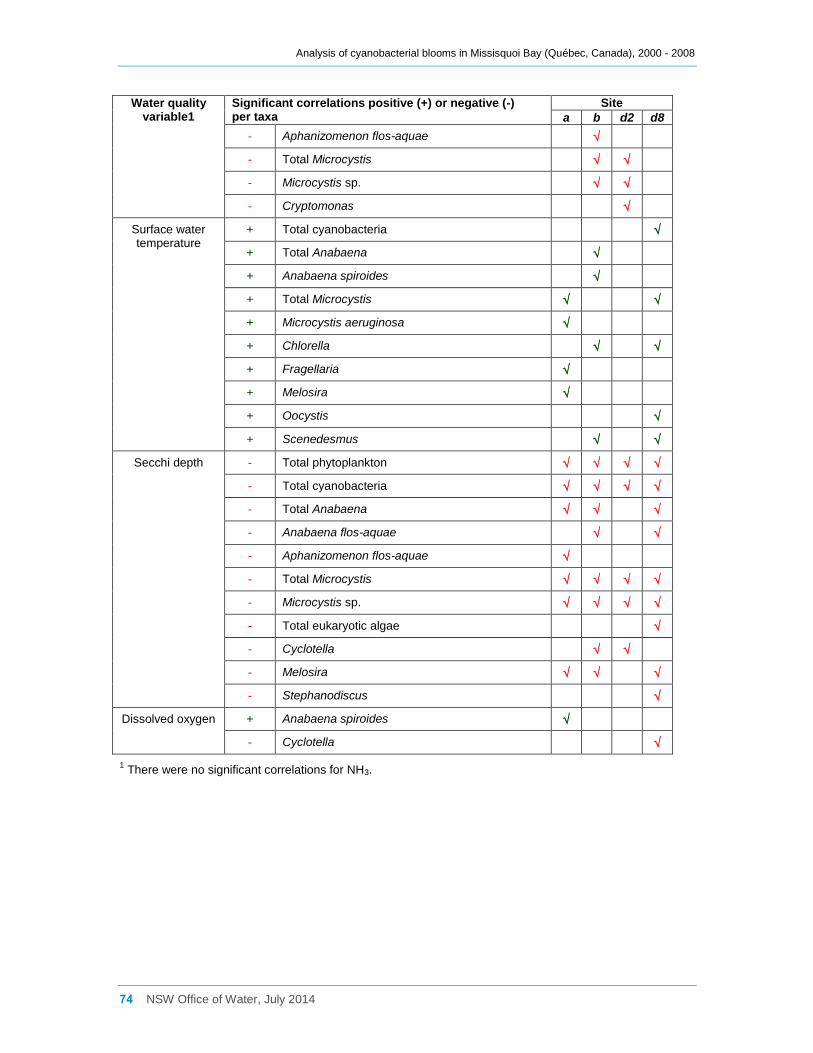
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
74 NSW Office of Water, July 2014
Water quality variable1
Significant correlations positive (+) or negative (-) per taxa
Site
a b d2 d8
- Aphanizomenon flos-aquae
- Total Microcystis
- Microcystis sp.
- Cryptomonas
Surface water temperature
+ Total cyanobacteria
+ Total Anabaena
+ Anabaena spiroides
+ Total Microcystis
+ Microcystis aeruginosa
+ Chlorella
+ Fragellaria
+ Melosira
+ Oocystis
+ Scenedesmus
Secchi depth - Total phytoplankton
- Total cyanobacteria
- Total Anabaena
- Anabaena flos-aquae
- Aphanizomenon flos-aquae
- Total Microcystis
- Microcystis sp.
- Total eukaryotic algae
- Cyclotella
- Melosira
- Stephanodiscus
Dissolved oxygen + Anabaena spiroides
- Cyclotella
1 There were no significant correlations for NH3.
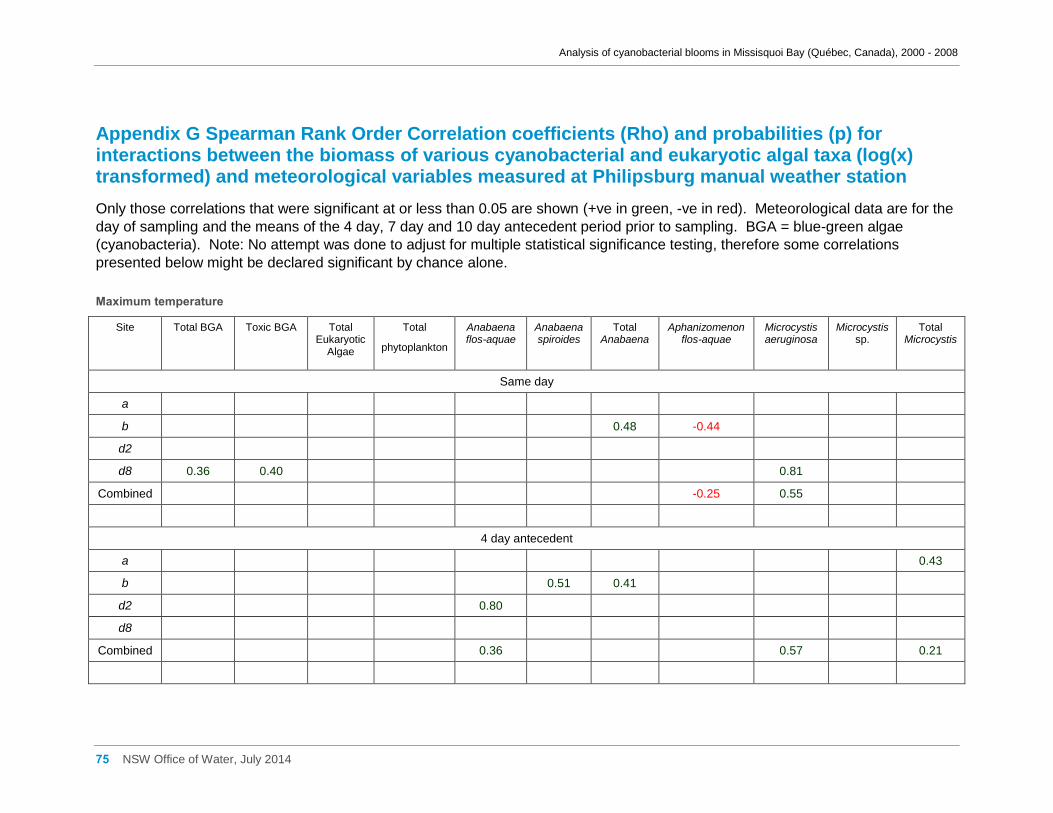
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
75 NSW Office of Water, July 2014
Appendix G Spearman Rank Order Correlation coefficients (Rho) and probabilities (p) for interactions between the biomass of various cyanobacterial and eukaryotic algal taxa (log(x) transformed) and meteorological variables measured at Philipsburg manual weather station
Only those correlations that were significant at or less than 0.05 are shown (+ve in green, -ve in red). Meteorological data are for the
day of sampling and the means of the 4 day, 7 day and 10 day antecedent period prior to sampling. BGA = blue-green algae
(cyanobacteria). Note: No attempt was done to adjust for multiple statistical significance testing, therefore some correlations
presented below might be declared significant by chance alone.
Maximum temperature
Site Total BGA Toxic BGA Total Eukaryotic
Algae
Total
phytoplankton
Anabaena flos-aquae
Anabaena spiroides
Total Anabaena
Aphanizomenon flos-aquae
Microcystis aeruginosa
Microcystis sp.
Total Microcystis
Same day
a
b 0.48 -0.44
d2
d8 0.36 0.40 0.81
Combined -0.25 0.55
4 day antecedent
a 0.43
b 0.51 0.41
d2 0.80
d8
Combined 0.36 0.57 0.21
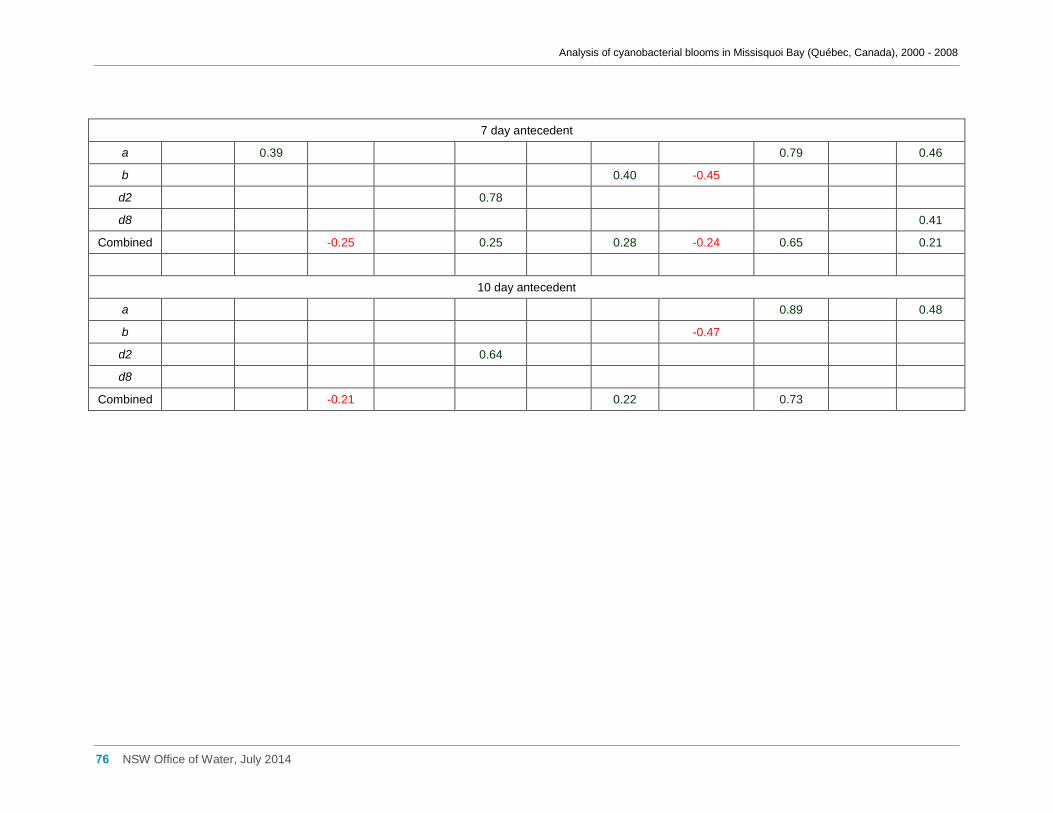
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
76 NSW Office of Water, July 2014
7 day antecedent
a 0.39 0.79 0.46
b 0.40 -0.45
d2 0.78
d8 0.41
Combined -0.25 0.25 0.28 -0.24 0.65 0.21
10 day antecedent
a 0.89 0.48
b -0.47
d2 0.64
d8
Combined -0.21 0.22 0.73
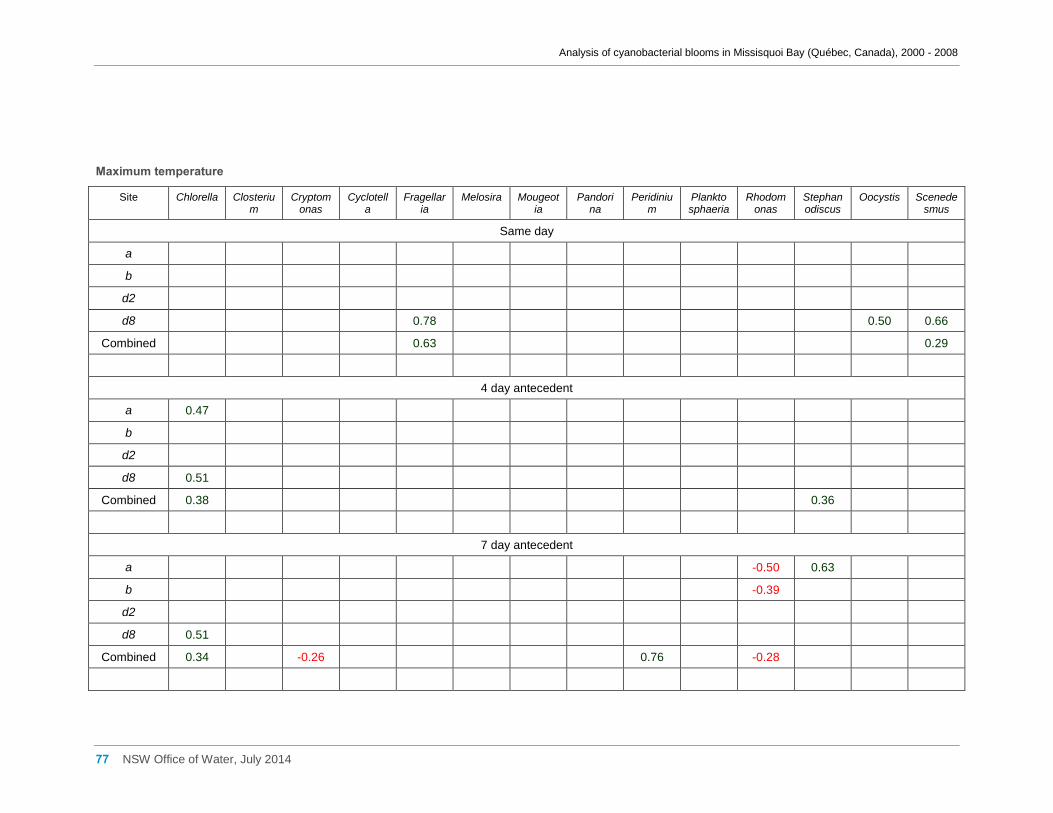
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
77 NSW Office of Water, July 2014
Maximum temperature
Site Chlorella Closterium
Cryptomonas
Cyclotella
Fragellaria
Melosira Mougeotia
Pandorina
Peridinium
Plankto sphaeria
Rhodomonas
Stephanodiscus
Oocystis Scenedesmus
Same day
a
b
d2
d8 0.78 0.50 0.66
Combined 0.63 0.29
4 day antecedent
a 0.47
b
d2
d8 0.51
Combined 0.38 0.36
7 day antecedent
a -0.50 0.63
b -0.39
d2
d8 0.51
Combined 0.34 -0.26 0.76 -0.28
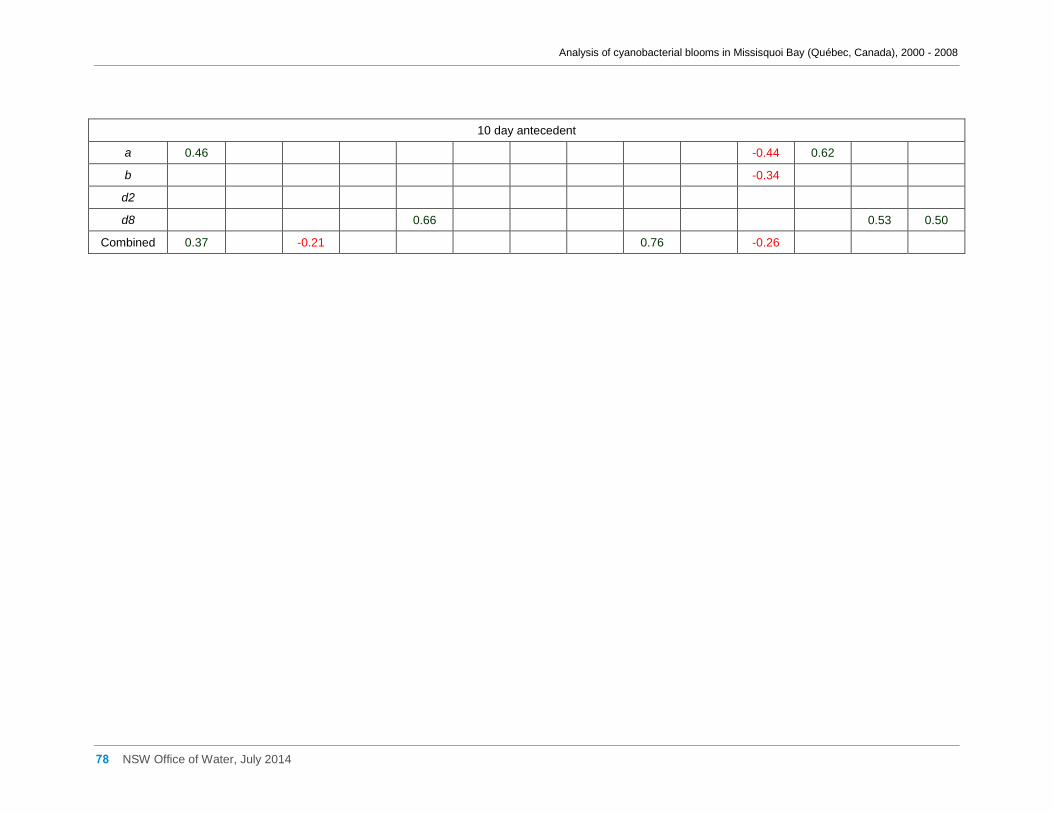
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
78 NSW Office of Water, July 2014
10 day antecedent
a 0.46 -0.44 0.62
b -0.34
d2
d8 0.66 0.53 0.50
Combined 0.37 -0.21 0.76 -0.26
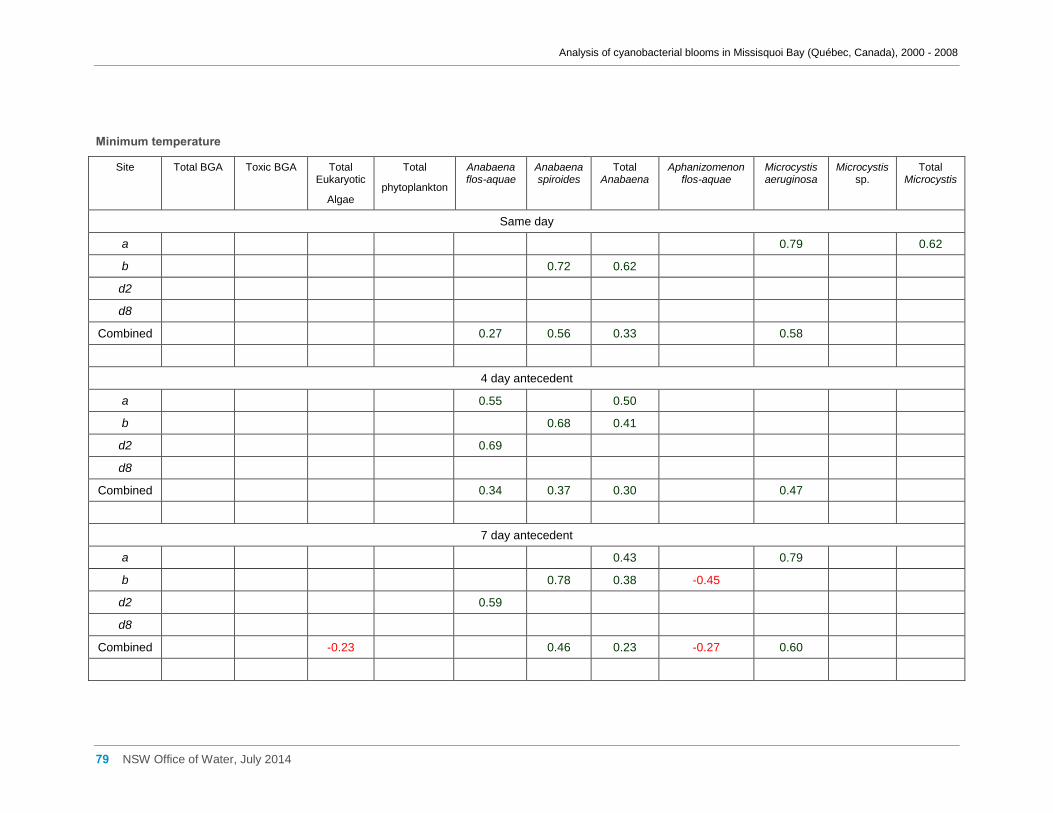
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
79 NSW Office of Water, July 2014
Minimum temperature
Site Total BGA Toxic BGA Total Eukaryotic
Algae
Total
phytoplankton
Anabaena flos-aquae
Anabaena spiroides
Total Anabaena
Aphanizomenon flos-aquae
Microcystis aeruginosa
Microcystis sp.
Total Microcystis
Same day
a 0.79 0.62
b 0.72 0.62
d2
d8
Combined 0.27 0.56 0.33 0.58
4 day antecedent
a 0.55 0.50
b 0.68 0.41
d2 0.69
d8
Combined 0.34 0.37 0.30 0.47
7 day antecedent
a 0.43 0.79
b 0.78 0.38 -0.45
d2 0.59
d8
Combined -0.23 0.46 0.23 -0.27 0.60
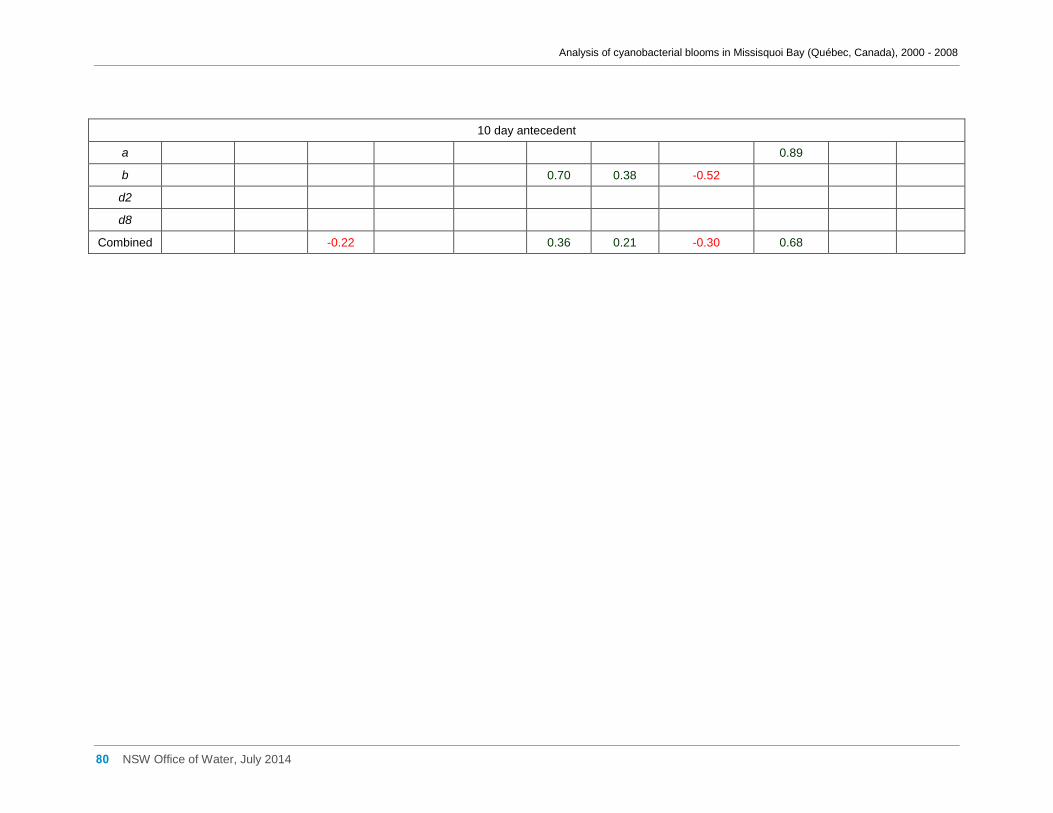
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
80 NSW Office of Water, July 2014
10 day antecedent
a 0.89
b 0.70 0.38 -0.52
d2
d8
Combined -0.22 0.36 0.21 -0.30 0.68
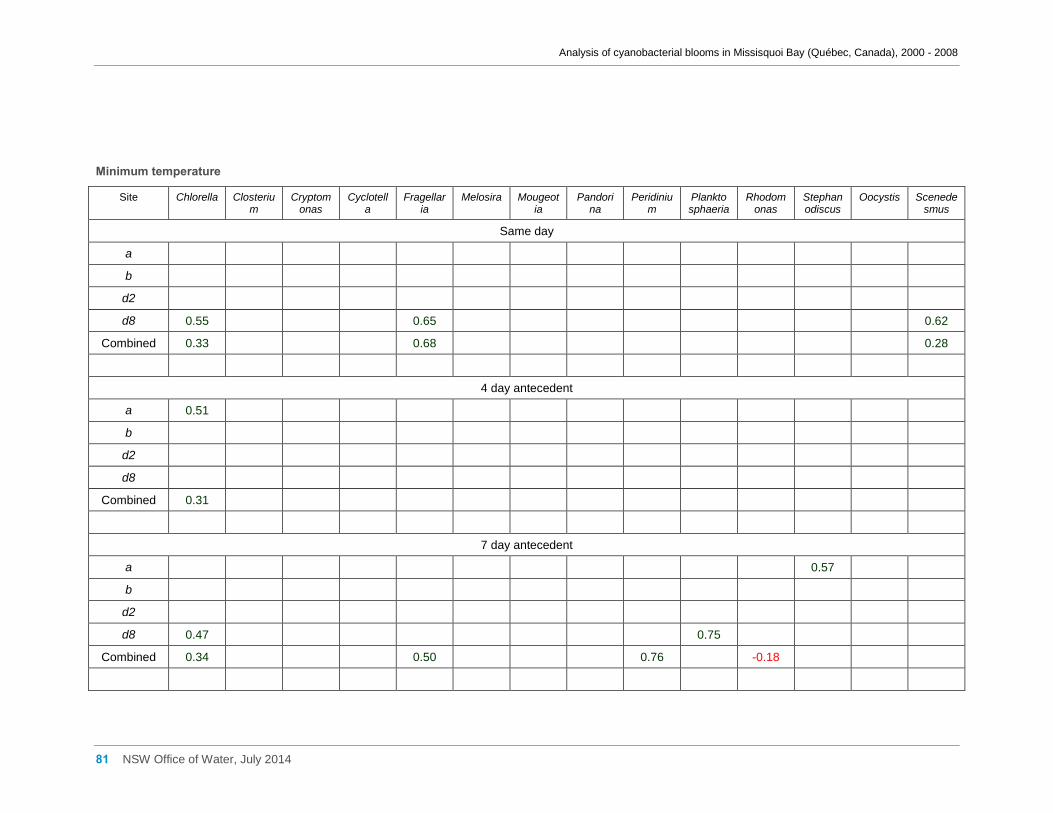
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
81 NSW Office of Water, July 2014
Minimum temperature
Site Chlorella Closterium
Cryptomonas
Cyclotella
Fragellaria
Melosira Mougeotia
Pandorina
Peridinium
Plankto sphaeria
Rhodomonas
Stephanodiscus
Oocystis Scenedesmus
Same day
a
b
d2
d8 0.55 0.65 0.62
Combined 0.33 0.68 0.28
4 day antecedent
a 0.51
b
d2
d8
Combined 0.31
7 day antecedent
a 0.57
b
d2
d8 0.47 0.75
Combined 0.34 0.50 0.76 -0.18
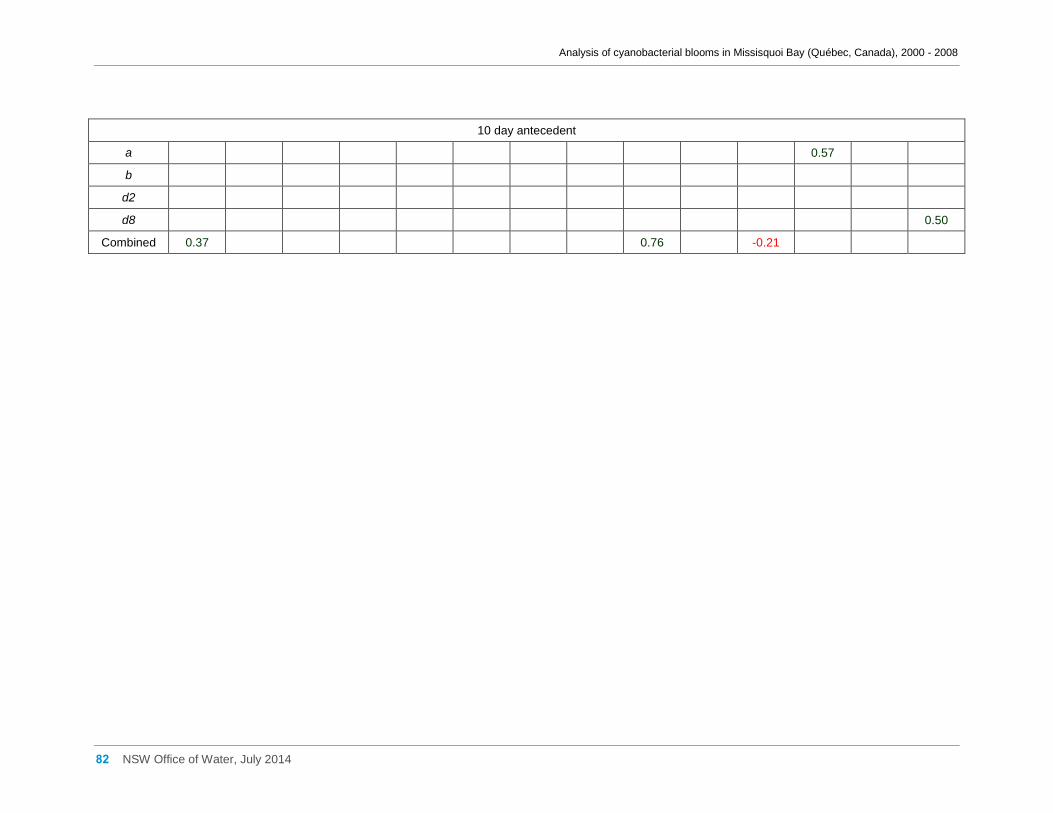
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
82 NSW Office of Water, July 2014
10 day antecedent
a 0.57
b
d2
d8 0.50
Combined 0.37 0.76 -0.21
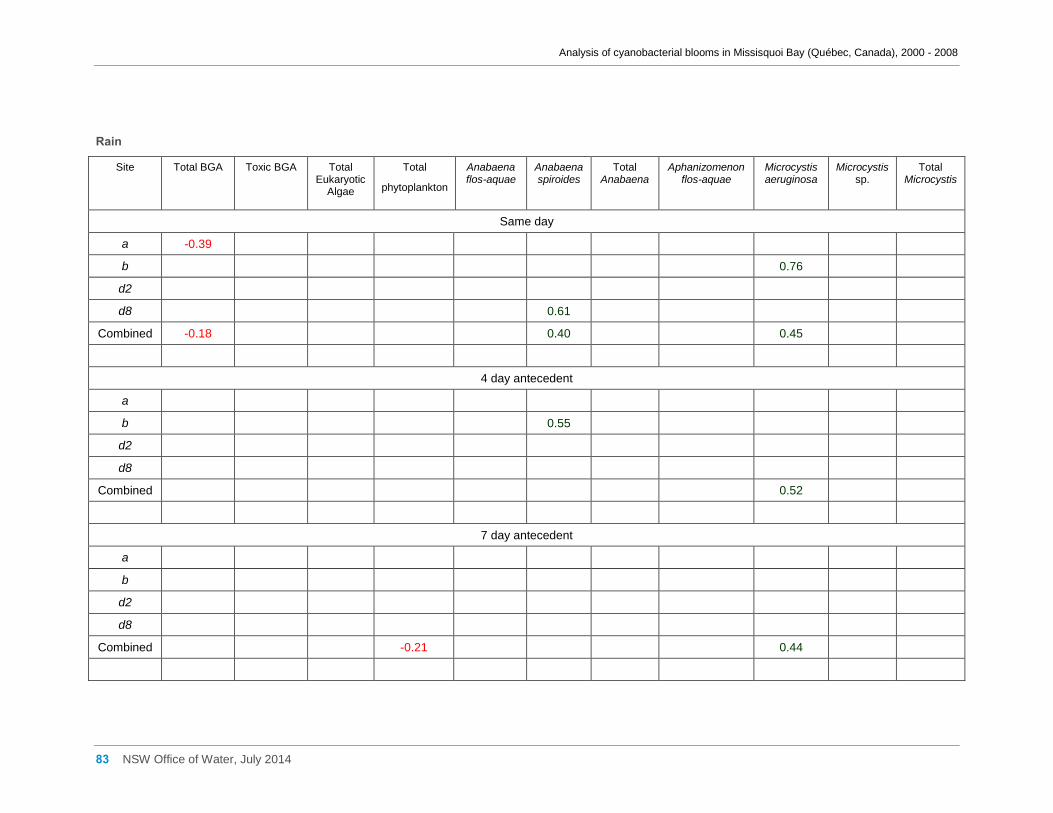
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
83 NSW Office of Water, July 2014
Rain
Site Total BGA Toxic BGA Total Eukaryotic
Algae
Total
phytoplankton
Anabaena flos-aquae
Anabaena spiroides
Total Anabaena
Aphanizomenon flos-aquae
Microcystis aeruginosa
Microcystis sp.
Total Microcystis
Same day
a -0.39
b 0.76
d2
d8 0.61
Combined -0.18 0.40 0.45
4 day antecedent
a
b 0.55
d2
d8
Combined 0.52
7 day antecedent
a
b
d2
d8
Combined -0.21 0.44
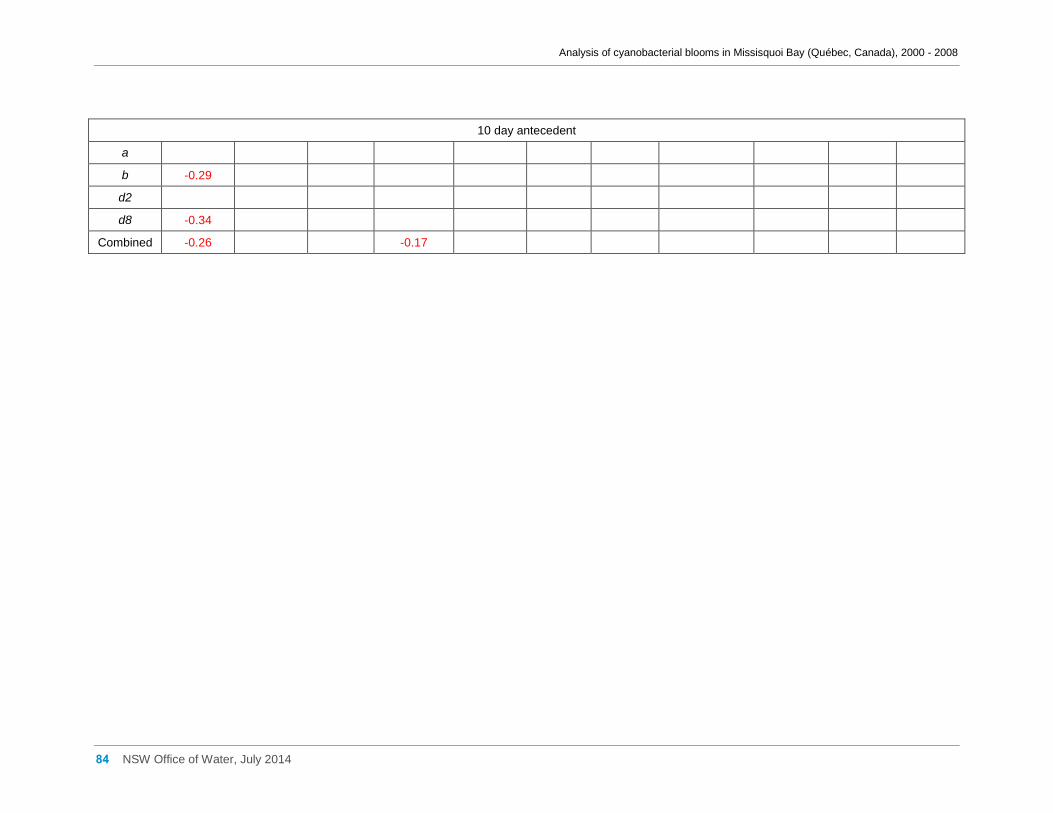
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
84 NSW Office of Water, July 2014
10 day antecedent
a
b -0.29
d2
d8 -0.34
Combined -0.26 -0.17
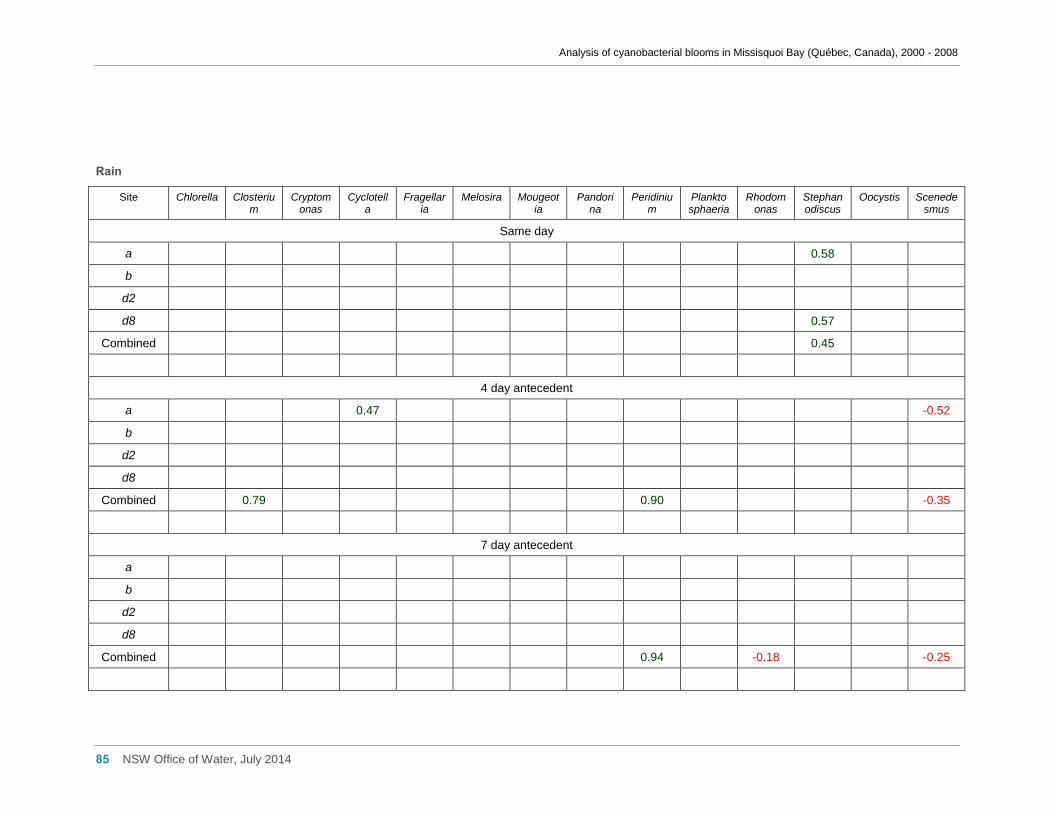
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
85 NSW Office of Water, July 2014
Rain
Site Chlorella Closterium
Cryptomonas
Cyclotella
Fragellaria
Melosira Mougeotia
Pandorina
Peridinium
Plankto sphaeria
Rhodomonas
Stephanodiscus
Oocystis Scenedesmus
Same day
a 0.58
b
d2
d8 0.57
Combined 0.45
4 day antecedent
a 0.47 -0.52
b
d2
d8
Combined 0.79 0.90 -0.35
7 day antecedent
a
b
d2
d8
Combined 0.94 -0.18 -0.25
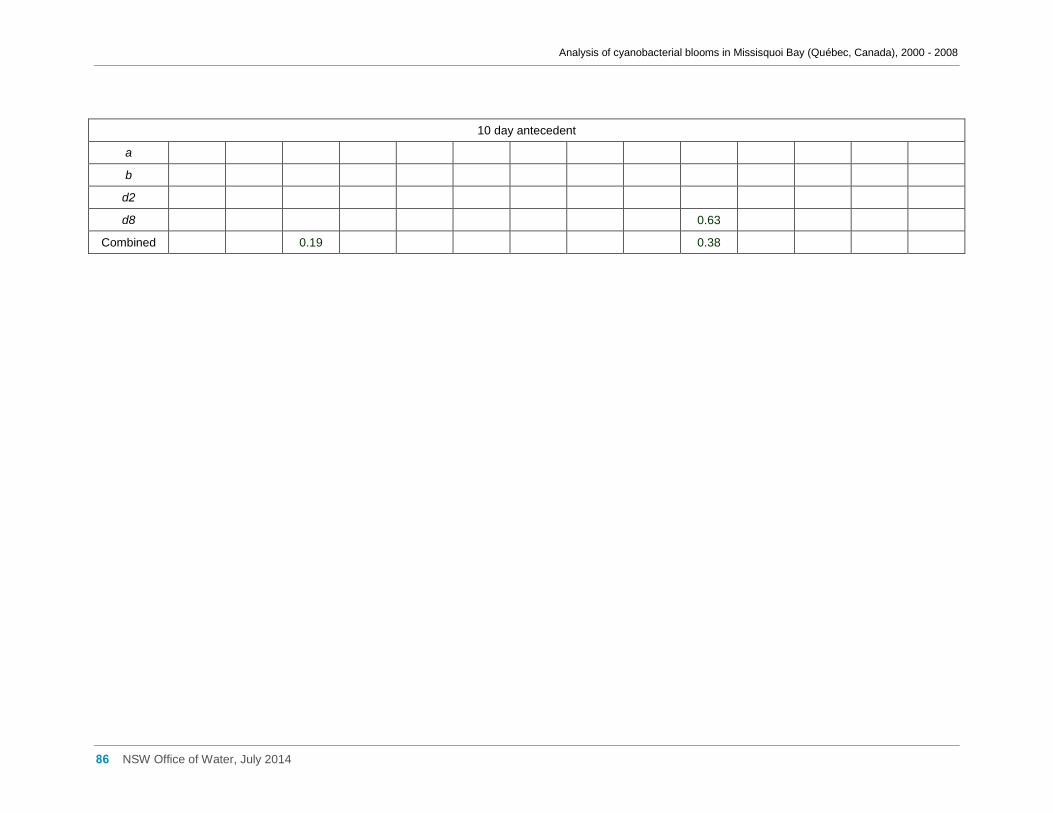
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
86 NSW Office of Water, July 2014
10 day antecedent
a
b
d2
d8 0.63
Combined 0.19 0.38
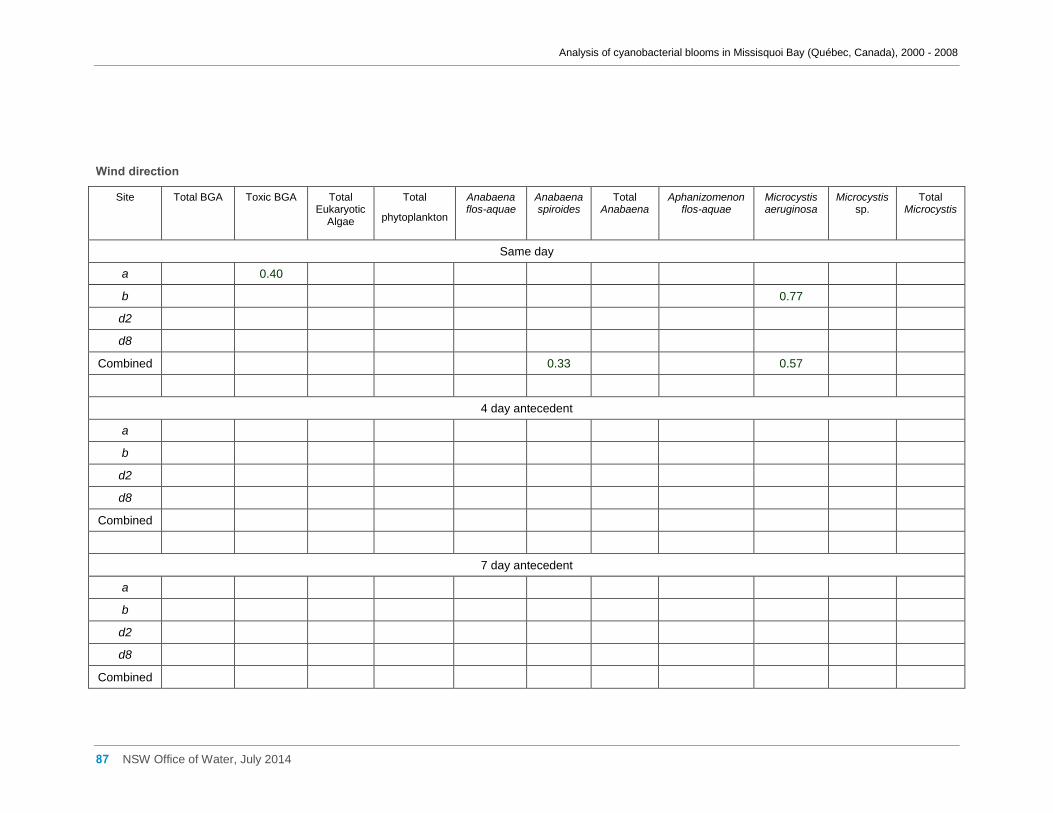
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
87 NSW Office of Water, July 2014
Wind direction
Site Total BGA Toxic BGA Total Eukaryotic
Algae
Total
phytoplankton
Anabaena flos-aquae
Anabaena spiroides
Total Anabaena
Aphanizomenon flos-aquae
Microcystis aeruginosa
Microcystis sp.
Total Microcystis
Same day
a 0.40
b 0.77
d2
d8
Combined 0.33 0.57
4 day antecedent
a
b
d2
d8
Combined
7 day antecedent
a
b
d2
d8
Combined
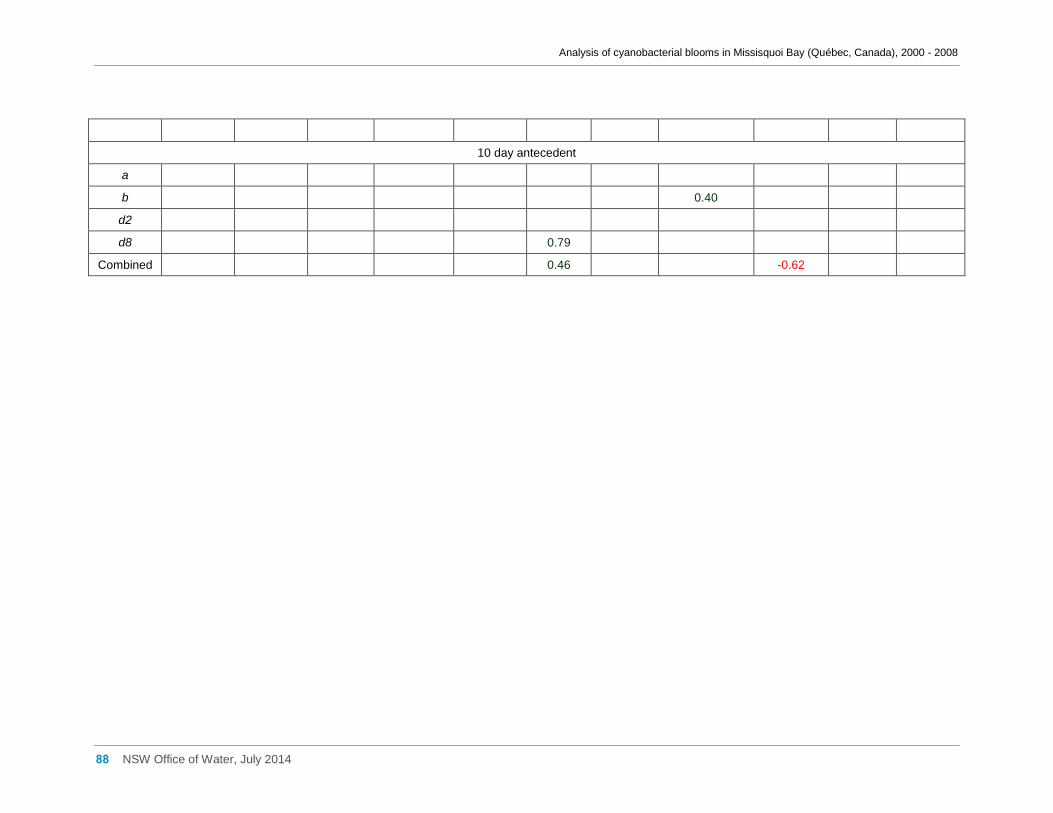
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
88 NSW Office of Water, July 2014
10 day antecedent
a
b 0.40
d2
d8 0.79
Combined 0.46 -0.62
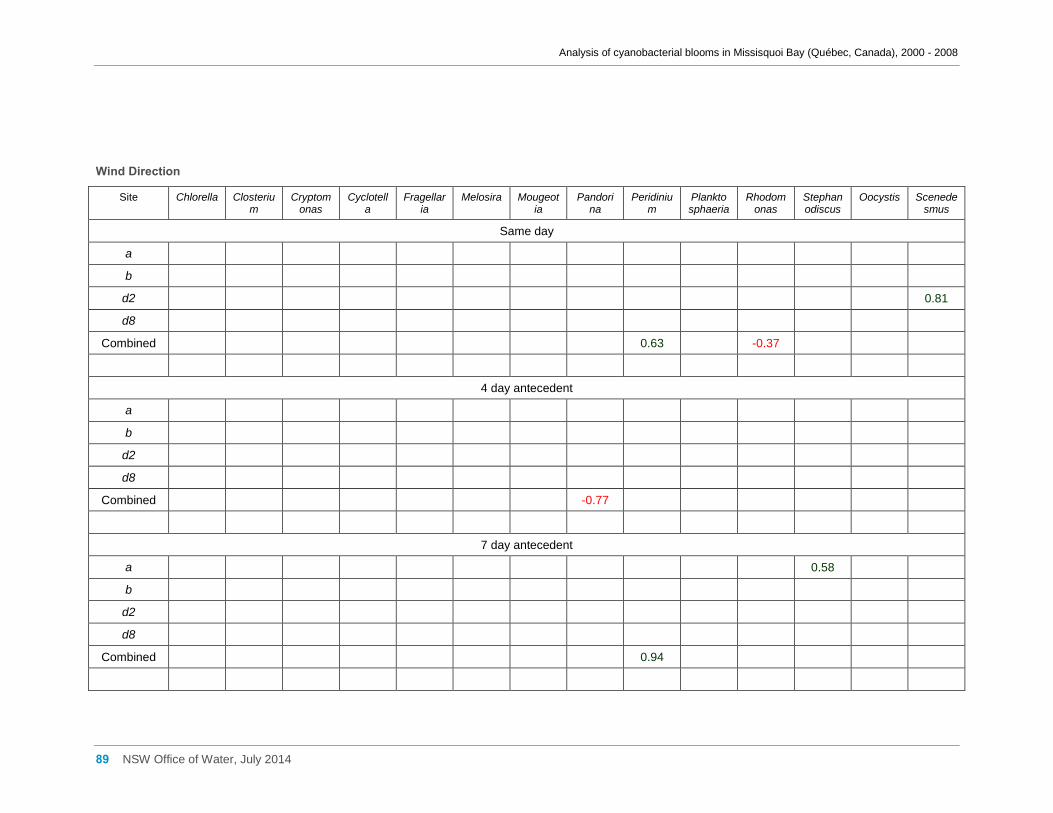
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
89 NSW Office of Water, July 2014
Wind Direction
Site Chlorella Closterium
Cryptomonas
Cyclotella
Fragellaria
Melosira Mougeotia
Pandorina
Peridinium
Plankto sphaeria
Rhodomonas
Stephanodiscus
Oocystis Scenedesmus
Same day
a
b
d2 0.81
d8
Combined 0.63 -0.37
4 day antecedent
a
b
d2
d8
Combined -0.77
7 day antecedent
a 0.58
b
d2
d8
Combined 0.94
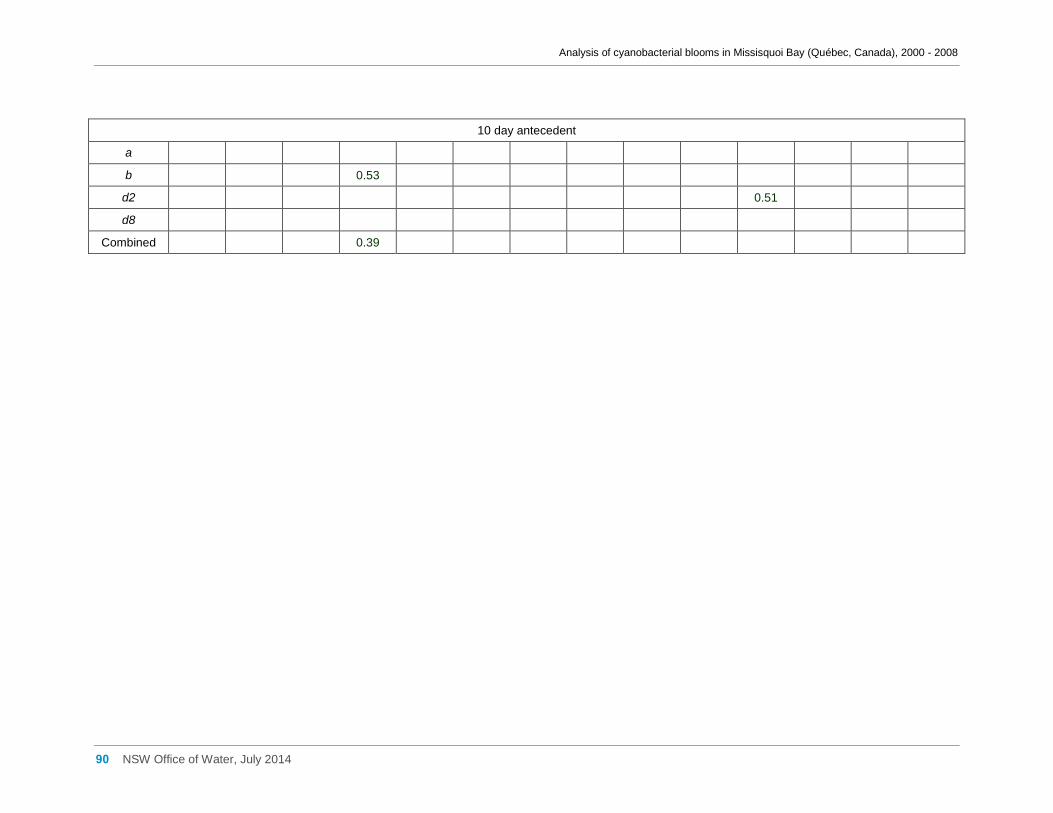
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
90 NSW Office of Water, July 2014
10 day antecedent
a
b 0.53
d2 0.51
d8
Combined 0.39
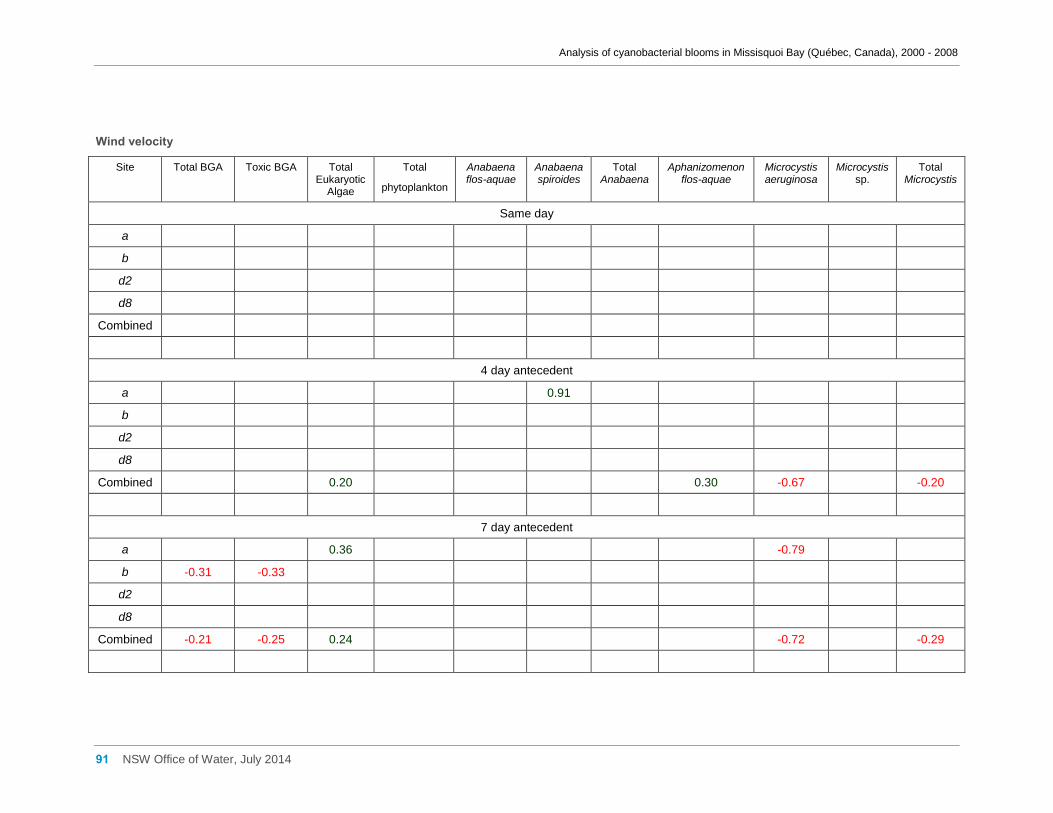
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
91 NSW Office of Water, July 2014
Wind velocity
Site Total BGA Toxic BGA Total Eukaryotic
Algae
Total
phytoplankton
Anabaena flos-aquae
Anabaena spiroides
Total Anabaena
Aphanizomenon flos-aquae
Microcystis aeruginosa
Microcystis sp.
Total Microcystis
Same day
a
b
d2
d8
Combined
4 day antecedent
a 0.91
b
d2
d8
Combined 0.20 0.30 -0.67 -0.20
7 day antecedent
a 0.36 -0.79
b -0.31 -0.33
d2
d8
Combined -0.21 -0.25 0.24 -0.72 -0.29
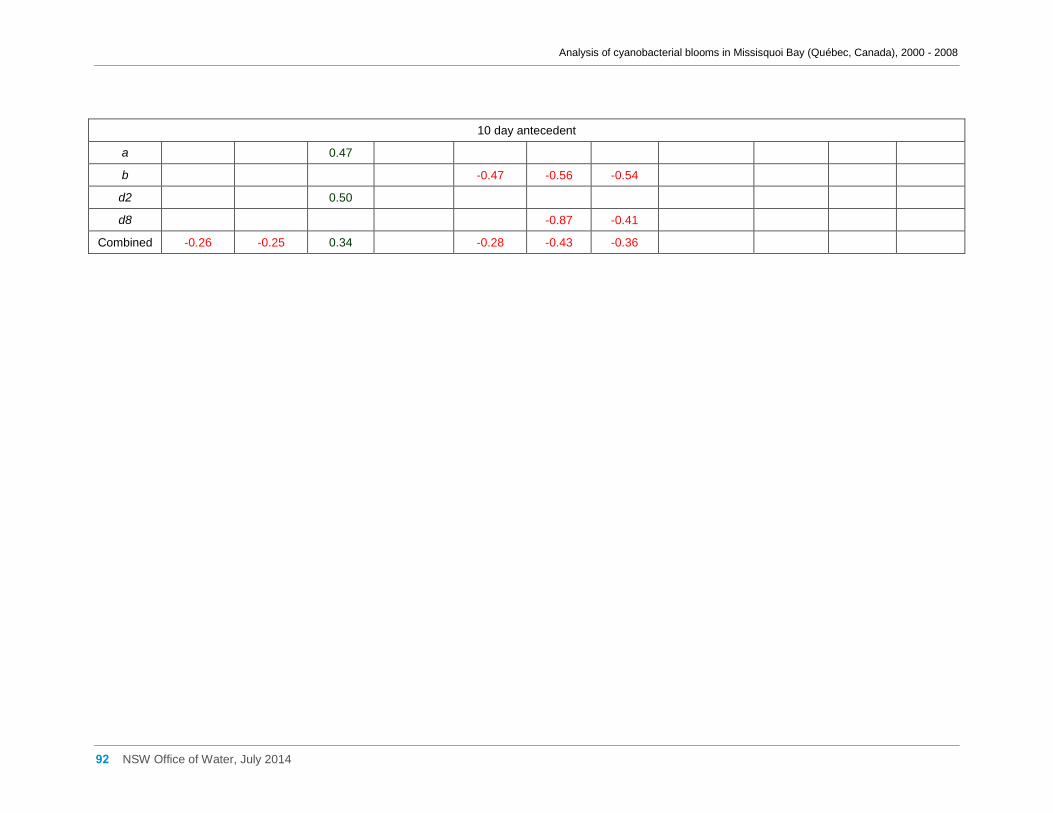
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
92 NSW Office of Water, July 2014
10 day antecedent
a 0.47
b -0.47 -0.56 -0.54
d2 0.50
d8 -0.87 -0.41
Combined -0.26 -0.25 0.34 -0.28 -0.43 -0.36
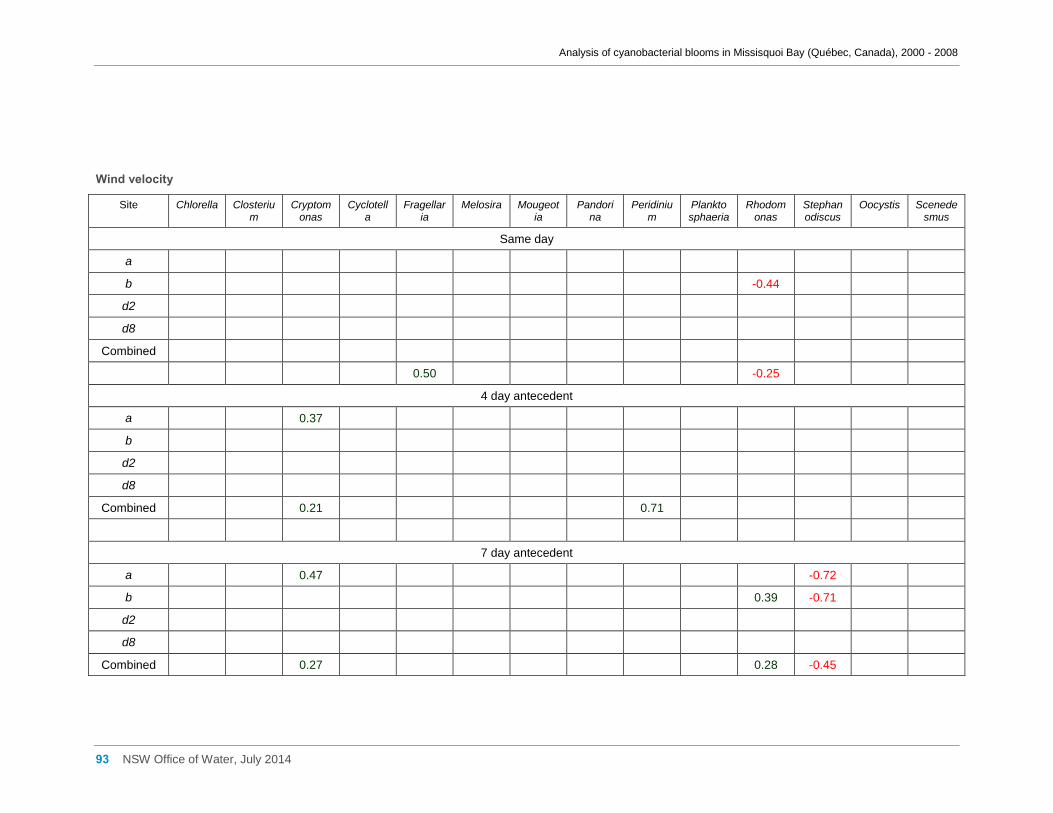
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
93 NSW Office of Water, July 2014
Wind velocity
Site Chlorella Closterium
Cryptomonas
Cyclotella
Fragellaria
Melosira Mougeotia
Pandorina
Peridinium
Plankto sphaeria
Rhodomonas
Stephanodiscus
Oocystis Scenedesmus
Same day
a
b -0.44
d2
d8
Combined
0.50 -0.25
4 day antecedent
a 0.37
b
d2
d8
Combined 0.21 0.71
7 day antecedent
a 0.47 -0.72
b 0.39 -0.71
d2
d8
Combined 0.27 0.28 -0.45
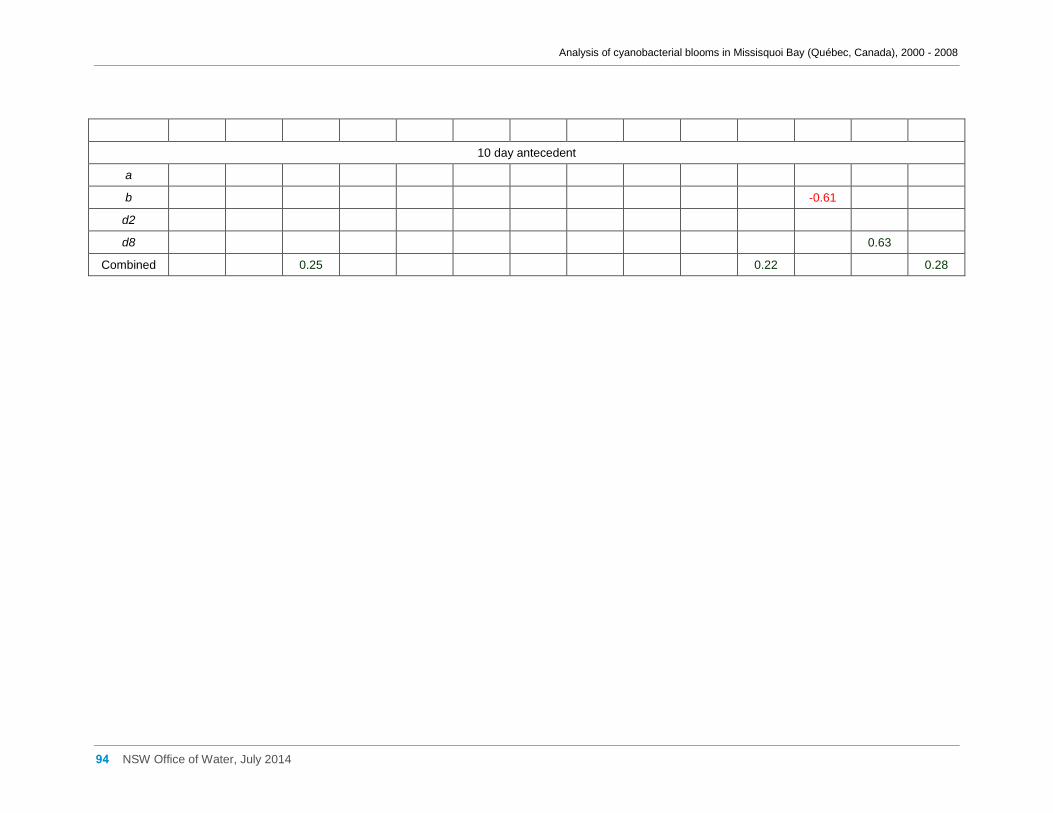
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
94 NSW Office of Water, July 2014
10 day antecedent
a
b -0.61
d2
d8 0.63
Combined 0.25 0.22 0.28
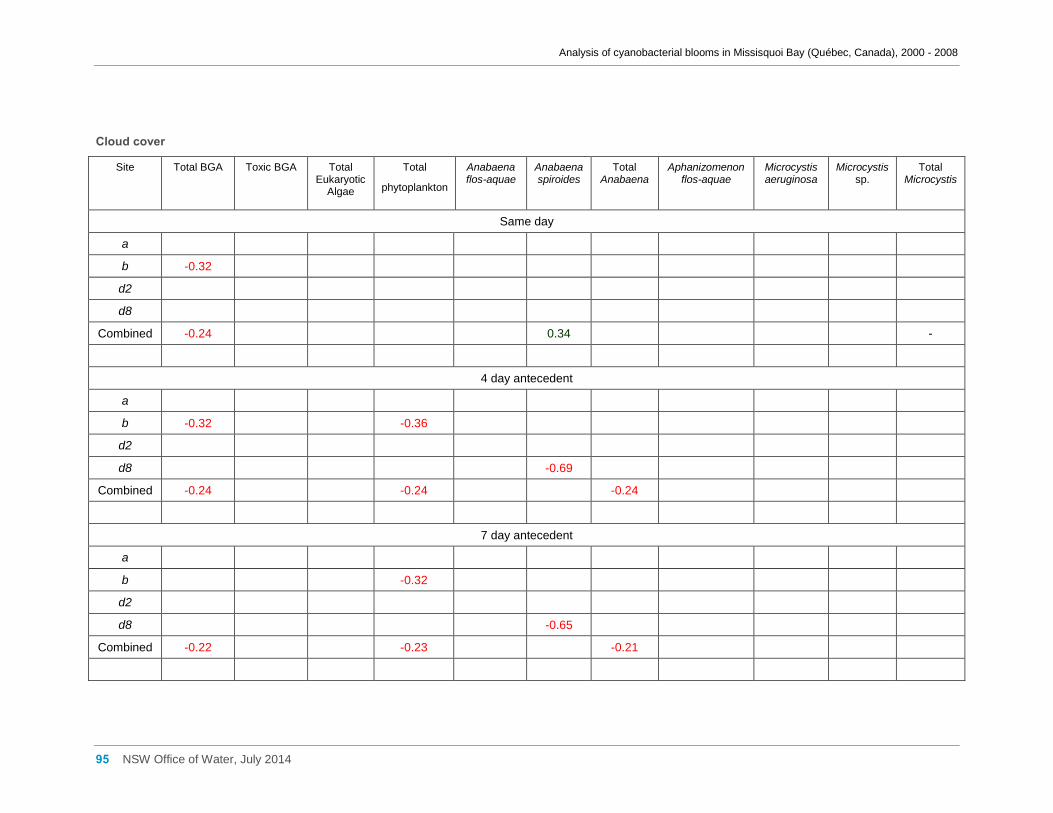
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
95 NSW Office of Water, July 2014
Cloud cover
Site Total BGA Toxic BGA Total Eukaryotic
Algae
Total
phytoplankton
Anabaena flos-aquae
Anabaena spiroides
Total Anabaena
Aphanizomenon flos-aquae
Microcystis aeruginosa
Microcystis sp.
Total Microcystis
Same day
a
b -0.32
d2
d8
Combined -0.24 0.34 -
4 day antecedent
a
b -0.32 -0.36
d2
d8 -0.69
Combined -0.24 -0.24 -0.24
7 day antecedent
a
b -0.32
d2
d8 -0.65
Combined -0.22 -0.23 -0.21
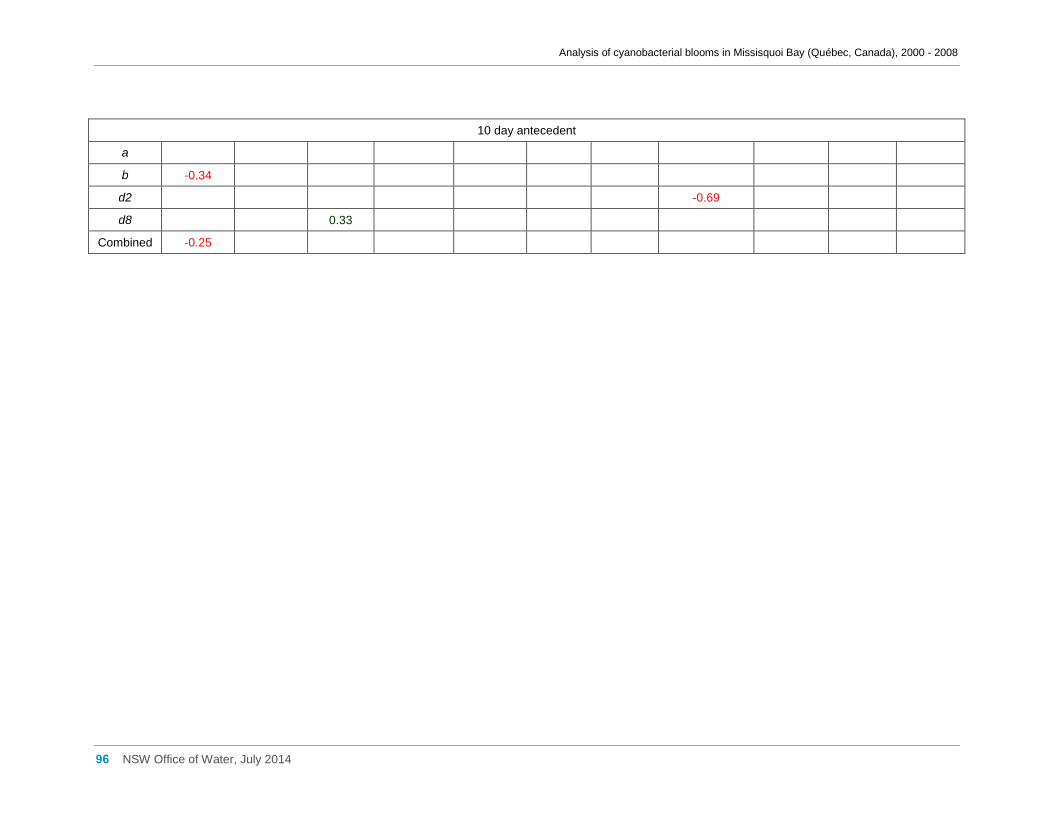
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
96 NSW Office of Water, July 2014
10 day antecedent
a
b -0.34
d2 -0.69
d8 0.33
Combined -0.25
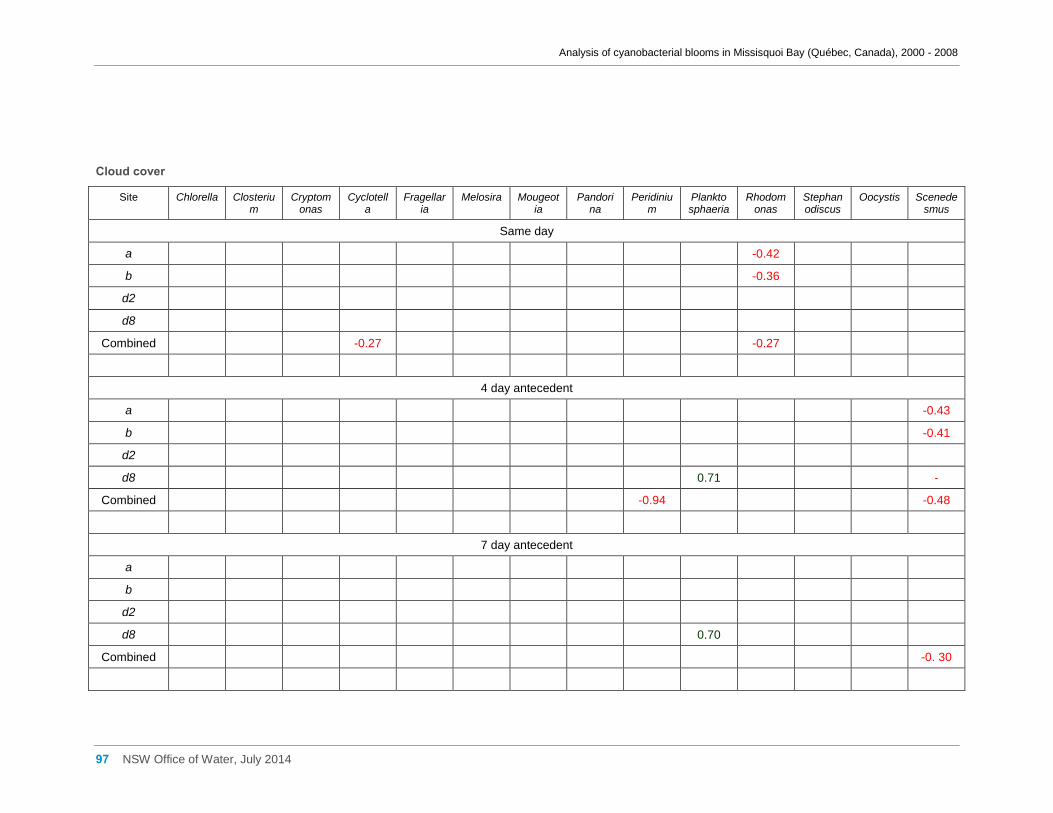
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
97 NSW Office of Water, July 2014
Cloud cover
Site Chlorella Closterium
Cryptomonas
Cyclotella
Fragellaria
Melosira Mougeotia
Pandorina
Peridinium
Plankto sphaeria
Rhodomonas
Stephanodiscus
Oocystis Scenedesmus
Same day
a -0.42
b -0.36
d2
d8
Combined -0.27 -0.27
4 day antecedent
a -0.43
b -0.41
d2
d8 0.71 -
Combined -0.94 -0.48
7 day antecedent
a
b
d2
d8 0.70
Combined -0. 30
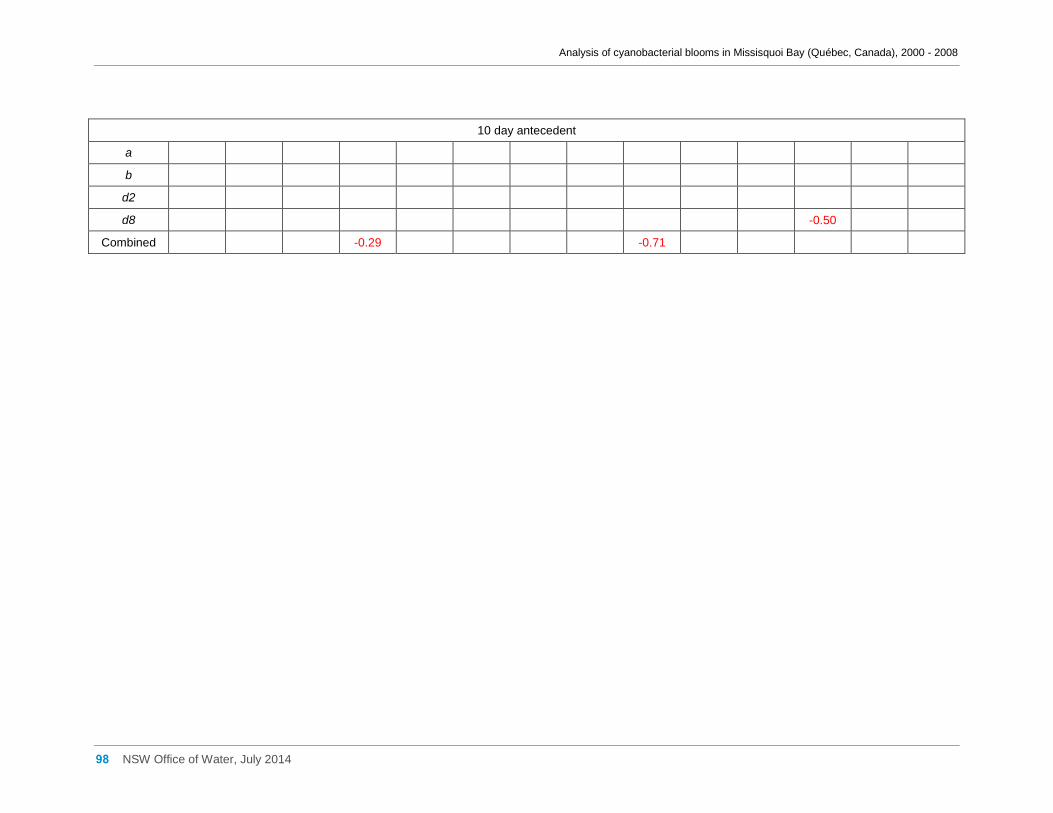
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
98 NSW Office of Water, July 2014
10 day antecedent
a
b
d2
d8 -0.50
Combined -0.29 -0.71
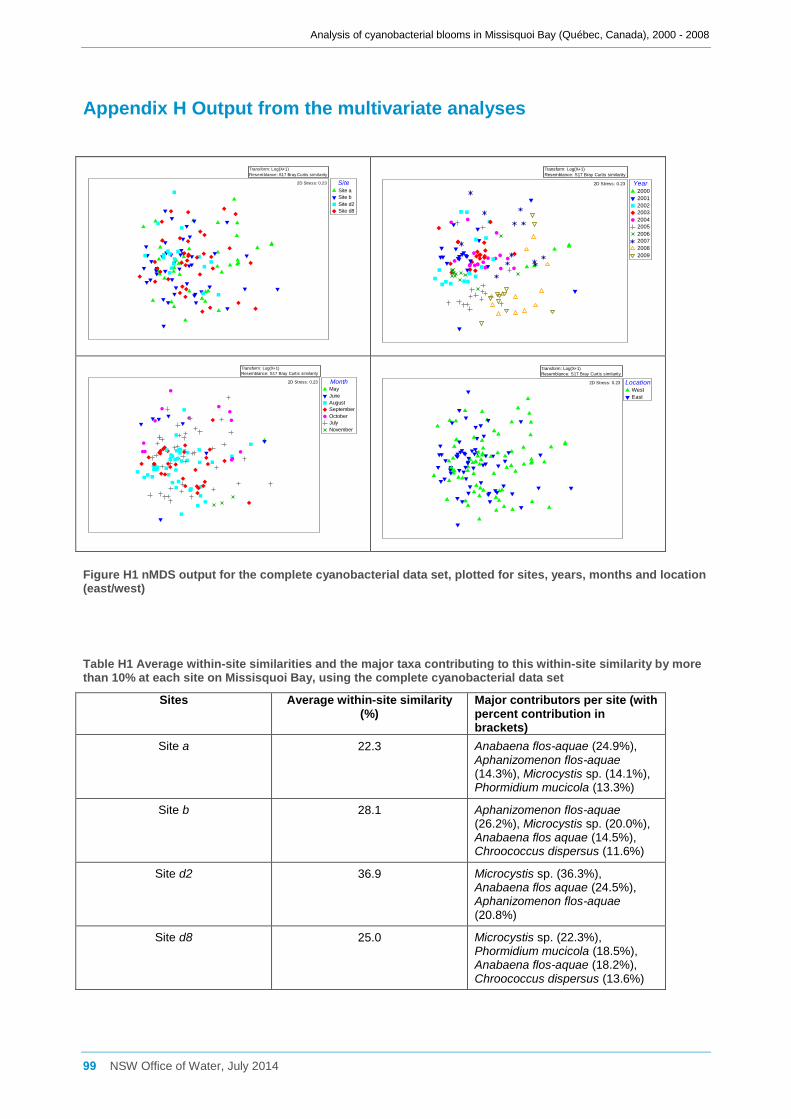
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
99 NSW Office of Water, July 2014
Appendix H Output from the multivariate analyses
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
SiteSite a
Site b
Site d2
Site d8
2D Stress: 0.23
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
Year2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2D Stress: 0.23
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
MonthMay
June
August
September
October
July
November
2D Stress: 0.23
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
LocationWest
East
2D Stress: 0.23
Figure H1 nMDS output for the complete cyanobacterial data set, plotted for sites, years, months and location (east/west)
Table H1 Average within-site similarities and the major taxa contributing to this within-site similarity by more than 10% at each site on Missisquoi Bay, using the complete cyanobacterial data set
Sites Average within-site similarity (%)
Major contributors per site (with percent contribution in brackets)
Site a 22.3 Anabaena flos-aquae (24.9%), Aphanizomenon flos-aquae (14.3%), Microcystis sp. (14.1%), Phormidium mucicola (13.3%)
Site b 28.1 Aphanizomenon flos-aquae (26.2%), Microcystis sp. (20.0%), Anabaena flos aquae (14.5%), Chroococcus dispersus (11.6%)
Site d2 36.9 Microcystis sp. (36.3%), Anabaena flos aquae (24.5%), Aphanizomenon flos-aquae (20.8%)
Site d8 25.0 Microcystis sp. (22.3%), Phormidium mucicola (18.5%), Anabaena flos-aquae (18.2%), Chroococcus dispersus (13.6%)
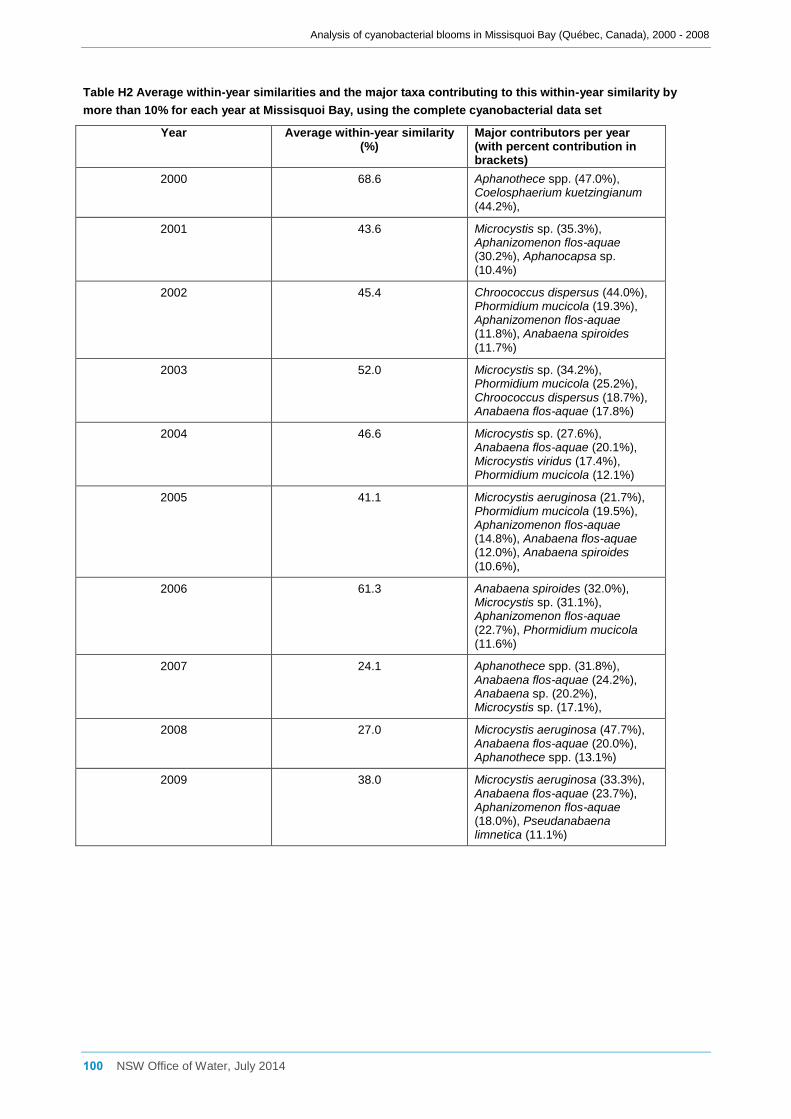
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
100 NSW Office of Water, July 2014
Table H2 Average within-year similarities and the major taxa contributing to this within-year similarity by
more than 10% for each year at Missisquoi Bay, using the complete cyanobacterial data set
Year Average within-year similarity (%)
Major contributors per year (with percent contribution in brackets)
2000 68.6 Aphanothece spp. (47.0%), Coelosphaerium kuetzingianum (44.2%),
2001 43.6 Microcystis sp. (35.3%), Aphanizomenon flos-aquae (30.2%), Aphanocapsa sp. (10.4%)
2002 45.4 Chroococcus dispersus (44.0%), Phormidium mucicola (19.3%), Aphanizomenon flos-aquae (11.8%), Anabaena spiroides
(11.7%)
2003 52.0 Microcystis sp. (34.2%), Phormidium mucicola (25.2%), Chroococcus dispersus (18.7%), Anabaena flos-aquae (17.8%)
2004 46.6 Microcystis sp. (27.6%), Anabaena flos-aquae (20.1%), Microcystis viridus (17.4%), Phormidium mucicola (12.1%)
2005 41.1 Microcystis aeruginosa (21.7%), Phormidium mucicola (19.5%), Aphanizomenon flos-aquae (14.8%), Anabaena flos-aquae (12.0%), Anabaena spiroides
(10.6%),
2006 61.3 Anabaena spiroides (32.0%), Microcystis sp. (31.1%), Aphanizomenon flos-aquae (22.7%), Phormidium mucicola (11.6%)
2007 24.1 Aphanothece spp. (31.8%), Anabaena flos-aquae (24.2%), Anabaena sp. (20.2%), Microcystis sp. (17.1%),
2008 27.0 Microcystis aeruginosa (47.7%), Anabaena flos-aquae (20.0%), Aphanothece spp. (13.1%)
2009 38.0 Microcystis aeruginosa (33.3%), Anabaena flos-aquae (23.7%), Aphanizomenon flos-aquae (18.0%), Pseudanabaena limnetica (11.1%)
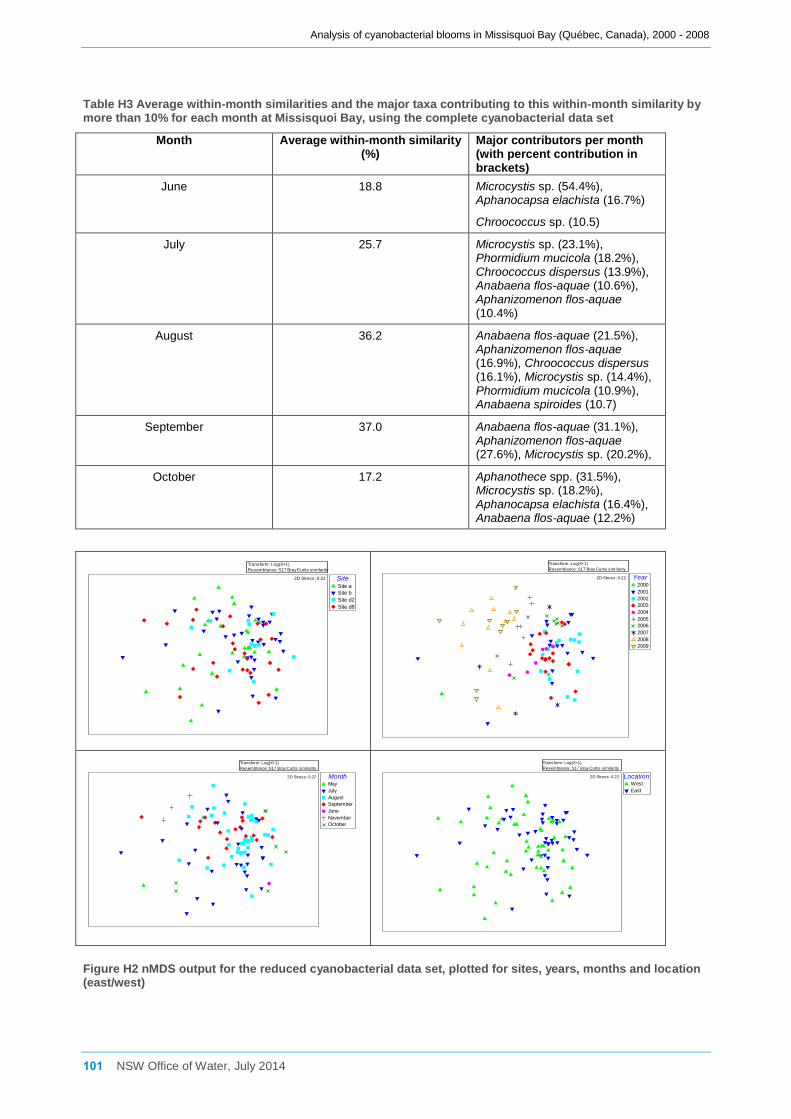
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
101 NSW Office of Water, July 2014
Table H3 Average within-month similarities and the major taxa contributing to this within-month similarity by more than 10% for each month at Missisquoi Bay, using the complete cyanobacterial data set
Month Average within-month similarity (%)
Major contributors per month (with percent contribution in brackets)
June 18.8 Microcystis sp. (54.4%), Aphanocapsa elachista (16.7%)
Chroococcus sp. (10.5)
July 25.7 Microcystis sp. (23.1%), Phormidium mucicola (18.2%), Chroococcus dispersus (13.9%), Anabaena flos-aquae (10.6%), Aphanizomenon flos-aquae (10.4%)
August 36.2 Anabaena flos-aquae (21.5%), Aphanizomenon flos-aquae (16.9%), Chroococcus dispersus (16.1%), Microcystis sp. (14.4%), Phormidium mucicola (10.9%), Anabaena spiroides (10.7)
September 37.0 Anabaena flos-aquae (31.1%), Aphanizomenon flos-aquae (27.6%), Microcystis sp. (20.2%),
October 17.2 Aphanothece spp. (31.5%), Microcystis sp. (18.2%), Aphanocapsa elachista (16.4%), Anabaena flos-aquae (12.2%)
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
SiteSite a
Site b
Site d2
Site d8
2D Stress: 0.22
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
Year2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2D Stress: 0.22
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
MonthMay
July
August
September
June
November
October
2D Stress: 0.22
Transform: Log(X+1)
Resemblance: S17 Bray Curtis similarity
LocationWest
East
2D Stress: 0.22
Figure H2 nMDS output for the reduced cyanobacterial data set, plotted for sites, years, months and location (east/west)
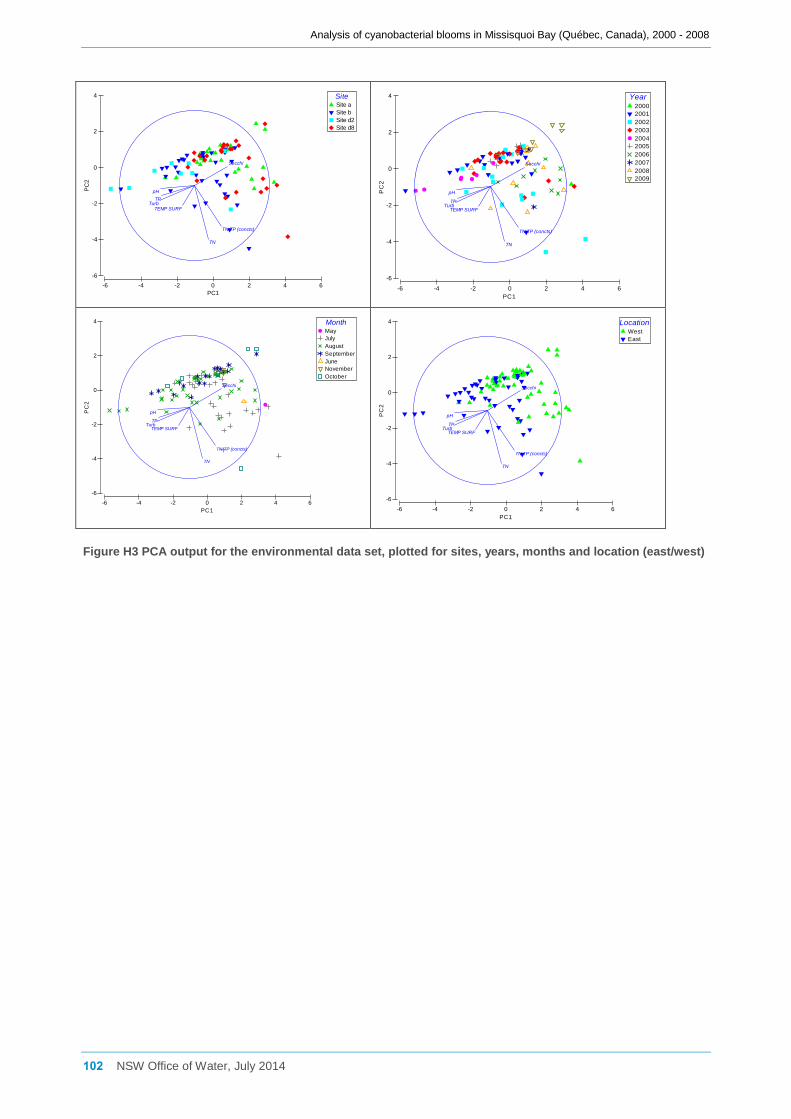
Analysis of cyanobacterial blooms in Missisquoi Bay (Québec, Canada), 2000 - 2008
102 NSW Office of Water, July 2014
-6 -4 -2 0 2 4 6
PC1
-6
-4
-2
0
2
4
PC
2
SiteSite a
Site b
Site d2
Site d8
TP
TN
TN:TP (concts)
pH
TurbTEMP SURF
Secchi
-6 -4 -2 0 2 4 6
PC1
-6
-4
-2
0
2
4
PC
2
Year2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
TP
TN
TN:TP (concts)
pH
TurbTEMP SURF
Secchi
-6 -4 -2 0 2 4 6
PC1
-6
-4
-2
0
2
4
PC
2
MonthMay
July
August
September
June
November
October
TP
TN
TN:TP (concts)
pH
TurbTEMP SURF
Secchi
-6 -4 -2 0 2 4 6
PC1
-6
-4
-2
0
2
4
PC
2
LocationWest
East
TP
TN
TN:TP (concts)
pH
TurbTEMP SURF
Secchi
Figure H3 PCA output for the environmental data set, plotted for sites, years, months and location (east/west)





























