Embed Size (px)
Citation preview

An analysis of otoliths from two eels (Anguilla australis and 1
Anguilla dieffenbachii) recovered from a hydroelectric dam 2
turbine in the Tarawera River, New Zealand 3
Paris Achenbach1 4
1. University of Canterbury, Christchurch New Zealand. email: [email protected] 5
6
ABSTRACT 7
With the onset of anthropogenic affects on aquatic life including climate change, over-fishing 8
and hydroelectric dam construction, it is more important than ever to understand fish 9
behaviour, life cycles, and migratory journeys. Otoliths provide a wealth of information 10
regarding their biological characteristics; however, they’re also difficult to analyze and 11
interpret, and despite the wealth of existing literature on otoliths, few have provided an all-12
encompassing look at otolith methodologies. The present study experiments on two pairs of 13
eel otoliths, one pair from Anguilla australis and one from Anguilla dieffenbachii, retrieved 14
from the Tarawera River, New Zealand, to examine the biological characteristics of the eels 15
(such as the lifespan and growth rate) using a Scanning Electron Microscope. It also 16
compiles a straightforward and effective approach to methodologies of eel otolith research, 17
from original findings, and from outside recent research. It is important to note that while this 18
paper serves as a review of otolith methodologies, it also presents several new questions 19
regarding the specific otoliths recovered from the Tarawera River. One surprising finding in 20
particular regarding the SEM images was a series of pores in the shortfin eel otolith, 21
suggesting the eel had a disease or other unknown condition. 22
23
INTRODUCTION 24
Worldwide freshwater eel (Anguilla species) populations are facing a rapid decline, posing a 25
problem for commercial fisheries, ecosystem health, and cultural diets (Guan et al. 2004; 26
Tesch 2003; Boubee et al. 2008; Chisnall and Hicks 1993; Kelly et al. 2007). Eel is a high 27
commodity for the fishing industry; in 1995, total world eel harvest amounted to about 28
205,000 tons of eel from fishing and aquaculture, which was valued at over US$3.1 billion 29
(Tesch 2003). Further than just being a problem economically, the eel population decline has 30
significant effects for countries that value eels as an important part of their culture, such as 31
China, Japan, and New Zealand (Guan et al. 2004). 32

33
In New Zealand, the local Maori people view eels an important part of their diet; not 34
coincidentally, eels naturally contain high amounts of proteins and antioxidants, which have 35
been shown to reduce the risk of heart disease and cancer (Ekanayake et al. 2005). There are 36
two endemic species of eel in New Zealand: the shortfin eel (Anguilla australis) and longfin 37
eel (Anguilla dieffenbachii), the latter of which lives longer and is solely found in New 38
Zealand (Northland Conservancy 2006). 39
40
Although it is understood that New Zealand eels are diadromous, meaning they spawn in the 41
ocean but mature in freshwater, little is known about the specifics of where they hatch and 42
travel, and similarly, how they migrate, and what cues they use for navigation. Recent 43
initiatives in discovering eel spawning locations involve satellite tagging, and radio and 44
acoustic telemetry studies (Jellyman 2006, 2009; Jellyman and Tsukamoto 2010; Aarestrup et 45
al. 2009), which have shown that New Zealand eels potentially spawn off the coast of New 46
Caledonia, putting their migration distance over 2,000 kilometers. One satellite in particular 47
showed longfin eels to swim at speeds ranging from 26 to 31 km per day and at depths of up 48
to 980m (Jellyman 2006). 49
50
A more common way to examine eel migration and life cycles, however, is by studying their 51
otoliths, which can reveal age and growth rate through back-calculation of annually deposited 52
rings (McCleave 2008; Yokouchi et al. 2011; Tzeng et al. 1993; Marohn et al. 2009; 53
Lecomte-Finiger 1992; Chisnall and Hicks 1993; Graynoth 1999; Jellyman 1995). Further, 54
the otoliths can be used to study the timing of the eel’s entrance into freshwater or saltwater, 55
by analyzing the chemical makeup of the bone (Arai et al. 2003; Tzeng et al. 2005; Tzeng 56
and Tsai 1994; Yokouchi et al. 2011). 57
58
There are numerous ways in which otoliths can be prepared and examined, including etching, 59
SEM imaging, staining, burn-and-breaking, sawing, using laser ablation, or a combination of 60
these procedures. This study examines two pairs of otoliths from the Tarawera River using a 61
combination of methods, including with etching and SEM imaging, to compare the 62
differences in ages and size between the two species, as well as understand the potential 63
variables that affect otolith size, morphology, and growth rate. 64
65
METHODS 66

Two pairs of eel otoliths were extracted from two eels that had died in a hydroelectric turbine 67
on the Tarawera River, New Zealand. One pair is from a female longfin eel (Anguilla 68
dieffenbachii), which measured to be 160 cm at 8.8 kg. The other pair is from a male shortfin 69
eel, A. australis, which measured to be 100 cm at 1.74 kg. Unfortunately one of the otoliths 70
from the shortfin eel broke in half in transit to the South Island, but it was still possible to 71
examine the two pieces. The otoliths were embedded in resin, then ground to reveal the 72
nucleus and rings, and finally polished on a lap with ½ µm diamond paste (check). They were 73
then examined under a Scanning Electron Microscope four times, under a variety of 74
conditions to obtain the clearest image possible: 75
-With a gold coating 76
-Painted with liquid carbon, drying into a solid conductive surface from the edge of 77
the resin to the edge of the otolith 78
-Etched with Hydrochloric acid for 2-3 minutes 79
After the primary gold covering, the otoliths were grinded again and re-polished, before 80
painting with liquid carbon. 81
82
Over the course of four SEM sessions, the otoliths were examined under backscatter and 83
secondary electron microscopy settings. Post-SEM session, images were studied to count the 84
annuli deposits and observe the relationship between the rings, otolith radius, and length and 85
weight of the eel. 86
87
RESULTS 88
89
The otoliths were difficult to view in the SEM under just a gold coating or painted with liquid 90
carbon; the image was too blurry to see any rings (figure 1a, 1b). However, the etching was 91
highly effective in producing a clear image on the SEM to examine the microstructure. 92
Unfortunately, this meant that the microchemistry of the otolith would not be able to be 93
examined, after altering it with hydrocholoric acid. 94
95
Otoliths from Anguilla deiffenbachii 96
The longfin otolith radius measured to be 2.5mm (figure 2) on its longest axis under the 97
SEM, and showed 20 major rings throughout that axis; however, there are clearly many more 98
rings within those rings outlined (figure 3a). On average, major rings occur once every 100 99

µm, but within those 100 µm transects, there are at least 6 or 7 distinct rings (figure 3b). This 100
would put the eel at an age much older than 20. 101
102
Otoliths from Anguilla australis 103
The shortfin eel otolith radius measured to be 1.9 mm (figure 4) on its longest axis under the 104
SEM, and showed 22 major rings from the nucleus to the edge, though the rings became 105
increasingly more blurred the further from the nucleus (figure 5a). This otolith too showed 106
multiple rings within rings, suggesting either the eel was older than 22, or that there were 107
false annuli deposits, or perhaps seasonal or daily deposits (figure 5b; figure 6a and 6b). 108
109
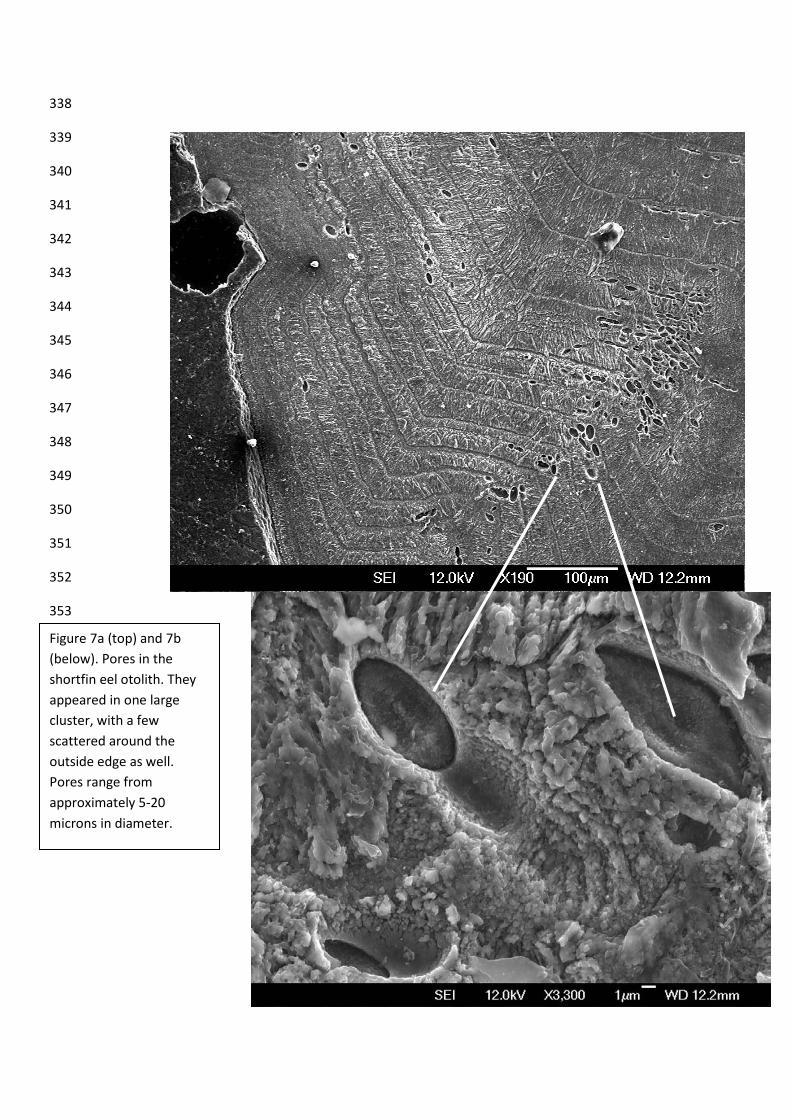
Pores in Anguilla australis 110
One of the more curious findings of this study was the presence of pores, or perhaps pits, in 111
both of the broken pieces of the Anguilla australis otolith (figure 7a and 7b). Due to lack of 112
time, there are only pictures from one of the pieces, but they were present in both. It’s unclear 113
if they were on opposing sides of the otolith or only on one side, but they clearly 114
conglomerated together in one particular spot on the otolith. 115
116
DISCUSSION 117
SEM settings: Backscatter vs Secondary Settings 118
Both the backscatter and secondary settings showed distinct rings after etching; the former 119
showing chemical differences in the otolith, the latter showing topographical differences. 120
Both settings showed rings in the same places (figure 8), indicating that a chemical change is 121
associated with a topographical change in otoliths. 122
123
Relationship between Otolith Radius and Eel Length 124
One primary use of studying otoliths is to gauge the relationship between eel length and 125
otolith radius, and find the average rate of growth. Interestingly, the present otoliths fell 126
closely in line with the otolith radius to eel length ratio from a study by Graynoth, which 127
involved 939 longfin and 791 shortfin eels, suggesting that the relationship between the two 128
is curvilinear (figure 9). 129
130
Age of Eels 131
Although rough estimates of the eel ages could be found with the images from the SEM, it 132
was difficult to find an accurate or precise age, because the annual bands were often 133

indiscernible from seasonal and daily growth rings. If only the larger bands are counted as 134
annual bands, it would put both eels at an age between 20 and 25 (figures 3a and 5a). 135
However, if the major rings (~6) within the larger rings are counted for the eels, it would put 136
their age at over 100 years old (figure 3b and 5b). 137
138
Otolith Pores 139
The otolith pores were clearly not a result of etching, because they did not appear in three of 140
the four otoliths examined which also underwent etching; furthermore, the circles are too 141
rounded to be a result of hydrochloric acid. Unfortunately, because of lack of time in the 142
SEM session, it was unclear whether or not the pits were on opposite sides of the otolith. If 143
they occurred only on one side, then it may be possible they formed around hair follicles 144
during later development, which is what one study found in otoliths from a white sea bass in 145
California (figure 10) (Checkley, unpublished). However, if they formed on both sides, then 146
it must be something else, perhaps a disease or another unknown condition. This is something 147
that should be further investigated. 148
149
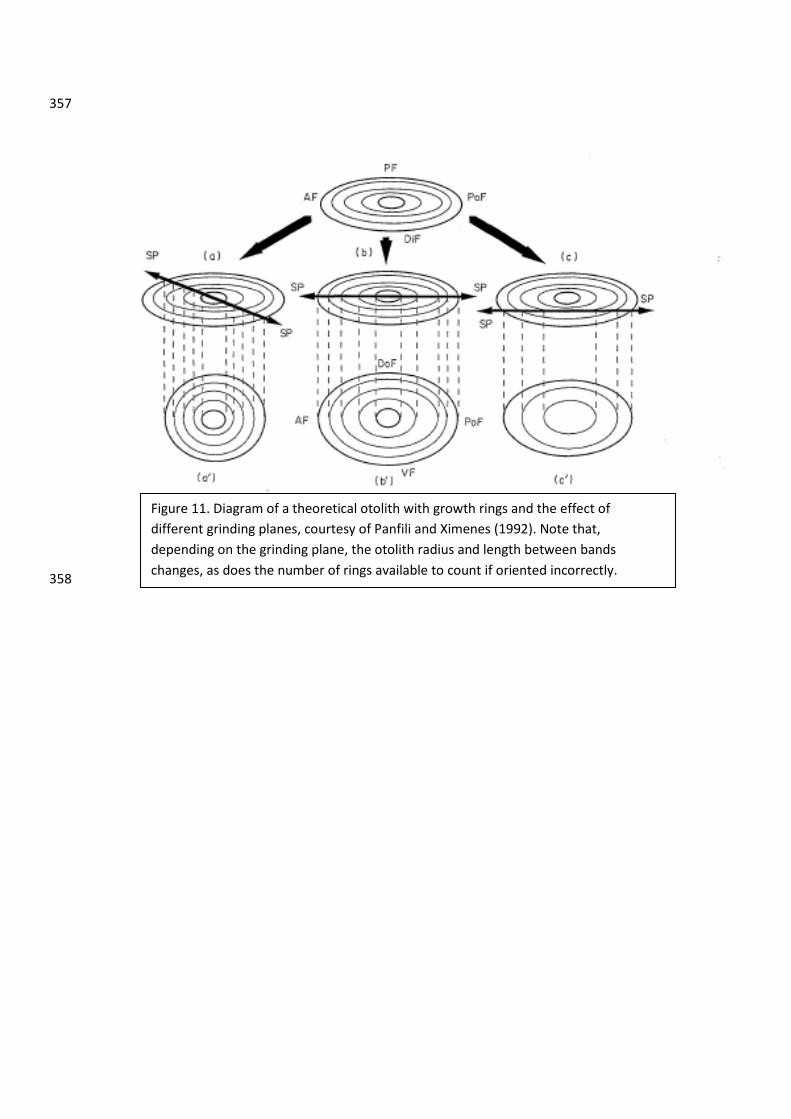
Problems in Interpretation: Methodologies and Otolith Preparation 150
Otolith interpretations can be easily affected by the process in which they are prepared and 151
studied. For example, depending on how you orient the otolith in resin and how far down you 152
grind and polish the otolith, you can get an inaccurate ring count, and thus age, of the otolith 153
(figure 11) (Panfili and Ximenes 1992). Similarly, eels in colder water grow slower than in 154
warm water, making rings closer together and difficult to discern from one another; this is 155
particularly a problem in glacial lakes of New Zealand, where eels are only growing about 156
9mm/year (especially difficult for really old eels, up to 106 years) (Jellyman 1995). Further, 157
the sampling techniques of otolith trace elements such as diameter of sampling spot could 158
affect the concentration of elements and reconstruction of migratory history of fishes 159
(Yokouchi et al 2011). Lastly, the otolith radius is measured on its longest axis in most 160
studies, but the longest axis depends on how you orient and grind them (figure 12) (Graynoth 161
1999). 162
163
In this particular study, it was difficult to find an accurate age count under the SEM for a 164
number of reasons, including: a) the charge buildup that occurs and causes black spots in 165
imaging after time, thus the examiner must operate with maximum efficiency to find the best 166
images as early as possible; b) photos later can be difficult to interpret; c) it’s difficult to get 167

an overall picture of larger otoliths (ie >2mm radius on its long axis) because of the narrow 168
scope of the SEM lens. 169
170
Problems in Interpretation: Complications in Ontogenic Development 171
In addition to preparation issues causing a conflict in otolith interpretation, there have been 172
numerous studies showing other biological variables affect otolith morphology and growth 173
rings. 174
1. Otolith rings are often thicker in one section than another, and often overlap or 175
coalesce as well (figure 3b, figure 12). Thus, it’s difficult to get an accurate length/age 176
to otolith radius ratio, ie a growth rate to otolith radius ratio, because the otolith radius 177
is inconsistent. 178
2. Bands, or checks, are formed by changes in calcium and protein deposition rates 179
due to sudden changes in temperature, and possibly other factors, as opposed to 180
simply annual deposits (Graynoth 1999, Jellyman 1995). Similarly, irregular variation 181
in width of summer bands could represent migrations between different habitats, 182
because rate of maturation is affected by temperature, pressure and swimming 183
(Jellyman and Tsukamoto 2010). Thus, rings could potentially be due to starvation, an 184
increase or decrease in swimming, or other biogenic or environmental variables, as 185
opposed to a temporal variable. 186
3. There’s a delay in the incorporation of different Sr:Ca ratios depending on ambient 187
water – time lag of at least 10 days and up to 30-60 days in Anguilla japonica; and 188
further, strontium incorporation can also be affected by biological factors like 189
temperature and growth (Yokouchi et al. 2011), both of which make it difficult to 190
assess an accurate timeline for eel migration using strontium-calcium ratios found 191
through chemical analysis of the otolith 192
4. The area of the otoliths can increase when carbon dioxide levels in the ocean water 193
increases due to acidification – an affect that would take place during the leptocephali 194
phase, and could potentially increase the area of the otolith by a significant percent, 195
thus throwing off the otolith radius to length/age ratio (Checkley et al 2009) 196
197
CONCLUSION 198
Many questions remain regarding spawning locations of both eel species, but particularly of 199
A. deiffenbachii, of which only one eel has been observed to travel to New Caledonia, and no 200
larvae have ever been found in the ocean (Kuroki et al. 2008; Jellyman and Tsukamoto 201

2010). Due to the rapid decline in their population numbers, it is imperative to understand eel 202
behaviour and migration if we want to set up conservation schemes and protect them from 203
potential obstacles to their migration, such as hydroelectric dams. 204
205
There has been a multitude of research published on eel otolith growth, microstructure, and 206
microchemistry, and yet the data is quite convoluted and makes it difficult to come up with 207
many accurate conclusions. Otolith microstructure studies provide an important base for our 208
understanding of eel migration and life cycles, but when it comes to preserving their 209
population, a more practical approach is necessary. As of now, satellite tagging and acoustic 210
and telemetry studies provide the best option for establishing effective protection measures, 211
as they can tell us a) the origin of eel spawning, b) their migratory paths (distance, location 212
etc), c) time taken during migration, d) eel movement and behaviour within the water 213
column, e) movement within lakes, and f) how much of their migration is due to using 214
oceanic currents for travelling efficiency, versus how much is due to actual navigation and 215
swimming. From there, it may even be possible to understand the environmental cues and 216
conditions that promote or guide their migration paths, whether they are lunar 217
phases/declination, water temperature, or magnetic navigation. Once we understand how and 218
why they are migrating, it may be possible to establish how anthropogenic affects such as 219
global climate change and hydroelectric dam construction may impact their migration and 220
ultimately survival. 221
222
ACKNOWLEDGEMENTS 223
224
I’d like to thank Kerry Swanson for the endless help with the four SEM sessions and otolith 225
preparation; Travis Horton and Nan Hauser for promoting my interest in animal migration; 226
Bill Kerrison and Mick Kearson for the otoliths and inspiration; Rob Spiers for the diamond 227
paste, sandpaper and entertaining stories; Don Jellyman at NIWA for taking time out of his 228
busy day to explain eel migration and otoliths; and Josh Blackstock for helping me all along 229
the way. 230
231
232
233
234

REFERENCES 235
Arai T, Kotake A, Lokman PM, Tsukamoto K (2003) Migratory history and habitat use by 236 New Zealand freshwater eels Anguilla dieffenbachia and A. australis, as revealed by otolith 237 microchemistry. Ichthyological Resesarch 50: 190-194 238
Boubee J, Jellyman D, Sinclair C (2008) Eel protection measures within the Manapouri 239 hydroelectric power scheme, South Island, New Zealand. Hydrobiologia 609:71-82 240
Checkley DM, Dickson AG, Takahashi M, Radich A, Eisenkolb N, Asch R (2009) Elevated 241 CO2 Enhances Otolith Growth in Young Fish. Science 342:1683. 242
Chisnall BL, Hicks BJ (1993) Age and growth of longfinned eels (Anguilla dieffenbachii) in 243 pastoral and forested streams in the Waikato River basin, and in two hydro-electric lakes in 244 the North Island, New Zealand. New Zealand Journal of Marine and Freshwater Research 27: 245 317-332 246
Ekanayake PM, Park GT, Lee YD, Kim SJ, Jeong SC, Lee J (2005) Antioxidant Potential of 247 Eel (Anguilla Japonica and Conger Myriaster) Flesh and Skin. Journal of Food Lipids 12:34-248 47 249 250 Graynoth, E (1999) Improved otolith preparation, ageing and back-calculation techniques for 251 New Zealand freshwater eels. Fisheries Research 42: 137-146 252 253 Guan R, Xuehong W, Guozhen K (1994) Age and growth of eels Anguilla japonica in a 254 Chinese river. Journal of Fish Biology 45: 653-660. 255
Jellyman D (1995) Longevity of longfinned eels Anguilla dieffenbachii in a New Zealand 256 high country lake. Ecology of Freshwater Fish 4:106-112 257
Jellyman D (2006) Tagging along when longfins go spawning. Water and Atmosphere 14 (1): 258 24-25. Retrieved from www.niwa.co.nz 259
Jellyman D (2009) A review of radio and acoustic telemetry studies of freshwater fish in New 260 Zealand. Marine and Freshwater Research 60: 321-327 261
Jellyman D, Tsukamoto K (2010) Vertical migrations may control maturation in migrating 262 female Anguilla dieffenbachii. Marine Ecology Progress Series 404: 241-247 263
Kelly DJ, Jellyman DJ (2007) Changes in trophic linkages to shortfin eels (Anguilla 264 australis) since the collapse of submerged macrophytes in Lake Ellesmere, New Zealand. 265 Hydrobiologia 579: 161-173 266
Kuroki M, Aoyama J, Miller MJ, Watanabe S, Shinoda A, Jellyman DJ, Feunteun E, 267 Tsukamoto K (2008) Distribution and early life-history characteristics of anguillid 268 leptocephali in the western South Pacific. Marine and Freshwater Research 59: 1035-1047 269
Lecomte-Finiger R (1992) The crystalline ultrastructure of otoliths of the eel (A anguilla L. 270 1758). Journal of Fish Biology 40:181-190 271

Marohn L, Prigge E, Zumholz K, Klugel A, Anders H, Hanel R (2009) Dietary effects on 272 multi-element composition of European eel (Anguilla anguilla) otoliths. Mar Biol 156:927-273 933 274
McCleave JD (2008) Contrasts between spawning times of Anguilla species estimated from 275 larval sampling at sea and from otolith analysis of recruiting glass eels. Mar Biol 155:249-276 262 277
Northland Conservancy (2006) New Zealand’s native freshwater fish. Department of 278 Conservation, Christchurch, NS0059. http://www.doc.govt.nz/upload/documents/about-279 doc/concessions-and-permits/conservation-revealed/nz-native-freshwater-fish-lowres.pdf 280
Panfili J, Ximenes MC (1992) Measurements on ground or sectioned otoliths: possibilities of 281 bias. Journal Fish of Biology 41: 201-207. 282
Tesch FW (2003) The Eel. Blackwell Science Ltd, Oxford, UK 283
Tzeng WN, Tsai YC (1992) Otolith microstructure and daily age of Anguilla japonica, 284 Temmink & Schlegel elvers from the estuaries of Taiwan with reference to unit stock and 285 larval migration. Journal of Fish Biology 49: 845-857 286
Tzeng WN, Tsai YC (1994) Changes in otolith microchemistry of the Japanese eel, Anguilla 287 japonica, during its migration from the ocean to the rivers of Taiwan. Journal of Fish Biology 288 45: 671-683 289
Tzeng WN, Severin KP, Wang CH, Wickstrom H (2005) Elemental composition of otoliths 290 as discriminator of life stage and growth habitat of the European eel, Anguilla anguilla. 291 Marine and Freshwater Research 56: 629-635 292
Yokouchi K, Fukuda N, Shirai K, Aoyama J, Daverat F, Tsukamoto K (2011) Time lag of the 293 response on the otolith strontium/calcium ratios of the Japanese eel, Anguilla japonica to 294 changes in strontium/calcium ratios of ambient water. Environ Biol Fish 92: 469-478 295
296
297

Figure 3a (top) and 3b (below): An overlook of the longfin otolith. The short, dark transects mark the major annual deposits, but smaller bands in between could represent annual rings as well. The opaque zone on the side could possibly be a result of incorrectly orienting the otolith in the resin. A close-up of rings on the longfin otolith. Major bands were outlined in black, totalling seven on the left. Note the crack running through the center, causing a discrepancy in the rings between the left and the right. This was probably biological in origin, as opposed to a result of the otolith preparation
298
299
300
301
302
303
304
305
306
307
308
309
310

311
312
313
314
315
Figure 5a (top) and 5b (below): An overlook of the shortfin otolith. Darker transect lines mark the annual deposits, 22 in total, though some of the lines are more ambiguous than others. The annual lines start at the first major dark deposit, which is understood to be when the eel migrates into freshwater, at around 10-11 months old. Within the approximate 10-20 micron-wide rings, there are anywhere from 5-10 more rings inside.

316
317
318
319
320
321
322
323
324
325
326
327
328
Figure 1a and 1b: An SEM photograph of the longfin otolith with a carbon coat, and without hydrochloric acid etching. Note some grinding and dust marks.
329
330
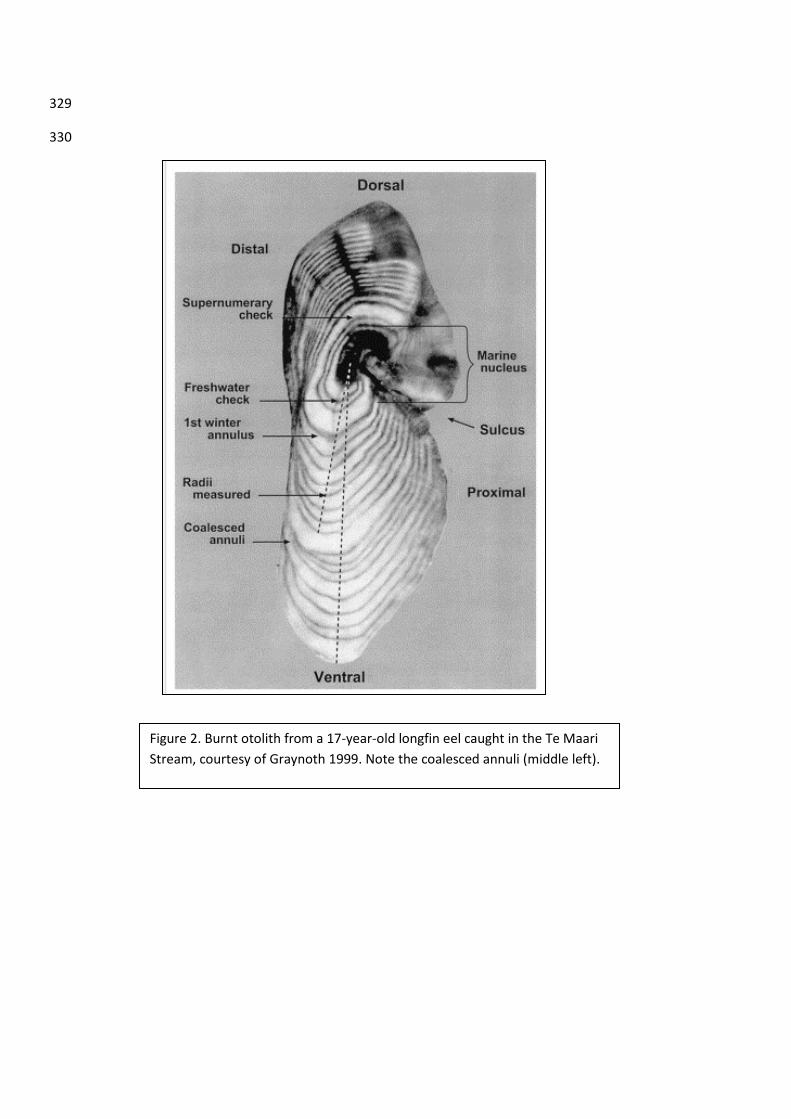
Figure 2. Burnt otolith from a 17-year-old longfin eel caught in the Te Maari Stream, courtesy of Graynoth 1999. Note the coalesced annuli (middle left).
331
332
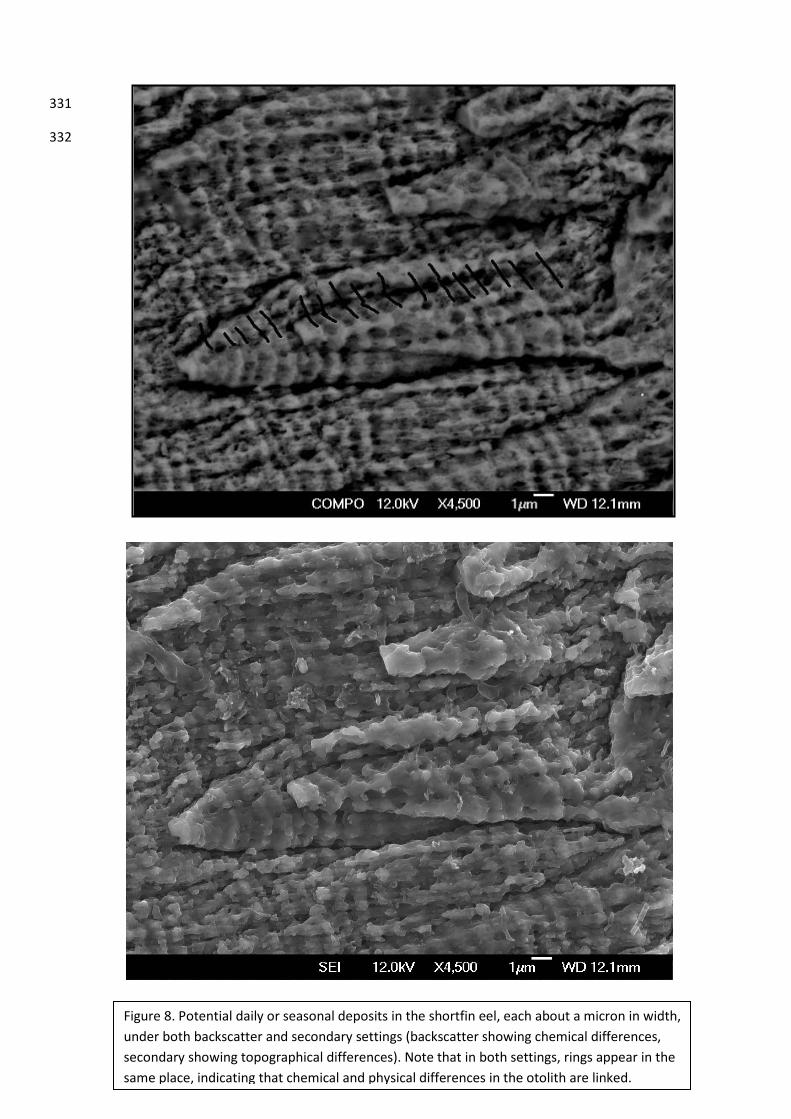
Figure 8. Potential daily or seasonal deposits in the shortfin eel, each about a micron in width, under both backscatter and secondary settings (backscatter showing chemical differences, secondary showing topographical differences). Note that in both settings, rings appear in the same place, indicating that chemical and physical differences in the otolith are linked.

333
334
Figure 6a (top) and 6b (bottom). Rings within annually deposited rings in the shortfin eel.

335
Figure 4. Otolith radius of the shortfin eel. Radius was taken on the longest axis.
Figure 2. Otolith radius of the longfin eel.
336
337
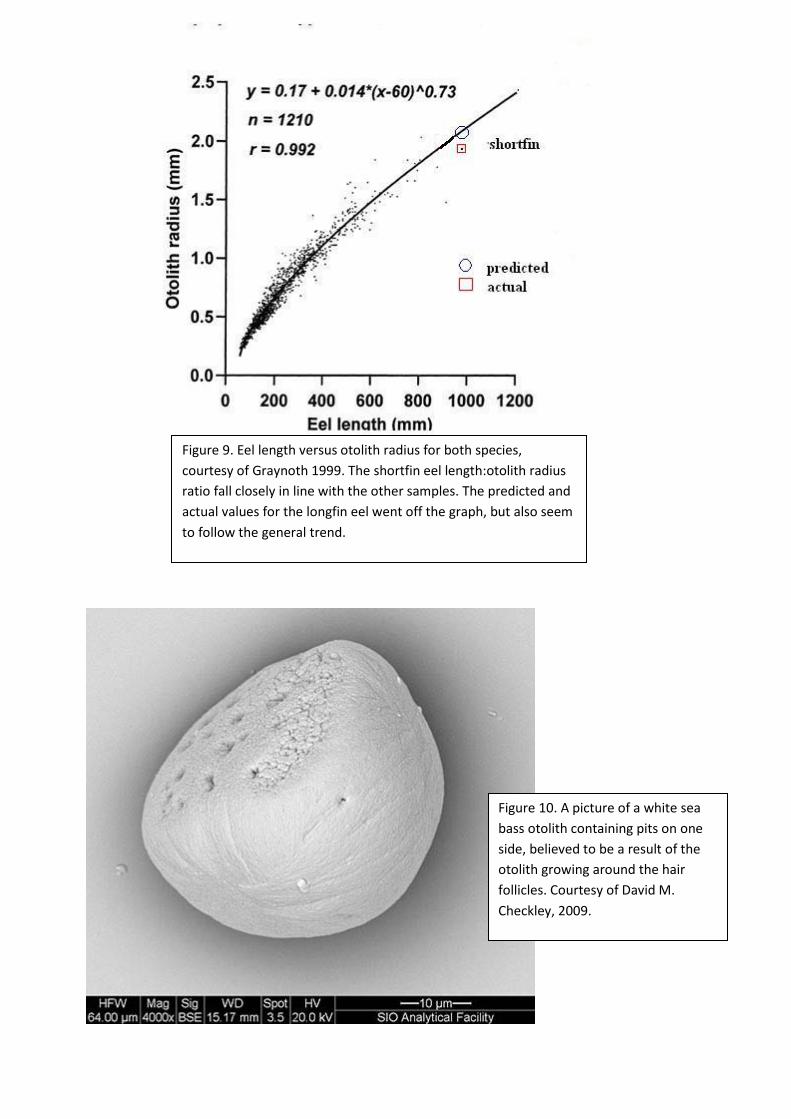
Figure 9. Eel length versus otolith radius for both species, courtesy of Graynoth 1999. The shortfin eel length:otolith radius ratio fall closely in line with the other samples. The predicted and actual values for the longfin eel went off the graph, but also seem to follow the general trend.
Figure 10. A picture of a white sea bass otolith containing pits on one side, believed to be a result of the otolith growing around the hair follicles. Courtesy of David M. Checkley, 2009.
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
Figure 7a (top) and 7b (below). Pores in the shortfin eel otolith. They appeared in one large cluster, with a few scattered around the outside edge as well. Pores range from approximately 5-20 microns in diameter.
357
358
Figure 11. Diagram of a theoretical otolith with growth rings and the effect of different grinding planes, courtesy of Panfili and Ximenes (1992). Note that, depending on the grinding plane, the otolith radius and length between bands changes, as does the number of rings available to count if oriented incorrectly.