Embed Size (px)
Citation preview

An Endophytic Sanguinarine-Producing Fungus from Macleayacordata, Fusarium proliferatum BLH51
Xue-Jun Wang • Chang-Li Min • Mei Ge •
Rui-Hua Zuo
Received: 4 September 2013 / Accepted: 11 September 2013 / Published online: 29 October 2013
� Springer Science+Business Media New York 2013
Abstract Fermentation processes using sanguinarine-
producing fungi other than Macleaya cordata may be an
alternative way to produce sanguinarine (SA), which is a
quaternary benzo[c]phenanthridine alkaloid possessing
antibacterial, anthelmintic, and anti-inflammatory proper-
ties. In this study, a SA-producing endophytic fungus strain
BLH51 was isolated from the leaves of M. cordata grown
in the Dabie Mountain, China. Strain BLH51 produced SA
when grown in potato dextrose liquid medium. The amount
of SA produced by this endophytic fungus was quantified
to be 178 lg/L by HPLC, substantially lower than that
produced by the host tissue. The fungal SA—which was
analyzed by thin layer chromatography and high-perfor-
mance liquid chromatography—was shown to be identical
to authentic SA. Strain BLH51 was identified as Fusarium
proliferatum based on the morphological characteristics
and nuclear ribosomal DNA ITS sequence analysis. To the
best of our knowledge, this is the first report concerning the
isolation and identification of endophytic SA-producing
fungi from the host plant, which further proved that
endophytic fungi are valuable reservoirs of bioactive
compounds.
Introduction
Macleaya cordata (Willd) R. Br., also known as plume
poppy or Bocconia cordata is a perennial plant of the Pap-
averaceae family. It has been used as one of traditional
Chinese medicines for a long time. This herb widely dis-
tributed throughout southeastern and northwest of China.
Sanguinarine (SA), a quaternary benzo[c] phenanthridine
alkaloid (QBA), isolated from M. cordata, Sanguinaria
canadensis, and Chelidonium majus are known to exert a
wide spectrum of biological activities, such as antibacterial
[1, 2], anthelmintic [3], antitumour [4, 5], and anti-inflam-
matory properties [6, 7]. QBA fractions from M. cordata
(SANGUIRITRIN) and S. canadensis (SANGUINARIA)
are used in toothpastes and mouthwashes as antiplaque
agents. SANGUIRITRIN is applied as a veterinary prepa-
ration for mastoiditis in cows [8]. SA is an active component
of the preparation Sangrovit� as an additive to animal feeds.
Recently, SA has gained increasing attention as potential
agents in the treatment of cancer.
Most of the SA currently used in herbal supplements and
medicines is extracted from M. cordata and some other
species in the Papaveracea. Although M. cordata actually
has abundant SA content, and it has a rich resources, but
indiscriminate felling of trees will destroy the ecological
balance of nature. Thus, increasing efforts have been made
to develop alternative means of SA production, such as
using complete chemical synthesis and Papaveracea plant
cell culture. However, thus far, the total chemical synthesis
of SA still cannot be chemically synthesized in an indus-
trially feasible manner, and the in vitro culture of opium
poppy has only met limited success [9].
In recent years, endophytic fungi from plants have been
widely accepted as major sources of drugs, and a large
number of compounds with new structures and various
X.-J. Wang � C.-L. Min (&) � R.-H. Zuo
College of Biotechnology and Pharmaceutical Engineering,
West Anhui University, Lu’an 237012, China
e-mail: [email protected]
X.-J. Wang � C.-L. Min
Research Center for Endophytic Fungi Resources of Dabie
Mountain, West Anhui University, Lu’an 237012, China
M. Ge
Shanghai Laiyi Center of Biopharmaceutical R&D,
Shanghai 201203, China
123
Curr Microbiol (2014) 68:336–341
DOI 10.1007/s00284-013-0482-7

bioactivities are continuously being isolated from them [10].
Thus, if a microbial source of SA is available, there is no need
to harvest and extract the M. cordata for this drug. As far as
we know, little work has been done concerning the endo-
phytic fungi associated with M. cordata. Therefore, this
study aimed to isolate the endophytic SA-producing fungus
in M. cordata plants collected from Anhui Province of China
by rDNA ITS sequences analysis, and the potential of strain
BLH51 for SA production was also evaluated.
Materials and Methods
Materials
Solvents used for chromatography were of high-perfor-
mance liquid chromatography (HPLC) grade, while sol-
vents used for extraction were of American Chemical
Society grade. Authentic SA (C98 % purity) was obtained
from the National Institute for the Control of Pharmaceu-
tical and Biological Products of China. All other chemicals
were purchased from China Medicine Shanghai Chemical
Reagent Co., Ltd. Polymerase chain reaction (PCR) prim-
ers were synthesized by Shanghai Sangon Biologic Engi-
neering Technology and Service Co., Ltd.
Samples of M. cordata were obtained from the natural
populations at Dabie Mountain in Anhui Province in cen-
tral China. All samples were placed in polyethylene bags,
immediately transported to the laboratory, and placed in a
refrigerator at 4 �C.
Isolation of Endophytic Fungi from Macleaya cordata
Strain BLH51 used in this study was 1 of 55 endophytic fungi
isolated from the leaves of M. cordata. Healthy leaves of
M. cordata were thoroughly washed in running tap water, then
sterilized by washing in 70 % ethanol for 2 min and 0.5 %
sodium hypochlorite for 2 min, and followed by washing with
70 % ethanol (v/v) for 5 s. Afterwards, the leaves were rinsed
four times in sterile distilled water, and then they were cut into
pieces (0.5 9 0.5 cm). The small pieces were placed in petri
dishes (9 cm diameter) on the surface of potato dextrose agar
(PDA) medium containing 0.5 g/L streptomycin sulfate. Petri
dishes were incubated at 28 �C in darkness. After several days,
fungi were observed growing from the leaf fragments. Indi-
vidual hyphal tips of the various fungi were removed and
placed on fresh PDA medium and incubated at 28 �C for at
least 10 days. Each fungal culture was checked for purity and
transferred to another PDA plate by the hyphal tip method. The
fungal isolates were numbered and stored on the surface of
PDA plate at 4 �C or as spores and mycelia in 15 % (v/v)
glycerol at -70 �C.
Fermentation and Preparation of Endophytic Fungi
Extracts
The endophytic fungi isolates were inoculated, respectively,
into 500 mL Erlenmeyer flasks containing 100 mL of potato
dextrose liquid medium and cultured at 28 �C with 200
recycles/minute for 10 days in a rotary shaker. The mycelia
were harvested by centrifugation at 12,0009g for 10 min and
dried at 50 �C overnight. Dried mycelia were crushed and
extracted with 85 % ethanol under reflux for 1 h at 80 �C,
repeated four times, the extracts were filtered and the filtrates
were evaporated under reduced pressure. The dry residue was
dissolved in 50 mL 1 % sulfuric acid–water, and then
exhaustively extracted with chloroform. In the last stage, the
combined chloroform extracts were evaporated to dryness.
The dry residues were dissolved in 1 mL of methanol (HPLC
purity grade). The methanolic extracts were filtered through a
0.45 lm filter prior to chromatographic separation.
Screening of SA-Producing Endophytic Fungi
The SA-producing endophytic fungi were screened by thin
layer chromatography (TLC). TLC analysis of the metha-
nolic extract of endophytic fungi as well as the methanolic
solution of SA standard was developed in a solvent system
(petroleum ether: methanol: at 15:1 v/v) by spotting on a
0.25 mm (10 9 20 cm) silica gel plate. SA was detected
under 365 nm ultraviolet light, which appeared as yel-
lowish spots. The SA spot was identified by comigration
with authentic SA.
High-performance liquid chromatography (HPLC) was
performed using a C18 column (5 lm, 4.6 9 150 mm)
(Agilent, USA). A 20 lL amount of each methanolic
extract was injected. The mobile phase was acetonitrile:
water (25: 75, v/v) at a flow rate of 1.0 mL/min. The
effluent was monitored at 270 nm. SA was quantified by
comparing the peak area of the samples with that of the
authentic SA.
Identification of Strain BLH51
After SA was determined in the culture of strain BLH51, it
was identified by microscopic morphologic characteristics
and ITS sequence analysis. The strain BLH51 was grown
on the surface of PDA medium at 25 �C for 2 weeks,
followed by identification based on the morphology of the
fungal colony, and the characteristics of the spores.
Mycelia and conidia of strain BLH51 were observed with a
light microscope (BA300, Motic, China).
The genomic DNA of strain BLH51 was extracted from
fresh mycelia according to the method described by Pirttila
X.-J. Wang et al.: Producing Fungus from Macleaya cordata 337
123
et al. [11]. The target rDNA region including ITS1, ITS2
regions and 5.8S gene was amplified using primers ITS1 (50-TCCGTAGGTGAACCTGCGG-30) and ITS4 (50-TCCTCC
GCTTATTGATATGC-30) [12]. Total volume was brought
to 25 lL with deionised water. Amplification of the ITS
region was performed as follows: 95 �C for 3 min, followed
by 40 cycles of 94 �C for 60 s, 50 �C for 60 s, and 72 �C for
120 s and a final extension at 72 �C for 10 min. The PCR
products were purified using the Gel extraction kit (Watson,
China) and sequenced by Shanghai Sangon Biologic Engi-
neering Technology and Service Co., Ltd.
The ITS sequence of strain BLH51 was compared to the
data available in NCBI using BLASTn search to estimate
the phylogenetic relationships of the endophytic fungi. The
resulting sequences were aligned with the Clustal X soft-
ware [13] with gaps treated as missing data. The phylo-
genetic tree was constructed using the neighbor-joining
method [14] and the Kimura two-parameter distance cal-
culation in mega software version 3.1 [15]. The bootstrap
was 1,000 replications to assess the reliable level to the
nods of the tree.
Results
Isolation of Endophytic Fungi
We obtained 55 endophytic fungi isolates flourishing in the
leaves of M. cordata from Dabie Mountain, China. Based
on colonial characteristics, the endophytic fungi were
related to Fusarium spp., Aspergillus spp., Penicillium
spp., Leptosphaeria spp., and unidentified strains. These
results confirm host specificity and geographic structure
affect biological diversity of endophytic fungi [16, 17].
Screening of SA-Producing Endophytic Fungi
The extracts of fungal cultures were examined for the
presence of SA by TLC under UV illumination. Results
indicated that strain BLH51 showed positive results for SA
production in potato dextrose liquid medium, and one of
the fungal compounds exhibited the same Rf values (0.69)
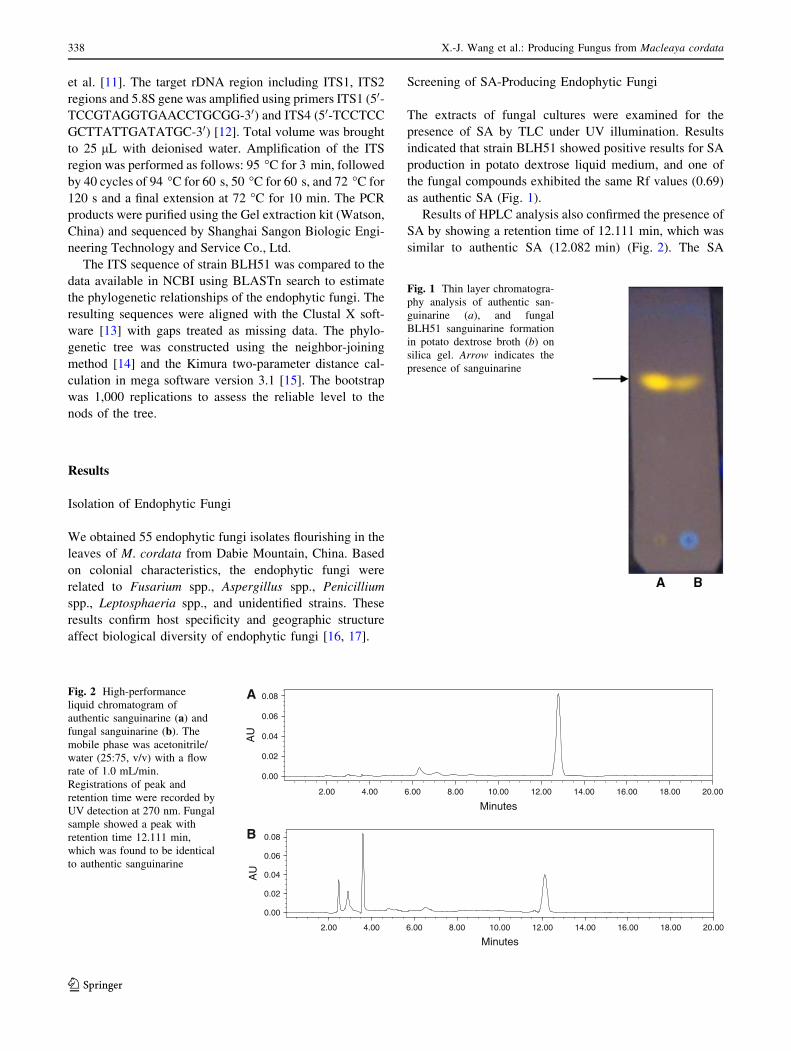
as authentic SA (Fig. 1).
Results of HPLC analysis also confirmed the presence of
SA by showing a retention time of 12.111 min, which was
similar to authentic SA (12.082 min) (Fig. 2). The SA
Fig. 1 Thin layer chromatogra-
phy analysis of authentic san-
guinarine (a), and fungal
BLH51 sanguinarine formation
in potato dextrose broth (b) on
silica gel. Arrow indicates the
presence of sanguinarine
AU
0.00
0.02
0.04
0.06
0.08
Minutes2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00
AU
0.00
0.02
0.04
0.06
0.08
Minutes2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00
A
B
Fig. 2 High-performance
liquid chromatogram of
authentic sanguinarine (a) and
fungal sanguinarine (b). The
mobile phase was acetonitrile/
water (25:75, v/v) with a flow
rate of 1.0 mL/min.
Registrations of peak and
retention time were recorded by
UV detection at 270 nm. Fungal
sample showed a peak with
retention time 12.111 min,
which was found to be identical
to authentic sanguinarine
338 X.-J. Wang et al.: Producing Fungus from Macleaya cordata
123
yield of strain BLH51 was about 178 lg/L or 135 lg/g (SA
per dry wt of mycelium) when it was cultured in 100 mL
potato dextrose liquid medium at 28 �C with 200 recycles/
minute shaking for 10 days. Our results showed the fungal
endophyte clearly produced SA.
Phenotypic Characterization of Strain BLH51
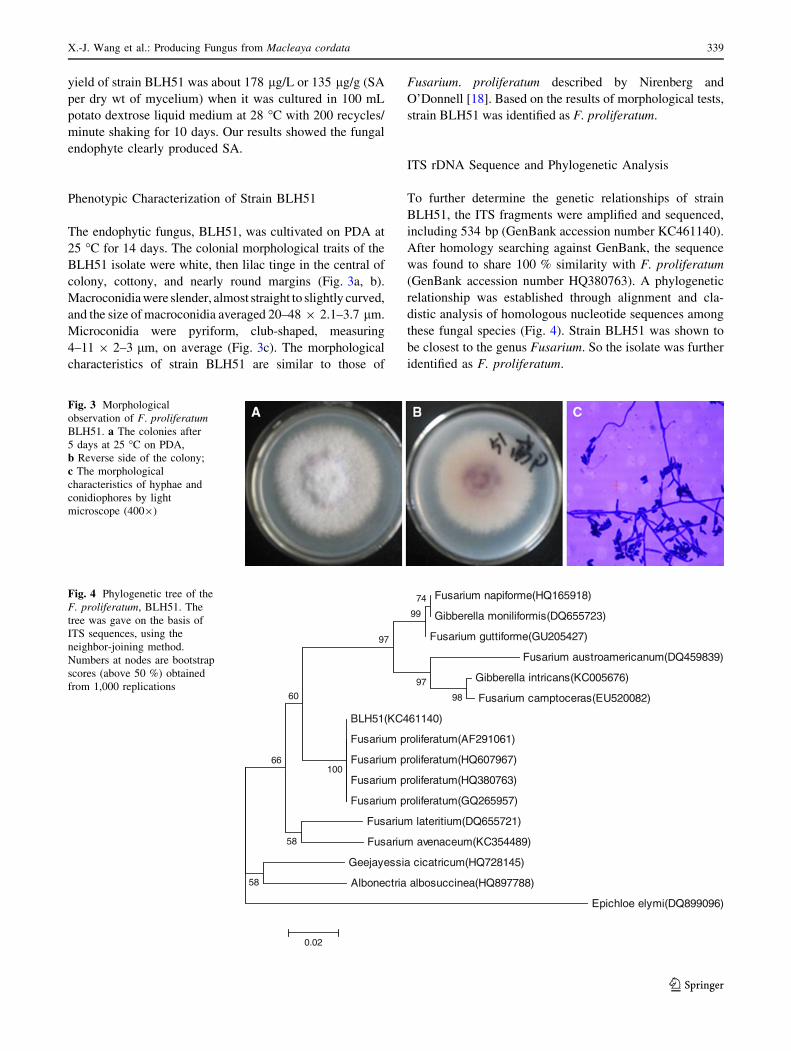
The endophytic fungus, BLH51, was cultivated on PDA at
25 �C for 14 days. The colonial morphological traits of the
BLH51 isolate were white, then lilac tinge in the central of
colony, cottony, and nearly round margins (Fig. 3a, b).
Macroconidia were slender, almost straight to slightly curved,
and the size of macroconidia averaged 20–48 9 2.1–3.7 lm.
Microconidia were pyriform, club-shaped, measuring
4–11 9 2–3 lm, on average (Fig. 3c). The morphological
characteristics of strain BLH51 are similar to those of
Fusarium. proliferatum described by Nirenberg and
O’Donnell [18]. Based on the results of morphological tests,
strain BLH51 was identified as F. proliferatum.
ITS rDNA Sequence and Phylogenetic Analysis
To further determine the genetic relationships of strain
BLH51, the ITS fragments were amplified and sequenced,
including 534 bp (GenBank accession number KC461140).
After homology searching against GenBank, the sequence
was found to share 100 % similarity with F. proliferatum
(GenBank accession number HQ380763). A phylogenetic
relationship was established through alignment and cla-
distic analysis of homologous nucleotide sequences among
these fungal species (Fig. 4). Strain BLH51 was shown to
be closest to the genus Fusarium. So the isolate was further
identified as F. proliferatum.
Fig. 3 Morphological
observation of F. proliferatum
BLH51. a The colonies after
5 days at 25 �C on PDA,
b Reverse side of the colony;
c The morphological
characteristics of hyphae and
conidiophores by light
microscope (4009)
Fusarium napiforme(HQ165918)
Gibberella moniliformis(DQ655723)
Fusarium guttiforme(GU205427)
Fusarium austroamericanum(DQ459839)
Gibberella intricans(KC005676)
Fusarium camptoceras(EU520082)
BLH51(KC461140)
Fusarium proliferatum(AF291061)
Fusarium proliferatum(HQ607967)
Fusarium proliferatum(HQ380763)
Fusarium proliferatum(GQ265957)
Fusarium lateritium(DQ655721)
Fusarium avenaceum(KC354489)
Geejayessia cicatricum(HQ728145)
Albonectria albosuccinea(HQ897788)
Epichloe elymi(DQ899096)
74
99
98
97
97
100
60
58
66
58
0.02
Fig. 4 Phylogenetic tree of the
F. proliferatum, BLH51. The
tree was gave on the basis of
ITS sequences, using the
neighbor-joining method.
Numbers at nodes are bootstrap
scores (above 50 %) obtained
from 1,000 replications
X.-J. Wang et al.: Producing Fungus from Macleaya cordata 339
123

Discussion
This study isolated a SA-producing endophytic fungus
BLH51 from the leaves of M. cordata obtained from Dabie
Mountain in Anhui Province in central China. Strain BLH51
was identified as F. proliferatum on the basis of its mor-
phology and the ITS sequence. Previous studies demonstrated
that Fusarium spp. are distributed worldwide, and are fre-
quently obtained from some trees and numerous crop plants
[19–21]. The genus Fusarium includes many species, with F.
proliferatum as one of the most common species. Fusarium
spp. was reported to produce several secondary metabolites,
including cyclic tetrapeptides, rohitukine, taxol, and other
antibiotics [19, 21–23]. To the best of our knowledge, the
endophytic fungus Fusarium spp. and other genus have never
been reported capable of producing SA. The present study is
the first to isolate, characterize, and identify SA-producing F.
proliferatum from Papaveraceae plants in China.
The discovery of SA-producing endophytic fungi asso-
ciated with Papaveraceae plants is valuable for industrial
interest and for basic research. The consistent production of
SA by F. proliferatum BLH51 further supports the theory
that during the long coevolution of endophytes and their host
plants, endophytes adapted to their special microenviron-
ments by genetic variation, including uptake of some plant
DNA into their own genomes [24]. This could have led to the
ability of certain endophytes to biosynthesize some phyto-
chemicals originally associated with the host plant [25].
The total amount of SA produced by F. proliferatum
BLH51 was 178 lg/L or 135 lg/g (SA per dry wt of
mycelium) when it is cultured under the conditions described
in this study. This suggests that F. proliferatum BLH51 is a
promising candidate for SA production. However, it is only a
wild strain, and we believe that the SA yield of F. prolifer-
atum BLH51 will be increased by strain improvement and
optimization of the fermentation media.
Acknowledgments This work was supported by National Natural
Science Foundation of China (Grant No. 31100019) and Anhui Nat-
ural Science Research Project of Colleges and Universities of China
(Grant Nos. KJ2013A266 and KJ2011Z398). We thank Prof. Yun-
jiang Min from College of Biotechnology and Pharmaceutical Engi-
neering, West Anhui University, China for helping with the identifi-
cation of the plant material. We also wish to thank the anonymous
reviewers for constructive comments.
References
1. Beuria TK, Santra MK, Panda D (2005) Sanguinarine blocks
cytokinesis in bacteria by inhibiting FtsZ assembly and bundling.
Biochemistry 44:16584–16593
2. Lovasoa RR, Havet JL, Catherine P, Henri F (2007) Solid-liquid
extraction of protopine from Fumaria officinalis L.-Analysis
determination, kinetic reaction and model building. Sep Purif
Technol 54:253–261
3. Satou T, Akao N, Matsuhashi R, Koike K, Fujita K, Nikaido T
(2002) Inhibitory effect of isoquinoline alkaloids on movement of
second-stage larvae of Toxocara canis. Biol Pharm Bull
25:1651–1654
4. Bai LP, Zhao ZZ, Cai ZW, Jiang ZH (2006) DNA-binding
affinities and sequence selectivity of quaternary benzophenan-
thridine alkaloids sanguinarine, chelerythrine, and nitidine. Bio-
org Med Chem 14:5439–5445
5. Luo XB, Chen B, Yao SZ (2005) Rapid determination of proto-
pine, allocryptopine, sanguinarine and chelerythrine in fruits of
Macleaya cordata by microwave-assisted solvent extraction and
HPLC-ESI/MS. Phytochem Anal 17:431–438
6. Saeed SA, Gilani AH, Majoo RU, Shah BH (1997) Anti-throm-
botic and anti-inflammatory activities of protopine. Pharmacol
Res 36:1–7
7. Zhou HY, Mineshita S (2000) The effect of berberine chloride on
experimental colitis in rats in vivo and in vitro. J Pharmacol Exp
Ther 294:822–829
8. Faddeeva MD, Beliaeva TN (1997) Sanguinarine and ellipticine
cytotoxic alkaloids isolated from well-known antitumor plants.
Intracellular targets of their action. Tsitologiia 39:181–208
9. Alcantara J, Bird DA, Franceschi VR, Facchini PJ (2005) San-
guinarine biosynthesis is associated with the endoplasmic retic-
ulum in cultured opium poppy cells after elicitor treatment. Plant
Physiol 138:173–183
10. Strobel G, Daisy B, Castillo U, Harper J (2004) Natural products
from endophytic microorganisms. J Nat Prod 67:257–268
11. Pirttila AM, Kamarainen T, Hirsikorpi M, Jaakola L, Hohtola A
(2001) DNA isolation methods for medicinal and aromatic plants.
Plant Mol Biol Rep 19:273a–273f
12. White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and
direct sequencing of fungal ribosomal RNA genes for phyloge-
netics. In: Innis MA, Gelfland DH, Sninsky JJ, White TJ (eds)
PCR protocols: a guide to methods and applications. Academic
Press, New York
13. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins
DG (1997) The Clustal_X windows interface: flexible strategies
for multiple sequence alignment aided by quality analysis tools.
Nucleic Acids Res 25:4876–4882
14. Saitou N, Nei M (1987) The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol Biol Evol
4:406–425
15. Kumar S, Tamura K, Nei M (2004) MEGA3: integrated software
for molecular evolutionary genetics analysis and sequence
alignment. Brief Bioinform 5:150–163
16. Higgins KL, Arnold AE, Miadlikowska J, Sarvate SD, Lutzoni F
(2007) Phylogenetic relationships, host affinity, and geographic
structure of boreal and arctic endophytes from three major plant
lineages. Mol Phylogenet Evol 42:543–555
17. Strobel G, Daisy B (2003) Bioprospecting for microbial endo-
phytes and their natural products. Microbiol Mol Biol Rev
67:491–502
18. Nirenberg HI, O’Donnell K (1998) New Fusarium species and
combinations within the Gibberella fujikuroi species complex.
Mycologia 90:434–458
19. Deng BW, Liu KH, Chen WQ, Ding XW, Xie XC (2009)
Fusarium solani, Tax-3, a new endophytic taxol-producing fun-
gus from Taxus chinensis. World J Microbiol Biotechnol
25:139–143
20. Marın S, Magan N, Serra J, Ramos AJ, Canela R, Sanchis V
(1999) Fumonisin B1 production and growth of Fusarium moni-
liforme and Fusarium proliferatum on maize, Wheat, and Barley
Grain. J Food Sci 64:921–924
21. Mohana Kumara P, Zuehlke S, Priti V, Ramesha BT, Shweta S,
Ravikanth G, Vasudeva R, Santhoshkumar TR, Spiteller M, Uma
Shaanker RU (2012) Fusarium proliferatum, an endophytic
340 X.-J. Wang et al.: Producing Fungus from Macleaya cordata
123

fungus from Dysoxylum binectariferum Hook.f, produces rohi-
tukine, a chromane alkaloid possessing anti-cancer activity. An-
tonie Van Leeuwenhoek 101:323–329
22. Cheng ZS, Tang WC, Su ZJ, Cai Y, Sun SF, Chen QJ, Wang FH,
Lin YC, She ZG, Vrijmoed LLP (2008) Identification of man-
grove endophytic fungus 1403 (Fusarium proliferatum) based on
morphological and molecular evidence. J For Res 19:219–224
23. Jin JM, Baek SR, Lee KR, Lee JK, Yun SH, Kang SC et al (2008)
Purification and phytotoxicity of apicidins produced by the
Fusarium semitectum KCTC16676. Plant Pathol J 24:417–422
24. Germaine K, Keogh E, Garcia-Cabellos G, Borremans B, Barac
T, Dowling DN et al (2004) Colonisation of poplar trees by gfp
expressing bacterial endophytes. FEMS Microbiol Ecol
48:109–118
25. Stierle A, Strobel G, Stierle D (1993) Taxol and taxane produc-
tion by Taxomyces andreanae, an endophytic fungus of pacific
yew. Science 260:214–216
X.-J. Wang et al.: Producing Fungus from Macleaya cordata 341
123





























