Embed Size (px)
Citation preview

Journal of Immunological Methods 330 (2008) 75–85www.elsevier.com/locate/jim
Research paper
An enzyme-linked immunosorbent assay (ELISA) for quantificationof mouse surfactant protein D (SP-D)
Soren Hansen a,⁎, Vivi Schmidt a, Maria Abildgaard Steffensen b, Pia Hoennerup Jensen c,Morten Gjerstorff b, Steffen Thiel d, Uffe Holmskov b
a Department of Immunology & Microbiology, University of Southern Denmark, Winsloewparken 21-1, DK-5000 Odense, Denmarkb Medical Biotechnology Center, University of Southern Denmark, Odense, Denmark
c Department of Biochemistry and Molecular Biology, University of Southern Denmark, Odense, Denmarkd Department of Medical Microbiology and Immunology, University of Aarhus, Aarhus, Denmark
Received 31 August 2007; received in revised form 5 October 2007; accepted 1 November 2007
Abstract
Surfactant protein D (SP-D) is a pattern recognition molecule of the collectin family of C-type lectins. It is found in the airways and atmucosal surfaces. SP-D is part of the innate immune systemwhere it neutralizes and leads to elimination of microorganisms. It regulates thefunctions of other innate immune cells, such asmacrophages and neutrophils. It alsomodulates the adaptive immune response by interactingwith antigen-presenting cells and Tcells.Monoclonal anti-mouse-SP-D antibodies were raised fromSP-D deficient mice using recombinantSP-D as antigen. Ten monoclonal antibodies were characterized and validated for use in sandwich enzyme-linked immunosorbent assay(ELISA). Based on two of these, we established an ELISA that allows for measurements of mouse SP-D in various body fluids. The finalELISAwas optimized and calibrated with a standard of purified recombinant mouse SP-D, which was analyzed by quantitative amino acidanalysis. The ELISAwas validated with respect to parallelism, recovery, intra- and inter-assay variation. The practical working range wasestimated to be 1.9–200 ng/ml. We measured SP-D in lung lavage to an average concentration of 435 ng/ml (3-ml lavage), and in mousevaginal fluid (1-ml lavage) to an average concentration of 94 ng/ml, but could not detect SP-D in the serum or plasma of healthy mice(b3.8 ng/ml). With this ELISA, it will be possible to study the regulation of SP-D in various mouse models and upon various stimuli.© 2007 Elsevier B.V. All rights reserved.
Keywords: Surfactant; Collectin; SP-D; Mouse; ELISA; Assay
1. Introduction
Lung surfactant protein D (SP-D) is a collagenous C-type lectin (collectin), which participates in the first, innateline of immune defence and subsequently modulates both
Abbreviations: CTLD, C-type lectin domain; SP-D, surfactantprotein D.⁎ Corresponding author. Tel.: +45 6550 3062; fax: +45 6591 5267.E-mail address: [email protected] (S. Hansen).
0022-1759/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jim.2007.11.001
the degree of inflammation and the adaptive immuneresponse (Wright, 2005; Hansen et al., 2007). SP-D is apattern recognition molecule, which recognizes and bindsto foreign patterns of glycoconjugates found onmicroorganisms, particulate antigens and apoptotic cells(Erpenbeck et al., 2005; Stuart et al., 2006). SP-D alsoparticipates in the homeostasis of phospholipids in the lung(Ikegami, 2006). Traditionally, SP-Dhas been thought of asa protein associated exclusively with lung surfactant andlung immunology but recent results suggest that SP-D

76 S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
exerts similar functions at extra pulmonary sites (Madsenet al., 2000; Leth-Larsen et al., 2004; Sorensen et al., 2006;Herias et al., 2007). Structurally the polypeptide chains ofSP-D and other collectins are composed of a collagen-likeregion and a C-terminal C-type lectin domain (CTLD).Polypeptide chains trimerize into subunits via interactionsin an alpha-helical coiled-coil neck and in the collagen-likeregion. Subunits oligomerize further by formation of inter-subunit di-sulfide bonds between cystein residues in the N-termini (Hansen and Holmskov, 1998; Holmskov et al.,2003). The highly organized structural clustering ofCTLDs sustains the necessary specificity and avidity ofSP-D to selectively recognize and bind to foreign or mo-dified glycoconjugates with a relatively high content ofmannose-like carbohydrates; and it thus establishes SP-Das a pattern recognition receptor. SP-D is conserved evolu-tionarily and found in various mammals, and some specieshave even evolved additional SP-D-like proteins (Hansenand Holmskov, 2002). In humans SP-D is likely to leakfrom the lung into the blood and depending on the twodifferent commercially available assays, serum levels arebetween 0.2 and 5.2 μg/ml or between 0.04 and 0.1 μg/ml,respectively (Leth-Larsen et al., 2003; Hartl and Griese,2006). Single nucleotide polymorphisms that lead to struc-tural variants of the N-termini and differences in averageserum levels have led to conflicting conclusions in terms ofthe chance of acquiring tuberculosis or bronchiolitis uponinfection with respiratory syncytical virus (Floros et al.,2000; Lahti et al., 2002; Leth-Larsen et al., 2005).
In association with pneumonia, the serum levels of SP-D are highest at day 5 of an infection, with hospitalizationand antibiotic treatment. The levels are increased 2–3-foldalthough there is great variability among patients. The peakof SP-D serum concentration occurs 4–5 days after C-reactive protein reaches its maximum, and it declines moreslowly than both C-reactive protein and total bloodleukocyte count (Leth-Larsen et al., 2003). Studies inrodents have shown that SP-D levels increase upon briefinhalations of air with a high oxygen content (95%) andupon stimulation with either IL-4, IL-13 or TNF-α(Aderibigbe et al., 1999; Ikegami et al., 2000; Homeret al., 2002; Fujita et al., 2005). The transcriptionalregulation of SP-D is only poorly understood. In vitrocell culture studies have shown that SP-D transcription isincreased upon stimulationwith glucocorticoids (Dulkerianet al., 1996; Rust et al., 1996).
In association with many human lung diseases andsyndromes, lavaging the lungs is impossible or associatedwith great risk. Hence, observations of SP-D regulation areoften based on the levels in blood. However, such studiesmay neither reflect the actual systemic regulation northe transcriptional regulation of SP-D in the lung. Lung
deterioration and subsequent increased permeability due todisease pathology are important aspects to consider. Insituations where the lungs can be lavaged safely, inter-personal comparison can be hard to perform due to:individual and practical difficulties of performing thelavage, different accessibility of the lung segments andgenetic differences. Obtaining lavage from mice in mousemodels of lung diseases or syndromes is a good alternativebut until now, SP-D levels have, with only a fewexceptions (Zhang et al., 2003; Fujita et al., 2005), beenbased on themRNA levels in the lung epithelia or based onsemi-quantitative estimates by Western blotting. Here wedescribe the development of an enzyme-linked immuno-sorbent assay (ELISA) for the measurement of mouse SP-D. With this novel ELISA, it is possible to study theregulation of SP-D in the mouse lung upon variousinfections, stimuli or models of human diseases (i.e.respiratory distress syndrome, acute respiratory distresssyndrome or lung intestinal diseases). It is also possible toaccess the leakage of SP-D from the lung to the blood uponthe same stimuli and diseases, and begin to access theimmuno-regulatory role of SP-D at extra pulmonary sitesin the relevant mouse models. The current ELISA isvalidated for measuring mouse SP-D in different bodyfluids with a working range of 1.9 to 200 ng/ml.
2. Materials and methods
2.1. Reagents, buffers, solutions and general ELISAprocedures
Unless otherwise stated reagents were obtained fromSigma-Aldrich Denmark, Brondby, Denmark.
Coating buffer, 15 mM Na2CO3, 35 mM NaHCO3,pH 9.6; substrate buffer, 50 mM citric acid titrated to pH5.0 with 1 M Na2HPO4; TBS, 10 mM Tris, 150 mMNaCl, pH 7.4; TBS/Ca2+/Tw, TBS with 1 mMCaCl2 and0.01% Tween 20, pH 7.4; TBS/EDTA/Tw, TBS with2 mMEDTA and 0.01% Tween 20; washing buffer, TBSwith 5 mM CaCl2 and 0.05% Tween 20.
Unless otherwise stated, detailed procedures of opti-mizing the ELISA and characterizing antibodies followedthe principles of the final ELISA described in Section 2.9.
2.2. Recombinant expression of mouse SP-D
A recombinant protein representing the neck/CTLDof mouse SP-D was expressed in Pichia pastoris GS115using the multi-copy gene plasmid pPIC9K (Invitrogen,Taastrup, Denmark), which provides the ability to growon histidine drop-out plates and resistance towards theantibiotic G418. Cloning took advantage of the SnaBI

77S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
and AvrII sites. Transformation and selection were per-formed according to the manufacturer's recommenda-tions. A clone with the ability to grow in the presence of1 mg of G418/ml was selected for further characteriza-tion, and after 5 days with methanol induction this cloneproduced approximately 15 μg mouse SP-D neck/CTLD per ml of minimal media. The recombinant andsecreted neck/CTLD of mouse SP-D comprised thefollowing amino residues: (EAEA)YV dsa alr qqm ealkgk lqr lev afs hyq kaa lfp dgr svg dki frt ads ekp fed aqemck qag gql asp rsa ten aai qql ita hnk aaf lsm tdv gtegkf typ tge plv ysn wap gep nnn gga enc vei ftn gqw ndkacg eqr lvi cef, where capital letters refer to vectorderived residues. Signal peptide retention and cleavageupon secretion is not consistent and the N-terminalresidues in parentheses indicate this variation. Recom-binant full-length mouse SP-D expressed in CHO cellswas a generous gift from Professor Jo Rae Wright,Department of Cell Biology, Duke University MedicalCenter, Durham, NC, USA.
2.3. Purification of mouse SP-D
Recombinant neck/CTLD mouse SP-D was expressedin Pichia for 5 days (see above). To chelate calcium ionsand release SP-D bound to the yeast, EDTAwas added tothe culture to a final concentration of 5 mM and incubatedat room temperature for 30 min. The culture was centri-fuged at 8000 ×g for 10 min at 4 °C and the supernatantfiltered through 44 μm filters. Formic acid was added to thesupernatant to a final concentration of 50 mM, the pH wasadjusted to 4.0 and the sample was loaded onto a SP-Sepharose column (Amersham Pharmacia Biotech, 20 mlbeads/l of culture). The columnwas washedwith 200ml ofbuffer made of 50 mM formic acid, 0.5 mMEDTA, 0.01%Tween 20 (pH 4.0) and eluted with a similar buffer with800 mM NaCl. The eluate was dialyzed against TBS andsubsequently against TBS/Ca2+/Tw. The dialysate wasloaded onto a maltose agarose column (5 ml/l of culture),and after washing with 200 ml TBS/Ca2+/Tw the columnwas eluted with TBS/EDTA/Tw. Full-length mouse SP-Dexpressed in CHO cells was purified by maltose affinitychromatography as previously described (Strong et al.,1998). Purity was estimated by SDS-PAGE and theconcentration was measured by quantitative amino acidanalysis.
2.4. Mouse monoclonal anti-mouse-SP-D antibodies
Four SP-D deficient mice on C57 black background(Botas et al., 1998) were immunized s.c. with 30 μg ofpurified mouse neck/CRD SP-D. The first immunization
was done using Freund's complete adjuvant, whereasthe three following s.c. immunizations at 3 week inter-vals were done using incomplete Freund's adjuvant. Ahigh responder mouse, as estimated by means of ELISAusing coating with neck/CTLD SP-D, was selected forsplenectomy; 3 days before the splenectomy, the mousewas immunized intravenously with 30 μg of the im-munogen in sterile TBS without any adjuvant. Spleno-cytes were fused with the SP2 cell line as previouslydescribed by Kohler and Milstein (1975). Selection foraminopterin sensitivity was done in the absence offeeder cells and began 24 h after the fusion. Hybridomaswere screened using antigen coated in 96-well plates(2 μg/ml) and bound antibodies were detected usingHRP-conjugated rabbit-anti-mouse-Ig (Dako Norden,Glostrup, Denmark). Hybridomas were cloned by limit-ing dilution and monoclonal antibodies were expressedin Hybridoma-SFM (Invitrogen) with 0.75% FCS, witha defined low content of bovine immunoglobulins(Invitrogen). Monoclonal antibodies were purified bymeans of affinity chromatography using a HiTrap Pro-tein G HP column (GE Healthcare) using previouslydescribed conditions and eluted with 50 mM glycine(pH 2.3) (Akerstrom and Bjorck, 1986).
2.5. SDS-PAGE
SDS-PAGE was done in a discontinuous buffersystem on pre-cast 4–12% gradient gels (Invitrogen)using the principles described by Laemmli (1970), andthe MES buffer system described by the manufacturer.Proteins were either stained with Coomassie BrilliantBlue R250 or with silver nitrate using a modifiedversion of the method developed by Nesterenko andcolleagues (Nesterenko et al., 1994; Hansen et al.,2000). Molecular weights were estimated by compar-ison with pre-stained marker proteins (Mark 12 fromInvitrogen).
2.6. Biotinylation of antibodies
Purified antibodies were biotinylated by means oftwo different methods to ensure optimal reactivity uponlabelling. Lysine residues were targeted by traditionalNHS-esters of biotin using biotin N-hydroxysuccini-mide ester according to the manufacturer's recommen-dations (Sigma). Glycosylations on the Fc portion of theantibodies were targeted after mild oxidation (to allowformation of aldehyde groups) with Biotin LC-hydra-zide (Pierce, Rockford, USA) according to the manufac-turer's recommendations using the principle describedby O'Shannessy et al. (1984).

78 S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
2.7. Characterization of monoclonal antibodies andselection for assay application
The relative affinity of each monoclonal antibody wasinvestigated by coating 96-well plates with a 2-fold serialdilution of neck/CTLD of mouse SP-D in coating buffer.Dilutions spanned from 2 μg/ml to 0.97 ng/ml. A low-density coating of antigen allows relative estimation of theinteraction between a single binding site of an antibodywith a single antigen epitope, and hence avoids avidityinterference due to multiple interactions. The monoclonalantibodies were applied to the wells at 2 μg/ml in washingbuffer with 1mgHSA/ml, and bindingwas detected usingsecondary HRP-anti-mouse-Ig (Dako Norden).
Calcium dependency was analyzed by coating of neck/CTLD of mouse SP-D (2 μg/ml) in 96-well plates andincubation with serial dilutions of biotinylated monoclonalantibodies in either washing buffer with 1mgHSA/ml or inTBS with 5 mM EDTA and 1 mg HSA/ml. Boundantibodies were detected by means of streptavidin-HRP.
Competitive analysis was performed to avoid thecatching and detecting antibody reacting with the sameepitope. For this, neck/CTLDmouse SP-D was coated at2 μg/ml and the biotinylated monoclonal antibodies werediluted into buffer containing an unlabelled monoclonalantibody at a concentration of 10 μg/ml. All combina-tions of unlabelled monoclonal antibodies vs. biotiny-lated monoclonal antibodies were tested separately.
Pair-wise optimizationwas used to analyze all combina-tions of coating and biotinylated detecting antibodies. Twomicrograms of antibody permillilitre of coating bufferwerecoated in 96-well plates and incubated with serial dilutionsof lung lavage, supernatant fromCHOcells expressing full-length mouse SP-D, or the neck/CTLD of mouse SP-Dexpressed in Pichia. All these combination of antigenswere analyzed to evaluate and ensure reactivity with nativeSP-D. Biotinylated monoclonal antibodies were used at aconcentration of 2 μg/ml of washing buffer with 1 mgHSA/ml, and streptavidin conjugated HRP and OPD wasused for the final development. Combinations of antibodieswere evaluated by their performance of recognizing bothnative and recombinant mouse SP-D, and finally by theirsignal-to-noise ratio.
2.8. Optimization
The assay was optimized according to the proceduresdescribed by Crowther (2000). All optimization stepswere evaluated by the signal-to-noise ratio. The opti-mization comprised: (a) various coating conditions andconcentrations of MAb 432; (b) various blocking con-ditions and reagents; (c) various buffers for incubating
samples; and (d) various concentrations of biotinylatedMAb 403 and streptavidin conjugated HRP.
2.9. ELISA for mouse SP-D
The assay is a non-competitive indirect sandwichELISA using MAb 432 for coating, biotinylated MAb403 (conjugated with hydrazine biotin) and streptavidinconjugated HRP for detection, and finally OPD conver-sion as the read-out parameter for enzyme activity.Certified 96-well Maxisorb plates (Nunc, Roskilde,Denmark) were coated overnight at 4 °C with 100 μl of2.0 μg MAb 432/ml coating buffer. Wells were emptied,and incubated for 1 h at room temperature with 200 μl ofwashing buffer with 1 mg HSA/ml. Wells were emptied,washed four times in washing buffer and incubatedovernight at 4 °C with 100 μl of calibrators (see below),quality controls (see below) or samples diluted inwashing buffer with 1 mg HSA/ml with an additional1 M NaCl. Wells were emptied, washed four times withwashing buffer and incubated for 1 h at room tempera-ture with 100 μl of 0.8 μg biotinylated MAb 403/mlwashing buffer with 1 mg HSA per ml. Wells wereemptied, washed four times in washing buffer and incu-bated at room temperature with 100 μl of 60 ng strep-tavidin conjugated HRP (Zymed by Invitrogen)/mlwashing buffer with 1 mg HSA/ml. Wells were emptied,washed four times and incubated at room temperaturefor 20 min with 100 μl of OPD (0.8 mg/ml, Kementec,Taastrup, Denmark) in substrate buffer with 0.03%H2O2, according to the manufacturer's recommenda-tions. Development was stopped by adding 100 μl of1 M H2SO4 and plates were read at OD 492 nm with OD600 nm as subtractive reference.
The calibratorswere derived fromCHOcells expressingfull-length mouse SP-D grown in the presence of completemedium (HAM-12, Invitrogen) with 10% FCS. Thesupernatant from these cells was diluted in washing bufferwith 1 mg HSA/ml. The highest calibrating concentrationused was 200 ng/ml and from this dilution a 7-step, 1.95-fold serial dilution was made. The resulting calibratingsamples comprised the following concentrations: 200 ng/ml, 103 ng/ml, 52.6 ng/ml, 27.0 ng/ml, 13.8 ng/ml, 7.09 ng/ml, 3.64 ng/ml, and 1.87 ng/ml. Calibrating and qualitycontrol samples were analyzed in a triplicate setup. A ‘fourparameter fit’model was applied to the calibrating samplesand used for estimating the concentration of unknownsamples. Three quality controls were included in the assay.Similar to the calibrating samples, they were derived fromCHO cells expressing full-length mouse SP-D. They weremade once and stored in aliquots of 350 μl at −20 °C, andwere ready for thawing and assaying without any further

Fig. 1. SDS-PAGE of recombinant mouse SP-D. The neck/CTLD(19 kDa) of SP-D was expressed in Pichia and full-length SP-D in CHOcells. Recombinant proteins were purified by means of carbohydrateaffinity chromatography and analyzed on 4–12% acrylamide gels. Proteinswere detected by means of Coomassie brilliant blue or silver staining. R,reduced; U, unreduced.
79S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
dilution at the following concentrations: 160, 19.4 and3.16 ng/ml. The assay was only accepted when themean ofthe measured triplicate quality control values diverged lessthan 10% from the expected values, and when the CVs ofthe triplicate estimates were also less than 10%. In general,lung lavage with a total volume of 3 ml was diluted 4–6-fold before analysis, vaginal lavage with a total volume ofapproximately 1 ml was diluted 3–4-fold, whereas plasmaand serum samples (and other samples) were diluted withan equal volume of buffer.
2.10. Assay validation
2.10.1. Parallelism and linearityDilutions of lung lavage, supernatant from CHO cells
expressing full-length mouse SP-D and mouse serum andplasma spiked with purified full-length mouse SP-D wereanalyzed in 2-fold dilutions at eight different concentra-tions. In the absence of spiking with purified SP-D, serumand plasma samples did not give any signals.
2.10.2. Detection limit and absolute correlationThe minimal detection limit was determined as the
average of ten zero samples plus two standard deviations.An absolute correlation of signal vs. concentration (ng/ml)was determined by diluting purified full-length mouse SP-D and a calibrator of the supernatant of CHO cellsexpressing the same protein. This calibrator was used forthe latter studies. The purified preparation had previouslybeen analyzed by quantitative amino acid analysis.
The working range was determined by triplicatedilutions of lung lavage samples and supernatant ofCHO cells expressing full-length mouse SP-D, serum andplasma spiked with purified SP-D, representing concen-trations ranging from 0.14 ng/ml to 260 ng/ml. The finalrange was determined by combined evaluation of plotting‘measured over mean’ as a function of the concentrationand by plotting CVs as a function of the concentration (seeSection 3).
Intra- and inter-assay variation was determined bydilution of pools of lung lavage samples and triplicatetesting on either the same plate or different plates, ac-cording to the ELISA conditions described above.Samples were diluted to span the entire working range.
3. Results
3.1. Recombinant expression
Recombinant mouse SP-D was produced in both P.pastoris and CHO cells. Two constructs representingeither the neck/CTLD alone or full-length SP-D were
made and purified to a high degree of purity by sugaraffinity chromatography (Fig. 1). The final yields afterpurification were approximately 4 and 1 μg/ml for thePichia and CHO system, respectively (not shown). Theneck/CTLD of SP-D served as antigen source for theproduction of monoclonal antibodies and the recombi-nant full-length SP-D for characterizing antibodies andfor validating the ELISA.
3.2. Characterization of monoclonal antibodies
Ten hybridomas were cloned by limiting dilution andthe reactivity of their secreted monoclonal antibodiescharacterized (Fig. 2). The relative affinity of each mono-clonal antibody was analyzed using a low-density coatingof antigen. Nine out of the ten monoclonal antibodiesbound coated neck/CTLD of SP-D with sufficient affinity(Fig. 2A). MAb 408, which failed, recognized and boundsufficiently (ODN0.5) only when more than 2 μg/ml wasapplied, and it was excluded for the subsequent analysis.All antibodies were of the IgG1kappa isotype (not shown).To determine which monoclonal antibodies had over-lapping epitopes a relative epitope analysis was performed.All monoclonal antibodies were biotinylated and analyzedseparately by inhibition ELISA using coated antigen andexcessive amounts of unlabelled antibody to inhibit(Fig. 2B). Even though two different biotin conjugationmethods were applied, some antibodies (MAb 445 and452) failed or reacted only poorly with the antigen afterbiotinylation. The analysis ofMAb 403 is shown in Fig. 2Band shows that the recognized epitope of MAb 403 over-laps with the epitopes recognized by MAb 404, 442 and
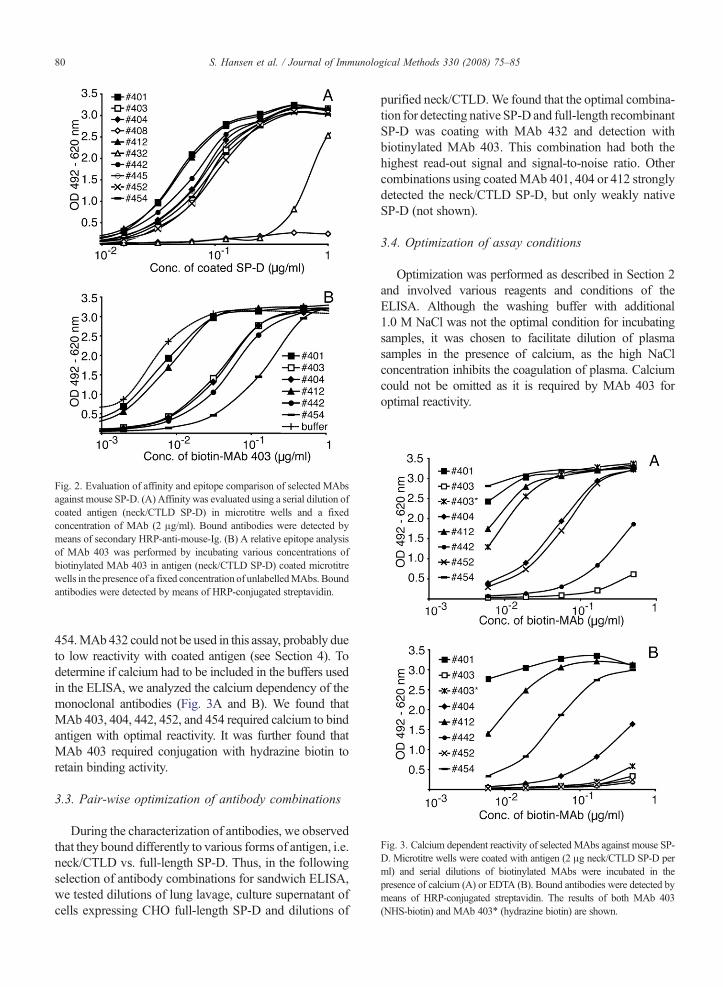
Fig. 2. Evaluation of affinity and epitope comparison of selected MAbsagainst mouse SP-D. (A) Affinity was evaluated using a serial dilution ofcoated antigen (neck/CTLD SP-D) in microtitre wells and a fixedconcentration of MAb (2 μg/ml). Bound antibodies were detected bymeans of secondary HRP-anti-mouse-Ig. (B) A relative epitope analysisof MAb 403 was performed by incubating various concentrations ofbiotinylated MAb 403 in antigen (neck/CTLD SP-D) coated microtitrewells in the presence of a fixed concentration of unlabelledMAbs. Boundantibodies were detected by means of HRP-conjugated streptavidin.
Fig. 3. Calcium dependent reactivity of selected MAbs against mouse SP-D. Microtitre wells were coated with antigen (2 μg neck/CTLD SP-D perml) and serial dilutions of biotinylated MAbs were incubated in thepresence of calcium (A) or EDTA (B). Bound antibodies were detected bymeans of HRP-conjugated streptavidin. The results of both MAb 403(NHS-biotin) and MAb 403⁎ (hydrazine biotin) are shown.
80 S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
454.MAb432 could not be used in this assay, probably dueto low reactivity with coated antigen (see Section 4). Todetermine if calcium had to be included in the buffers usedin the ELISA, we analyzed the calcium dependency of themonoclonal antibodies (Fig. 3A and B). We found thatMAb 403, 404, 442, 452, and 454 required calcium to bindantigen with optimal reactivity. It was further found thatMAb 403 required conjugation with hydrazine biotin toretain binding activity.
3.3. Pair-wise optimization of antibody combinations
During the characterization of antibodies, we observedthat they bound differently to various forms of antigen, i.e.neck/CTLD vs. full-length SP-D. Thus, in the followingselection of antibody combinations for sandwich ELISA,we tested dilutions of lung lavage, culture supernatant ofcells expressing CHO full-length SP-D and dilutions of
purified neck/CTLD.We found that the optimal combina-tion for detecting native SP-D and full-length recombinantSP-D was coating with MAb 432 and detection withbiotinylated MAb 403. This combination had both thehighest read-out signal and signal-to-noise ratio. Othercombinations using coatedMAb 401, 404 or 412 stronglydetected the neck/CTLD SP-D, but only weakly nativeSP-D (not shown).
3.4. Optimization of assay conditions
Optimization was performed as described in Section 2and involved various reagents and conditions of theELISA. Although the washing buffer with additional1.0 M NaCl was not the optimal condition for incubatingsamples, it was chosen to facilitate dilution of plasmasamples in the presence of calcium, as the high NaClconcentration inhibits the coagulation of plasma. Calciumcould not be omitted as it is required by MAb 403 foroptimal reactivity.
81S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
3.5. Calibration and determination of working range
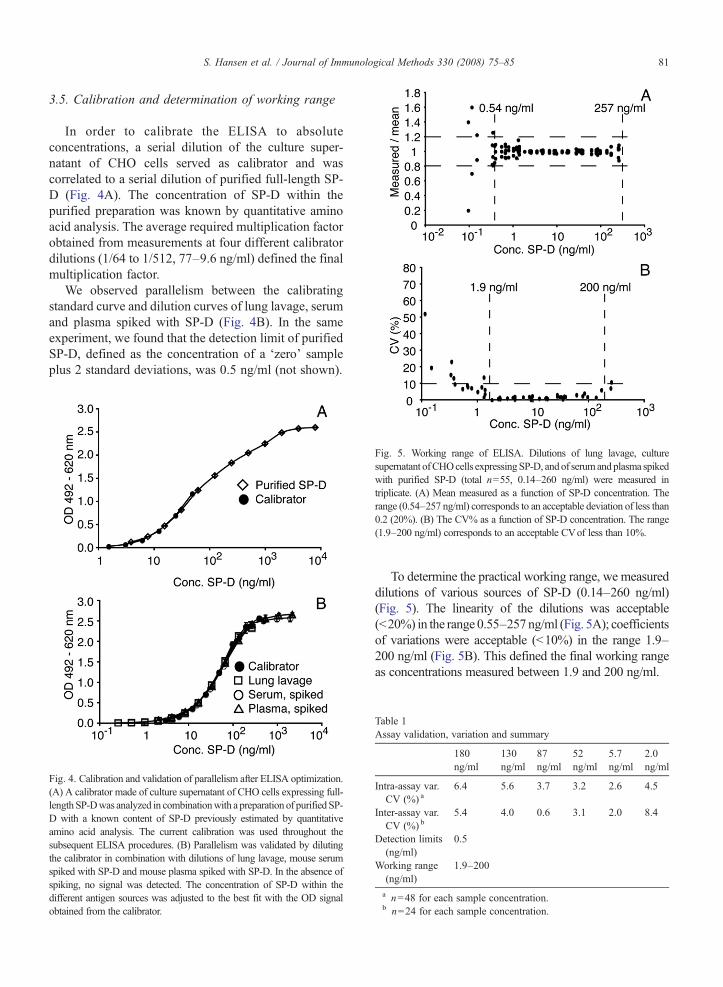
In order to calibrate the ELISA to absoluteconcentrations, a serial dilution of the culture super-natant of CHO cells served as calibrator and wascorrelated to a serial dilution of purified full-length SP-D (Fig. 4A). The concentration of SP-D within thepurified preparation was known by quantitative aminoacid analysis. The average required multiplication factorobtained from measurements at four different calibratordilutions (1/64 to 1/512, 77–9.6 ng/ml) defined the finalmultiplication factor.
We observed parallelism between the calibratingstandard curve and dilution curves of lung lavage, serumand plasma spiked with SP-D (Fig. 4B). In the sameexperiment, we found that the detection limit of purifiedSP-D, defined as the concentration of a ‘zero’ sampleplus 2 standard deviations, was 0.5 ng/ml (not shown).
Fig. 4. Calibration and validation of parallelism after ELISA optimization.(A) A calibrator made of culture supernatant of CHO cells expressing full-lengthSP-Dwas analyzed in combinationwith a preparation of purifiedSP-D with a known content of SP-D previously estimated by quantitativeamino acid analysis. The current calibration was used throughout thesubsequent ELISA procedures. (B) Parallelism was validated by dilutingthe calibrator in combination with dilutions of lung lavage, mouse serumspiked with SP-D and mouse plasma spiked with SP-D. In the absence ofspiking, no signal was detected. The concentration of SP-D within thedifferent antigen sources was adjusted to the best fit with the OD signalobtained from the calibrator.
Fig. 5. Working range of ELISA. Dilutions of lung lavage, culturesupernatant ofCHOcells expressingSP-D, andof serumandplasma spikedwith purified SP-D (total n=55, 0.14–260 ng/ml) were measured intriplicate. (A) Mean measured as a function of SP-D concentration. Therange (0.54–257 ng/ml) corresponds to an acceptable deviation of less than0.2 (20%). (B) The CV% as a function of SP-D concentration. The range(1.9–200 ng/ml) corresponds to an acceptable CVof less than 10%.
To determine the practical working range, we measureddilutions of various sources of SP-D (0.14–260 ng/ml)(Fig. 5). The linearity of the dilutions was acceptable(b20%) in the range 0.55–257ng/ml (Fig. 5A); coefficientsof variations were acceptable (b10%) in the range 1.9–200 ng/ml (Fig. 5B). This defined the final working rangeas concentrations measured between 1.9 and 200 ng/ml.
Table 1Assay validation, variation and summary
180ng/ml
130ng/ml
87ng/ml
52ng/ml
5.7ng/ml
2.0ng/ml
Intra-assay var.CV (%) a
6.4 5.6 3.7 3.2 2.6 4.5
Inter-assay var.CV (%) b
5.4 4.0 0.6 3.1 2.0 8.4
Detection limits(ng/ml)
0.5
Working range(ng/ml)
1.9–200
a n=48 for each sample concentration.b n=24 for each sample concentration.
Table 2Assay validation, recovery
Sample with SP-D (ng/ml) 150 (±2.5) 148 (±4.9) 139 (±7.2) 117 (±1.3) 63.5 (±1.0) 38.2 (±1.3) 11.8 (±0.1) 8.9 (±0.3)Spiking with SP-D (ng/ml) 35.5 (1.5) a 35.5 (1.5) a 35.5 (1.5) a 35.5 (1.5) a 35.5 (1.5) a 35.5 (1.5) a 35.5 (1.5) a 35.5 (1.5) a
Expected (ng/ml) 186 184 175 153 99.0 73.7 47.3 44.4Measured (ng/ml) 182 (±0.1) 186 (±3.9) 177 (±5.3) 146 (±2.4) 105 (±0.4) 78.0 (±1.1) 47.3 (±0.7) 46.1 (±1.9)Recovery (%) 97.8 (±1.0) 101 (±2.2) 101 (±3.1) 95 (±1.1) 106 (±0.1) 106 (±1.3) 106 (±1.5) 104 (±4.1)a SEM (n=25).
82 S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
3.6. Validation of ELISA
The estimated intra- and inter-assay variations (Table 1)were a maximum of 6.4% and 8.4%, respectively. Wevalidated the recovery at varying working concentrationsby spiking different samples of lung lavage with a fixedamount of purified full-length mouse SP-D (Table 2). Werecovered 97–106% of the expected amounts at workingconcentrations spanning 8.9–186 ng/ml.
3.7. Levels of SP-D in murine body fluids and lavages
Using the newly established ELISA, the levels of SP-Dwere accessed in various murine body fluids and lavages.The average of SP-D in lungs lavaged with a total of 3 mlof saline was 435 ng/ml (143–726 ng/ml) (Table 1). Theonly extra pulmonary source of SP-D we found was thevaginal lavage, inwhichwe observed an average of 91 ng/ml (7–180 ng/ml), using a lavage volume of 1 ml (onlyapproximate value). No SP-D (b3.8 ng/ml) was detectedin either serum or plasma. The specificity of the ELISAwas confirmed by analysis of samples from SP-D defi-cient mice (SP-D−/−). No SP-D was detected in the lunglavage obtained from SP-D deficient mice.
3.8. Effect of freezing and thawing
Fifteen samples of lung lavage, serum and plasmaspiked with SP-D were frozen (−20 °C) and thawed (roomtemperature). After a single freeze thawing procedure, werecovered 71% (SD 8.3) of the SP-D in lung lavage, 72%(SD 3.1) in serum and 81% (SD 0.52) in plasma. Anadditional eight rounds of freezing and thawing did notaffect the recovery in serum and plasma, but lowered therecovery in lung lavage to 46% and resulted in a highdegree of individual variation (SD 18) (results not shown).
4. Discussion
In general, SP-D is associated with lung surfactant andmucus at extra pulmonary sites but in humans SP-D is alsopresent in the blood, albeit in lower concentrations than inlung lavage.We have successfully developed monoclonalanti-mouse-SP-D antibodies (MAb 432 and MAb 403)
and established an ELISA for mouse SP-D in variousbody fluids. To our surprise, and in contrast to thesituation in humans, there is no SP-D in the blood ofmice.Besides the obvious presence of SP-D in mouse lunglavage, we also found SP-D in the mouse vaginal lavage.
Two different sources of antigens were used to raiseantibodies, for testing specificity, and for setting up theELISA. The same sources of antigens in combinationwithmouse lung lavage were used for validating the ELISA.The two most suitable antibodies in terms of reactivitywith native SP-D and appliance in ELISAwere selected.Although MAb 432, in comparison with the other mono-clonal antibodies, only binds to coated antigen with arelatively low affinity (Fig. 2A), it was selected as catch-ing antibody. Its reactivity with soluble antigen, in com-bination with MAb 403 as detecting antibody, was thebest of all the antibodies tested (not shown). The detectingantibody, MAb 403, recognized coated antigen (Fig. 2A)but its reactivity depended critically on the presence ofcalcium (Fig. 3). However, the binding site on MAb 403was sensitive to labelling with NHS-biotin, which targetsthe lysine residues of the primary amine groups. Goodreactivity of labelled MAb 403 was achieved by bio-tinylation using hydrazine biotin, which targets oxidizedcarbohydrates on the Fc portion of the antibody (Fig. 3).As the epitope recognized by MAb 432 is sensitive tocoating, and this is not the case for the epitope recognizedby MAb 403, we expected that the two MAbs wouldrecognize different epitopes on SP-D. However, we werenot able to verify this by inhibition ELISA (Fig. 2B) as thereactivity ofMAb 432with coated antigen is probably toolow to inhibit binding of the labelled antibody. Usingunlabelled MAb 432 for inhibition did not inhibit any ofthe tested labelled MAbs, including labelled MAb 432itself (not shown).
As the combined use of MAb 432 and MAb 403resulted in the best sensitivity, this further supported thenotion that MAb 432 and MAb 403 recognize differentepitopes. It was also obvious from the pair-wise opti-mization studies that neither the selected combination ofMAbs nor other combinations recognized neck/CTLD aswell as native and full-length SP-D (not shown). Ingeneral, the absorbance using saturating amounts of neck/CTLD resulted in only 10–50% of the signal obtained

Table 3Measurements of mouse SP-D in body fluids and lavages
WTconcentration(ng/ml)
n a SP-D−/−
concentration(ng/ml)
n
Lung lavage (∼3 ml) 143–726 56 ND 19Serum ND 35 ND 10Plasma ND 25 ND 9Eye lavage (∼40 μl) ND 21 ND 19Peritoneum
lavage (∼2 ml)ND 11
Vagina lavage (∼1 ml) 7.0–180 11
ND, not detectable, b3.8 ng/ml.a Various strains.
83S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
from lung lavage or full-length SP-D. We expect that thecapture of trimeric neck/CTLD, in comparison withdodecameric SP-D (native and full-length), decreases thenumber of captured and available epitopes for the detectingantibodies. The current ELISAwas only validated for usewith native and full-length SP-D.
The practical working range of the final ELISA wasfound to be 1.9–200 ng/ml. Previously, an ELISA basedon both catching and detection with the same rabbit anti-mouse-SP-D polyclonal antibody reported sensitivitylevels of 4–20 ng/ml (Zhang et al., 2003). No practicalworking range was estimated for this ELISA. In com-parison with the performance of an ELISA formeasuring human SP-D (16–520 ng/ml), the workingrange of the anti-mouse-SP-D ELISA developed in ourstudy is comparable and acceptable (Leth-Larsen et al.,2003). We deliberately calibrated the ELISA with puri-fied SP-D (recombinant) that had been analyzed byquantitative amino acid analysis (Fig. 4A) and foundthat this resulted in 2–3-fold lower concentrations thanwhat we initially expected from the quantification of thesame preparation using the absorbance at 280 nm (notshown). This might explain some of the differences seenin the estimations of concentrations and also the dif-ferences in the working ranges of the different ELISAs.We expect that non-protein contaminants such as endo-toxins and detergents may interfere with the estimationbased on optical absorbance.
We also observed parallelism between the supernatantof CHO cells expressing full-length SP-D and lung lavage(Fig. 4B). Similar parallelisms were observed withdilutions of spiked serum and plasma (Fig. 4B), anddemonstrate that the current ELISA setup can be used tomeasure SP-D concentrations and regulation in the blood.The inter- and intra-assay variations (b8.4%, Table 1)were acceptable (b10%) and the deviation of recoveriesupon spikingwith SP-D (Table 2)were also acceptable anddiverged only up to 6% from the expected concentrations.During the initial calibration procedure, we observed thatthe assay was influenced by repeated freezing and thawingand prolonged storage at 4 °C of the calibrator. After arecalibration of the ELISA applying new calibrators andnew standard preparations of purified SP-D (quantified byamino acid analysis) that had only been frozen once andused immediately upon thawing, we performed a numberof additional freeze thawing experiments. We found thatafter a single freeze thawing procedure samples loseapproximately 25% of the SP-D, but the loss appears to benearly uniform for all samples (SD 0.5–8.3). Furtherrounds of freezing and thawing affected themeasurementsof SP-D in lung lavage in particular. Thus, we concludethat storage of both samples and calibrators at at least
−20 °C is essential and that repeated freezing and thawingshould be avoided. We expect that enzymatic degradationis responsible for the loss.
The specificity of the final ELISA was best demon-strated by the negative measurements of SP-D in lunglavage, serum and plasma from SP-D deficient mice(Table 3). When the samples were diluted with an equalvolume of buffer, the content of measured SP-D waslower than the minimal working limit of the assay(b1.9); and upon back calculation, this resulted in lessthan 3.8 ng/ml. To further test the assay we measuredSP-D in body fluids from wild type mice (Table 3). Thisconfirmed what we observed during the test for paral-lelism. SP-D is absent in mouse serum and plasma(b3.8 ng/ml). Our observations confirm the previouswork based on an ELISA using polyclonal-anti-mouse-SP-D (Zhang et al., 2003; Fujita et al., 2005). In thesestudies, negligible, low serum concentrations of SP-Dranging from 4.3 to 8.3 ng/ml in various strains of micewere found. In addition, serum concentrations werefound to be up regulated by the presence of lung tu-mours, airway inflammation due to instilled bleomycinor aerozolized OVA in sensitized mice.
In the lung lavage from mice, we found on average435 ng/ml (±292 ng/ml, n=56) when the lungs wereeither lavaged using 3 ml of PBS or the measuredconcentration was adjusted to this lavage volume. Themeasured samples were derived from various cohortsand strains of mice at varying ages, and some of thesamples had been frozen and thawed up to three times.This might explain the relatively large variation.
In humans, RNA and immunohistochemical studieshave shown that SP-D is synthesized by epithelial cellsoften associated with mucosal surfaces (Madsen et al.,2000; Stahlman et al., 2002). Similar studies have shownthat SP-D is also associated with human endothelial cells(Sorensen et al., 2006). In contrast, the expression of SP-D in mice appears to be restricted to the airways, the

84 S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
lacrimal gland, the uterus and the ovary (Akiyama et al.,2002). We were able to measure SP-D in lavage from thevagina, and thereby support the observations that SP-D issynthesized in the female genital tract. This supports thenotion that SP-Dmight play a role in the immune defenseand regulation of the female genital tract (Akiyama et al.,2002; Leth-Larsen et al., 2004). We were not able tomeasure SP-D in lavage from mouse eye, but as we didnot directly examine its presence in tear fluid, the pres-ence and importance of SP-D synthesized by the lacrimalgland is not entirely excluded (Akiyama et al., 2002; Niet al., 2005).
In summary,we have described an immunological assayfor measuring the concentration of mouse SP-D in differentbody fluids. This assay specificallymeasuresmurine SP-D.This method produces low backgrounds, has sensitive de-tection over a physiological range of concentrations, and issuitable for screeningmultiple samples in research settings.The method represents a marked improvement over pub-lished assays for the detection of mouse SP-D due tostandardized reagents, an absolute calibration and valida-tion for measuring SP-D in various body fluids.
Acknowledgements
We thank Professor Jo Rae Wright (Department of CellBiology, Duke University Medical Center, Durham, NC,USA) for theCHOcells expressing full-lengthmouse SP-Dand Professor Sam Hawgood (Department of Pediatrics,Cardiovascular Research Institute, University of California,San Francisco, CA, USA) for donating the SP-D deficientmice. This work was supported by the Danish MedicalResearch Council, The Carlsberg Foundation and theLundbeck Foundation.
References
Aderibigbe, A.O., Thomas, R.F., Mercer, R.R., Auten, R.L., 1999. Briefexposure to 95% oxygen alters surfactant protein D and mRNA inadult rat alveolar and bronchiolar epithelium. Am. J. Respir. CellMol. Biol. 20, 219.
Akerstrom, B., Bjorck, L., 1986.A physicochemical study of proteinG, amolecule with unique immunoglobulin G-binding properties. J. Biol.Chem. 261, 10240.
Akiyama, J., Hoffman, A., Brown, C., Allen, L., Edmondson, J., Poulain,F.,Hawgood, S., 2002. Tissue distribution of surfactant proteins a andd in the mouse. J. Histochem. Cytochem. 50, 993.
Botas, C., Poulain, F., Akiyama, J., Brown, C., Allen, L., Goerke, J.,Clements, J., Carlson, E., Gillespie, A.M., Epstein, C., Hawgood, S.,1998. Altered surfactant homeostasis and alveolar type II cellmorphology in mice lacking surfactant protein D. Proc. Natl. Acad.Sci. U. S. A. 95, 11869.
Crowther, J.R., 2000. The ELISA guidebook. Methods Mol. Biol. 149(III–IV), 1.
Dulkerian, S.J., Gonzales, L.W., Ning, Y., Ballard, P.L., 1996. Regulationof surfactant protein D in human fetal lung. Am. J. Respir. Cell Mol.Biol. 15, 781.
Erpenbeck, V.J., Malherbe, D.C., Sommer, S., Schmiedl, A., Steinhilber,W., Ghio, A.J., Krug, N.,Wright, J.R., Hohlfeld, J.M., 2005. Surfactantprotein D increases phagocytosis and aggregation of pollen-allergenstarch granules. Am. J. Physiol., Lung Cell. Mol. Physiol. 288, L692.
Floros, J., Lin, H.M., Garcia, A., Salazar, M.A., Guo, X., DiAngelo, S.,Montano,M., Luo, J., Pardo, A., Selman,M., 2000. Surfactant proteingeneticmarker alleles identify a subgroup of tuberculosis in aMexicanpopulation. J. Infect. Dis. 182, 1473.
Fujita, M., Shannon, J.M., Ouchi, H., Voelker, D.R., Nakanishi, Y., Mason,R.J., 2005. Serum surfactant proteinD is increased in acute and chronicinflammation in mice. Cytokine 31, 25.
Hansen, S., Holmskov, U., 1998. Structural aspects of collectins andreceptors for collectins. Immunobiology 199, 165.
Hansen, S., Holmskov, U., 2002. Lung surfactant protein D (SP-D) andthemolecular diverted descendants: conglutinin, CL-43 andCL-46.Immunobiology 205, 498.
Hansen, S., Thiel, S., Willis, A., Holmskov, U., Jensenius, J.C., 2000.Purification and characterization of twomannan-binding lectins frommouse serum. J. Immunol. 164, 2610.
Hansen, S., Lo, B., Evans, K., Neophytou, P., Holmskov, U., Wright, J.R.,2007. Surfactant protein D augments bacterial association butattenuates major histocompatibility complex class II presentation ofbacterial antigens. Am. J. Respir. Cell Mol. Biol. 36, 94.
Hartl, D., Griese, M., 2006. Surfactant protein D in human lung diseases.Eur. J. Clin. Investig. 36, 423.
Herias, M.V., Hogenkamp, A., van Asten, A.J., Tersteeg, M.H., vanEijk, M., Haagsman, H.P., 2007. Expression sites of the collectinSP-D suggest its importance in first line host defence: power ofcombining in situ hybridisation, RT-PCR and immunohistochem-istry. Mol. Immunol. 44, 3324.
Holmskov, U., Thiel, S., Jensenius, J.C., 2003. Collections and ficolins:humoral lectins of the innate immune defense. Annu. Rev. Immunol.21, 547.
Homer, R.J., Zheng, T., Chupp,G., He, S., Zhu, Z., Chen,Q.,Ma, B., Hite,R.D., Gobran, L.I., Rooney, S.A., Elias, J.A., 2002. Pulmonary type IIcell hypertrophy and pulmonary lipoproteinosis are features of chronicIL-13 exposure. Am. J. Physiol., Lung Cell. Mol. Physiol. 283, L52.
Ikegami, M., 2006. Surfactant catabolism. Respirology 11, S24 (Suppl).Ikegami, M., Whitsett, J.A., Chroneos, Z.C., Ross, G.F., Reed, J.A.,
Bachurski, C.J., Jobe, A.H., 2000. IL-4 increases surfactant andregulates metabolism in vivo. Am. J. Physiol., Lung Cell. Mol.Physiol. 278, L75.
Kohler, G., Milstein, C., 1975. Continuous cultures of fused cellssecreting antibody of predefined specificity. Nature 256, 495.
Laemmli, U.K., 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680.
Lahti, M., Lofgren, J., Marttila, R., Renko, M., Klaavuniemi, T., Haataja,R., Ramet, M., Hallman, M., 2002. Surfactant protein D genepolymorphism associated with severe respiratory syncytial virusinfection. Pediatr. Res. 51, 696.
Leth-Larsen, R., Nordenbaek, C., Tornoe, I., Moeller, V., Schlosser, A.,Koch, C., Teisner, B., Junker, P., Holmskov, U., 2003. Surfactantprotein D (SP-D) serum levels in patients with community-acquiredpneumonia small star, filled. Clin. Immunol. 108, 29.
Leth-Larsen, R., Floridon, C., Nielsen,O.,Holmskov,U., 2004. Surfactantprotein D in the female genital tract. Mol. Hum. Reprod. 10, 149.
Leth-Larsen, R., Garred, P., Jensenius, H., Meschi, J., Hartshorn, K.,Madsen, J., Tornoe, I., Madsen, H.O., Sorensen, G., Crouch, E.,Holmskov, U., 2005. A common polymorphism in the SFTPD gene

85S. Hansen et al. / Journal of Immunological Methods 330 (2008) 75–85
influences assembly, function, and concentration of surfactant proteinD. J. Immunol. 174, 1532.
Madsen, J., Kliem, A., Tornoe, I., Skjodt, K., Koch, C., Holmskov, U.,2000. Localization of lung surfactant protein D onmucosal surfacesin human tissues. J. Immunol. 164, 5866.
Nesterenko,M.V., Tilley,M., Upton, S.J., 1994.A simplemodification ofBlum's silver stain method allows for 30minute detection of proteinsin polyacrylamide gels. J. Biochem. Biophys. Methods 28, 239.
Ni, M., Evans, D.J., Hawgood, S., Anders, E.M., Sack, R.A., Fleiszig,S.M., 2005. Surfactant protein D is present in human tear fluid andthe cornea and inhibits epithelial cell invasion by Pseudomonasaeruginosa. Infect. Immun. 73, 2147.
O'Shannessy, D.J., Dobersen, M.J., Quarles, R.H., 1984. A novelprocedure for labeling immunoglobulins by conjugation to oligosac-charide moieties. Immunol. Lett. 8, 273.
Rust, K., Bingle, L., Mariencheck, W., Persson, A., Crouch, E.C., 1996.Characterization of the human surfactant protein D promoter:transcriptional regulation of SP-D gene expression by glucocorticoids.Am. J. Respir. Cell Mol. Biol. 14, 121.
Sorensen, G.L., Madsen, J., Kejling, K., Tornoe, I., Nielsen, O., Townsend,P., Poulain, F., Nielsen, C.H., Reid, K.B., Hawgood, S., Falk, E.,
Holmskov, U., 2006. Surfactant protein D is proatherogenic in mice.Am. J. Physiol., Heart Circ. Physiol. 290, H2286.
Stahlman, M.T., Gray, M.E., Hull, W.M., Whitsett, J.A., 2002.Immunolocalization of surfactant protein-D (SP-D) in human fetal,newborn, and adult tissues. J. Histochem. Cytochem. 50, 651.
Strong, P., Kishore, U., Morgan, C., Lopez Bernal, A., Singh, M., Reid,K.B., 1998. A novel method of purifying lung surfactant proteins AandD from the lung lavage of alveolar proteinosis patients and frompooled amniotic fluid. J. Immunol. Methods 220, 139.
Stuart, L.M., Henson, P.M., Vandivier, R.W., 2006.Collectins: opsonins forapoptotic cells and regulators of inflammation. Curr. Dir. Autoimmun.9, 143.
Wright, J.R., 2005. Immunoregulatory functions of surfactant proteins.Nat. Rev., Immunol. 5, 58.
Zhang, F., Pao, W., Umphress, S.M., Jakowlew, S.B., Meyer, A.M.,Dwyer-Nield, L.D., Nielsen, L.D., Takeda,K., Gelfand, E.W., Fisher,J.H., Zhang, L., Malkinson, A.M., Mason, R.J., 2003. Serum levelsof surfactant protein D are increased in mice with lung tumors.Cancer Res. 63, 5889.





























