Embed Size (px)
DESCRIPTION
comparison of the performance
Citation preview

Appl Microbiol Biotechnol (1994) 41:62-72 App//ed
Microbiology Biotechnology © Springer-Verlag 1994
An interlaboratory comparison of the performance of ethanol-producing micro-organisms in a xylose-rich acid hydrolysate B. Hahn-H~igerdal 1, H. Jeppsson 1, L. Olsson 1, A. Mohagheghi 2
1 Department of Applied Microbiology, Lund Institute of Technology/University of Lund, P.O. Box 124, S-22100 Lund, Sweden 2 Bioprocess and Fuels Engineering Research Branch, National Renewable Energy Laboratory, 1617 Cole Boulevard, Golden, CO 80401, USA
Received: 30 June 1993/Received revision: 29 September 1993/Accepted: 30 September 1993
Abstract. A xylose-rich, dilute-acid-pretreated corn- cob hydrolysate was fermented by Escherichia coli ATCC 11303, recombinant (rec) E. coli B (pLOI 297 and KOl l ) , Pichia stipitis (CBS 5773, 6054 and R), Saccharomyces cerev&iae isolate 3 in combination with xylose isomerase, rec S. cerev&&e (T J1, H550 and H477) and Fusarium oxysporum VTT-D-80134 in an interlaboratory comparison. The micro-organisms were studied according to three different options: (A) fermentation under consistent conditions, (B) fermen- tation under optimal conditions for the organism, and (C) fermentation under optimal conditions for the or- ganism with detoxification of the hydrolysate. The highest yields of ethanol, 0.24 g/g (A), 0.36 g/g (B) and 0.54 g/g (C), were obtained from rec E. coli B, KOl l . P, stipit& and F. oxysporum were sensitive to the inhi- bitors present in the hydrolysate and produced a maxi- mum yield of 0.34 g/g (C) and 0.04 g/g (B), respective- ly. The analysis of the corn-cob hydrolysate and as- pects of process economy of the different fermentation options (pH, sterilization, nutrient supplementation, adaptation, detoxification) are discussed.
Introduction
As a renewable energy resource, fuel ethanol has re- ceived considerable scientific interest since the oil cri- sis in the mid 1970s. Fuel ethanol can be produced from different sources, e.g. fast-growing wood, agricul- tural or forestry plant residues, and appropriate muni- cipal waste (Kosaric et al. 1983; Magee and Kosaric 1985; Parisi 1989; Hahn-H/igerdal et al. 1991; Olsson and Hahn-Hfigerdal 1993). The raw material, lignocel- lulose, has to be degraded to a fermentable hydroly- sate from which ethanol can be produced. Fuel ethanol is a bulk product with a world market price of 0.35-
Correspondence to: B. Hahn-H~igerdal
0.45 US dollars (May 1993), which requires an efficient fermentation process to be economically feasible. Fur- thermore, environmental benefits and political deci- sions are of great importance for the extent of fuel ethanol appliCations (Keim and Venkatasubramanian 1989; Lynd et al. 1991). Xylitol, which is of higher eco- nomic value than ethanol, can also be produced by fer- mentation of lignocellulose hydrolysates. It is widely used in the food industry as a sweetener in the Nordic countries. Xylitol is anticariogenic and suitable as a su- gar substitute for diabetics (M/~kinen 1979; Hyv6nen et al. 1982). A fermentation process for xylitol production could be advantageous compared to the chemical proc- ess used today (Melaja and H/~m/~l~iinen 1977) because the downstream processing is expected to be cheaper (Ojamo et al. 1989).
Depending on the raw material and pretreatment, the lignocellulose hydrolysates contain many different and variable amounts of monosaccharides (e.g. glu- cose, galactose, mannose, xylose and arabinose), disac- charides (e.g. cellobiose) and polysaccharides (e.g. xy- lans and mannans) (Magee and Kosaric 1985). Special attention has been given to the fermentation of xylose as it is the most difficult to ferment (Jeffries 1983; Schneider 1989; Hahn-H/~gerdal et al. 1993). The pre- treatment and hydrolysis of lignocellulose result in the formation of a number of degradation products and ex- tracts. These compounds, e.g. furfural, 5-hydroxyme- thylfurfural, levulinic acid, vanillin, syringaldehyde and acetic acid are known to inhibit micro-organisms (Clark and Mackie 1984; Ando et al. 1986; Tran and Chambers 1986; Nishikawa et al. 1988; Sanchez and Bautista 1988; van Zyl et al. 1991). Moreover, the products of the fermentation, ethanol and organic acids, are also inhibitory to micro-organisms (Maiorel- la et al. 1983; Jones 1989). Attempts have been made to increase the fermentability of the hydrolysates (i.e. to detoxify the hydrolysate) using different pretreat- merits, e.g. ion-exchange (Clark and Mackie 1984; Fein et al. 1984; Frazer and McCaskey 1989), calcium hy- droxide treatment (Strickland and Beck 1985; van Zyl et al. 1988), molecular sieves (Tran and Chambers

63
1986), s t eam str ipping (Yu et al. 1987) and ion exclu- sion (Bucher t et al. 1990).
A n economic fuel e thanol p roduc t i on process re- quires a mic ro -organ i sm with the fol lowing qualities: efficient uti l ization of the raw mater ia l (high yield), high final e thanol concen t ra t ion ( m i n i m u m distillation costs), and fast e thanol f e rmen ta t ion ra te (high pro- ductivity). In addit ion, these demands should be ful- filled wi thout the need for extra chemicals and process steps, such as complex nutr ient supplements , detoxifi- cat ion chemicals, detoxif icat ion p rocedures and sterili- zation. Genera l ly , a high yield and a high e thanol con- cent ra t ion are cons idered to have the greates t impact on process economy , bu t because the e thanol p roduc- t ion process is so complex, the process needs detai led invest igat ion to achieve a meaningfu l economic evalua- t ion (Wright 1988; Zacch i et al. 1988; H i n m a n et al. 1989).
The presen t s tudy was u n d e r t a k e n as a col labora- tive pro jec t be tween different g roups involved with fuel e thanol research (Table 1), in o rder to c o m p a r e the results ob ta ined f r o m fe rment ing al ternat ive prepa- rat ions of one par t icular l ignocellulose hydro lysa te substrate with Escherichia coli, r e c om bi na n t (rec) E. coli, Saccharomyces cerevisiae + xylose i somerase (XI) , rec S. cerevisiae, Pichia stipitis and Fusarium ox- ysporum. For this s tudy a xylose-r ich hydro lysa te de- r ived f rom corn cobs after dilute acid p r e t r ea tmen t was chosen since the f e rmen ta t ion of the xylose f ract ion p roduces mos t problems. T he hydro lysa te was distri- bu ted to the part ic ipants at p H 1.5 to assure tha t no metabol ic activity occur red dur ing t ranspor ta t ion . The invest igat ion was divided into three exper imenta l cate- gories. First, the f e rmen ta t ion p e r f o r m a n c e of the in- vest igated micro-organisms was c o m p a r e d unde r con- sistent condi t ions concern ing pH, oxygenat ion , tem- pe ra tu re and nutrients. This was cons idered as the ba- sic exper imen t because it a l lowed the compar i son of a b r o a d range of organisms unde r identical condit ions. Second, the organisms were c o m p a r e d unde r wha t was cons idered to be the opt imal condi t ions for each indi-
Table 1. Participants in the interlaboratory collaboration
Name Affiliation
S. Amartey/T. W. Jeffries
D. Beall/L. O. Ingram
B. L. Boynton/J. D. McMillan
B. Hahn-Hggerdal/H. Jeppsson/T. Linddn/N. Meinander/L. Olsson
H. G. Lawford/J. Rousseau
J. Pourquie
T. Seki/T. Yoshida M.-L. Suihko
Forest Products Laborato- ry, Madison, USA
University of Florida, USA
National Renewable En- ergy Laboratory, Gold- en, USA
Lund Institute of Tech- nologyAJniversity of Luud, Sweden
University of Toronto, Canada
Ministdre de l'Agricul- ture, INA, Paris, France
Osaka University, Japan VTT, Espoo, Finland
vidual o rgan ism with respect to oxygena t ion and nu- tr ient supplementa t ion . Third, the organisms were c o m p a r e d in a subst ra te where detoxif icat ion was used to reduce the con ten t o f any inhibitors and in some cases in combina t ion with adap ta t ion of the micro-or- ganism to the hydrolysate . The basic exper imen t repre- sents the f e rmen ta t ion al ternat ive requir ing a mini- m u m of addi t ional costs whereas the two o the r gener- ally represen t m o r e cost ly f e rmen ta t ion alternatives. In addit ion, the accuracy of the analysis o f the corn-cob hydro lysa te was evaluated.
Materials and methods
Hydrolysate and pretreatments. The hydrolysate was prepared by dilute acid pretreatment of corn cobs, obtained locally at the Na- tional Renewable Energy Laboratory in Golden, Colo., USA. The dry corn material was knife-milled to pass through a 2-mm rejection screen. The corn-cob particles were pretreated with di- lute (0.83%, w/w) sulphuric acid solutions (yielding a hydrolysis pH of ].35-1.4) in a 2-gallon stainless-steel Parr reactor (Parr, Moline, Ill., USA). A low-solid slurry (10%) of biomass in deion- ized water was heated to 160 ° C, the acid was then injected and, after being heated at 160°C for 10 min, the pretreated material was cooled to 90 ° C. The mixture was then immediately filtered through a Buchner funnel to recover the hydrolysate at a final pH of approximately 1.5.
Experimental design. Experiment category A - the basic experi- ment: with minor deviations the following procedure was used. The hydrolysate was supplemented with 2.5 g/1 of yeast extract, 0.25 g/1 of (NH4)zHPO4, 0.025 g/1 of MgSO4"7HzO and 0.1 M so- dium phosphate. The pH was adjusted to 5.5 for the yeasts and 7.0 for the bacteria. The yeast fermentations were performed un- der Oz-limited conditions, in half-filled (50/100, v/v) flasks at 30°C and 150 rpm. The bacterial fermentations were performed under anoxic conditions at 30 ° C. The fermentations were inocu- lated to give a final cell mass concentration of approximately 5 g/1 (dry weight). Samples were taken regularly and the pH was ad- justed each time a sample was taken.
Experiment categories B and C were performed under opti- mal conditions for each type of micro-organism without and with detoxification, respectively. The conditions are reported in the Results.
Micro-organisms and cell mass production. Each type of micro- organism was grown under the conditions given below and har- vested in the log phase by centrifugation. The cells were washed twice with physiological sodium chloride solution (9 g/l), if not otherwise stated, and were then used as the inoculum for the fer- mentation of the corn-cob hydrolysate under the conditions re- ferred to as categories A, B and C.
E. coli ATCC 11303 was grown in a medium containing 10 g/1 of bactotryptone, 5 g/1 of yeast extract, 5 g/1 of sodium chloride, 10 g/1 of glucose, 10 g/1 of xylose and 10 g/1 of galactose, with the pH adjusted to 7.5. The medium was inoculated with a 2.0% (v/v) inoculum of an overnight culture. One-litre baffled erlenmeyer flasks containing 200-ml aliquots of this culture were incubated in a rotary shaking water bath at 37 ° C.
E. coli B (ATCC 11303) carries the plasmid pLOI297, con- taining the genes pdc and adhB coding for the enzymes pyruvate decarboxylase and alcohol dehydrogenase II from Zymomonas mobilis (Alterthum and Ingrain 1989). Cukures were stored ( - 10 ° C) in LB (Luria broth; 10 g/1 of tryptone, 5 g/1 of yeast ex- tract, 5 g/1 of sodium chloride)/glycerol-citrate and were plated on selective media (LB + 20 g/1 of agar + 10 mg/1 of tetracycline and 40 rag/1 of ampicillin). Cell mass was produced overnight in buff
64
fered LB containing antibiotics (ampicillin and tetracycline) and 30 g/1 of xylose as the carbon source.
E. coli KO 11 carries the genes pdc and adhB from Z. mobilis integrated into the chromosome. In addition, other genetic changes were carried out to minimize by-product formation (Ohta et al. 1991). The inoculum was prepared by transferring cells from a single colony to 125-ml flasks containing 50 ml fer- mentation broth (LB + 50 g/1 of xylose). Cultures were incu- bated overnight at 30°C without agitation.
F. oxysporum VTT-D-80134 was grown in a medium consist- ing of 50 g/1 of xylose, 1.0 g/1 of yeast extract, 3.4 g/1 of NaNO3, 2.0 g/1 of KH2PO4, 0.4 g/1 of CaC12"2H20, 0.3 g/1 of MgSO4"7H20 and 5.0 ml trace metal solution. The composition of the trace metal solution was 2.0 g/1 of H3BO3, 0.2 g/1 of COSO4"7H20 , 0.8 g/1 of CuSO4"5H20, 2.0 g/1 of ZnSO4"7H20, 0.6 g/1 of MnSO4-H20, 0.2 g/1 of KI, 0.4 g/1 of FeSO4.7H20 and 0.6 g/1 of A12(SO4)3. The mycelial mass was washed once with wa- ter before inoculation. The pH was 5.0 and temperature 30 ° C.
P. stipitis CBS 5773 (NRRL Y-7124) (a) was grown aerobical- ly in four 1-1 DeLong culture flasks containing 250 ml buffered yeast nitrogen base (YNB, Difco) supplemented with 2 g/1 of glu- cose, 20 g/1 of xylose, 20 g/1 of arabinose and 20 g/1 of galactose as carbon sources. The YNB was buffered at pH 5.5 using KH2PO4 and Na2HPO4 at concentrations of 10 g/1 and 0.5 g/l, respective- ly.
P. stipitis CBS 5773 (/3) was obtained from a continuous cul- ture grown aerobically at pH 4.5 on purified xylose (12 g/l) as the sole carbon source. The medium composition was not defined further.
P. stipitis CBS 5773 (9,) was produced at 30 ° C, pH 6.0, in 500- ml Sakaguchi flasks containing 50 ml medium containing 20 g/1 of xylose, 20 g/l of polypeptone and 10 g/1 of yeast extract. The cells were washed once with 10 g/1 of sodium chloride solution prior to inoculation.
P. stipitis CBS 6054 (6) was grown in a medium containing 10 g/1 of xylose, 10 g/1 of glucose, 10 g/1 of galactose, 3 g/1 of yeast extract, 3 g/l of malt extract, 5 g/1 of bactopeptone, 19 g/1 of KH2PO4, 3 g/1 of (NH4)2HPO4 and 1.1 g/1 of MgSO4'7H20. The growth medium was inoculated directly from agar slants. One- litre baffled erlenmeyer flasks containing 200-ml aliquots of this culture were incubated in a rotary shaking water bath at 30 ° C.
P. stipitis CBS 6054 (e) was grown in a medium containing 1.7 g/1 of yeast nitrogen base (without amino acids and ammon- ium sulphate), 1.0 g/1 of casamino acids, 2.2 g/1 of urea, 40 g/1 of glucose, 50 g/1 of xylose and 10 g/1 of arabinose. The pH was ad- justed to 5.0. Cell mass was produced at 30°C and 150 rpm in a 125-ml erlenmeyer flask filled to 50 ml.
P. stipitis R, a strain that has been adapted to lignocellulose hydrolysates (Parekh et al. 1986), was grown as for P. stipitis CBS 6054 (6).
S. cerevisiae Isolate 3, a strain isolated from a spent sulphite liquor fermentation plant (Lind6n et al. 1992), was grown as for P. stipitis CBS 6054 (3).
S. cerevisiae H477 is a strain carrying the gene for the enzyme xylose reductase from P. stipitis CBS 6054. S. cerevisiae H477 was constructed as described by Hallborn et al. (1991), although tt/e parent strain was S. cerevisiae H308 (his3-dell, leu2-3, leu2-112, trpl-289, ura3-52, cir +, Gal +) (Hallborn, personal communica- tion). It was grown in a medium containing 6.7 g/1 of yeast nitrog- en base without amino acids, 20 g/1 of glucose, 0.0135 g/1 of ade- nine, 0.348 g/1 of arginine, 0.266 g/1 of aspartic acid, 0.058 g/1 of histidine, 0.036 g/1 of inositol, 0.525 g/1 of isoleucine, 0.091 g/1 of lysine, 0.149 g/1 of methionine, 0.083 g/1 of phenylalanine, 0.105 g/1 of serine, 0.119 g/1 of threonine, 0.082 g/1 of tryptophan, 0.018 g/1 of tyrosine, 0.022 g/1 of uracil and 0.117 g/1 of valine. The cells were grown in two stages, first in a 50-ml volume and then in a 500-ml volume at 30 ° C and 200 rpm.
S. cerevisiae H550 is S. cerevisiae H308 transformed with the genes for the enzymes xylose reductase (XR) and xylitol dehy- drogenase (XDH) from P. stipitis CBS 6054 on the plasmids pUA103 and pYEP-XYL2 (Hallborn et al. 1992; Hallborn, per-
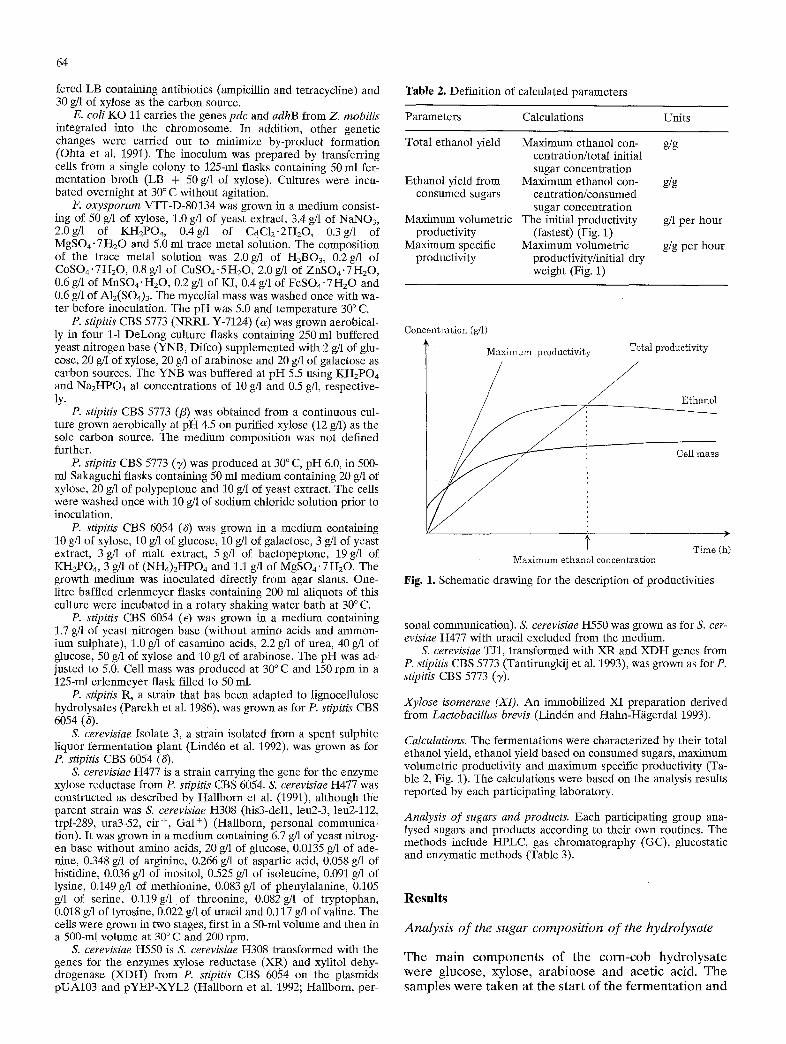
Table 2. Definition of calculated parameters
Parameters Calculations Units
Total ethanol yield Maximum ethanol con- g/g centration/total initial sugar concentration
Ethanol yield from Maximum ethanol con- g/g consumed sugars centration/consumed
sugar concentration Maximum volumetric The initial productivity g/1 per hour
productivity (fastest) (Fig. 1) Maximum specific Maximum volumetric g/g per hour
productivity productivity/initial dry weight (Fig. 1)
Concentration (g/t)
Maximum productivity Total productivity
Cell mass
Time (h) Maximum ethanol concentration
Fig. 1. Schematic drawing for the description of productivities
sonal communication). S. cerevisiae H550 was grown as for S. cer- evisiae H477 with uracil excluded from the medium.
S. cerevisiae TJ1, transformed with XR and XDH genes from P. stipitis CBS 5773 (Tantirungkij et al. 1993), was grown as for P. stipitis CBS 5773 (';/).
Xylose isomerase (XI). An immobilized XI preparation derived from Lactobacillus brevis (Lind6n and Hahn-H~igerdal 1993).
Calculations. The fermentations were characterized by their total ethanol yield, ethanol yield based on consumed sugars, maximum volumetric productivity and maximum specific productivity (Ta- ble 2, Fig. 1). The calculations were based on the analysis results reported by each participating laboratory.
Analysis of sugars and products. Each participating group ana- lysed sugars and products according to their own routines. The methods include HPLC, gas chromatography (GC), glucostatic and enzymatic methods (Table 3).
Results
A n a l y s i s o f the sugar compos i t ion o f the hydro lysa te
T h e m a i n c o m p o n e n t s o f t h e c o r n - c o b h y d r o l y s a t e w e r e g lucose , xy lose , a r a b i n o s e a n d ace t ic acid. T h e s a m p l e s w e r e t a k e n at t he s t a r t o f t he f e r m e n t a t i o n a n d
65
©
© ?
O
©
© ,
©
<
©
e~
O
©
O
©
r~
<
I I I I I l [ l l q II 0 0 0
r
~ d ~ S d d d d
o o o o o o o o . . . ~
N x x
c~
x~
©
+1 ~
+l
~ o
+1 .~
cq .,-.
+1 c,I ,d ©
O
c r
©
~N3
U ©
e~
0 c~
d
c~
cj
0 © d
N ©
.H x
x~ o a

66
addition of cell mass and nutrients according to option A caused a dilution of 2%. Compiling the analysis re- sults of the hydrolysate from the different laboratories (Table 3), the mean value of total amount of sugar was found to be 43.6 g/1 with a standard deviation of 5.2 g/1 and a concentration range of 31.%54.7 g/1. For glucose a mean value of 6.2 g/1 with a standard deviation of 1.5 g/1 and a concentration range of 3.6-8.1 g glucose/1 was found. Xylose gave a mean value of 32.6 g/1 with a standard deviation of 4.0 g/1 and a concentration range of 23.2-40.1 g xylose/1. For arabinose a mean value of 4.8 g/1 with a standard deviation of 1.8 g/1 and a con- centration range of 3.0-7.7 g arabinose/1 was found. A mean value of 4.3 g/1 with a standard deviation of 0.6 g/1 and a concentration range of 3.4-4.9 g/1 was ob- tained for acetic acid. One reported analysis result has been omitted because of unexplainable low figures for sugars, 3.0 g glucose/l, 16.0 g xylose/1 and 2.0 g arabi- nose/1 (both from Table 3 and the statistical compari- sons).
Fermentations
The fermentation performance of the bacterium E. coli, natural as well as recombinant, naturally occur- ring yeasts (P. stipitis and S. cerevisiae), recombinant yeasts (S. cerevisiae) and a fungus (F. oxysporum) were investigated.
Option A
The fermentations performed under option A were consistent with respect to nutrient supplementation, pH, temperature, and aeration in order to compare the performance of different micro-organisms in the same hydrolysate.
Naturally occurring micro-organisms. E. coli ATCC 11303 produced 2.9 g/1 of biomass but no ethanol or other products (Tables 4, 5 and 6). None of the yeasts P. stipitis CBS 5773 (oz), P. stipitis CBS 5773 (~), P. sti- pitis CBS 6054 (6) and P. stipitis R produced any etha- nol, biomass or by-products (Tables 4, 5, and 6). P. sti- pitis CBS 5773 (y) produced 8.4 g/1 of ethanol with a yield of 0.18 g/g and a maximum volumetric productiv- ity of 0.20 g/l per hour, in addition to 0.19 g/g of xylitol (Tables 4, 5, and 6). P. stipitis CBS 6054 (E) gave a yield of 0.03 g/g, the maximum volumetric productivity was 0.18 g/1 per hour and the maximum specific pro- ductivity was 0.04 g/g per hour: xylitol formation was 0.02 g/g (Tables 4, 5 and 6). S. cerevisiae, bakers' yeast, is not able to utilise xylose, but can utilise the isomeric form of xylose, xylulose. The bacterial enzyme XI cata- lyses the isomerisation of xylose to xylulose. When us- ing S. cerevisiae isolate 3 in combination with XI, the yield was 0.06 g/g, the maximum volumetric productivi- ty 1.04 g/l per hour, the maximum specific productivity 0.31 g/g per hour and the xylitol formation was 0.22 g/g (Tables 4, 5 and 6).
Table 4. Experimental conditions and resulting yields
Organism Option pH Temperature Initial dry Growth Total sugar Ethanol Yield (g/g) Yield (g/g) (° C) weight (g/l) (g/l) (g/l) (g/l) (total) (consumed)
Escherichia coli A T C C 11303 A 7.0 30 5.5 2.9 40.9
E. coli B, pLOI 297 A 7.0 30 4.0 2.5 40.6 E. coli B, pLOI 297 A(a)" 7.0 30 3.0 3.5 41.6 E. coli B, pLOI 297 A(a) a 7.0 30 0.5 0.5 51.6 E. coli B, K O l l A 7.0 30 5.0 n.s. 39.3
Pichia stipitis CBS 5773 a A 5.5 30 5.0 0.0 31.7 P. stipitis CBS 5773/3 A 4.5 28 6.4 0.0 54.7 P. stipitis CBS 5773 y A 6.0 30 2.6 2.9 47.6 P. stipitis CBS 6054 8 A 5.5 30 4.8 0.0 41.3 P. stipitis CBS 6054 • A 5.0 30 4.7 0.8 44.1 P. stipitis R A 5.5 30 4.6 0.0 41.7
Saccharomyces cerevisiae 3 + XI A 5.5 30 3.4 5.6 42.4
S. cerevisiae TJ1 ( X R + X D H ) A 6.0 30 2.3 1.2 47.8 S. cerevisiae H550 ( X R + X D H ) A 5.5 30 5.0 2.9 42.2
E. coli B, K O l l B 7.0 30 5.0 n.s. 39.3
P. stipitis CBS 5773 fi B(b) b 4.5 28 6.4 0.0 54.7 P. stipitis CBS 5773/3 B(c) b 4.5 28 6.4 0.0 54.7 P. stipitis CBS 6054 • B 5.0 30 5.6 0.9 44.8
S. cerevisiae H550 ( X R + X D H ) B 5.5 30 5.0 3.2 39.8
Fusarium oxysporum VTT-D-80134 B 5.0 30 5.3 0.0 21.0 E. coli B, pLOI 297 C 7.0 30 0.5 0.5 49.8 E. coli B, K O l l C 6.0 30 5.0 n.s. 39.3 P. stipitis CBS 5773/3 C c 4.5 28 6.4 2.0 24.8 P. stipitis CBS 6054 • C 5.0 30 7.0 2.0 43.5
0.0 0.00 0.00
2.5 0.06 0.08 9.5 0.23 0.25 7.9 0.15 0.21 9.4 0.24 (96 h) n.s.
0.0 0.00 0.00 0.0 0.00 0.00 8.4 0.18 (72 h) 0.21 (72 h) 0.0 0.00 0.00 1.5 0.03 0.21 0.0 0.00 0.00
2.7 0.06 0.06
3.5 0.07 (72 h) 0.13 (72 h) 2.9 0.07 0.31
14.0 0.36 (96 h) n.s.
0.0 0.00 0.00 0.0 0.00 0.00 1.9 0.04 0.24 3.4 0.09 0.44
0.8 0.04 (92 h) 0.20 (92 h) 7.3 0.15 0.22
21.4 0.54 0.55
8.3 0.33 (44 h) 0.39 (44 h) 13.3 0.31 0.41
For options, see Materials and methods; XI, xylose XR, xylose reductase; XDH, xylitol dehydrogen~ise a No washing of inoculum prior to inoculation
isomerase; b For details, see text c Fed-batch, no detoxification of the hydrolysate
Table 5. Resulting productivities
Organism Option Maximum volumetric Maximum specific productivity (g/l/h) productivity (g/g/h)
67
E. coli ATCC 11303 A 0.00 0.00 E. coli B, pLOI 297 A 0.02 < 0.01 E. coli B, pLOI 297 A(a) a 0.15 0.05 E. coli B, pLOI 297 A(a) a 0.10 0.20 E. coli B, K O l l A 0.88 0.18
P. stipitis CBS 5773 a A 0.00 0.00 P. stipitis CBS 5773/3 A 0.00 0.00 P. stipitis CBS 5773 y A 0.20 0.08 P. stipitis CBS 6054 6 A 0.00 0.00 P. stipitis CBS 6054 • A 0.18 0.04 P. stipitis R A 0.00 0.00
S. cerevisiae 3 +XI A 1.04 0.31
S. cerevisiae TJ1 (XR + XDH) A 0.05 0.02 S. cerevisiae H550 (XR+XDH) A 0.76 0.15
E. coIi B, K O l l B 1.20 0.24
P. stipitis CBS 5773/3 B(b) b 0.00 0.00 P. stipitis CBS 5773/3 B(c) b 0.00 0.00 P. stipitis CBS 6054 • B 0.20 0.04
S. cerevisiae H550 (XR+XDH) B 0.83 0.17
F. oxysporum VTT-D-80134 B < 0.01 < 0.01
E. coli B, pLOI 297 C 0.11 0.22 E. coli B, K O l l C 1.32 0.26
P. stipitis CBS 5773/3 C ° n.s. n.s. P. stipitis CBS 6054 • C 0.32 0.05
~° See footnotes to Table 4
Table 6. By-products produced
Organism Option Lactate Lactate Xylitol Xylitol (g/g total sugar) (g/g sugar consumed) (g/g total sugar) (g/g sugar consumed)
E. coli ATCC 11303 A
E. coli B, pLOI 297 A 0.23 0.30 E. coli B, pLOi 297 A(a) a 0.31 0.34 E. coli B, pLOI 297 A(a) a 0.18 0.25 E. coli B, K O l l A P. stipitis CBS 5773 a A P. stipitis CBS 5773/3 A P. stipitis CBS 5773 3' A P. stipitis CBS 6054 6 A P. stipitis CBS 6054 e A P. stipitis R A
S. cerevisiae 3 + XI A
S. cerevisiae TJ1 (XR + XDH) A S. cerevisiae H550 (XR + XDH) A E. coli B, K O l l B
P. stipitis CBS 5773/3 B(b) b P. stipitis CBS 5773/3 B(c) b P. stipitis CBS 6054 • B S. cerevisiae H550 (XR +. XDH) B
F. oxysporum VTT-D-80134 B
E. coli B, pLOI 297 C 0.17 0.25 E. coli B, K O l l C P. stipitis CBS 5773/3 C c P. stipitis CBS 6054 e C
0.00 0.00 0.00 0.00 0.19 0.22 0.00 0.00 0.02 0.11 0.00 0.00 0.22 0.22
0.23 0.41 0.04 0.21
0.00 0.00 0.00 0.00 0.02 0.11
0.04 0.19
0.03 0.03
a-~ See footnotes to Table 4

68
Recombinant micro-organisms. The washing of E. coli B, pLOI 297 cells severely affected both yield and pro- ductivity; washing prior to inoculation was omitted un- der option A (a). For E. coli B, pLOI 297 the yield was 0.06g/g under option A and 0.23g/g under option A (a) (Table 4). The maximum volumetric productivity was 0.02 g/1 per hour and 0.15 g/1 per hour, respectively (Table 5). The yield for E. coli B, K O l l was 0.24 g/g, the maximum volumetric productivity was 0.88 g/1 per hour (Tables 4 and 5). For all fermentations with E. coli B, KOl l , the medium was filter-sterilized before use. Both recombinant S. cerevisiae strains gave a yield of 0.07 g/g (Table 4). The maximum volumetric pro- ductivity of ethanol was 0.05 g/1 per hour for S. cerevi- siae TJ1 and 0.76 g/1 per hour for S. cerevisiae H550, and the formation of xylitol was 0.23 g/g and 0.04 g/g, respectively (Tables 5 and 6).
Option B
Fermentations under option B were performed under optimal conditions for each type of micro-organism chosen by each participating laboratory. The fermenta- tions were performed without detoxification of the hy- drolysate. Nutrient supplementation and the amount of available 02 were changed. Under option B, the nat- urally occurring micro-organisms P. stipitis CBS 5773 (/3), P. stipitis CBS 6054 (e) and F. oxysporum VTT- D-80134 were investigated, and among the recombi- nants E. coli K O l l and S. cerevisiae H550 were investi- gated.
Naturally occurring micro-organisms. P. stipitis CBS 5773 (/3) was investigated under two conditions B (b) and B (c). Options B (b) and B (c) did not result in any improvement compared to option A, i.e. no ethanol, biomass or by-products were produced. Under option B (b) P. stipitis CBS 5773 (/3) fermented a hydrolysate that had been supplemented with the same chemicals as under option A and thereafter autoclaved, and un- der option B (c) the hydrolysate was supplemented with IFP mineral medium and thereafter autoclaved. IFP mineral medium was obtained by blending 5 ml so- lution A + 5 ml solution B + 50 ml solution C + 1.3 g yeast extract. Solution A contained 1.2 g/1 of CaO, 0.4 g/I of ZnO, 5.4 g/1 of iron(III) chloride, 0.35 g/1 of MgO, 0.25 g/1 of CuSO4" 5 H20, 0.24 g/1 of CaC12" 5 HzO, 0.06 g/1 of orthoboric acid and 1.3 ml/1 of concentrated HC1. Solution B contained 10.1 g/1 of MgO and 45 ml/1 of concentrated HC1, and solution C contained 64 g/1 of urea, 12 g/1 of KHzPO4 and 1.8 g/1 of disodium hydrogen phosphate. IFP mineral medium (5 ml) was mixed with 45 ml autoclaved hydrolysate. P. stipitis CBS 6054 (e) was fermented in hydrolysate sup- plemented with 1.7 g/1 of YNB (without amino acids and ammonium sulphate), 1.0 g/1 of casamino acids and 2.2 g/1 of urea. This resulted in only marginal changes in yield, maximum volumetric productivity, maximum specific productivity and xylitol formation compared to option A (Tables 4, 5, and 6). F. oxysporum VTT-D-
80134 produced ethanol with a yield of 0.04 g/g and a maximum specific productivity of less than 0.01 g/g per hour when the hydrolysate was supplemented with the same nutrients as in the cultivation media (Tables 4 and 5). In addition the fermentation was performed under an N2 atmosphere.
Recombinant micro-organisms. For E. coli B, K O l l the yield increased by 50% to 0.36 g/g, the maximum volumetric productivity and the maximum specific pro- ductivity increased by 35% to 1.20 g/1 per hour and to 0.24 g/g per hour, respectively (Tables 4 and 5) when the hydrolysate was supplemented with 1 ml of corn steep liquor/50 ml hydrolysate. Oz-limited conditions (25 ml of the hydrolysate in a 25 ml flask, sealed with a rubber stopper supplied with a cannula for COz and stirred gently with a magnetic stirrer) did not improve the yield for recombinant S. cerevisiae H550 (Table 4), but the maximum volumetric and specific productivity increased by 34% to 1.02 g/1 per hour and 0.20 g/g per hour (Table 5).
Option C
Fermentations under option C were performed with detoxification of the hydrolysate and/or adaptation of the micro-organism to the hydrolysate in addition to the optimal nutrient and oxygenation conditions pre- sented under option B. The organisms investigated un- der this option were P. stipitis CBS 5773 (~), P. stipitis CBS 6054 (e), E. coli B, pLOI 297 and E. coli B, KOll . The detoxification method, over-liming with Ca(OH)z, was chosen by each individual laboratory. In one case a form of adaptation, fermentation of a diluted hydro- lysate in fed-batch mode, was performed.
Naturally occurring micro-organisms. When the fer- mentation with P. stipitis CBS 5773 (13) was performed as a fed-batch fermentation, started from 0.6 1 of pro- duction culture cell mass and then adding 0.5 1 of auto- claved hydrolysate in a fed-batch mode, the yield was 0.33 g/g, which should be compared to the two pre- vious conditions where no ethanol was produced at all (Table 4). This procedure allowed P. stipitis CBS 5773 (/3) to ferment a diluted hydrolysate thereby avoiding pronounced inhibition. Fed-batch fermentations may be considered as a way of adapting the micro-organ- isms to the hydrolysate. P. stipitis CBS 6054 (e) was adapted to the hydrolysate by successively subculturing it in 25%, 50% and 75% hydrolysate for 48 h at 30 ~ C before fermenting a hydrolysate detoxified by overlim- ing. This resulted in an increased yield by almost a fac- tor of ten [0.04 g/g to 0.31 g/g (Table 4], but only a mar- ginal increase in the maximum productivity (Table 5).
Recombinant micro-organisms. E. coli B, pLOI 297 and E. coli B, K O l l were fermented in a detoxified hydrolysate (over-limed). This resulted in a yield o f 0.15 g/g for E. coli B, pLOI 297 compared to 0.54 g/g for E. coli B, K O l l (Table 4). The maximum volumet-

ric productivities were 0.11 g/1 per hour and 1.32 g/1 per hour, respectively, and the maximum specific produc- tivities were 0.22 g/g per hour and 0.26 g/g per hour, respectively (Table 5). The initial cell mass concentra- tion was 0.5 g/1 (E. coli B, pLOI 297) and 5 g/1 (E. coli B, KOll ) , which to some extent explains the maximum volumetric productivity difference between the two re- combinant E. coli. E. coli B, pLOI 297 produced 0.24 g lactate/g sugar (mean value) under option A and A (a) and 0.17 g/g under option C (Table 6), compared to no reported lactate production for E. coli B, KOll . Clear- ly, E. coli B, pLOI 297 has a different fermentation pattern compared to E. coli B, KOll . In addition, the fermentation conditions for the two recombinant E. coli differed with respect to pH, nutrient Supplementa- tion and initial cell dry weight (Beall and Ingrain 1992; Lawford and Rousseau 1992).
Xylitol production
During ethanol fermentation xylitol formation is nor- mally an unwanted by-product but xylitol per se is used in the food industry as an anticariogenic sweetener as well as a sugar substitute for diabetics. Production by naturally occurring xylose-utilising yeasts cannot be used in the food industry since they are not classified as GRAS (Generally Recognized As Safe) organisms. S. cerevisiae is a GRAS organism and therefore a xyli- tol-producing recombinant S. cerevisiae may be more easily accepted in the food industry. The recombinant S. cerevisiae H477 harbouring the gene coding for XR from P. stipitis CBS 6054 is an organism constructed for the production of xylitol (Hallborn et al. 1991). The xylitol yield for S. cerevisiae H477 was 0.08 g xylitol/g xylose under option A (Table 7). Under option B, in which glucose was added to the fermentation in a fed- batch pattern, the yield was 0.72 g xylitol/g xylose. Conversion of xylose to xylitol does not produce any energy for the cells, therefore a co-substrate (glucose) was needed for growth and for cofactor regeneration (Hallborn et al., unpublished data). The xylitol yield in grams per gram of xylose consumed was 0.79 g/g under option A, and 0.99 g/g under option B.
Discussion
The sugar and product analysis data are of crucial im- portance for the significance of the reported fermenta-
69
tion parameters. Since eight different laboratories ana- lysed the same hydrolysate (corn-cob hydrolysate, dis- tributed at pH 1.5 to avoid sugar loss due to metabolic activity) this study gives an idea of the problems and the accuracy when analysing complex substrates. The main components in the corn-cob hydrolysate were glucose, xylose, arabinose and acetic acid with a mean value of 6.2 g/l, 32.6 g/l, 4.8 g/1 and 4.3 g/l, respectively. For the total amount of sugar the concentration range was as much as 31.7-54.7 g/1 (Table 3). In a fermenta- tion where 18.1 g/1 of ethanol is obtained, the corre- sponding yield ranges from 0.33 g/g to 0.57 g/g when the yields are recalculated using the lowest and the highest value of total amount of sugar, respectively. This indicates an uncertainty in the comparison of re- sults from different investigations. Despite the scatter in analysis results, the conclusions drawn from this study are valid because the fermentation results for dif- ferent organisms and for different fermentation op- tions differ enough to allow conclusions to be drawn.
Several participants pointed out the difficulties in- volved with analysing lignocellulose hydrolysates due to the complex matrix of the hydrolysate, lack of suita- ble analytical instruments and lack of trained person- nel for interpretation of chromatograms. Linddn and Hahn-H~igerdal (1989a) developed a separation meth- od for the analysis of lignocellulose hydrolysates using spent sulphite liquor as the model substrate. The sepa- ration was performed using a precolumn and two HPX-87H (Biorad) columns coupled in series to en- hance the resolution. The use of a precolumn results in a longer life-time of the main columns and an in- creased stability of the analysis system. This separation column system is suitable for separation of glucose, xy- lose and arabinose, which were the main sugar compo- nents in the corn-cob hydrolysate. When a hydrolysate also contains galactose and mannose, common in soft- wood hydrolysates, the sugars must be separated on an HPX-87P column (BioRad). More selective analysis methods for sugar and product analysis in complex ma- trices are presently under development (Marko-Varga et al. 1990; Buttler et al. unpublished data). An addi- tional control of the accuracy of the analytical proce- dure would have been to carry out a complete carbon balance of individual fermentations (Skoog and Hahn- H~igerdal 1990; Senac and Hahn-H~igerdal 1991; Ols- son and Hahn-H~igerdal 1993). A carbon balance can also provide information about the metabolic path- ways. In the present study it was not possible to make carbon balances due to the incomplete analysis results reported.
Table 7. Xylitol production by S. cerevisiae H477
Organism Option Initial dry Growth pH Temperature Xylitol yield Xylitol yield weight (g/l) (g/l) (° C) (g/g xylose) (g/g xylose consumed)
S. cerevisiae H477 (XR) A 5 2.2 5.5 30 0.08 0.79 S. cerevisiae H477 (XR) B 5 5.7 5.5 30 0.72 0.99

70
A micro-organism suitable for the fermentation of lignocellulose hydrolysates should have the following qualities: (i) be able to ferment all sugar components in the hydrolysate to ethanol at a reasonable rate and (ii) be tolerant of potential inhibitors in the hydrolysate. The study was designed to test the investigated micro- organisms under three consistent options: a basic ex- perimental fermentation and two fermentations where additional media components and detoxification of the hydrolysate were used. The latter two options general- ly represent more costly alternatives.
In the economic evaluation of fuel ethanol produc- tion, by far the most important factor is the total yield as the raw material cost is of prime importance (Wright 1988; Hinman et al. 1989). Yields should be compared to the theoretical yield, which is generally considered to be 0.51 g/g, but other possible theoretical yields have been discussed (Slininger et al. 1987).
The total volumetric productivity (Fig. 1) deter- mines the capital investment cost, i.e. the size of fer- mentors. Almost any volumetric productivity can be achieved by increasing the concentration of cell mass, although at the cost of carbon. The specific productivi- ty (Fig. 1, Table 2), on the other hand, gives a measure of the efficiency of the fermenting micro-organism. Productivities (volumetric and specific) are often used in the literature as an indication of the efficiency of dif- ferent micro-organisms under given fermentation con- ditions. The value of comparing productivities ob- tained in different studies is highly questionable since they can be calculated differently especially when batch processes are considered. It is particularly un- clear when a batch process should be considered ended. In order to compare productivities the calcula- tions have to be clearly defined. In the present study no total productivity figures are reported because not enough data were reported and/or the sampling points were too infrequent to identify the time at which max- imum ethanol concentration was reached. Additional- ly, in the literature the maximum productivity is often reported together with the maximum ethanol concen- tration, although in typical batch fermentations these values are seldom connected (Fig. 1).
Not only yield and productivity are of importance, but also chemicals and process steps that increase pro- duction costs. A low media pH (Esser and Karsch 1984; Bj6rling and Lindman 1989) and a fast produc- tion time reduce the contamination risk and therefore contribute to process economy, whereas the use of me- dium sterilisation and sterile techniques add extra costs to the process. Detoxification with molecular sieves and mixed-bed ion resins have been reported to re- move sugars from the hydrolysate (Tran and Cham- bers 1986), but contradictory results (detoxification with calcium hydroxide, extraction with organic sol- vents, treatment with ion-exchange and adsorption re- sins) have also been reported (Frazer and McCaskey 1989). The Ca(OH)2 detoxification treatment used in the present study is not economically sound since both chemicals and equipment add costs. Detoxification of the hydrolysate costs 0.40 SEK in the chemical cost
alone of Ca(OH)2 (based on the price for a consump- tion of 3500 metric tons per year, Chematur, Sweden, 1993), which should be compared with the world mar- ket price of ethanol, 2.6-3.4 SEK (1 SEK equals 0.13 US dollars). Detoxification methods such as organic solvent extraction and ion exchange have been consid- ered to be even more costly than Ca(OH)2-pretreat- merit (Frazer and McCaskey 1989).
After adaptation and dilution of the hydrolysate, P. stipitis CBS 5773 (/3) gave a yield of 0.34 g/g, and after detoxification in combination with adaptation P. stipi- tis CBS 6054 (e) gave a yield of 0.31 g/g. Detoxification and/or adaptation is a prerequisite for obtaining rea- sonable yields with P. stipitis, indicating that this mi- cro-organism is highly sensitive to inhibitors present in the corn-cob hydrolysate. The inhibitor sensitivity is in agreement with earlier results with P. stipitis (van Zyl et al. 1988; Wilson et al. 1989). None of the other P. stipitis strains nor the fungus F. oxysporum or the bac- terium E. coli ATCC 11303 produced more than 0.04 g ethanol/g total sugars (Table 4). The poor results ob- tained with these micro-organisms make them unsuita- ble for fuel ethanol production according to this corn- cob hydrolysate study. E. coli B, K O l l was the most promising ethanol producer in the corn-cob hydroly- sate. This organism gave the highest yield in all fer- mentation options [0.24g/g (A), 0,36g/g (B) and 0.54 g/g (C), Table 4]. Recombinant E. coli has the ad- vantage of giving high total volumetric productivities even at low cell densities (0.033-330mg dry cell weight/l), at least in laboratory substrates (Beall et al. 1991). However, for maximum fermentation perform- ance this organism needed detoxification of the corn- cob hydrolysate. E. coli B, pLOI 297 has given reason- able yields for sQme hydrolysates without detoxifica- tion (Lawford and Rousseau 1993). A disadvantage for bacteria in general is the requirement for a fermenta- tion pH of 6.0 to 7.0, which increases the contamina- tion risk. In the present study the fermentations with rec E. coli B, K O l l were performed in sterilized media. Yeast, on the other hand, can ferment well at low pH (3.0-4.0) (Esser and Karsch 1984; Bj6rling and Lind- man 1989). S. cerevisiae is well known to exhibit high tolerance to inhibitors present in non-detoxified ligno- cellulose hydrolysates (Lind6n and Hahn-H~igerdal 1989b; Olsson and Hahn-H~gerdal 1993) and tolerance to high ethanol concentrations (Brown et al. 1981). En- zymatic conversion of xylose to xylulose with XI fol- lowed by fermentation of xylulose by S. cerevisiae has been successfully performed in both a laboratory sub- strate and a lignocellulose hydrolysate (Gong et al. 1981; Lind6n et al. 1992). However, with the hydroly- sate used in this study, S. cerevisiae isolate 3 in combi- nation with XI did not perform well. This is presently under investigation. In the xylose fermentation using the recombinant S. cerevisiae carrying the gene for XR, xylose was converted to xylitol with an efficiency of 0.99 g xylitol/g xylose consumed when a co-substrate was added. The recombinant S. cerevisiae strains carry- ing the genes for XR and XDH showed poor ethanol production from xylose, which is in agreement with

71
earlier studies (Tant i rungki j et al. 1993; K6t te r and Ciriacy 1993). This indicates fur ther blocks in the me- tabol ism of xylose by S. cerevisiae no t earlier ob- served.
A comparab l e eva lua t ion of the suitability of micro- organisms for the f e rmen ta t ion of a di lute-acid-pre- t rea ted hydro lysa te tha t is r ich in xylose was pe r fo rm- ed. This w o r k gave rise to suggest ions for fu ture w o r k concern ing d e v e l o p m e n t of s tandard analysis me thods for l ignocellulose hydrolysates , eva lua t ion of the eco- nomic feasibility o f steril ization and different detoxifi- cat ion methods , and fur ther compar i son of micro-or- ganisms fe rmen t ing o the r l ignocellulose hydrolysates .
Acknowledgements. This study was performed within Interna- tional Energy Agency (IEA), subgroup Biomass utilization (Task X). The xylose fermentation research at the Department of Ap- plied Microbiology, Lund Institute of Technology/University of Lurid has been supported financially by The Swedish Natural Science Research Council (NFR), Swedish National Board for Industrial and Technical Development (NUTEK), Swedish Na- tional Board for Technical Development (STU), National Swe- dish Energy Administration (STEV), Swedish Ethanol Develop- ment Foundation (SSEU) and The Nordic Fund for Technology and Industrial Development (NIF). The participating groups have contributed with experimental data as well as valuable dis- cussions for which they are gratefully acknowledged.
References
Alterthum F, Ingram LO (1989) Efficient ethanol production from glucose, lactose and xylose by recombinant Escherichia coli. Appl Environ Microbiol 55:1943-1948
Ando S, Arai I, Kiyoto K, Hanai S (1986) Identification of aro- matic monomers in steam-exploded poplar and their in- fluences on ethanol fermentation by Saccharomyces cerevi- siae. J Ferment Technol 64:567-570
Beall DS, Ingram LO (1992) Conversion of hydrolysates of corn cobs and hulls into ethanol by recombinant Escherichia coli B containing integrated genes for ethanol production. Biotech- nol Lett 14: 857-862
Beall DS, Ohta K, Ingram LO (1991) Parametric studies of etha- nol production from xylose and other sugars by recombinant Escherichia coli. Biotechnol Bioeng 38:296-303
BjOrling T, Lindman B (1989) Evaluation of xylose-fermenting yeasts for ethanol production from spent sulfite liquor. En- zyme Microb Technol 11:240-246
Brown SW, Oliver SG, Harrison DEF, Righelato RC (1981) Ethanol inhibition of yeast growth and fermentation: differ- ences in the magnitude and complexity of the effect. Eur J Appl Microbiol Biotechnol 11:151-155
Buchert J, Niemel~ K, Puls J, Poutanen K (1990) Improvement in the fermentability of steamed hemicellulose hydrolysate by ion exclusion. Process Bioehem 25 : 176-180
Clark TA, Mackie KL (1984) Fermentation inhibitors in wood hydrolysates derived from the softwood Pinus radiata. J Chem Technol Biotechnol 34B:101-110
Esser K, Karsch T (1984) Bacterial ethanol production: advan- tages and disadvantages. Process Biochem 19:116-121
Fein JE, Tallim SR, Lawford GR (1984) Evaluation of D-xylose fermenting yeasts for utilization of a wood-derived hemicellu- lose hydrolysate. Can J Microbiol 30:682-690
Frazer FR, McCaskey TA (1989) Wood hydrolysate treatments for improved fermentation of wood sugars to 2,3-butanediol. Biomass 18:31-42
Gong C-S, Chen L-F, Flickinger MC, Chiang L-C, Tsao GT (1981) Production of ethanol from D-xylose by using D-xylose isomerase and yeasts. Appl Environ Microbiol 41:430-436
Hahn-Hggerdal B, Lind6n T, Senac T, Skoog K (1991) Ethanolic fermentation of pentoses in lignocellulose hydrolysates. Appl Biochem Biotechnol 28/29:131-144
Hahn-H~igerdal B, Hallborn J, Jeppsson H, Olsson L, Skoog K, Walfridsson M (1993) Pentose fermentation to alcohol. In: Saddler J (ed) Bioconversion of forest and agricultural plant residues. CAB International, Wallingford, UK, pp 231-290
Hallborn J, Walfridsson M, Airaksinen U, Ojamo H, Hahn-H~i- gerdal B, Penttil~i M, Ker~inen S (1991) Xylitol production by recombinant Saccharomyces cerevisiae. Bio/Technology 9:1090-1095
Hallborn J, Walfridsson M, Airaksinen U, Ker~nen S, Hahn-H~i- gerdal B, Penttil~ M (1992) Xylitol dehydrogenase activity in recombinant Saccharomyces cerevisiae. In: Breitenbach M, Schweyen RJ, Pammer M, R~ith J (eds) 16th International Conference on Yeast Genetics and Molecular Biology, Yeast vol 8 Spec. Iss. August 15-21, 1992. J Wiley & Sons Ltd., Chi- chester, pp s580
Hinman ND, Wright JD, Hoagland W, Wyman CE (1989) Xylose fermentation. An economic analysis. Appl Biochem Biotech- nol 20/21 : 391-401
HyvOnen L, Koivistoinen P, Voirol F (1982) Food technological evaluation of xylitol. Adv Food Res 28: 373-403
Jeffries TW (1983) Utilization of xylose by bacteria, yeasts and fungi. Adv Biochem Eng Biotechnol 27:1-32
Jones RP (1989) Biological principles for the effects of ethanol. Enzyme Microb Technol 11:130-153
Keim CR, Venkatasubramanian K (1989) Economics of current biotechnological methods of producing ethanol. TIBTECH 7: 22-29
Kosaric N, Wieczorek A, Cosentino GP, Magee RJ, Prenosil JE (1983) Ethanol fermentation. In: Dellweg H (ed) Biotechno- logy, vol 3. Verlag Chemie, Weinheim, pp 257-385
K~Stter P, Ciriacy M (1993) Xylose fermentation by Saccharo- myces cerevisiae. Appl Microbiol Biotechnol 38:776-783
Lawford HG, Rousseau JD (1992) Fuel ethanol from corn resi- due prehydrolysate by a patented ethanologenic Escherichia coli B. Biotechnol Lett 14:421-426
Lawford HG, Rousseau JD (1993) Improved pentose utilization efficiency using genetically engineered Escherichia coli. In: Klass DL (ed) 16th Annual Conference in Energy from Bio- mass and Wastes XVI. Institute of Gas Technology, Chicago, Ill, pp 559-597
Lind6n T, Hahn-H~gerdal B (1989a) HPLC determination of xy- lulose formed by enzymatic xylose isomerization in lignocellu- lose hydrolysates. Biotechnol Tech 3:189-192
Linddn T, Hahn-H~gerdal B (1989b) Fermentation of lignocellu- lose hydrolysates with yeasts and xylose isomerase. Enzyme Microb Technol 11:583-589
Lind6n T, Peetre J, Hahn-H~igerdal B (1992) Isolation and char- acterization of acetic acid-tolerant galactose-fermenting strains of Saccharomyces cerevisiae from a spent sulfite liquor fermentation plant. Appl Environ Microbiol 58:1661-1669
Lind6n T, Hahn-Hfigerdal B (1993) Activity and stability of xy- lose isomerase preparations from whole cells of Lactobacillus brevis in spent sulfite liquor. Enzyme Microb Technol 15: 321-325
Lynd LR, Cushman JH, Nichols RJ, Wyman CE (1991) Fuel ethanol from cellulosic biomass. Science 251 : 1318--1323
Magee RJ, Kosaric N (1985) Bioconversion of hemicellulosics. Adv Biochem Eng Biotechnol 32:61-93
Maiorella B, Blanch HW, Wilke CR (1983) By-product inhibition effects on ethanolic fermentation by Saccharomyces cerevi- siae. Biotechnol Bioeng 25:103-121
Marko-Varga G, Dominguez E, Hahn-H~igerdal B, Gorton L (1990) Selective determination of carbohydrates in fermenta- tion broths with on-line sample clean-up and catalytic reac- tion detection. Chromatographia 30:591-594
Melaja A, H~im~l~iinen L (1977) Process for making xylitol. US patent no. 4.008.285

72
Mfikinen KK (1979) Xylitol and oral health. Adv Food Res 25:137-158
Nishikawa NK, Sutcliffe R, Saddler JN (1988) The influence of lignin degradation products on xylose fermentation by Kleb- siella pneumoniae. Appl Microbiol Biotechnol 27:549-552
Ohta K, Beall DS, Mejia JP, Shanmugam KT, Ingrain LO (1991) Genetic improvement of Escherichia coli for ethanol produc- tion: chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase II. Appl Environ Microbiol 57:893-900
Ojamo H, Ylinen L, Linko M (1989) Mikrobiologinen valmistus- menetelm~i. Finnish patent no. 76377
Olsson L, Hahn-Hfigerdal B (1993) Fermentative performance of bacteria and yeasts in lignocellulose hydrolysates. Process Biochem 28: 249-257
Parekh SR, Yu S, Wayman M (1986) Adaptation of Candida she- hatae and Pichia stipitis to wood hydrolysates for increased ethanol production. Appl Microbiol Biotechnol 25:300-304
Parisi F (1989) Advances in lignocellulosic hydrolysis and in the utilization of the hydrolyzates. Adv Biochem Eng 38:53-87
Sanchez B, Bautista J (1988) Effects of furfural and 5-hydroxy- methylfurfural on the fermentation of Saccharomyces cerevi- siae and biomass production from Candida guilliermondii. Enzyme Microb Technol 10: 315-318
Schneider H (1989) Conversion of pentoses to ethanol by yeasts and fungi. CRC Crit Rev Biotechnol 9:1-40
Senac T, Hahn-H~igerdal B (1991) Effects of increased transaldo- lase activity on D-xylulose and D-glucose metabolism in Sac~ charomyces cerevisiae cell extracts. Appl Environ Mierobiol 57:1701-1706
Skoog K, Hahn-H~igerdal B (1990) Effect of oxygenation on xy- lose fermentation by Pichia stipitis. Appl Environ Microbiol 56: 3389-3394
Slininger PJ, Bolen PL, Kurtzman CP (1987) Pachysolen tanno- philus: properties and process considerations for ethanol pro- duction from D-xylose. Enzyme Microb Technol 9:5-15
Strickland RC, Beck MJ (1985) Effective pretreatment alterna- tives for the production of ethanol from hemicellulosic hard- wood hydrolyzates. In: Klass DL (ed) Symposium on Energy from Biomass and Wastes IX. pp 915-938
Tantirungkij M, Nakashima N, Seki T, Yoshida T (1993) Con- struction of xylose-assimilating Saccharomyces cerevisiae. J Ferment Bioeng 75: 83-88
Tran AV, Chambers RP (1986) Ethanol fermentation of red oak acid prehydrolysate by the yeast Pichia stipitis CBS 5776. En- zyme Microb Technol 8:439-444
Wilson JJ, Deschatelets L, Nishikawa NK (1989) Comparative fermentability of enzymatic and acid hydrolysates of steam- pretreated aspenwood hemicellulose by Pichia stipitis CBS 5776. Appl Microbiol Biotechnol 31 : 592-596
Wright JD (1988) Ethanol from biomass by enzymatic hydrolysis. Chem Eng Prog 84:62-74
Yu S, Wayman M, Parekh SK (1987) Fermentation to ethanol of pentose-containing spent sulphite liquor. Biotechnol Bioeng 29:1144-1150
Zacchi G, Skoog K, Hahn-H~igerdal B (1988) Economic evalua- tion of enzymatic hydrolysis of phenol-pretreated wheat straw Bioteehnol Bioeng 32: 460-466
Zyl C van, Prior BA, Preez JC du (1988) Production of ethanol from sugar cane bagasse hemicellulose hydrolyzate by Pichia stipitis. Appl Bioehem Bioteclinol 17: 357-369
Zyl C van, Prior BA, Preez JC du (1991) Acetic acid inhibition of D-xylose fermentation by Pichia stipitis. Enzyme Microb Technol 13: 82-86





























