Embed Size (px)
Citation preview

Journal of Reproductive Immunology ELSEVIER 31 (1996) 109-123
Ana C. Abadia- olina”, ConcepcGn
“hidad de Imunologia. Departamentn de Bioquirniea y Siologia Molecultw. Facultud de Medicina, Universidad de Granada, Avda. de Madrid s/n. 18012 Granada, Spain
bResearclt Group in Human Reproductive Immunobiology, Department qf Pathology, University of Cambridge, Cambridge, UK
‘Institute de Biotecnologia, Avda. de Madrid s/n, 18012 Granada, Spain
Received 20 November 1995; revised 1 March 1996; accepted 12 March 1996
Abstract
Flow cytometric data were used to compare the phenotype of term decidual lymphocytes and peripheral blood lymphocytes. Unlike peripheral blood lymphocytes. a significant percentage of CD3 +, CD4+, CDS’ and CD16+ term decidual lymphocyte populations expressed the CD69 activation marker. The relative proportions.of CD38 in CD3 + , CD4 + and CDS+ populations were more than twice as large in term decidual lymphocytes as in peripheral blood lymphocytts. As reported for early decidual lymphocytes, the expression of CD38 and CD69 by term decidua! lymphocytes suggests that these cells are also regionally activated. However, term deciduai lymphocytes showed no spontaneous cytotoxicity against normal trophoblast or its tumoral counterpart, JEG cells. After stimulation with interleukin- 2, these lymphocytes became qrtotoxic, 3s did peripheral blood lymphocytes. The relevance of this latter result to the immune control& the physiological and pathological invasion of the decidua by the trophoblast IS discus,ed.
Keywords: Human decidual lymphocytes: Maternal-fetal interface; NK cells: CD38; CD69
* Corresponding author.
0165-0378/96/$15.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved PIZ SO165-0378(96)00965-5

110 A.C. Abadia-Moha et ai. 1 Joma/ of Reproductive Inmtur~ology 31 (1996) 109-123
1. Introduction
During normal human pregnancy, the fetal trophoblasa invades the maternal uterine decidua. It has been proposed that this invasion could be controlled by an immunological mechanism, a hypothesis which was strengthened by the detection in early decidua of a high proportion of large granular lymphocytes (LGL) that showed natural killer (NK) activity against K562 target cells (King et al., 1989a). These lymphocytes were CD3-CD56+ but, unlike peripheral blood lymphocytes (PBL), they did not express CD16 (Starkey et al., 1988; King et al., 1989b; Bulmer et al., 199lb). However, when the spontaneous cytotoxic activity of early decidual lymphocytes (EDL) was tested against normal and tumoral trophoblast, no activity was found against either cell type (King et al., 1989a; King and Loke, 1990). King and Loke (1990) nonetheless found that EDL became highly cytotoxic against trophoblast when they were previously stimulated with interleukin-2 (IL-2). Furthermore, treatment with this cytokine also induced the expression of CD16 in EDL (King et al., 1992). A relationship between the expression of CD16 and high cytotoxic activity has also been demonstrated in EDL clones generated by culture with IL-2 (Christmas et al., 1990; Deniz et al., 1994). In a previous study, we demonstrated that a significant proportion of lymphocytes from human term decidua (term decidual lymphocytes, TDL) expressed CD16 (Vargas et al., 1993). The present study was designed to characterize further the TDL populations and to investigate their cytotoxic activity against normal and tumoral tro- phoblast .
2. Materials and methods
2.1. Extraction of term deciduai lymphocytes
Normal term placentae were collected from healthy pregnancies at the Departamento de Obstetricia y Ginecologia, Hospital Universitario de Granada. The method of extraction has been described elsewhere (King et al., 1989a). Briefly, samples of term decidua were dissected from the maternal-facing surface, identified macroscopically, and extensively washed in phosphate-buffered saline solution (PBS). Decidual fragments were finely minced between two scalpels in a small volume of RPM1 1640 medium and then pushed through a 53-pm sieve (Gallenkamp, Loughborough, UK). The resultant cell suspension was washed with RPM1 and layered on an equivalent volume of Lymphoprep (Flow Laboratories, Herts, UK) at room

temperature, and centrifuged for 20 m cells were collected from the interface, suspended in e cells were then suspended in complete culture medium ( I 1640, PO% fetal calf serum (FCS), 100 U/ml penicillin and 50 pg/mI gentamicin), and incubated for 2 h at 37°C in an atmosphere of 5% CO2 to allow adherent cells to attach to the plastic. Supernatant containing TDL was then c ed. Term decidual lymphocytes were #washed and suspended either in for flow cytometric analysis or in complete culture medium for stimulation with IL-2.
To obtain peripheral blood lymphocytes, blood sa s were taken from healthy volunteers. The blood was e volume of EDTA 0.2%, and the sampIes centri on Lymphoprep. Thereafter we used the same steps were used as in
Cells were labeiled by direct staining with monoclonal antibodies (Table 1). One hundred microliters of a suspension of 1 x lO’/ml decidual lymphocytes in PBS were incubated with 10 or 20 ~1 of the appropriate monoclonal antibody for 30 min at 4°C in the dark. As a negative control, cells were stained with FITC, PE-labelled mouse IgGl or IgG2a (Ortho Diagnostic Systems, Raritan, NJ). Cells were washed, suspended in 1 ml PBS and immediately analyzed in a flow cytometer. Although the isolated cells were mostly lymphocytes, cells were studied in the electrically gated lymphocyte cluster. The percentage of cells that were antibody positive was calculated by comparison with the appropriate control. Student’s t-test was used to compare data fro*m TDL and PBL.
2.3. Trophobht
First trimester placental tissue was obtained from vaginal termination of pregnancy as described previously (Loke and Burland, 1988). Chorionic villi were minced into small pieces and incubated for 10 min at 37°C in 0.25% trypsin and 0.02% EDTA. The resultant cell suspension was filtered through a muslin and then centrifuged at 400 x g for 5 min. The pellet was suspended in PBti, c layered onto Lymphoprep and centrifuged at 800 x g for 15 min. The band of cells was resuspended in Ham’s F12 medium supplemented with L-glutamine, antibiotics and 20% FCS, adjusted to 10’ cells/ml, and plated out onto 35-mm culture dishes precoated for 45 min with 20 pg/ml laminin (Sigma, St. Louis, MO). The dishes were incubated at 37°C in an atmosphere of 5% CO? for 2 days. This method selects
112 A.C. Abadia-Molina et al. / Journal of Reproductive Inmunology 31 (1996) 109- 123
extravillous cytotrophoblast, the type of trophoblast that invades decidua (Loke and Burland, 1988)
2.4. JEG choriocarcinoma cell line
Cells of the JEG choriocarcinoma cell line were maintained in complete culture medium RPM1 1640 containing 5% FCS.
2.5. Culture of lymphocytes with IL-2
Term decidual lymphocytes or PBL were cultured for 4 days in complete medium containing 100 U/ml of IL-2 (Sigma, St. Louis, MO). After this period of incubation, cells were washed and suspended in complete culture medium for cytotoxicity studies.
Table 1 Monoclonal antibodies (mAbs) used in this study
mAb Specificity Clone lg subclass Conjugated Obtained with” fromb
OKTl I OKT3 Anti-TCR-
a8- I Anti-TCR-
y&1 0KT4 OKT8 OKB20 OKM14 OKM15 OK-NK CD56 OKT26a Leu-17 Leu-23 OKT9 OKDR
CD2 CD3 TCRclP
TCRyG
CD4 CD8 CD20 CD14 CD15 CD16 CD56 CD25 CD38 CD69 CD71 HLA-DR
OKTll 0KT3 wT31
llF2
OKT4 OKT8 KU-B2 BKM14 OKM15 OK-NK NKl-nbl-1 OKT26a EBVCS-5 HB-7 OKT9 SC-2
lgG2a lgG2a lgG1
lgG1
lgG2b lgG2a lgG2b lgG2a
IgM lgM IgGl lgG2a IgGl IgGl IgGl lgG2a
FITC FITC, PE FITC
FITC BD
FITC FITC FITC FITC FlTC FITC PE FITC PE PE FITC PE
OD OD BD
OD OD OD OD OD OD CALTAG OD BD BD OD OD
“FITC, fluorescein-isothiocyanate; PE, phycoerythrin. bOD Ortho-Diagnostic System, Raritan, NJ; BD, Becton-Dickinson, Erembodegem-Aalst, Belgium; CALTAG, San Francisco, CA.

A.C. Abadia-Molirza et ul. 1 Journal of Reproducriw Inmunology 31 (1996) lO9- 123 113
2.6 Cy to toxicity studies
Trophoblast, cultured as indicated above, was removed fro with 0.25% trypsin/0.02% EDTA, washed and suspended in medium. Cells (2 x 104) were added to each well. of a 96-well flat plate (Becton ickinson, San Jose, CA) that had n pre-coated with 20 pg/ml laminin nd left to incubate overnight at 3 plates were washed twice with medium and 3 PCi of N Bucks, UK) in 35 ~1 of culture medium was ad to each well. After an overnight labelling period in a moist 37”C, 5% z atmosphenti, the cells were washed three times and then 100 ~1 of complete culture medium was added. JEG cells were also la overnight with 3 FCi Nai’CrO,/well. Effector cells from decidua or were added in different proportions in a volume of 100 pl culture medium. After 5 or 16 h of incubation at 37”C, 100 ~1 of supernatant were removed from each well and counted with a gamma counter. T e percentage of cytotoxicity was calculate the following formula:
% cytotoxicity
(test connts/min - spontaneous counts/min) = (rnaxifl?urIg T&n ts /min - spontaneous cow2 ts jmin)
x 10
3.1. Lymphocyte populations measured with single-iabelling fiow cytometry in term decidua and peripheral blood
Fig. 1 shows the proportions of lymphocytes in term decidua and peripheral blood, as determined with single-labelling flow cytometry. CD3 + lymphocytes were the most abundant type of lymphocyte in term decidua, although the proportion of these cells was smaller than that in PBL. Most T cells of term decidua expressed TCRorP and only a small proportion of T cells similar to the proportion in PBL were TCRyG + . The percentage of CD4+ TDL was lower than in PBL; however, no significant differences were found for CDS + lymphocytes. The proportion of CD20+ B cells was lower in TDL than in PBL. The amount of CD14’ and CD1 5+ cells was negligible in both types of lymphocyte. The percentages of C CD56+ lymphocytes were higher in TDL than in differences were found for CD25, CD71 and HLA-D

CD2 (6) __--
CD3 (13) .
TCRat3 (4) ~- i-- +t
TCRy3 (4)
CD4 (11)
CD8 (10)
CD20 (9)
CD14 (6)
CD15 (6)
CD16 (14)
0066 (5)
CD26 (6)
CDS2 (13) -----, ***
CD69 (12)
CD71 (5)
HLA.OR (7)
---, !..: PBL
TDL
cl 10 20 30 40 50 60 70 00 90 100
% stained cells
Fig. 1. Comparison between the proportions of populations of peripheral blood lymphocytes (PBL) and term decidual lymphocytes (TDL) by single-labelling flow cytometric analysis. Numbers in parentheses show the number of q:unples of each type of lymphocyte. *P <
0.05. **p < 0.01. **+p < 0.001.
CD38+ and CD69+ lymphocytes were more abundant in TDL than in PBL.
3.2. Lymphocyte populations measured with double-labelliug jlow cytometry in term decidua and peripheral blood
As in PBL, CD16+ TDL atso co-expressed CD56 (Table 2, Fig. 2). CD16+ TDL and CD3 + TDL were found to be virtually independent populations, as only a negligible proportion of cells were labeled with both antigens (n = 7, x = 0.2 f 0.4). Similar results were obtained for CD56+ and CD3+ TDL (n = 4, x = 1.2 + 0.9). Unlike EDL, in which CD56+ TCRyS+ cel!s and CD3 -TCRyS + cells were found (Mincheva- Nilsson et al., 1992), these types of lymphocytes were not detected in term decidua (not shown),
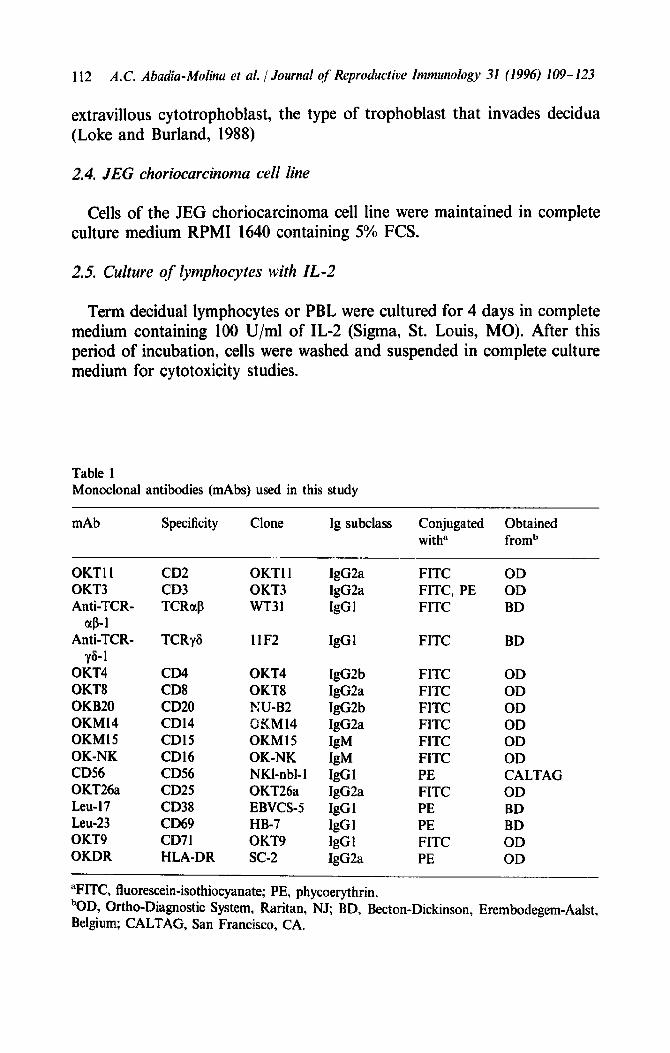
The relative proportions of CD38 + in CD3+, .CD4+ and CDS + lymphocytes were more than twice as large in TDL as in PBL, although no significant difference was found in the relative proportion of CD38 in CD16+ lymphocytes (Table 2, Fig. 3). CD8+CD38+ cells of both TDL and PBL were distributed in two populations: CDIIdimCD38+ and CD8b’igh’CD38 + (Fig. 3). As reported for PBL (Perussia et al., 1983), CD8dim cells were thought to be NK cells, whereas CD8bright cells were

Table 2 Cell subsets in different lymphocyte populations of term decidua and peripheral blood
Lymphocyte population
Cell subset T PBL lla P-value
CD3 CD3+CD38+ 38.95 + 15.50 13.04 & 7.07 7 CO.005 CD3 CD3+CD69+ 10.86 + 5.69 0.62 + 1.62 9 <O.OOl CD3 CD3+DR+ 6.63 f 4.34 3.98 f 2.64 7 NSb CD4 CD4+CD38+ 37.69 + 15.05 18.50 f. 5.80 7 to.05 CD4 CD4+CD69+ 7.82 + 5.88 0.23 & 0.34 6 co.05 CDS CD8+CD38+ 57.42 f 13.34 24.78 f 13.32 IO <O.OOl CDS CD8+CD69+ 11.89 + 8.88 0.48 & 0.28 5 <0.05 CD16 CD16+CD38+ 85.61 + 20.63 74.31 + 15.17 IO NS CD16 CD16+CD69+ 18.24 ~fr 9.07 1.34 + 1.35 7 <O.OOl CD16 CD16+DR + 6.58 + 3.53 14.87 _+ 10.31 5 NS CD16 CDl6+CD56+ 92.79 + 1.3 94.42 + 9.1 4 NS
The figures in the columns headed TDL (term decidual lymphocytes) and PBL (peripheral blood lymphocytes) are expressed as percentages of double-stained cells (second column) compared to the corresponding lymphocyte population {first column). “Number of samples of each type of lymphocyte studied. bNS, not significant.
considered to be cytotoxic T cells. ouble staining of PBL and TDL with anti-CD3 and anti-CDS mAb2 detected, in both types of lymphocytes, the presence of CD3 + CDSbrighr T cells together with a smaller population of CD3 - CD8”‘” 1 ymphocytes which were probably NK cells (not shown).
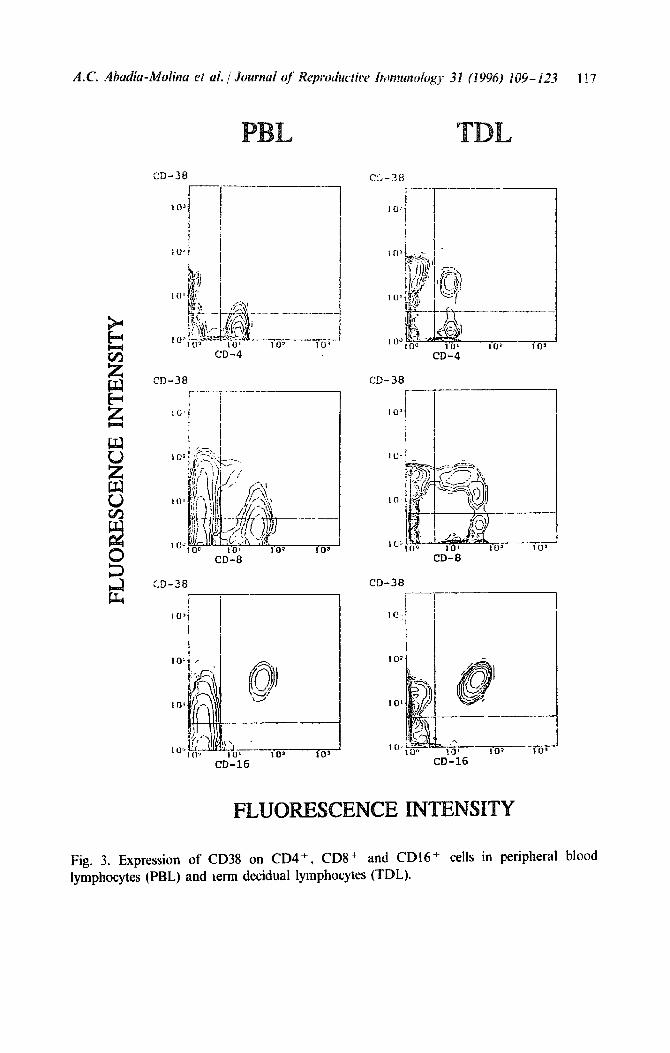
CD69, although expressed weakly, was significantly detected on CD3 +, CD4 + , CD8 + and CD1 6 + TDL, whereas its expression on the different populations of PBL was negligible (Table 2, Fig. 4). Nevertheless, double staining cytograms revealed the presence of a discrete lymphocyte popula- tion(s) of CD4 - CD8 - CD16- TDL which expressed CD69 more strongly (Fig. 4). No differences were found between TDL and PBL in the relative proportions of HLA-DR antigens in CD3 + and CD16 + lymphocytes (Table 2).
3.3. Anti-trophoblast cytotoxicity of term decidual lymphocytes and peripheral blood lymphocytes
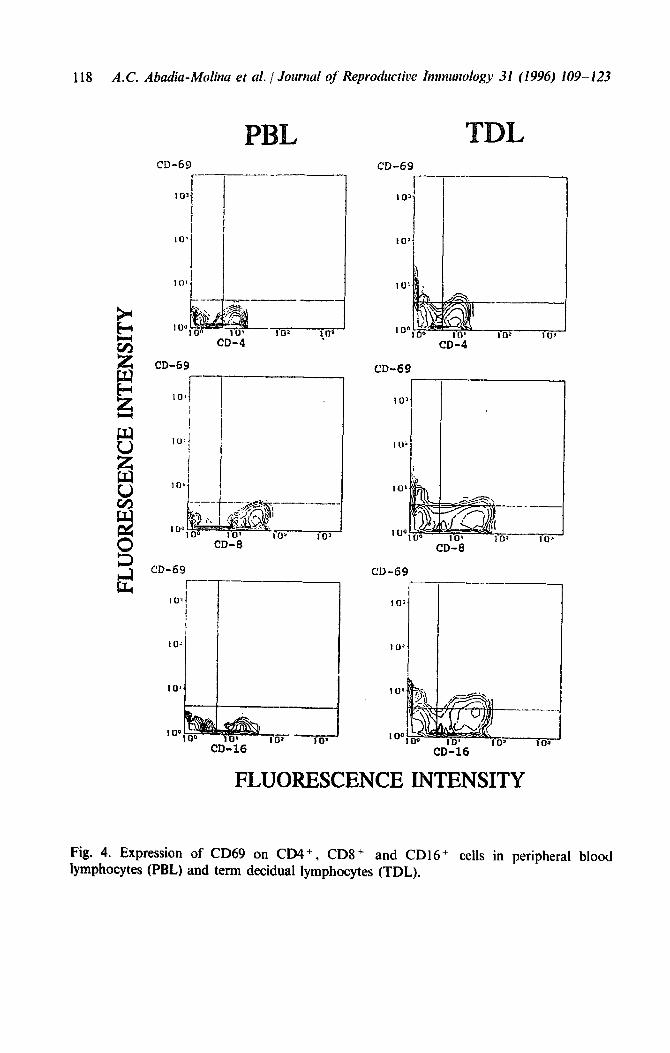
Neither TDL nor PBL showed spontaneous cytotoxicity against normal trophoblast or against its tumoral counterpart, the JEG cell line, in 5- and

116 A.C. Abadia-Molina et al. 1 Journal of Reproductive Immunology 31 (1996) 109-123
16-h incubation assays (Fig. 5). Both types of lymphocyte, however, became cytotoxic against normal trophoblast in a 5-h assay when they were previously incubated with IL-2. Although lymphokine-activated killer cells (LAK) of PBL also killed JEG cells, the activity of TDL-derived LAK was weak (Fig. 5).
4, Discussion
It has been widely reported that in early human decidua most lymphocytes are CD%+CD16- (Starkey et al., 1988; King et al., 1989b; Bulmer et al., 1991b). Although some authors observed that most T cells were TCRaP + and few or none expressed TCRyG in this tissue (Chernyshov et al., 1993), others found a large number of TCRyG+ cells, but smaller numbers of TCRccP + cells (Mincheva-Nilsson et al., 1992). In term decidua, however, the presence of significant proportions of TCRaP + cells, CD1 6 + CD56 + lymphocytes, and a small percentage of TCRyG + cells (Figs. 1 and 2) shows that TDL population profile is closer to that of PBL than EDL (Vargas et al., 1993). This similarity between TDL and PBL was not due to the presence of maternal blood in samples of term decidua as special care was taken to wash thoroughly the samples before processing them (see Section 2, Materials and methods). Furthermore, we previously observed a profile similar to that of the cell suspensions in term decidual sections (Vargas et al., 1993), which further confirms that our findings were not due
CD
PBL -56
I
103 “--T----7 1 I ,
!
I 10:
i I
i /
10.
------.--
IOC
CD-16
CD-56
i0'
FLUORESCENCE INTENSIT
Fig. 2. Co-expression of CD16 and CDS6 on peripheral blood lymphocytes (PBL) and term decidual lymphocytes (TDL).
I
I I I
CD-38
I i I
CD-38
CD-38
Fig. 3. Expression of CD38 on CD4’, CD8 + and CD16 + cells in peripheral blood
lymphocytes (PBL) and kerm decidual lymphocytes (TDL).
118 A.C. Abadia-Molina et al. / Journal of Reproductic)e Ittttnuttology 31 (1996) 109-123
CD-63
CD-69
CD-69
l----- 10~
i
! I
IO-‘
Fig. 4. Expression of CD69 on CD4+, lymphocytes (PBL)
CDS + and CD16 + cells in peripheral blood and term decidual lymphocytes (TDL).
CD-69
r-1
CD-4
L’D-69
100
CD-8
CD-69 1-
101,
-
IO'
IO’
CD-16
FLUORESCENCE INTENSITY
to contaminant blood. PBL: the proportion of appeared to be due tc the lo
Fig. 5. Cytotoxic activity against normal trophoblast and JEG cells in 5 and 16 h “Cr-release assays of peripheral Vood lymphocytes ( Cl- I’) and term decidual lymphocytes (
) with
and without stimulation with IL-2. Lymphokin-activated killer (LAK) activity was measured in 5 h assays. Spontaneous release in 5 h assays was 16 & 3% (average from 6 experiments) for trophoblast and 9 f 3 (average from 8 experiments) for JEG cells. Spontaneous release in 16 h assays was 65 f 5% (average from 3 experiments) for trophoblasts and 40 f 7% (average from 3 experiments) for JEG cells.

120 A.C. Abud’a-Molka et al. I Journal of’ Reprociuctiw In~t~~unology 31 (1996) 109- 123
In contrast, the percentages of CD16+ and CD56+ cells were higher in TDL than in PBL (Fig. 1). More importantly, we also found differences in the expression of activation markers such as CD38 and CD69. The relative proportions of CD69 in CD3+, CD4’ and CD8 + and CDl6+ lymphocytes, and of CD38 in CD3 + , CD4 + and CD8 + lymphocytes, were higher in TDL than in PBL (Table 2, Figs. 3 and 4).
The expression of activation markers seems to be a general characteristic of different types of decidual lymphocytes as these markers have also been described by several authors in EDL (King et al., 1991; Saito et al., 1992; Chernyshov et al., 1993; Mincheva-Nilsson et al., 1994). This status of immune activation appears to be paradoxical in a tissue in which immuno- suppression should predominate in order to guarantee maternal tolerance toward the fetus. Some authors have speculated that activated decidual lymphocytes play an immunosuppressive role (Mincheva-Nilsson et al., 1992). Others (e.g. Saito et al., 1992), in accordance with Wegmann’s immunotrophic theory (Athanassakis et al., 1987), suggest that these cells may synthesize cytokines thought to promote trophoblast growth. Nonethe- less, it is possible that activated cells control the invasion of trophoblast into the decidua and protect against the development of tumors such as moles or choriocarcinoma.
The expression of CD16 by cells of the NK lineage has been considered to reflect a state of maturation or activation (Nagler et al., 1989). In fact, CD3 - CD56 + CD 16 + lymphocyte clones obtained from early decidua were much more cytotoxic against both NK-sensitive and NK-resistant target cell lines than CD3 - CD56 + CD 16 - lymphocyte clones (Christmas et al., 1990; Deniz et al., 1994). Furthermore, in vitro experiments showed that CD56+ CDl6- EDL were unable to kill trophoblast in vitro (King et al., 1989a). However, when these EDL were treated with IL-2 they became cytotoxic to trophoblast cells (King and Loke, 1990). Interleukin-2 also induced the expression of CD16 on CD56+ EDL, suggesting that the CD56+CDl6+ lymphocytes are responsible for trophoblast killing (King et al., 1992). In this connection the detection of CDl6+ CD56+ lymphocytes not only at term (Fig. 2) but even from as early as mid-gestation (Erbach et al., 1993) suggested initially that these lymphocytes were involved in cytotoxic control of trophoblast invasion into the decidua, principally at the later stages of pregnancy when the mass of trophoblast becomes larger. The signs of activation in TDL (Table 2, Figs. 3 and 4), and the presence at term of a large proportion of CD8 + CD38 + cells (Fig. 3) - lymphocytes that, at less in peripheral blood, are related to NK cells and cytotoxic T cells (Perussia et al., 1983; Vanham et al., 1991) - also suggested that different popula- tions of d&dual lymphocytes may be involved in cytotoxic control of trophoblast invasion. However, this hypothesis is not supported by our

results on the cytotoxic activity of TDL were unable spo
was suggested al., 1991a) that the apparent absence of oblast killing b mphocytes was due to the fact that this
activity had been studied in short-term (4 or 5 h) incubation experiments, and that a longer incubatio of decid ual lymphocytes.
time might reveal the anti-trophoblast activity owever, we were also unable to detect this
activity when TDL and PBL (and also EDL, not shown) were incubated for 16 h with normal trophoblast and JEG cells (Fig. 5).
Although we cannot exclude the possibility that the high spontaneous release in the long-term assays may have decreased the sensitivity of our method, the results suggest that if decidual lymphocytes exert any control on trophoblast invasion they probably do so via a more physiological mechanism (e.g. tokines) than through cytotoxicity. However, TDL, and also (King and Loke, 1990) became against trophoblast when they were previously stimulated with though we cannot rule out that cytotoxic activity by PBL-derived LA against trophoblast reflects the possible artefactual nature of the LA phenomenon, this finding may well reproduce what occurs in pathological situations such as spontaneous abortion (Yokoyama et al., 1994), which may involve both decidual and peripheral blood lymphocytes as well as stimulation of both types of lymphocytes.
We thank Professor GonzBlez-Gbmez from the Departatnento de Obste- tricia y Ginecologia, Facultad de Medicina, Universidad de Granada, for supplying samples of human decidua. We also thank Ms. K. Shashok for improving the style of the manuscript. This work was supported by grants 91/0577 and 94/0711 from the Fondo de Investigaciones Sanitarias de la Seguridad Social (Spanish Ministry of Health).
References
Athanassakis, I.. Bleackley, KC., Paetkau, V., Guilbert. L.. Barr. P.J. and Wegmann, T.G. (1987) The immunostimulatory effect of T cells and T cell lymphokines on murine fetally derived placental cells. J. Immunol. 138. 37-44.
Bulmer, J.N., Morrison, L.. Longfellow. M. and Ritson, A. (199la) Leucocytes in human decidua: investigation of surface markers and function. In: Cellular and Molecular Biology of the Materno-Fetal Relationship (Chaouat. G. and Mowbray, J., eds.). pp. 189- 196. John Libbey, Eurotext Ltd, Pari 3.

122 A.C. Alr,cldiu-Molina et al. / Journa; oj’ Rqwductiw I~~mtunology 31 (1996) 109- 123
Bulmer, J.N., Morrison, L., Longfellow, M., Ritson, A. and Pace, D. (1991b) Granulated lymphocytes in human endometrium: histochemical and immunohistochemical studies.
Hum. Reprod. 6, 791-798. Chernyshov, V.P., Slukvin, 1.1. and Bondarenko, G.I. (1993) Phenotypic characterization of
CD7 + , CD3 + , and CD8 + lymphocytes from 1 st trimester human decidua using 2-color flow cytometry. Am. J. Reprod. Immunol. 29, 5-16.
Christmas, S.E., Bulmer, J.N., Meager, A. and Johnson, P.M. (1990) Phenotypic and functional analysis of human CD3 - decidual leucocyte clones. Immunology 71, 182- 189.
Deniz, G., Christmas, SE. Brew, R. and Johnson, P.M. (1994) Phenotypic and functional cellular differences betneen human CD3- decidual and peripheral blood leukocytes. J.
Immunol. 152, 4255-4261. Erbach, G.T., Semple. J.F., Mildfor, E.. Goguen, J., Osathanondh, R. and Kurnick, J.T.
(1993) Phenotypic characteristics of lymphocyte populations isolated from middle gesta- tion human placenta. J. Reprod. Immunol. 25, I- 13.
King, A., Birkby, C. and Loke. Y.W. (1989a) Early human decidual cells exhibit NK activity against K562 line but not against first trimester trophoblast. Cell. Immunol. 118, 337-344.
King, A., Wellings, V., Gardner, L. and Loke. Y.W. (1989b) Immunocytochemical charac- terization of the unusual large granular lymphocytes in human endometrium throughout the menstrual cycle. Hum. Immunol. 24, 195-205.
King, A. and Loke, Y.W. (1990) Human trophoblast and JEG choriocarcinoma cells are sensitive to lysis by IL-Zstimulated decidual NK cells. Cell. Immunol. 122, 435-448.
King, A., Balendran. N.. Wooding, P., Carter, N.P. and Loke. Y .W. ( 1991) CD3 leukocytes present in the human uterus during early placentation: phenotypic and morphologic characterization of the CD56+ + population. Dev. Immunol. 1, 169- 190.
King, A., Wheeler, R., Carter, N.P., Francis, D.P. and Loke, Y.W. ( 1992) The response of human decidual leukocytes to IL-2. Cell. Immunol. 141. 409-421.
Loke, Y.W. and Burland, K. (1988) Human trophoblast cells cultured in modified medium and supported by extracellular matrix. Placenta 9, 173- 182.
Mincheva-Nilsson, L., Hammarstriim, S. and Hammarstrom, M.L. (1992) Human decidual leukocytes from early pregnancy contain high numbers of y 6 + cells and show selective down-regulation of alloreactivity. J. lmmunol. 149. 2203 - 22 11.
Mincheva-Nilsson. L., Baranov, V., Yeung, M.M., Hammarstrom, S. and Hammarstriim. ML. (1994) Immunomorphologic studies of human decidua-associated lymphoid cells in normal early pregnancy. J. Immunol. 152, 2020-2032.
Nagler, A., Lewis, L., Cwirla, S. and Phillips, J.H. (1989) Comparative studies of human FcRlII-positive and negative natural killer cells. J. Immunol. 143, 3183-3191.
Perussia. B., Fanning, V. and Trinchieri, G. (1983) A human NK and K cell subset shares with cytotoxic T cells expression of the antigen recognized by antibody OKT8. J. Immunol. 131, 223-231.
Saito, S., Nishikawa, K., Morii, T., Narita, N., Enomoto, M. and Ichijo, M. (1992) Expression of activation antigens CD69, HLA-DR, interleukin-2 receptor-alpha (IL-2Rrx) and IL-2RP on T cells of human decidua at an early stage of pregnancy. Immunology 75. 710-712.
Starkey, P.M., Sargent, IL. and Redman, C.W.G. (1988) Cell populations in human early pregnancy: characterization and isolation of large granular lymphocytes by flow cytome- try. Immunology 65, 129- 134.
Vanham, G., Kestens, L., Penne, G., Goliav. C.. Gigase, P., Colebunders, R., Vandenbru- acne. M.. Goeman, J., van der Groen. G. and Ceuppens, J.L. ( 1991) Subset markers of

CD8( + ) cells and their relation to enhanced cytotoxic T-cell activity during human immunodeficiency virus infection. 9. Clin. Immunol. 11. 345-356.
Vargas, M.L., Santos. J.L., Ruiz, C., Montes, M.J.. Aleman. P.. Garcia-Tortosa, C. and Garcia-Ohvares, E. (1993) Comparison of the proportions of leukocytes in early and term human decidua. Am. J. Reprod. Immunol. 29, 1355140.
Yokoyama, M., Sano, M., Sonoda K., Nozaki, M., Nakamura, 6. and Nakano, J-J. (1994). Cytotoxic cells directed against placental cells detected in human habitual abortions by an in vitro terminal labelling assay. Am. J. Reprod. Immunol. 31, 197-204.





























