Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, JUIY 1994, p. 2450-2455 Vol. 60, No. 70099-2240/94/$04.00+0Copyright C 1994, American Society for Microbiology
Anaerobic Degradation of Methylmercaptan andDimethyl Sulfide by Newly Isolated Thermophilic
Sulfate-Reducing BacteriaYUICHI TANIMOTO* AND FRIEDHELM BAKt
Max-Planck-Institut fiir Terrestrische Mikrobiologie, D-35032 Marburg (Lahn), Federal Republic of GernanyReceived 15 December 1993/Accepted 20 April 1994
The complete oxidation of methylmercaptan (MSH) and dimethyl sulfide (DMS) with sulfate or nitrate aselectron acceptors was observed in enrichment cultures and dilution series using thermophilic fermentorsludge as the inoculum. Three new strains of thermophilic sulfate reducers were isolated in pure culture(strains MTS5, TDS2, and SDN4). Strain MTS5 grew on MSH and strain TDS2 grew on DMS whereas strainSDN4 grew on either MSH or DMS. The cellular growth yields were 2.57 g (dry weight)/mol of MSH for strainMTS5 and 6.02 g (dry weight)/mol of DMS for strain TDS2. All strains used sulfate, sulfite, or thiosulfate aselectron acceptors, but only strain SDN4 used nitrate. DMS and MSH were oxidized to CO2 and sulfide witheither sulfate or nitrate as the electron acceptor. Sulfate was stoichiometrically reduced to sulfide while nitratewas reduced to ammonium. All strains were motile rods, required biotin for growth, lacked desulfoviridin, hadDNA with G+C contents of 48 to 57 mol%, and probably belonged to the genus Desulfotomaculum. This is thefirst report of the oxidation of MSH and DMS by pure cultures of sulfate-reducing bacteria.
Volatile methylated sulfur compounds are intensively stud-ied since they play an important role in the global cycle ofsulfur and constitute climatically relevant trace gases in theatmosphere (1, 6). Natural and industrial sources of methyl-ated sulfur compounds have been found. Methylmercaptan(MSH) and dimethyl sulfide (DMS) are emitted from bothterrestrial and marine ecosystems (1). Methylated sulfur com-pounds are also found in kraft pulping drainage, being formedfrom lignin-derived aromatic compounds (11, 15). MSH- andDMS-forming anaerobes have been isolated from both fresh-water and marine sediments (2). The degradation of MSH andDMS by anaerobic microorganisms has also been observed.Several DMS-utilizing methanogens have been isolated fromdifferent habitats (9, 16, 17), and Finster et al. (9) isolated amethanogenic bacterium which grew on both MSH and DMS.The utilization of MSH and DMS in anoxic salt marshsediments appeared to be linked to sulfate reduction (13), butno sulfate reducers which degrade MSH or DMS have beenisolated so far. This paper reports the enrichment, isolation,and characterization of three new strains of anaerobic sulfatereducers which utilize MSH and/or DMS in freshwater me-dium.
MATERIALS AND METHODS
Source of organisms. All strains were isolated from thermo-philic fermentor sludge which was obtained from laboratory-and precommercial-scale methanogenic fermentors. The labo-ratory-scale fermentors were fed mainly a chemical complexmedium in tap water. On the other hand, the precommercial-scale fermentor was fed an evaporator condensate of kraftpulping waste, which contained 0.6 M methanol, 2.0 mM MSH,
* Corresponding author. Mailing address: Institute of Technology,Shimizu Corporation, No. 4-17, Etchujima 3-Chome, Koto-ku, Tokyo135, Japan. Phone: 81 33643 4311. Fax: 81 3 3643 7260. Electronic mailaddress: [email protected].
t Friedhelm Bak died on 27 December 1992, a sudden end to aresearch career of great promise.
18 mM DMS, and 2.2 mM dimethyl disulfide. Both laboratory-and precommercial-scale fermentors were operated at -53 to55°C. Strain MTS5 was isolated from a dilution series infreshwater liquid medium inoculated with sludge, while strainsTDS2 and SDN4 were isolated from enrichment cultures. Adilution series consisted of five tubes with successive 10-folddilutions. Desulfotomaculum thermobenzoicum DSM 6193 andDesulfotomaculum orientis DSM 765 were obtained from theDeutsche Sammlung von Mikroorganismen, Braunschweig,Federal Republic of Germany.
Media, isolation, and culture conditions. Defined, HCO3-C02-buffered, and sulfide-reduced media were used through-out the study (modified from the media of Widdel and Bak[24]). The medium contained the following (in grams per liter):NaCl, 1.0; MgCl2 . 6H20, 0.4; CaCl2 - 2H20, 0.15; KH2PO4,0.2; KCl, 0.5; NH4Cl, 0.25; NaHCO3, 2.5; and Na2S * 9H2O,0.24. Trace element solution (9) and three different vitaminsolutions were added to this medium.Two milliliters of nonchelated, acidic, trace element solution
was used for isolation and characterization except for stoichio-metries. For determination of stoichiometries, 1 ml of achelated trace element solution with EDTA was added insteadof the nonchelated, acidic, trace element solution. The com-position of the chelated trace element solution with EDTA wasas follows (in milligrams per liter): EDTA, 3,000; FeSO4.7H2O, 1,100; H3BO3, 300; CoCl2 - 6H2O, 190; MnCI2 - 4H2O,50; ZnCl2, 42; NiCl2- 6H2O, 24; NaMoO4 * 2H20, 18; andCuCl2 * 2H2O, 2. Three different vitamin solutions were sepa-rately prepared for stabilization of the vitamins in the solu-tions. One was sodium phosphate (50 mM; pH 7.0)-bufferedseven-vitamin solution, which contained the following (inmilligrams per liter): 4-aminobenzoate, 40; folic acid, 30; lipoicacid, 10; nicotinic acid, 100; calcium D-(+)-pantothenic acid,50; pyridoxamine-dihydrochloride, 100; and D-(+)-biotin, 10.The other two vitamin solutions were nonbuffered cyanocobal-amin solution (50 mg/liter) and acidic sodium phosphate (200mM; pH 3.4)-buffered thiamine hydrochloride solution (0.3mM/liter). Vitamin solutions were filter sterilized and added ata concentration of 1 ml/liter of medium. Organic complex
2450
on April 13, 2019 by guest
http://aem.asm
.org/D
ownloaded from

ANAEROBIC DEGRADATION OF MSH AND DMS 2451
growth factors (for example, yeast extract and casein hydroly-sate) were not used throughout this study except for the puritycheck test. Acetate was also not added as a carbon sourceexcept where indicated. Small amounts of sodium dithionite(.200,uM) were used as an additional reductant and addeddirectly before inoculation by using either a small dry crystal ora freshly prepared solution (24).Enrichment and isolation. Enrichments were carried out in
screw-cap bottles (50 ml) which were completely filled with thedefined medium and anoxic sludge (2%) as the inoculum.Isolation was also performed by using serial 10-fold dilutions ofanoxic sludge samples in the defined medium. The dilutionswere made in 16-ml test tubes sealed' with butyl rubberstoppers. Dilution series were gassed with an 02-free N2-C02(75:25, vol/vol) mixture. All cultures were incubated at 55°C inthe dark. The bacteria were isolated by repeated application ofdeep-agar dilutions (24) prepared with Gelrite instead of agar.
Purity and maintenance. Purity was checked by growthexperiments with complex medium containing 0.5% (wt/wt)yeast extract, fumarate (5 mM), pyruvate (5 mM), and glucose(or fructose) (5 mM). Addition cultures were microscopicallyexamined. Stock cultures were kept at 4°C in the dark andtransferred every 2 months.
Nutritional studies. Nutritional studies were performed induplicate, using butyl septum stopper-sealed test tubes con-taining 10 ml of the defined medium. Growth was determinedby measuring the optical density at 500 nm with a Bausch &Lomb Spectronic 70 spectrophotometer. Cultures were alsochecked by microscopy.
Determination of the GC content of DNA. The extraction ofDNA and determination of GC content were performed by theDeutsche Sammlung von Mikroorganismen. Cell disruptionwas done with a French pressure cell, and the DNA waspurified on hydroxyapatite (5). The DNA was hydrolyzed withP1 nuclease, and the nucleotides were dephosphorylized withbovine alkaline phosphatase (14). The resulting deoxyribo-nucleosides were analyzed by high-performance liquid chro-matography (20).
Determination of stoichiometries. Stoichiometries duringthe conversion of MSH and DMS were determined in batchcultures by endpoint determinations. Experiments were con-ducted with calibrated 120-ml serum bottles which were filledwith 55 ml of defined medium and inoculated with 5 ml of aculture grown under a gas phase of N2-CO2 (75:25, vol/vol).These serum bottles were sealed with black butyl rubberstoppers (Geo-Microbial Technologies, Inc., Ochelata, Okla.).MSH and DMS were injected through the stoppers at thebeginning of the experiment. MSH and DMS in the headspaceof the bottles were measured by gas chromatography. Theconcentrations of MSH and DMS were adjusted by usingnoninoculated standards, because some absorption of DMSinto the butyl rubber stopper was observed. Concentrations ofMSH and DMS were computed by using experimentally cal-culated distribution coefficients (liquid concentration/vaporconcentration) of 8.56 for MSH and 15.5 for DMS. The sulfideconcentration was measured after injection of 10 ml of a 2 wt%zinc acetate solution through the stoppers in order to trap thesulfide as zinc sulfide. Zinc sulfide suspensions were collectedby centrifugation. Aliquots were then withdrawn for spectro-photometric analysis of sulfide by the methylene blue method(7). Stoichiometries of DMS and nitrate used by strain SDN4were measured by a time course experiment. Periodically, gasand liquid samples were withdrawn with syringes for determi-nation of sulfur gases, ion content, sulfide, and optical density.The sampled volumes were replaced by the same volumes of02-free N2-CO2.
Determination of molar growth yield. One-liter cultureswere grown in the defined medium with MSH or DMS as thesubstrate. After growth, the bottles were gently purged withCO2 gas to remove dissolved sulfide in order to preventprecipitation of elemental sulfur. The cells were harvested bycentrifugation and resuspended in 10 mM sodium phosphatebuffer (pH 7.0) and then transferred to a small glass vial fordrying at 60°C to a constant weight. The values were correctedto that of cell-free controls.
Analytical techniques. MSH and DMS were measured in agas chromatograph (model GC6000; Carlo Erba Instruments,Milan, Italy) equipped with a flame ionization detector and aChromosil 330 column (Supelco Deutschland GmbH, BadHomburg, Federal Republic of Germany). The oven, injectionport, and detector temperatures were 65, 115, and 125°C,respectively. Nitrogen gas was used as the carrier gas, with aflow rate of 20 ml/min. Sulfate and nitrate were measured byion chromatography (Sykam GmbH, Gilching, Federal Repub-lic of Germany) with an A09 anion-exchange column (3).Ammonium was measured by the salicylate-hypochloritemethod (4).
Chemicals. All chemicals were of reagent-grade quality andpurchased from commercial suppliers. MSH and DMS werepurchased from E. Merck AG (Darmstadt, Federal Republicof Germany). Gelrite was obtained from the Karl Roth Co.
RESULTS
Dilution series and isolation of strain MTS5. The fermen-tative sludge of a laboratory-scale, methanogenic fermentorwas diluted serially in five tubes with defined minimal mediumsupplemented with 20 mM methanol, 2 mM MSH, and 10 mMsodium sulfate. A 250 ,uM concentration of 2-bromoethane-sulfonate was added to suppress growth of methanogens. After1 or 2 months of incubation at 55°C, growth and sulfideproduction were observed only in the highest-dilution tubes(10-4 and 10-5). Microscopic examinations showed slightlymotile and rod-shaped bacteria. In the next transfer, the MSHconcentration in the medium was increased to 5 mM instead ofthe methanol concentration being lowered to 1 mM. Theculture was then transferred to the methanol-free medium, inwhich the organism continued to grow with MSH as the soleelectron donor. MSH was completely consumed within 1 week.The isolate obtained from this procedure was designated strainMTS5.
Enrichments and isolations of strains TDS2 and SDN4.Enrichment cultures (50 ml) supplemented with 5 mM DMS,10 mM sodium sulfate, and 250 ,uM 2-bromoethanesulfonatewere inoculated with 2% fermentative sludge. Enrichmentcultures were also initiated with 5 mM DMS and 5 mM sodiumnitrate as an electron acceptor without 2-bromoethanesulfo-nate. After 3 weeks, the smell of DMS had disappeared andgrowth was observed. Microscopy revealed predominantlyslightly motile and rod-shaped bacteria. Cultures supple-mented with sulfate grew within 1 week while those withnitrate grew within 3 days. A sulfate-utilizing and a nitrate-utilizing isolate, designated strains TDS2 and SDN4, respec-tively, were used for further studies. The three isolates weredeposited in the Deutsche Sammlung von Mikroorganismenand registered under the numbers 7474 (strain MTS5), 7475(SDN4), and 7476 (TDS2).
General characteristics of strains MTS5, TDS2, and SDN4.The major characteristics of the three strains isolated (MTS5,TDS2, and SDN4) are summarized in Table 1. All strains werethermophilic, rod-shaped motile cells (Fig. 1) and requiredbiotin as a growth factor. The three strains formed central
VOL. 60, 1994
on April 13, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2452 TANIMOTO AND BAK
TABLE 1. Characteristics of the new isolatesa
Strain G+C content Length Width Electron acceptors Temp ('C) pH(mol%) (Im) (Im) Optimum Growth Optimum Growth
MTS5 56.6 1.8-4.5 0.5-0.8 Sulfate, sulfite, thiosulfite 55-60 55-60b 6.9 6.3-7.4TDS2 57.0 2.5-5.5 0.5-0.7 Sulfate, sulfite, thiosulfate 55-63 50-65 6.8 6.3-7.6SDN4 48.3 2.8-5.0 0.6-0.8 Sulfate, sulfite, thiosulfate, nitrate 55-60 55-63* 7.0 6.4-7.5
" All isolates were motile rods, required biotin for growth, and required 0 to 7 g of NaCl per liter.b SIOW growth at 50'C.
spores on rare occasions. Desulfoviridin was not detected inany of the strains (19). All the strains were sulfate reducers andalso had the ability to utilize sulfite and thiosulfate. Nitrate wasutilized only by strain SDN4. No growth was observed in saltconcentrations higher than 7 g/liter. Strain MTS5 had a narrow
growth temperature range, between 55 and 60°C, and an
optimum pH at 6.9. The growth temperature range of TDS2was wider, i.e., 50 to 65°C, and its optimum pH was 6.8; no
growth was observed below pH 6.3 or above pH 7.6. StrainSDN4 had a temperature range similar to that of strain MTS5but grew at temperatures up to 63°C. Growth pH ranged from6.4 to 7.5, with the optimum at pH 7.0. Determination of theDNA base composition of strains MTS5, TDS2, and SDN4yielded contents of 56.6, 57.0, and 48.3 mol% guanine pluscytosine, respectively. Strain SDN4, grown on DMS and/orMSH plus nitrate, exhibited the fastest growth of the threestrains, but the bacteria rapidly lost their activity and thecultures became yellowish. Strains TDS2 and MTS5 grew moreslowly (about 1 week) but maintained their activity for longerperiods.
Utilization of substrates. Electron donors and carbon
FIG. 1. Photomicrograph of isolate strain MTS5 grown with 3,4,5-trimethoxybenzoate. Bar = 10 ,.m.
sources for the three strains are summarized in Table 2.Complete oxidation of acetate with sulfate was performed byall strains. All the strains were able to grow by fermentation ofpyruvate in the absence of an external electron acceptor.Strains TDS2 and SDN4 were able to grow by fermentingfumarate. Strain MTS5 grew on MSH, strain TDS2 grew onDMS, and strain SDN4 grew on both MSH and DMS. StrainMTS5 was able to utilize MSH but not methanol. Various fattyacids were used by all the strains. Filter-sterilized glucose andfructose were utilized by strains MTS5 and TDS2. Autotrophicgrowth on hydrogen by all strains was confirmed by at leastthree transfers in acetate-free medium. Restricted methoxy-lated aromatic compounds were utilized by all the strains.Syringate and 3,4,5-trimethoxybenzoate were utilized by all thestrains, while sinapic acid was used only by strain MTS5.Ferulic acid and anisol were not utilized.
Stoichiometry of DMS oxidation with nitrate as the electronacceptor. The stoichiometry of DMS oxidation by nitrate
TABLE 2. Substrate utilization by strains MTS5, TDS2, and SDN4
Compound tested as electron Result for:donor (mM)a MTS5 TDS2 SDN4
MSH + - +DMS - + +H2 (autotrophically) + + +CO + acetate (1) + + +Formate (10) + acetate (1) + + +Acetate (10) + + +Propionate (10) + + +Butyrate (10) + + +Isobutyrate (5) + + +3-Methylbutyrate (5) - +Valerate (5) + + +Caproate (3) + + +Caprylate (1) + +Alanine (5) + +Lactate (10) + + +Pyruvate (10) + + +Fumarate (10) + + +Malate (10) + + +Succinate (5) + + +Methanol (5) - + +Ethanol (5) + + +Propanol (5) + + +Glucose (5) + +Fructose (5) + +Syringate (2) + + +3,4,5-Trimethoxybenzoate (2) + + +Sinapic acid (2) +
a The following substrates and concentrations (millimolar) were tested but notutilized: methane, trimethylamine (5), methylamine (5), benzoate (5), phenol(0.5), gallate (2), feruric acid (0.5), anisol (2), phloroglucinol (2), 4-hydroxyben-zoate (2), and thioglycolate (5). All tests were performed with sulfate as anelectron acceptor.
APPL. ENVIRON. MICROBIOL.
on April 13, 2019 by guest
http://aem.asm
.org/D
ownloaded from
ANAEROBIC DEGRADATION OF MSH AND DMS 2453
a~~~~~~~~~~~~~ir- tv e
C)~~~~~~~~~~~~~~~~~~~~~~~~~~J
CD~~~~~~~~~~~~~~~~~c
~0.2
0
Sulfide
0 50 100 150Time (hrs)
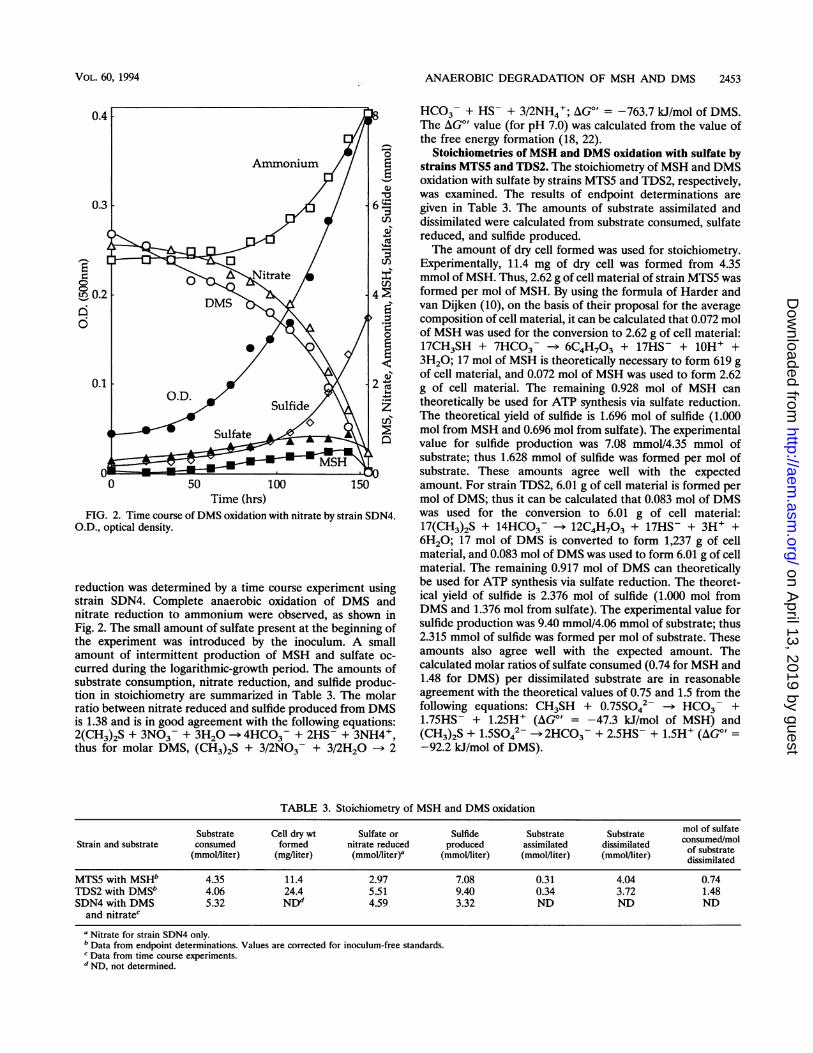
FIG. 2. Time course of DMS oxidation with nitrate by strain SDN4.O.D., optical density.
reduction was determined by a time course experiment usingstrain SDN4. Complete anaerobic oxidation of DMS andnitrate reduction to ammonium were observed, as shown inFig. 2. The small amount of sulfate present at the beginning ofthe experiment was introduced by the inoculum. A smallamount of intermittent production of MSH and sulfate oc-
curred during the logarithmic-growth period. The amounts ofsubstrate consumption, nitrate reduction, and sulfide produc-tion in stoichiometry are summarized in Table 3. The molarratio between nitrate reduced and sulfide produced from DMSis 1.38 and is in good agreement with the following equations:2(CH3)2S + 3NO3- + 3H20 -- 4HCO3- + 2HS- + 3NH4+,thus for molar DMS, (CH3)2S + 3/2NO3 + 3/2H20 -> 2
HCO3- + HS- + 3/2NH4+; AG"' = -763.7 kJ/mol of DMS.The AG"' value (for pH 7.0) was calculated from the value ofthe free energy formation (18, 22).
Stoichiometries of MSH and DMS oxidation with sulfate bystrains MTS5 and TDS2. The stoichiometry of MSH and DMSoxidation with sulfate by strains MTS5 and TDS2, respectively,was examined. The results of endpoint determinations are
given in Table 3. The amounts of substrate assimilated anddissimilated were calculated from substrate consumed, sulfatereduced, and sulfide produced.The amount of dry cell formed was used for stoichiometry.
Experimentally, 11.4 mg of dry cell was formed from 4.35mmol of MSH. Thus, 2.62 g of cell material of strain MTS5 was
formed per mol of MSH. By using the formula of Harder andvan Dijken (10), on the basis of their proposal for the average
composition of cell material, it can be calculated that 0.072 molof MSH was used for the conversion to 2.62 g of cell material:17CH3SH + 7HCO3-> 6C4H703 + 17HS- + 10H+ +
3H20; 17 mol of MSH is theoretically necessary to form 619 g
of cell material, and 0.072 mol of MSH was used to form 2.62g of cell material. The remaining 0.928 mol of MSH can
theoretically be used for ATP synthesis via sulfate reduction.The theoretical yield of sulfide is 1.696 mol of sulfide (1.000mol from MSH and 0.696 mol from sulfate). The experimentalvalue for sulfide production was 7.08 mmol/4.35 mmol ofsubstrate; thus 1.628 mmol of sulfide was formed per mol ofsubstrate. These amounts agree well with the expectedamount. For strain TDS2, 6.01 g of cell material is formed permol of DMS; thus it can be calculated that 0.083 mol of DMSwas used for the conversion to 6.01 g of cell material:17(CH3)2S + 14HCO3- -> 12C4H703 + 17HS- + 3H+ +6H20; 17 mol of DMS is converted to form 1,237 g of cellmaterial, and 0.083 mol of DMS was used to form 6.01 g of cellmaterial. The remaining 0.917 mol of DMS can theoreticallybe used for ATP synthesis via sulfate reduction. The theoret-ical yield of sulfide is 2.376 mol of sulfide (1.000 mol fromDMS and 1.376 mol from sulfate). The experimental value forsulfide production was 9.40 mmol/4.06 mmol of substrate; thus2.315 mmol of sulfide was formed per mol of substrate. Theseamounts also agree well with the expected amount. Thecalculated molar ratios of sulfate consumed (0.74 for MSH and1.48 for DMS) per dissimilated substrate are in reasonableagreement with the theoretical values of 0.75 and 1.5 from thefollowing equations: CH3SH + 0.75So42- -> HCO3 +1.75HS- + 1.25H+ (AG"' = -47.3 kJ/mol of MSH) and(CH3)2S + 1.5SO42 -* 2HCO3- + 2.5HS- + 1.5H+ (AG"' =
-92.2 kJ/mol of DMS).
TABLE 3. Stoichiometry of MSH and DMS oxidation
Substrate Cell dry wt Sulfate or Sulfide Substrate Substrate mol of sulfateStrain and substrate consumed formed nitrate reduced produced assimilated dissimilated ofsubstrate
(mmol/liter) (mg/liter) (mmol/liter)" (mmol/liter) (mmol/liter) (mmol/liter) dissimilated
MTS5 with MSHb 4.35 11.4 2.97 7.08 0.31 4.04 0.74TDS2 with DMSb 4.06 24.4 5.51 9.40 0.34 3.72 1.48SDN4 with DMS 5.32 NDd 4.59 3.32 ND ND NDand nitrateC
a Nitrate for strain SDN4 only.b Data from endpoint determinations. Values are corrected for inoculum-free standards.c Data from time course experiments.d ND, not determined.
VOL. 60, 1994
on April 13, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2454 TANIMOTO AND BAK
DISCUSSION
MSH and DMS utilization by strains MTS5, TDS2, andSDN4. The newly isolated strains MTS5, TDS2, and SDN4 arethe first reported sulfate-reducing bacteria which have theability to break down MSH and DMS. No information aboutthe utilization of MSH and DMS by the genus Desulfotomacu-lum was previously available. Two known strains of the genusDesulfotomaculum, i.e., D. orientis DSM 765 and D. thermo-benzoicum DSM 6193, were tested for the inability to grow withMSH or DMS. Neither MSH nor DMS supported the growthof these species. Therefore, the ability to utilize MSH andDMS was specific for the newly isolated strains rather than forall species of the genus Desulfotomaculum.
Physiology and taxonomic status of strains. The threestrains were thought to belong to the genus Desulfotomaculum,since all spore-forming sulfate reducers belong to this genus(23). However, the isolates made central internal spores a fewtimes, and the conditions under which sporulation occurs havenot yet been determined. Some new characteristics for thegenus Desulfotomaculum were presented by the new isolates,for example, glucose utilization and the GC content of -56 to57% in strains MTS5 and TDS2. The difference between theGC contents of strains MTS5 and TDS2 and those reported forthe other Desulfotomaculum species was more than 10%,which means the strains can be considered as another genus.The three strains have a narrow growth temperature range,especially strain MTS5, and we do not know why. However, allthe strains were isolated from fermentor sludge, from afermentor that operated at a constant temperature of -53 to55°C (15). Thus, natural selection of microorganisms mighthave occurred in the source of the isolates.The three strains were able to grow using 3,4,5-trimethoxy-
benzoate, which was considered one precursor of the methyl-ated sulfur compounds, but we could not find any evidence ofthe formation of methylated sulfur compounds by the newisolates. Thus, the methoxy group seemed to be directlyconverted to CO2 in the presence of sulfate.
D. thermobenzoicum has been reported previously as anitrate reducer (21), but no details were given. In this study,strain SDN4 was found to be a utilizer of nitrate as an electronacceptor, and the product was ammonium. Small amounts ofsulfate and MSH were observed during the time courseexperiment with strain SDN4 with DMS and nitrate. Thereason sulfate and MSH concentrations increased during DMSdegradation by nitrate is not clear, especially the increase insulfate. However, the reason for MSH production during DMSdegradation is probably that DMS catabolism is proceeding viaMSH, which was reported for methylotrophic methanogenduring DMS degradation (9, 12).The enzymatic pathway in strains TDS2 and SDN4 would be
different. Both organisms degrade DMS, because strain TDS2was not able to grow on MSH. Also, DMS degradation instrain TDS2 probably did not proceed via MSH.
Ecological aspects. Methylated sulfur compounds, mainlyDMS, are thought to play an important role in the global sulfurcycle (6). DMS is the major organic sulfur compound which isdischarged from the biological environment. The methylatedsulfur compound emissions from the natural terrestrial envi-ronments are thought to be a very small amount comparedwith the coastal areas. The emissions from vegetation, incontrast, are much greater than those from the land (1). Theemission from vegetation is primarily from crops and is alsoproduced during microbiological degradation of plant mate-rial. However, the microbiological degradation of DMS andrelated sulfur compounds has not been clarified (8), especially
in the terrestrial microbiological ecosystem, because no DMS-degrading microorganisms (including methanogenic or sulfatereducing, etc.) have been isolated. Furthermore, no majorresearch on the measurement of anaerobic microbial activity inDMS or MSH sinks has been conducted. Several methano-genic bacteria have been isolated from anaerobic marinesediments; however, no sulfate reducers have been isolated inspite of many observations regarding DMS sinks.For practical use, the main tool is peat biofilters, which
oxidize and remove methylated sulfur compounds from the air,using aerobic bacteria. The main flux of artificial methylatedsulfur compounds is undoubtedly due to the pulping industries.However, anaerobic treatment of pulp waste has not become amajor method yet; the main treatments are burning or aerobictreatment. Thus, it is still unclear whether mesophilic bacteriaor methanogenic bacteria exist. We also tried to isolate metha-nogenic bacteria by the same method, but all efforts failed.Hence, it appears that mainly sulfate reducers are responsiblefor the degradation of MSH and DMS in a thermophilicfreshwater environment.The isolates described here will give us a new consideration
for the sulfur cycle, even though the isolates cannot grow in anatural environment, because they are the first reported sulfatereducers which can be grown with methylated sulfur compounds.
ACKNOWLEDGMENTS
Thanks are extended to R. K. Thauer and his laboratory at theUniversity of Marburg for the loan of the gas chromatograph used forthe methylated sulfur gas analysis and also for the use of manyexperimental materials. We thank F. Widdel for critical comments onthe manuscript. Anonymous reviewers are thanked for their criticalcomments and suggestions which helped improve the manuscript. Y.Tanimoto thanks R. Conrad and the members of his laboratory fortheir great hospitality and encouragement during this study.
REFERENCES1. Aneja, V. P., and W. J. Cooper. 1989. Biogenic sulfur emissions, p.
2-13. In E. S. Salzman and W. J. Cooper (ed.), Biogenic sulfur inthe environment. American Chemical Society, Washington, D.C.
2. Bak, F., K. Finster, and F. Rothfuss. 1992. Formation of dimethylsulfide and methanthiol from methoxylated aromatic compoundsand inorganic sulfide by newly isolated anaerobic bacteria. Arch.Microbiol. 157:529-534.
3. Bak, F., G. Scheff, and K. H. Jansen. 1991. A rapid and sensitiveion chromatographic technique for the determination of sulfateand sulfate reduction rates in freshwater lake sediments. FEMSMicrobiol. Ecol. 85:23-30.
4. Bower, C. E., and T. Holm-Hansen. 1980. A salicylate-hypochloritemethod for determining ammonia in sea water. Can. J. Fish.Aquat. Sci. 37:794-798.
5. Cashion, P., M. A. Holder-Franklin, J. McCully, and M. Franklin.1977. A rapid method for the base ratio determination of bacterialDNA. Anal. Biochem. 81:461-466.
6. Charlson, R. J., J. E. Lovelock, M. 0. Andreae, and S. G. Warren.1987. Oceanic phytoplankton, atmospheric sulphur, cloud albedoand climate. Nature (London) 326:655-661.
7. Clinde, J. D. 1969. Spectrophotometric determination of hydrogensulfide in natural waters. Limnol. Oceanogr. 14:454-458.
8. De Zwart, J. M. M., and J. G. Kuenen. C,-cycle of sulfurcompounds. Biodegradation, in press.
9. Finster, K., Y. Tanimoto, and F. Bak. 1992. Fermentation ofmethanethiol and dimethylsulfide by a newly isolated methano-genic bacterium. Arch. Microbiol. 157:425-430.
10. Harder, W., and J. P. van Dijken. 1976. Theoretical considerationson the relation between energy production and growth of meth-ane-utilizing bacteria, p. 403-428. In Symposium on microbialproduction and utilization of atmospheric trace gases. Goltze,Gottingen, Germany.
11. Hynninen, P. 1971. On the reactions of methylmercaptan and its
APPL. ENVIRON. MICROBIOL.
on April 13, 2019 by guest
http://aem.asm
.org/D
ownloaded from

ANAEROBIC DEGRADATION OF MSH AND DMS 2455
oxidation products in air- or steam-stripping of sulfate mill con-densates. Paper och trae. specialnummer 4a:159-169.
12. Kiene, R. P., R. S. Oremland, A. Catena, L. M. Miller, and D. G.Capone. 1976. Metabolism of reduced methylated sulfur com-pounds in anaerobic sediments and by a pure culture of anestuarine methanogen. Appl. Environ. Microbiol. 52:1037-1045.
13. Kiene, R. P., and P. T. Visscher. 1987. Production and fate ofmethylated sulfur compounds from methionine and dimethylsul-foniopropionate in anoxic salt marsh sediments. Appl. Environ.Microbiol. 53:2426-2434.
14. Mesbah, M., U. Premachandran, and W. Whitman. 1989. Precisemeasurement of the G+C content of deoxyribonucleic acid byhigh-performance liquid chromatography. Int. J. Syst. Bacteriol.39:159-167.
15. Minami, K., K. Okamura, S. Ogawa, and T. Naritomi. 1991.Continuous anaerobic treatment of wastewater from a kraftpulpmill. J. Ferment. Bioeng. 71:270-274.
16. Ni, S., and D. R. Boone. 1991. Isolation and characterization of adimethyl sulfide-degrading methanogen, Methanolobus siciliaeH1350, from an oil well, characterization of M. siciliae T4/MT, andemendation of M. siciliae. Int. J. Syst. Bacteriol. 41:410-416.
17. Oremland, R. S., R. P. Kiene, I. Mathrani, M. J. Whiticar, andD. R. Boone. 1989. Description of an estuarine methylotrophicmethanogen which grows on dimethyl sulfide. Appl. Environ.Microbiol. 55:994-10)02.
18. Pedley, J. B., and J. Rylance. 1977. Sussex-N.P.L. computeranalyzed thermochemical data: organic and organometallic com-pounds. University of Sussex, Falmer, Brighton, Sussex, UnitedKingdom.
19. Postgate, J. R. 1979. The sulfate reducing bacteria. CambridgeUniversity Press, New York.
20. Tamaoka, J., and K. Komagata. 1984. Determination of DNAbase composition by reversed-phase high-performance liquidchromatography. FEMS Microbiol. Lett. 25:125-128.
21. Tasaki, M., Y. Komagata, K. Nakamura, and E. Mikami. 1991.Isolation and characterization of a thermophilic benzoate-degrad-ing, sulfate-reducing bacterium, Desulfotomaculuim therrnobenzoi-cumn sp. nov. Arch. Microbiol. 155:348-352.
22. Thauer, R. K., K. Jungermann, and R. K. Decker. 1977. Energyconservation in chemotrophic anaerobic bacteria. Bacteriol. Rev.41:100-180.
23. Widdel, F. 1992. The genus Desulfotomaculum, p. 1792-1799. In A.Balows, H. G. Truper, M. Dworkin, W. Harder, and K. H.Schleifer (ed.), The prokaryotes, 2nd ed., vol. 2. Springer-Verlag,New York.
24. Widdel, F., and F. Bak. 1992. Gram-negative mesophilic sulfate-reducing bacteria, p. 3352-3378. In A. Balows, H. G. Triuper, M.Dworkin, W. Harder, and K. H. Schleifer (ed.), The prokaryotes,2nd ed., vol. 4. Springer-Verlag, New York.
VOL. 6(1, 1994
on April 13, 2019 by guest
http://aem.asm
.org/D
ownloaded from





























