Embed Size (px)
Citation preview

YEAST VOL. 8: 559-567 (1 992)
Analysis of the MSS.51 Region on Chromosome XI1 of Saccharomyces cerevisiae MICHEL SIMONt, FLAVIO DELLA SETA*, FREDERIC SOR AND GERARD FAYE
lnstitut Curie-Biologie, Brit. 110,91405 Orsay, France *Laboratoire d'Enzymologie et de Gknie Gknktique, Faculti des Sciences, BP 239,54506 Vandoeuvre-les-Nancy, France
Received 25 September 1991; accepted 6 February 1992
We have localized gene MSS5I on chromosome XI1 of Saccharomyces cerevisiae between the RDNl and CDC42 loci. 'Head to head' with MSSSl is another gene, QRI5, the function of which is unkown. However, the proximity of these genes, the structure of the intergenic region and the presence of an ABFl binding site right in the middle of this region suggest that the MSS5l and QR15 expressions are submitted to a common regulatory process.
KEY WORDS - MSS51; QRI5; chromosome XII: ABFl binding site; Saccharomyces cerevisiae.
INTRODUCTION primary transcripts must be cleaved, spliced and translated (because some intron-encoded proteins The Yeast (cYtochrome b, and coxz (cytochrome Oxidase subunit I) genes are are necessary for splicing) before becoming mature messenger RNAs. In addition to the translation interrupted by introns and belong to multigenic
transcription units (Christianson et al., 1983; Simon mitochondrial mRNAs require mRNA- specific translation activators (Costanzo and and '9g4; Grivell, 1989)* The 1990). Almost all of the proteins involved in these
tcorresponding author. processes are encoded ih the nucleus. We have
Table 1. Yeast strains
Strain Genotype Source/Reference
D273-10B/A umet6 A. Tzagoloff E4-2 18 amet6 mss5l-1 A. Tzagoloff E4-218- 1 B aleu2 his4 mss51-1 Faye & Simon (1983) 5 1 -R2G aleu2 his4 GLY+ revertant This work 4870-6B GRFl8 aleu2-112 leu2-3 his3 A-I canR T. Donahue AH22 aleu2-112 leu2-3 his4 canR A Hinnen DGlOl GRF18 X AH22 This work 1758 Klapholz and Esposito (1982) GF268-16A This work 1762 Klapholz and Esposito (1982) GF272-4D ahis3 mssSl-1 This work lB-TR-ZD/Fl 1 aleu2 trpl mssSl-1 rho" This work FDC33- 13D ahis3 sdc25::HIS3 cdc25-5 uru3 E. Boy-Marcotte C82-1857 ailv5-1 ura4 asp5 met1 argl gal2 YGSC DJDM2-7C acdc42-1 ura3 his4 leu2 gal2 RDNI::LEU2 D. Johnson MH41-7B aade2 his1 H. Fukuhara
agal10-4 suc ma1 his5 trpl ura4 met8 ade5.7 leu2 lysl is01 ilv3 canl D. Hawthorne
aspoll ura3 canl cyh2 ade2 his7 hom3 tyrl aspoll cyh2 his7 or 4 leu2 hom3 ura3 tyrl mss51-1 aspoll ura3 ade6 arg4 aro7 asp5 met14 lys2petl7 trpl
0749-503X/92/070559-09 $09.50 0 1992 by John Wiley & Sons Ltd

560 M. SIMON ETAL. - 1 Kb
Bg Ba
:::::I, 4D -* OR15 MSS51
Figure 1 . Restriction endonuclease cleavage map of the MSS52 region. The94 kb long HindIII-Hind111 DNA fragment contain- ing the MSS-52 gene is drawn. The positions of some restriction sites are indicated. R, EcoRI; Bg, BgII; Ba, BumHI; S , SsrI; H, HindIII. The portion of the HindIII-Hind111 fragment which was sequenced is shown by a shaded bar. Black arrows represent open reading frames. QRI means ‘quasi-renownless information’ and MSS ‘mitochondria1 synthesis system’.
recently shown that one of them, the MSS.51 gene product, is needed for the translation of the COX1 mRNA (Decoster et al., 1990).
In order to obtain more information about the function of MSS.51, we have undertaken two studies. First, we searched for extragenic sup- pressors of mss.51 mutants. Second, we studied a newly identified gene (Faye and Simon, 1983) named QRZ.5, transcribed in the opposite direction. The ATG start codons of the MSS.51 and QRZ.5 genes are separated by 310 bp and their divergent promoters seem to originate from inverted-repeat sequences with an unique axis of symmetry which is included in a putative ABFl binding site (Della Seta et al., 1990b). Since it sometimes appears that genes arranged ‘head to head’ have related function (Beck and Warren, 1988), we have tried to identify the role of QRZ.5. Finally, the chromosomal assign- ment and genetic mapping of the MSS.51 gene were determined.
Our results, as a whole, contribute to extend our knowledge of the genetic information contained in chromosome XII.
MATERIALS AND METHODS
Yeast strains and media A list of the yeast strains used is given in Table 1.
All genetic procedures and media were as described by Sherman et al. (1983). Yeast transformation, DNA preparation and DNA manipulation were as described (Faye and Simon, 1983; Maniatis et al., 1982).
Cloning and sequencing Total DNA ofstrains D273-10B/A or E4-218 was
digested by HindIII restriction enzyme; DNA frag- ments were ligated at the HindIII sites of 1728 or
hL47 (Murray et al., 1977; Loenen and Brammar, 1980). h plaques harboring HindIII-Hind111 frag- ment containing the MSS.51 gene were screened using plasmid pB51-Bal as a probe (pB51-Bal is pBR322 carrying the HindIII-BamHI-BamHI-Ban fragment of YEp51) (Faye and Simon, 1983). The D273-10B/A or E4-218 HindIII-Hind111 fragments were digested with appropriate restriction enzymes and the fragments thereof were cloned in pBR328 or pBR322. DNA sequencing was performed as described by Maxam and Gilbert (1980).
Northern experiment and S1 mapping
Total RNA was extracted from strain D273-10B/ A and poly(A)+ mRNA was purified on an oligo(dT) cellulose column. RNA transfer to nitrocellulose fil- ter and RNA-DNA hybridization were performed according to the Southern method as modified by Thomas (1980).
S1 mapping was carried out as described earlier (Faye and Simon, 1983). pBR328 harboring the 1660 bp EcoRI-EcoRI DNA fragment (from strain D273-10B/A) was cleaved by AvaII. The 5’ ends were labelled with T4 polynucleotide kinase and [y3’P]ATP, then the labelled fragments were digested with EcoRI restriction endonuclease. The DNA strands of the A vaII-EcoRI fragment, overlapping the 5’-end of the QRZ5 gene, were separated on a polyacrylamide gel. The labelled strand was used to map the 5’-end of the QRZ5 mRNA. The same plas- mid was cleaved with HinfI endonuclease; the 3‘- ends of DNA fragments were 3’-end labelled with Klenow DNA polymerase and [a3*P]-dATP. The strands of the 329 bp DNA fragment covering the 3‘-end of gene QRZ.5 were separated and used for S1 mapping (cf. Figure 4).
Gene disruption
The SalI-EcoRV segment of pBR328 was deleted in order to eliminate its SphI site, before cloning the 1660 bp EcoRI-EcoRI fragment harboring the QRZ.5 gene. The plasmid obtained was digested with Sphl and the ends were made blunt using S 1 nuclease and Klenow DNA polymerase before inserting the 2.4 kb HpaI-HpaI fragment containing the LeU2’ gene. The plasmid obtained was digested with EcoRI and the fragment bearing the LEU2 gene was inserted in place of one of the two chromosomal copies of the QRZ5 gene of strain DGlOl by the one-step disruption method described by Rothstein (1985).

ANALYSIS OF MSSSI REGION ON CHROMOSOME XI1 OF S. CEREVISIAE 56 1
Gel retardation experiment
The MSS.51 and RPC40 probes (Della Seta et al., 1990a) were 3'-end labelled with Klenow polym- erase and purified by gel electrophoresis. For com- petition studies, a 21 bp oligonucleotide including the ABFl DNA binding site upstream from the sequence of RPC40 promoter (position -1 32 to - 1 18) (Della Seta et al., 1990a) (cf. Figure 5) and two vari- ants of this sequence, were synthesized chemically and treated as previously described (Della Seta et al., 1990a).
ABFl factor was isolated by DNA affinity col- umn and the analysis of protein-DNA complexes was carried out as described (Della Seta et al., 1990a). For the titration experiments, 14 ng/20 p1 of pBR322 DNA were added and used as non-specific competitor. For the competition experiments, the total DNA concentration was kept constant (60 ng or 300 ng/20 pl) by adding appropriate quantities of pBR322 DNA.
RESULTS AND DISCUSSION
An extragenic suppressor of the mss51-1 mutant
Search for suppressors Protein MSS51 seems to interact with the small subunit of the mitoribosome to promote the translation of the COX1 mRNA. However, we do not know its exact role in this pro- cess. It should be possible, starting from mutants in the MSS.51 gene, to isolate extragenic suppressors which might bring new clues as to the function of MSS.51. Cells of strain E4-218- 1 B,harboring allele mss.51-1 (Decoster et al., 1990) were UV-irradiated, then plated on glycerol as a carbon source. After 2 weeks' incubation, only one revertant, named 51- R2G, was obtained. Using genetic tests we con- cluded that this suppressor is not linked to mss51-1 (data not shown).
Nature of the mss51-1 mutation Extragenic sup- pressors may be informational (e.g. nonsense sup- pressors). Before isolating the gene affected in revertant 5 1-R2G, we wanted to know the nature of the mss.51-1 mutation. The MSS51 gene is included in a 9.8 kb long HindIII-Hind111 DNA fragment (Figure 1). This fragment was isolated from the two isonuclear strains D273- 10B/A and E4-2 18 harbor- ing the wild-type MSS.51 gene and the mss.51-1 allele, respectively. After subcloning, we determined the sequence of the HincII-EcoRV DNA fragments containing the MSS51 region. Compared to the wild-type MSS.51 gene, the mss.51-1 mutant presents
a T to C transition in codon number 21, changing a serine into a proline, and a G to A transition in codon number 64, which generates a TGA stop codon (Figure 2). The comparison of the sequence of the MSS.51 gene of strain D273-10B/A with that obtained from strain AB320 (Faye and Simon, 1983) revealed 14 differences. Twelve are located in the MSS.51 coding phase, with five of them leading to amino-acid substitutions. This high degree of polymorphism between strains D273- lOB/A and AB320 is surprising.
The .51-R2G revertant gains an opal suppressor Since the mss.51-1 mutation consists of two alterations, one of which gives rise to a TGA codon, revertant 51-R2G might be an opal suppressor. To test this hypothesis, 51-R2G was crossed with strain 4870- 6B, which contains several amber, ochre and opal mutations in different genes. Diploids were isolated and forced to sporulate. As a control, strain 4870- 6B wascrossed with strain E4-218-18. The resultsof tetrad analyses are shown in Table 2. The presence and the proportion of 3'11- and 4'10- tetrads observed with the gall0 and ade5-7 alleles clearly demonstrate that 51-R2G harbors a TGA sup- pressor. On the other hand, since the activity of this suppressor restores the respiratory competence of strain 51-R2G, the mss-51-1 alteration in codon 21 (serine into proline) is not damaging for the MSS5 1 protein.
In conclusion, the extragenic suppressor obtained is not of the type we desired. The analysis of other mutants in the MSS.51 gene might allow us to locate the conjectured suppressors.
The QRI5 gene
Sequence In sequencing the 1660 bp long EcoRI- EcoRI DNA fragment ending in gene MSS.51, we completed the sequence of the QRZ.5 gene, which is transcribed divergently from gene MSS51 (Figure 2). QRZ5 encodes a 11 1 amino acid long protein. Its calculated molecular weight is 12,774 daltons. It is very basic, especially in its C-terminal end (Figure 3) (20 out of the last amino acids are arginine or lysine), a particularity suggesting that it might inter- act with nucleic acids. No protein sequence signifi- cantly homologous with QRI5 was found in data banks.
Sequencing also revealed the C-terminal 106 codons of a putative gene which stretches over the other end of the 1660 bp long EcoRI-EcoRI DNA fragment.

- 120 E F L A T F H K P P L L Q Q F I N N I H E N I H K E P C T I L S Y C H V L Y L A G A A T T C T T G G C T A C G T T C C A T A A G C U L C C G C T A C T A C A G C G
240 L F A G G K L I R S N L Y R R L G L F P N F E K L S Q K E L V K K G T N F F T F C T T T T C G C C G G C G C G A m G A A ~ T A C
360 S D L G P T E E T R L K W E Y K K N Y E L A T R R N A G C G A T C T G G G C C C C A ~ G A A ~ C A ~ ~ T ~ T A ~ W U \ G A A ~ A ~ ~ C A C C A ~ C ~ ~ C C G A A G C A ~ G ~ ~ T T A G C G T C G C A G A A G
480 G C A T P T T T G A T T G G A A C T G T T G C A ~ T T ~ ~ ~ T C G m G C ~ ~ A R ~ ~ G C ~ G ~ T A ~ A C G T A C ~ T ~ G A A G A A ~ T G ~ C A A C A
HinfI a. 600 A A ~ ~ T T A C T A G A A W C A C P I C G G ~ T G C T G C T ~ ~ ~ T ~ A T T A ~ G C G A ~ ~ ~ A ~ ~ A A A G A A G A G ~ T T A G C A T A G T A T A A C G T A T A A
720 ~ G T C T A A A T A C A R I \ C I \ T I \ A T A T A T A ~ T ~ T A A T A T ~ T A T G A A G A A T A ~ ~ C A ~ ~ ~ C A ~ T G A G A ~ C ~ A C A ~ C C T A T T A T C T A C C
R G
HinfI 840
Q S L K R R E A K E R K R R K R L K H K K M K K K R K R M V S D L Q M V I N S G
sphr 960 G m A A T G G C T C P I C T G A C T G T G C C P I G T G A T C C T T G T T G C C G T A G C C G T ~ T C P I T C A ~ ~ T A C T ~ T G C C ~ C T T G C A T ~ ~ T T G T A G ~ ~ A A ~ ~ m ~ f f i T N L P E S V T G T I R T A T A T M M M M T S L M G E Q M P P M T T N I A T Q F T
CTGAW\TAGCTTTCTPC~C~;CCTTCCCTC~CTTC~TCCC
AvaII 1080 T C T G ~ A A A ~ A ~ C ~ C T T ~ ~ ~ ~ C T T A A C ~ C C G ~ C T A G T A A ~ C C A G C C P I C C ~ ~ ~ T G C C C ~ C ~ ~ ~ ~ A T C ~ ~ ~ ~ G ~ T G T ~ C G T C
T~;GCTGWULCTCAACCGCCCGCTCTT~TATTATAI AJp$kT~GCGE*GAAGJWGAA
R N F Y S G C S P K K V V T R T I G L W G P R L A R G L M
1200 -
*tt
fndfng site 1320
TC-GGAG~~GAGACAULATAGTAGATA~~cTACTAAGGGCGGGCCTITGGTn;TAGOACPI-ATAAGC~WU\~~TA~TA~GCWU\GGTGGA HinCII - *** ** C
~ L Y A P S G A T Q L Y F H L L R K ! P H N R L V V 1440
C T T A A A G T A G m U ; C A A A C T T G T A C T P C C A G A A C T A T C A C C T C
1560 S H Q T R R H L M G F V R N A L G L D P P P S P E D P T P . E N R F H P W D Q S P T C T C A C ~ C P I C ~ G T C ~ T ~ G T T A ~ C G C C C ~ ~ ~ C ~ C ~ C A W U \ ~ ~ ~ A ~ C C G G A G A A ~ ~ C A ~ ~ ~ G ~ T C C C
@ 1680 S V D L R E R R A K I R T L A H C P V T G K D I N Y T C P L S G I P T H H S R E T C A G n ; W L C ~ G T W C G C ~ G A ~ A G A A C ~ A ~ C A ~ C ~ C C ~ G A T A ~ A R C T A C A ~ ~ ~ C ~ ~ ~ C A C G A G A G
1800 A W E M D K A Y H D S K K Y E I L K K V N I Y E H D L R S G R P F P E F D F P Q Q T G A ~ T ~ ~ A T R T C A C G A T P C T A A R A A A T
1920 Q Q G Y D K A V N L T N W D L F F Y T R S F Y S M D T E F Q L A A V T K M L S Y C A G C A A G G C T A C G A ~ G G C C G T T A A C ~ A A ~ ~ ~ ~ ~ ~ C A ~ G A T ~ T T ~ A ~ C A ~ T A ~ G ~ ~ ~ A G C C ~ ~ ~ G A T G C T A A G C T A T
I 2040 P I T I G S L L H K F S P Y S L N P K G P M T L E G L K S L A A L R Y T L Y P L C C A A T T A C G A T T G G G m m G C T G C A C A A P l m T C C C C G T A T T G
2160 E N R S L P T T T K N R A M R I F I L G A R A E A Q L P G H V W K Q L Q F L F P B G A A T A G A T C m A C C A A C C P I C A U L A C T A R G A A C C G ~ A T G A G A A ~ C A T C C T A ~ C ~ G A A ~ ~ C ~ C P I C ~ C P I G C ~ C A G ~ C ~ C C C P I
2280 E Q S F E I H F I G P E C L Y K R D K Q E Y V K S T T P V V Q R V D E T L K F I W I G C A W L G T T T C ~ T A C G G A C C C G A A T ~ ~ T A T ~ T A A A C A ~ G T A C G ~ G ~ T A C T A C G C C ~ A G T A ~ ~ ~ A G A C G A G A C T ~ ~ C P I ~
0
A -I
EaoRl
C A
G
C
T C S 2400
Y R T N F F E V F H E A Q D F F P Y D P Y M D V F F T F H P G Y A S P E S H G S T A T C G T A C A A A C m r P C G A A C T A T T C U L T W L G G C T C A A W \ r r
T G A X 2520
W M G E T M K A L L E T K C A I F T T G F N K K D L T D D I N L V K S K Y G K E T G G A T G G G T W V \ A C T A ~ ~ ~ A ~ ~ ~ G C T A m ~ C A U L A C ~ ~ T ~ ~ G A C f f A R C G G A C ~ C A T A A A ~ ~ ~ T A C ~ A A G G A A
T X 2640
M D V L M E P V R N V F G S T K W E L N D M N P Q E V Y Q F N M Y I A G F R G K A T G G A T G T ~ A A T G G A G C C C G ~ ~ C G T A ~ ~ G T A C G A A G ~ ~ A A A ~ C A ~ T C - G A A G ~ A ~ ~ ~ ~ T A T A ~ C ~ ~ ~ ~ ~
A

ANALYSIS OF MSSSJ REGION ON CHROMOSOME XI1 OF S. CEREVISZAE
Table 2. Tetrad analysis of sporulation products of diploids 5 1-R2G x 4870-6B* and E4-2 18- 1 B x 4870-6B
563
Number of tetrads 0+/4- l f /3 - 2+/2- 3+/1- 4+/0-
Cross: 51-R2G X4870-68 UGA gall0
mss51 ade.5- 7 leu2
UAA ilv3 lysl
UAG trpl Mating type Cross: E4-218-18 X 4870-68 UGA gall0
mssS1 ade5- 7 leu2
UAA ilv3
0 0 0 0 0 0 7 16 0 0 0 3 0 8 0 0
0 1 0 1 0 4
31 0 1 3
5 21 8 17 3 19 6 0
28 1 26 0 21 0 27 2
30 0 30 0 27 0 0 0
26 1
3 4 7 0 0 0 0 0
0 0 0 0 0
*Strain 4870-6B harbors opal (UGA), ochre (UAA) and amber (UAG) mutations. + , wild-type phenotype; -, mutated phenotype.
Northern blotting and SI mapping A Northern experiment was carried out to determine the size of the QRI5 mRNA. Purified poly(A)+ mRNA was hybridized with the 32P-labelled HinfI-AvaII DNA fragment (161 bp long) (Figure 2). A 560 bp long mRNA was detected (Figure 4c).
The ends of the QRZ.5 mRNA were delimited by S1 mapping experiments as shown in Figure 4. The AvaII-EcoRI and HinfI -HinfI probes were pro- tected on 104-105 and 252 base long segments, respectively. This suggests that the QRI5 mRNA possesses a poly(A) tail of about 40 residues (560 - (252 + 16 1 + 105)).
Both results demonstrate that QRZ5 is an active gene.
Gene disruption The QRI5 gene was disrupted in diploid strain DGlOl (cf. Materials and Methods)
M L G R A L R P G W L G I T R T V V K K P S C G S Y F N R T F Q T A I N T T M P P M Q E G M L S T M M M M T A T A T R I T G T V S E P L N G S N I V M Q L D S V M R K R K K K M K K H K L R K R R K R E K A E R R K L S Q G R
Figure 3. The QRl5 protein sequence. Contrary to Figure 2, the QRl5 protein sequence is shown in the usual orientation, i.e. printed from its N- to its C-terminal end.
and after sporulation, tetrads were analysed. The four spores of each tetrad were viable and respira- tory competent, meaning that QRI5 is not involved in mitochondria biogenesis. Thus, the identification of its function cannot shed any light on that of MSS51.
ABFI binding site The 5' end of the QRZ5 mRNA and that of the major transcript of the MSS5I gene are separated by 166 nucleotides; inverted repeat
Figure 2. Sequence of the MSSSI region. The sequence presented from nucleotide number one to the AvuII site was obtained with the DNA of strain D273-10B/A, whereas the rest of the sequence is from the DNA of strain AB320 (Faye and Simon, 1983). The HincII-EcoRV fragment was also sequenced using the DNA of the former strain. Fourteen nucleotide changes, which were found when the two sequences were compared, are indicated by the letters G, A, T or C printed under the nucleotide sequence, and the flve amino-acid substitutions by a letter drawn above the MSS5l amino acid sequence. The letters indicating the two mutations present in the MSSSl gene of strain E4-218 are enclosed within a circle. The triangle means that this T residue is deleted in D273-10B/A. Horizontal arrows show the direction of transcription of the genes, asterisks the 5'-ends of the mRNA of the MSSSl and QRlS genes and solid circles the 3'-ends of the same genes. Inverted repeat sequences between QRIS and MSSSl are shown by lines drawn above them and their symmetry axis by two short vertical lines, between nucleotides 1171 and 1172. The ABFI binding site is located in this region. Some restriction sites relevant to our studies are mentioned.

564 M. SIMON ET AL.
scale I A 200 b
R 4 R
A \b .r,
B 252 161 104 -1 05
R 4 I R I
H H H - a
1 2 3 4
1631-
51 7 , 506-
396-
344-
298-
221- 2 20-
154-
1 2 3 4
C A
Hind I II
-329 -252
564-
t105
ANALYSIS OF MSS5il REGION ON CHROMOSOME XI1 OF S. CEREVISIAE 565
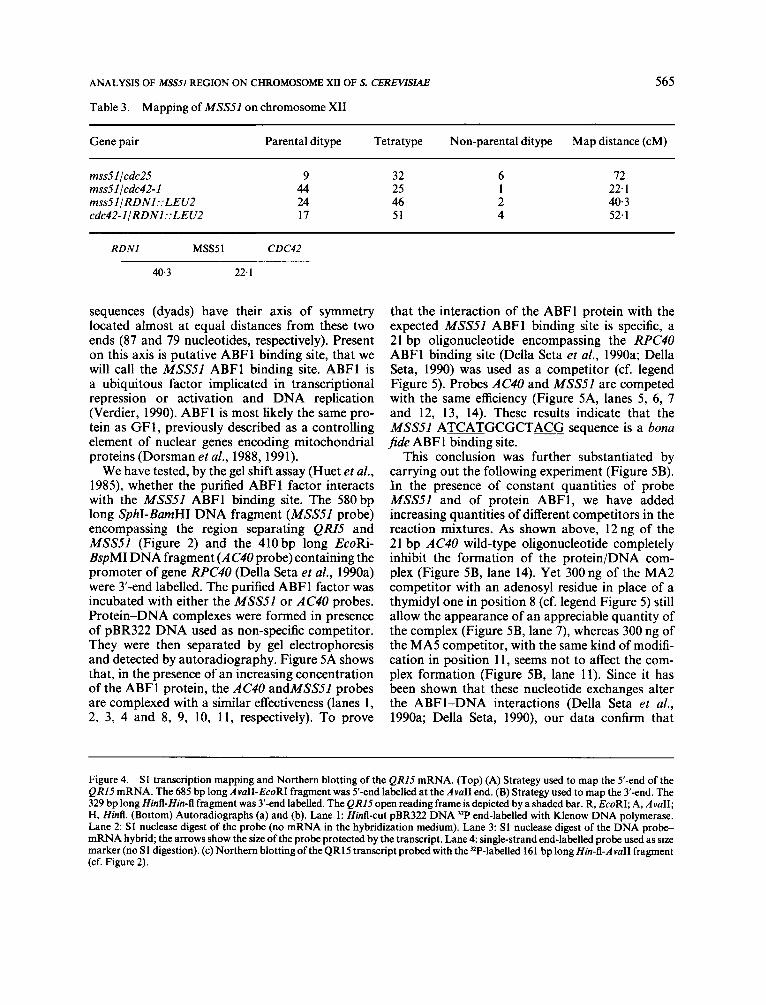
Table 3. Mapping of MSS51 on chromosome XI1
Gene pair Parental ditype Tetratype Non-parental ditype Map distance (cM)
m s d l lcdc25 mss51 lcdc42-1 mss51/RDNl::LEU2 cdc42-1IRDNl::LEU2
9 32 44 25 24 46 17 51
72 22.1 40.3 52.1
RDNl MSS51 CDC42
40.3 22.1
sequences (dyads) have their axis of symmetry located almost at equal distances from these two ends (87 and 79 nucleotides, respectively). Present on this axis is putative ABFl binding site, that we will call the MSS.51 ABFl binding site. ABFl is a ubiquitous factor implicated in transcriptional repression or activation and DNA replication (Verdier, 1990). ABFl is most likely the same pro- tein as GF1, previously described as a controlling element of nuclear genes encoding mitochondria1 proteins (Dorsman et al., 1988, 1991).
We have tested, by the gel shift assay (Huet et al., 1985), whether the purified ABFl factor interacts with the MSS.51 ABFl binding site. The 580 bp long SphI-BamHI DNA fragment (MSS.51 probe) encompassing the region separating QRZ5 and MSS.51 (Figure 2) and the 410bp long EcoRi- BspMI DNA fragment (AC40 probe) containing the promoter of gene RPC40 (Della Seta et al., 1990a) were 3'-end labelled. The purified ABFl factor was incubated with either the MSS.51 or AC40 probes. Protein-DNA complexes were formed in presence of pBR322 DNA used as non-specific competitor. They were then separated by gel electrophoresis and detected by autoradiography. Figure 5A shows that, in the presence of an increasing concentration of the ABFl protein, the AC40 andMSS.51 probes are complexed with a similar effectiveness (lanes 1, 2, 3, 4 and 8, 9, 10, 11, respectively). To prove
that the interaction of the ABFl protein with the expected MSS.51 ABFl binding site is specific, a 2 1 bp oligonucleotide encompassing the RPC40 ABFl binding site (Della Seta et al., 1990a; Della Seta, 1990) was used as a competitor (cf. legend Figure 5). Probes AC40 and MSS.51 are competed with the same efficiency (Figure 5A, lanes 5, 6, 7 and 12, 13, 14). These results indicate that the MSS.51 A T C A T G C G C T E sequence is a bona fide ABFl binding site.
This conclusion was further substantiated by carrying out the following experiment (Figure 5B). In the presence of constant quantities of probe MSS.51 and of protein ABFl, we have added increasing quantities of different competitors in the reaction mixtures. As shown above, 12ng of the 2 1 bp A C40 wild-type oligonucleotide completely inhibit the formation of the protein/DNA com- plex (Figure 5B, lane 14). Yet 300ng of the MA2 competitor with an adenosyl residue in place of a thymidyl one in position 8 (cf. legend Figure 5) still allow the appearance of an appreciable quantity of the complex (Figure 5B, lane 7), whereas 300 ng of the MA5 competitor, with the same kind of modifi- cation in position 11, seems not to affect the com- plex formation (Figure 5B, lane 11). Since it has been shown that these nucleotide exchanges alter the ABFl-DNA interactions (Della Seta et al., 1990a; Della Seta, 1990), our data confirm that
Figure 4. SI transcription mapping and Northern blotting of the QRZ5 mRNA. (Top) (A) Strategy used to map the 5'-end of the QRZ5 mRNA. The 685 bp long AvaII-EcoRI fragment was 5'-end labelled at the AvaII end. (B) Strategy used to map the 3'-end. The 329 bp long Hinfl-Hin-fl fragment was 3'-end labelled. The QRl5 open reading frame is depicted by a shaded bar. R, EcoRI; A, AvaII; H, Hinfl. (Bottom) Autoradiographs (a) and (b). Lane 1: Hinfl-cut pBR322 DNA 32P end-labelled with Klenow DNA polymerase. Lane 2: S1 nuclease digest of the probe (no mRNA in the hybridization medium). Lane 3: S1 nuclease digest of the DNA p r o b e mRNA hybrid; the arrows show the size of the probe protected by the transcript. Lane 4: single-strand end-labelled probe used as size marker (no S1 digestion). (c) Northern blotting of the QRl5 transcript probed with the '*P-labelled 161 bp long Hin-fl-AvaII fragment (cf. Figure 2).

M. SIMON ET AL. 566
A AC40 protein ABF1, by interacting with the MSS.51 ABFl binding site, could control the expression of MSSSl I QRZ5 and MSS.51.
1 1 I 2 1 3 I 4 1518 I 7 81 1 t10111112113114 Genetic mapping of the MSS5 1 gene
I A 32P-labelled MSS5I DNA probe was hybrid-
C- -c ized with the yeast chromosomes of strain MH41- 7B separated by pulsed-field gel electrophoresis (Carle and Olson, 1984; Sor, 1988). Gene MSS51 appeared to be located on chromosome IV or XII, since both chromosomes comigrate under our experimental conditions. Using the genetic method described by Klapholz and Esposito (1982), we found that gene MSS5I is on chromosome XII. We mapped MSSSI by tetrad analysis using markers known to belong to this chromosome. The strains used are described in Table 1 and the results are presented in Table 3. Gene MSS.51 is located on the I I MA2 I I WT I rightarmbetweentheRDNIandtheCDC42lociat 40.3 and 22.1 cM, respectively. The tetrad distri- bution obtained for the gene pair cdc42-IIRDNI:: LEU2 is very similar to those previously published (Mortimer e t al., 1989).
d- -d
B 1 I 2 I 3 4 I 5 I@ I 7 8 I llolll 12113114
,c
,d
AC40 WT TCGAGAGTCACTATAAACGGC
MA2 TCGAGAGaCACTATAAACGGC
MA5 TCGAGAGTCAaTATAAACGGC
C
Figure 5 . Gel retardation experiments. (A) Protein-DNA com- plexes formed between proteins ABFl and either the AC40 (lanes 1-7) or MSS51 probes (lanes 8-14). The concentration ofprobes was maintained constant. In lanes 1 4 the amount of protein ABFl was 0, 2.7, 6.75 and 13.5 ng, respectively. The same pro- gression was followed in lanes 8-1 1. In the presence of 13.5 ng of protein ABFl in lanes 5-7 and lanes 12-14, 1.2, 12 or 60ng of AC40 wild-type oligonucleotide were added to complete with either the AC40 or MSSSI probes. Protein-DNA complexes were analysed by gel electrophoresis. (c) Position of DNA-pro- tein complexes; (d) position of DNA probes. (B) Effect of mutated oligonucleotides in competition with the MSSSI probe. The cohcentration of the MSSSI probe was maintained constant in every lane. In lanes 1-3, the quantity of protein ABFl was 0, 13.5 and 27 ng, respectively; no competitor was present. In lanes 4-14, the amount of protein ABFl was 13.5 ng. In lanes 4-7, the quantity of MA2 competitor was 12,30,120 and 300 ng, respect- ively. In lanes 8-1 1, similar quantities of MA5 competitor were added. In lanes 12-14, 1.2.3 and 12 ng of AC40 wild-type oligo- nucleotide were added. (C) Sequences of the competitors used.
ACKNOWLEDGEMENTS We thank E. Boy-Marcotte, M. Brandriss, D. Johnson, P. Brisco and D. Hawthorne for strains and A. Besnard and J. G. Valay for excellent assist- ance. This work was supported by the CNRS, the Institut Curie (contrat cooperatif no. 90-25) and the Association pour la Recherche sur le Cancer (contrat no. 6087).
REFERENCES Beck, C. F. and Warren, R. A. J. (1988). Divergent
promoters, a common form of gene organization. Microbial. Rev. 52,3 18-326.
Carle, G. F. and Olson, M. V. (1984). Separation of chro- mosomal DNA molecules from yeast by1 orthogonal- field-alternation gel electrophoresis. Nucl. Acids Res.
Christianson, T., Edwards, J., Mueller, D. and Rabinowitz, M. (1983). Identification of a single tran- scription initiation site for the glutamic tRNA and COB genes in yeast mitochondria. Proc. Natl. Acad. Sci. USA 80,5564-5568.
Costanzo, M. and Fox, T. (1990). Control of mitochon- drial gene expression in Saccharomyces cerevisiae. Ann Rev. Genet. 24,91-113.
Decoster, E., Simon, M., Hayat, D. and Faye, G. (1990). The MSS.51 gene product is required for the translation of the COX1 mRNA in yeast mitochondria. Mol. Gen. Genet. 224,111-1 18.
12,5647-5664.

ANALYSIS OF MSSSI REGION ON CHROMOSOME XI1 OF S. CEREVZSZAE 567
Della Seta, F. (1990). Le facteur ABFl de S. cerevisiae et son interaction avec les gknes RPC40, RPC160, L2A et LZB. Purification, reconnaissance de L’ADN et r81e dans la transcription. ThQe de 1’UniversitC de Paris- Sud, Centre d’Orsay, 12 Septembre 1990.
Della Seta, F., Treich, I., Buhler, J. M. and Sentenac, A., (1990a). ABFl binding sites in yeast RNA polymerase genes. J. Biol. Chem. 265,15168-15175.
Della Seta, F., Ciafre, S. A., Marck, C., Santoro, B., Presutti, C., Sentenac, A. and Bozoni, I. (1990b). The ABFl factor is the transcriptional activator of the L2 ribosomal protein genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 10,2437-2441.
Dorsman J. C., van Heeswijk, W. C. and Grivell, L. A. (1988). Identification of two factors which bind to the upstream sequences of a number of nuclear genes coding for mitochondrial proteins and to genetic el- ements important for cell division in yeast. Nucl. Acids Res. 16,7287-7301.
Dorsman, J. C., Gozdzicka-Jozefiak, A., Van Heeswijk, W. C. and Grivell, L. A. (1991). Multi-functional DNA proteins in yeast: the factors GFI and GFII are identical to the ARS-binding factor ABFl and the centromere-binding factor CPF 1 respectively. Yeast 7,
Faye, G. and Simon. M. (1983). Analysis of a yeast nuclear gene involved in the maturation of mitochon- drial pre-messenger RNA of the cytochrome oxidase subunit 1. CeIl32,77-87.
Grivell, L. A. (1989). Nucleo-mitochondria1 interactions in yeast mitochondrial biogenesis. Eur. J . Biochem. 182, 477-493.
Huet, J., Cottrelle, P., Cool M., Vignais, M. L., Thiele, D., Marck, C., Buhler, J. M., Sentenac, A. and Fromageot, P. (1985). A general upstream binding factor for genes of the yeast translational apparatus. EMBO J. 4, 3539-3547.
401-412.
Klapholz, S. and Esposito, R. (1982). A new mapping method employing a meiotic rec- mutant of yeast. Genetics 100,387-412.
Loenen, W. and Brammar, W. (1980). A bacteriophage lambda vector for cloning large DNA fragments made with several restriction enzymes. Gene 10,
Maniatis, T., Fristch, E. and Sambrook, J. (1982). Mol- ecular Cloning: A Laboratory Manual. Cold Spring Harbour Laboratory Press. Cold Spring Harbor, New York.
Maxam, A. and Gilbert, W. (1980). Sequencing end- labeled DNA with base-specific chemical cleavages. Methods Enzymol. 65,499-560.
Mortimer, R. K., Schild, D., Contopoulou, C. R. and Kans, J. A. (1989). Genetic map of Saccharomyces cerevisiae. Edition 10. Yeast 5,321-403.
Murray, N., Brammar, W. and Murray K. (1977). Lambdoid phages that simplify the recovery of in vitro recombinants. Mol. Gen. Genet. 150,53-61.
Rothstein, R. (1985). Cloning in yeast. In Glover, D. M. (Ed.), DNA Cloning, vol. 1 1, A Practical Approach. IRL Press, New York, pp. 45-66.
Sherman, F., Fink, G. and Hicks, J. (1983). Methods in Yeast Genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Simon, M. and Faye, G. (1984). Organization and pro- cessing of the mitochondrial oxi3/oli2 multigenic transcript in yeast. Mol. Gen. Genet. 196,266-274,
Sor, F. (1 988). A computer program allows the separation of a wide range of chromosome sizes by pulsed field gel electrophoresis. Nucl. Acids Res. 16,4853-4863.
Thomas, P. (1980). Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proc. Natl Acad. Sci. USA 77,5201-5205.
Verdier, J. M. (1990). Regulatory DNA-binding proteins in yeast: an overview. Yeast 6,271-297.
249-2 59.





























