-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
1/10
RESEARCH ARTICLE
Anatomy and Aging of the Amygdala and Hippocampus in
AutismSpectrum Disorder: An In Vivo Magnetic Resonance Imaging
Studyof Asperger Syndrome
Clodagh M. Murphy, Q. Deeley, E.M. Daly, C. Ecker, F.M. OBrien,
B. Hallahan, E. Loth, F. Toal, S. Reed,S. Hales, D.M. Robertson,
M.C. Craig, D. Mullins, G.J. Barker, T. Lavender, P. Johnston, K.C.
Murphy,and D.G. Murphy
It has been proposed that people with autism spectrum disorder
(ASD) have abnormal morphometry and development ofthe amygdala and
hippocampus (AH). However, previous reports are inconsistent,
perhaps because they included peopleof different ASD diagnoses,
ages, and health. We compared, using magnetic resonance imaging,
the in vivo anatomy ofthe AH in 32 healthy individuals with
Asperger syndrome (1247 years) and 32 healthy controls who did not
differsignificantly in age or IQ. We measured bulk (gray1white
matter) volume of the AH using manual tracing (MEASURE).We first
compared the volume of AH between individuals with Asperger
syndrome and controls and then investigatedage-related differences.
We compared differences in anatomy before, and after, correcting
for whole brain size. There wasno significant between group
differences in whole brain volume. However, individuals with
Asperger syndrome had asignificantly larger raw bulk volume of
total (Po0.01), right (Po0.01), and left amygdala (Po0.05); and
when correctedfor overall brain size, total (Po0.05), and right
amygdala (Po0.01). There was a significant group difference in
aging of
left amygdala; controls, but not individuals with Asperger
syndrome, had a significant age-related increase in volume(r50.486,
Po0.01, and r50.007, P50.97, z51.995). There were no significant
group differences in volume or age-related effects in hippocampus.
Individuals with Asperger syndrome have significant differences
from controls in bulkvolume and aging of the amygdala. Autism Res
2011,4:xxxxxx. & 2011 International Society for Autism
Research,Wiley Periodicals, Inc.
Keywords:Asperger syndrome; autism; amygdala; hippocampus;
age
Introduction
It has been suggested that the biological basis of autism
spectrum disorder (ASD) may include anatomical abnorm-
alities in the amygdala and hippocampus [Baron-Cohen,
Ring et al., 2000; Schultz, 2005]. To date, however,
researchfindings have been variable [Cody, Pelphrey et al.,
2002].
Classic lesion studies [e.g. see Kluver, 1939; Rosvold,
Mirsky et al., 1954] and a review [Bachevalier, 1994] have
demonstrated a crucial role of the amygdala in primate
social and emotional behaviors. In humans, the amygda-
la contributes to reward and motivation [Baxter &
Murray, 2002; Gottfried, ODoherty et al., 2003; Holland
& Gallagher, 2004], processing of faces [Haxby, Hoffman,
et al., 2002], recognition of emotions [Adolphs, 2003],
emotional memory [McGaugh, 2004], detecting threat
[Amaral, 2003; Amaral & Bauman, 2003], fear and
anxiety [Davis, Walker et al., 2003; LeDoux, 2003], and
modulates social brain regions [such as fusiform-
extrastriate cortices [Deeley, Daly et al., 2007]. The
amygdala also interacts reciprocally with the hippocam-
pus to encode emotional memories [Richardson, Strangeet al.,
2004]. The hippocampus, in addition to its role in
emotional processing and memory [Bannerman, Rawlins
et al., 2004; Vargha-Khadem, Gadian et al., 1997], is also
involved in spatial learning [Maguire, Gadian et al.,
2000] and verbal novelty detection [Grunwald, Lehnertz
et al., 1998]. Given the difficulties of individuals with
ASD in responding to, and processing, socioemotional
cues, it has been suggested that its biological basis
includes abnormalities in the development and function
of limbic structures, including the amygdala and
INSAR Autism Research 4: 110, 2011 1
Received October 6, 2010; accepted for publication July 20,
2011
Published online in Wiley Online Library
(wileyonlinelibrary.com)
DOI: 10.1002/aur.227
&2011 International Society for Autism Research, Wiley
Periodicals, Inc.
Additional Supporting Information may be found in the online
version of this article.Clodagh M. Murphy and Q. Deeley are first
authors.
From the Kings College London, Institute of Psychiatry,
Department of Forensic and Neurodevelopmental Science, London,
United Kingdom (C.M.M.,
Q.D., E.M.D., C.E., B.H., E.L., F.T., S.R., S.H., D.M.R.,
M.C.C., D.M., T.L., P.J., D.G.M.); Royal College of Surgeons in
Ireland, Dublin, Ireland (F.M.O.,
K.C.M.); Department of Clinical Neuroscience, Kings College
London, Institute of Psychiatry, Centre for Neuroimaging Sciences,
London, United
Kingdom (G.J.B.)
Address for correspondence and reprints: Clodagh M. Murphy,
Department of Forensic and Neurodevelopmental Science, Institute of
Psychiatry,
De Crespigny Park, PO Box 50, Denmark Hill, London SE5 8AF, UK.
E-mail: [email protected]
Grant sponsor: Medical Research Council.
35: 312, 2012 3
21 September 2011 in Wiley Online Library
(wileyonlinelibrary.com)
2012,5:312. 2011 International Society for Autism Research,
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
2/10
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
3/10
Controls were recruited locally by advertisement.
Individuals with Asperger syndrome were recruited withthe
support of the Medical Research Council (MRC) UK
Autism Imaging Multicentre Study (A.I.M.S) (MRC AIMS)
program. Diagnosis of Asperger syndrome was made by
two Consultant Psychiatrists (D.G.M. and D.R.) and a
Nurse Specialist (S.R.), using ICD-10 research diagnostic
criteria [1994]. All individuals fulfiled the diagnostic
criteria of childhood autism, except that they did not
have a history of language delay; they were therefore
subtyped as having Asperger syndrome (International
Classification of Diseases, 10th Edition ICD-10, F84.5).
Where permission was given by individuals with Asperger
syndrome, additional diagnostic measures were sought.
The Autism Diagnostic Interview-Revised (ADI-R) [Lord
et al., 1994] was completed for all 32 individuals with
Asperger syndrome (13 young people and 19 adults).
Thirty of the 32 reached the ADI algorithm cut-offs for
autism in all three domains (social, communication, and
restricted and stereotyped behaviors). One adult and one
young person just failed to reach the ADI Communication
algorithm cut-off for autism by one point. Six adults with
Asperger syndrome agreed to complete the Autism
Diagnostic Observation Schedule (ADOS) [Lord, Rutter
et al., 1989], and were above cut-off for autism on all
ADOS domains.
Ethical approval was obtained from the Institute ofPsychiatry
and Bethlem and Maudsley Hospitals Trust
Research Ethics Committee. All participants gave written
informed consent, including each childs parent (where
relevant).
All participants underwent a structured physical and
psychiatric examination to exclude comorbid medical and
psychiatric disorders, and biochemical, hematologic or
chromosomal abnormalities (including Fragile X syn-
drome) possibly affecting brain function. Participants were
excluded if they had a comorbid psychiatric or medical
disorder affecting brain development (e.g. epilepsy or
psychosis), history of head injury, genetic disorder asso-
ciated with autistic spectrum disorder (e.g. tuberous
sclerosis or Fragile X syndrome), or an IQo70. Intelligence
quotients were measured using the Wechsler Adult Intelli-
gence ScaleRevised (WAIS-R) short form [Weschler, 1999].
MRI Data Acquisition
MRI data were obtained using a GE Sigma 1.5T
Neuro-optimised MR system (General Electric, Milwaukee,
Wisconsin). Whole head coronal three-dimensional (3D)
Inversion recovery prepared spoiled gradient echo (IR-SPGR
images) (repetition time513.8 msec, echo time52.8msec,
inversion time5450 msec, 256192 acquisition matrix,
reconstructed as a 256256 matrix, over a 220220 field of
view, 1241.5-mm slices) were obtained from all subjects.
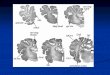
ROI Approach
Manual tracing of the bulk volume (i.e. both gray andwhite
matter) of total, left, and right amygdala and
hippocampus was carried out as previously described
[Cutter, Daly et al., 2006] by a single rater. In brief,
manual tracing was performed on IR-SPGR data sets,
using both MEASURE Image Analysis software [Barta
et al., 1997] (Johns Hopkins University, Baltimore,
Maryland) and published anatomical definitions [van
Amelsvoort, Daly et al., 2001]. The anatomical defini-
tions of the regions were defined using a modified
version of Watsons guidelines [Watson, Andermann
et al., 1992]. Images were realigned parallel to the sylvian
fissure. The most posterior hippocampus measurement
started on the slice displaying the aqueduct of sylvius.
Continuing anteriorly, the superior border of the hippo-
campus merges with the inferior border of the amygdala
and the regions are delineated by white matter and the
temporal horn of the lateral ventricle. If the delineation
is unclear, the inferior border of the posterior amygdala is
arbitrarily marked as a horizontal line drawn medially
from the head of the temporal stem to the medial border
of the amygdala; the hippocampus taken to be gray
matter inferior to the line. The anterior boundary of the
amygdala measurement was made on the slice with
closure of the lateral sulcus (i.e. temporal lobe is joined
to frontal lobe by the limen insulae). Total (i.e. left
plusright) and individual left and right amygdala and
hippocampus were measured. As manually traced regio-
nal brain volumes are affected by brain size, we also
measured total whole brain volume (WBV). The volume
of each region was calculated by multiplying the summed
pixel cross-sectional areas measured by the slice thick-
ness. The rater was blind to subject status and intrarater
reliabilities were obtained for all regions identified. On a
separate test data set of ten scans, the rater had achieved
Table I. Participant Demographics
Controls,N5 32
(31 male, 1
female)
Asperger syndrome,
N5 32 (30 male, 2
female)
Age in years, mean7SD (range) 23711 (1049) 23711 (1247)
FSIQ, mean7SD 111715 108713
Autistic Diagnostic Interview (ADI), N532 (30 male, 2
female)
Social (mean7SD) 1976
Communication (mean7SD) 1375
Restricted, repetitive behavior
(mean7SD)
673
Autistic Diagnostic Observation
Schedule (ADOS), N56 (5
male, 1 female)
Communication 571
Social 872
Total (social and communication) 1373
Stereotyped behaviors and
restricted interests
272
INSAR Murphy et al./Anatomy and aging of the amygdala 35
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
4/10
intrarater and interrater reliabilities of r40.9 for all
regions traced [Bartko & Carpenter, 1976].
In order to control for the relationship of brain volume
and head size, amygdala and hippocampus volumes were
expressed as raw (uncorrected) volumes, and when
normalized, as a percentage of traced WBV. Statistical
analyses were carried out on both raw and normalizedbrain
volumes.
Statistical Analysis
SPSS 15.0 for Windows (SPSS Inc, Chicago, IL) was used for
all statistical analyses. All demographic data (age and IQ)
and brain volume measurements were normally distributed.
We corrected for multiple comparisons (Bonferroni). Those
regions that did survive a corrected Bonferroni (P50.0038)
are marked as a in Table II (Summary of results). Those
regions that did not survive are marked as b. Level
of statistical significance was defined as Po0.05. Results
in the discussion are based on findings with a level
ofstatistical significance ofPo0.05.
Because regional brain volumes may be affected by
overall brain size, and some have suggested that people
with ASD have differences in WBV (i.e. macrocephaly);
statistical analyses were performed on both the raw bulk
volume of the AHC, and after being expressed as a
percentage of WBV (% WBV).
To test our main hypothesis, we first compared people
with Asperger syndrome with the controls. Between-group
differences in age, IQ, and brain volumes were calculated
using independent-samples t-tests.
We then investigated our subsidiary hypothesis, that
there would be no group difference in the effect of age,
using a correlational approach. Prior to comparing differ-
ences in age-related correlations, the data were tested for
(i) linearity, (ii) independence, (iii) homoscedasticity,
and(iv) normality error distribution. None of these assump-
tions was violated. Hence, we initially calculated Pearson
productmoment correlations to investigate within-group
age-related differences in AHC volumes. Then, in order to
examine between-group differences in brain aging, we
converted the relevant Pearsons r coefficient into Fishers
Z-score to test the statistical significance of between
group
differences in correlations [Pallant, 2005].
In addition, we further investigated the relationship
between age and total amygdala hippocampus volume by
calculating a partial correlation to control for the effect
of
WBV. This did not change our results and even enhancedthe effect
size of our correlation coefficients.
ResultsEffect of Group: Asperger Syndrome (N532) and
Controls(N532)
There was no significant between group difference in:
age, Full-Scale IQ, WBV, or bulk volume of hippocampus.
Table II. Summary of Results: Mean AH Volumes and Significant
Differences
Controls (N5 32), mean7SD Asperger syndrome (N5 32), mean7SD
WBV
Total WBV (ml) 1,095.837104.80 1,117.507134.12 t(62)50.720, P5
0.474
Amygdala
Total
Raw (ml) 5.1670.50a 5.5870.66a t(62)52.863, P5 0.006a,b
Corrected (%WBV) 0.4770.04a 0.5070.07a t(62)52.306, P5
0.024a,b
Right
Raw (ml) 2.5270.26a 2.7870.36a t(62)53.300, P5 0.002a,c
Corrected (%WBV) 0.2370.02a 0.2570.04a t(62)52.719, P5
0.008a,b
Left
Raw (ml) 2.6470.30a 2.8070.34a t(62)52.053, P5 0.044a,b
Corrected (%WBV) 0.2470.02 0.2570.03 t(62)51.516, P5 0.135
Hippocampus
Total
Raw (ml) 5.6770.78 5.8070.86 t(62)50.623, P5 0.535
Corrected (%WBV) 0.5270.06 0.5270.07 t(62)50.253, P5 0.801
Right
Raw (ml) 2.9470.44 3.0270.47 t(62)50.691, P5 0.492
Corrected (%WBV) 0.2770.03 0.2770.04 t(62)50.223, P5 0.824
Left
Raw (ml) 2.7370.38 2.7870.43 t(62)50.456, P5 0.650
Corrected (%WBV) 0.2570.03 0.2570.04 t(62)50.069, P5 0.946
WBV, whole brain volume; AH, amygdala and
hippocampal.aSignificant between group difference on independent
samplest-test.bDid not survive Bonferroni.cSurvived Bonferroni.
4 Murphy et al./Anatomy and aging of the amygdala INSAR6
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
5/10
However, individuals with Asperger syndrome had a
significantly larger uncorrected bulk volume of total
(left plus right) (t (62)52.863, Po0.01), right
(t (62)53.300, Po0.01), and left amygdala
(t(62)52.053, Po0.05) and in the corrected (% WBV)
total (t (49.335) 52.306, Po0.05) and right amygdala
(t(50.066)52.719, Po0.01).
Age and Volume of Amygdala and Hippocampus
Increased corrected (% WBV) volume of total (r50.483,
Po0.01) and left amygdala (r50.486, Po0.01) in con-
trols, but not individuals with Asperger syndrome, was
significantly positively correlated with increasing age.
Further, controls had a significantly larger age-related
increase in corrected (% WBV) volume of left amygdala
than people with Asperger syndrome (z51.995) (z5 sig-
nificant ifr1.96 or Z1.96).
No significant correlations between age and volume of
hippocampus were identified in either group.
Increasing age in controls, but not individuals with
Asperger syndrome, was significantly negatively corre-lated with
WBV (r50.549, Po0.01). There was a trend
toward controls having a significantly larger age-related
decrease in WBV than people with Asperger syndrome
(z51.953; Fig. 1) (See Supplementary data Appendix
Table A).
Discussion
We compared the bulk volume and age-related differ-
ences of the amygdala and hippocampus in physically
healthy individuals with Asperger syndrome and
controls, who did not differ significantly in age and IQ.
Volume of the Amygdala and Hippocampus
Our finding that people with Asperger syndrome have a
significantly larger volume of total, left, and right
amygdala and no difference in volume of hippocampus,
is in agreement with earlier findings of amygdala
enlargement in individuals with ASD as measured using
VBM [Abell, Krams et al., 1999], ANALYZE [Howard,
Cowell et al., 2000], and manual tracing [Mosconi,
Cody-Hazlett et al., 2009]. It is also in agreement with
previous reports of no significant difference in hippo-
campal volume as measured using ANALYZE [Howard
et al., 2000] and manual tracing [Piven, Bailey et al.,
1998]. Furthermore, our finding of an enlarged amygdala
was true for both uncorrected (total, left, and right
amygdala) and corrected (total and right amygdala)
values.
Our results are, nevertheless, in contrast to others who
found no significant difference in bulk volume of
amygdala [Haznedar et al., 2000; Palmen et al., 2006],
or reduced volume of amygdala [Nacewicz, Dalton et al.,
2006] as measured using manual tracing. Palmens study
cohort was only very slightly smaller than ours but it
included much younger (725 years old) groups of mixed
diagnoses (both autism and Asperger syndrome). Simi-
larly, Nacewiczs study included a younger (825 years
old) group of 11 males with autism and 5 with Aspergersyndrome
or pervasive developmental disorder. Likewise,
Haznedars study included ten individuals with autism
and seven individuals with Asperger syndrome, who
ranged in IQ from 55 to 125. In contrast, we included a
slightly larger sample of people with Asperger syndrome
from a wider age group, who had an IQ above 70, and
they did not significantly differ from controls in IQ or
age. Distinct differences between brain anatomy of
individuals with Asperger and autism have been reported
[McAlonan, Cheung et al., 2009; McAlonan, Suckling
et al., 2008]. Such differences in autistic subtypes, age,
and IQ may partially explain the differences in our
findings.
Development of the Amygdala and Hippocampus
There are relatively few in vivo neuroimaging studies of
normal amydala and hippocampal development, and
most previous studies have been cross-sectional [Giedd
et al., 1996]. Nevertheless, those that are available
suggest
that amygdala and hippocampus volume in typical
human development changes with age, and that the
Figure 1. The relationship between age and corrected bulk
volume of the left amygdala. NB: Controls had a
significantly
larger age-related difference in corrected bulk volume of the
left
amygdala (significant between group difference: z51.995).
r5 correlation co-efficient; correlation is significant at
the
0.01 level; strength of relationship: r50.10.295 small,
r50.300.495medium, and r50.501.05 large; z5 signifi-
cant if o1.96 or41.96; AS, Asperger syndrome.
INSAR Murphy et al./Anatomy and aging of the amygdala 57
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
6/10
developmental trajectory may differ between amygdala
and hippocampus. For example, in boys left amygdala
volume increases between 4 and 18 years of age [Giedd
et al., 1996], but changes relatively little in early
adulthood [Pruessner, Collins, et al., 2001]. In contrast,
male hippocampal volume does not change during
adolescence [Giedd et al., 1996; Schumann et al., 2004],
increases in young adulthood (1921 years old, [Suzuki,
Hagino et al., 2005] but decreases in later life and old
age[Liu, Lemieux et al., 2003; Murphy, DeCarli et al., 1996;
Pruessner et al., 2001; Raz, Lindenberger et al., 2005].
However, rates of volume loss may be significantly less
marked than in parietal and frontal areas [Grieve, Clark
et al., 2005] suggesting that the timing of age-related
changes may differ across brain regions.
Schumanns [Schumann et al., 2004, 2009] investiga-
tion of the amygdala and hippocampus further high-
lights the importance of age in amygdala hippocampal
development. Schumanns [Schumann et al., 2004] study
suggested that differences in bulk volume of the
amygdala and hippocampus (measured using manualtracing) between
people with ASD and controls vary
by age, diagnostic subtype, and anatomical location.
Schumanns group first investigated their sample as one
age group (718 years old) and found amygdala enlarge-
ment only in young people with low-functioning autism
and hippocampal enlargement in young people with
both low- and high-functioning autism. No differences
were found in young people with Asperger syndrome.
In controls, age positively correlated with amygdala
volume, whereas no correlation between age and volume
was found for individuals with autism or Asperger
syndrome. As such, Schumann et al. further investigated
the relationship between age and amydala by dividing
their sample into children and adolescents; amygdala
enlargement was specific to young children (712 years
old) with autism (both low and high functioning),
although a trend toward enlarged right amygdala was
found in young children with Asperger syndrome
(N511, P50.06). No significant difference in amygdala
volume was found between adolescents (12.7518.5 years
old) with autism, Asperger syndrome (N513), and
controls (N511). While this study was a very valuable
first step, subjects were separated into categorical age
groups (i.e. children and adolescents), rather than
using a continuous approach. However, neuroimagingevidence
suggests that postnatal phases of brain devel-
opment do not begin, or end, at specific chronological
ages, but extend across age ranges [Giedd et al., 1996].
Age and Volume of the Amygdala and Hippocampus
We therefore investigated the continuous relationship
between age and volume of the amygdala and hippocampus
from childhood into adulthood. We found a significantly
stronger association of age and left amygdala volume in
controls relative to people with Asperger syndrome. Other
studies have previously reported the presence of amygdala
enlargement at an early age in childhood autism [Mosconi
et al., 2009; Schumann et al., 2004, 2009; Sparks et al.,
2002]
and in adolescence [Groen et al., 2010]. Our findings extend
that work and suggest that amygdala enlargement in
individuals with Asperger syndrome persists into adult-
hood. Also, they suggest that although the amygdalavolume of
people with Asperger syndrome may be
significantly enlarged at an early age, it does not continue
to increase in volume with age. In contrast, our finding of
increasing amygdala volume with age in controls is in
keeping with previous reports of typical amygdala develop-
ment [Giedd et al., 1996; Schumann et al., 2004]. In sum,
our findings support earlier reports of amygdala enlarge-
ment in young children and adolescents with ASD
[Mosconi et al., 2009; Schumann et al., 2004, 2009; Sparks
et al., 2002] and also suggest that, in adolescent and adult
males with Asperger syndrome, the amygdala continues to
be significantly larger compared with age-matched
controls.Furthermore, the development of the amygdala appeared
to
be different across groups. The amygdala of controls
continued to increase with age, whereas the amygdala of
individuals with Asperger syndrome did not. Our results
suggest that both the volume and aging of the amygdala is
significantly different in individuals with Asperger syn-
drome from controls. Future longitudinal studies are
required to help clarify the development and aging of the
amygdala and hippocampus across the lifespan; and to
determine if this differs across the diagnostic subtypes of
ASD.
Potential Causes
The causes of the subtle differences in morphometry and
development of the amygdala and hippocampus that we
found are not known, but probably include a complex
geneenvironment interaction. For example, the amyg-
dala plays a central role in fear and anxiety [Davis et al.,
2003; LeDoux, 2003]; people with an ASD have a
significantly increased risk of experiencing stress and
anxiety disorders [for review, see White, Oswald et al.,
2009]; and stress in turn impacts on amygdala develop-
ment [McEwen, 2007]. For instance, in animals, acute
and chronic stress increases dendrite growth in amygdala[Vyas,
Mitra et al., 2002] and it has been suggested that
stress-related hypertrophy of amygdala may contribute to
anxiety [Vyas, Bernal et al., 2003]. Hence, it is possible
that increased stress in individuals with Asperger syn-
drome may have contributed to, or result from, the
increased volume of amygdala we found. However,
genetic influences may also contribute to differences in
amygdala and hippocampus volume. Preliminary support
for this suggestion is provided by studies that reported
6 Murphy et al./Anatomy and aging of the amygdala INSAR8
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
7/10
differences in amygdala hippocampal volume within the
relatives of individuals with ASD [Dalton et al., 2007;
Rojas et al., 2004].
The specific genes/gene systems that may underpin the
differences we found are unknown. Nevertheless, some
have reported that cortical gray matter overgrowth,
including within the temporal lobes, in young people
[Wassink, Hazlett et al., 2007] (but not adults [Raznahan,
Pugliese et al., 2009] with autism may be associated with
functional variation in the serotonin transporter gene.
This suggestion is supported by other work demonstrating
that serotonin is crucial to brain development, including
synaptic modeling, neurogenesis, dendritic organization
and axon myelination [Whitaker-Azmitia, 2001], and
abnormal brain serotonin synthesis has been found in
children with autism [Chugani, Muzik et al., 1999]; and a
significant reduction in brain 5-HT2A receptor density
has been reported in adults with ASD [Murphy, Daly
et al., 2006] and their relatives [Goldberg, Anderson
et al., 2009]. Hence, genetically determined differences in
serotonin metabolism may contribute toward our find-ings. There
are however, other potential candidate genes.
For example, there is increasing support from our group
and others for an association between ASD and genetic
variation in the glutamatergic and GABAergic systems; in
nonautistic populations glutamate and GABA modify
neuronal growth, connectivity and function, and we have
previously reported that physically healthy, normal IQ,
medication nave adults with ASD have a significant
increase in the glutamate/glutamine (Glx) concentration
of amygdala and hippocampus complex [Page, Daly
et al., 2006]. Future studies are required to replicate our
work, and to investigate potential genetic and environ-
mental factors associated with differences in morphome-
try of the amygdala and hippocampus.
Finally, we do not suggest that people with Asperger
syndrome only have differences in the anatomy of the
amygdala and hippocampus, as both metabolic [Endo,
Shioiri et al., 2007; Otsuka, Harada et al., 1999; Page
et al., 2006] functional [Ashwin, Baron-Cohen, et al.,
2007; Baron-Cohen, Ring et al., 1999; Critchley, Daly
et al., 2000; Grelotti, Klin et al., 2005; Pierce et al.,
2001], and subtle neural network differences [Ecker,
Rocha-Rego et al., 2010] in the amygdala and hippo-
campus of people with ASD have also been reported.
Hence, our findings add to the body of evidence that
individuals with ASD have complex differences from
controls in the structure, function, and metabolism of
the amygdala and hippocampus. However, the relation-
ship between these differences is unknown, and further
research is required to clarify this.
Limitations. This study was cross-sectional in design,and
specific to individuals with Asperger syndrome.Hence, our findings
describe age-related differences, not
individual changes across time, and our findings may
notgeneralize to others within the autism spectrum.Nevertheless, we
were able to examine age-relateddifferences across an age-span of
39 years (1049 years ofage), which would not have been practically
possible usinga longitudinal design. Also, the study was limited to
apredominantly male sample and further studies are neededto
ascertain whether these findings are also present infemales [Craig,
Zaman et al., 2007]. ADOS assessmentswere not available for all
subjects. However, ADI-Rs werecompleted on all subjects (N532), and
ICD-10 researchdiagnoses were confirmed by two Consultant
psychiatristsand a Nurse Specialist trained in the ADI and
ADOS.
We also carried out multiple statistical tests. However,
we corrected (Bonferroni testing) for the increased risk of
Type 1 errors due to multiple testing. Moreover, we used a
relatively large sample of one clearly defined diagnostic
group (healthy individuals with Asperger syndrome),
who had no other medical history that may adversely
affect brain development, and we controlled for head
size. Finally, due to the nature of our study, we cannot
determine if our findings of differences in volume ofamygdala
are a cause or a consequence of social
communication difficulties and their associated stress
and anxiety. Hence, our findings must be viewed as
preliminary.
Conclusion
Individuals with Asperger syndrome have significant
differences from controls in volume and aging of the
amygdala. The cause of these differences is unknown, and
most likely includes a complex interaction between a
primary difference in brain development and abnormal
interactions between the affected individual and their
environment. Future studies of amygdala and hippocampus
development, combining structural and functional imaging
with clinical measures of behavior are warranted in ASD.
Acknowledgments
We thank all the individuals and their families who
participated in this study and our colleagues for their
help in recruiting subjects. This study was supported by a
grant from the Medical Research Council (MRC UK
A.I.M.S research program). None of the authors reported
any financial interests or potential conflicts of interests
associated with this study. The authors declare no
conflict of interest.
References
Abell, F., Krams, M., Ashburner, J., Passingham, R., Friston,
K.,
et al. (1999). The neuroanatomy of autism: A voxel-based
INSAR Murphy et al./Anatomy and aging of the amygdala 79
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
8/10
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
9/10
Grieve, S.M., Clark, C.R., Williams, L.M., Peduto, A.J.,
&
Gordon, E. (2005). Preservation of limbic and paralimbic
structures in aging. Human Brain Mapping, 25, 391401.
Groen, W., Teluij, M., Buitelaar, J., & Tendolkar, I.
(2010).
Amygdala and hippocampus enlargement during adolescence
in autism. Journal of American Academy of Child Adolescent
and Psychiatry, 49, 552560.
Grunwald, T., Lehnertz, K., Heinze, H.J., Helmstaedter, C.,
&
Elger, C.E. (1998). Verbal novelty detection within thehuman
hippocampus proper. Proceedings of the National
Academy of Sciences USA, 95, 31933197.
Haug, J.S., Goldner, C.M., Yazlovitskaya, E.M., Voziyan, P.A.,
&
Melnykovych, G. (1994). Directed cell killing (apoptosis)
in human lymphoblastoid cells incubated in the presence
of farnesol: Effect of phosphatidylcholine. Biochimica
Biophysica Acta, 1223, 133140.
Haxby, J.V., Hoffman, E.A., & Gobbini, M.I. (2002).
Human
neural systems for face recognition and social communica-
tion. Biological Psychiatry, 51, 5967.
Haznedar, M.M., Buchsbaum, M.S., Wei, T.C., Hof, P.R.,
Cartwright, C., et al. (2000). Limbic circuitry in patients
with
autism spectrum disorders studied with positron
emissiontomography and magnetic resonance imaging. American
Journal of Psychiatry, 157, 19942001.
Herbert, M.R., Ziegler, D.A., Deutsch, C.K., OBrien, L.M.,
Lange, N., et al. (2003). Dissociations of cerebral cortex,
subcortical and cerebral white matter volumes in autistic
boys. Brain, 126, 11821192.
Holland, P.C., & Gallagher, M. (2004). Amygdala-frontal
inter-
actions and reward expectancy. Current Opinion in Neuro-
biology, 14, 148155.
Howard, M.A., Cowell, P.E., Boucher, J., Broks, P., Mayes, A.,
et al.
(2000). Convergent neuroanatomical and behavioural evi-
dence of an amygdala hypothesis of autism. Neuroreport, 11,
29312935.
Kluver, H.a.B.P. (1939). Preliminary analysis of functioning
of
the temporal lobes in monkeys. Archives of Neurological
Psychiatry, 42, 9791000.
LeDoux, J. (2003). The emotional brain, fear, and the
amygdala.
Cellular and Molecular Neurobiology, 23, 727738.
Liu, R.S., Lemieux, L., Bell, G.S., Sisodiya, S.M., Shorvon,
S.D.,
et al. (2003). A longitudinal study of brain morphometrics
using quantitative magnetic resonance imaging and differ-
ence image analysis. Neuroimage, 20, 2233.
Lord, C., Rutter, M., Goode, S., Heemsbergen, J., Jordan, H., et
al.
(1989). Autism diagnostic observation schedule: A standar-
dized observation of communicative and social behavior.
Journal of Autism and Developmental Disorders, 19,
185212.
Lord, C., Rutter, M., & Le Couteur, A. (1994). Autism
Diagnostic
Interview-Revised: A revised version of a diagnostic
interview
for caregivers of individuals with possible pervasive
develop-
mental disorders. Journal of Autism and Developmental
Disorders, 24, 659685.
Maguire, E.A., Gadian, D.G., Johnsrude, I.S., Good, C.D.,
Ashburner, J., et al. (2000). Navigation-related structural
change in the hippocampi of taxi drivers. Proceedings of
the National Academy of Sciences USA, 97, 43984403.
McAlonan, G.M., Suckling, J., Wong, N., Cheung, V.,
Lienenkaemper, N., et al. (2008). Distinct patterns of grey
matter abnormality in high-functioning autism and Asper-
gers syndrome. Journal of Child Psychology and Psychiatry,
49, 12871295.
McAlonan, G.M., Cheung, C., Cheung, V., Wong, N., Suckling,
J.,
et al. (2009). Differential effects on white-matter systems
in
high-functioning autism and Aspergers syndrome. Psycho-
logical Medicine, 39, 18851893.McEwen, B.S. (2007). Physiology
and neurobiology of stress and
adaptation: Central role of the brain. Physiological
Reviews,
87, 873904.
McGaugh, J.L. (2004). The amygdala modulates the consolida-
tion of memories of emotionally arousing experiences.
Annual Review of Neuroscience, 27: 128.
Mosconi, M.W., Cody-Hazlett, H., Poe, M.D., Gerig, G.,
Gimpel-Smith, R., et al. (2009). Longitudinal study of
amygdala
volume and joint attention in 2- to 4-year-old children with
autism. Archives of General Psychiatry, 66, 509516.
Murphy, D.G., DeCarli, C., McIntosh, A.R., Daly, E., Mentis,
M.J.,
et al. (1996). Sex differences in human brain morphometry
and metabolism: An in vivo quantitative magnetic resonance
imaging and positron emission tomography study on the
effect of aging. Archives of General Psychiatry, 53, 585594.
Murphy, D.G., Daly, E., Schmitz, N., Toal, F., Murphy, K., et
al.
(2006). Cortical serotonin 5-HT2A receptor binding and
social communication in adults with Aspergers syndrome:
An in vivo SPECT study. American Journal of Psychiatry, 163,
934936.
Nacewicz, B.M., Dalton, K.M., Johnstone, T., Long, M.T.,
McAuliff, E.M., et al. (2006). Amygdala volume and non-
verbal social impairment in adolescent and adult males with
autism. Archives of General Psychiatry, 63, 14171428.
Nicolson, R., DeVito, T.J., Vidal, C.N., Sui, Y., Hayashi,
K.M.,
et al. (2006). Detection and mapping of hippocampal
abnormalities in autism. Psychiatry Research, 148, 1121.Otsuka,
H., Harada, M., Mori, K., Hisaoka, S., & Nishitani, H.
(1999). Brain metabolites in the hippocampus-amygdala
region and cerebellum in autism: An 1H-MR spectroscopy
study. Neuroradiology, 41, 517519.
Page, L.A., Daly, E., Schmitz, N., Simmons, A., Toal, F., et
al.
(2006). In vivo 1H-magnetic resonance spectroscopy study of
amygdala-hippocampal and parietal regions in autism.
American Journal of Psychiatry, 163, 21892192.
Pallant, J. (2005). SPSS survival manual. A step by step guide
to
data analysis using SPSS for Windows. Buckingham, Maiden-
head, UK: Open University Press.
Palmen, S.J., van Engeland, H., Hof, P.R., Schmitz, C., et al.
(2004).
Neuropathological findings in autism. Brain, 127,
25722583.Palmen, S.J., Hulshoff Pol, H.E., Kemner, C., Schnack,
H.G.,
Sitskoorn, M.M., et al. (2005). Brain anatomy in
non-affected
parents of autistic probands: A MRI study. Psychological
Medicine, 35, 14111420.
Palmen, S.J., Durston, S., Nederveen, H., & Van Engeland,
H.
(2006). No evidence for preferential involvement of medial
temporal lobe structures in high-functioning autism. Psycho-
logical Medicine, 36, 827834.
Pierce, K., Muller, R.A., Ambrose, J., Allen, G., &
Courchesne, E.
(2001). Face processing occurs outside the fusiform face
INSAR Murphy et al./Anatomy and aging of the amygdala 911
-
8/13/2019 Anatomy and Aging of Amygdala and Hippocampus in
ASD
10/10
area in autism: Evidence from functional MRI. Brain, 124,
20592073.
Piven, J., Bailey, J., Ranson, B.J., & Arndt, S. (1998). No
difference
in hippocampus volume detected on magnetic resonance
imaging in autistic individuals. Journal of Autism and
Developmental Disorders, 28, 105110.
Pruessner, J.C., Collins, D.L., Pruessner, M., & Evans, A.C.
(2001).
Age and gender predict volume decline in the anterior and
posterior hippocampus in early adulthood. Journal of
Neuroscience, 21, 194200.
Raymond, G.V., Bauman, M.L., & Kemper, T.L. (1996).
Hippo-
campus in autism: A Golgi analysis. Acta Neuropathology, 91,
117119.
Raz, N., Lindenberger, U., Rodrigue, K.M., Kennedy, K.M.,
Head, D., et al. (2005). Regional brain changes in aging
healthy adults: general trends, individual differences and
modifiers. Cereberal Cortex, 15, 16761689.
Raznahan, A., Pugliese, L., Barker, G.J., Daly, E., Powell, J.,
et al.
(2009). Serotonin transporter genotype and neuroanatomy in
autism spectrum disorders. Psychiatric Genetics, 19, 147150.
Richardson, M.P., Strange, B.A., Dolan, R.J. (2004). Encoding
of
emotional memories depends on amygdala and hippocampus
and their interactions. Nature Neuroscience, 7, 278285.Rojas,
D.C., Smith, J.A., Benkers, T.L., Camou, S.L., Reite, M.L.,
et al. (2004). Hippocampus and amygdala volumes in parents
of children with autistic disorder. American Journal of
Psychiatry, 161, 20382044.
Rosvold, H.E., Mirsky, A.F., & Pribram, K.H. (1954).
Influence of
amygdalectomy on social behavior in monkeys. Journal of
Comparative and Physiological Psychology, 47, 173178.
Saitoh, O., Karns, C.M., & Courchesne, E. (2001).
Development of
the hippocampal formation from 2 to 42 years: MRI evidence
of smaller area dentata in autism. Brain, 124, 13171324.
Salmond, C.H., Ashburner, J., Connelly, A., Friston, K.J.,
Gadian, D.G., et al. (2005). The role of the medial temporal
lobe in autistic spectrum disorders. European Journal of
Neurosciences, 22, 764772.
Schultz, R.T. (2005). Developmental deficits in social
perception
in autism: The role of the amygdala and fusiform face area.
International Journal of Developmental Neurosciences, 23,
125141.
Schumann, C.M., & Amaral, D.G. (2005). Stereological
estima-
tion of the number of neurons in the human amygdaloid
complex. Journal of Comparative Neurology, 491, 320329.
Schumann, C.M., & Amaral, D.G. (2006). Stereological
analysis
of amygdala neuron number in autism. Journal of Neu-
roscience, 26, 76747679.
Schumann, C.M., Hamstra, J., Goodlin-Jones, B.L., Lotspeich,
L.J.,
Kwon, H., et al. (2004). The amygdala is enlarged in
children
but not adolescents with autism; the hippocampus is enlargedat
all ages. Journal of Neuroscience, 24, 63926401.
Schumann, C.M., Barnes, C.C., Lord, C., & Courchesne, E.
(2009). Amygdala enlargement in toddlers with autism
related to severity of social and communication impairments.
Biological Psychiatry, 66, 942949.
Sparks, B.F., Friedman, S.D., Shaw, D.W., Aylward, E.H.,
Echelard, D., et al. (2002). Brain structural abnormalities
in
young children with autism spectrum disorder. Neurology,
59, 184192.
Suzuki, M., Hagino, H., Nohara, S., Zhou, S.Y., Kawasaki, Y., et
al.
(2005). Male-specific volume expansion of the human
hippocampus during adolescence. Cereberal Cortex, 15,
187193.
Sweeten, T.L., Posey, D.J., Shekhar, A., & McDougle, C.J.
(2002).
The amygdala and related structures in the pathophysiology
of autism. Pharmacology, Biochemistry, and Behaviour, 71,
449455.
The ICD10 classification of mental and behavioural
disorders.
Clinical descriptions and diagnostic guidelines. (1994).
Geneva: World Health Organization.
van Amelsvoort, T., Daly, E., Robertson, D., Suckling, J., Ng,
V.,
et al. (2001). Structural brain abnormalities associated
with
deletion at chromosome 22q11: Quantitative neuroimaging
study of adults with velo-cardio-facial syndrome. British
Journal of Psychiatry, 178: 412419.
Vargha-Khadem, F., Gadian, D.G., Watkins, K.E., Connelly, A.,Van
Paesschen, W., et al. (1997). Differential effects of early
hippocampal pathology on episodic and semantic memory.
Science, 277, 376380.
Vyas, A., Mitra, R., Shankaranarayana Rao, B.S., &
Chattarji, S.
(2002). Chronic stress induces contrasting patterns of
dendritic remodeling in hippocampal and amygdaloid neu-
rons. Journal of Neuroscience, 22, 68106818.
Vyas, A., Bernal, S., & Chattarji, S. (2003). Effects of
chronic
stress on dendritic arborization in the central and extended
amygdala. Brain Research, 965, 290294.
Wassink, T.H., Hazlett, H.C., Epping, E.A., Arndt, S., Dager,
S.R.,
et al. (2007). Cerebral cortical gray matter overgrowth and
functional variation of the serotonin transporter gene in
autism. Archives of General Psychiatry, 64, 709717.
Watson, C., Andermann, F., Gloor, P., Jones-Gotman, M.,
Peters, T., et al. (1992). Anatomic basis of amygdaloid and
hippocampal volume measurement by magnetic resonance
imaging. Neurology, 42, 17431750.
Weschler, D. (1999). Weschler abbreviated scale of
intelligence.
Psychological Corporation. San Antonio, TX: Harcourt Press.
West, M.J., Slomianka, L., & Gundersen, H.J. (1991).
Unbiased
stereological estimation of the total number of neurons in
the
subdivisions of the rat hippocampus using the optical
fractionator. The Anatomical Record, 231, 482497.
Whitaker-Azmitia, P.M. (2001). Serotonin and brain develop-
ment: Role in human developmental diseases. Brain Research
Bulletin, 56, 479485.White, S.W., Oswald, D., Ollendick, T.,
& Scahill, L. (2009).
Anxiety in children and adolescents with autism spectrum
disorders. Clinical Psychology Reviews, 29, 216229.
10 Murphy et al./Anatomy and aging of the amygdala INSAR12