Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 375:378-392 (1996)
Anatomy of Rat Semaphorin III/ Collapsin-1 mRNA Expression and
Relationship to Developing Nerve Tracts During Neuroembryogenesis
ROMAN J. GIGER, DAVID P. WOLFER, GERARD MJ. DE WIT, AND JOOST VERHAAGEN
Graduate School Neurosciences Amsterdam, Netherlands Institute for Brain Research, 1105 AZ Amsterdam-ZO, The Netherlands (R.J.G., G.M.J.d.W., J.V.); Rudolf Magnus
Institute for Neurosciences, University Utrecht, 3584 CG Utrecht, The Netherlands (J.V.); Institute of Anatomy, University of Zurich, CH-8057 Zurich, Switzerland (D.P.W.)
ABSTRACT Semaphorin III/collapsin- 1 (semaIII/coll-1) is a chemorepellent that exhibits a repulsive
effect on growth cones of dorsal root ganglion neurons. To identify structures that express semaIII/coll-1 in developing mammals, we cloned the rat homologue and performed in situ hybridization on embryonic, neonatal, and adult rats. The relationship between semaIII/coll-1 mRNA distribution and developing nerve tracts was studied by combining in situ hybridization with immunohistochemistry for markers of growing nerve fibers. At embryonic day 11, semaIII/coll-1 expression was restricted to the olfactory pit, the basal and rostra1 surface of the telencephalic vesicle, the anlage of the eye, the epithelium of Rathke’s pouch, and the somites. At later developmental stages, semaIII/coll-1 mRNA was found to be widely distributed in neuronal as well as in mesenchymal and epithelial structures outside the nervous system. Strong expression was found in the olfactory bulb, retina, lens, piriform cortex, amygdalostria- tal area, pons, cerebellar anlage, motor nuclei of cranial nerves, and ventral spinal cord. After birth, mesenchymal staining decreased rapidly and expression became progressively restricted to specific sets of neurons in the central nervous system (CNS). In the mature CNS, semaIII/coll-1 mRNA remains detectable in mitral cells, neurons of the accessory bulb and cerebral cortex, cerebellar Purkinje cells, as well as a subset of cranial and spinal motoneurons. The temporal and spatial expression pattern of semaIII/coll-1 mRNA and its relationship to emerging nerve tracts suggests that semaIIIicol1- 1 is involved in guiding growing axons towards their targets by forming a molecular boundary that instructs axons to engage in the formation of Specific nerve tracts.
Indexing terms: axon guidance, chemorepulsion, motoneurons, spinal cord, B-50, GAP-43
o 1996 Wiley-Liss, Inc.
The formation of specific connections in the developing nervous system requires axon extension, growth cone guid- ance, and recognition of target cells. Pathway finding and target recognition are mediated by a number of different processes, including contact-mediated attraction and repul- sion (Goodman and Shatz, 1993; Kuhn et al., 19951, chemoattraction (Serafini et al., 1994; Kennedy et al., 19941, and chemorepulsion (Luo et al., 1993; Schwab et al., 1993; Tessier-Lavigne, 1994; Puschel et al., 1995; Messer- smith et al., 1995; Wright et al., 1995; Shepherd et al., 1996). Until recently, the primary candidates for axonal pathfinding were thought to be adhesion and extracellular matrix molecules. However, increasing attention has been focused on new molecules that influence the growth cone by
chemoattraction or chemorepulsion (for reviews, see Dodd and Schuchardt, 1995; Kolodkin, 1996; Culotti and Kolod- kin, 1996). Collapsin-1, the first cloned chemorepellent, is a 100 kDa glycoprotein of the chick that has been shown to induce growth cone collapse and paralysis of dorsal root ganglia (DRG) neurons in vitro (Luo et al., 1993). Since that time, it has become evident that chicken collapsin-1 is a member of a new gene family, given the designation “semaphorins” (Kolodkin et al., 1993; Puschel et al., 1995).
Accepted August 1,1996. Address reprint requests to Joost Verhaagen, Netherlands Institute for
Brain Research, Meibergdreef 33, 1105 AZ Amsterdam ZO, The Nether- lands.
o 1996 WILEY-LISS. INC.

SEMAPHORIN III/COLLAPSIN-1 EXPRESSION IN THE RAT 379
The vertebrate semaphorins are characterized by a con- served domain of approximately 500 amino acids containing one consensus N-glycosylation site and 13 conserved cyste- ine residues. Molecular screening for new members of the semaphorin family, based on the original collapsin and semaphorin sequences, has so far identified 13 members of this gene family, of which two proteins are encoded by viral genes (reviewed by Dodd and Schuchardt, 1995; Inagaki et al., 1995). Furthermore, there exist different-sized tran- scripts of the semaIII/coll-1 gene (Luo et al., 19931, and alternative splicing is likely to occur (Puschel et al., 1995). Semaphorin proteins are either secreted or transmembrane molecules (Dodd and Schuchardt, 1995). However, the surface association of secreted collapsin-1 in brain mem- brane preparations and transfected cells (Luo et al., 1993) suggests that its diffusion in the tissue may be limited.
Semaphorin I (formerly designated " fasciclin IV") is a transmembrane glycoprotein of the grasshopper that has been implicated in guiding sensory axons in the peripheral nervous system (PNS) during embryogenesis (Kolodkin et al., 1992). Semaphorin I1 has been shown to inhibit selec- tively motor neuron growth cones from forming normal synaptic terminal arborizations on their target muscles in transgenic Drosophila (Matthes et al., 1995). Mouse sema- phorin (D)III has been suggested to pattern DRG projec- tions in the spinal cord by selectively repelling nerve growth factor (NGF)-responsive cutaneous afferents (terminating in the dorsal part of the spinal cord) but not the neuro- trophin-3-responsive muscle afferents, which grow into the ventral part of the spinal cord where semaphorin I11 is expressed (Wright et al., 1995; Messersmith et al., 1995).
As a first step in correlating nerve tract formation, axonal growth, and semaIII/coll-1 expression in vivo, we have analyzed the temporal and spatial expression patterns of semaIII/coll-1 mRNA during rat neuroembryogenesis and in adult rats. To determine the relationship between the cellular distribution of semaIII/coll- 1 and the development of nerve fiber tracts, we performed double staining for semaIII/coll-1 mRNA and two established marker proteins for growing nerve fiber tracts, neuron-specific class I11 p-tubulin (TuJ1; Moody et al., 1987; Easter et al., 1993) and the growth-associated protein B50/GAP43 (Biffo et al., 1990; Dani et al., 1991).
MATERIALS AND METHODS RNA isolation and RT-PCR cloning
of rat semaIII/coll-1 cDNA Total RNA from embryonic day (E) 15 brain, adult
piriform cortex, and adult cerebellum of Wistar rats was prepared by tissue homogenization in RNAzol (Cinna/ Biotecx). The homogenate was extracted with chloroform and centrifuged at 12,OOOg for 15 minutes. From the colorless aqueous phase, RNA was precipitated with isopro- panol at -2O"C, resuspended in water, and finally precipi- tated with two volumes of ethanol at - 20°C.
First strand cDNA synthesis was carried out in a volume of 20 p1 by using 1 pg RNA from adult brain, 100 ng oligo(dT) primers, and 200 U Superscript RNaseH- reverse transcriptase (Gibco BRL) following the manufacturer's protocol. Polymerase chain reaction (PCR) was performed with 1 pl aliquots of the cDNA synthesis mixture, 500 pmol of each primer, and a mixture of rTth and Vent DNA polymerases (XL-PCR Kit; Perkin Elmer) at a final MgClz concentration of 1.1 mM. The PCR primers were based on
the cDNA sequence of human semaphorin I11 (Kolodkin et al., 19931, flanking the coding sequence and containing EcoRI and XbaI sites for subcloning. Sense primer 5'- TATGGAATTCCCTGCAGCATGGGCTGG-3' and anti- sense primer 5'-GAGGTTTCTAGAGGTAATGCAGCT- CAGACACTCC-3' EcoRI and XbaI sites are italicized. The reaction was cycled 40 times in a cycle profile of 1 minute at 93"C, 1 minute at 60°C, and 6 minutes at 72"C, followed by a final extension step for 15 minutes at 72°C. The amplified product was digested with EcoRI and XbaI restriction endonucleases (Boehringer Mannheim), purified, and sub- cloned in pBluescript K%+/ -) vectors (Stratagene, La Jolla, CAI. Single strand DNA sequencing was performed by the dideoxy-chain termination method using the Se- quenase 2.0 kit (United States Biochemical Corp.). The entire coding sequence of rat semaIIIicol1-1 was assembled and examined for an open reading frame using the Genetic Computer Group (GCG) software.
Preparation of digoxigenin-labeled cRNA probes
Riboprobes identical (sense) and complementary (anti- sense) to the coding region of the semaIII/coll-1 mRNA were obtained by in vitro transcription of the cloned rat semaIIIicol1-1 cDNA. A 20 p1 reaction mix contained 1 pg linear template DNA, 40 mM Tris-HC1 pH 8.0, 6 mM MgC12, 10 mM dithiothreitol (DTT), 2 mM spermidine, 10 mM NaCl, 2 p1 lox digoxigenin (DIG) RNA labeling mixture (Boehringer; 10 mM ATP, 10 mM GTP, 10 mM CTP, 6.5 mM UTP, and 3.5 mM DIG-11-UTP), 10 U RNase inhibitor, and either 20 U T3 (sense) or T7 (antisense) RNA polymerases (Boehringer). After 2 hours at 37"C, transcrip- tion was terminated by adding 20 U of RNase-free DNase I for 15 minutes. To improve tissue penetration, cRNA probes were partially hydrolyzed for 1 hour at 60°C in 10 mM DTT, 200 mM NaHC03/Na2C03, pH 11, to an average length of 100-250 nucleotides (Schaeren-Wiemers and Gerfin-Moser, 1993). Hydrolysis was stopped by neutraliza- tion with 100 mM acetic acid, and the cRNA probes were precipitated with 1/10 volume of 4 M LiCl and 2.5 volumes of ethanol at -80°C. Precipitated cRNA fragments were recovered in 20 p1 diethylpyrocarbonate (DEPC)-treated water.
Tissue preparation All aspects of animal care and treatment were carried out
according to the local guidelines of the experimental animal care committee. Timed-pregnant Wistar rats used in this study were purchased from Harlan CPB-Zeist, The Nether- lands. The plug day was considered as day zero (EO). Pregnant females were killed by intraperitoneal injection of a lethal dose of HypnormTM (0.1 ml/lOO g body weight) and decapitated. The embryos were rapidly removed via cesar- ean section. E l l , E13, E15, E17, and E l 9 embryos were cooled on wet ice, covered with Tissue-Tek (Miles, Elkhart, IN), and frozen in dry ice-cooled 2-methylbutane. At least five animals were processed for each stage. The developmen- tal stage of embryos was confirmed by assessing the crown- rump length according to Hebel and Stromberg (1986). The day of birth was designated postnatal day zero (PO). New- born rat pups were anesthetized with halothane, quickly decapitated, and also frozen in 2-methylbutane. Coronal and horizontal sections of the head and coronal sections of the whole body at cervical and thoracic levels were prepared from PI , P6, and P12 pups. Brain, spinal cord, samples of

380
muscle tissue, and liver of adult (5-6 months old) rats were dissected and processed separately.
In situ hybridization Cryostat sections of 20 pm were cut at - 15°C and thaw
mounted on polylysine-coated microscope slides coated with 50 pg polylysine per 1 ml of diethylpyrocarbonate (DEPC)-treated water. Sections were air dried at room temperature (rt) and fixed for 20 minutes in freshly pre- pared 4% paraformaldehyde in 0.1 M phosphate-buffered isotonic saline (PBS; pH 7.4), rinsed three times for 5 minutes in PBS, and rinsed once briefly in DEPC-treated water. After acetylation for 10 minutes with 0.25% acetic anhydride in 1% triethanolamine and two washing steps in PBS, sections were rinsed in 2x standard saline citrate (SSC) for 5 minutes and prehybridized for 6 hours at room temperature in 50% formamide, 5 x Denhardt's solution, 5x SSC, 250 pg/ml bakers yeast tRNA, and 500 ygiml sheared and heat-denatured herring sperm DNA. One hundred microliters of hybridization mixture containing 200 ng/ml DIG-labeled riboprobe were applied per slide, sealed with a coverslip, and hybridized overnight at 55°C.
On the following day, sections were washed under high stringency: for 5 minutes at 55°C in 5~ SSC, 5 minutes at 55°C in 2 x SSC, 30 minutes at 55°C in 50% formamide containing 0 . 2 ~ SSC, and 5 minutes in 0 . 2 ~ SSC at room temperature. Sections were then washed in Tris-buffered saline (TBS; 100 mM Tris-HC1, pH 7.5, 150 mM NaCl) for 5 minutes and blocked for 1 hour in a 1% solution of blocking reagent (Boehringer) in TBS. Digoxigenin (DIG)-labeled RNA hybrids were detected for 1 hour with an anti-DIG Fab fragment conjugated to alkaline phosphatase (anti-DIG- AP; Boehringer) in a dilution of 1:2,500, and sections were washed twice for 15 minutes in TBS solution. To block endogenous phosphatase activity, 0.24 mgiml levamisole (Sigma) was added to the color reagents, 300 pg/ml nitro- bluetetrazolium chloride (Sigma) and 170 pg/ml5-bromo-4- chloro-3-indolylphosphate (Sigma) in 100 mM NaC1, 5 mM MgC12, and 100 mM Tris-HC1, pH 9.5. The color reaction was performed overnight at room temperature. Sections were rinsed extensively in 10 mM Tris-HC1, pH 8.0, 5 mM EDTA; counterstained with cresylviolet if desired; dehy- drated in a graded series of ethanol; cleared in xylene; and coverslipped with Histomount (National Diagnostics, At- lanta, GA).
Double staining of semaIII/coll-1 mRNA and growing nerve fiber tracts
In situ hybridization was carried out first, as described above. After color reaction with alkaline phosphatase, the sections were rinsed twice with TBS for 15 minutes and blocked with 1% blocking reagent in TBS for 30 minutes. B50/GAP43 and TuJ l were detected with monoclonal antibodies (anti-B50/GAP43 antibody diluted 15,000 and anti-TuJ1 antibody diluted 1:1,000). Sections were incu- bated overnight at room temperature, washed with TBS for 30 minutes, and incubated with a rabbit anti-mouse immu- noglobulins antibody conjugated to horseradish peroxidase (DAKO; diluted 1 : l O O ) . Visualization of the secondary antibody was performed using diaminobenzidine staining as described in the manufacturer's protocol.
Northern blot analysis RNA samples (approximately 30 yg) prepared from E l 5
and adult rat brain tissue were size separated by
R.J. GIGER ET AL.
electrophoresis in a 0.8% agarose formaldehyde gel, trans- ferred to a positively charged nylon membrane (Boeh- ringer), and crosslinked to the membrane with UV light (Giger et al., 1995). The blot was screened with sense and antisense cRNA probes obtained by in vitro transcription of the semaIIIicol1-1 cDNA (see above). Prehybridization with 0.2 ml/cm2 DIG Easy Hyb solution (Boehringer) was performed for 30 minutes at 65"C, followed by hybridiza- tion overnight with 200 ngiml denatured probe at 65°C. After stringency washes (twice for 5 minutes in 2~ SSC, 0.1% SDS at room temperature and twice for 15 minutes in 0 . 1 ~ SSC, 0.1% SDS at 65"C), the blot was subjected to immunological detection using an anti-DIG-AP conjugate in a dilution of 1:5,000, followed by a color reaction as described under In situ hybridization.
RESULTS SemaIII/coll-1 is the rat homologue
of chicken collapsin-1, mouse semaphorin D, and human semaphorin I11
Based on the cDNA sequence of human semaphorin 111 (Kolodkin et al., 1993), oligonucleotide primers were de- signed for RT-PCR amplification of the rat homologue. PCR was performed on first strand cDNA obtained by reverse transcription of total RNA isolated from adult rat brain. Sequencing of the subcloned PCR product revealed a full-length cDNA clone of rat semaIIIicol1-1 with an open reading frame spanning over 2,316 bp.' Alignment of the nucleotide sequence of the newly amplified cDNA to nucleo- tide sequences in GenBank revealed an identity of 95% to mouse semaphorin D (Puschel et al., 19951, 89% to human semaphorin I11 (Kolodkin et al., 19931, and 81% to chicken collapsin-1 (Luo et al., 1993) at the nucleotide level. In all three species the next most related sequences had 53% identity or less to rat semaIII/coll-1, confirming that the cloned cDNA is the rat homologue of the chemorepulsive molecules collapsin- 1, semaphorin D, and semaphorin 111.
Northern blot analysis To examine the specificity of the semaIIIicol1-1 cRNA
probes used for in situ hybridization, we performed North- ern blot analysis on total RNA isolated from E l 5 brain, adult piriform cortex, and adult cerebellum. The antisense semaIII/coll-1 probe hybridized to three different tran- scripts with lengths of approximately 12 kb, 7.5 kb, and 4 kb, found in both embryonic and adult tissue (Fig. 1). No signal was obtained with the parallel-processed sense probe. The 12 kb transcript was less prominent than the tran- scripts of 7.5 kb and 4 kb. However, the relative abundance of the three transcripts appeared to be independent of the developmental stage. All three transcripts were large enough to encode the cloned 2,316 bp open reading frame of rat semaIII/coll-1 .
Expression profile of semaIII/coll-1 mRNA in the developing and adult rat
A DIG-labeled antisense cRNA probe complementary to the entire coding region of rat semaIII/coll-1 was generated by in vitro transcription and used for in situ hybridization
'The nucleotide sequence of the cloned semaphorin IIIicollapsin-1 cDN.4 from Rattus noruegicus has been sent to the GenBankiEMBL Data Bank and is available under the accession number X95286.

SEMAPHORIN III/COLLAPSIN-1 EXPRESSION IN THE RAT
1 2 3
Fig. 1. Expression of semaIIIicol1-1 by Northern blot analysis. Total RNA was separated on a 1% agarose-formaldehyde gel and subjected to blot hybridization with a digoxigenin (DIG)-labeled ribo- probe specific for rat semaIIIicol1-1. Lane 1: E l 5 brain. Lane 2: Adult piriform cortex. Lane 3: Adult cerebellum. Three different transcripts with estimated lengths of 4 kb, 7.5 kb, and 12 kb were detected in embryonic and adult tissue. The length of the semaIIIicol1-1 tran- scripts was calculated using the 28s and 18s rRNA as molecular weight markers.
histochemistry to examine the distribution of semaIII/ coll-1 mRNA in the developing and adult rat. Staining was absent from control sections, which were processed in parallel with a DIG-labeled sense cRNA probe. Thus, a positive stain was considered as expression of semaIII/ coll-1 message. Figure 2 gives a summary of the time course of semaIII/coll-1 mRNA expression in selected structures. Descriptions of the results are illustrated in Figures 3-7, organized on the basis of the developmental pattern of expression in specific anatomical structures. In addition, Figure 8 shows double staining of semaIII/coll-1 mRNA and developing nerve fiber tracts.
Mesenchymal and epithelial labeling outside the CNS. Nonneuronal semaIII/coll-1 mRNA expression was first detected in E l l embryos. The brachial arches and the anlage of the heart were lined by a thin band of epithelial staining (not shown). In agreement with a previous report (Shepherd et al., 1996), weak to moderate expression was found in the epidermis and the somitic mesoderm (as described below under Spinal cord and somites). The epithe- lium of Rathke’s pouch was moderately stained (Table 1). By E13, staining had become more widespread. It reached a maximum in intensity and was most widely distributed in E l 5 embryos. Its distribution at this age can best be appreciated in low-magnification photomicrographs (Fig. 3A-C). Several instances of epithelial labeling were ob- served. Staining continued in the anlage of the anterior pituitary as it lost contact with the pharyngooral cavity. Moreover, the epithelium of the pharyngooral cavity was in part stained, most prominently along the dorsal surface of the tongue (Fig. 3A,C). Groups of very strongly labeled cells were also detected in the epithelium of the otic vesicle (not shown). The notochord and virtually all parts of the developing skeleton were themselves unstained but en- sheathed by densely stained mesenchyme. The forming vertebrae, intervertebral disks, skull base, ribs, and bones of fore- and hindlimbs were all demarcated by a rim of prominently labeled mesenchyme (Fig. 3A-D). High levels of expression were also found in the mesenchyme around the nasal cavity and in the anlage of the larynx (Fig. 3A,C).
381
Sema-Ill / collapsin-1 mRNA
Structures with persisting expression in the adult
cortical plateicerebral cortex piriform cortex
mitral cells / olfactory bulb accessory olfactoly bulb
medial amygdaloid nucleus Purkinje cells
motor neurons
Spinal and cerebral neuroepithelia
cortical neuroepithelium 0 pontrne neuroepithelium 0
spinal basal plate neuroepithelium 0 0 . - m m r - m . - L D N C s ~ ~ ~ ~ a a - =
Neuronal structures with temporal expression
baso-lateral amygdaloid nucleus x x
spinal intermediate zone
Z 2 D S Z z g 2 3 n o m w w w w w
Non-neuronal structures with temporal expression
somites/spine mesenchyme cribrrform plate mesenchyme
Rathkeladenohypophysis
-2LT:z . - m c u - - L 5 w w w w a n i i 2
staining levels: 4 -maximal, very intense 3 -strong, intense 2 - moderate
0 1 -weak 0 -background, no signal
Fig. 2. Overview of the time course of semaIII/coll-1 mRNA expression in the developing and adult rat. Columns represent develop- mental stages, starting at E l l and ending with 6-month-old adult animals. Rows indicate selected regions as indicated. Intensity of shading of each square represents a semiquantitative evaluation of the expression level of semaIIIicol1-1 mRNA, assigned to one of the five staining levels listed at the bottom. The x indicates that the structure was not yet present or no longer exists.
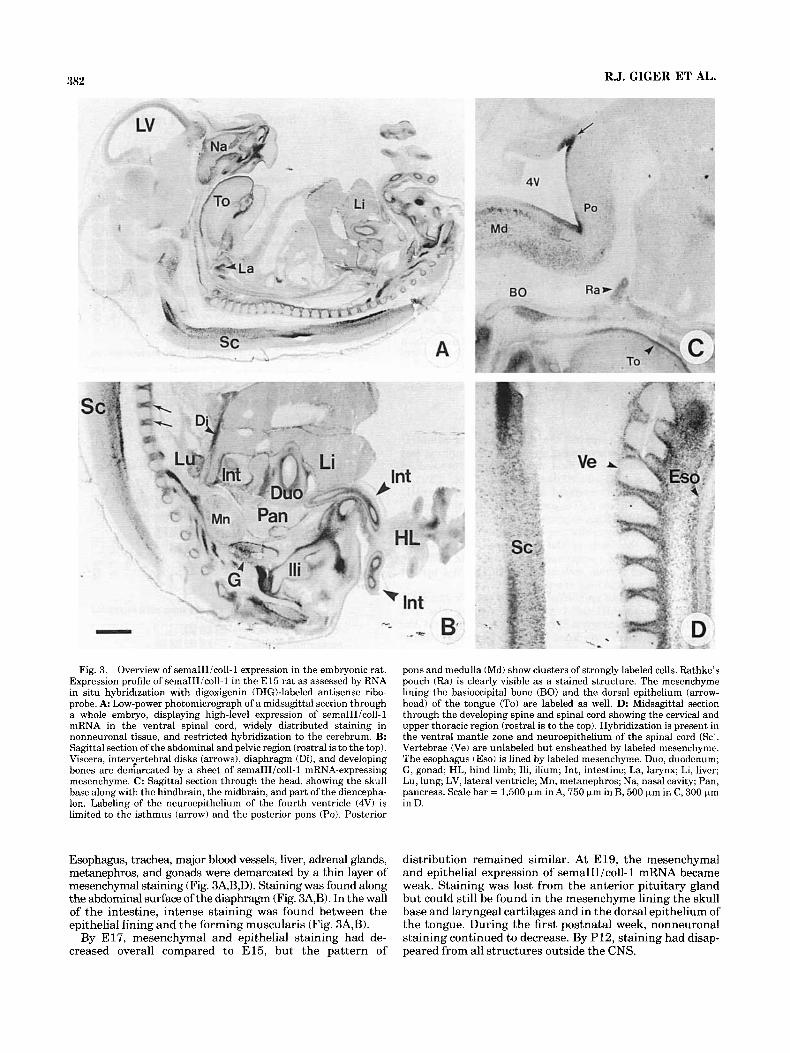
382 R.J. GIGER ET AL.
Fig. 3. Overview of semaIIIicol1-1 expression in the embryonic rat. Expression profile of semaIIIicol1-1 in the E l 5 rat as assessed by RNA in situ hybridization with digoxigenin (DIGblabeled antisense ribo- probe. A Low-power photomicrograph of a midsagittal section through a whole embryo, displaying high-level expression of semaIIIicol1-1 mRNA in the ventral spinal cord, widely distributed staining in nonneuronal tissue, and restricted hybridization to the cerebrum. B: Sagittal section of the abdominal and pelvic region (rostral is to the top). Viscera, intervertebral disks (arrows), diaphragm (Di), and developing bones are demarcated by a sheet of semaIIIicol1-1 mRNA-expressing mesenchyme. C: Sagittal section through the head, showing the skull base along with the hindbrain, the midbrain, and part of the diencepha- Ion. Labeling of the neuroepithelium of the fourth ventricle (4V) is limited to the isthmus (arrow) and the posterior pons (Po). Posterior
Esophagus, trachea, major blood vessels, liver, adrenal glands, metanephros, and gonads were demarcated by a thin layer of mesenchymal staining (Fig. 34B,D). Staining was found along the abdominal surface of the diaphragm (Fig. 3A,B). In the wall of the intestine, intense staining was found between the epithelial lining and the forming muscularis (Fig. 3A,B).
By E17, mesenchymal and epithelial staining had de- creased overall compared to E15, but the pattern of
pons and medulla (Md) show clusters of strongly labeled cells. Rathke’s pouch (Ra) is clearly visible as a stained structure. The mesenchyme lining the basioccipital bone (BO) and the dorsal epithelium (arrow- head) of the tongue (To) are labeled as well. D: Midsagittal section through the developing spine and spinal cord showing the cervical and upper thoracic region (rostral is to the top). Hybridization is present in the ventral mantle zone and neuroepithelium of the spinal cord (Sc:. Vertebrae (Ve) are unlabeled but ensheathed by labeled mesenchyme. The esophagus (Eso) is lined by labeled mesenchyme. Duo, duodenum; G, gonad; HL, hind limb; Ili, ilium; Int, intestine; La, larynx; Li, liver; Lu, lung; LV, lateral ventricle; Mn, metanephros; Na, nasal cavity; Pan. pancreas. Scale bar = 1,500 pm in A, 750 pm in B, 500 pm in C, 300 pm in D.
distribution remained similar. At E19, the mesenchymal and epithelial expression of semaIIIicol1-1 mRNA became weak. Staining was lost from the anterior pituitary gland but could still be found in the mesenchyme lining the skull base and laryngeal cartilages and in the dorsal epithelium of the tongue. During the first postnatal week, nonneuronal staining continued to decrease. By P12, staining had disap- peared from all structures outside the CNS.

Fig. 4. Olfactory system. SemaIIIicoll-1 mRNA expression was detected with a digoxigenin-labeled antisense riboprobe. A: Parasagit- tal section of the head of an E l l embryo. Labeled cells are detected at the surface of the telencephalic vesicle (TE, arrows) facing the olfactory placode (OP). The mesenchyme lining the OP shows intense staining. B: Detail of a coronal section through the head of an E l 9 fetus. The midline (arrow) is to the left. Strong staining is visible in the accessory olfactory bulb (AOB), whereas the main olfactory bulb (OBI reveals weak expression in the mitral cell layer (MC, arrowheads). There is also weak expression in the adjacent granular cell layer (triangle). Scattered labeled cells are found in the orbital cortical plate (CxP). The mesen- chyme of the cribriform plate (CP) shows moderate staining inter- rupted by unlabeled areas. C: Coronal section through an E l9 snout. The nasal septum ( S ) is lined by a thin layer of stained mesenchyme. The sensory epithelium of the vomeronasal organ (VNO) is labeled with very high expression in the epithelium facing the lumen. The respira-
tory epithelium (RE) is devoid of staining, whereas the olfactory epithelium (OE) shows weak staining. D Coronal section of a P1 brain at the level of the amygdala, showing projection areas of mitral and tufted cells. Medial (MA) and basolateral (BLA) amygdaloid nuclei express semaIII/coll-1 transcripts. Staining is also detected in the piriform cortex (arrows) and in the pallidum (Pa). Layers ViVI of the neocortex (circle) show a spotted distribution of moderately to strongly labeled cells. E: Horizontal section through the OB of an adult rat. In the mitral cell layer (MC), a majority of cells is labeled. Between MC and the olfactory glomeruli (GL), individual tufted cells (T) are found to be positive (arrowheads). F: High-power magnification of the MC in the OB of an adult rat. Expression remains detectable in the cytoplasm of individual cells (arrowhead), whereas the nuclei are spared. G: Coronal section through the adult forebrain, showing expression in the acces- sory olfactory bulb (AOB). Scale bar = 250 pm inA,B,E,G, 1,000 pm in C,D, 30 pm in F.

384 R.J. GIGER ET AL.
Figure 5

SEMAPHORIN III/COLLAPSIN-1 EXPRESSION IN THE RAT 385
Olfactory system. At E l l , moderate expression of semaIIUcol1-1 mRNA was found at the basal and rostral surface of the telencephalic neuroepithelium, facing the olfactory placode. The placode itself was unstained but delineated by intensely stained mesenchyme (Fig. 4A). At E13, neuroepithelial labeling was still detectable along the base of the telencephalic vesicle but was weaker in intensity than on E l l . At E15, staining had disappeared from telencephalic neuroepithelium, but a weak to moderate, more diffuse expression became apparent in the differenti- ating zone of the olfactory bulb (Fig. 3A). The mesenchyme around the nasal cavities expressed very high levels of semaIII/coll-1 message (Fig. 3A) that were maintained until E17. At E19, the mitral and granular cell layers in the developing main olfactory bulb exhibited weak staining (Fig. 4B), whereas the accessory olfactory bulb contained densely packed, very strongly labeled cells. Mesenchymal staining in the septum and around the nasal cavity had decreased compared to earlier stages but was still promi- nent in the region of the cribriform plate ensheathing the unstained olfactory fila (Fig. 4B,C). The olfactory epithe- lium exhibited weak to moderate staining (Fig. 4 0 , but the respiratory areas of the epithelium remained devoid of labeling (Fig. 4C, see also Fig. 8C). Near the anterior end of the nasal septum, prominent hybridization was associated with the vomeronasal organ (VNO). The sensory epithe- lium of the VNO showed overall moderate staining, with a zone of very intense labeling on its concave aspect facing the lumen (Fig. 4C). A number of projecting areas of mitral cells and tufted cells outside of the bulb were found to be positive for semaIII/coll-1 mRNA. At E19, the developing piriform cortex and endopiriform nucleus were densely packed with very strongly stained cells, as were several nuclei of the amygdala and the amygdalostriatal area. During early postnatal development (P1 and P6), staining persisted in mitral cells of the main olfactory bulb and in putative relay neurons of the accessory bulb. In the telencephalon of the P1 rat, staining was most conspicuous in the piriform cortex, olfactory tubercle, and medial and basolateral amyg- daloid nuclei (Fig. 4D). In the mature olfactory system of 6-month-old animals, moderate levels of expression were found in mitral cells, tufted cells, neurons of the accessory olfactory bulb, layer I1 of the piriform cortex (Figs. 4E-G, 6A), and medial amygdaloid nucleus (not shown).
Expression in the olfactory system is described separately (see above). SemaIII/coll-1 expression first appeared in distinct zones of the telencephalic neuro- epithelium. At E l l , moderate labeling was detected in the
Telencephalon.
neuroepithelium of the amygdala and basal ganglia (Fig. 4A). By E15, weak labeling became apparent also in the neocortical neuroepithelium (Fig. 3A). By E17, the first labeled cells appeared in the cortical plate. On E19, the cortical and hippocampal neuroepithelia still showed some weak labeling along the ventricular surface (Fig. 5C,D). Labeling in the differentiating cortical plate had become much more prominent and showed a medial to lateral gradient of increasing intensity. Although only weakly labeled cells were found in the cingulate cortex, frontal and parietal regions showed moderate labeling. Expression increased further toward the insular cortex and reached a maximum in the piriform cortex (Fig. 5A-D). Within the developing neocortex, labeled cells were found mainly in the lower plate (future layers V and VIa) and in the subplate (Fig. 5C,D). Weak to moderate staining was detected in the anlage of the dentate gyrus (Fig. 5D) and in the supracom- missural hippocampus (Fig. 5B). In E l 9 brains, labeling was also detected in several nuclei of the basal forebrain. The highest levels of staining were found in the globus pallidus, but also the corpus striatum revealed patches of moderate semaIII/coll-1 expression (Fig. 5B). The amyg- dala showed very strong staining of the medial nucleus, whereas the basolateral nucleus was moderately to strongly stained. At P1, the labeling of the medial amygdaloid nucleus had not changed, but staining was less pronounced in the basolateral amygdaloid nucleus, globus pallidus, and piriform cortex and had disappeared from the striatum (Fig. 4D). Labeling in the neocortex was confined to layers V/VI, where strongly expressing cells could be found (Fig. 4D). In 6-month-old animals, neocortical expression re- mained detectable in layer VI (not shown).
Expression was first detected on E l 1 in the retinal sheet of the forming optic cup and later in the lens placode. Later, labeling of the optic cup shifted to the developing ganglion cell layer, where it reached a maximum at E15. At this age, virtually all cells in the ganglion cell layer showed moderate to strong hybridization (Fig. 8D,E). Staining in the anlage of the lens was very strong and was concentrated in its anterior aspect. Weak labeling was apparent in the lid mesenchyme and in the mesenchymal sheet of the eye anlage as well as in the lid epithelium (Fig. 8D). As development proceeded, the gan- glion cells in the central retina gradually lost staining, whereas those in the peripheral retina retained strong staininguntil E l 9 (Fig. 5A). In the E l 9 orbita, the Harade- rian gland was found to be weakly stained (Fig. 5A). At E19, staining in the lens was still very strong but was limited to
Diencephalon and eye.
Fig. 5. Expression profile of semaIII/coll-1 mRNA in the embryonic head. A Coronal section through the head of an E l 9 embryo. The midline is to the right. In the eye, the ganglion cell layer (G) of the retina along with cells in the anterior epithelium and the equatorial zone (EZ) of the lens show prominent hybridization. The Harderian gland (arrow) is moderately stained. In the rostral telencephalon, labeled cells are scattered through deep layers of the cortical plate (CxP). In the anlagen of the teeth (asterisk), the inner dental epithe- lium is strongly labeled. B: Coronal section of the E l 9 forebrain at the level of the anterior commissure (LV indicates the lateral ventricle). Highest levels of expression are present in the developing piriform cortex (Pi) and globus pallidus (Pa). Patches of moderate labeling are found in the striatum (St). The arrow points to the most rostral, supracommissural portion of the hippocampal formation (induseum griseum), which shows moderate labeling. C: Nissl staining of a coronal section of the telencephalon of an E l 9 embryo, taken from a more caudal level than in B. It shows the layering of the cortical anlage: molecular layer (l), cortical plate (CxP), subplate ((2x3, intermediate
zone (ICx), subventricular zone (SubV), and ventricular zone (cx). The forming dentate gyrus of the hippocampus is indicated by (DG). D: Coronal section adjacent to that shown in C, stained for semaIII/coll-1 mRNA. Weak labeling is present in the neuroepithelium (arrowheads) along the lateral ventricle (LV). Labeled cells are found in the lower cortical plate and in the subplate. They are loosely scattered in the cingulate cortex (CC) and become more numerous toward parietal regions of the neocortex (arrow). The formingdentate gyrus (DG) of the hippocampus shows weak to moderate staining. E: Sagittal section of the developing cerebellum of an E l7 embryo (4V indicates the fourth ventricle). Moderate labeling is found in the differentiating zone, with some strongly labeled cells in the cortical transitory zone (CAI. F: Coronal section through the midbrain, pons, and medulla of an E l 9 embryo. Very intense labeling occurs in the oculomotor (31, trigeminal motor (5), and facial motor (7) nuclei. G In the vestibular ganglion (VeG) at E15, virtually all cells show very high levels of expression. The cochlear duct (CD) is indicated by arrows. Scale bar = 1,000 pm in A,F, 750 pm in B, 250 pm in C,D, 150 pm in E, 75 pm in G.

386 RJ. GIGER ET AL.
Fig. 6. Selected structures of the adult rat nervous system. SemaIIIi coll-1 mRNA expression was detected with a digoxigenin-labeled anti- sense riboprobe. A Detail of a coronal section showing the piriform cortex. SemaIIIicoll-1 transcripts are found only in layer 11; layers I and 111 are not labeled. B: Detail of acoronal section through the cerebellum showing moderate staining in Purkinje cells (Pc). C: Detail of a coronal
section through the ventral medulla oblongata showing moderate labeling of facial motoneurons. D Detail of a coronal section through the dorsal medulla oblongata. Hypoglossal motoneurons express se- maIIIicol1-1 transcripts. 4V indicates fourth ventricle. Scale bar = 250 pm in A, 150 km in B-D.
the anterior epithelium and the equatorial zone (Fig. 5A). During late embryogenesis and early postnatal period, expression gradually decreased as well in the ganglion cell layer of the peripheral retina and became undetectable by the end of the first postnatal week. Labeling in the lens declined somewhat more slowly and was lost by P12.
In E l 3 embryos, the neuroepithe- lium of the medulla and pontine region showed moderate expression. Very strong staining was found in the anlage of the vestibulocochlear ganglion (Fig. 5G) between E l 3 and E15. At E15, strong neuroepithelial staining was present in the region of the isthmus and posterior pons (Fig. 3C, arrow). The basal plate of the posterior pons and medulla, as well as the forming midbrain tegmentum, contained clusters of strongly labeled cells, which represent emerging
Mid- and hindbrain.
cranial motor nuclei (Fig. 3C). Less strongly labeled and more scattered cells were found in the zone of the develop ing medullar and pontine reticular formation (Fig. 3A,C). At E17/E19, staining in the pontine and medullar reticular formation had somewhat decreased compared to E 15, but motor nuclei, including the hypoglossal, facial (Fig. 5F), and motor trigeminal (Fig. 5F) nuclei, showed very intense staining comparable to spinal motor neurons. In the cerebel- lar anlage, staining was weak to moderate in differentiating zones, with some strongly labeled cells in the medial cortical transitory zone (Fig. 5E). The developing mesencephalon was virtually devoid of labeling, with the exception of the strongly labeled oculomotor (Fig. 5F) and trochlear nuclei. Labeling in the mid- and hindbrain gradually declined during postnatal life. The adult pattern of expression was

Fig. 7. Spinal cord and somites. SemaIIIicoll-1 mRNA expression was detected with a digoxigenin-labeled antisense rihoprohe. A: Parasag- ittal section through the trunk of an E l l embryo (rostral is to the right). Transcripts are seen in the posterior part of the sclerotome (arrowheads), whereas the anterior part is virtually devoid of staining. B: Transversal section of an E l 3 embryo at a thoracic level. In the spinal cord, staining is present in the basal plate neuroepithelium and in a number of cells in the ventral horn. No expression is found in the dorsal horn and in dorsal root ganglia (Drg). The mesenchyme ensheath- ing the notochord (arrow) is weakly labeled. C: Transverse section of an E l 5 thoracic spinal cord. Groups of very intensely stained cells are found in the lateral motor pool of the ventral horn. Individual strongly stained cells also appear in the medial part of the ventral horn and the intermediate grey matter. The dorsal horn is devoid of labeling. Strong staining is present in the basal plate neuroepithelium, whereas the floorplate (FP) and dorsal plate neuroepithelium are not labeled. D Detail of a transverse section of the spinal cord on E l 7 (midline is to the right). In the lateral ventral horn, clusters of motoneurons (LM) with maximal labeling are visible. Individual positive cells are scattered
through the medial ventral horn and the intermediate grey matter. E: Transverse section of a P1 cervical spinal cord. Clusters of motoneu- rons in the ventral horn show strong hybridization. In addition, weakly to moderately stained cells are dispersed across the ventral horn and intermediate grey matter. F: Transverse section of the upper thoracic spinal cord at P12. Arrow points to the central canal. SemaIII/coll-1 mRNA is expressed by a number of motoneurons located in the ventral horn (VH). Weak expression occurs in individual cells scattered through- out the ventral and intermediate zone. The dorsal horn (DH) shows no labeling. G Transverse section of adult spinal cord at the upper lumbar level. On the left side, the grey matter is outlined by a dashed line. SemaIII/coll-1-positive motoneurons are detected in the lateral (LM) and medial (MM) motor pool of theventral horn. The intermediate zone and the dorsal horn (DH) show no staining. H High-power photomicro- graph of a transverse section through an adult spinal cord at the upper thoracic level. Clusters of labeled motoneurons in lamina IX of the ventral horn are shown. Scale bar = 250 Fm in A, 150 Fm in B-D, 500 pm in E-G, 150 Fm in H.

Fig. 8. Double staining of semaIIUcol1-1 transcripts and growing nerve fiber tracts. In situ hybridization with a riboprobe specific for semaIII/coll-1 mRNA was followed by immunohistochemistry with monoclonal antibodies specific for TuJ l or B50iGAP43. SemaIIIicoll-1 transcripts were visualized using alkaline phosphatase (purple stain- ing), TuJ l and B50/GAP43 with peroxidase-catalyzed histochemistry (brown staining). A Sagittal section of a head at E l l . SemaIIIicoll-1 expression occurs in the mesenchyme lining the olfactory placode (OP) and in the telencephalic vesicle (TE) along its basal surface. The primary olfactory pathway, visualized with an anti-B50/GAP43 anti- body, extends from the OP towards the TE and does not grow behind the rim of semaIIIicol1-1 mRNA-expressing cells (arrows). B: High- power magnification of a parasagittal section through the forming olfactory bulb at E14.5. SemaIIIicoll-1 mRNA expression in the telencephalic neuroepithelium (TEN) is hardly detectable. B50iGAP43- positive axom of the primary olfactory pathway (POP) grow along the surface of the TEN; some of them have grown into the forming bulb (arrows). C: Coronal section through the developing olfactory bulb (OBI and epithelium at E17. S indicates the nasal septum. Strong expression of semaIIIicol1-1 is found in the mesenchyme lining the cribriform plate; weak expression is present in the olfactory bulb (OB) and olfactory epithelium (arrowhead). Axons of primary olfactory neurons stained with an anti-B50/GAP43 antibody penetrate the cribriform plate through the gaps in the semaIIIicol1-1-expressing mesenchyme
(arrows) and extend into the OB. D: Coronal section through the eye anlage at E15. SemaIIIicoll-1 mRNA expression is found in the ganglion cell layer (G), in the anterior part of the lens (Le), and to a lesser degree in the mesenchymal sheet of the eye anlage (Sl) and in the eyelid (asterisk). TuJ l immunoreactivity is found in the optic fiber layer and in the optic nerve. E: High-power magnification of an El5 central retina stained exclusively for semaIII/coll-1 transcripts. The ganglion cell layer (G) contains heavily stained cells. The mesenchymd sheet of the eye anlage (S1) shows weak labeling. F: Parasagittal section through the trunk of an E l l embryo (rostra1 is to the right). SemaIIIi coll-1 mRNA expression in the sclerotome of individual somites shows a gradient increasing from anterior to posterior. Arrowheads indicate the labeled posterior part. TuJ1-positive axons of primary sensory neurons are located exclusively in the anterior half of each successive somite (arrow). G Parasagittal section showing dorsal root ganglia (asterisk) of an E l 3 embryo. TuJ1-positive, highly fasciculated axons originate from the ganglia and are flanked by semaIIUcol1-1-expressing mesen- chyme ensheathing developing ribs and vertebrae (arrowheads). H: Sagittal section through the mesencephalic flexure (MeF) at E 15. SemaIIIicoll- 1-expressing cells of the oculomotor nucleus demarcate TuJ1-positive nerve fibers turning at the tip of the MeF. Fibers rarely cross the line defined by the semaIII/coll-1-positive cells (arrowheads). Scale bar = 250 pm in A,G, 50 pm in B,E, 150 bm in C,D,F, 75 pm in H.

SEMAPHORIN III/COLLAPSIN-1 EXPRESSION IN THE RAT 389
largely established by P12. Purkinje cells in the adult cerebellum retained moderate levels of staining (Fig. 6B). Moderate semaIII/coll-1 expression persisted also in the hypoglossal (Fig. 6D), ambiguus, facial (Fig. 6C), motor trigeminal, trochlear, and oculomotor nuclei.
Spinal cord and somites. At E l l , moderate to strong expression of semaIII/coll-1 mRNA was found in the dermamyotome and the posterior sclerotome of somites, whereas the anterior sclerotome was virtually devoid of staining (Fig. 7A). No signal was detected in the spinal cord at this developmental stage. Expression in the spinal cord was first detected on E l3 at the cervical and thoracic levels. Moderate labeling was present in the basal plate neuroepi- thelium and in clusters of moderately labeled cells in the forming ventral horn (Fig. 7B). DRG and the dorsal mantle zone were devoid of staining. A similar, but more intense, pattern of expression was observed in the E l5 spinal cord (Fig. 7 0 . In the lateral motor pool of the ventral horn, high expression was detected along the entire rostrocaudal length. The medial part of the ventral horn and the intermediate zone showed individual cells with very strong staining. Expression was fading in the intermediate grey matter, from ventral to dorsal, and was absent from the dorsal horn as well as from DRGs. High levels of expression were also found in the spinal neuroepithelium in a longitudinal stripe just ventral to the limiting sulcus. This stripe showed a sharp border against the unlabeled dorsal half of the neuroepithelium, whereas, ventrally, staining faded more gradually towards the floor plate, which itself was devoid of labeling. At E17, staining had disappeared from the spinal neuroepithelium, but the staining pattern in the grey matter remained very similar (Fig. 7D).
After birth, expression in the spinal cord became gradu- ally restricted to a subset of presumptive motoneurons in the ventral horn (Fig. 7E). At P12, strongly stained indi- vidual motoneurons were clearly visible in the ventral horn, and some weakly labeled cells were scattered throughout the intermediate grey matter (Fig. 7F). In the adult rat, staining was restricted to groups of motoneurons in the ventral horn. In the cervical spinal cord, clusters of labeled motoneurons were found mainly in layer IX (Molander et al., 1989) of the ventral horn (Fig. 7H). In the lumbar region, semaIII/coll-1 mRNA-positive motoneurons were located in the ventrolateral and ventromedial motor pool of the spinal cord (Fig. 7G).
Double staining for semaIII/coll-1 mRNA and growing nerve fiber tracts
During neuronal development, semaIII/coll- 1 might func- tion as a chemorepulsive molecule, preventing distinct populations of axons from innervating structures that ex- press and release this chemorepellent. As a first step in identify- ing fiber systems that could potentially be influenced by this chemorepellent, we combined in situ hybridization for semaIII/ coll-1 mRNA with immunohistochemistry for neuron-specific class I11 P-tubulin (TuJ1) and B50/GAP43, marker pro- teins for growing nerve fibers (Moody et al., 1987; Biffo et al., 1990). Assuming that growing axons of responsive neurons avoid or navigate around semaIII/coll-1-express- ing structures, we predicted a complementary distribution of semaIII/coll-1 mRNA and fiber tracts sensitive to this chemorepellent .
In the E l l rat embryo, the formation of the primary olfactory pathway (POP) has just begun (Santacana et al., 1992; Gong and Shipley, 19951, and the first primary
olfactory pioneer fibers are about to approach the telence- phalic vesicle. In double stained sections of E l l embryos, TuJ1- and B50/GAP43-positive primary olfactory pioneer fibers stalled at the periphery of the telencephalic vesicle where semaIIIicol1- 1 mRNA-expressing cells were found (Fig. 8A). Axons did not grow behind the line defined by the semaIII/coll-1-positive cells. Before making contact they turned abruptly by 90" and grew parallel to the surface of the telencephalic vesicle. At later stages (E14.5), semaIII/ coll-1 expression at the rostrobasal surface of the telencepha- lon decreased, and some axons began to penetrate deep into the weakly labeled neuroepithelium (Fig. 8B). Stained mesenchyme lining the cribriform plate was interrupted by semaIII/coll-1-negative areas. Double staining revealed that these areas were occupied by TuJ1-positive fila olfactoria entering the CNS (Fig. 7C).
Another example of complementary staining between semaIII/coll-1 and developing nerve tracts was observed in the somites and is illustrated in Figure 8F. Double staining in E l l embryos revealed that the anterior half of the sclerotome, which is penetrated by TuJ1-labeled axons, showed virtually no expression of semaIII/coll-1 mRNA. Expressing cells were restricted to the posterior part of the sclerotome, which is strictly avoided by exiting fibers. At E13, double staining for growing axons revealed highly fasciculated fibers projecting between semaIII/coll-1 mRNA- positive mesenchyme ensheathing vertebrae and ribs (Fig. 8G). No defasciculation of axon bundles was observed as long as fibers were flanked by labeled mesenchyme. In double stained sections of El5 embryos, a similar situation was observed in the mesencephalic flexure. TuJ1-positive fibers turned abruptly before they reached the tip of the flexure and did not penetrate the differentiating midbrain tegmentum that contained clusters of semaIII/coll-1- expressing motoneurons (Fig. 8H).
We also observed several instances in which fiber tracts were apparently not affected by semaIII/coll-1-expressing cells. For example, motoneurons in the brainstem and spinal cord and ganglion cells in the retina (Fig. 8D,E) were positive for semaIII/coll-1 transcripts and showed overlap- ping staining with growing nerve fiber tracts as visualized with the anti-TuJ1 antibody (Fig. 8D). The overlapping expression may indicate that retinal ganglion neurons, which express semaIII/coll-1 themselves, are not sensitive to the growth cone collapse activity of this chemorepellent. This would be consistent with cell culture studies, which showed that growth cones of these neurons are not sensi- tive to chicken collapsin-1 (Luo et al., 1993).
DISCUSSION To study the anatomical distribution of mammalian
semaIII/coll-1, we isolated a full-length cDNA clone from adult rat brain RNA by RT-PCR and used in situ hybridiza- tion techniques to identify expressing structures. The cDNA sequence of rat semaIII/coll-1 shows a high level of similarity to human semaphorin 111, mouse semaphorin D, and chicken collapsin-1, indicating that it encodes the homologue growth cone collapse protein of the rat. The specificity of the riboprobe used for in situ hybridization was demonstrated by Northern blot analysis. The antisense cRNA probe recognized three transcripts. Transcripts of variable size from this gene family have also been found for chicken collapsin-1 (Luo et al., 1993). Alternative splicing as suggested by Puschel et al. (1995) and/or the use of

390
alternative transcription initiation sites at the 5’ end or additional polyadenylation signals at the 3’ end are other possible explanations for the occurrence of the three differ- ent-sized transcripts.
Expression of semaIII/coll-1 is developmentally regulated
In situ hybridization in embryonic and neonatal rats revealed a widely distributed and dynamic expression pat- tern of semaIII/coll-1 mRNA. Stainingwas not restricted to the nervous system but was also observed in distinct epithelial and mesenchymal structures. Whereas mesenchy- ma1 staining was already evident on E l l and became most intense and widely distributed on E15, expression of se- maIII/coll-1 in the brain was most prominent around birth and declined more slowly than in nonneuronal structures.
In several areas of the CNS, expression of semaIII/coll-1 mRNA begins in precursors in the ventricular zone and seems to be maintained as neurons migrate into the mantle zone and begin to differentiate and extend axons. Neocorti- cal neurons originate from the proliferative neuroepithe- lium that lines the lateral ventricle and migrate along processes of radial glia towards the pial surface where they settle to form the layers of the cortex (Rakic, 1982). Moderate semaIIIicol1- 1 expression begins in the neuroepi- thelium of the prospective cortex before the onset of migration on E15. Expression of semaIII/coll-1 mRNA in the presumptive neocortex starts around E l 7 and contin- ues during cortical development in layers V and VI; by the end of the first postnatal week, a significant reduction of cortical semaIII /coll- 1 expression was observed. A similar observation was made for the spinal cord neuroepithelium, which generates spinal cord motor neurons that retain expression of semaIIIicol1-1 into adult life. In the E l 3 spinal cord, semaIIIicol1- 1 mRNA expression is restricted to the basal plate neuroepithelium and is not detectable in the floor plate. Thus, expression is confined to those parts of the spinal neuroepithelium that produce neurons that form the ventral horn, including motor neurons. Motor neurons in the lateral motor pool express the highest levels of semaIIIicol1-1 mRNA throughout development and re- main positive in the adult animal.
Comparison of the expression profile of chicken collapsin-1 and rat
semaIII/coll-1 mRNA A detailed study of collapsin-1 mRNA expression, focus-
ing on the embryonic development of the chick, has been recently published by Shepherd et al. (1996). Comparison with the expression profile of the embryonic rat revealed overlapping expression patterns, which further confirms that the cloned rat semaIIIicol1-1 cDNA is the species homologue of the chicken growth cone collapse molecule collapsin-1. However, we also found instances in which the two labeling patterns deviate. In the developing chicken olfactory bulb, collapsin- 1 message was strongly associated with the glomerular layer by stage 36 (El01 and later also with the developing granular cell layer stage 45 (E20). In the olfactory bulb of the embryonic rat, semaIII/coll-1 mRNA was weakly expressed in the mitral and granular cell layer at E l 7 and E19. After birth, expression became stronger in mitral cells and tufted cells, whereas no expres- sion was observed in the glomerular layer. In the developing neural retina, rat semaIII/coll- 1 mRNA was exclusively found in the ganglion cells layer, with the highest expres-
R.J. GIGER ET AL.
sion levels around E15. No staining in other layers of the retina was observed at any developmental stage. In the embryonic chicken at stage 41 (E15), strong in situ hybrid- ization signals were present within all three nuclear layers of the neural retina. At stage 24 (E4.5) of the developing chicken, collapsin-1 mRNA expression was found over a short time window in DRG (Shepherd et al., 1996). In cross sections of rat embryos at a comparable developmental stage (E11-E13), no labeled DRGs were found. Because semaIII/coll-1 mRNA shows a highly regulated and very dynamic expression pattern during embryogenesis, the chosen time frame of 2 day intervals may not have been fine enough to reveal short time expression in the DRG of the developing rat embryo.
Expression of semaIII/coll-1 persists in adulthood
Previous studies have suggested that semaIII/coll-1 exhib- its repulsive effects on restricted sets of neurons during axonal pathfinding (Luo et al., 1993; Messersmith et al., 1995; Puschel et al., 1995; Wright et al., 1995, Shepherd et al., 1996). The continued expression of semaIII/coll-1 in the adult CNS poses new questions about its function. Synaptic plasticity does not stop a t birth but continues throughout adulthood and allows an organism to respond to environ- mental changes by modifying the structure of dendrites and axonal arbors as well as the strength of synapses (see, e.g., Adams and Jones, 1982; Bailey and Chen, 1989). Because semaIII/coll-1 is an extremely potent chemorepellent that is active at concentrations around 10 pM (Luo et al., 19931, the observed expression in adult rat CNS invites the speculation that this protein may impose limits on the remodelling of synaptic connections. In the piriform cortex, for example, the persistent expression of semaIII/coll-1 in layer I1 may prevent aberrant ingrowth of projections from layer I or 111, where semaIII/coll-1 mRNA is absent, thereby potentially regulating remodelling of neuronal connections during adulthood. Thus, semaIII/coll- 1 could contribute to the maintenance of the integrity of existing neuronal networks in adult animals by restricting the formation of new connections in those areas where its expression is maintained during adulthood.
Persistent expression of semaIII/coll-1 in the adult CNS may inhibit regeneration of semaIII/coll-1-sensitive neu- rons after lesion or trauma. Together with other members of the semaphorin gene family, which exhibit different expression patterns (Luo et al., 1995; Messersmith et al., 1995) and which probably act on different populations of neurons, semaIII/coll-1 may form a potential group of inhibitors of regeneration in the CNS. In this context it is of interest that semaIII/coll-1 mRNA is absent from the adult PNS.
SemaIII/coll-1 expression in somites shows an anterior-posterior gradient
Double staining for semaIII/coll-1 transcripts and grow- ing spinal nerves a t E l l revealed a remarkable complemen- tarity. Whereas B50/GAP43- and TuJ1-stained axons were found in the anterior half of sclerotome, semaIII/coll-1- expressing cells were confined to the posterior half. Previ- ous studies suggested that the segmented pattern of periph- eral spinal nerves is orchestrated by segmentation in the paraxial (somite) mesoderm lining the neural tube (Det- wiler, 1934). Furthermore, in higher vertebrate embryos, segmentation is determined by the subdivision of each

SEMAPHORIN III/COLLAl'SIN-1 EXPRESSION IN THE RAT 391
somite into anterior (cranial) and posterior (caudal) halves (Keynes and Stern, 1984, 1985; Tonsey, 1988). At each segmental level, sensory axons sprout from DRG and motoneurons grow from the ventral neural tube exclusively within the anterior half of the sclerotome. When spinal nerve axons contact the proximal side of the somite, they have developed into the outer dermatome-myotome (respec- tively, prospective dermis and skeletal muscle) and inner sclerotome (prospective vertebral column). Inhibitory cues for growing axons located in the posterior sclerotome have previously been reported by Davies et al. (1990). Further- more, growth cones of cutaneous afferents of DRG neurons have been shown to be sensitive to semaIII/coll-1 (Luo et al., 1993; Messersmith et al., 1995). An anterior-posterior gradient of semaIII/coll-1 expression might offer an expla- nation for the strong preference of spinal nerve axons to grow into the anterior half of the sclerotome. Our observa- tions are consistent with the study by Wright et al. (1995) in which DiI labeling was employed to visualize the pathway of primary sensory axons.
Later in development, semaIII /coll- 1 expression was found in mesenchyme ensheathing vertebrae, ribs, and a number of other structures that demarcate the pathway of sensory spinal nerve axons as they grow towards their targets in the periphery. DRG neurons begin to extend axons on E l l and by E15/E16 the first reflex responses to cutaneous stimulation can be obtained (Vaughn and Grie- shaber, 19731, indicating that growing axons have made contact with their target tissue. The expression pattern of semaIII/coll-1 in nontarget structures demarcating the pathway of sensory axons is most remarkable and wide- spread during the same time, starting around E l l in somites and disappearing rapidly from labeled mesenchyme after E15. The temporal correlation of semaIII/coll-1 expres- sion in the periphery and the outgrowth of sensory axons suggests that nontarget structures might be deflected from innervation by expression and secretion of this chemorepel- lent. It will be interesting to see whether other members of the semaphorin gene family have similar effects on popula- tions of neurons that do not respond to semaIIUcol1-1.
SemaIII/coll-1 may be involved in the induction of the olfactory bulb and the
formation of olfactory projections Olfactory bulb development is dependent on the olfactory
placode (Stout and Graziadei, 1980; Byrd and Burd, 1991; Gong and Shipley, 1995). In the developing olfactory sys- tem, axons begin to grow at E l l from the olfactory placode towards the telencephalic vesicle. Upon reaching the periph- ery of the neuroepithelium, they turn abruptly by 90" and grow parallel to the surface of the telencephalic vesicle (Gong and Shipley, 1995). At the same time, a rim of semaIII/coll- 1-expressing cells is present at the periphery of the telencephalic vesicle in the marginal layer opposite the olfactory pit. Immunostaining of the forming pri- mary olfactory tract with a B50iGAP43 antibody showed that this fiber tract does not penetrate the rim of semaIII/ coll-1-expressing cells at the surface of the telencephalic neuroepithelium opposing the olfactory placode. During the next days, the expression of semaIII/coll-1-positive cells in the telencephalic rim decreases, and, at E14-El5, POP axons start to extend and penetrate deep into the telence- phalic neuroepithelium. These axons, which probably in- duce the formation of the olfactory bulb, have been de- scribed previously (Santacana et al., 1992; Gong and Shipley,
1995). The time of bulb induction correlates precisely with a reduction of the semaIII/coll-1 level at the surface of the telencephalic vesicle, which would subsequently allow axons of the POP to cross the marginal layer of the telencephalic vesicle and grow deep into the neuroepithelium. A predic- tion that follows from these observations is that down- regulation of semaIII/coll-1 expression may be a crucial event for the induction of the olfactory bulb by primary olfactory axons. Expression of semaIII/coll-1 is also present in the mesenchyme lining the cribriform plate. Interest- ingly, double staining of semaIII/coll-1 mRNA and T u J l in the olfactory system revealed that the fila olfactoria strictly use the semaIII/coll-1-free gaps in the developing cribri- form plate to grow towards the developing olfactory bulb. Mitral cells are the main target neurons of primary olfac- tory neurons, and they project to the piriform cortex, the amygdala, and the entorhinal cortex (Brunjes and Frazier, 1986). Interestingly, these projection areas of the mitral cells strongly express the semaIII/coll-1 message during development, suggesting that semaIII/coll-1 plays a role in patterning projections to the olfactory cortex by preventing other afferents from entering these areas. Recently, Shep- herd et al. (1996) showed that olfactory bulb neurons are not sensitive to semaIII/coll-1 in vitro, which is in line with this hypothesis. In the adult bulb, mitral cells continue to express semaIII/coll-1 mRNA. This may suggest that mi- tral cell-derived semaIII/coll-l may function as a stop signal for new primary olfactory nerve fibers that grow into the olfactory bulb glomeruli throughout adulthood.
The interpretation of the double staining experiment has certain limitations, because semaIII/coll- 1 expression was detected at the RNA level by in situ hybridization and not at the protein level by immunostaining. For example, motoneurons in the spinal cord may express semaIII/coll-1 on their cell soma or transport the protein to axonal terminals in the periphery and/or dendrites in the spinal cord. For the nonneuronal cells, which express semaIII/ coll-1 mRNA at the periphery of the telencephalic vesicles, the mesenchymal lining of the cribriform plate, and the posterior part of sclerotome, the localization of the encoded chemorepellent is easier to predict: Although semaIII/ coll-1 can be secreted, studies on brain membrane prepara- tions indicate that a significant proportion of the molecules remains associated with the cell membrane (Luo et al., 1993).
CONCLUSIONS This study provides the first complete mapping of the
spatial and temporal expression of semaIII/coll-1 in embry- onic and neonatal rats and documents the continued expres- sion of this growth cone collapse protein in subsets of neurons in the adult mammalian CNS. The observed dynamic expression pattern of semaIII/coll-1 mRNA and its striking complementary relationship to a number of growing fiber tracts suggests an important role for this chemorepellent in growth cone guidance and nerve tract formation.
ACKNOWLEDGMENTS We thank Corbert van Eden, Robert E. Baker, Guus
Wolswijk, Dominique de Kleijn, and Mercedes McClean for their valuable comments on the contents and for proofread- ing the manuscript. The monoclonal anti-TuJ1 antibody

392
was kindly provided by Dr. A. Frankfurter (University of Virginia), and B50iGAP43 antibodies were a kind gift of Dr. A.B. Oestreicher (University of Utrecht). For technical assistance, we thank Roman Hofer (animal work) and Gerben van der Meulen (images). This work was supported by NWO-GMW Pioneer grant 030-94-142.
R.J. GIGER E T AL.
LITERATURE CITED Adams, I., and D.G. Jones (1982) Quantitative ultrastructural changes in rat
cortical synapses during early-, mid-, and late-adulthood. Brain Res. 239:349-363.
Bailey, C.H., and M. Chen (1989) Structural plasticity at identified synapses during long term memory in Aplysia. J. Neurobiol. 20:356-372.
Biffo, S., J. Verhaagen, L.H. Schrama, P. Schotman, W. Danho, and F. Margolis (1990) B-50iGAP43 expression correlates with processes out- growth in the embryonic mouse nervous system. Eur. J. Neurosci. 2487-499.
Brunjes, P.C., and L.L. Frazier (1986) Maturation and plasticity in the olfactory system ofvertebrates. Brain Res. Rev. 11.1-45.
Byrd, C.A., and G.D. Burd (1991) Development of the olfactory bulb in the clawed frog, Xenopus laeuis: A morphological and quantitative analysis. J. Comp. Neurol. 314:79-90.
Culotti, J.G., and A.L. Kolodkin (1996) Functions of netrins and semaphor- ins in axon guidance. Curr. Opin. Neurobiol. 6:81-88.
Dani, J.W., D.M. Armstrong, and L.I. Benowitz (1991) Mapping the develop- ment of the rat brain by GAP-43 immunocytochemistry. Neuroscience 40277-287.
Davies, J.A., G.M. Cook, C.D. Stern, and R.J. Keynes (1990) Isolation from chick somites of a glycoprotein fraction that causes collapse of dorsal root ganglion growth cones. Neuron 2.11-20.
Detwiler, S.R. (1934) An experimental study of spinal nerve segmentation in Ambystoma with reference to the plurisegmental contribution to the brachial plexus. J. Exp. Zool. 67:395441.
Dodd, J., and A. Schuchardt (1995) Axon guidance: A compelling case for repelling growth cones. Cell 8k471-474.
Easter, S.S., S.L. Ross, and A. Frankfurter (1993) Initial tract formation in the mouse brain. J. Neurosci. I3:285-299.
Giger, R.J., L. Vogt, R.A. Zuellig, C. Rader, A. Henehan-Beatty, D.P. Wolfer, and P. Sonderegger (1995) The gene of chicken axonin-1: Complete structure and analysis of the promoter. Eur. J. Biochem. 227:617-628.
Gong, Q., and M.T. Shipley (1995) Evidence that pioneer olfactory axons regulate telencephalon cell cycle kinetics to induce the formation of the olfactory bulb. Neuron 14:91-101.
Goodman, C.S., and C.J. Shatz (1993) Developmental mechanisms that generate precise patterns of neural connectivity. Cell 72iNeuron 10 (Suppl.):77-98.
Hebel, R., and M.W. Stromberg (1986) Anatomy and embryology of the laboratory rat. Guenzburg, Germany: BioMed Verlag Woerthsee, Appel- Druck Donau-Verlag GmbH.
Inagaki, S., T. Furuyama, and Y. Iwahashi (1995) Identification of a member of mouse semaphorin family. FEBS Lett. 370.269-272.
Kennedy, T.E., T. Serafini, R.J. de la Torre, and M. Tessier-Lavigne (1994) Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell 78.425435.
Keynes, R.J., and C.D. Stern (1984) Segmentation in the vertebrate nervous system. Nature 310:786-789.
Keynes, R.J., and C.D. Stern (1985) Segmentation and neuronal develop- ment in vertebrates. Trends Neurosci. 8220-223.
Kolodkin, A.L. (1996) Semaphorins: Mediators of repulsive growth cone guidance. Trend. Cell Biol. 6:15-22.
Kolodkin, A.L., D.J. Matthes, T.P. O’Connor, N.H. Patel, A. Admon, D. Bentley, and C.S. Goodman (1992) Fasciclin IV: Sequence, expression, and function during growth cone guidance in the grasshopper embryo. Neuron 91831-845.
Kolodkin, A.L., D.J. Matthes, and C.S. Goodman (1993) The semaphorin genes encode a family of transmembrane and secreted growth cone guidance molecules. Cell 75:1389-1399.
Kuhn, T.B., M.F. Schmidt, and S.B. Kater (1995) Laminin and fibronectin guideposts signal sustained but opposite effects to passing growth cones. Neuron 14275-285.
Luo, Y., D. Raible, and J.A. Raper (1993) Collapsin: A protein in brain that induces the collapse and paralysis of neuronal growth cones. Cell 752 17-227.
Luo, Y., I. Shepherd, J. Li, M.J. Renzi, S. Chang, and J.A. Raper (1995) A family of molecules related to collapsin in the embryonic chick nervous system. Neuron 14: 113 1-1 140.
Matthes, D.J., H. Sink, A.L. Kolodkin, and C.S. Goodman (1995) Sema- phorin I1 can function as a selective inhibitor of specific synaptic arborizations. Cell 81 :631-639.
Messersmith, E.K., D.E. Leonardo, C.J. Shatz, M. Tessier-Lavigne, C.S. Goodman, and A.L. Kolodkin (1995) Semaphorin 111 can function as a selective chemorepellent to pattern sensory projections in the spinal cord. Neuron 14:949-959.
Molander C., Q. Xu, C. Rivero-Melian, and G. Grant (1989) Cytoarchitec- tonic organization of the spinal cord in the rat: 11. The cervical and upper thoracic cord. J. Comp. Neurol. 289.375-385.
Moody, S.A., M.S. Quigg, and A. Frankfurter (1987) Development of the peripheral trigeminal system in the chick revealed by an isotype-specific anti-beta-tubulin monoclonal antibody. J. Comp. Neurol. 279.567-580.
Puschel, A.W., R.H. Adams, and H. Betz (1995) Murine semaphorin Dicollapsin is a member of a diverse gene family and creates domains inhibitory for axonal extension. Neuron 14:941-948.
Rakic, P. (1982) Early developmental events: Cell lineages, acquisition of neuronal positions, and areal and laminar development. In P. Rakic and P.S. Goldman-Rakic (eds): Development and Modifiability of the Cere- bral Cortex. (Neurosci. Res. Progr. Bull., Vol. 20). London: MIT Press, pp. 43-51,
Santacana, M., M. Heredea, and F. Valverde (1992) Transient pattern of exuberant projections of olfactory axons during development in the rac. Dev. Brain Res. 70:213-222.
Schaeren-Wiemers, N., and A. Gerfin-Moser (1993) A single protocol to detect transcripts of various types and expression levels in neural tissue and cultured cells: In situ hybridization using digoxigenin-labeled cRN.4 probes. Histochemistry 100:431440.
Schwab, M.E., J.P. Kapfhammer, and C.E. Bandtlow (1993) Inhibitors of neurite growth. Annu. Rev. Neurosci. 16.565-595.
Serafini, T., T.E. Kennedy, M.J. Galko, C. Mirzayan, T.M. Jessell, and hl. Tessier-Lavigne (1994) The netrins define a family of axon outgrowth- promoting proteins homologous to C. elegans UNC-6. Cell 78:409-424.
Shepherd, I., Y. Luo, J.A. Raper, and S. Chang (1996) The distribution of collapsin-1 mRNA in the developing chicken nervous system. Dev. Bid. 173.185-199.
Stout, R.P., and P.P.C. Graziadei (1980) Influence of the olfactoryplacode on the development of the brain in Xenopus Zueuis (Daudini. I. Axonal growth and connections of the transplanted olfactory placode. Neurosci- ence 52175-2186.
Tessier-Lavigne, M. (1994) Axon guidance by diffusible repellents and attractants. Curr. Opin. Genet. Dev. 4596-601.
Tonsey, K.W. (1988) Proximal tissues and patterned neurite outgrowth at the lumbosacral level of the chick embryo: Partial and complete deletion of the somite. Dev. Biol. 127%-286.
Vaughn, J.E., and J.A. Grieshaber (1973) A morphological investigation of an early reflex pathway in developing rat spinal cord. J. Comp. Neurol. 148:177-209.
Wright, D.E., F.A. White, R.W. Gerfen, I. Silos-Santiago, and W.D. Snider (1995) The guidance molecule semaphorin 111 is expressed in regions of spinal cord and periphery avoided by growing sensory axons. J. Comp. Neurol. 361.321-333.