Embed Size (px)
Citation preview

I lMOUNIZATON AGAINST BRUCELLA INFECTON
IX. THE RESPONSE OF THE GuNA PIG Tro T IMZuruzN STRm(REv.1) OF BRucUA mELrrENsis
JOAN MCCAMIEH, PHD.,* D SAIFOD S. EEXG, PH.D.
From the Dqrtment of Bacteiology, University of California, Berkley, Calif.
The great susceptibility of the guinea pig to infection by species ofBrucela has been utilized traditionally to test the virulence of a givenstrain and to evaluate the efficacy of various means of immunization.Very little has been added to the description of the histologic events inthe guinea pig following inoculation with virulent B. abortus since theclassic paper by Fabyan.1 He descnbed the intracellular location of theorganisms, the racteristic reticuloendothelial response, the succes-sion of cell types, the formation of typical granulomatous lesions, andthe occasional occurrence of giant cells or microscopic abscesses. Thesimilarity of the lesions provoked by B. melitensis and by B. abortus wasrecognized by Meyer, Shaw, and Fleischner.2 Recent work has correlatedthe microscopic lesions with the grosser signs of disease3 and with thefunction of specific organs in the infection.4 The process, causes, andeffects of granuloma formation have also received special attention.'6-Investigations of a histologic nature have been made on two vaccinestrains, BA i9 7* and SMd.10The relatively poor immunogenicity of strain SMd of B. melitensis
prompted studies on the derived mutant strain, B. mditensis Rev. i. Theeffectiveness of the Rev. i mutant as a vaccine has been shown in miceand guinea pigs,11 pregnant and nonpregnant goats,"-" ewes,17 andmonkeys.18 There is now sufficient information available on the highimmunogenicity of B. mditensis Rev. i to warrant its consideration foremployment on a wider scale. It is desired to record, therefore, the de-tailed study made of the guinea pig's response to Rev. i as a guide indefining the extent and limits of the virulence of the strain, thereby sup-plementing the earlier published studies on Rev. i (vide supra).
MATERIALS AND METHODSGuinea Pigs
Female animal of the Hartley strain, about 3 months old at the time of injection,were used.
This work was supported by Grant (E22) from the National Institutes of Health.Accepted for publiation, July 31, I96I.
* Present address: Department of Bacteriology, University of Kansas Lawrence, Kans
77

MC CAMISH AND ELBERG
BacteriaA smooth clone of BruceUa melitensis Rev. i, cultured on Albimi Brucella agar,*
was harvested in sterile saline. The viable count was determined.
Inoculaion of AnimalsGroup I, 44 guinea pigs, received io bacterial cells.Group II, 43 guinea pigs, received ioo bacterial cells.Group m, 44 guinea pigs, received i,ooo bacterial cells.Controls, I4 guinea pigs, were not inoculated.All inoculums were contained in 0.2 ml. of o.85 per cent saline. The injections were
given subcutaneously in the lower right quadrant of the abdomen, approximatelyone-half inch above the position of the right inguinl lymph node.
Sacrifice of AnimalsEach guinea pig, at the time of sacrifice, received an intraperitoneal injection of
0.5 ml. of Beuthanasia.t Just before the death of the animal, 2 ml. of blood was with-drawn from the heart for serum agglutination test and culture. A portion of the skinoverlying the site of injection as well as any adherent muscle, the spleen, the rightsuperficial inguinal lymph node(s), and the left axillary lymph node(s) were removedaseptically. Approximately one half of each tissue or organ was cultured for thepresence of brucellae, and the remaining portion was placed in Technicon fixative4Two guinea pigs from each of the 3 test groups were sacrificed at the following
times after the inoculation: o, 6, and I2 hours; 2, 4, 7, 9, II, 14, i6, i8, 2I, 23 and25 days; 4, 5, 6, 7, 8, 9, I2, and i6 weeks.At each of the time periods o hours and 2, 4, 6, 8, 12 and I6 weeks after the
inoculation of the test animals, 2 of the control guinea pigs were also sacrificed. Cul-tures were not always made of the tissues in the normal animals, but all the otherspecimens were obtained from them in the same way as from the test animals.
Tissue SectionsThe portions of tissue which had been fixed in Technicon fixative were dehydrated
and cleared by passage through Technicon dehydrants and embedded in paraffin.Sections were cut at 6 p on a rotary microtome and fixed to slides by floating froma warmed dilute gelatin-K2Cr307 solution. For histologic examiation, they werestained with hematoxylin and eosin and with Giemsa stain. Representative slideswere also stained with Ehrlich's plasma cell stain.
Agglutination TestsDilutions of serum in o.85 per cent saline were mixed with equal amounts of the
standard antigen, Agricultural Research Service, United States Department of Agri-culture. The test mixtures were incubated 24 hours at 370 C. and then were allowedto stand at room temperature for 48 hours. The titer recorded represented the highestdilution of serum in which visible agglutination occurred.
RESULTSSerum Agglutination Titers
The time of the appearance of agglutinins in the serum of the cardiacblood was to some extent dependent on the number of organisms in the
* Albimi Laboratories, Inc, Brooklyn, N.Y.t H. C. Burns Company, Inc, Oakland, Calif.$ Technicon Chemical Company, Inc., Chauncey, N.Y.
78 Vol. 4o, No. z
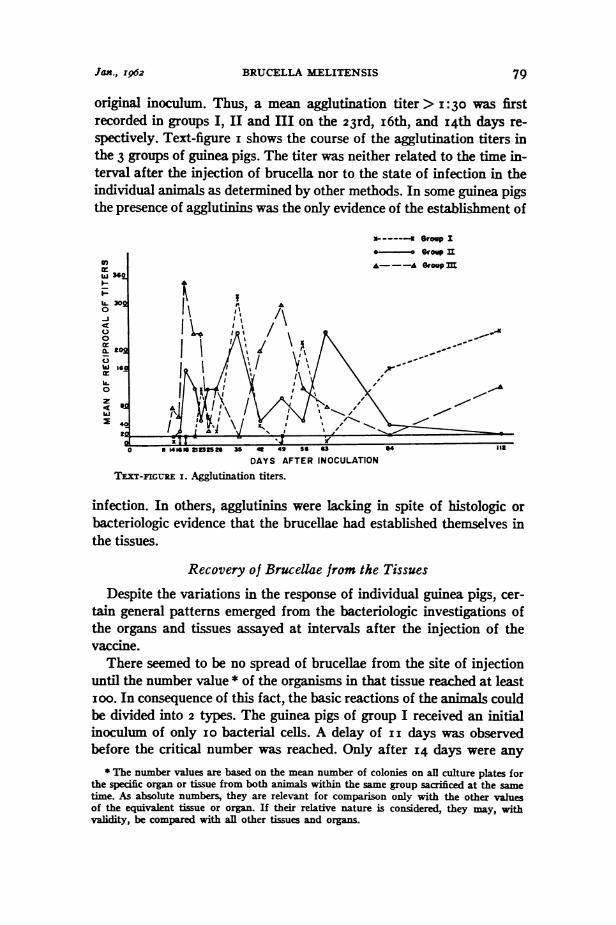
Jan., 1962 BRUCELLA MELITENSIS 79
original inoculum. Thus, a mean agglutination titer > I:30 was firstrecorded in groups I, II and III on the 23rd, i6th, and 14th days re-spectively. Text-figure i shows the course of the agglutination titers inthe 3 groups of guinea pigs. The titer was neither related to the time in-terval after the injection of brucella nor to the state of infection in theindividual animals as determined by other methods. In some guinea pigsthe presence of agglutinins was the only evidence of the establishment of
-------E GBrp I
a)
wI-
LL0f-j
0c-J
4U0.0
IL.0
zhii
DAYS AFTER INOCULATION
TEXT-FIGURE I. Agglutination titers.
infection. In others, agglutinins were lacking in spite of histologic orbacteriologic evidence that the brucellae had established themselves inthe tissues.
Recovery of Brucellae from the Tissues
Despite the variations in the response of individual guinea pigs, cer-tain general patterns emerged from the bacteriologic investigations ofthe organs and tissues assayed at intervals after the injection of thevaccine.
There seemed to be no spread of brucellae from the site of injectionuntil the number value * of the organisms in that tissue reached at leastioo. In consequence of this fact, the basic reactions of the animals couldbe divided into 2 types. The guinea pigs of group I received an initialinoculum of only io bacterial cells. A delay of ii days was observedbefore the critical number was reached. Only after I4 days were any
* The number values are based on the mean number of colonies on all culture plates forthe spedfic organ or issue from both animal within the same group sacrificed at the sametime. As absolute numbers, they are relevant for comparison only with the other valuesof the equivalent issue or organ. If their relative nature is considered, they may, withvalidity, be compared with all other tissues and organs.
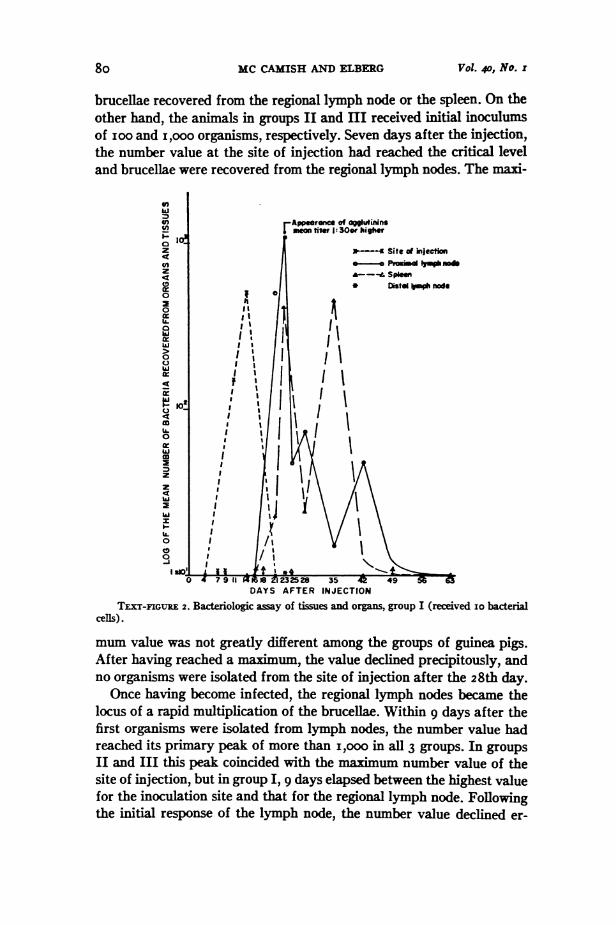
MC CAMISH AND FTBRERG Vol. 40, No. r
brucellae recovered from the regional lymph node or the spleen. On theother hand, the animals in groups II and HI received initial inoculimsof ioo and Ii,ooo organisms, respectively. Seven days after the injection,the number value at the site of injection had reached the critical leveland brucellae were recovered from the regional lymph nodes. The maxi-
kJ
U,
a
CA
0
0
"a
Li.
0
at
2L
4t
LA.01
DAYS AFTER INJECTION
ITXT-FIGURE 2. Bacteriologic assay of tissues and organs, group I (received io bacterialcels).
mum value was not greatly different among the groups of guinea pigs.After having reached a maximum, the value declined precipitously, andno organisms were isolated from the site of injection after the 28th day.Once having become infected, the regional lymph nodes became the
locus of a rapid multiplication of the brucellae. Within 9 days after thefirst organisms were isolated from lymph nodes, the number value hadreached its primary peak of more than I,oo in all 3 groups. In groups
II and III this peak coincided with the maximum number value of thesite of injection, but in group 1,9 days elapsed between the highest valuefor the inoculation site and that for the regional lymph node. Followingthe initial response of the lymph node, the number value declined er-
8o
is
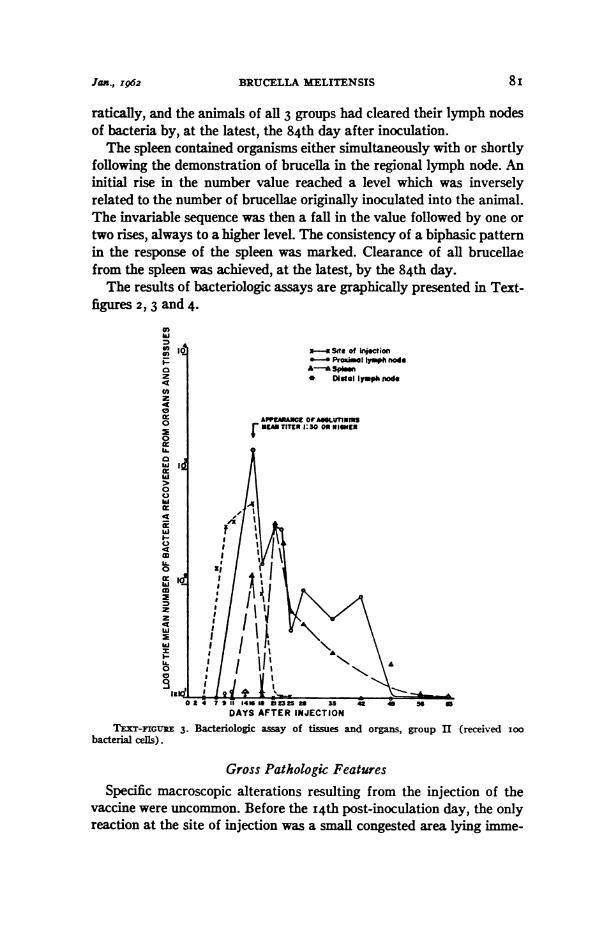
Jax., I962 BRUCELLA MELITENSIS 8i
ratically, and the animals of all 3 groups had cleared their lymph nodesof bacteria by, at the latest, the 84th day after inoculation.The spleen contained organisms either simultaneously with or shortly
following the demonstration of brucella in the regional lymph node. Aninitial rise in the number value reached a level which was inverselyrelated to the number of brucellae originally inoculated into the animal.The invariable sequence was then a fall in the value followed by one ortwo rises, always to a higher level. The consistency of a biphasic patternin the response of the spleen was marked. Clearance of all brucellaefrom the spleen was achieved, at the latest, by the 84th day.The results of bacteriologic assays are graphically presented in Text-
figures 2,3 and 4.0)ki
U'02P
z4t
0
0U.ai4
02C.hi02
4c
Ii.C
-onlymphi node
meph nodeo
A
DAYS AFTER INJECTION
TEXT-FIGUIE 3. Bacteriologic assay of tissues andbacterial cells).
organs, group II (received Ioo
Gross Pathologic FeaturesSpecific macroscopic alterations resulting from the injection of the
vaccine were uncommon. Before the I4th post-inoculation day, the onlyreaction at the site of injection was a small congested area lying imme-
I
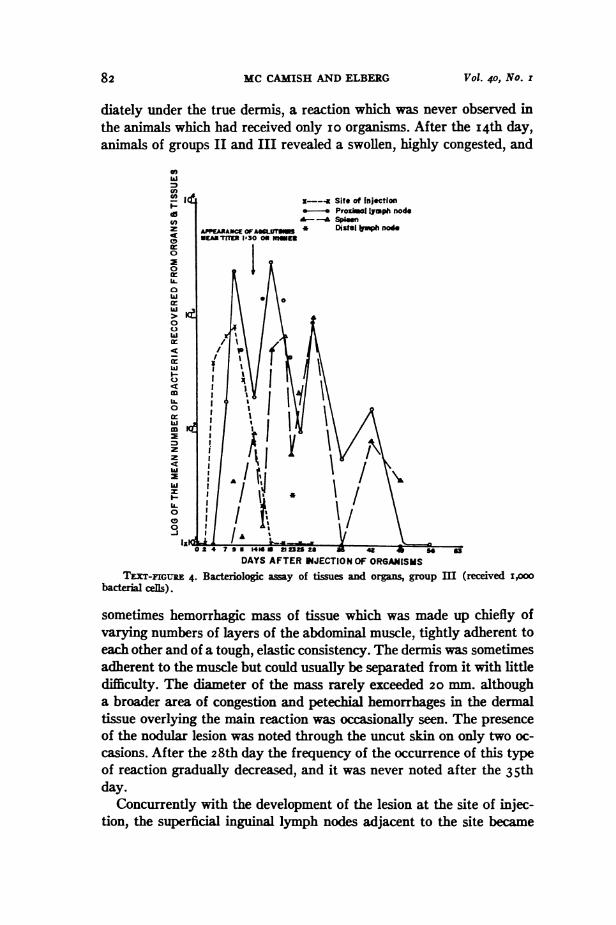
82 MC CAMISH AND ELBERG Vol. 40, No. I
diately under the true dermis, a reaction which was never observed inthe animals which had received only io organisms. After the I4th day,animals of groups II and III revealed a swollen, highly congested, and
@7
hia,7
UI-z
01
>k
0
0
hl
I-cr
0
0
z
hi-
I.-
m
IL.0
0
-JlIC,
X--- Site of Injection- v ProawI lymph node--A Spin
APPEARA*= OFAx UT * DistaV1ph noeEASTITEr 1'30 OR Mteu
DAYS AFTER INJECTION OF ORGANISMS
TEXT-FIGURE 4. Bacteriologic away of tissues and organs, group m (received i,ooobaceria cefs).
sometimes hemorrhagic mass of tissue which was made up chiefly ofvarying numbers of layers of the abdominal muscle, tightly adherent toeach other and of a tough, elastic consistency. The dermis was sometimesadherent to the muscle but could usually be separated from it with littledifficulty. The diameter of the mass rarely exceeded 20 mm. althougha broader area of congestion and petechial hemorrhages in the dermaltissue overlying the main reaction was occasionally seen. The presenceof the nodular lesion was noted through the uncut skin on only two oc-casions. After the 28th day the frequency of the occurrence of this typeof reaction gradually decreased, and it was never noted after the 35thday.
Concurrently with the development of the lesion at the site of injec-tion, the superficial inguinal lymph nodes adjacent to the site became
0

BRUCELLA MELITENSIS
more prominent. They enlarged to a diameter of as much as I2 mm. andassumed a dull gray color. The supporting tissue and the surroundingfat showed gross signs of congestion. The hypertrophy of the regionallymph nodes did not regress with the general decline of the infection.Many of them were still very prominent at the end of the experiment,I I2 days after the initial inoculation.The spleens of some of the guinea pigs were enlarged, in some cases
to as much as i 12 times their normal size. Enlarged spleens were seenat random times in both the inoculated and the uninoculated animals.
In a great majority of the animals, focal lesions were seen in thelungs, visible both through the uncut pleura and throughout the paren-chymal tissue of the sectioned organ. They were found in both normaland infected guinea pigs.
Histologic ObservationsWith the exception of the reaction at the site of injection, the micro-
scopic alterations which occurred in guinea pigs in response to the in-jection of living cells of the Rev. i strain of B. melitensis did not differqualitatively among the 3 groups of animals, despite the differences inthe number of organisms originally inoculated. The quantitative differ-ences were only matters of frequency and timing; the maximum possi-ble intensity of any individual reaction was equivalent whether the ini-tial inoculum consisted of IO, ioo, or i,ooo bacterial cells.
Histologic Features of the Infection. The guinea pig's response to theinoculation of IO brucella cells (Rev. i vaccine strain, B. melitensis)consisted almost entirely of a proliferative reaction affecting cells of thereticuloendothelial system. Fibroblast activity, with minor perivascularreticular cell hyperplasia, constituted the major alterations seen at thesite of injection. These reached their climax by the I4th day, by whichtime a diffuse proliferation of the reticular elements of the regionallymph node was also typically seen. In the lymph node the stages ofthe intensification of the response followed in rapid succession so thatby the 23rd day typical granulomatous nodules, containing various de-grees of small cell infiltration and necrosis, were always found. An oc-casional nodular focus of epithelioid cells could be seen in the spleenby the 28th day. In the following period of 3 weeks the lesions remainedmore or less unchanged in all 3 tissues. Sections exaiined on the 56thday indicated an apparent increase in the frequency and severity of thereaction in the lymph node and spleen, but thereafter all tissues appearednormal or showed definite signs of resolution of those lesions present.The injection of ioo bacterial cells was followed within 4 days by
hyperplasia of the reticular elements both at the site of injection and
83Jan., I962

MC CAMISH AND ELBERG
in the regional lymph node. In addition, a granulomatous lesion wasdeveloping in the connective tissue at the site of injection. By the i6thday a fully developed foreign body reaction was present at the inocula-tion site. The reaction remained at its maximum during the succeeding5 days and then was rapidly resolved, leaving, by the 35th day, onlyresidual loose fibrous tissue and an occasional localized cell focus. Thelymph node response reached its maximum by the 23rd day, after whichoccurred a slow, steady regression of the abnormalities. Specific lesionswere seen in the spleen as early as the I4th day, developed maximallyby the 35th day, and then diminished rapidly, not to be seen after the49th day.The order of events at the site of the inoculation of i,ooo bacteria
paralleled very closely those of group II. The height of the reaction wasseveral days earlier, starting on the eleventh day, as was the beginningof resolution, starting on the i6th day. The remainder of the course ofresolution was nearly identical with that of group II. The spleen andlymph node reactions also approximated those of the previous group inthe early period of the experiment. After the 23rd day, however, theregional lymph nodes maintained the signs of most intense reactivitythrough the 56th day, and thereafter the recovery of the node was veryrapid. The time sequence of the alterations in the spleens of group IIIwas the same as in group II. The severity of the maximum reaction wasgreater but, as in group II, was observed on the 35th day.
In order better to visualize the course of the infection as it was re-flected in histologic changes, a scheme of evaluating the lesions wasdevised. For this purpose different stages in their development were as-signed cardinal numbers, arbitrarily, but yet with a presumed relation-ship to their actual degree of intensity of response. Since the criteriadiffered at least slightly in the various tissues, separate value scales weredeveloped for each one. They are the following:
Site of injection:minor vascular nges Iedema, congestion 2minor cellular infiltration 3cellular foci (without connective tissue reaction) 4minor connective tissue reaction 5major connective tissue reaction 6localization of lesions or scar 4residuum with cellular foci 3residuum without cellular foci 2
84 Vol. 4o, No. z

BRUCELLA MELITENSIS
Regional lymph node:diffuse RE cell proliferation I"streaming" RE cell proliferation 2epithelloid cell nodules 3cellular infiltration of nodules 4fusion or central necrosis 5abscess formation or extreme granuloma reaction 6lymphocyte replacement 4
Spleen:epithelioid cell nest 2epithelloid cell nodule 3cellular infiltration of nodule 4central necrosis 5hemorrhage 6
Liver:
large or mixed focus Ifocus with epithelioid cells 2
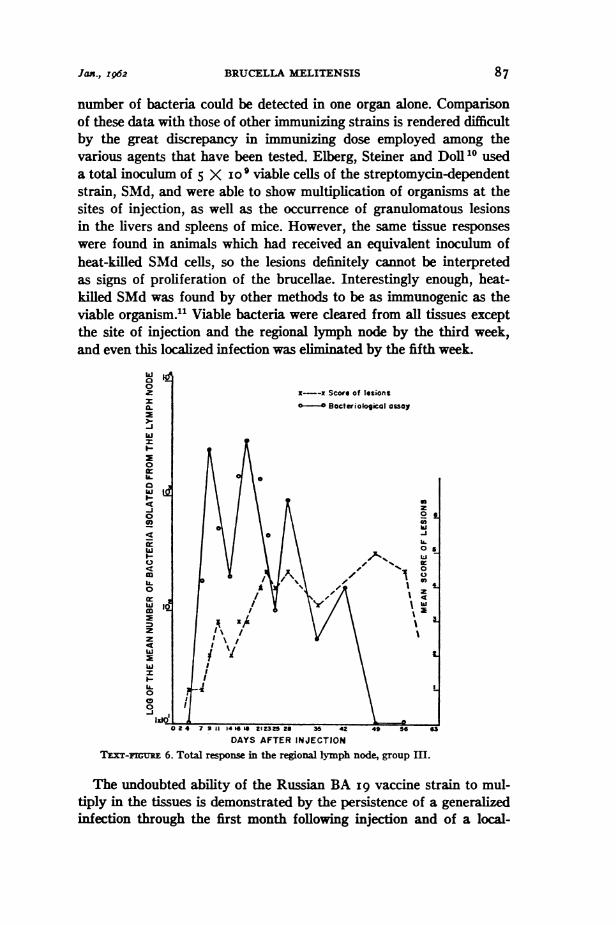
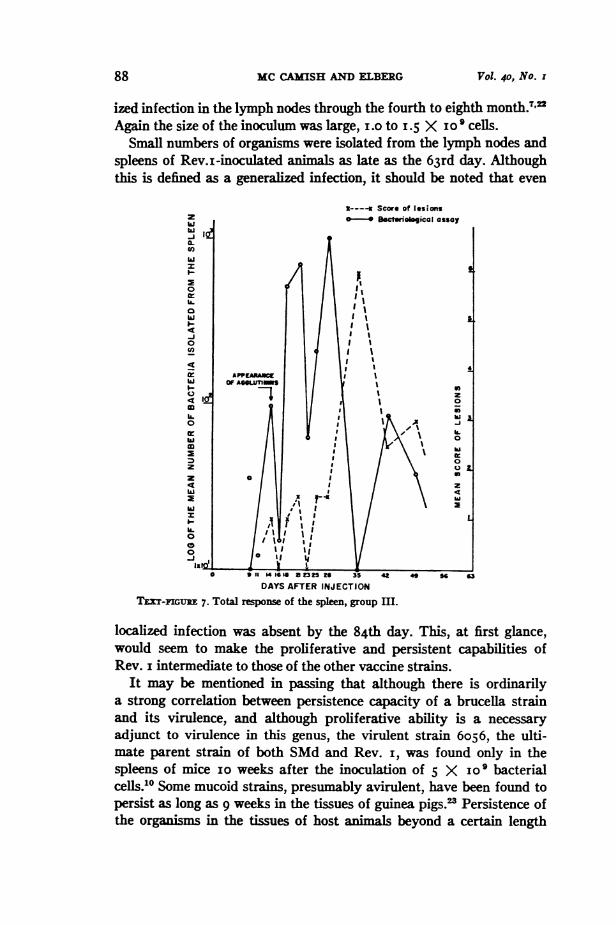
The highest relevant number was assigned to each tissue of eachanimal. The mean of the values obtained within the separate groups foreach test interval was used for plotting the figures. Examples of therelation observed between the tissue responses and the course of bac-terial proliferation in the tissues is presented in Text-figures 5, 6 and 7.The data in Text-figure 7 indicate that the brucellae initially multi-
plied successfully in the spleen only to undergo a drastic reductionin numbers immediately after the appearance of serum agglutinins.The course of events leads to the interpretation that some humoralfactor, appearing at the same time as the agglutinins, is effective inridding the organ of a majority of bacteria but is ineffective againstorganisms located intracellularly.1l21 These survivors may well serveas the generators of the second massive involvement of the spleen whilethey and their host cells provide the initial foci for a granulomatousreaction. Extensive multiplication of brucellae within the granulomasis strongly indicated by the combined data. This second line cellulardefense is, however, eventually successful in eliminating bacteria fromboth the spleen and the lymph nodes.The possibility exists that the second rise in the bacterial count in
the spleen is the sign of re-infection of that organ as a consequenceof a secondary bacteremia resulting, for example, from an overwhelming
Jax., I962
MC CAMISH AND FLBERG
infection in the regional lymph node. The main evidence against this
is the coexistence of granulomas with the rise, contrasted with thefact that the development of granulomatous lesions generally requiresseveral days.Serum agglutinins, of a titer of I:40 or greater, were produced in
z
0
ki-z
is
i-i
09w:
0It
-J
4t
hi
0
4
IL.0
hi
z
z
4
hi
I--
0
0
0
DAYS AFTER INJECTION
TXT-FIGuRE S. Total response at the site of injection, group m.
78 per cent of the animals sacrificed after these antibodies first ap-peared in individuals of the various groups and were found throughoutthe rining period of the experiment. The time of their first appear-ance and the frequency of their occurrence was directly related to thenumber of orga in the original inoculum, but the maximum titerattained was independent of that factor.
DIsCUSSION
The results amply demonstrate the ability of B. melitensis Rev. i
to multiply in the tissues of guinea pigs. As few as io organisms were
capable of eliciting an infection in which a hundredfold increase in the
86 Vol. 4o, No. z
Jaxg., I962 BRUCELLA MELITENSIS 87
number of bacteria could be detected in one organ alone. Comparisonof these data with those of other immunizing strains is rendered difficultby the great discrepancy in immunizing dose employed among thevarious agents that have been tested. Elberg, Steiner and Doll10 useda total inoculum of 5 X i0o viable cells of the streptomycin-dependentstrain, SMd, and were able to show multiplication of organisms at thesites of injection, as well as the occurrence of granulomatous lesionsin the livers and spleens of mice. However, the same tissue responseswere found in animals which had received an equivalent inoculum ofheat-killed SMd cells, so the lesions definitely cannot be interpretedas signs of proliferation of the brucellae. Interestingly enough, heat-killed SMd was found by other methods to be as immunogenic as theviable organism.1 Viable bacteria were cleared from all tissues exceptthe site of injection and the regional lymph node by the third week,and even this localized infection was eliminated by the fifth week.
wC)0zz
IL
a--J
I~-
0
03dL.-4
m
1-J
0
hi
0
0)
zco
C
J°00r0-J
x---x Score of lesionsB- cteriologcal assay
DAYS AFTER INJECTION
TExT-rGuzE 6. Total response in the regional lymph node, group m.
The undoubted ability of the Russian BA i9 vaccine strain to mul-tiply in the tissues is demonstrated by the persistence of a generalizedinfection through the first month following injection and of a local-
MC CAMaSH AND ELBERG Vol. 4o, No. I
ized infection in the lymph nodes through the fourth to eighth month.7'2Again thesize of the inoculum was large, I.O tO I.5 X io9 ceUs.
Small numbers of organisms were isolated from the lymph nodes andspleens of Rev.i-inoculated animals as late as the 63rd day. Althoughthis is defined as a generalized infection, it should be noted that even
X----x Score of lesions2 b -A ..
J
al
-J
hi
n
S
I~-
m
0
iL0
w
hi
0
In
4
C.)
0
z
4tula
IL
0
0
0
DAYS AFTER INJECTION
IExT-IGURE 7. Total reponse of the spleen, group m.
localized infection was absent by the 84th day. This, at first glance,would seem to make the proliferative and persistent capabilities ofRev. i intermediate to those of the other vaccine strains.
It may be mentioned in passng that although there is ordrilya strong correlation between persistence capacity of a brucella strainand its virulence, and although proliferative ability is a necessary
adjunct to virulence in this genus, the virulent strain 6056, the ulti-mate parent strain of both SMd and Rev. i, was found only in thespleens of mice io weeks after the inoculation of 5 X io0 bacterialcells.'0 Some mucoid strains, presumably avirulent, have been found topersist as long as 9 weeks in the tissues of guinea pigs.' Persistence ofthe organisms in the tissues of host animals beyond a certain length
88

BRUCELLA MELITENSIS
of time may with validity be assumed to indicate proliferation, but itsunqualified use as a critenron of virulence may reasonably be calledinto question.
Attempts to assess virulence by means of histologic alterations areno less subject to lack of clarity. The initial cellular response at thesite of injection was considered to be a sensitive indicator of the ani-mal's ability to deal with the inoculated orgnis.m An extreme neutro-phil reaction was evoked by the avirulent SMd strain in normal animals,whereas the large inoculum of virulent 6056 called forth a rapid re-sponse, but less marked and of a slightly less suppurative nature.The local reaction to the injection of large numbers of the BA organ-isms seems to be similar to the reaction to 6056, though less pronounced.The only signs of reaction at the site of injection of small numbersof Rev. i cells were those of a barely perceptible inflammation. Plasmacells were frequently observed in the tissues of the animals inocu-lated with SMd and with BA. They were noticed especially at the edgesof the abscess formed at the site of injection.10 Chernysheva8 founda period of plasma cell increase I5 to 30 days after inoculation ofBA. Plasma cells did not seem to be stimulated by the injection ofRev. i. They, and the equally rarely appearing neutrophils, were seenin quantity in only a few sections and scattered sparsely in some others.The sequels in the other organs after the primary reaction at the
injection site seem to justify the assumption of Elberg, Steiner andDoll.10 Thus, SMd infection was characterized by the production ofonly minor epithelioid lesions in the other organs, while the lesionsarising in the infections due to the other vaccine strains were those of atypical brucella infection in which there was occasional central ne-crosis of the nodule. Unfortunately the differences in the host speciesused in the SMd experiments and the experiments with BA and Rev. ipreclude the possibility of absolute comparison of the tissue responses.However, these data, conjoined with the available knowledge of theimmunogenic capacities of the respective strains, do indicate that anefficiency in the removal of bacteria from the site of injection is notan advantage in the production of the immune state. Organisms ofthe two relatively more virulent vaccine strains are afforded moreopportunity for establishment and proliferation in the reticuloendo-thelial elements of the internal tissues. The implications of this factare in harmony with the generally accepted view of the importance ofcellular immunity in brucella infection.The in vitro investigations involving the interactions of brucella or
tubercle bacilli with tissue cells have been numerous and in some casescontradictory. For ready presentation of some of the data which may
89fax., z962

MC CAMISH AND ELBERG
be relevant to the immune mechanism in vivo, the following summaryhas been prepared. Because of the strong evidence for the similaritiesin the behavior of the different bacteria and mammalian cell types, anddespite the realization that too easy transfer of data may be invalid,reactions involving both Mycobacterium tuberculosis and brucella spe-aes with monocytes, embryonic fibroblasts, and blood phagocytes havebeen included. The differences in normal and immune cells have not beennoted in the summary. The reports have been synthesized into a hypo-thetically chronologic order, inferring the transition from normal tomune.Virulent bacteria inhlibit, apparently nonspecifically, the migration of
leukocytes.2' Few virulent bacteria are found in cells at first, whichmay slow the rate of their multiplication to the level at which the hostcell bursts.' Or, conversely, virulent bacteria bring about degenerationof the host cell.2" Immune serum protects the host cell from degen-eration.7 One or both factors keep the virulent organisms in an intra-cellular situation, in which position they are protected from the detri-mental effects of serum.2-'l The presence of the extracellular serummay serve to prevent the spread of the infection."' The bacteria aredestroyed in the host cell.282
Avirulent bacteria fail to influence the migration of leukocytes,24and their subsequent rapid ingestion by phagocytes allows them to reachquickly the population level at which the host cell bursts.15 In vivostudies indicate the presence of bacteriostatic substances in the tissuefluids which can act on these exposed bacteria.30 Or, conversely, aviru-lent bacteria fail to induce degeneration of the host cell26 and aredestroyed intracellularly.The data presented in this paper suggest that Rev. i in guinea pigs
behaves to a certain degree like virulent organms in vitro. Especiallyin the spleen, the number of bacteria increases rapidly until the ap-pearance of immune serum factors. Either because they have not beenphagocytized to any great extent or because, in the absence of immuneserum, they have caused degeneration of the cells which had phago-cytized them, large numbers are exposed to the action of anti-brucellafactors when they do appear. Thereafter the few which manage to sur-vive because of a perhaps transient intracellular location are protectedfrom the further effect of extracellular damaging agents and enter asecond phase of multiplication.
It is at this stage, perhaps, that the essentially attenuated stateof the virulence of Rev. I is evidenced. Multiplication of the bacteriaand extension of the lesions proceed for a time with parallel intensity.Disappearance of bacteria from the tissues occurs with relative rapidity,
l'01. 4o, N.O.rgo

Jan., 1962 BRUCELLA MELITENSIS 9I
and it is suggested that the intrinsic nature of the bacterial cell isresponsible for the host cell's eventual success in ridding itself of theparasite. A possible mechanism of the intracellular destruction is astepwise alteration of the bacterial cell wall followed by dissolutionof the cellular integrity of the bacteria by a lysozyme-like enzymewhich has been isolated from monocytic cells.31'32 Demonstration of theenzyme activity is not as readily accomplished with a virulent strain,60I5, of Brucella melitensis as it is with Rev. i.
Bacteriologic, histologic, and serologic responses of guinea pigs tosmall graded doses of the immunizing strain, Rev. i, of BruceUa meli-tensis have been reported.The tissue response to the inoculum is the development of a general-
ized granulomatous inflammation, particularly characterized by the vir-tual absence of neutrophils. Epithelioid nodules, some of which devel-oped microscopic abscesses in their centers, were formed in the lymphnodes proximal to the site of injection and in the spleen. Extensiveformation of giant cells occurred in only one case.
After the establishment of the infection in either the lymph node orthe spleen, the multiplication of bacteria and the extension of the lesionsproceeded with parallel intensity. The multiplication of bacteria withinthe lesion is postulated.
Clearance of bacteria from the tissues was followed, more gradually,by the resolution and disappearance of the lesions.The reaction of the host to Brucella melitensis Rev. i has been com-
pared to the reactions of host animals to two other immunizing strains,SMd and BA.The relation of the in vivo response to viable brucellae to the in
vitro reactions of tissue cells to granuloma-producing organisms hasbeen discussed. The conclusion was reached that B. melitensis Rev. ibehaves as a virulent organism until it is fully established intracellularly.Its success as an intracellular parasite is limited. This propitious order-ing of its properties of virulence and avirulence makes it particularlysuitable as an immunizing agent against the disease caused by B. meli-tensis.
REFERENCES
I. FABYAN, M. A contribution to the pathogenesis of Brucella abortus. J. Med.Res., I9I2-I9I3, 26, 44I-487.
2. MEYER, KL F.; SHAw, E. B., and FLEIsCHNER, E. C. Studies on the genusBrucella Nov. Gen. The pathogenicity of B. melitensis and B. abortus forguinea pigs. J. Infect. Dis., I922, 3I, I59-I97.

92 MC CAMISH AND ELBERG Vol. 4o, No. I
3. BRAuE, A. I. Studies in the pathology and pathogenesis of experimental brucel-losis. I. A comparison of the pathogenicity of Brucella abortus, BruceUamelitensis, and BruceUa suis for guinea pigs. J. Infect. Dis., i95i, 89, 79-86.
4. BRAUDE, A. I., and SpiK, W. W. Studies in the pathology and pathogenesisof experimental brucellosis. m. Investigations pertaining to the functionof the spleen. J. Infect. Dis., 195I, 89, 272-276.
S. BRAuDE, A. I. Studies in the pathology and pathogenesis of experimental brucel-losis. II. The formation of the hepatic granuloma and its evolution. J. Infect.Dis., I95I, 89, 87-94.
6. MOULTON, J. E., and MEYER, M. E. The pathogenesis of Brucella suis infec-tion in guinea pigs; lesions of the spleen, liver, testis and articulations. CorneUVet., I958, 48, I65-I95.
7. VERSEHLOVA, P. A., and KOKORIN, I. N. Morphologic and bacteriologic char-acteristics of vaccinal process in brucellosis. Zhur. mikrobiol. epidemiol. iimmunol., I954, (T) 7-13.
8. CHERNYSHEVA, M. I. Histochemical characteristics of morphological reactionsin brucellar infection. Biul. eksp. biol. med., 957, 44(9), 109-II4.
9. CHERNYsmmvA, M. I. The cytological method of investigation of vaccinal im-munogenesis in brucellosis. Zhur. mikrobiol. epidemiol. i immunol., 1958,29(II), 20-25.
IO. ELBERG, S. S.; STEINER, P. E., and DOLL, J. P. Tmmunization against brucellainfection. V. Histopathologic appraisal of immunity induced in mice by astreptomycin-dependent mutant of Brucella melitensis. Am. J. Path., I955,3I, IO65-I075.
I i. HERZBERG, M., and ELBERG, S. S. Immunization against brucella infection. m.Response of mice and guinea pigs to injection of viable and nonviable sus-pensions of a streptomycin-dependent mutant of Brucella melitensis. J. Bact.,I955, 69, 432-435.
12. ELBERG, S. S., and FAuNCE, K, JR. Immimization against brucella infection. VI.Immunity conferred on goats by a nondependent mutant from a streptomycin-dependent mutant strain of Brucella melitensis. J. Bact., I957, 73, 2I1-2I7.
13. ELBERG, S. S., and MEYER, K F. Caprine immunization against brucellosis. Asummnary of experiments on the isolation, properties and behavior of a vaccinestrain. Bull. World Health Org., I958, I9, 71I-724.
I4. JoNEs, L. M.; THOMPSON, P. D., and ALTON, G. G. Production of immunityagainst experimental Br. melitensis infection in goats; a comparison of twokilled and one living vaccine. J. Camp. Path., I958, 68, 274-287.
i5. ALTON, G. G. Brucellosis in Malta I. Brit. Vet. J., I959, I15, 25I-260.I6. ELBERG, S. S. Immunization against brucella infection. 7. Tmmunological and
epidemiological studies in Cordoba, Spain. Bull. World Health Org., I959,20, IO33-IO52.
I7. RENOux, G. ttudes sur la brucellose ovine et caprine. XX. Vaccination desbrebis contre l'infection a Brucella melitensis. Comparaison de trois vaccines.Arch. Inst. Pasteur, Tunis, 1958, 35, 25I-274.
i8. ELBERG, S. S., and FAUNCE, W. K., JR. Immunization against brucella infec-tion. VIII. Response of monkeys, guinea pigs, and goats to immunzation.Bull. World Health Org., I962. (In press)
I9. MAGoFFN, R. L., and SPiK, W. W. The protection of intracellular brucellaagainst streptomycin alone and in combination with other antibiotics. J. Lab.& Clin. Med., 195I, 37, 924-930.
20. SHAFFER, J. M.; KucERA, C. J., and SPiNx, W. W. The protection of intra-

Jan., 1962 BRUCELLA MELITENSIS 93
cellular brucelLa against therapeutic agents and the bactericidal action ofserum. J. Exper. Med., I953. 97, 77-90.
2I. HOLLAND, J. J., and PICKETT. M. J. Intracellular behavior of brucella variantsin chick embryo cells in tissue culture. Proc. Soc. Exper. Biol. & ,Med., I956,93, 476-479.
22. ZDRODOWSKI. P.; VERSHILOVA, P., and KOTLAROVA. H. Recherches immuno-logiques sur la brucellose et immunisation humaine contre cette infection aumoyen d'un vaccin vivant attenue. Resv. d'immunol., I956, 20, 85-99.
23. JoNEs. L. M., and BERMAN. D. T. The pathogenicity of mucoid variants ofbrucellae for guinea pigs. J. Infect. Dis., I95i. 89, 2I4-223.
24. ELBERG, S. S., and SCHNEIDER, P. Directed leucocyte migration in response toinfection and other stimuli. J. Infect. Dis., I953. 93, 36-42.
25. BRAU-N, W.; POMALES-LEBRON, A., and STINEBRING, W. R. Interactions be-tween mononuclear phagocytes and Brucella abortus strains of different viru-lence. Proc. Soc. Exper. Biol. & Med., I958, 97, 393-397.
26. FONG, J.; SCHNEIDER, P., and ELBERG, S. S. Studies on the tubercle bacillus-monocyte relationship. II. Induction of monocyte degeneration by bacteriaand culture filtrate: specificity of serum and monocyte effects on resistanceto degeneration. J. Exper. Med., 1957, I05, 2 5-37.
27. FONG, J.; SCHNEIDER. P., and ELBERG. S. S. Studies on the tubercle bacillus-monocyte relationship. I. Quantitative analysis of the effect of serum ofanimals vaccinated with BCG upon the bacterium-monocyte system. J. Exper.Med., I956, I04, 455-465.
28. ELBERG, S. S.; SCHNEIDER, P., and FONG, J. Cross-immunity between Brucellamelitensis and Mycobacterium tuberculosis; intracellular behavior of Brucellamelitensis in monocytes from vaccinated animals. J. Exper. Med., I957, io6,545-554.
29. PomALEs-LEBR6ON, A., and STINEBRING, W. R. Intracellular multiplication ofBrucella abortus in normal and immune mononuclear phagocytes. Proc. Soc.Exper. Biol. & Med., I957, 94, 78-83.
30. LuRIE, M. B. Studies on the mechanism of immunity in tuberculosis; the roleof extracellular factors and local immunity in the fixation and inhibition ofgrowth of tubercle bacilli. J. Exper. Med., I939, 69, 555-578.
3I. RALSTON, D. J., and ELBERG, S. S. Action of glycine and a lysozyme-like agentfrom rabbit monocytes in the destruction of brucella. Proc. Soc. Exper. Biol.& Med., I960, I04, 464-467.
32. RALSTON, D. J., and ELBERG, S. S. Intramonocytic destruction of brucella.J. Infect. Dis., 196I, 1O9, 7I-80.







![f V a V a l Journal of Vaccines & Vaccination · but it had the ability to induce abortions and teratogenesis in pregnant ewes, goats and cows [3,16,17]. To overcome these drawbacks,](https://img.pdfslide.net/doc/110x75/5d635f3188c9936c668b7cc5/f-v-a-v-a-l-journal-of-vaccines-vaccination-but-it-had-the-ability-to-induce.jpg)





















