Embed Size (px)
Citation preview

EXPANDED DISTRIBUTION OF THE BAMBOO SPIDER MITE,
SCHIZOTETRANYCHUSLONGUS (ACARI: TETRANYCHIDAE),
AND PREDATION BY NEOSEIULUS FALLA CIS (ACARI: PHYTOSEIIDAE)
SCHIZOTETRANYCHUS
LONGUS
NEOSEIULUS FALLACIS
BAMBOOORNAMENTAL
SCHIZOTETRANYCHUS
LONGUS
NEOSEIULUS FALLA CIS
BAMBUS
ORNAMENTAL
BY P. D. PRATT * and B. A. CROFT**
SUMMARY:Schizotetranychus longus Saito is a common pest of bamboo in Japan.We report its new occurrence in the Willamette Valley, Oregon, on bamboo.We assessed the ability of adult females of Neoseiulus fallacis (Garman), anative predaceous mite, to invade S. longus nests after construction times of5 or 10 days. Frequency of predators in nests decreased from 5 to 10 days,although the predator laid most of its eggs in nests irrespective of constructiontime. In choice tests, spider mite webbing was more attractive to N.fallacis thaneggs, but similar to feces. Predators searched more in proximity to feces thaneggs, but resting sites were equally common near both. Survival, activity andreproduction of N. fallacis when given excess of mixed life stages of S. longus,Tetranychusurticae Koch, pollen of Tulipagesneriana L. or Pseudotsuga menzesii(Mirbel), or no food were measured by holding single adult female N.fallacis for7 days. Survival, activity, oviposition and immature production of predatorswere alike for both spider mites, but lower (or higher activity) with pollens orwhen starved. To see if N. fallacis would suppress S. longus under normalgrowing conditions, predators were added to infested Sasaella hidaensis (Makinoand Uchida) var. 'Murai' plants, and mites were monitored thereafter for5 weeks. N. fallacis significantly reduced levels of S. longus and the rates that itinfested bamboo leaves; it nearly eliminated S. longus from plants at the end ofthe test.
ZUSAMMENFASSUNG:Die Spinnmilbe Schizotetranychus longus Saito ist einweitverbreiteter Schiidling an Bambus in Japan und semi-tropischen RegionenNordamerikas (Kalifornien, Georgia und Florida). Erstmals wird iiber dasAuftreten dieser Art an Bambus im Willamette- Valley, Oregon, berichtet. Eswurde untersucht, ob adulte Weibchen der (in Oregon heimischen) RaubmilbeNeoseiulus fallacis (Garman) fahig sind, 5 oder 10 Tage alte Gespinste von S.longus zu durchbrechen und in deren Nester einzudringen. Mit zunehmenderGespinstdichte nahm die Anzahl der Raubmilben in den Spinnmilbennestern ab.Unabhiingig von der Gespinstdichte legten die Raubmilben den Grossteil derEier in den Nestern ab. In Wahlversuchen bevorzugte N. fallacis Spinnmilben-gespinste und -kot gegeniiber Spinnmilbeneiern. Nahrungs~uchende Raubmil-benweibchen wurden hiiufiger in der Niihe von Spinnmilbenkot als in derNiihevon Spinnmilbeneiern gefunden, rastende Raubmilbenweibchen zeigten
* USDA-ARS, 3205 College Ave., Ft. Lauderdale, Fl. 3314, U.S.A. E-mail: [email protected] Department of Entomology, Oregon State University, Corvallis, Oregon 97331-2907, U.S.A. E-mail: [email protected].
Acarologia, vol. XL, rase. 2, 1999.
- --- ------

- 192 -
keine Priiferenz. Die Oberlebensfahigkeit, Aktivitiit und Reproduktion voneinzeln gehaltenen adulten N fallacis Weibchen wurde mit den Beutetieren S.longus und Tetranychus urticae Koch bzw. mit Pollen von Tulipa gesneriana L.und Pseudotsuga menzesii (Mirbel) iiber einen Zeitraum von 7 Tagen gemessen.Als Kontrolle wurden Weibchen ohne Nahrung gehalten. Der Prozentsatz aniiberlebenden Weibchen, Aktivitiit, Oviposition und Nachkommenszuwachswaren gleich hoch mit beiden Spinnmilbenarten als Nahrung, jedoch geringer(und die Aktivitiit hoher) mit Pollen bzw. ohne Nahrung. Urn zu testen, ob S.longus unter natiirlichen Bedingungen durch N fallacis bekiimpft werden kann,wurden die Raubmilben an spinnmilbenbesetzten Bambuspflanzen, Sasaellahidaensis (Makino & Uchida) var., Murai', ausgesetzt und das Riiuber/BeuteVerhiiltnis iiber einen Zeitraum von 5 Wochen kontrolliert. N fallacis reduziertesowohl die Populationsdichte der Schadmilben, als auch die Anzahl der durch S.celarius befallenen Blatter signifikant. Gegen Versuchsende war S. longus bei-nahe vollstiindig von den Pflanzen verschwunden.
INTRODUCTION
Establishment of an exotic pest may depend on theabsence of native predators that are adapted to it(GOEDEN& LOUDA,1976). Also, changes in localenvironmental conditions caused by a pest canimprove microclimates or retard predators, and there-by enhance its establishment. Schizotetranychus lon-gus Saito is a pest of bamboo in Japan (SAITO,1990b).Upon finding a feeding site, this spider mite builds adensely webbed nest within which it reproduces.SAITOsuggested that the dense webbing and fecaldeposits of S. longus may protect it from predators(SAITO, 1983), although webbing of some spidermites is attractive to many predaceous phytoseiidmite species (SCHMIDT,1976). SAITO(1990) reportedfeeding and adaptations of Typhlodromus (Antho-seius) bambusae Ehara, a native phytoseiid asso-ciated with Schizotetranychus species in Japan andconcluded that it had coevolved with S. celarius to
feed and maneuver within the nest of the pest. Nostudies to date have reported any endemic phytoseiidsin the USA that are associated with S. longus.
Recent inspections of bamboo revealed infesta-tions of an unknown spider mite in the coastal-temperate regions of Oregon and Washington USA,where ornamentals are widely grown. Preliminaryobservations of the pest indicated that it was S. lon-gus. We sought to confirm this identification and, inthe absence of T. bambusae, to measure the ability ofa native phytoseiid mite to exploit the nest habitat
and suppress this spider mite. In temperate-humidareas of the USA, Neoseiulus fallacis (Garman) iswidely released on ornamentals and other crops tocontrol spider mites (PRATT& CROff, 1998). Objec-tives of this study were to measure: 1) propensities ofN fallacis to enter the nest of S. longus, 2) attractive-ness to N fallacis of webbing, feces or eggs ofS. longus, 3) ability of N fallacis to reproduce onS. longus and 4) whether N fallacis could suppressS. longus on bamboo plants under normal growingconditions.
MATERIALS AND METHODS
Identification ofS. longus and predator mite cultures
Initially, an unknown spider mite was collectedfrom commercial bamboo near Coos Bay and Port-land, OR, USA. Adults of both sexes were mountedon glass slides and sent to Z. Q. ZHANG,LandcareResearch, Auckland, New Zealand for identification.Predators for releases (N fallacis) had been collectedfrom crops in the Willamette Valley, Oregon (HADAMet al., 1986). These cultures had been held for sixyears or more with yearly additions of field-collectedmites. Predator cultures were held at 25 (:i: 5)OC,16:8L:D (light:dark), and 75-95% RH, and fed mixed lifestages of the common spider mite, Tetranychus urti-cae Koch, three times per week. Only gravid femalepredators were selected for use in tests. To adjust for

possible dissimilar levels of hunger among predatormites, all N. fallacis adult females were held withoutfood for 24 hours just prior to feeding experiments(below).
Invasion of nests by N. fallacis and attraction to pro-ducts ofS. longus
To quantify the ability of N. fallacis to enterwebbed nests of S. longus, 2.5 x 2.5 cm arenas werecreated on bamboo leaves (underside up) using awater-soaked cotton barrier (SAITO,1990a). To allownest construction, 3-4 S. longus adult females wereadded to each of 16 arenas held at 25 (:I: ltC, 75(:I: 10) RH, and 16:8 L:D. We randomly selected 8arenas and placed a single gravid N. fallacis on eachafter 5 days of nest construction. The other 8 arenasreceived a single female N.fallacis after 10days. Sitesof entrance of the predator into the nest, restinglocations of N. fallacis and number and sites of pre-dator eggs were measured at 2, 4, 6, 8, 10, 12,24, and36 hours after transfer of predators. Mode ofentrance was scored as either entering via existingopenings on either side of the nest or boring anentrance into the nest through the webbing. To com-pare frequencies of these two binomial measuredattributes, we used a binomial test with the null hypo-thesis frequency of 0.5. A P-value <0.10 was consi-dered evidence that the null hypothesis was false.
To test attraction and arrestment of N. fallacis,dual choice arenas were used (SCHMIDT,1976). Are-nas consisted of 2 x 2 cm tile substrate ringed withwater-soaked cotton and replicated 18 times(MACRAE& CROFT,1993). S. longus eggs, webbing orfeces were removed with different forceps from bam-boo leaves [Sasaella hidaensis (Makino and Uchida)var. 'Murai'] and placed near two diagonal comersand the other S. longus product pair was placed nearthe remaining two comers. Treatments were eggs vs.webbing, webbing vs. feces or eggs vs. feces. UnlikeT. urticae, S. longus defecates outside the nest and anymixing of fecal pellets and webbing can be avoided.A single N. fallacis adult female was placed in thecenter of each arena and location of the predatorwhile searching or resting and number and locationof predator eggs were monitored at 1-12,24 and 36hours after transfer of the predator. To compare
193 -
relative attractiveness of the spider mite products, weused a binomial test and significant levels as before.
Feeding and ability ofN. fallacis to control S. longuson bamboo
To measure the ability of N. fallacis to feed,reproduce and develop on S. longus we constructed2.5 x 2.5 cm waterproof arenas ringed with a stickymaterial (Tanglefoot@, The Tanglefoot Co., GrandRapids MI 49504) and replicated tests eight times(MONETTI& CROFT,1997). Three adult female preda-tors of about the same age were transferred to eachindividual arena and excess mixed life stages of S.longus were provisioned every 24 h. For comparisons,N. fallacis was also held with excess amounts ofTulipa gesneriana L. and Pseudotsuga menzesii (Mir-bel) pollen grains. Arenas with mites and treatmentswere held at 25 (:I: ltC, 70 ( :I:5) RH, and 16:8 L:Dfor 7 days. Assessments of survivorship, activity(ambulation), oviposition per female per day, andproduction of immatures (larvae, protonymphs, deu-tonymphs) per female per day were measured every24 hours. An index for survivorship of immatures wascalculated on day 3-7 by dividing the number ofimmatures by the number of eggs present two daysprior to the sampling of immatures (CROFTet al.,1998). We compared our results with reported valuesfor N. fallacis when held under identical conditionswith the optimal prey T. urticae or no food (PRATTetal., 1999). Means of each measured attribute andfood type were compared by analysis of variance(AN OVA) and TUKEY'SHSD.
Preliminary tests showed that N. fallacis wouldfeed, reproduce and develop on S. longus. Because N.fallacis is often used to control pest mites in otherornamental plant systems, we were interested in theability of N.fallacis to suppress S. longus on bamboo.In May, 1998, 10, two-year S. hidaensis 'Murai', of 64(:I: 17.4) culms each, were potted in 4-liter plasticcontainers and inoculated with 50 (:I: 12) adultfemale S. longus. Plants were placed in a shadednursery bed and randomly assigned either release ofthree adult female N.fallacis per plant or no predatorrelease (control). By June 11, S. longus averaged 9.6(:!::3.1) colonies per plant and three adult female N.fallacis were released onto each plant (STRONG&

-------
- 194 -
TABLE I: Invasions of Schizotetranychus longus nests by the phytoseiid predator Neoseiulus fallacis." Duration of nest construction period after inoculation of 3-4 adult female S. longusonto a 2.5 x 2.5 em arena. bAttributes of N.fallacis measured after introduction
into colonies of S. longus. C P-value calculated from binomial test, null hypothesis frequency =0.5.
CROFT,1995). Infestation of new leaves after release(or non-release) of N fallacis were estimated by mark-ing each S. longus colony and scanning for new colo-nies every 7 days. The initially marked colonies of S.longus and 10 randomly selected (with replacement)uninfested leaves per plant were monitored for Nfallacis every 7 days. To adjust for sampling the samepopulations over time, data were analyzed by repea-ted measures ANOVA (VONENDE,1993).
RESULTS
Identification ofS. longus
The unknown spider mite that was collected was S.longus. Although infestations by this pest on orna-mental bamboo had been observed for about 5 yearsbefore this study was begun, this was the first taxono-mic identification for these temperate-coastal regionsof Oregon. Also, no phytoseiids were associated withS. longus within Oregon.
Invasion of nests by N. fallacis and attraction to pro-ducts ofS. longus
N fallacis readily invaded nests of S. longus whenwebbing was minimal but was somewhat less invasivewhen webbing was denser (Table 1).When measuringthe entrance into nests, N fallacis used both natural
openings and created new holes equally as often after5 or 10 days of web construction. N fallacis laidsignificantly more eggs within the nest than without,irrespective of the nest construction time (Table I).When comparing attraction to prey products, N fal-lacis was associated more with webbing than eggseither when searching (P= 0.082) or resting (P=0.052). With feces and eggs, N fallacis searched morenear feces (P=0.059) although it was equally probablefor them to rest among either product (P=0.855).Webbing and feces were equally attractive to N falla-cis when either searching or resting (P=l.O, 0.2 res-pectively).
Feeding and ability ofN. fallacis to control S. longuson bamboo
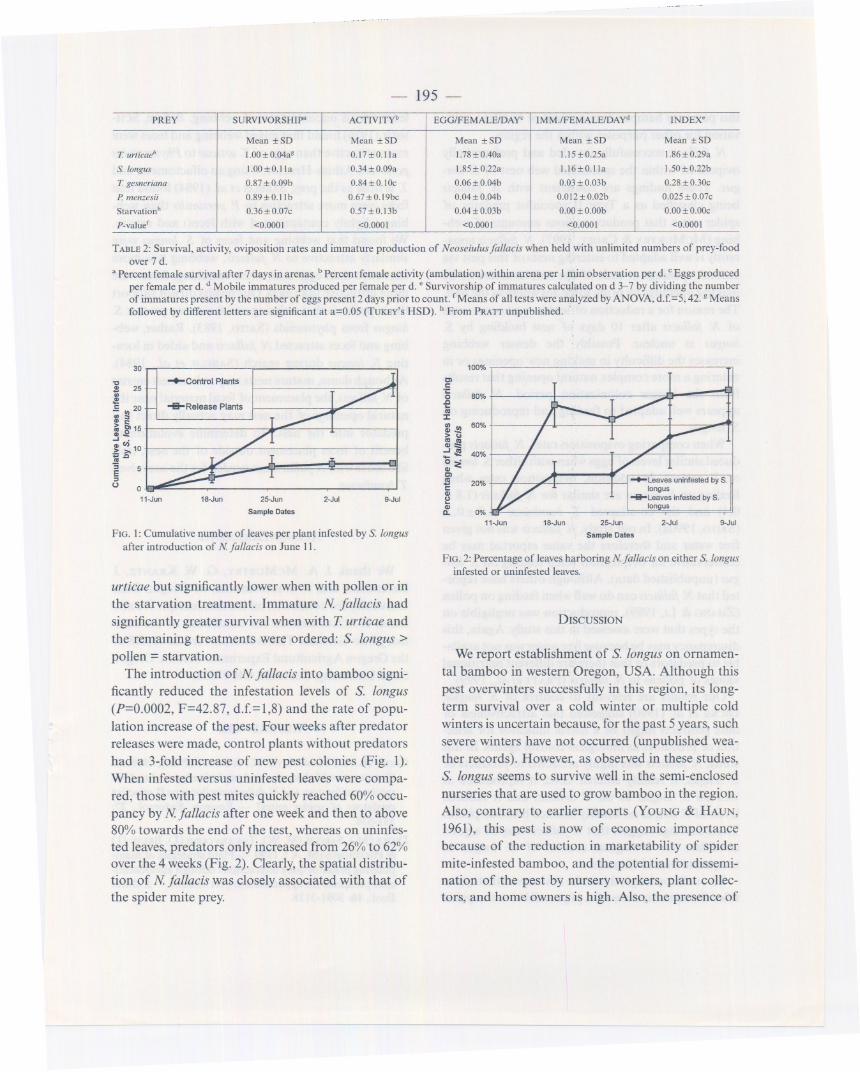
N fallacis survived equally well when held witheither the optimal prey T. urticae or with S. longus,but significantly lower survival occurred when heldwith other treatments (Table 2). Activity of Nfallaciswas similar between the two species of spider mitesbut significantly lower when with pollen or starved.Activity of N fallacis was higher when with pollen ascompared to the starvation treatment, which may bedue to the increased morbidity and mortality of thepredator near the end of the 7 d test (PRATT& CROFT,1998). Oviposition rates and immature productionwere similar when held with either S. longus or T.
Nest ConstructionAttribute of N. fallacisb Proportion P-Valuec
Period"
5 days Observed location Within Nest 0.65
On Leaf 0.35 0.08
10 days Within Nest 0.47
On Leaf 0.53 0.72
5 days Mode of Entrance Natural Opening 0.33
Break-in 0.67 0.68
10 days Natural Opening 0.73
Break-in 0.27 0.22
5 days Egg Placement Within Nest 0.85
Without Nest 0.15 0.02
10 days Within Nest 1.00
Without Nest 0.00 <0.001
- 195 -
TABLE 2: Survival, activity, oviposition rates and immature production of Neoseiulus fal/acis when held with unlimited numbers of prey-foodover 7 d.
a Percent female survival after 7 days in arenas. bPercent female activity (ambulation) within arena per I min observation per d. C Eggs producedper female per d. d Mobile immatures produced per female per d. .Survivorship of immatures calculated on d 3-7 by dividing the numberof immatures present by the number of eggs present 2 days prior to count. fMeans of all tests were analyzed by ANOYA, d.f.=5, 42. g Meansfollowed by different letters are significant at a=0.05 (TUKEY'S HSD). h From PRAIT unpublished. .
30
Control Plants
Release Plants
18-Jun 25-Jun
SampleDates
2-.1ul 8-Jul
FIG. I: Cumulative number of leaves per plant infested by S. longus
after introduction of N. fal/acis on June II.
urticae but significantly lower when with pollen or inthe starvation treatment. Immature N. fallacis hadsignificantly greater survival when with T urticae andthe remaining treatments were ordered: S. longus >pollen =starvation.
The introduction of N. fallacis into bamboo signi-ficantly reduced the infestation levels of S. longus(P=0.0002, F=42.87, d.f.=1,8) and the rate of popu-lation increase of the pest. Four weeks after predatorreleases were made, control plants without predatorshad a 3-fold increase of new pest colonies (Fig. I).When infested versus uninfested leaves were compa-red, those with pest mites quickly reached 60% occu-pancy by N.fallacis after one week and then to above80% towards the end of the test, whereas on uninfes-ted leaves, predators only increased from 26% to 62%over the 4 weeks (Fig. 2). Clearly, the spatial distribu.tion of N. fallacis was closely associated with that ofthe spider mite prey.
- -- ---
100%
80%
60%
40%
20%
0%11-Jun 9-Jul18-Jun 25-Jun
Sample Date.
2-.1ul
FIG. 2: Percentage of leaves harboring N. fal/acis on either S. longusinfested or uninfested leaves.
DISCUSSION
We report establishment of S. longus on ornamen-tal bamboo in western Oregon, USA. Although thispest overwinters successfully in this region, its long-term survival over a cold winter or multiple coldwinters is uncertain because, for the past 5years, suchsevere winters have not occurred (unpublished wea-ther records). However, as observed in these studies,S. longus seems to survive well in the semi-enclosednurseries that are used to grow bamboo in the region.Also, contrary to earlier reports (YOUNG& HAuN,1961), this pest is now of economic importancebecause of the reduction in marketability of spidermite-infested bamboo, and the potential for dissemi-nation of the pest by nursery workers, plant collec-tors, and home owners is high. Also, the presence of
PREY SURVIVORSHIP" ACTIVITYb EGG/FEMALEIDAY. IMM.lFEMALEIDAY. INDEX.
Mean :t: SD Mean :t: SD Mean :t: SD Mean :t: SD Mean :t: SD
T. urlica.!' l.oo:t: 0.04a8 0.17:t:0.1Ia I. 78 :t: 0.40a 1.15:t: 0.25a 1.86:t: 0.29a
S. longus 1.00:t:0.1Ia 0.34:t: 0.09a 1.85 :t:0.22a 1.16:t:0.1Ia 1.50:t: 0.22b
T. gesneriana 0.87 :t: 0.09b 0.84:t: O.IOc 0.06:t: O.04b 0.03 :t: 0.03b 0.28 :t:O.3Oc
P. menzesii 0.89:t:0.1Ib 0.67:t: 0.19bc 0.04:t: O.04b 0.012:t: 0.02b 0.025 :t:0.07c
Starvationh 0.36:t: 0.07c 0.57:t: 0.13b 0.04:t: 0.03b 0.00:t: O.OOb 0.00 :t:0.00c
P-valuef <0.000 I <0.0001 <0.0001 <0.0001 <0.0001

this pest may hamper development of bamboo culti-vation for other purposes within the region.
N Jallacis successfully invaded and preferentiallyoviposited within the specialized web nest of S. lon-gus. These findings are consistent with N Jallacisbeing classified as a Type II specialist predator ofspider mites that produce copious amounts of web-bing (McMuRTRY & CROFT,1997). N Jallacis appa-rently is well adapted to entering nests of this pest vianatural openings at ends of the nest structure or bymaking openings by brute force anywhere in the nest.The reason for a reduction of within-nest occurrence
of N Jallacis after 10 days of nest building by S.longus is unclear. Possibly the denser webbingincreases the difficulty in making new openings or inentering a more complex natural opening that resultsfrom the longer colonization period. N Jallacisappears well adapted to feeding and reproducing onS. longus.
When comparing oviposition rates, N Jallacis pro-duced similar levels of eggs when with either S. longusor T urticae. In addition, oviposition rates whenfeeding on S. longus are similar for N Jallacis (1.8:1:0.4) and the co-adapted T bambusae (1.7:1:0.2)(SAITO,1990a). In our study, N Jallacis was not givenfree water and therefore the value reported may beconservative for oviposition when feeding on S. lon-gus (unpublished data). Although others have repor-ted that N Jallacis can do well when feeding on pollen(ZHANG& LI, 1989), reproduction was negligible onthe types that were assessed in this study. Again, thisdiscrepancy may be because free water was not availa-ble to predators or that there are different nutritionalvalues of pollens to N Jallacis (PRATTet al., 1999).
Our results are relevant to control of S. longus.DICKEet al. (1990) suggested that attractiveness ofmite products might be a useful indicator for selec-tion of predators as biological control agents. In thisstudy, N Jallacis was highly attracted to products ofS. longus, it readily entered nests, it reproduced aswell on S. longus as on T urticae, and it reducedlevels of S. longus on densely infested bamboo plants.These findings suggest that N Jallacis may be a goodcandidate for biological control of S. longus intemperate-humid climates.
To our knowledge, this is the first report that quan-tifies the attractiveness of a phytoseiid to tetranychid
-- -. --. ... .-.--.
196 -
feces versus uncontaminated webbing. Before, SCH-MIDT(1976) found that mixed webbing and feces weremore attractive than eggs of T urticae to Phytoseiuluspersimilis Athias-Henriot. Using an olfactometer andT urticae as the prey, SABELISet al. (1984) found thatfeces were more attractive to P persimilis than web-bing (slightly contaminated with feces) and exuvia.We found that webbing and feces of S. longus weresimilarly attractive to N Jallacis, webbing was moreattractive than eggs and, when searching, feces weremore attractive than eggs. Our results do not supportthe hypothesis that webbing and feces may protect S.longus from phytoseiids (SAITO,1983). Rather, web-bing and feces attracted N Jallacis and aided in loca-ting S. longus during search (SABELISet al., 1984).Although dense, mature nests may reduce nest entriesof N Jallacis, the placement of fecal material near thenatural openings of the nest may actually direct thispredator into the nest. To determine evolutionarybenefit of feces placement outside of the nest, testslike those with N Jallacis are needed for the coevolvedT bambusae.
ACKNOWLEDGEMENTS
We thank 1. A. McMuRTRY, G. W. KRANTZ,1.DEANGELISand P. SCHAUSBERGERof Oregon StateUniversity for comments on the manuscript. We alsothank P. SCHAUSBERGERfor translation of the sum-
mary into German. This is Journal Article 11,418 ofthe Oregon Agricultural Experiment Station. OregonState University, Corvallis, OR 97331.
REFERENCES
CROFf (B. A.), MONETTI(L. N.) & PRATT(P. D.), 1998.-Comparative life histories and predation types: Are Neo-seiulus califomicus and N fallacis similar type II selectivepredators of spider mites? - Environ. Entomol., 27:531-538.
DICKE (M.), SABELIS(M. W.), TAKABAYASHI(1.), BRUIN (1.)& POSTHUMUS(M. A.), 1990. - Plant strategies of mani-
pulating predator-prey interactions through allelochemi-cals: prospects for application in pest control. - 1. chern.Ecol., 16: 3091-3118.

GOEDEN(R. D.) & LoUDA(S. M.), 1976.- Biotic interfer-ence with insects imported for weed control. - Ann. Rev.Entomol., 325-342.
HADAM(1.1.),ALINIAZEE(M. T.) & CROff (8. A.), 1986.-Phytoseiid mites of major crops in Willamette Valley,Oregon, and pesticide resistance in Typhlodromuspyri. -Environ. Entomol., 15: 1255-1263.
JEPPSON(L. R.), KEIFER(H. H.) & BAKER(E. W), 1975.-Mites Injurious to Economic Plants. - Univ. of Califor-nia Press, Berkeley and Los Angeles, 614 p.
MACRAE(I. Y.) & CROff (8. A.), 1993. - Influence oftemperature on interspecific predation and cannibalismby Metaseiulus occidenta/is and Typhlodromus pyri (Aca-rina: Phytoseiidae). - Environ. Entomol., 22: 770-775.
McMURTRY(1.A.) & CROff (8. A.), 1997.- Life-styles ofphytoseiid mites and their roles in biological control. -Ann. Rev. Entomol., 42: 291-321.
MONETTI(L. N.) & CROff (8. A.), 1997. - Neoseiuluscalifomicus and Neoseiulus fallacis: Larval responses toprey and humidity, nymphal feeding drive and nymphalpredation on phytoseiid eggs. - Exp. appl. Acarol., 21:225-234.
PRATT(P. D.) & CROff (8. A.), 1998.- Panonychus citri onornamental Skimmia in Oregon, with assessment of pre-dation by native phytoseiid mites. - Pan-Pacif. Ento-mol., 74(3): 163-168.
PRATT (P. D.), SCHAUSBERGER (P.) & CROff (8. A.), 1999.-
Prey-food types of Neoseiulusfallacis and prey-food esti-mates for five representative phytoseiid species. - Exp.appl. Acarol., 23: 551-565.
SABELIS(M. W), AFMAN(8. P.) & SLIM(P. 1.), 1984. -Location of distant spider mite colonies by Phytoseiulus
197
persimi/is: localization and extraction of a kairomone. -Acarology, 6(1): 431-440.
SAITO(Y), 1990a. - Life-history and feeding habit ofTyphlodromus bambusae, a specific predator of Schizote-tranychus celarius. - Exp. appl. Acarol., 10: 45-51.
SAITO(Y.), 1990b. - Two new spider mite species of theSchizotetranychus celarius complex (Acari: Tetranychi-dae). - App. Entomol. Zoo., 25(3): 389-396.
SAITO(Y.), 1983. - Theconceptof lifetypesin Tetranychi-nae. - An attempt to classifythe spinningbehaviourofTetranychinae. - Acarologia, 24: 377-391.
SCHMIDT(Y.G.), 1976. - Influence of traces left behind byits prey on searching behaviour and searching successof Phytoseiulus persimilis. - 1. appl. Entomol., 82: 216-218.
STRONG(W B.) & CROff (8. A.), 1995. - Inoculativerelease of phytoseiid mites (Acarina: Phytoseiidae) intothe rapidly expanding canopy of hops for control ofTetranychus urticae (Acarina: Tetranychidae). - Envi-ron. Entomol., 24: 446-453.
VONENDE(c. N.), 1993. - Repeated-measures analysis:growth and other time-dependent measures. - Pp. 113-137, in S. M. SCHEINER& 1.GUREVITCH(eds) Design andAnalysis of Ecological Experiments. Chapman & Hall,New York, New York, USA.
YOUNG(R. A.) & HAUN(1. R.), 1961. - Bamboo in theUnited States: description, culture, and utilization. -USDA Handbook 193.174 pp.
ZHANG(N.) & LI (Y.), 1989. - Rearing of the predaciousmite, Amblyseius fallacis with plant pollen. - Chin. 1.BioI. Control., 5: 60-63.

.





























