Embed Size (px)
Citation preview

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 1/19
Comparative morphology of populations of Monstera Adans.
( Araceae) from natural forest fragments in Northeast Brazilusing elliptic Fourier Analysis of leaf outlines
I. M. Andrade1, S. J. Mayo2, D. Kirkup2 & C. Van Den Berg1
Summary. A comparative study of the leaf outline morphometrics of Monstera adansonii var. klotzschiana , M. adan-
sonii var. laniata and M. praetermissa was carried out. The study focused on populations in isolated montane humid(brejo) forests of Ceará state in Northeast Brazil and compared them with populations from Amazonia and theBrazilian Atlantic Forest. Digitised outlines were prepared from a total of 1,695 field-collected leaf images from 20populations, and elliptic Fourier analysis was used to generate matrices of coef ficients, from which six shape
variables (principal components) were extracted using Principal Components Analysis. Intra-population variability and inter-population differences were analysed with multivariate distance methods. Separate analyses were carriedout for each of three leaf size classes (juvenile, submature, mature) because of the strong heteroblasty typical of this genus. Juvenile leaves were the least variable size class within populations of M. adansonii var. klotzschiana . Theshape variables expressed very similar types of variation in all three size classes. The Ceará brejo populations of M.
adansonii var. klotzschiana showed significant differences between mature leaf outlines in all pairwise comparisons;the Pacatuba population was the most distinct. The Ceará populations did not cluster together exclusively. In allthree size classes, populations clustered together into their taxonomic groups, most clearly so in mature leaves. Nocorrelation between morphological and geographic distance matrices was found, nor between morphological andmolecular distance. The study showed that leaf outline shape is a practicable and useful quantitative trait forstudying morphological variability at species, varietal and population levels.
Key Words. Araceae , Brazil, brejo forest, Ceará, elliptic Fourier Analysis, Monstera , morphometrics.
IntroductionThe context of this study is the “brejo forest ” — amosaic of isolated humid forest areas in Northeast Brazil scattered within the region’s predominant semi-arid caatinga biome wherever rainfall brought by humid winds from the Atlantic Ocean providessuitable microclimates in mountainous terrain (Saleset al. 1998; Rodal et al. 1998; Sampaio et al. 2002;Borges-Nojosa & Caramaschi 2003; Pôrto et al. 2004; Andrade et al. 2007). The brejo forests are today classified as part of the Brazilian Atlantic forest in the
category of seasonal semi-deciduous montane forest (IBGE 1985, 2005). In Ceará State there are sevensuch areas (Figueiredo 1997) situated roughly halfway between Amazonia and the Atlantic forest, Brazil’smajor humid forest regions.
The brejo forests are of particular biogeographicinterest because they are thought to be natural fragmentsrepresenting relicts of previous expansions of the Atlanticand Amazon forests (Ab’Sáber 1982; Andrade-Lima1982; Borges-Nojosa & Caramaschi 2003). The resultsof Oliveira et al. (1999), in a study of a Quaternary pollen profile from central Bahia (relatively near thefocus of our study in Ceará), suggest that humid andcooler climates existed in Northeast Brazil until around4,500 yrs BP, and they discussed two hypotheses whichpostulate corridors of humid forest linking the Amazon
and Atlantic forests; one along the northeastern coast of Brazil from Maranhão to the region around Salvador inBahia and the other across central Brazil through anetwork of gallery forests. The existence of suchconnections and associated patterns of gene flow over
Accepted for publication June 2008.1 Programa de Pós-Graduação em Botânica, Departamento de Ciências Biológicas, Universidade Estadual de Feira de Santana (UEFS), Avenida Universitária s/n,
44031-460, Feira de Santana, Bahia, Brazil.2 Herbarium, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK. e-mail: [email protected]
Electronic supplementary material The online version of this article (doi:10.1007/s12225-008-9032-z ) contains supplementary material, which is available toauthorized users.
KEW BULLETIN VOL. 63: 193–211 (2008)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 2/19
wide areas of eastern South America is consistent withrecent results from lizard and amphisbaenid biogeogra-phy (Borges-Nojosa & Caramaschi 2003), and our ownresults using AFLP molecular markers in Monstera
adansonii Schott var. klotzschiana (Schott) Madison(Andrade et al. 2007).
This paper is part of a population level study aimed at comparing morphological and molecular patterns in indi- vidual species of the family Araceae occurring in brejoforests in the Brazilian state of Ceará and in the Atlanticand Amazon forests; a molecular study of Monstera
adansonii var. klotzschiana has already been published(Andrade et al. 2007).
Monstera adansonii var. klotzschiana was chosen as atypical representative of the herbaceous brejo flora forthree main reasons. Firstly as a root-climber of humidtropical forests it is intolerant of low humidity and isconfined ecologically to this habitat — it has not beenobserved in caatinga or seasonally dry forests. On the
other hand, it is rarely found in undisturbed primary forest, occurring typically in secondary moist tropicalforest with a well-developed canopy. Flowering is most conspicuous where light levels are higher, at the edgeof clearings and along forest edges. We observed that in Ceará this taxon only occurs near waterfalls andstreams that are permanent throughout the year.Second, this taxon is widespread in the eastern half of tropical South America (Madison 1977), allowing comparison of the Ceará populations with those in Amazonia and the Atlantic forest. It is one of severalspecies of Araceae which show a disjunct biogeo-graphical range between these two major humid forest
biomes (Mayo 1983). Finally, previous work (Andrade& Mayo 1998) provided a good understanding of themorphology of the plant in its natural habitat.
Our aim was to estimate quantitatively morpho-logical divergence between the populations with a view to comparison with genetic patterns estimated withmolecular AFLP markers. Using leaf outline morpho-metrics as data for estimating morphological variation(see Methods for further explanation) we focused on thefollowing specific objectives: 1. Comparison of the Ceará brejo populations of Monstera adansonii var. klotzschiana ;2. Estimation of the morphological differences betweenthese populations; 3. Comparison of the Ceará popula-
tions with those in the Atlantic and Amazon forests; 4.Comparison of leaf outline morphology in var. klotzschi-
ana with that of the related taxa in Northeast Brazil,M. adansonii var. laniata (Schott) Madison and M.
praetermissa E. G. Gonç. & Temponi.
Materials and Methods
Leaf outline morphometrics. Two constraints led tothe choice of leaf outline data for the following
reasons. Firstly, to obtain results comparable with ourmolecular study (Andrade et al. 2007), adequately sampled morphological data from the same popula-tions were needed in a form which could be analysedusing similar quantitative techniques. Second, limitedresources for fieldwork and the large geographical
distances between study populations meant that many localities could be sampled only once. These factorsfavoured leaves since, unlike inflorescences, they werenearly always available in suf ficient numbers and easily sampled and preserved. Using a digital camera, leaf outline shapes and dimensions were easily captured inthe field. Digitisation of the leaf outlines from imagesand obtaining mathematical descriptions (using ellip-tic Fourier Analysis) from the resulting coordinatedata was made easy by freely available software(TpsDig: Rohlf (2004); Morpheus et al.: Slice (2008);SHAPE: Iwata & Ukai (2002); download from Rohlf 2006).
Taxonomic determination of Monstera species nor-mally demands observation of characters from inflor-escences as well as vegetative organs (Madison 1977).Fertile voucher specimens were gathered from sam-pled populations whenever found, and identifiedusing the standard revision (Madison 1977); these were deposited permanently as herbarium specimens(see Appendix and Electronic Supplementary Material).Combinations of vegetative characters were used toidentify field-collected specimens of the three taxa of Monstera studied. M. praetermissa is easily distinguishedfrom M. adansonii in non-flowering plants as follows:the mature plant is altogether smaller in stature; the
leaves become dark brown to blackish after drying; theinternodes are much more slender, darker green andlack the abundant whitish speckling of M. adansonii —
instead the epidermis is shallowly verrucate, the verrucae being the same colour as the rest of theinternode. The petiole sheath margins are very quickly marcescent, decomposing into fragile black membra-naceous fragments which are eventually deciduous; inM. adansonii the sheath margins are persistent, greenand herbaceous. The leaf perforations in M. praetermissa
are relatively few and large, extending almost to the very margin of the blade, which often becomes partly pinnatifid by the rupture of the remaining marginal
strip. In M. adansonii leaf perforation patterns are very variable, ranging from absent to extremely abundant and with the individual perforations varying widely insize and shape. However, they never extend right to the very margin and the blade thus does not becomepinnatifid.
Within Monstera adansonii , the differentiation of var.klotzschiana and var. laniata is problematic in fragmen-tary herbarium material, but easier in living fieldpopulations. The most reliable diagnostic charactersare in the flowers: in var. klotzschiana the stigma is
194 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 3/19
elliptic and raised up on a shallow but distinct stylarprominence at the gynoecium apex, whereas in var.laniata the stigma is linear and sessile on a truncate,flattened gynoecial apex. Living non-flowering plantscan be distinguished in the field as follows: var.klotzschiana prefers more shaded and protected hab-
itats under forest canopies and has leathery matureleaf blades which are more than twice as long as wideand ovate-elliptic to oblong-elliptic in shape; the leaf blade base is less asymmetric, usually acute on oneside and obtuse to rounded on the other. Var. laniata
typically grows in more open exposed habitats such ascoastal scrub on rocks or even on fence posts in humidtropical pasture; the mature leaf blades are rather thinin texture, less than twice as long as wide and broadly ovate to ovate-elliptic, with strongly asymmetric leaf blade bases acute on one side and usually truncate tosubcordate on the other. Var. laniata is quite rare innon-Amazonian Brazil, its main range being Central
America, Pacific tropical South America, western Amazonia, Venezuela, the Guianas and northern Amazonian Brazil. Madison (1977) cited no recordsfor eastern or northeastern Brazil, but subsequent fieldwork uncovered a form of var. laniata in the Atlantic forest of southern Bahia and Pernambuco;the Bahian plants are notable for their very numerousand irregular leaf perforations.
The degree to which leaf outline shape is geneti-cally determined is an important issue from a taxo-nomic standpoint. The experimental studies neededto investigate this, involving comparative growth trialsin an experimental garden, were beyond the scope of
this project. However, there is evidence which suggeststhat leaf shape has a genetic basis both in general andin the particular case of Monstera . Kessler & Sinha(2004) reviewed studies of genetic control of leaf shape in model organisms and discussed a range of genetically mediated processes involved in establishing the mature shape of the leaf blade. Studies by Furuta et
al. (1995), Yamanaka et al. (2001) and Iwata et al.
(2002) used principal component shape variablesderived from matrices of Elliptic Fourier coef ficientsto describe leaf shape quantitatively in selectedcultivars and varieties of soybean and citrus. All thesestudies found significant genetic components in the
shape variables, and were furthermore able to show that the variables which describe symmetric changes inleaf shape have a much higher genetic component that the asymmetrical ones. Yoshioka et al. (2004)obtained similar results using petal shape in primulacultivars. The heteroblasty shown by all Monstera
species also indicates a major role for genetic deter-mination in leaf shape in these plants. Juvenile leavesalways differ considerably in shape, size and presenceor pattern of perforations and undergo a predictableseries of shape changes as the plant develops from
seedling to adult; this was used by Madison (1977) asthe basis of his sectional classification of the genus.
Leaf outline capture. 1,695 leaves were sampled fromnatural populations at 32 sites in Brazil and FrenchGuiana, grouped into 20 populations (Fig. 1, Table 1,
Electronic Supplementary Material). At each site,leaves at various stages of the growth cycle werecollected according to availability. To avoid possiblebias caused by clonal growth, care was taken to samplefrom plants that were widely distant and to avoidsampling from plants on the same host tree; thisreduced the scope for making larger samples in somepopulations. Digital images were made using a FujiFinepix A303 digital camera while the leaves were stillfresh. After trimming off most of the petiole, theleaves were attached with map pins to a backboardmade of rubberised carpet tiles with the abaxial sideexposed and photographed using a 15 cm measuring
rule as a scale. One source of variation in shapecapture arose from the fact that the leaves were not strictly two-dimensional, but undulated to variousdegrees especially along the margin. To reduce thiserror, the pinning was carried out so that the leaf undulation was evenly distributed around the marginand the leaves were photographed directly overhead.
The images were prepared for outline digitisationusing the Gimp software package (GIMP 2006).Monstera leaves are distichous and normally oblique,i.e. one half of the leaf blade is wider than the other, with successive leaves showing mirror-symmetry. Eachleaf was rotated so that the apex faced to the left with
the midrib more-or-less horizontal and flipped whennecessary to bring the broadest half of the blade ontothe lower side. The paint tool was used to adjust theimage to provide suf ficient uniformity of tone along the margin. Minor damage to the margin caused by herbivores was also edited out.
The prepared images were then opened in tpsDig (Rohlf 2004, ver. 1.40). The petiole-leaf blade junctionand leaf blade apex were digitised as landmarks 1 and2 respectively. To remove fine scale “noise” along themargin, the image was smoothed 8 to 10 times using the “Smooth image” tool. The thresholded image was viewed and checked using tpsDig ’s Image tools to
capture the outline as precisely as possible by optimis-ing the threshold value. Editing-out of shadows wassometimes necessary at this stage. After these steps,the outline was digitised using the “Outline object ”tool. All the outlines of a population were thus writteninto a single “.tps” file for further processing.
Size classification of leaves. A methodological compli-cation was the conspicuous change in leaf shape and size(heteroblasty) observed in Monstera adansonii during development from juvenile to adult plants (Andrade &
195COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 4/19
Mayo 1998), which is even more pronounced in someother species of the genus (Madison 1977; Ray 1983).The total sample of leaves was therefore divided into
three size classes for analysis so that leaves at approxi-mately the same heteroblastic stage would be compared.
In Monstera adansonii , this shape change is gradualduring the progression from the smallest to largest leaves, but in M. praetermissa a sharp change in shape was observed at about 15 cm length. Using this as aguide, we selected 15 cm leaf blade length as theboundary between juvenile and submature leaves. Theboundary between submature and mature was set at 30 cm leaf blade length after examination of 167 fertileherbarium specimens at the Kew herbarium (K) of M.
adansonii var. klotzschiana and var. laniata , to establish an
approximate lower limit to the leaf blade size in matureflowering shoots.
Elliptic Fourier Analysis. Elliptic Fourier Analysis (EFA)is a method for quantitatively characterising closedoutline shapes and has been used widely in many biological fields, including taxonomy (e.g. Rohlf & Archie 1984; Ferson et al. 1985; McLellan 1993; Cannon& Manos 2001; Rumpunen & Bartish 2002; Jensen et al.
2002; Tatsuta et al. 2004; Yoshioka et al. 2004; Ravelosonet al. 2005). It is particularly useful for shapes whichpossess few clearly homologous landmarks, as in thepresent case, where only the petiole-leaf blade junctionand the leaf blade apex can be regarded as consistent,biologically meaningful homologous sites. We used EFA
KBAS, LBAIlh,
LBAUb
KCEma, KCEpa
KFG, LFG
KCEub, PCEib, PCEub
KPA
KPER, LPET
KBAR, PBAR
KESli, KESst
KRJ
KSP
N
1000 km
PCEbat
Fig. 1. Geographical locations of Monstera populations studied in South America. Monstera adansonii var. klotzschiana. KBAR:Bahia, Recôncavo; KBAS: Bahia, Ubaitaba; KCEma: Ceará, Serra de Maranguape; KCEpa: Ceará, Serra de Aratanha, Pacatuba;KCEub: Ceará, Serra de Ibiapaba; KESli: Espírito Santo, Linhares; KESst: Espírito Santo, Santa Tereza; KFG: French Guiana; KPA:Pará, near Belém; KPER: Pernambuco, Recife; KRJ: Rio de Janeiro, near Rio de Janeiro city; KSP: São Paulo, Bertioga. Monsteraadansonii var. laniata. LPET: Pernambuco, Tapacurá; LBAIlh: Bahia, Ilhéus; LBAUb: Bahia, Ubaitaba; LFG: French Guiana. Monstera
praetermissa. PBAR: Bahia, Serra da Copioba; PCEbat: Ceará, Serra de Baturité; PCEib: Ceará, Serra de Ibiapaba (Ibiapina); PCEub:Ceará, Serra de Ibiapaba (Ubajara).
196 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 5/19
Table 1. Geographical details of sampled populations (20 populations, 1695 leaves) of Monstera adansonii var. klotzschiana, M. adanso
Code and taxon Latitude longitude Population detailsCountry orBrazilian state Total
KBAR Monstera adansonii var. klotzschiana 12°50 ′20 ″S 39°16 ′19 ″ W Recôncavo (mean value for 2 localities) Bahia 43 KBAS " 14°18 ′02 ″S 39°19 ′01 ″ W Ubaitaba Bahia 13 KCEma " 3°48 ′18 ″S 38°42 ′46 ″ W Serra de Maranguape (mean value for
2 localities)Ceará 62
KCEpa " 3°57 ′21 ″S 38°37 ′44 ″ W Serra de Aratanha, Pacatuba Ceará 75 KCEub " 4°01 ′19 ″S 40°51 ′43 ″ W Serra de Ibiapaba (Ibiapina, São Benedito,
Ubajara) (mean value for 3 localities)Ceará 179
KESli " 19°9 ′3 ″S 40°4 ′15 ″ W Linhares Espírito Santo 14 KESst " 19°57 ′57 ″S 40°31 ′58 ″ W Santa Tereza Espírito Santo 40 KFG " 4°37 ′26 ″N 52°32 ′05 ″ W Petit Saut, Kaw, Matoury, Arataí (mean
value for 4 localities)French Guiana 189
KPA " 1°30 ′32 ″S 48°17 ′59 ″ W Acará, Marituba Pará 110 KPER " 8°2 ′45 ″S 34°47 ′00 ″ W Recife (mean value for 2 localities) Pernambuco 61 KRJ " 22°58 ′08 ″S 43°14 ′38 ″ W Rio de Janeiro city area (mean value for
2 localities)Rio de Janeiro 114
KSP " 23°51 ′14 ″S 46°8 ′20 ″ W Bertioga São Paulo 71
LBAIlh Monstera adansonii var. laniata 14°46 ′19 ″S 39°13 ′19 ″ W Ilhéus Bahia 21 LBAUb " 14°18 ′2 ″S 39°19 ′1 ″ W Ubaitaba Bahia 44 LFG " 4°54 ′32 ″N 52°16 ′26 ″ W Cayenne, Fort Diamant, Colline de
Montravel (mean value for 3 localities)French Guiana 316
LPET " 8°0 ′0 ″S 35°10 ′58 ″ W Tapacurá Pernambuco 37
PBAR Monstera praetermissa 12°46 ′41 ″S 39°03 ′38 ″ W Serra da Copioba Bahia 8 PCEbat " 4°16 ′33 ″S 38°57 ′26 ″ W Serra de Baturité Ceará 39 PCEib " 3°55 ′51 ″S 40°52 ′33 ″ W Serra de Ibiapaba (Ibiapina) Ceará 171 PCEub " 3°49 ′58 ″S 40°56 ′23 ″ W Serra de Ibiapaba (Ubajara) Ceará 88
©T h e B o a r d of T r u s t e e s of t h e R
o y a l B o t a ni c G a r d e n s ,K e w ,2 0 0 8

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 6/19
to produce a matrix of 160 coef ficients from which sixshape descriptor variables, the first six principal com-ponents, were derived.
The Fourier analysis was carried out using theMorpheus et al. package (Slice 2008). Before running the Elliptic Fourier Analysis module, the outlines were
standardised using the option “Bookstein superimposi-tion”, which requires that the orientation, scale, star-tingPt and location settings be switched off. The outlinesare then superimposed using two landmarks placed at the base and apex of the leaf blade which equalises thelength and orientation of the interval between theselandmarks. In contrast, the default standardisation inMorpheus involves equalising the length and orienta-tion of the first ellipse of all outlines. The justificationfor using Bookstein superimposition is that the twolandmarks at either end of the midrib represent themost consistent homologies recognisable in the leaves.
The Fourier analysis was carried out to a limit of 40
harmonics, resulting in a matrix of 164 elliptic Fouriercoef ficients. The Fourier procedure reconstructs theoutlines to a level of approximation limited only by the number of pixels used in the original digitisation,so the decision on how many harmonics to set is tosome extent arbitrary (Rohlf & Archie 1984). Aftercomparing the reconstructed outlines by eye with theoriginals we considered that with 40 harmonics, noobvious artefactual distortions would be introducedinto the analyses. The first 4 coef ficients (harmonic 0)are discarded because they correspond to an arbitrary starting point of the outline tracing sequence ratherthan any feature of the leaf shape, so the final matrix
consisted of 160 variables (1st to 40th harmonic, eachharmonic consisting of 4 values).
Multivariate data analyses. Multivariate analyses of theresulting coef ficient matrices were carried out using NTSYSpc ver. 2.20d (Rohlf 2005). Principal component analysis of the coef ficient matrices was used to reducethe dimensionality of the data to a much smallernumber of uncorrelated shape descriptor variables. Eachof these variables was reified using the proceduredescribed by Yoshioka et al. (2004). First, the mean vector of the Fourier coef ficients was computed. Eacheigenvector was then multiplied by integer factors ( f )from 1 to 5. The factored eigenvectors were thenseparately added or subtracted from the mean vector.The mean vector corresponds to the mean shape of thedata set. The effect of adding or subtracting a particularfactored eigenvector is to exaggerate or diminish itseffect on the mean shape while holding all the othereigenvectors constant. The change in principal compo-nent values corresponds to a change of f standarddeviations on the principal component concerned,provided the principal component values have beensuitably scaled. The resulting vectors were then plotted
using the “Fourplot ” module in NTSYSpc, which recon-structs the outline from the set of Fourier coef ficientsprovided. These outlines visualise the shape changesexpressed independently by each of the shape variables(principal components) we used. Figures 2, 3 and 4 show this transformation from −2 to + 2 standard deviations.
We selected the first six principal components (PCs)to be treated as variables in a Canonical Variates Analysis(CVA); together these accounted for 96.6% of the variance in the original covariance matrix. The reduc-tion to six PCs was necessary because once the leaf samples had been divided into their size classes, somepopulation samples were reduced, in one case to only 8 individuals. The aim of the CVA, carried out using NTSYSpc, was to show the maximum separation of thecentroids (multivariate means) of each population. Theresulting eigenvectors were used to project the matrix of principal component scores for all the leaves analysed,onto canonical variate axes. The matrix of pairwise
generalised distance between the population centroids was used to generate Neighbor-Joining (NJ) trees, alsousing NTSYSpc. As a comparison with the pattern basedon generalised distance, the mean vectors of the sixshape variables were computed for each population andtheir pairwise Euclidean distances computed withNTSYSpc. The resulting distance matrix was then usedto carry out a Principal Coordinates Analysis (PCoA),incorporating a minimum spanning tree (MST).
To estimate intra-population variability, the Euclid-ean distance between all pairs of individuals for eachpopulation was computed, using the matrix of 6principal component scores from the combined PCA
of all populations. Analyses of each size class werecarried out separately. The pairwise inter-individualdistances of each population were then summarised by calculating the mean, standard deviation and coef fi-cient of variation (Raveloson et al. 2005). The pairwisedifferences in median Euclidean distance in the threeCeará populations were tested using the non-paramet-ric Kruskal-Wallis test as implemented in PAST (Ham-mer et al. 2001).
Similarity relationships between pairs of popula-tions (based on the matrix of principal component scores) were tested using multivariate proceduresimplemented in PAST: NPMANOVA is a non-paramet-
ric analogue of MANOVA which calculates an F value with significance values computed by 5000 replicatepermutations of group membership (Table 2).
Results
The shape variables. In all three size classes, the first three principal components accounted for over 90%of the total variance (Mature: 92.29%, Submature:
198 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 7/19
1 X
- 0 . 4
9
- 0 . 2
4
0 . 0
1
0 . 2
5
0 . 5
0
- 0 . 1
9
- 0 . 0
9
0 . 0
1
0 . 1
1
0 . 2
1
1 X
- 0 . 4
8
- 0 . 2
3
0 . 0
1
0 . 2
6
0 . 5
0
0 . 2
2
0 . 1
0
0 . 0
1
0 . 1
2
0 . 2
4
1 X
- 0 . 4
7
- 0 . 2
3
0 . 0
2
0 . 2
6
0 . 5
1
0 . 2
5
0 . 1
2
0 . 0
1
0 . 1
4
0 . 2
7
1 X
- 0 . 4
7
- 0 . 2
2
0 . 0
3
0 . 2
7
0 . 5
2
- 0 . 2
9
- 0 . 1
4
0 . 0
1
0 . 1
5
0 . 3
0
1 X
- 0 . 4
9
- 0 . 2
3
0 . 0
3
0 . 2
9
0 . 5
4
- 0 . 3
2
- 0 . 1
6
0 . 0
0
0 . 1
7
0 . 3
3
1 X
- 0 . 5
0
- 0 . 2
5
- 0 . 0
1
0 . 2
4
0
. 4 8
0 . 1
0
0 . 0
4
0 . 0
2
0 . 0
8
0 . 1
4
1 X
- 0 . 4
9
- 0 . 2
4
0 . 0
0
0 . 2
4
0
. 4 9
0 . 1
5
0 . 0
7
0 . 0
2
0 . 1
1
0 . 1
9
1 X
- 0 . 4
6
- 0 . 2
2
0 . 0
3
0 . 2
7
0
. 5 2
0 . 2
8
0 . 1
4
0 . 0
1
0 . 1
5
0 . 2
9 - 0
. 4 8
- 0 . 2
3
0 . 0
1
0 . 2
6
0
. 5 0
0 . 2
2
0 . 1
0
0 . 0
1
0 . 1
3
0 . 2
4 - 0 . 4
7
- 0 . 2
2
0 . 0
3
0 . 2
8
0
. 5 3
0 . 3
4
0 . 1
7
0 . 0
0
0 . 1
7
0 . 3
5
1
- 0 . 5
0
- 0 . 2
5
- 0 . 0
1
0 . 2
4
0 . 4
9
- 0 . 1
0
- 0 . 0
4
0 . 0
1
0 . 0
7
0 . 1
2
1 X
- 0 . 4
9
- 0 . 2
4
0 . 0
0
0 . 2
5
0 . 4
9
- 0 . 1
4
- 0 . 0
6
0 . 0
2
0 . 1
0
0 . 1
8
1 X
- 0 . 4
6
- 0 . 2
2
0 . 0
2
0 . 2
7
0 . 5
1
- 0 . 2
6
- 0 . 1
2
0 . 0
1
0 . 1
5
0 . 2
8 - 0
. 4 8
- 0 . 2
3
0 . 0
1
0 . 2
6
0 . 5
0
- 0 . 2
0
- 0 . 0
9
0 . 0
1
0 . 1
2
0 . 2
3
1 X
- 0 . 4
6
- 0 . 2
1
0 . 0
3
0 . 2
7
0 . 5
2
- 0 . 3
2
- 0 . 1
5
0 . 0
1
0 . 1
7
0 . 3
3
- 1 s.d.- 2 s.d. Mean shape + 2 s.d.+ 1 s.d.
Mature (54.67% of total variance)
Submature (71.28% of total variance)
Juvenile (62.85% of total variance)
Fig. 2. Reification of principal component 1 as shape variable Anisotropy . Derived from principal component analysis of ellipticFourier coefficient matrices of leaf outlines of Monstera adansonii var. klotzschiana, M. adansonii var. laniata and M. praetermissa.Each outline is a 40-harmonic reconstruction from elliptic Fourier analysis data. Each row shows the trend of shape change alongprincipal component 1, in separate PCAs of mature (>=30 cm long), submature (15 to 29.9 cm long) and juvenile (< 15 cm long)leaves. From left to right the outlines represent the principal component scores corresponding to: (mean − 2 standard deviations),(mean − 1 s.d.), mean, (mean + 1 s.d.), (mean + 2 s.d.). Computed with NTSYSpc ver. 2.2.
199COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 8/19
S u b m a t ur e ( 2 1 .4
0 % of t o t al v ar i an c e )
J uv eni l e ( 2 9 .2 3
% of t o t al v ar i an c e )
-0.47 -0.22
-0.47 -0.23
-0.47 -0.23
-0.47 -0.22
0.47 -0.22
1
-0.47 -0.22 0.03 0.29 0.54
.19
.09
.02
.13
.24
1
X
-0.47 -0.23 0.02 0.27 0.51
.20
.09
.02
.13
.24
-0.48 -0.23 0.01 0.26 0.50
.22
.10
.01
.13
.24
1
-0.48 -0.23 0.01 0.26 0.51
.23
.11
.01
.13
.25
-0.48 -0.23 0.02 0.27 0.52
.25
.12
.00
.12
.25
1
X
-0.48 -0.22 0.03 0.28 0.53
-0.19
-0.08
0.02
0.13
0.23
1
-0.47 -0.23 0.02 0.26 0.51
-0.19
-0.09
0.02
0.12
0.23
1
X
-0.48 -0.23 0.01 0.26 0.50
-0.20
-0.09
0.01
0.12
0.23
1
X
-0.48 -0.23 0.02 0.26 0.51
-0.21
-0.10
0.01
0.12
0.23
1
X
-0.49 -0.23 0.02 0.27 0.53
-0.23
-0.11
0.00
0.11
0.23
F i g.
3 .
R e i fi c a t i on of p
r i n c i p a l c om p on e n t 2 a s s h a p e v a r i a b l e
D i mi d i a t e .D e r i v e d f r om
pr i n c i p a l c om p o
n e n t a n a l y s i s of e l l i p t i c
F o ur i e r c o e f fi c i e n t m a t r i c e s of l e a f o u t l i n e s of M on s t e r a a d a n s oni i v a r .k l o t z s c h i a n a ,M. a d a n s oni i v a r .l a ni a
t a a n d M. pr a e t e r mi s s a .
F or f ur t h e r e x pl a n a t i on
s e e F i g.2 .
©T h e B o a r d of T r u s t e e s of t h e R o y a l B o t a ni c G a r d e n s ,K e w ,2 0 0 8

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 9/19
S u b m a t ur e ( 2
.4 5 % of t o t al v ar i an c e )
J uv eni l e ( 3 . 3
3 % of t o t al v ar i an c e )
1
X
-0.49 -0.25 -0.00 0.24 0.49
-0.19
-0.08
0.02
0.13
0.23
1
X
-0.48 -0.24 0.00 0.25 0.49
-0.19
-0.09
0.02
0.12
0.23
1
X
-0.48 -0.23 0.01 0.26 0.50
-0.20
-0.09
0.01
0.12
0.23
1
X
-0.47 -0.22 0.02 0.26 0.51
-0.21
-0.10
0.01
0.12
0.23
1
X
-0.46 -0.21 0.03 0.27 0.52
-0.21
-0.10
0.01
0.12
0.24
1
X
-0.49 -0.25 -0.00 0.24 0.48
.21
.09
.02
.14
.25
1
X
-0.48 -0.24 0.00 0.25 0.49
.21
.10
.02
.13
.25
1
X
-0.48 -0.23 0.01 0.26 0.50
.22
.10
.01
.13
.24
1
X
-0.47 -0.22 0.02 0.27 0.51
.22
.10
.01
.13
.24
1
X
-0.46 -0.22 0.03 0.28 0.52
.22
.11
.01
.13
.24
-0.46 -0.21
-0.46 -0.22
-0.47 -0.23
-0.48 -0.23
-0.49 -0.24
.24
.12
.01
.14
.26
F i g.4 .R e i fi c a t i on o
f pr i n c i p a l c om p on e n t 3 a s s h a p e v a r i a b l e Ov a t e .D e r i v e d f r om pr i n c i p a l c om p on e
n t a n a l y s i s of e l l i p t i c F o ur i e r
c o e f fi c i e n t m a t r i c e s
of l e a f o u t l i n e s of M on s t e r a a d a n s oni i v a r .k l o t z s c h i a n a ,M. a d a n s oni i v a r .l a ni a t a a n d M. pr a e t e r mi s s a .F or
f ur t h e r e x pl a n a t i on
s e e F i g.2 .
©T h e B o a r d of T r u s t e e s of t h e R o y a l B o t a ni c G a r d e n s ,K e w ,2 0 0 8

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 10/19
95.13%, Juvenile: 95.41%) and described distinct andeasily recognised trends of leaf outline shape change which are illustrated in Figs. 2, 3 and 4. The fourth,fi
fth and sixth principal components expressed ob-scure shape variations that were not easily interpreted visually.
The first principal component is named here theAnisotropy shape variable (Fig. 2), because its most conspicuous characteristic is the increase in length/ width ratio. This shape variable also expresses anasymmetry at the leaf base which becomes morepronounced in the transition from juvenile to mature
leaves. The second principal component, here termed Dimidiate shape variable (Fig. 3) is characterisedmainly by the increasing difference in the width and
area of the two sides of the leaf blade, one sideeventually becoming very narrow. A secondary featureis a slight basiscopic migration of the axis of widest width. The third principal component, the Ovate
shape variable (Fig. 4), is characterised chiefl y by themigration of the widest width axis giving a trend fromelliptic to ovate or vice versa (the orientation of theprincipal component axes between analyses is arbi-trary). A slight inverse dimidiate trend is also evident,
Table 2. Inter-population differences between nine populations of Monstera adansonii var. klotzschiana based on matrix of sixshape variables derived from elliptic Fourier analysis of mature leaf outlines.
KBAR KBAS KCEma KCEpa KCEub KFG KPA KPER KSP
KBAR –
KBAS 0.009 –
KCEma 0.016 n.s. –
KCEpa 0.000 n.s. 0.000 –
KCEub 0.048 0.027 0.003 0.000 –
KFG n.s. 0.011 0.011 0.000 0.009 –
KPA n.s. 0.027 0.011 0.000 n.s. n.s. –
KPER n.s. 0.006 n.s. 0.000 0.002 0.020 0.042 –
KSP n.s. 0.002 0.000 0.000 n.s. 0.004 n.s. 0.001 –
Values shown are p -values (probability that two groups are the same) derived from Non-Parametric MANOVA implemented in PAST ver.1.34 (Hammer et al. 2001); 5,000 replicates computed. See Table 1 for key to population codes.
0
0.001
0.002
0.003
0.004
0.005
0.006
0.007
0.008
PCEub PCEib LFG KCEpa KCEub KFG KPER LPET KCEma KPA LBAUb KBAR KSP
Populations
M e a n i n t e r - i n d i v i d u a l E u c l i d e a n d i s t a n c e p e r p o p u l a t i o n
mature submature juvenile
Fig. 5. Intra-population variability of leaf shape in Monstera adansonii var. klotzschiana, M. adansonii var. laniata and M. praetermissa. For each population, the mean Euclidean distance between pairs of individuals is shown for three size classes (left bar :mature leaves; middle bar : submature leaves; right bar [when data available]: juvenile leaves). Data is matrix of individual scores onsix shape variables (principal components) derived from EFA coefficients of leaf outlines. 95% confidence limits shown. See Fig. 1for key to population codes.
202 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008
8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 11/19
i .e. the more ovate the leaf becomes, the less
dimidiate: this secondary trend is more distinct inthe submature and juvenile size classes.
Comparison of intrapopulational variability in Ceará
populations of Monstera adansonii var. klotzschiana . Juvenile leaves were only available for the Ubajarapopulation (KCEub), so comparisons of the three Ceará populations were made with submature and matureleaves. Using mean Euclidean distance between pairs of individuals as a measure of population variability,submature leaves were more variable than mature leaves(Fig. 5), with the largest discrepancy in size class variability shown in the Pacatuba population (KCEpa).
Comparison of the mature leaves showed significantly greater variability in the Maranguape (KCEma) popu-lation compared to the other two (Pacatuba: KCEpa;Ubajara: KCEub), and non-significant difference in the variability of the Pacatuba and Ubajara populations.
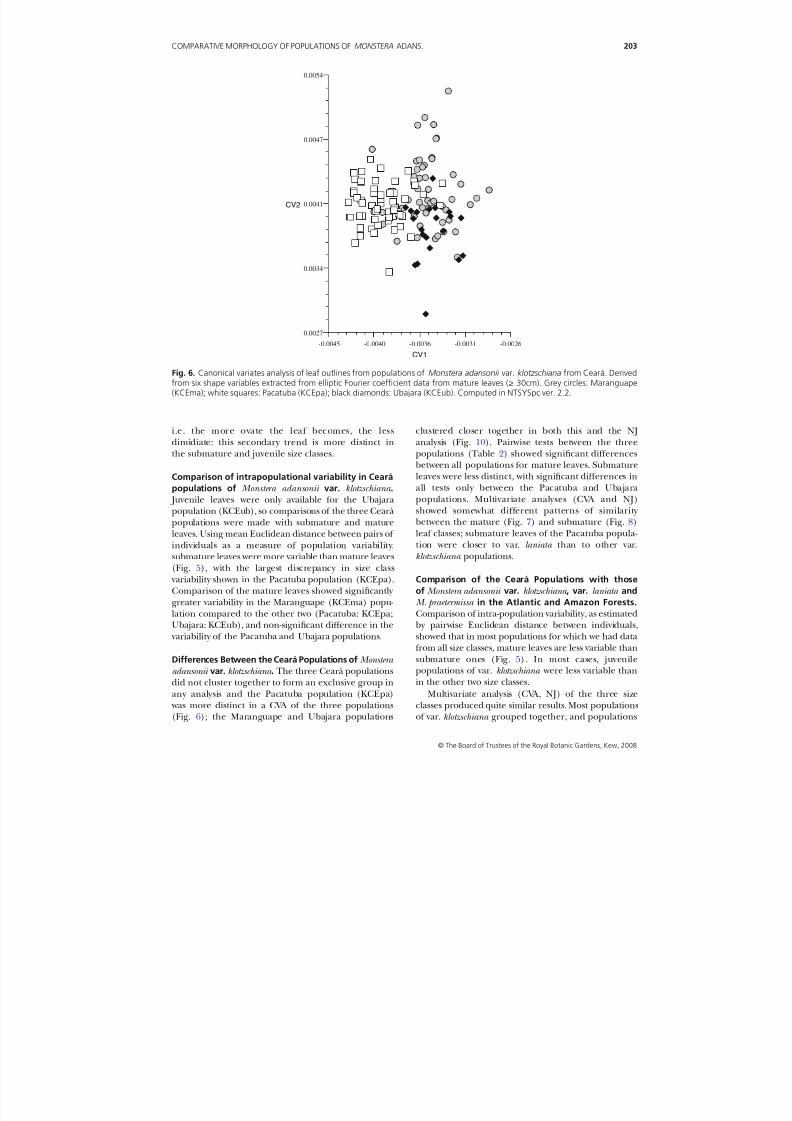
Differences Between the Ceará Populations of Monstera
adansonii var. klotzschiana . The three Ceará populationsdid not cluster together to form an exclusive group inany analysis and the Pacatuba population (KCEpa) was more distinct in a CVA of the three populations(Fig. 6); the Maranguape and Ubajara populations
clustered closer together in both this and the NJ
analysis (Fig. 10). Pairwise tests between the threepopulations (Table 2) showed significant differencesbetween all populations for mature leaves. Submatureleaves were less distinct, with significant differences inall tests only between the Pacatuba and Ubajarapopulations. Multivariate analyses (CVA and NJ)showed somewhat different patterns of similarity between the mature (Fig. 7) and submature (Fig. 8)leaf classes; submature leaves of the Pacatuba popula-tion were closer to var. laniata than to other var.klotzschiana populations.
Comparison of the Ceará Populations with those
of Monstera adansonii var. klotzschiana , var. laniata andM. praetermissa in the Atlantic and Amazon Forests.
Comparison of intra-population variability, as estimatedby pairwise Euclidean distance between individuals,showed that in most populations for which we had datafrom all size classes, mature leaves are less variable thansubmature ones (Fig. 5). In most cases, juvenilepopulations of var. klotzschiana were less variable thanin the other two size classes.
Multivariate analysis (CVA, NJ) of the three sizeclasses produced quite similar results. Most populationsof var. klotzschiana grouped together, and populations
CV1
-0.0045 -0.0040 -0.0036 -0.0031 -0.0026
CV2
0.0027
0.0034
0.0041
0.0047
0.0054
Fig. 6. Canonical variates analysis of leaf outlines from populations of Monstera adansonii var. klotzschiana from Ceará. Derivedfrom six shape variables extracted from elliptic Fourier coefficient data from mature leaves (≥ 30cm). Grey circles: Maranguape(KCEma); white squares: Pacatuba (KCEpa); black diamonds: Ubajara (KCEub). Computed in NTSYSpc ver. 2.2.
203COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 12/19
CV1
-3.95 -2.56 -1.17 0.21 1.60
CV2
-2.50
-1.52
-0.55
0.42
1.40
KBAR
KBAS
KCEma
KCEpa
KCEub
KFGKPA
KPER
LPET
KSP
LBAUb
LFG
PBAR
PCEib
PCEub
1
- 0 . 4
7
- 0 . 2 2
0 . 0 2
0 . 2 7
0 . 5
2
5 1 4 7
1 X
- 0 . 4
7
- 0 . 2
2
0 . 0
3
0 . 2 7
0 . 5
2
. 2 3
. 1 1
. 0 2
. 1 4
. 2 6
- 0 . 4 6
- 0 . 2
2
0 . 0 3
0 . 2 8
0 .
. 3 0
. 1 5
. 0 0
. 1 5
. 3
1
- 0 . 4 6
- 0 . 2 1
0 . 0 4
0 . 2 8
0 . 5
3
- 0 . 2
8
- 0 . 1
4
0 . 0
0
0 . 1
5
0 . 2
9
1 X
- 0 . 4 7
- 0 . 2 2
0 . 0 2
0 . 2 7
0 . 5 1
. 2 8 . 1 4
. 0 0 . 1 5
. 2 9
1
- 0 . 4
6
- 0 . 2 2
0 . 0
3
0 . 2 8
0 . 5
3
9 5 5
1 X
- 0 . 4 7
- 0 . 2
3
0 . 0
2
0 . 2
7
0 . 5 1
2 5 1 2 0 1 1 3 2 6
1
- 0 . 4
8
- 0 . 2
3
0 . 0
2
0 . 2
7
0 . 5
2
. 2 6 . 1 3
. 0 0 . 1 4
. 2 7
1 X
- 0 . 4
8
- 0 . 2 3
0 . 0
2
0 . 2
7
0 . 5
2
- 0 . 2
5
- 0 . 1
2
0 . 0
1
0 . 1
3
0 . 2
6
1 X
- 0 . 4
8
- 0 . 2
3
0 . 0
1
0 . 2 6
0 . 5
1
0 . 2
2
0 . 1
1
0 . 0
1
0 . 1
2
0 . 2
4
1
- 0 . 4
7
- 0 . 2
2
0 . 0 2
0 . 2
7
0 . 5 1
2 2
1 0 2
1 4
2 6
- 0 . 4 8
- 0 . 2
4
0 . 0 1
0 . 2 6
0 . 5 0
6 1 3 0
1 4 7
1 X
- 0 . 4
8
- 0 . 2
4
0 . 0
1
0 . 2 6
0 . 5 0
3 1 1 3 5
1 X
- 0 . 4
7
- 0 . 2
2
0 . 0
2
0 . 2
7
0 . 5
1
. 2 5
. 1 3
0 . 0
0 . 1
3 . 2
6
1 X
- 0 . 4 8
- 0 . 2 3
0 . 0 2
0 . 2 7
0 . 5 1
Y
- 0 . 2 4
- 0 . 1 2
0 . 0 1
0 . 1 3
0 . 2 6
CV1
-3.87 -1.94 -0.01 1.91 3.84
CV2
-2.88
-1.85
-0.82
0.21
1.25
PC1
PC2
PC3
PC4
PC5
PC6
Fig. 7. Canonical variates analysis of mature leaf outlines in nine populations of Monstera adansonii var. klotzschiana (KBAR, KBAS,KCEma, KCEpa, KCEub, KFG, KPA, KPER, KSP), three populations of M. adansonii var. laniata (LBAUb, LFG, LPET) and threepopulations of M. praetermissa (PBAR, PCEib, PCEub). Plot shows mean population shapes. CVA computed from six shape variables(principal components) extracted by principal components analysis from a matrix of elliptic Fourier coefficients. Lower figure showsvector diagram of eigenvectors in relation to the first two canonical variates axes. Computed with NTSYSpc ver. 2.2.
204 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 13/19
CV1
-1.95 1.03 2.51 4.00
CV2
-1.90
0.14
1.16
2.1
8
KBAR
KCEma
KCEpa
KCEub
KESli
KESst
KFG
KPA
KPER
LPET
KRJ
KSP
LBAIlh
LBAUb
LFG
PCEbat
PCEib
PCEub
X
- 6 . 8
2
- 3 . 3
5
0 . 1
1
3 . 5 8
7 . 0
4
0 7 6 8 1
1 X
- 5 . 2
4
- 2 . 5
3
0 . 1
8
2 . 8
9
5 . 6
0
6 0 3 4 1 8
1 X
- 6 . 4
4
- 3 . 0
1
0 . 4
2
3 . 8
5
7 . 2
8
7 2 3 9 4
- 4 3
. 1 0
- 2 1
. 0 4
1 . 0
2
2 3
. 0 8
4 5
. 1 3
6
- 0 . 4 8
- 0 . 2 3
0 . 0 1
0 . 2
6
0 . 5
1
3 1 1 3 5
X
- 0 . 4
8
- 0 . 2
4
0 . 0
1
0 . 2 6
0 . 5
0
0 0 1 1 2
X
- 0 . 4
7
- 0 . 2
2
0 . 0
2
0 . 2
7
0 . 5
2
4 1 0 1 3 6
1 X
- 0 . 4
8
- 0 . 2
4
0 . 0
1
0 . 2
6
0 . 5
1
4 1 1 3 5
1
- 0 . 4
7
- 0 . 2
2
0 . 0
3
0 . 2
7
0 . 5
2
3 1 1 4 6
1 X
- 0 . 4
9
- 0 . 2
5
0 . 0
0
0 . 2
5
0 . 5
0
. 1 1
. 0 4
. 0 2
. 0 9
. 1 6
1 X
- 0 . 4
8
- 0 . 2 4
0 . 0 1
0 . 2
6
0 . 5 0
. 1 6 . 0 6
. 0 3 . 1 2
. 2 1
1 X
- 0 . 4 8
- 0 . 2
4
0 . 0 1
0 . 2 6
0 . 5
0
6 7 2 2 1
- 0 . 4
9
- 0 . 2 4
0 . 0 0
0 . 2 5
0 . 5
0
1
- 0 . 4
9
- 0 . 2
4
0 . 0
1
0 . 2
5
0 . 5
0
9 9 1 1 1
1
- 0 . 4
9
- 0 . 2
4
0 . 0
0
0 . 2
5
0 . 5
0
7 0 8
0 1 0 9
1 X
- 0 . 4
9
- 0 . 2
4
0 . 0
1
0 . 2
5
0 . 5
0
0 9 1 2 2
- 0 . 4
6
- 0 . 2
2
0 . 0
3
0 . 2
7
0 . 5
2
9 4 1 6 1
1 X
- 0 . 4
6
- 0 . 2
1
0 . 0
3
0 . 2
8
0 . 5
3
7 3 1 4 8
CV1
-1.95 -0.46 1.03 2.51 4.00
CV2
-1.90
-0.85
0.20
1.25
2.30
PC1
PC2
PC3
PC4
PC5
PC6
Fig. 8. Canonical variates analysis of submature leaf outlines in eleven populations of Monstera adansonii var. klotzschiana (KBAR,KCEma, KCEpa, KCEub, KESli, KESst, KFG, KPA, KPER, KSP), four populations of M. adansonii var. laniata (LBAIlh, LBAUb, LFG,LPET) and three populations of M. praetermissa (PCEbat, PCEib, PCEub). For further explanation see Fig. 7.
205COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 14/19
CV1
-3.80 -2.50 -1.19 0.11 1.41
CV2
-2.95
-1.54
-0.13
1.29
2.70
KBAR
KCEub
KESst
KFG
KPA
KRJ
KSP
LBAUb
LFG
PCEbat
PCEib
1 X
- 0 . 4
9
- 0 . 2
4
0 . 0
1
0 . 2 6
0 . 5
0
0 . 1 7
0 . 0 8
0 . 0 2
0 . 1 1
0 . 2 0
X
- 0 . 4 7
- 0 . 2
2
0 . 0
2
0 . 2 7
0 . 5 1
2 3 1 1 0 1 1 2 2 4
1 X
- 0 . 4
9
- 0 . 2
4
0 . 0
1
0 . 2
6
0 . 5
0
1 8
0 8
0 1
1 0
1 9
1 X
- 0 . 4
8
- 0
. 2 3
0 . 0
1
0 . 2
6
0 . 5
0
2 1 1 1
1 3 5
- 0 . 4
9
- 0 . 2 4
0 . 0
1
0 . 2
5
0 . 5 0
1 3
0 6
0 1
0 8
1 6
X
- 0 . 4
8
- 0 . 2
4
0 . 0
1
0 . 2 6
0 . 5 0
0 1 X
- 0 . 4
8
- 0 . 2
4
0 . 0
1
0 . 2 6
0 . 5
0
1 X
- 0 . 4
7
- 0 . 2 2
0 . 0
2
0 . 2 7
0 . 5 1
2 1 1 0 0 1 1 3 2 4
X
- 0 . 4 6
- 0 . 2
1
0 . 0
4
0 . 2
9
0 . 5 3
2 7
1 3
0 1
1 5
2 9
1 X
- 0 . 4 9
-
0 . 2 4
0 . 0
0
0 . 2 5
0 . 5 0
1 2 0 5 0 2 0 9 1 6
1 X
- 0 . 4
7
- 0 . 2
3
0 . 0
2
0 . 2
6
0 . 5
1
1 8
0 8
0 2
1 2
2 2
CV1
-3.65 -2.12 -0.59 0.93 2.46
CV2
-2.87
-1.52
-0.17
1.18
2.53
PC1
PC2
PC3
PC4
PC5
PC6
Fig. 9. Canonical variates analysis of juvenile leaf outlines in seven populations of Monstera adansonii var. klotzschiana (KBAR,KCEub, KESst, KFG, KPA, KRJ, KSP), two populations of M. adansonii var. laniata (LBAUb, LFG) and two populations of M.
praetermissa (PCEbat, PCEib). For further explanation see Fig. 7.
206 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 15/19
of var. laniata and Monstera praetermissa formed sepa-rate groups (Figs. 7, 8, 9 and 10). These taxonomically correlated clusters were most distinct in mature leaves(Fig. 7), with all populations of var. klotzschiana
forming a single cluster.
Pairwise NPMANOVA tests of mature-leaf popula-tions of var. klotzschiana (Table 2) showed the most significant differences in pairs involving the Pacatubapopulation (KCEpa). The results of these tests werebroadly consistent with a PCoA ordination of themean vectors of the six shape variable scores for each
of the nine var. klotzschiana populations (Fig. 11). Thelatter plot emphasised the disparity between the threebrejo populations in Ceará.
Mantel tests computed in NTSYSpc ver. 2.2 foundno significant correlation between leaf outline shape(measured as Euclidean distance between populationmean vectors of the six shape variables) and geo-graphical distance between populations [10,000 ran-dom permutations; p (random Z <= observed Z ) =0.6305, p (random Z >= observed Z ) = 0.3695; matrixcorrelation (normalised Mantel statistic Z ): r = 0.06712]or between leaf outline and genetic distance [data from
Andrade et al. 2007, genetic distance estimated aspairwise F ST ; 10,000 random permutations; p (randomZ <= observed Z ) = 0.3308, p (random Z >= observedZ ) = 0.6692; matrix correlation (normalised Mantelstatistic Z ): r = −0.1158].
DiscussionNo strong geographical patterns emerged from theanalyses of var. klotzschiana populations. This contrasted with the results obtained in our AFLP analysis (Andrade
et al. 2007), which showed a clear indication of greatergenetic diversity in Northeast Brazilian and Amazonianpopulations and a significant correlation of genetic andgeographical distance. The Ceará brejo populations of var. klotzschiana emerge from this study as distinct fromone another, at least using mature leaves. Althoughthere is considerable overlap (Fig. 5), their shapes aresignificantly different (Table 2). The Maranguapepopulation is more variable but the Pacatuba popula-tion is the most distinct within Ceará (Figs. 5, 8, 10).The study shows that in var. klotzschiana , leaf outlinemorphology expresses a degree of inter-population
Coefficient
0.00 0.65 1.29 1.94 2.58
KBAR
KPA
KSP
KBAS
KCEub
KCEma
KPER
KFG
KCEpa
LPET
LBAUb
LFG
PBAR
PCEib
PCEub
Fig. 10. Inter-population morphological distance of mature leaf outlines in nine populations of Monstera adansonii var.klotzschiana (KBAR, KBAS, KCEma, KCEpa, KCEub, KFG, KPER, KSP), three populations of M. adansonii var. laniata (LBAUb, LFG,LPET) and three populations of M. praetermissa (PBAR, PCEib, PCEub). Neighbor-Joining tree computed from a matrix ofgeneralised distance between population centroids derived from canonical variates analysis of six shape variables. Shape variables(principal components) computed by principal components analysis from a matrix of elliptic Fourier coefficients. Computed withNTSYSpc ver. 2.2.
207COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 16/19
divergence which is often significant (Table 2). All themost significant inter-population differences involvedCeará populations, and the degree of morphologicaldivergence between the Ceará brejo populations isgreater than between most population pairs from themajor forest blocks in Amazonia and the AtlanticForest). The Ceará populations do not cluster togeth-er as an exclusive group, but rather are mixed with
populations from Amazonia and the Atlantic forest without an obvious pattern (Fig. 10). Broadly speak-ing, the leaf outline data was successful as a markerfor the taxa (species and varieties) but showed noclear geographical patterns at population level within the best sampled taxon, var. klotzschiana . Thiscontrasts with our AFLP results (Andrade et al. 2007),in which the molecular data revealed clear geograph-ical structure.
This study demonstrates that leaf outline shapealone is suf ficiently effective as a taxonomic marker tocluster the populations of these three taxa into theircorrect hierarchical taxonomic order successfully,
using mature leaves. Although population differences were weaker, significant divergence was observed. Leaf outline shape, treated quantitatively can be a usefultool for taxonomic comparisons down to populationlevel. This is somewhat surprising to anyone familiar with the great variability of the leaves of Monstera
adansonii . Even more surprising is that the smaller leaf size classes are almost as successful in discriminating the taxa.
In var. klotzschiana the generally lower variability of juvenile leaf shape, combined with its power todiscriminate the three taxa, is potentially of practical
value. In the damaged brejo forests, we observed that where the forest had lost its continuous canopy through human disturbance (e.g. São Benedito),root-climbers like Monstera were represented almost entirely by terrestrial and rupicolous juvenile forms,because of the absence of suitable host trees. In thissituation, discrimination of juvenile forms of different species is of practical importance in making invento-
ries of endangered biodiversity.Clearly, a wider range of quantitative characters isneeded to “capture” the morphology more completely.Leaf shape plays a significant (but by no meansexclusive) role in the taxonomic definition of the taxa we studied and therefore our finding that leaf outlineshape discriminates the taxa could be seen as a circularargument. In our defence however, we would point out that our field identification of vegetative individualsrelied on a range of other characters including habitat,stem morphology, leaf texture, leaf perforation pat-terns and colour of the dried leaves. Furthermore thisstudy used quantitative estimates of variability and
difference in leaf shape and thus at the very least it provides verification of grouping behaviour of amorphological feature that is usually deployed intui-tively in alpha taxonomy. Working from field-collecteddigital images proved to be a practical and effective way to gather large samples from which quantitative datacould be captured semi-automatically. If extended toother quantitative characters recoverable from images,this approach could bring detailed and sophisticatedmorphological studies at population level within thereach of many workers, making it possible to work fromlarge samples without permanently damaging popula-
PCo1
-0.0026 -0.0014 -0.0003 0.0009 0.0021
PCo2
-0.0021
-0.0011
-0.0002
0.0008
0.0018
KBAR
KBAS
KCEma
KCEpa
KCEub
KFG
KPA KPER
KSP
Fig. 11. Inter-population morphological distance of mature leaf outlines in nine populations of Monstera adansonii var.klotzschiana (KBAR, KBAS, KCEma, KCEpa, KCEub, KFG, KPER, KSP). Principal coordinates analysis (PCoA) with minimum spanningtree superimposed computed from a matrix of Euclidean distance between population mean vectors, derived from a matrix of sixshape variables. Shape variables (principal components) computed by principal components analysis from a matrix of elliptic Fourier
coefficients. Computed with NTSYSpc ver. 2.2.
208 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 17/19
tions by removing significant numbers of plants forherbarium vouchering. This is increasingly a concernfor biologists who study surviving populations in smalland threatened habitats like the brejo forests of Northeast Brazil.
Acknowledgements We are most grateful to the following organisations andpersons for their essential help: FUNCAP (FundaçaoCearense de Amparo à Pesquisa), FAPESB (Fundaçãode Amparo a Pesquisa do Estado da Bahia, WWF Brasil,Kew Latin America Research Fellowships Programme(Royal Botanic Gardens, Kew), Margaret Mee Fellow-ships Programme (Fundação Botânica Margaret Mee,Rio de Janeiro, Royal Botanic Gardens, Kew), Minis-tério do Meio Ambiente (IBAMA, permit number 001/2004, 042/04), Conselho Nacional de Desenvolvi-mento Científica e Tecnológica (CNPq, MCT permit
number, 168/01), Dr Ana Maria Giulietti, Dr RobynCowan, Anna Haigh, Dr Jean-Jacques de Granville andthe staff of the CAY herbarium in French Guiana,Bertrand Goguillon and the staff of IRD (FrenchGuiana) for permission to collect, Dr Denis Barabé,Thales Alves Ribeiro, Sr. José Teodoro Soares, Rectorof the Universidade Estadual do Vale do Acaraú (UVA),Sobral-CE.
Appendix
List of voucher specimens.
Voucher specimens are deposited in the following herbaria: EAC = Universidade Federal do Ceará,Fortaleza, Ceará, Brazil; HUEFS = Universidade Estadualde Feira de Santana, Bahia, Brazil; K = Royal BotanicGardens, Kew, United Kingdom; UVA = Herbarium of the Universidade Estadual Vale do Acaraú, Sobral, Ceará.Population codes are given in bold.
Monstera adansonii Schott var. klotzschiana (Schott)MadisonKBAR : Brazil, Bahia, Serra da Copioba: I. M. Andrade
2771 (HUEFS), I. M. Andrade 2772 (HUEFS), I. M.
Andrade 2783 (HUEFS), I. M. Andrade 2784 (EAC),I. M. Andrade 1996 (HUEFS).KBAS: Brazil, Bahia, Ubaitaba: I. M. Andrade 2075(HUEFS), I. M. Andrade 2077 (EAC), I. M. Andrade
2078 (HUEFS), I. M. Andrade 2079 (HUEFS).KCEma: Brazil, Ceará, Maranguape: I. M. Andrade 867(UVA).KCEpa : Brazil, Ceará, Pacatuba: I. M. Andrade 685(HUEFS), I. M. Andrade 706 (EAC), I. M. Andrade 707(HUEFS).KCEub: Brazil, Ceará, São Benedito: I. M. Andrade 602(EAC), I. M. Andrade 605 (EAC), I. M. Andrade 2613
(HUEFS). Ubajara: I. M. Andrade 2677 (HUEFS), I. M.
Andrade 2680 (HUEFS), I. M. Andrade 2709 (HUEFS),I. M. Andrade 2722 (HUEFS).KESli: Brazil, Espírito Santo, Linhares: I. M. Andrade
1618 (EAC), I. M. Andrade 1622 (EAC), I. M. Andrade
1628 (EAC).
KESst: Brazil, Espírito Santo, Santa Tereza: I. M. Andrade 764 (HUEFS), I. M. Andrade 767 (HUEFS).KFG: French Guiana, Petit Saut: I. M. Andrade 2800(EAC, HUEFS, K), I. M. Andrade 2802 (EAC, HUEFS, K).KPA : Pará, Acará: I. M. Andrade 2385 (HUEFS), I. M.
Andrade 2388 (HUEFS).KPER : Brazil, Pernambuco, Recife: I. M. Andrade 1881(HUEFS).KRJ: Brazil, Rio de Janeiro, Vista Chinesa: I. M. Andrade
2222 (HUEFS), I. M. Andrade 2228 (HUEFS), I. M.
Andrade 2230 (HUEFS).KSP: Brazil, São Paulo, Bertioga: I. M. Andrade 2121(HUEFS), I. M. Andrade 2137 (HUEFS).
Monstera adansonii Schott var. laniata (Schott) MadisonLBAIlh: Brazil, Bahia, Ilhéus, Centro de Pesquisas doCacau (CEPEC): I. M. Andrade 1983 (UVA).LBAUb: Brazil, Bahia, Ubaitaba: I. M. Andrade 1962(UVA).LFG: French Guiana, Cayenne: I. M. Andrade 2815 (K).LPET: Brazil, Pernambuco, Tapacurá: I. M. Andrade
1933 (UVA).
Monstera praetermissa E. G. Gonç. & TemponiPBAR: Brazil, Bahia, Serra da Copioba: I. M. Andrade
2740 (UVA).PCEbat: Brazil, Ceará, Serra de Baturité, I. M. Andrade
s.n. (UVA).PCEib: Brazil, Ceará, Ibiapina, I. M. Andrade 2529 (UVA).PCEub: Brazil, Ceará, Ubajara, I. M. Andrade 2630 (UVA).
References Ab’Sáber, A. N. (1982). The paleoclimate and paleo-
ecology of Brazilian Amazônia. In: G. T. Prance(ed.), Biological diversification in the Tropics.Columbia University Press, New York. Pp. 41 – 59.
Andrade, I. M. & Mayo, S. J. (1998). Dynamic shoot morphology in Monstera adansonii Schott var.klotzschiana (Schott) Madison (Araceae ). Kew Bull.532: 399 – 417.
____, ____, Van den Berg, C., Fay, M., Chester, M.,Lexer, C. & Kirkup, D. (2007). Genetic variation inpopulations of Monstera adansonii Schott (Araceae )from natural forest fragments in Northeast Brazilestimated with AFLP molecular markers. Ann. Bot.100: 1143 – 1154.
Andrade-Lima, D. de (1982). Present-Day Forest Refuges in Northeastern Brasil. In: G. T. Prance
209COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 18/19
(ed.), Biological Diversification in the Tropics.Columbia University Press, New York. Pp. 245 –
251.Borges-Nojosa, D. M. & Caramaschi, U. (2003). Com-
posição e Análise Comparativa da Diversidade e das A finidades Biogeográ ficas dos Lagartos e Anfisbení-
deos (Squamata) dos Brejos Nordestinos. In: I. Leal, J. M. C. Silva & M. Tabarelli (eds.), Ecologia eConservação da Caatinga. ed. 1, 1: 489 – 540. Recife.
Cannon, C. H. & Manos, P. S. (2001). Combining andcomparing morphometric shape descriptors with amolecular phylogeny: the case of fruit type evolu-tion in Bornean Lithocarpus ( Fagaceae ). Syst. Biol. 50(6): 860 – 880.
Ferson, S., Rohlf, F. J. & Koehn, R. K. (1985).Measuring shape variation of two-dimensional out-lines. Syst. Zool. 34(1): 59 – 68.
Figueiredo, M. A. (1997). Vegetação. In: FundaçãoInstituto de Planejamento do Ceará (IPLANCE).
Atlas do Ceará. Fortaleza. Pp. 28 – 29.Furuta, N., Ninomiya, S., Takahashi, S., Ohmori, H. &
Ukai, Y. (1995). Quantitative evaluation of soybean(Glycine max (L.) Merr.) leaflet shape by principalcomponent scores based on elliptic Fourier descrip-tors. Breeding Sci. 45: 315 – 320.
GIMP (2006). http://www.gimp.org . Version 2.2.4 by S. Kimball & P. Mattis.
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. (2001).PAST: Palaeontological Statistics software package foreducation and data analysis. Palaeontologia Electron-ica 41: 9; http://folk.uio.no/ohammer/past
IBGE (1985). Atlas nacional do Brasil. IBGE. Rio de
Janeiro. ____ (2005) http://www.ibge.gov.br/home/presidencia/
noticias/noticia_impressão.php?id_noticia 06/07/2005
Iwata, H. & Ukai, Y. (2002). SHAPE: A computerprogram package for quantitative evaluation of biological shapes based on elliptic Fourier descrip-tors. J. Hered. 93: 384 – 385.
____, Nesumi, H., Ninomiya, S., Takano, Y. & Ukai, Y.(2002). The evaluation of genotype x environment interactions of citrus leaf morphology using imageanalysis and elliptic Fourier descriptors. Breeding Sci. 52: 243 – 251.
Jensen, R. J., Ciofani, K. M. & Miramontes, L. C. (2002).Lines, outlines, and landmarks: morphometric analy-ses of leaves of Acer rubrum , Acer saccharinum
(Aceraceae ) and their hybrid. Taxon 51: 475 – 492.Kessler, S. & Sinha, N. (2004). Shaping up: the genetic
control of leaf shape. Curr. Opin. Pl. Biol. 7: 65 – 72.Madison, M. (1977). A revision of Monstera (Araceae ).
Contr. Gray Herb. 207: 1 – 101.Mayo, S. J. (1983). Aspectos da fitogeografia das Araceae
bahianas. In: Anais XXXIV Congresso Nacional de
Botânica do Brasil, Porto Alegre, Brasil 2: 215 –
227. Diretoria da Sociedade, Porte Alegre.McLellan, T. (1993). The roles of heterochrony and
heteroblasty in the diversification of leaf shapes inBegonia dregei (Begoniaceae ). Amer. J. Bot. 807: 796 –
804.
Oliveira, P. E., Barreto, A. M. F. & Suguio, K. (1999).Late Pleistocene/Holocene climatic and vegeta-tional history of the Brazilian caatinga: the fossildunes of the middle São Francisco River. Palaeo-geogr., Palaeoclimatol., Palaeoecol. 152: 319 –
337.Pôrto, K. C., Cabral, J. J. P. & Tabarelli, M. (2004).
Brejos de altitude em Pernambuco e Paraíba:história natural, ecologia e conservação. Ministériodo Meio Ambiente, Brasília.
Raveloson, H., Le Minor, J.-M., Rumpler, Y. &Schmittbuhl, M. (2005). Shape of the lateral mandib-ular outline in Lemuridae : a quantitative analysis of
variability using elliptical Fourier analysis. FoliaPrimatol. 76: 245 – 261.
Ray, T. (1983). Monstera tenuis (Chirravaca, Mano deTigre, Monstera). Pp. 278 – 280 In: D. H. Janzen,Costa Rican Natural History. University of ChicagoPress, Chicago.
Rodal, M. J. N., Sales, M. F. & Mayo, S. J. (1998).Florestas Serranas de Pernambuco: Localização dosremanescentes dos brejos de altitude. ImprensaUniversitária, Universidade Federal Rural de Per-nambuco, Recife.
Rohlf, F. J. (2004). TpsDig Version 1.40. Digitizing software. http://life.bio.sunysb.edu/morph/
____ (2005). NTSYSpc: Numerical Taxonomy System ver. 2.20d, Exeter Publishing, Ltd. Setauket, NY.http://www.exetersoftware.com/cat/ntsyspc/ntsyspc.html
____ (2006). Morphometrics at SUNY Stony Brook: website http://life.bio.sunysb.edu/morph/
____ & Archie, J. W. (1984). A comparison of Fouriermethods for the description of wing shape inmosquitoes ( Diptera: Culicidae ). Syst. Zool. 33(3):302 – 317.
Rumpunen, K. & Bartish, I. V. (2002). Comparison of differentiation estimates based on morphometric andmolecular data, exemplified by various leaf shape
descriptors and RAPDs in the genus Chaenomeles (Rosaceae ). Taxon 51: 69 – 82.
Sales, M. F., Mayo, S. J. & Rodal, M. J. N. (1998).Plantas vasculares das florestas serranas de Pernam-buco: Um checklist da flora ameaçada dos brejosde altitude, Pernambuco Brasil. Imprensa Universi-tária, Universidade Federal Rural de Pernambuco,Recife.
Sampaio, V. S. B., Giulietti, A. M., Virginio, J. & Gamarra-Rojas, C. F. L. (2002). Vegetação e flora da caatinga.
210 KEW BULLETIN VOL. 63(2)
© The Board of Trustees of the Royal Botanic Gardens, Kew, 2008

8/12/2019 Andrade Et Al. - 2008 - Comparative Morphology of Populations of Monstera Adans-libre
http://slidepdf.com/reader/full/andrade-et-al-2008-comparative-morphology-of-populations-of-monstera-adans-libre 19/19
Associação Plantas do Nordeste (APNE), CentroNordestino de Informação Sobre Plantas (CNIP),Recife.
Slice, D. (2008). Morpheus et al. Multiplatform softwarefor morphometric research. http://www.morphometrics.org/id6.html
Tatsuta, H., Mizota, K. & Akimoto, S.-I. (2004).Relationship between size and shape in the sexually dimorphic beetle Prosopocoilus inclinatus (Coleoptera:
Lucanidae ). Biol. J. Linn. Soc. 81: 219 – 233.
Yamanaka, N., Ninomiya, S., Hoshi, M., Tsubokura, Y., Yano, M., Nagamura, Y., Sasaki, T. & Harada, K.(2001). An informative linkage map of soybeanreveals QTLs for flowering time, leaflet morphology and regions of segregation distortion. DNA Res. 8:61 – 72.
Yoshioka, Y., Iwata, H., Ohsawa, R. & Ninomiya, S.(2004). Analysis of petal shape variation of Primula
sieboldii by elliptic Fourier descriptors and principalcomponent analysis. Ann. Bot. 94: 657 – 664.
211COMPARATIVE MORPHOLOGY OF POPULATIONS OF MONSTERA ADANS.





























