Embed Size (px)
Citation preview

ANKARA ÜNİVERSİTESİ
FEN BİLİMLERİ ENSTİTÜSÜ
YÜKSEK LİSANS TEZİ
BAZI EKSTREM TERMOFİL BAKTERİLERİN AMİLAZLARININ
ÖZELLİKLERİNİN BELİRLENMESİ
GÜLAY FUKARA
GIDA MÜHENDİSLİĞİ ANABİLİM DALI
ANKARA
2007
Her hakkı saklıdır

i
ÖZET
Yüksek Lisans Tezi BAZI EKSTREM TERMOFİL BAKTERİLERİN AMİLAZLARININ ÖZELLİKLERİNİN
BELİRLENMESİ
Gülay FUKARA
Ankara Üniversitesi Fen Bilimleri Enstitüsü
Gıda Mühendisliği Anabilim Dalı
Danışman: Prof. Dr. Sedat DÖNMEZ
Bu çalışmada, çeşitli su ve toprak örneklerinden izole edilmiş olan bazı ekstrem termofil anaerobik bakteriler amilolitik aktiviteleri bakımından incelenmişlerdir. Test edilen 19 suştan, P/2, P/4, 70/2 ve CRK olarak isimlendirilen dört bakteri, eksraselüler α-amilaz ve ekstraselüler ve intraselüler α-glikozidaz üretim kapasitelerinin ve bu enzimlerin bazı özelliklerinin belirlenmesi amacıyla seçilmiştir. P/2, P/4, 70/2 ve CRK logaritmik gelişimlerini inkübasyonun sırasıyla 55., 48., 31. ve 31. saatlerinde tamamlamışlardır. Gelişme sırasında maksimum α-amilaz aktivitesi P/2 için 55. saatte 0.265 U/ml olarak, P/4 için 30. satte 0.254 U/ml olarak, 70/2 için 78. satte 0.278 olarak ve CRK için 55. satte 0.324 olarak belirlenmiştir. Enzimlerin sıcaklık ve pH optimumları; P/2’nin α-amilazı için 90 oC ve pH 7.0, P/4’ün α-amilazı için 70 oC ve pH 7.0, 70/2 ve CRK’nın α-amilazları için 70 oC ve pH 6.0 olarak belirlenmiştir. 70 oC’de 1 saat inkübasyondan sonra P/2, P/4, 70/2 ve CRK α-amilazları aktivitelerinin sırasıyla %30, %32, %25 ve %20’ini kaybetmişlerdir. Bakteriler, karbon kaynağı olarak nişasta ve azot kaynağı olarak tripton bulunan besiyerlerinde maksimum gelişme göstermişlerdir. Maksimum α-amilaz üretimi P/2 için, maltoz ve tripton, P/4 için nişasta ve tripton, 70/2 için sakkaroz ve tripton ve CRK için fruktoz ve pepton içeren besiyerlerinde meydana gelmiştir. α-amilaz aktivitesi, Mn+2 ve Ca+2 artırmış, Mg+2, Zn+2,Fe+2, Cu+2, Hg+2 ve EDTA ise azaltmıştır. Bakterilerde, ekstraselüler ya da intraselüler α-glikozidaz enzim aktivitesi belirlenememiştir. 2007, 57 Sayfa Anahtar Kelimeler : Ekstrem termofil, anaerob, termostabil, α-amilaz, α-glikozidaz, ekstraselüler, intraselüler, amilolitik aktivite

ii
ABSTRACT
Master Thesis
DETERMINATION OF THE SPECIFICATIONS OF THE AMYLASES OF SOME EXTREMELY THERMOPHILIC BACTERIA
Gülay FUKARA
Ankara University
Graduate School of Natural and Applied Sciences Department of Food Engineering
Supervisor: Prof. Dr. Sedat DÖNMEZ
In this study, some extremely thermophilic anaerobic bacteria isolated from various water and soil samples, were examined for amylolytic activity. Of the 19 strains tested, four bacteria called P/2, P/4, 70/2 and CRK were selected for determination of thermostable extracellular α-amylase and extracellular and intracellular α-glucosidase production capabilities and properties of these enzymes. P/2, P4, 70/2 and CRK finished the logarithmic growth at 55th, 48th, 31th and 31th hours of incubation, respectively. During the growth, maximum α-amylase activity was established as 0.265 U/ml at 55th hour for P/2, 0.254 at 31th hour for P/4, 0.278 U/ml at 78th hour and 0.324 U/ml at 55th hour for CRK. Temperature and pH optima were determined as 90 oC and 7.0 pH for P/2’s α-amylase, 70 oC and 7.0 pH for P/4’s α-amylase and 70 oC and 6.0 pH for 70/2 and CRK’s α-amylases. After 1 hour incubation at 70 oC and 6.0 pH α-amylases of P/2, P4, 70/2 and CRK lost 32%, 30%, 25% and 20% of their activities, repectively. Bacteria exhibited maximum growth in mediums that include starch as a carbon source and tyrptone as a nitrogen source. Maximum α-amylase production was occured in a medium containing maltose and tryptone for P/2, starch and tyrptone for P/4, sucrose and tyrptone for 70/2 and fructose and peptone for CRK. Mn+2 and Ca+2 increased, Mg+2, Zn+2,Fe+2, Cu+2, Hg+2 and EDTA decreased the α-amylase activity. Extracellular or intracellular α-glucosidase enzyme activity could not be determined in these bacteria. 2007, 57 pages Key Words : Extremely thermophilic, anaerobic, thermostable, α-amylase, α-glucosidase, extracellular, intracellular, amylolytic activity

iii
TEŞEKKÜR
Bana araştırma olanağı sağlayan ve çalışmamın her aşamasında yakın ilgi ve önerilerini
ile beni yönlendiren danışmanım Sayın Prof. Dr. Sedat DÖNMEZ (Ankara Üniversitesi
Mühendislik Fakültesi Gıda Mühendisliği Bölümü)’e, desteklerini gördüğüm Arş. Gör.
Safiye KARAAĞAÇ (Ankara Üniversitesi Mühendislik Fakültesi Gıda Mühendisliği
Bölümü) ve sevgili aileme teşekkürlerimi sunarım.
Gülay Fukara
Ankara, Nisan 2007

iv
İÇİNDEKİLER
ÖZET.................................................................................................................................i
ABSTRACT.....................................................................................................................ii
TEŞEKKÜR....................................................................................................................iii
ŞEKİLLER DİZİNİ.........................................................................................................v
ÇİZELGELER DİZİNİ..................................................................................................vi
1. GİRİŞ...........................................................................................................................1
2. KURAMSAL TEMELLER.......................................................................................4
2.1 Ekstrem Termofil ( Ekstremofil) Mikroorganizmaların Genel
Özellikleri..................................................................................................................4
2.2 Termofil Mikroorgazimalar....................................................................................5
2.2.1 Termofillerin yüksek sıcaklık direnci..................................................................7
2.3 Termostabil Enzimler...............................................................................................8
2.4 Amilazlar...................................................................................................................9
2.5 Amilazların Sınıflandırılması................................................................................11
2.6 Termostabil Amilaz Kaynakları............................................................................14
3. MATERYAL ve YÖNTEM.....................................................................................18
3.1 Deneyde Kullanılan Mikroorganizmalar.............................................................18
3.2 Besiyeri ve Çözeltiler..............................................................................................18
3.2.1 Besiyeri.................................................................................................................18
3.2.2 Analizlerde kullanılan çözeltiler.........................................................................19
3.3 Mikroorganizmaların Geliştirilmesi ve Muhafazası...........................................20
3.4 Mikroorganizma Gelişmesinin İzlenmesi.............................................................20
3.5 Enzim Aktivitelerinin Belirlenmesi.......................................................................20
3.5.1 αααα-amilaz aktivitesi...............................................................................................20
3.5.2 αααα-glikozidaz aktivitesi.........................................................................................21
3.6 Sıcaklık Optimumunun Belirlenmesi....................................................................22
3.7 Termal Stabilitenin Belirlenmesi..........................................................................23
3.8 Enzimlerin pH Optimumunun Belirlenmesi........................................................23
3.9 Karbon Kaynaklarının Bakteri Gelişimi ve Enzim Aktivitesine Etkisinin
Belirlenmesi............................................................................................................23

v
3.10 Azot Kaynaklarının Bakteri Gelişimi ve Enzim Aktivitesine Etkisinin
Belirlenmesi............................................................................................................23
3.11 Bazı Metallerin Enzim Aktivitesine Etkisini Belirlenmesi................................24
4. ARAŞTIRMA BULGULARI..................................................................................25
4.1 Mikroorganizmaların Gelişimlerinin İzlenmesi..................................................25
4.2 Mikroorganizmaların Enzim Aktiviteleri............................................................26
4.3 Enzimlerin Sıcaklık Optimumları.........................................................................27
4.4 Enzimlerin Termal Stabiliteleri............................................................................28
4.5 Enzimlerin pH Optimumları.................................................................................30
4.5 Farklı Karbon Kaynaklarının Bakteri Gelişmesi ve Enzim
Üretimine Etkisi......................................................................................................31
4.7 Azot Kaynaklarının Bakteri Gelişmesi ve Enzim Üretimine Etkileri...............32
4.8 Bazı Metallerin Enzim Aktivitesine Etkileri........................................................34
5. SONUÇ VE TARTIŞMA.........................................................................................36
KAYNAKLAR...............................................................................................................49
ÖZGEÇMİŞ...................................................................................................................57

vi
ŞEKİLLER DİZİNİ
Şekil 2.1 Amilolitik enzimlerin nişastayı hidroliz şekilleri ve
oluşturdukları ürünler.....................................................................................13
Şekil 4.1 a. P/2, b. P/4, c. 70/2 ve d. CRK izolatlarının 65 oC’de inkübasyon
sırasındaki optik yoğunluk (OD) ve pH değişimi...........................................25
Şekil 4.2 (a) P/2, (b) P/4, (c) 70/2 ve (d) CRK izolatlarının 65 oC’de inkübasyon
sırasındaki α-amilaz aktiviteleri......................................................................27
Şekil 4.3 (a) P/2, (b) P/4, (c) 70/2 ve (d) CRK izolatlarının α-amilazlarının farklı
sıcaklıklardaki oransal aktiviteleri...................................................................28
Şekil 4.4 (a) P/2, (b) P/4, (c).70/2 ve (d) CRK izolatlarının α-amilazlarının 70 oC
ve pH 6.0’daki termal stabiliteleri..................................................................29
Şekil 4.5 (a) P/2, (b) P/4, (c) 70/2 ve (d) CRK izolatlarının α-amilazlarının farklı
pH değerlerindeki oransal aktiviteleri.............................................................30

vii
ÇİZELGELER DİZİNİ
Çizelge 1.1 Mikrobiyal enzimlerin yıllık kullanım değerleri..........................................2
Çizelge 2.1 Ekstremofiller ve bulundukları ekstrem çevre koşulları...............................4
Çizelge 2.2 Bazı ekstremozimler ve uygulama alanları……………........................5
Çizelge 2.3 Türkiye’de ithalat yoluyla sağlanan toplam enzim miktarları
ve ödenen para değerleri............................................................................10
Çizelge 2.4 Bazı amilazların Ca iyonu gereksinimleri...................................................11
Çizelge 2.5 Termostabil amilaz üreten bazı mikroorganizmaların
doğadaki kaynakları....................................................................................14
Çizelge 2.6 Nişasta hidroliz eden enzimlerin kaynak mikroorganizmaları
ve özellikleri...............................................................................................16
Çizelge 4.1 P/2 ve P/4 izolatlarının farklı karbon kaynağı içeren besiyerindeki
gelişimleri ve α-amilaz aktiviteleri.............................................................31
Çizelge 4.2 70/2 ve CRK izolatlarının farklı karbon kaynağı içeren besiyerindeki
gelişimleri ve α-amilaz aktiviteleri.............................................................31
Çizelge 4.3 P/2 ve P/4 izolatlarının farklı azot kaynağı içeren besiyerindeki
gelişimleri ve α-amilaz aktiviteleri.............................................................33
Çizelge 4.4 70/2 ve CRK izolatlarının farklı azot kaynağı içeren besiyerindeki
gelişimleri ve α-amilaz aktiviteleri.............................................................33
Çizelge 4.5 Metallerin P/2, /4, 70/2 ve CRK α-amilazlarının
aktivitesine etkileri.....................................................................................35

1
1. GİRİŞ
Endüstride kullanılan enzimler, genellikle mikroorganizmalardan elde edilmekte, çok az
bir kısmı ise bitkisel ve hayvansal kaynaklardan sağlanmaktadır. Mikroorganizmaların
enzim kaynağı olarak kullanılmasının nedeni, aktivitelerinin yüksek olması, daha az yan
ürün oluşturmaları, daha stabil ve ucuz olmaları, büyük boyutlarda ve yüksek saflıkta
üretilebilmeleridir (Kıran ve Çömlekçioğlu 2003).
Mikrobiyal enzimlerin biyoteknolojik süreçlerle daha ekonomik olarak üretilmesi,
ayrıca suda çözünmeyen matrikslere bağlanarak immobilize edilerek daha uzun süre
kullanılabilmesi, endüstriyel enzim kullanımındaki artışın temel nedenleri arasındadır.
Enzimler, canlı hücrelerdeki tüm metabolik olayları yöneten spesifik katalizörlerdir.
Organik kimyanın alışılmış yöntemleri ile gerçekleştirilmesi zor olan pek çok
reaksiyonun, uygun enzimin varlığında, büyük bir özgünlükle (spesifiklik) ve kolayca
başarılabilmesi, çeşitli enzimlerin canlı hücrelerden izolasyonu ve canlı dışında çeşitli
amaçlar için kullanılabilmesi konusundaki araştırmaların güncelliğini sağlamaktadır.
Enzimatik proseslerin geleneksel proseslere göre daha az atık oluşturması, daha az
çevre kirliliğine yol açması, daha ılımlı koşullarda ve ekonomik olarak
gerçekleştirilebilmesi, enzim kullanımını giderek artırmaktadır (Gümüşel 2002).
Mikrobiyal enzimlerin yıllık kullanım değerlerinin verildiği çizelge1.1’de dünyada
kullanılan tüm enzimler içerisinde alkali proteazların %25, diğer proteazların %21,
amilazların %18, bir protez olan rennini %10, tripsin’in %3, lipaz’ın %3, diğer
karbonhidrat parçalayan enzimlerin %10 oranında kullanıldığı görülmektedir. 1985
yılında yapılan bir değerlendirmede, dünyadaki enzim satışının 450 milyon doları
bulduğu belirtilmiştir, günümüzde ise bu değerin 2-3 katı olduğu bilinmektedir ( Kıran
ve Çömlekçioğlu 2003).

2
Çizelge 1.1 Mikrobiyal enzimlerin yıllık kullanım değerleri (Kıran ve Çömlekçioğlu 2003)
Enzim Yıllık Kulanım Oranı(%)
Proteaz 25
Diğer Proteazlar 21
Amilaz 18
Rennin 10
Tripsin 3
Lipaz 3
Endüstriyel önemli birçok kimyasal proses, yüksek sıcaklık ve basınç gibi sert
koşullarda gerçekleştiğinden, bunlara alternatif yöntemler için bu ekstrem koşullara
dayanıklı enzimlere gerek duyulmaktadır. Günümüzde endüstriyel enzimlerin büyük
çoğunluğu mezofilik mikroorganizmalardan sağlanmakta ve bir çok avantaja sahip
olmalarına karşın, proseslerdeki ekstrem pH, sıcaklık ve iyon konsantresi v.b.
uygulamalarına dayanıksız olmaları nedeni ile kullanımları sınırlı kalmaktadır. Öte
yandan, ekstrem termofil (ekstremofil) mikroorganizmalardan elde edilen enzimler
(ekstremozimler) ekstrem koşullara daha dayanıklıdırlar. Bu nedenle son 20 yılda
ekstremofil ve ekstremozimlere olan ilgi artmış ve ektremozimler endüstriyel
biyoteknolojik uygulamalarda yaygın olarak kullanılmaya başlanmıştır.
Yaklaşık olarak son 40 yıldır, sert endüstriyel proseslere daha uygun olmaları nedeni ile
ekstremozimler endüstriyel ve biyolojik uygulamalarda ilgi çekmektedirler. Bu enzimler
ayrıca, protein mühendisliğinde termostabilitenin ve termoaktivitenin anlaşılabilmesi
için yapılan çalışmalarda model olarak da kullanılmaktadır (Kumar and Nussinov 2001,
Sterner and Lieble 2001).
Bugüne kadar, ekstremofil ve hipertermofil mikroorganizmalardan polisakkarit
parçalayan bir çok enzim (selülaz, amilaz, pullulanaz, lipaz, esteraz ,fitaz v.d.)
karakterize edilmiş ve endüstriyel prosesler için yeni olanaklar sağlanmıştır (Haki and
Rakshit 2003).

3
Termofilik organizmalardan elde edilen termostabil enzimlerin, çok yüksek
termostabiliteleri nedeni ile bir çok ticari uygulaması bulunmaktadır. Örneğin,
termostabil enzimlerin bir çoğu deterjan, gıda, yem, nişasta, tekstil, deri, kağıt ve ilaç
v.b. endüstrilerde kullanılmaktadırlar.
Günümüzde endüstriyel üretimlerde en çok kullanılan termostabil enzimler, nişasta
endüstrisinde kullanılan amilazlar’dır ve bunlar endüstriyel enzimlerin %25’ini
oluşturmaktadır (Poonam and Dalel 1995, Crab and Mitchinson 1997, Rao et al. 1998,
Sarıkaya et al. 2000). Nişasta endüstrisi, nişastanın glikoz ve diğer ürünlere
parçalanması ve modifikasyonu için termostabil amilolitik enzimlerin (amilaz,
glikoamilaz, izoamilaz ve pullulanaz) en yoğun kullanıldığı alandır. Nişasta
endüstrisinde ekonomik proseslerin sağlanabilmesi için kullanılan amilazların,
nişastanın jelatinizasyonu (100-110 oC) ve sıvılaştırma (80-90 oC) sıcaklıklarına
dayanıklı olmaları gerekmekte, bu nedenle termofilik ve termostabil amilazlara gerek
duyulmaktadır (Shindhu et al. 1997). Amilazlar ayrıca tekstil, gıda, içki, kağıt ve bira
v.d. endüstrilerde yaygın olarak kullanıldığı gibi klinik, medikal ve analitik kimya
alanlarında da kullanılmaktadırlar (Pandey et al. 2000).
Ekstremofillerden ekstremozimlerin üretimleri ile ilgili pekçok problem bulunmakta ve
bunların çoğu ekstrem termofil genlerinin mezofilik hücrelere aktarılması ile
aşılabilmektedir. Bu yolla bazı ekstremofillerde düşük düzeylerde bulunan bazı
enzimlerin mezofilik mikroorganizmalarda daha yüksek oranlarda üretilmeleri
sağlanmaktadır. Yeni ekstremofil mikroorganizmaların keşfi ve genetik mühendisliği
yöntemleri ile biyokataliz ve biyotransformasyon reaksiyonlarında yeni fırsatlar
sağlanacağı belirtilmektedir (Gomes and Steiner 2004).
Bu çalışmada çeşitli kaynaklardan izole edilen Thermoanaerobacter cinsine ait bazı
bakterilerin endüstriyel açıdan önemli olan termostabil amilaz enzimlerinin ve bu
enzimlerin biyoteknolojik üretim potansiyellerinin incelenmesi amaçlanmıştır.Bu yolla
özellikle gıda endüstrisinde enzim kullanımı ile proseslerde kolaylık sağlanacı gibi
üretim maliyetleri de ucuzlayacaktır.

4
2. KURAMSAL TEMELLER
2.1 Ekstrem Termofil Mikroorganizmaların ( Ekstremofiller) Genel Özellikleri
Gelişmeleri için ekstrem çevre koşullarına gerek duyan mikroorganizmalara
ekstremofiller ve bu mikroorganizmaların ürettiği enzimler ekstremozim’ler olarak
adlandırılmaktadır.Ekstremofiller, diğer bir çok mikroorganizmanın yaşayamayacağı
ortamlarda gelişebilen mikroorganizmalardır.Ekstrem çevre koşulları; çok yüksek (55-
121 oC) ya da çok düşük (-2-20 oC) sıcaklık, yüksek tuzluluk (2-5 M NaCl) ve çok
yüksek alkali (pH > 8) ya da çok yüksek asitli( pH < 4) koşullardır. Çeşitli
ekstremofiller, yüksek basınç, yüksek radyasyon ya da toksik bileşik içeren ortamlarda,
yeryüzünden çok derinde bulunan kayalarda ya da su seviyesi ve besin miktarı çok
düşük olan kuru ortamlarda yaşamlarını sürdürebilen mikroorganizmalardır. Ekstrem
termofilik mikroorganizmalar genellikle Arke’lere dahil bakterilerdir (Madigan and
Marrs 1997, Rothschild and Manicinelli 2001). Ekstremofillerin optimum gelişme
sıcaklıkları ve bulunabildikleri çevreler çizelge 2.1’de gösterilmektedir.
Çizelge 2.1 Ekstremofiller ve bulundukları ekstrem çevre koşulları (Woese et al. 1990) Fenotip Ortam Mikroorganizma
Termofilik 55-80 oC Methanobacterium, Thermoplasma, Thermus ve bazı Bacillus
türleri
Hipertermofilik 80-113 oC Aquifex, Archaeoglobus, Hydrogenobacter, Methanothermus,
Pyrococcus, Pyrodictium, Pyrolobus, Sulfolobus, Thermococcus,
Thermoproteus, Thermotoga
Psikrofilik -2- 20 oC Alteromonas, Psychrobacter
Halofilik 2- 5 M NaCl Haloarcula, Halobacterium, Haloferax, Halorurum
Asidofilik pH < 4 Acidianus, desulfurolobus, Sulfolobus, Thiobacillus
Alkafilik pH> 9 Natronobacterium, Natronococcus ve bazı Bacillus türleri
Son yıllarda ekstremofillere duyulan ilginin başlıca nedeni bunların ekstrem koşullarda
gelişmeleri ve biyoteknolojik uygulamalarda kullanılan enzimleri üretebilmeleridir
(Wiegel 1998). Ekstremofillerin ekstrem koşullarda yaşamlarını sürdürebilmelerinden
sorumlu olan enzimleri ve yapısal proteinleri üzerinde geniş kapsamlı araştırmalar
yapılmaktadır. Ekstremozimler, ekstrem pH, sıcaklık, iyonik kuvvet gibi uygulamalara

5
dayanıklılık gösteren enzimlerdir. Yapılan son araştırmalar, endüstriyel uygulamalar
için kullanılabilecek ekstremozimler üzerine yoğunlaşmıştır.Bu enzimler; esterazlar,
lipazlar, amilazlar, aldolazlar, nitrilazlar, amidazlar, fosfatazlar ve rasemazlardır.
Çizelge 2.2’ de endüstriyel uygulamalarda kullanılan ekstremozimler gösterilmektedir.
Çizelge 2.2 Bazı ekstremozimler ve uygulama alanları (Demirjian et al. 2001). Ekstremofil Enzim Endüstriyel uygulamalar
Termofil (55- 113 oC)
Amilazlar Ksilanazlar Proteazlar DNA polimerazlar
Tatlandırıcılar için glikoz ve fruktoz Kağıt beyazlatma Fırıncılık, bira, deterjan Genetik mühendisliği
Psikrofil (-2- 20oC)
Proteazlar Dehidrogenzlar Amilazlar
Peynir olgunlaştırılması, süt üretimi Biyosensörler Deterjanlarda polimer degradasyonu
Asidofil (pH< 3) Sülfür oksidasyonu Kömür desülfürizasyonu Alkafil (pH> 8) Selülazlar Deterjanlarda polimer degradasyonu Halofil (2– 5 M NaCl)
- Poli (α-glutamik asit )(PGA) üretimi Poli (β-hidroksi butirik asit)(PHB) üretimi
Barofil (Yüksek basınç) Tüm mikroorganizma Jellerin ve nişasta granüllerinin oluşturulması Metalofil (Yüksek metal konsantrasyonu)
Tüm mikroorganizma Maden cevheri elde edilmesi, biomineralizasyon
Psikrofilik enzimler, düşük sıcaklıklarda etkinliklerini sürdürebilmektedirler ve çeşitli
endüstriyel uygulamalarda kullanılmaktadırlar.Yüksek tuz konsantrasyonlarına
dayanıklı halofilik enzimler düşük su içeriğine sahip ortamlardaki biyokataliz
reaksiyonlarında model olarak kullanılmaktadırlar. Termofilik enzimler proteazlara,
deterjanlara ve organik çözücülerin etkilerine karşı dayanıklıdırlar (Sellek and
Chaudhuri 1999).
2.2 Termofil Mikroorgazimalar
Son yıllarda yapılan bir çok çalışmada, ekstremofil mikroorganizmalar içinde en çok
termofilik bakteriler dikkat çekmektedir ve bunların biyokatalitik potansiyelleri ve
enzimleri üzerinde birçok araştırma yapılmıştır (Gomes and Steiner 2004). Termofilik
mikroorganizmaların mezofilik mikroorganizmalara göre, yüksek üreme hızları, son
ürünün kolay kazanılması, yüksek proses stabilitesi ve verimi, nişasta selüloz gibi doğal
polimerleri doğrudan fermente edebilmeleri gibi önemli avantajları vardır

6
Yüksek sıcaklıklarda (60- 110 oC) gelişebilen termofilik mikroorganizmalar, volkanik
ve jeotermal kaynaklar örneğin, solfatarik alanlar, nötral sıcak su kaynakları ve deniz
dibi sıcak su kaynakları v.b. yüksek sıcaklığa sahip deniz ve karasal ortamlarında doğal
olarak bulunurlar. 50-60 oC arasındaki sıcaklıklarda optimum olarak çoğalabilen bu
mikroorganizmalar ılımlı termofillerdir, bu mikroorganizmaların bir çoğu; fungi,
protozoa, alg, streptomycetes ve cyanobacter gibi çeşitli prokaryotik ve ökaryotik
taksonomik gruplara dahildirler. 60-80 oC arasındaki sıcaklıklarda gelişebilen ekstrem
termofiller genellikle Bacillus, Clostridium, Thermoanaerobacter, Thermus,
Fervidobacterium, Thermotoga ve Aquifex cinslerine aittirler. 80-110 oC arasındaki
sıcaklıklarda gelişebilen hipertermofiller; Aquifex ve Thermotoga cinsi bakteriler ile
arke bakterilerden; Desulfurococcus, Sulfolobus, Pyrodictium, Pyrobaculum,
Thermoproteus, Methanopyrus, Pyrococcus, Thermococcus, Methanococcus ve
Archaeoglobus’u içermektedir.
Bazı termofil aerobik ve anaerobik sporlu bakteriler, 1940’lardan önce sıcak karasal
ortamlarından izole edilmişlerdir (Brock 1986). Yaklaşık 20 yıl önce, termal
kaynaklardan spor oluşturmayan aerobik bakteriler elde edilmiştir ve ilk termofil
organizma olan ve 80 oC’de optimum gelişme gösteren Sulfolobus acidocaldarius (arke
bakteri) izole edilmiştir (Brock et al. 1972). İlk olarak tanımlanan, spor oluşturmayan
termofil anaerob bakteri Methanobacterium thermoautotrophicum’dur (Zeikus and
Wolfe 1972). Hyperthermus butylicus ve Fervidobacterium islandicum gibi bazı
mikroorganizmalar max. gelişme gösterdikleri sıcaklıklardan daha yüksek
sıcaklıklardaki kaynaklardan elde edilebilirken, Archaeoglobus profundus gibi bazı
mikroorganizmalar da gelişme sıcaklıklarından daha düşük sıcaklıklardaki kaynaklardan
izole edilmişlerdir (Burggraf et al. 1990, Zillig et al. 1990).
Anaerobik termofiller, aerobik termofillere göre daha çok çeşitlilik göstermektedir ve
ekstrem yüksek sıcaklıklarda geliştirilmeleri daha kolaydır. Zorunlu anaerobik
mikroorganizmalar yüksek tuz konsantrasyonu ve düşük pH‘da adaptasyon
göstermektedirler (Lowe et al. 1993). Anaerobik termofil bakterilerin çoğunun
kemoorganotrof olduğu, genel olarak, anaerobik bakterilerin çevresel strese
adaptasyonlarının aerobik bakterilerden farklı faktörlerin etkisi ile olduğu anlaşılmıştır.

7
Bunlardan birincisi; anaerobik bakterilerin kemoorganotrofik gelişme boyunca
enerjilerinin kısıtlı olması, ikincisi ise bir çok kemoorganotrof anaerobik bakterinin
(methanojenler hariç) gelişmesi sonucu toksik ürünler (organik asit, alkol, H2S gibi)
meydana getirmesi ve bu ürünlere tolerans gösterebilmeleri için çeşitli adaptasyon
mekanizmaları geliştirmek zorunda olmalarıdır..
Termofillerin en ilgi çeken grubu hipertermofillerdir çünkü, bu organizmaların
izolasyonu, mikroorganizmalar için uygun habitatların yeniden değer kazanmasına ve
yaşamın bilinen sıcaklık limitlerinin yükselmesine neden olmuştur. Hipertermofiller
metabolizmaları açısından oldukça çeşitlidirler; metanogenler, sülfat indirgeyenler,
nitrat indirgeyenler ve hatta aerobik solunum yapanlar olmak üzere gruplara ayrılırlar.
Günümüzde bilinen hipertermofillerin büyük çoğunluğu anaerob heteretrofik sülfat
indirgeyen türlerdir. Arkeler içinde üç grup dikkat çekmektedir, bunlar karasal
solfatarik alanlarda bulunan asidofil ekstremofiller; kok şeklinde, zorunlu ve fakültatif
aerobiktirler ve asidik pH’ya (optimum pH 3.0) ihtiyaç duyarlar. Filogenetik olarak,
Sulfolobus, Metallosphaera, Acidianus ve Desulfurolobus cinslerine aittirler. Öte
yandan, ılımlı asidofil ve nötrofil termofiller solfatarik alanlar ve deniz hidrotermal
kaynaklarında bulunurlar ve hepsinin de zorunlu anaerob olduğu bilinmektedir.. Bu tür
ekstrem bölgelerde bulunan Thermoproteus, Pyrobaculum, Thermophilum,
Desulfurococcus ve Methanothermus cinslerine ait bakteriler izole edilmiştir (Kengen et
al. 1996, Stetter 1996).
2.2.1 Termofillerin yüksek sıcaklık direnci
Mikroorganizmalar, diğer tüm canlılar gibi yaşamak zorunda oldukları ortama uyum
sağlar ve yaşamlarını sürdürürler. Termofil mikroorganizmaların diğer canlılar gibi aynı
enzimleri içerirdikleri ancak bunların enzimlerinin daha termostabil, proteoliz ve
denatürasyona dayanıklı olduğu anlaşılmıştır (Kumar and Nussinova 2001).
Termofillerin hücre membranlarındaki doymuş yağ asitlerinini oranının fazla olduğu,
yağ asitlerinin hücre için hidrofobik bir ortam oluşturduğu ve yüksek sıcaklıklarda
yaşayabilmek için hücreyi daha dayanıklı hale getirdikleri anlaşılmıştır (Herbert and
Sharp 1992). Bir çok hipertermofilik bakteri içeren arkelerin hücre duvarlarında eter

8
bağı ile bağlanmış yağ asitleri bulunmakta, bu yapı doymuş yağ asitleri içeren
membranlara göre daha yüksek sıcaklıklarda bile bozulmadan kalabilmektedir (De Rosa
et al. 1994). Termofillerin DNA’larının pozitif süper sarmallar oluşturan ters DNAgiraz
içerdiği ve bu yapının, DNA’nın erime noktasını yükselttiği ve böylece
mikroorganizmayı daha yüksek sıcaklıklara dayanıklı hale getirdiği saptanmıştır.
Termofillerin ayrıca, disülfit bağları ve hidrosfobik etkileşimler ile sıcaklığa dayanıklı
hale gelebildiği belirlenmiştir (Lopez 1999, Kumar and Nussinov 2001).
2.3 Termostabil Enzimler
Termostabil enzimler, ekstremozimler içindeki en önemli enzimlerdir, bunlar protein
stabilitesinin anlaşılması için model olarak kullanılmalarının yanında çok önemli
biyoteknolojik potansiyellere de sahiptirler. Ekstremozim’lerin bu kadar önemli
olmalarının diğer nedeni ise yüksek sıcaklıklardaki biyoteknolojik proseslerin
avantajlarıdır. Örneğin, organik solvent içerisinde kimyasal reaksiyonların hızlarındaki
artış, yüksek sıcaklıklarda ortam viskozitesindeki azalma ve difüzyon katsayısındaki
artıştan kaynaklanmaktadır (Becker et al. 1997, Krahe et al. 1996 ). Çözünürlüğü düşük
olan hidrofobik bileşiklerin katıldığı çeşitli proseslerde, sıcaklığın yükseltilerek
çözünürlüğün artırılması ile reaksiyon hızının artması sağlanmakta ayrıca yüksek
sıcaklık, biyolojik hidroliz reaksiyonlarının hızlarını artırmakta ve kontaminasyon
riskini de azaltmaktadır.
Termofilik enzimlerin, etki mekanizmalarının mezofilik olanlarla benzer oldukları
belirtilmiştir (Adams 1993, Cowan 1992, Danson and Hough 1998). Bazı termostabil
enzimlerin mezofilik konak hücrede ekspresyonu ile termostabilitelerinin bozulmadığı
(Niehaus et al. 1999), bu nedenle termofilik enzimlerin yüksek sıcaklıklara adaptasyon
için geliştirdikleri moleküler stratejilerin genetik bir özellik olduğu belirtilmiştir
(Jaenicke and Bohm 1998).
Termofilik mikroorganizmalardan elde edilebilen termostabil enzimlerin,
mikroorganizmaların gelişme sıcaklıklarından daha yüksek sıcaklıklarda bile stabil
oldukları ve termofil enzim proteinlerinin daha sağlam oldukları belirlenmiştir (Saboto

9
et al. 1999). Fitter ve arkadaşları (2001) mezofilik ve termofilik amilazların yapısal
farklılıklarının aktiviteye etkisi üzerine yaptıkları çalışmada, termofilik proteinin
mezofilik proteine göre daha yüksek yapısal esneklik gösterdiğini belirlemişler ve
termofilik enzimin yüksek termostabiliteden sorumlu olduğunu kabul etmişlerdir.
2.4 Amilazlar
Amilazlar bilinen en önemli ve eski endüstriyel enzimlerdir. Nişasta molekülünü
hidroliz ederek dekstrin ve glikoz birimlerinden meydana gelen bir karışım oluştururlar.
Bu enzimler günümüzde büyük bir öneme sahiptirler ve gıda, fermentasyon, tekstil ve
kağıt v.b endüstrilerinde kullanılmaktadırlar. Endüstriyel alanda kullanılan amilazlar
bitkisel, hayvansal ve mikrobiyal kökenli olmakla birlikte ağırlıklı olarak
mikroorganizmalardan elde edilmektedirler. Bunun nedeni, mikroorganizma kaynaklı
enzimlerin katalitik aktivitelerinin yüksek olması, istenmeyen yan ürün
oluşturmamaları, daha stabil ve ucuz olmaları, büyük miktarda ve yüksek saflıkta elde
edilebilmeleridir.
Amilazların, toplam enzim pazarının %30’unu oluşturduğu belirtilmiştir (Van Der
Maarel et al. 2002). Nişastanın enzimatik dönüşümü; çirişlenme (jelatinizasyon),
sıvılaşma (liquefaction) ve şekerlenme (sakkarifikasyon) olmak üzere üç aşamada
meydana gelir. Jelatinizasyon; nişasta granüllerinin çözünmesi ve viskoz bir
süspansiyon oluşturması, sıvılaşma; nişastanın kısmi hidroliz ile viskozitenin azalması
olayıdır. Şekerlenme ise nişastadan glikoz ve fruktoz oluşmasıdır. Jelatinizasyon,
nişastanın su ile ısıtılması ile oluşur ve nişasta, sadece yüksek sıcaklıkta suda
çözünebilir (Rakshit 1998). Amilaz uygulamalarında, nişastanın hidrolizinin
jelatinizasyondan sonra hızla olabilmesi için kullanılan enzimin termostabil özellikte
olması gerekmektedir. Amilazların tarihi 1811 yılında Kirchhoff tarafından nişasta
parçalayıcı enzimlerin bulunması ile başlamaktadır. Bunu takiben sindirim amilazları ve
malt amilazları ile ilgili çalışmalar yapılmış, 1930 yılında Ohlsson maltta nişasta
parçalayan enzimleri, reaksiyonda ürettikleri şeker tipine göre α ve β- amilazlar olmak
üzere ikiye ayırmıştır (Ohlsson 1930).

10
Amilazlar, endoamilazlar ve ekzoamilazlar olmak üzere iki gruba ayrılabilirler,
endoamilazlar nişasta molekülünün iç kısmını rastgele noktalardan parçalar ve değişik
zincir uzunluğunda düz ve dallanmış oligosakkaritleri oluştururlar. Ekzoamilazlar,
ise,nişasta molekülünü indirgen olmayan ucundan parçalarlar. Günümüzde nişastayı
değişik ürünlere parçalayan birçok enzim bilinmektedir ( Gupta et al. 2003).
Amilazlar, asidik ve bazik koşullarda etki etmelerine göre asidik ve bazik karakterli
olabilmekte, bunlardan bazik pH’da aktivite gösterenler deterjan endüstrisinde önem
kazanmaktadırlar. Bazı basillerin pH 10 veya daha yukarısında aktivite gösteren α-
amilazları olduğu belirtilmiştir (Gupta et al. 2003). Endüstriyel proseslerde örneğin,
dekstroz üretiminde pH 4-5 aralığında etkili olabilen asidik amilazlar kullanılmaktadır.
Farklı kaynaklardan izole edilen bakteri endoamilazlarının optimum 50- 75 oC’de ve
3.5- 4.5 pH aralığında aktivite gösterdikleri saptanmıştır (Buonocore et al. 1976).
Amilaz üretimi ile ilgili teknik bilgiler, geniş boyutlarda araştırılmış ise de üretim
teknolojisi çoğunlukla patent altına alındığından yeterince açıklanmamıştır. Ülkemizde
giderek gelişen gıda sanayii, amilaz enzimi gereksinimini giderek artırmıştır (Üstünes
ve Güvenç 1985). Türkiye’de ithalat yoluyla sağlanan çeşitli enzimlerin toplam
miktarları ve dolar değerleri çizelge 2.3 ’de gösterilmektedir. Amilaz enzimleri de bu
enzimler içinde yer almakla beraber, bu enzimlerle ilgili yeterli bilgi edinilememiştir
(Dış Ticaret Müsteşarlığı).
Çizelge 2.3 Türkiye’de ithalat yoluyla sağlanan toplam enzim miktarları ve ödenen
para değerleri (Dış Ticaret Müsteşarlığı)
Yıl Miktar (kg) Dolar değeri 2004 6.749.850 55.909.914 $ 2005 8.006.471 62.372.989 $ 2006 (ocak-mart) 1.604.874 11.891.902 $
Hacmi giderek artan enzim tüketimine karşın ülkemizde enzim üretimi yapan sadece bir
kuruluş vardır (Orba A.Ş.). Kuruluş orta ölçekli bir işletme olup tümüyle araştırma ve
geliştirme çalışmaları sonucu tasarımlanmış ve faaliyete geçirilmiştir. Gıda ve tekstil
sektörü için amilaz enzimini ürettiği bildirilmektedir
(http://www.tubitak.gov.tr/btpd/btspd/platform/bio/bol2.html).
11
Dünyada amilaz ve diğer mikrobiyal enzimlerin üretimini yapan yüzlerce kuruluş
bulunmakla beraber, her ülke kendi gereksinimlerinin en az belirli bir bölümünü
üretmeye çalışmakta ve bu hemen hemen her zaman ithal edilene göre daha ekonomik
olmaktadır (Üstünes ve Güvenç 1985).
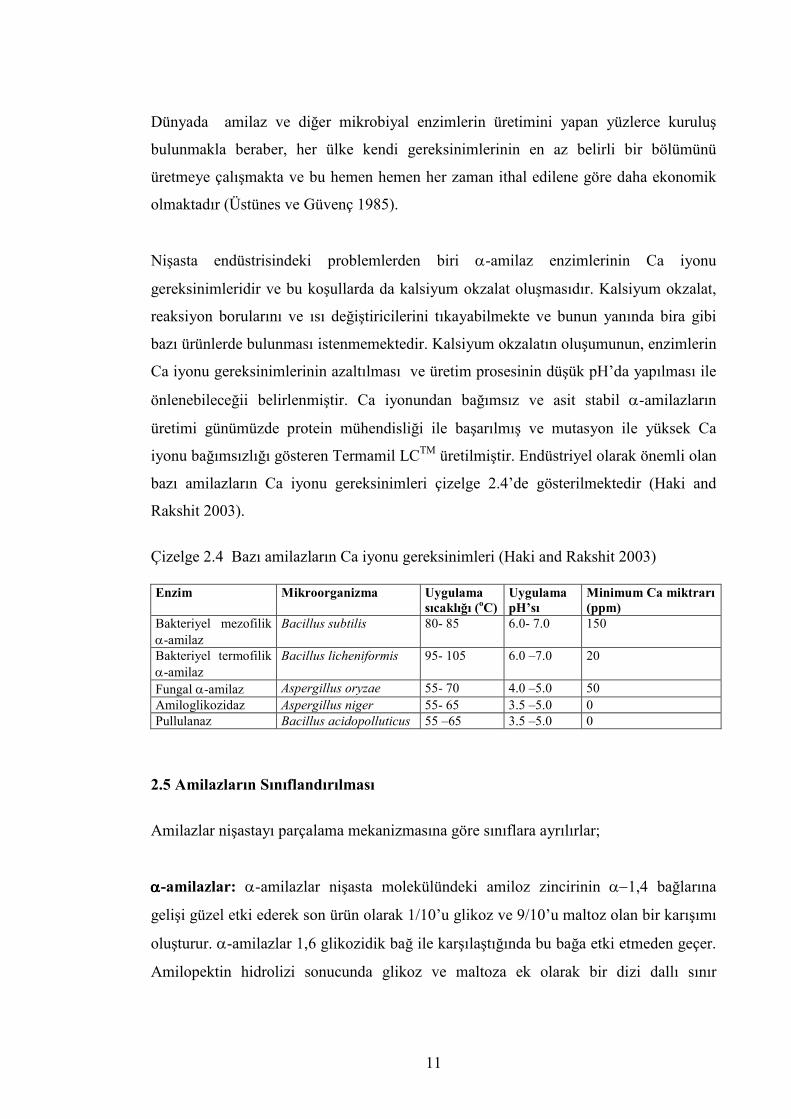
Nişasta endüstrisindeki problemlerden biri α-amilaz enzimlerinin Ca iyonu
gereksinimleridir ve bu koşullarda da kalsiyum okzalat oluşmasıdır. Kalsiyum okzalat,
reaksiyon borularını ve ısı değiştiricilerini tıkayabilmekte ve bunun yanında bira gibi
bazı ürünlerde bulunması istenmemektedir. Kalsiyum okzalatın oluşumunun, enzimlerin
Ca iyonu gereksinimlerinin azaltılması ve üretim prosesinin düşük pH’da yapılması ile
önlenebileceğii belirlenmiştir. Ca iyonundan bağımsız ve asit stabil α-amilazların
üretimi günümüzde protein mühendisliği ile başarılmış ve mutasyon ile yüksek Ca
iyonu bağımsızlığı gösteren Termamil LCTM üretilmiştir. Endüstriyel olarak önemli olan
bazı amilazların Ca iyonu gereksinimleri çizelge 2.4’de gösterilmektedir (Haki and
Rakshit 2003).
Çizelge 2.4 Bazı amilazların Ca iyonu gereksinimleri (Haki and Rakshit 2003) Enzim Mikroorganizma Uygulama
sıcaklığı (oC) Uygulama pH’sı
Minimum Ca miktrarı (ppm)
Bakteriyel mezofilik α-amilaz
Bacillus subtilis 80- 85 6.0- 7.0 150
Bakteriyel termofilik α-amilaz
Bacillus licheniformis 95- 105 6.0 –7.0 20
Fungal α-amilaz Aspergillus oryzae 55- 70 4.0 –5.0 50 Amiloglikozidaz Aspergillus niger 55- 65 3.5 –5.0 0 Pullulanaz Bacillus acidopolluticus 55 –65 3.5 –5.0 0
2.5 Amilazların Sınıflandırılması
Amilazlar nişastayı parçalama mekanizmasına göre sınıflara ayrılırlar;
αααα-amilazlar: α-amilazlar nişasta molekülündeki amiloz zincirinin α−1,4 bağlarına
gelişi güzel etki ederek son ürün olarak 1/10’u glikoz ve 9/10’u maltoz olan bir karışımı
oluşturur. α-amilazlar 1,6 glikozidik bağ ile karşılaştığında bu bağa etki etmeden geçer.
Amilopektin hidrolizi sonucunda glikoz ve maltoza ek olarak bir dizi dallı sınır

12
dekstrinler oluşmaktadır. Dört veya daha fazla glikoz içerikli bu dekstrinler orijinal
yapının 1,6 glikoz bağlarını içermektedirler. Belirli sayıda glikoz ünitesi kalınca
dallanma noktasına amilazlar etki edemediğinden bu dekstrinlere sınır dekstrin (limit
dekstrin) denilmektedir.
Nişastanın enzimatik hidrolizi sonunda oluşan ürünler α-optik konformasyonu
gösterdiğinden bu enzimlere α-amilazlar denilmektedir. Ayrıca bu enzimler nişastanın
iç kısımlarındaki α-1,4 bağlarına etki yaptıkları için endoenzimlerdir.
ββββ-amilazlar: β-amilazlar, nişasta molekülünün indirgen olmayan ucunda ardarda gelen
maltoz birimlerini uzaklaştıran enzimlerdir. Etkileri, α-1,6 bağlarına gelince durur.
Böylece sınır dekstrinler meydana gelmektedir. Hidroliz sonucu açığa çıkan ürünler β-
optik konformasyonu gösterdiğinden bu enzimlere de β-amilazlar adı verilmiştir.Genel
olarak β-amilazlar nişastanın dış kısmındaki 1,4 bağlarına etki ettikleri için
ekzoenzimlerdir.
Amiloglikozidazlar (Glikoamilazlar): Nişasta zincirinin ucundaki indirgen olmayan
kısımdan, ardışık olarak glikoz birimlerini uzaklaştıran bir ekzoenzimdir.
Amiloglikozidazların ürün olarak sadece glikoz oluşturması bu enzimleri, α ve β-
amilazdan ayırmaktadır. Hem α-1,4 bağlarını etkiler hem de yavaş bir hızla da olsa
α-1,6 bağlarını hidroliz eder.
Pullulanazlar: Substrat molekülünün sadece α-1,6 bağları üzerinde spesifik aktivite
gösterdikleri için ‘sınır dekstranazlar’ olarak da adlandırılırlar.
αααα- glukozidazlar: Amiloz ve amilopektinin parçalanma ürünleri olan kısa zincirli
oligosakkaritlerdeki α-1,4 bağlarını hidroliz ederek serbest glikoz birimleri oluşturan
enzimlerdir.

13
Şekil 2.1 Amilolitik enzimlerin nişastayı hidroliz şekilleri ve oluşturdukları ürünler (Bertoldo and Antranikan 2002)
CGTazlar: Nişastayı, siklodekstrin adı verilen ve 6,7 veya 8 glikoz ünitesinden oluşan
siklik D-glikozil birimleri oluşturan enzimlerdir. Bacillus macerans amilazları bunlara
örnektir (Aiyer 2005). Şekil 2.1’de amilazların nişastayı hidroliz şekilleri ve
oluşturdukları ürünler görülmektedir.

14
2.6 Termostabil Amilaz Kaynakları
Termostabil amilazlar, termofilik ve mezofilik bakterilerden ve funguslardan elde
edilebilirler ancak yüksek sıcaklıklardaki termostabiliteden termofil mikroorganizmalar
sorumludur. Bazı termostabil amilaz üreten mikroorganizmaların yaşam alanları çizelge
2.5’de gösterilmektedir.
Çizelge 2.5 Termostabil amilaz üreten bazı mikroorganizmaların doğadaki kaynakları (Haki and Rakshit 2003) Kaynak Mikroorganizma Enzim Sıcak su kaynağı Thermus sp. α-amilaz Sıcak su kaynağı Bacillus sp. WN.11 α-Amilaz Derin deniz hidrotermal kaynağı Staphilothermus marinus α-Amilaz Deniz solfatarik alanı Thermococcus litoralis Pullulanaz Kore tuzlu fermente ançüez Bacillus sp. KYJ963 β-amilaz
Sıcak su kaynaklarının sedimentleri Bacillus sp. 3183 α- amilaz
Derin deniz hidrotermal kaynaklarında nişasta bulunmamasına karşın bu bölgelerdeki
bazı mikroorganizmalardan nişasta parçalayan enzimlerin izolasyonu bu
mikroorganizmaların glikojeni hidroliz ettiklerini göstermektedir. Glikojen, hayvan
hücreleri ve bazı mikroorganizmalarda depo bileşiği olarak bulunur. Bazı arke
bakterilerinde, glikojen hücrede depo materyali olarak işlev görür, nişasta parçalayan
enzimlerin termofil bakterilerde fazla miktarda bulunması çok önemli role sahip
olduklarını gösterir. Polisakkaritlerin enzimatik hidrolizi ve modifikasyonları
biyoteknolojide çok ilgi görmektedir. Amilaz, pullulanaz ve glikoamilaz gibi
termostabil nişasta hidroliz eden enzimler gıda, kimya ve ilaç endüstrisinde bir çok
amaçla kullanılmaktadır. Birçok hipertermofilik mikroorganizmada bulunan amilazların
miktarı biyoteknolojik uygulamalar için düşük düzeydedir ancak termofillerden
mezofillere amilolitik enzim kodlayan genlerin aktarılması ile bu enzimlerin miktarının
önemli ölçüde arttığı belirlenmiştir. Bir çok durumda mezofilik hücrelerde üretilen bu
termostabil enzimler termostabilitelerini devam ettirirler ve düşük sıcaklığa rağmen
termostabilitelerinde artış gözlenir. Bu enzimler konak hücrenin proteolizine karşı
dirençlidirler ve ısıl denatürasyon ile konak hücrenin mezofilik enzimlerinden
kolaylıkla ayrılabilirler. Enzimin saflık derecesi birçok endüstriyel uygulama için
uygundur (Bertoldo and Antranikian 2002).

15
Nişasta endüstrisinin gereksinimlerini karşılamak amacıyla, değişik kaynaklardan
amilolitik enzimlerin izolasyonu ve karakterizasyonu için bir çok çalışma yapılmıştır.
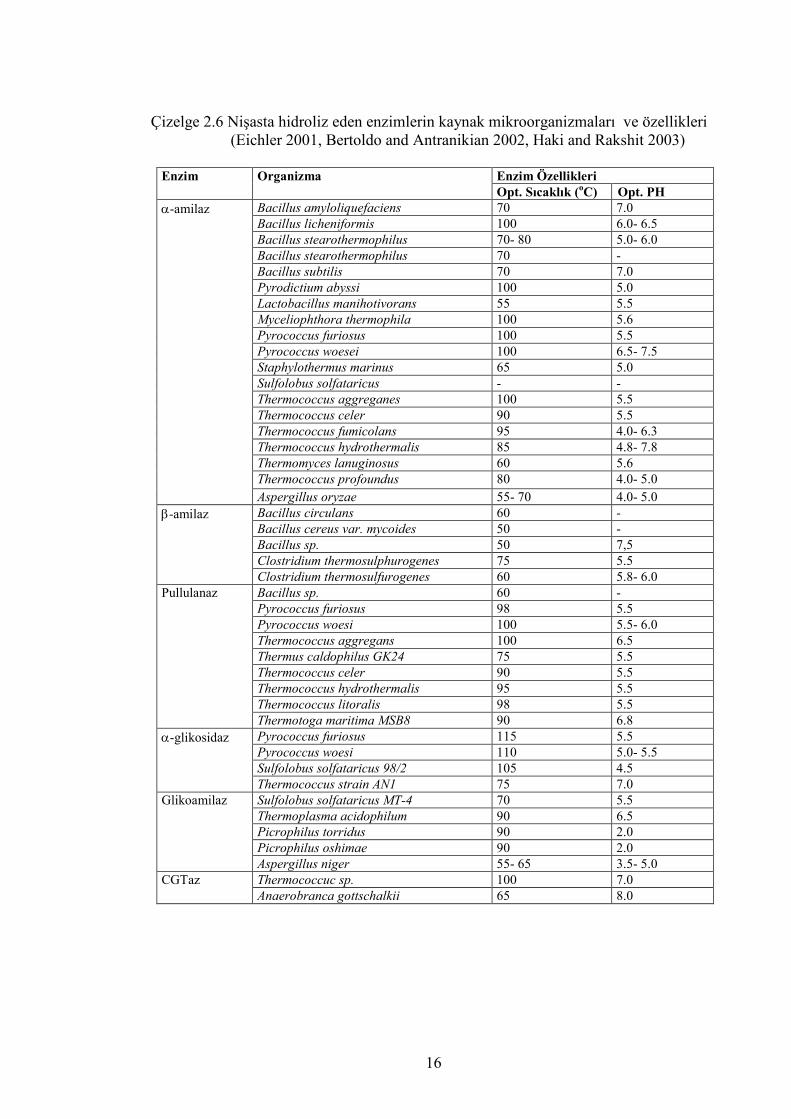
Bu çalışmalardan elde edilen bazı amilolitik enzimler ve bazı özellikleri çizelge 2.6’da
verilmiştir.
Termostabil α- amilazlar çok önceleri Bacillus subtilis, Bacillus licheniformis ve
Bacillus amyloliquefaciens’den ve 60 oC’ye kadar katalitik aktiviteye sahip olan β-
amilazlar çeşitli Bacillus türlerinden izole edilmişlerdir. Bu enzimler saf maltoz
şuruplarını elde etmek için pullulanazlarla birlikte gıda endüstrisinde çeşitli amaçlarla
kullanılabilirler.
Bacillus licheniformis, Myceliophthora thermophila, Pyrococcus wosei, Pyrococcus
furiosus ve Thermococcus aggreganes α-amilazları termofilik α-amilazlardır ve
optimum sıcaklıkları 100 oC’dir, ayrıca mezofillerin aksine Pyrococcus enzimleri Ca
iyonu varlığına da ihtiyaç duymamaktadırlar.
Bir anaerobik bakteri olan Thermococcus profoundus 80oC’de aktivite gösteren
termostabil α-amilaz üretmektiği ve bu amilazın maksimum termostabilite için Ca
iyonlarına gerek duyan S amilaz ya da daha asidik koşullarda etkili olan L amilaz
olduğu saptanmıştır. Bunlara ek olarak 90-100 oC arasında aktivite gösteren bir amilaz
enziminin Thermoproteales, Thermococcales ve Pyrodictiales’ ta bulunduğu
belirtilmiştir (Eichler 2001).
Termostabil bir β-amilaz enzimi için optimum sıcaklığın 50-75 oC ve pH’nın 5.5-7.5
arasında olduğu, en termostabil β-amilazın C. thermosulphurogenes’den elde edildiği ve
75 oC’de stabil olduğu belirlenmiştir (Haki and Rakshit 2003).
Termostabil pullulanaz enzimi için optimum sıcaklık aralığının 60-100 oC ve optimum
pH aralığının ise 5.5-6.8 olduğu, Pyrococcus wosei, Pyrococcus furiosus,
Thermococcus aggregans ve Thermococcus litoralis pullulanazları en termofil
pullulanazlar olduğu saptanmıştır (Haki and Rakshit 2003).
16
Çizelge 2.6 Nişasta hidroliz eden enzimlerin kaynak mikroorganizmaları ve özellikleri (Eichler 2001, Bertoldo and Antranikian 2002, Haki and Rakshit 2003)
Enzim Özellikleri Enzim Organizma Opt. Sıcaklık (oC) Opt. PH
Bacillus amyloliquefaciens 70 7.0 Bacillus licheniformis 100 6.0- 6.5 Bacillus stearothermophilus 70- 80 5.0- 6.0 Bacillus stearothermophilus 70 - Bacillus subtilis 70 7.0 Pyrodictium abyssi 100 5.0 Lactobacillus manihotivorans 55 5.5 Myceliophthora thermophila 100 5.6 Pyrococcus furiosus 100 5.5 Pyrococcus woesei 100 6.5- 7.5 Staphylothermus marinus 65 5.0 Sulfolobus solfataricus - - Thermococcus aggreganes 100 5.5 Thermococcus celer 90 5.5 Thermococcus fumicolans 95 4.0- 6.3 Thermococcus hydrothermalis 85 4.8- 7.8 Thermomyces lanuginosus 60 5.6 Thermococcus profoundus 80 4.0- 5.0
α-amilaz
Aspergillus oryzae 55- 70 4.0- 5.0 Bacillus circulans 60 - Bacillus cereus var. mycoides 50 - Bacillus sp. 50 7,5 Clostridium thermosulphurogenes 75 5.5
β-amilaz
Clostridium thermosulfurogenes 60 5.8- 6.0 Bacillus sp. 60 - Pyrococcus furiosus 98 5.5 Pyrococcus woesi 100 5.5- 6.0 Thermococcus aggregans 100 6.5 Thermus caldophilus GK24 75 5.5 Thermococcus celer 90 5.5 Thermococcus hydrothermalis 95 5.5 Thermococcus litoralis 98 5.5
Pullulanaz
Thermotoga maritima MSB8 90 6.8 Pyrococcus furiosus 115 5.5 Pyrococcus woesi 110 5.0- 5.5 Sulfolobus solfataricus 98/2 105 4.5
α-glikosidaz
Thermococcus strain AN1 75 7.0 Sulfolobus solfataricus MT-4 70 5.5 Thermoplasma acidophilum 90 6.5 Picrophilus torridus 90 2.0 Picrophilus oshimae 90 2.0
Glikoamilaz
Aspergillus niger 55- 65 3.5- 5.0 Thermococcuc sp. 100 7.0 CGTaz Anaerobranca gottschalkii 65 8.0

17
Pyrococcus wosei ve Pyrococcus furiosus α-glikosidazlarının en yüksek sıcaklık
derecesine sahip olan α-glikozidazlar olduğu, enzimin optimum aktivite sıcaklığının
105- 115 oC olduğu ve ayrıca metal iyonlarının varlığına gereksinim duyduğu
belirlenmiştir. İntraselüler bir α-glikosidazın Sulfolobus solfataricus 98/2’den elde
edildiği, enzimin optimum sıcaklığının mikroorganizmanın gelişme sıcaklığından
yaklaşık 20oC fazla olan 105oC olduğu bulunmuştur. Ayrıca Thermococcus
AN1’dentermostabil ekstraselüler bir α-glikozidaz elde edilmiştir (Eichler 2001).
Thermoplasma acidophilum, Picrophilus torridus ve Picrophilus oshimae’nin
termostabil ve asit stabil bir glikoamilaz ürettiği, bu termostabil glikoamilazın optimum
aktivitesinin pH: 2.0’de ve 90 oC’de olduğu anlaşılmıştır (Bertoldo and Antranikian
2002).
Termostabil CGTaz’ların bazı Thermoanaerobacter türlerinden,
Thermoanaerobacterium thermosulfirigens ve Anaerobranca gottschalkii’den üretildiği,
A. gottschalkii CGTaz’ının geniş bir pH aralığında (pH 4.0- 10.5) ve optimum
65 oC’ de aktif olduğu saptanmıştır. Nişastanın substrat olarak kullanıldığında bu
enzimin, α-siklodekstrin ve uzun süre inkübasyondan sonra β ve γ- siklodektrin
oluşturduğu, optimum 100 oC’de aktivite gösteren bir CGTazın, yeni izole edilmiş bir
arke olan Themococcus sp.’den elde edildiği açıklanmıştır (Bertoldo and Antranikian
2002).

18
3. MATERYAL ve YÖNTEM
3.1 Kullanılan Mikroorganizmalar
Bu çalışmada, çeşitli toprak ve su örneklerinden izole edilerek, gliserin içinde –65 oC’de
saklanan ekstrem termofil anaerob bakteri izolatları kullanılmıştır. Optimum olarak
65 oC’de gelişebilen 19 bakteri izolatı içerisinden amilaz aktivitesi en yüksek olan ve
P/2, P/4, CRK ve 70/2 olarak kodlanan Thermoanaerobacter cinsine ait bakteriler
kullanılmıştır. P/2 ve P/4 G(+), sporsuz ve basil; 70/2 ve CRK G(-), sporsuz ve basil
şeklindeki ekstrem termofil anaerobik bakterilerdir.
3.2 Besiyeri ve Çözeltiler
3.2.1 Besiyeri
Bu çalışmada, mikroorganizma gelişimi ve enzim üretimi için
Thermoanaerobacterium’lar için uygun olan ve aşağıda bileşimi verilen besiyeri
kullanılmıştır ( DSMZ-Medium 144). Bileşiminde litrede; NH4Cl, 0.9 g; NaCl, 0.9 g;
MgCl2.H2O, 0.4 g; KH2PO4, 0.75 g; K2HPO4, 1.5 g; FeSO4.7H2O, 1.0 ml (3.0 mg/ml);
resazurin, 1.0 ml (1mg/l); Na2S.9H2O, 1.0; maya özütü, 3.0 g; tripton, 10.0 g, karbon
kaynağı olarak son konsantrasyonu %1 (ağ/h) olacak şekilde glikoz ve 1000 ml distile
su içermektedir ancak bu çalışmada glikoz yerine nişasta ve tripton yerine bacto pepton
kullanılmıştır Besiyerine soğuk sterilize edilmiş 5 ml/l vitamin çözeltisi ve 9 ml/l iz
element çözeltisi eklenmiştir. Sterilizasyondan önce besiyeri pH’sı 1.0 N NaOH ile 7.2-
7.4’e ayarlanmıştır.
Vitamin çözeltisi (mg/l demineralize su); Biyotin, 2.0; folik asit, 2.0; piridoksin-HCl,
10.0; tiyamin-HCl.2H2O, 5.0; riboflavin, 5.0; nikotinik asit, 5.0; D-kalsiyum
pantotenat, 5.0; vitamin B12, 1.0, p-aminobenzoik asit, 5.0, lipoik asit, 5.0.
İz element çözeltisi (g/l demineralize su); FeCl2.7H2O, 1.0; nitrilotriasetik asit, 1.5;
MgSO4.7H2O, 3.0; MnSO4.2H2O, 0.5; NaCl. 1.0; CoSO4.7H2O, 0.18; CaCl2.2H2O, 0.1;

19
ZnSO4.7H2O, 0.18; CuSO4.5H2O, 0.01; KAl (SO4).12H2O, 0.02; H3BO3, 0.01;
Na2MO4.2H2O, 0.01; NiCl2.6H2O, 0.025; Na2SeO3.5H2O, 0.3x10-3. Nitrilotriasetik asit
belirli miktarda saf suda çözünmüş ve pH KOH’la 6.5’e ayarlanmıştır. Daha sonra diğer
mineraller de eklenerek, pH, KOH ile 7.0’a ayarlanmıştır (DSMZ-Medium 141).
Besiyeri, bir redoks inkibatörü olan resazurinden ileri gelen mavi renk pembe renge
dönüşünceye kadar kaynatılmış ve N2 atmosferi altında soğutulduktan sonra Hungate
tüplerine (Bellco Inc 2047-16000) 7.0 ml dağıtılmış ve tüpler kapatıldıktan sonra 121 oC’de 15 dakika sterilize edilmiştir.
3.2.2 Analizlerde kullanılan çözeltiler
•••• Sodyum fosfat tampon: 0.1 M, pH 6.0 sodyum fosfat tamponu α-amilaz aktivitesi
tayininde; 0.1 M, pH 7.0 sodyum fosfat tamponu α-glikozidaz aktivitesi tayininde;
0.1 M pH 5.0, 6.0, 7.0, 8.0 olan sodyum fosfat tamponları α-amilaz enziminin pH
optimumunun belirlenmesi amacıyla kullanılmıştır. Sodyum fosfat tamponu; 0.1 M
Na2HPO4.2H2O çözeltisi (15.6 g Na2HPO4.2H2O + 1000 ml destile su ) ile 0.1 M
NaH2PO4.2H2O çözeltisinin (17.8 g NaH2PO4.2H2O + 1000 ml destile su) belirli
oranlarda karıştırılması ile hazırlanmıştır.
• Glisin- NaOH tampon: Glisin-NaOH tamponu α-amilaz enziminin pH optimumunun
belirlenmesi amacıyla kullanılmıştır. PH’sı 9.0 ve 10.0 olan 0.2 M glisin-NaOH
tamponları, 0.2 M glisin ile 0.2 N NaOH çözeltilerinin belirli oranlarda karıştırılmasıyla
hazırlanmıştır.
• Tris-HCl tampon: 25 mM, pH 7.5 Tris-HCl tampon, EDTA ve metal çözeltilerinin
hazırlanmasında kullanılmıştır. 1.97 g Tris-HCl 500.0 ml saf suda çözüldükten sonra
1.0 N NaOH ile pH 7.5’e ayarlanmıştır.
• DNS (3,5-Dinitrosalisilik asit) çözeltisi: α-amilaz aktivitesi belirlenmesi sırasında
indirgen şeker tayininde kullanılmıştır. 1.0 g DNS (Sigma D-1510) 50.0 ml saf suda
çözüldükten sonra üzerine yavaş yavaş 30.0 mg sodyum potasyum tartarat (Merck) ilave

20
edilmiştir. Daha sonra 20.0 ml 2 N NaOH eklenerek son hacim saf su ile 100 ml’ ye
tamamlanmış ve hazırlanan çözelti en fazla 2 hafta süreyle kullanılmıştır (Worthington
1988).
• Na2CO3 çözeltisi: α-glikozidaz aktivitesi tayininde kullanılmıştır. Çözelti; 21.2 g
Na2CO3’ün 1000 ml saf suda çözülmesi ile hazırlanmıştır.
3.3 Mikroorganizmaların Geliştirilmesi ve Muhafazası
Bakterilerin izolasyonları, DSMZ-144 sıvı besiyerinde 65 oC’de geliştirilen kültürlerin
anaerobik ortamda petri kutularındaki DSMZ-144 besiyerine yayılması ve 65 oC’de
inkübasyon sonucu oluşan tek kolonilerin tekrar sıvı besiyerine inoküle edilmesi ile
yapılmıştır. Sıvı besiyerinde geliştirilen kültürler her üç günde bir taze besiyerine
aktarılarak çalışma süresince oda sıcaklığında muhafaza edilmişlerdir.
3.4 Mikroorganizma Gelişmesinin İzlenmesi
Mikroorganizma gelişmesi, 600nm dalga boyu ışıkta absorbans (O.D) ölçümü ile
izlenmiştir. Örnekler 15 ml’lik Hungate tüplerindeki 7.0 ml besiyerinde 65 oC’de 48
saat süreyle geliştirildikten sonra 50 ml besiyeri içeren 100 ml’lik serum şişelerine % 10
(h/h) oranında inoküle edilmiş ve tekrar 65 oC’de inkübasyona bırakılmıştır. İnkübasyon
sırasında belirli aralıklarla alınan örneklerin optik yoğunlukları spektrofotometrede
(Shimadzu UV-1208) 600nm’de ve tanık olarak taze besiyerine karşı ölçülmüştür. pH
ölçümü ise pH metre (GP-353 ATC) ile yapılmış, optik yoğunluk ve pH değerleri,
zamana karşı grafiklenerek gelişme eğrileri oluşturulmuştur.
3.5 Enzim Aktivitelerinin Belirlenmesi
3.5.1 αααα-amilaz aktivitesi
Geliştirilen bakterilerden belirli aralıklarla alınan 1.5 ml’lik örneklerin 10.000 rpm’de 5
dakika süre ile santrifüjlenmesiyle (Heraus Biofuge B) elde edilen berrak sıvı, hücre dışı

21
enzim aktivitesi tayininde enzim çözeltisi olarak kullanılmıştır.
Hücre dışı α-amilaz aktivitesinin tayin edildiği karışım 0.5 ml enzim çözeltisi ve 0.1 M,
pH 6.0 sodyum fosfat tamponu içerisinde hazırlanmış 0.5 ml %0.1’lik çözünür nişasta
(Merck F 309053) çözeltisi içermektedir. Substrat kontrolü, 0.5 ml enzim çözeltisi ile
0.5 ml sodyum fosfat tamponu; enzim kontrolü ise 0.5 ml sodyum potasyum tamponu
ile 0.5 ml çözünür nişasta solüsyonu içermektedir. Reaksiyon, 70 oC’de 30 dakika
süreyle gerçekleştirilmiş ve süre sonunda tüpler buz banyosuna içerisine alınarak
durdurulmuştur. 1 ml DNS eklendikten sonra tüpler 100 oC’de 5 dakika tutulmuş,
soğutulmuş ve gerekli seyreltme yapıldıktan sonra spektrofotometrede 540 nm’de
okuma yapılarak standart glikoz eğrisine göre glikoz oranı belirlenmiştir. Standart
eğrinin hazırlanmasında, sodyum fosfat tamponu içinde hazırlanan ve konsantrasyonu
%0-0.1 (ağ/h) aralığında glikoz çözeltileri ve tanık olarak sodyum fosfat tamponu
kullanılmıştır Enzim aktivitesi 3 paralel olarak belirlenmiş ve elde edilen sonuçların
ortalaması alınmıştır (Çoral ve Çolak 1998, Legin et al. 1998).
Bir α-amilaz enzim ünitesi (1U); 1 dakikada 1 µmol indirgen şeker (glikoz) oluşturan
enzim miktarı olarak tanımlanmış ve enzim aktivitesi aşağıdaki gibi hesaplanmıştır
(Çoral ve Çolak 1998, Legin et al. 1998).
Serbestleşen glikoz miktarı (µmol) Enzim aktivitesi (U/ml) =
Örnek miktarı (ml) x inkübasyon süresi (dk)
3.5.2 αααα-glikozidaz aktivitesi
α-glikozidaz aktivitesi yukarıdaki reaksiyona dayanarak saptanmış hücre dışı
(ekstraselüler) α-glikozidaz aktivitesi için enzim çözeltisi α-amilazda olduğu gibi
hazırlanmıştır. Hücre içi (intraselüler) enzim aktivitesi için enzim çözeltisi aşağıdaki
şekilde hazırlanmıştır.

22
48 saatlik sıvı kültürden alınanan 1.5 ml örnek, soğutmalı santrifüjde (Hettich D-78532)
4 oC’de 10.000 rpm’de 10 dakika süreyle santrifüj edilmiştir. Elde edilen hücre peleti,
7.0 pH’da 0.1 M sodyum fosfat tamponu ile yıkanarak tekrar santrifüjlenmiştir ve aynı
tampon içerisinde çözündürülmüştür. Soğuk su banyosu içinde 5 dakika sonifikasyon
(Sonics Vibra-cell VC 130) uygulaması ile hücreler parçalanmış ve örnek tekrar
santrifüjlenerek üstteki berrak sıvı kısmı enzim çözeltisi olarak kullanılmıştır (Suzuki et
al. 1975, Kocabaş ve Dizbay 1997).
Hücre içi ve hücre dışı enzim aktivitesinin ölçüldüğü karışım 0.5 ml enzim çözeltisi, 0.5
ml 20 mM PNPG (p-nitrophenyl-α-D-glucoside, sigma) çözeltisi (603 mg PNPG / 100
ml saf su) ve 1.0 ml 0.1 M sodyum fosfat tamponu (pH 7.0) içermektedir. Enzim
kontrolünde reaksiyon karışımındaki 0.5 ml PNPG çözeltisi yerine 0.5 ml saf su ;
substrat kontrolünde ise reaksiyon karışımındaki 0.5 ml enzim çözeltisi yerine 0.5 ml
sodyum fosfat tamponu konulmuştur. Reaksiyon, 70 oC’de 30 dakika sürmüş ve 2.0 ml
0.2 M Na2CO3 eklenerek durdurulmuştur. Kalan PNP miktarı, 400 nm’de tanığa (1.5 ml
tampon + 0.5 ml saf su) karşı O.D ölçülerek ve hazırlanan standart PNP (p-nitrophenyl)
eğrisi kullanılarak belirlenmiştir. Standart PNP eğrisinin hazırlanmasında, saf su içinde
hazırlanan ve konsantrasyonu %0-0.01 (ağ/h) aralığında olan PNP çözeltileri ve tanık
olarak saf su kullanılmıştır. Enzim aktivitesi 3 paralel olarak belirlenmiş ve elde edilen
sonuçların ortalaması alınmıştır.
Bir α-glikozidaz enzim ünitesi (1U), 1 dakikada 1 µmol PNP oluşturan enzim miktarı
olarak tanımlanmış ve enzim aktivitesi aşağıdaki gibi hesaplanmıştır (Çoral ve Çolak
1998, Legin et al. 1998, Toyobo enzymes AGH-201).
Serbestleşen PNP miktarı (µmol) Enzim aktivitesi (U/ml) =
Örnek miktarı (ml) x inkübasyon süresi (dk)
3.6 Sıcaklık Optimumunun Belirlenmesi
Bakterilerin α-amilazlarının sıcaklık optimumları, 50, 60, 70, 80, 90 ve 95 oC’de α-
amilaz enzim aktivitesi ölçülerek belirlenmiştir (Saha et al. 1990).

23
3.7 Termal Stabilitenin Belirlenmesi
Bakterilerin α-amilazlarının termal stabilitelerini belirlemek amacıyla, enzim çözeltileri
pH 6.0’da ve 70 oC’de 1 saat süreyle su banyosunda inkübe edilmiş, inkübasyon
sırasında belirli aralıklarla alınan örneklerin enzim aktiviteleri ölçülerek termal
stabiliteleri belirlenmiştir (Hyun and Zeikus 1985).
3.8 Enzimlerin pH Optimumunun Belirlenmesi
Bakterilerin α-amilazlarının optimum pH değerlerini belirlemek amacıyla 0.1M, pH 5.0,
6.0, 7.0, 8.0’de sodyum fosfat ve 0.2 M pH 9.0, 10.0’da glisin-NaOH tampon
çözeltilerinde hazırlanan nişasta çözeltisi substrat olarak kullanılmış ve enzim
aktiviteleri ölçülmüştür (Kligeberg et al. 1995).
3.9 Karbon Kaynaklarının Bakteri Gelişimi ve Enzim Aktivitesine Etkisinin
Belirlenmesi
Bu amaçla, besiyerine karbon kaynağı olarak %1 (ağ/h) oranında nişasta, fruktoz,
laktoz, ksiloz, maltoz, galaktoz, dekstrin, riboz, glikoz ve sakkaroz ilave edilmiştir
(Goyal et al. 2005). Değişik karbon kaynağı içeren besiyerlerinde 48 saat süreyle
geliştirilen bakterilerin 600 nm’deki optik yoğunlukları ve enzim aktiviteleri
ölçülmüştür.
3.10 Azot Kaynaklarının Bakteri Gelişimi ve Enzim Aktivitesine Etkisinin
Belirlenmesi
Bu amaçla, besiyerine azot kaynağı olarak %1 (ağ/h) oranında pepton, tripton, maya
özütü ve amonyum sülfat ilave edilmiştir (Lin et al. 1998). Değişik azot kaynağı içeren
besiyerlerinde 48 saat süreyle geliştirilen bakterilerin gelişmeleri ve enzim aktiviteleri
saptanmıştır.

24
3.11 Bazı Metallerin Enzim Aktivitesine Etkisinin Belirlenmesi
Bu amaçla, enzim çözeltilerine son konsantrasyonu 10mM olacak şekilde MgCl2,
MnCl2, CaCl2, ZnCl2, FeCl2, CuCl2, HgCl2 çözeltileri eklenmiştir. Metal içeren enzim
çözeltileri 70 oC’de 15 dakika süreyle inkübe edilmiştir ve bu süre sonunda enzim
aktiviteleri belirlenmiştir. Metal çözelti içermeyen enzim çözeltisi kontrol olarak
kullanılmıştır (Saha et al. 1990).
25
4.ARAŞTIRMA BULGULARI
4.1 Mikroorganizmaların Gelişimlerinin İzlenmesi
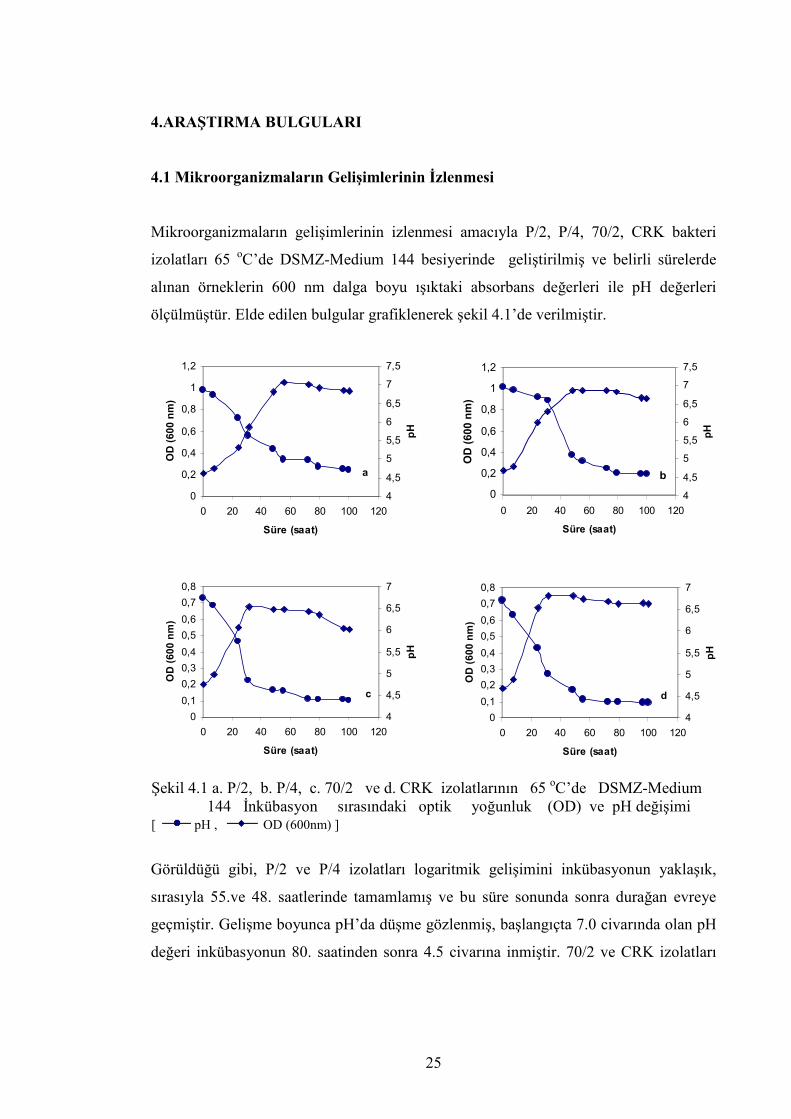
Mikroorganizmaların gelişimlerinin izlenmesi amacıyla P/2, P/4, 70/2, CRK bakteri
izolatları 65 oC’de DSMZ-Medium 144 besiyerinde geliştirilmiş ve belirli sürelerde
alınan örneklerin 600 nm dalga boyu ışıktaki absorbans değerleri ile pH değerleri
ölçülmüştür. Elde edilen bulgular grafiklenerek şekil 4.1’de verilmiştir.
Şekil 4.1 a. P/2, b. P/4, c. 70/2 ve d. CRK izolatlarının 65 oC’de DSMZ-Medium 144 İnkübasyon sırasındaki optik yoğunluk (OD) ve pH değişimi [ pH , OD (600nm) ]
Görüldüğü gibi, P/2 ve P/4 izolatları logaritmik gelişimini inkübasyonun yaklaşık,
sırasıyla 55.ve 48. saatlerinde tamamlamış ve bu süre sonunda sonra durağan evreye
geçmiştir. Gelişme boyunca pH’da düşme gözlenmiş, başlangıçta 7.0 civarında olan pH
değeri inkübasyonun 80. saatinden sonra 4.5 civarına inmiştir. 70/2 ve CRK izolatları
a
0
0,2
0,4
0,6
0,8
1
1,2
0 20 40 60 80 100 120
OD (600 nm)
4
4,5
5
5,5
6
6,5
7
7,5
0 20 40 60 80 100 120
Süre (saat)
pH
b
0
0,2
0,4
0,6
0,8
1
1,2
0 20 40 60 80 100 120
OD (600 nm)
4
4,5
5
5,5
6
6,5
7
7,5
0 20 40 60 80 100 120
Süre (saat)
pH
c
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0 20 40 60 80 100 120
OD (600 nm)
4
4,5
5
5,5
6
6,5
7
0 20 40 60 80 100 120
Süre (saat)
pH
d
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0 20 40 60 80 100 120
OD (600 nm)
4
4,5
5
5,5
6
6,5
7
0 20 40 60 80 100 120
Süre (saat)
pH

26
logaritmik gelişimini yaklaşık 31 saat sonra tamamlamış ve başlangıçta 7.0 civarında
olan pH değeri inkübasyonun 60.saatinden sonra 4,3 değerine kadar düşmüştür.
P/2 ve P/4, 70/2 ve CRK’ya göre yavaş fakat daha yoğun bir gelişme göstermiş, 600
nm’de ölçülen en yüksek yoğunluk değerleri; P/2 için 55. saatte 1.05, P/4 için 48. saatte
0.987, 70/2 için 31. saatte 0.675 ve CRK için 31.saatte 0.750 olarak ölçülmüştür.
Ayrıca, dört bakteri de besiyeri pH’sını logaritmik evre boyunca hızlı bir şekilde
düşürmüş, durağan evre süresince ise pH’daki düşüş yavaşlamıştır.
4.2 Mikroorganizmaların Enzim Aktiviteleri
Bu amaçla, P/2, P/4, 70/2 ve CRK izolatları 65 oC’de inkübe edilmiş ve belirli sürelerde
alınan örneklerde α-amilaz ve α-glikozidaz aktiviteleri ölçülmüş, elde edilen bulgular
grafiklenerek şekil 4.2’de gösterilmiştir.
Yapılan çalışmalar sonucunda dört bakteride de ekstraselüler ya da intraselüler
α-glikozidaz aktivitesine rastlanamamıştır.
Şekil 4.2’den anlaşıldığı gibi, bakterilerin izolatlarının dördünde de α-amilaz
üretiminde inkübasyonun ilk saatlerinde önemli bir artış gözlenmemiştir. P/2 izolatının,
inkübasyonun 55. saatinde yani logaritmik evrenin son saatlerinde max. enzim aktivitesi
gerçekleştirmiş ve α-amilaz aktivitesi 0.265 U/ml olarak heaplanmıştır. Bu saatten sonra
durağan evre süresince enzim aktivitesinde düşüş meydana gelmiştir. P/4 izolatı,
inkübasyonun 31. saatinde yani logaritmik gelişme fazında en yüksek α-amilaz
aktivitesine ulaşmış ve α-amilaz aktivitesi 0.254 U/ml olarak ölçülmüştür. Logaritmik
evrenin sonunda ve durağan evre boyunca enzim aktivitesinde düşüş gözlenmiştir. 70/2
izolatının α-amilaz aktivitesi logaritmik evre süresince artmış ve en yüksek enzim
aktivitesi 78. saatte yani durağan evrede 0.278 U/ml olarak ölçülmüştür. 80. saatte
enzim aktivitesi 0.267 U/ml olarak ölçülmüştür ve bu saatten sonra enzim aktivitesinde
düşüş gözlenmiştir. CRK izolatının α-amilaz aktivitesi inkübasyonun 55. saatine kadar
artmış ve en yüksek değeri olan 0.324 U/ml’ye ulaşmıştır. CRK, P/2 ve 70/2

27
izolatlarında gibi en yüksek enzim aktivitesi çoğalmanın durağan evresinde
gerçekleşmiştir (şekil 4.1 ve şekil 4.2).
Şekil 4.2 a. P/2, b. P/4, c. 70/2 ve d. CRK izolatlarının 65 oC’de DSMZ-Medium 144 besiyerinde inkübasyon sırasındaki α-amilaz aktiviteleri
4.3 Enzimlerin Sıcaklık Optimumları
Bakteri izolatlarının α-amilaz enzimlerinin optimum aktivite gösterdikleri sıcaklık
derecelerinin belirlenmesi amacıyla, 65 oC’de geliştirilen P/2, P/4, 70/2 ve CRK’nın α-
amilaz aktiviteleri 50, 60, 70, 80, 90, 95 oC’de ölçülmüştür. Elde edilen bulgular şekil
4.3’de gösterilmiş, oransal aktiviteler, en yüksek aktivite %100 kabul edilerek
hesaplanmıştır.
Şekil 4.3’te görüldüğü gibi P/2’den elde edilen α-amilaz enziminin aktivitesi 90 oC’ye
kadar sıcaklık artışı ile birlikte artış gösterdiği, 90 oC’de maksimum olduğu ve 95 oC’de
oransal aktivitenin %94 olduğu belirlenmiştir. P/4, 70/2 ve CRK izolatlarının α-amilaz
a
0
0,05
0,1
0,15
0,2
0,25
0,3
0 20 40 60 80 100 120
Süre (saat)
Enzim aktivitesi (U/ml)
b
0
0,05
0,1
0,15
0,2
0,25
0,3
0 20 40 60 80 100 120
Süre (saat)
Enzim aktivitesi (U/ml)
c
0
0,05
0,1
0,15
0,2
0,25
0,3
0 20 40 60 80 100 120
Süre (saat)
Enzim aktivitesi (U/ml)
d
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0 20 40 60 80 100 120
Süre (saat)
Enzim aktivitesi (U/ml)

28
aktiviteleri ise 70 oC’ye kadar sıcaklıkla orantılı olarak artmış, 70 oC’nin üzerindeki
sıcaklıklarda ise düşüş meydana gözlenmiş ve 95 oC’deki oransal aktiviteler sırasıyla
%91.7, %43 ve %62 olarak hesaplanmıştır.
d
0
20
40
60
80
100
120
40 50 60 70 80 90 100
Sıcaklık (oC)
Oransal Aktivite (%)
Şekil 4.3 a. P/2, b. P/4, c. 70/2 ve d. CRK izolatlarının α-amilazlarının farklı sıcaklıklardaki oransal aktiviteleri
4.4 Enzimlerin Termal Stabiliteleri
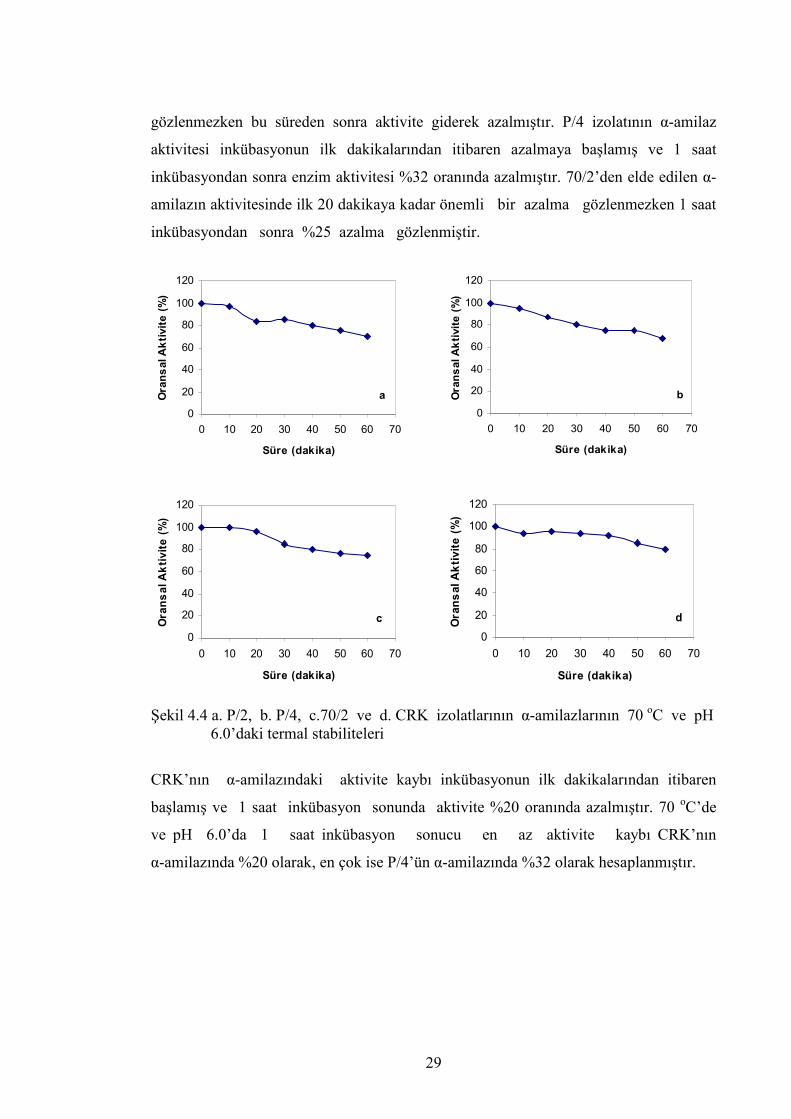
P/2, P/4, 70/2 ve CRK’nın α-amilazlarının termal satbilitelerini belirlemek amacıyla
enzim çözeltileri 70 oC’de ve pH 6.0’da 1 saat süreyle bekletilmiş ve her 10 dakikada
bir alınan örneklerde enzim aktivitesi ölçülmüştür. Elde edilen bulgular grafiklenerek
şekil 4.4’de gösterilmiş, oransal aktivite, en yüksek aktivite %100 kabul edilerek
hesaplanmıştır.
Şekil 4.4’den anlaşılacağı gibi, P/2 izolatından elde edilen α-amilaz, 1 saat inkübasyon
sonunda aktivitesinin %30’unu kaybetmiş, 10. dakikaya kadar aktivitede bir azalma
c
0
20
40
60
80
100
120
40 50 60 70 80 90 100
Sıcaklık (oC)
Oransal Aktivite (%)
a
0
20
40
60
80
100
120
40 50 60 70 80 90 100
Sıcaklık (oC)
Oransal Aktivite (%)
b
0
20
40
60
80
100
120
40 50 60 70 80 90 100
Sıcaklık (oC)
Oransal aktivite (%)
29
gözlenmezken bu süreden sonra aktivite giderek azalmıştır. P/4 izolatının α-amilaz
aktivitesi inkübasyonun ilk dakikalarından itibaren azalmaya başlamış ve 1 saat
inkübasyondan sonra enzim aktivitesi %32 oranında azalmıştır. 70/2’den elde edilen α-
amilazın aktivitesinde ilk 20 dakikaya kadar önemli bir azalma gözlenmezken 1 saat
inkübasyondan sonra %25 azalma gözlenmiştir.
Şekil 4.4 a. P/2, b. P/4, c.70/2 ve d. CRK izolatlarının α-amilazlarının 70 oC ve pH 6.0’daki termal stabiliteleri
CRK’nın α-amilazındaki aktivite kaybı inkübasyonun ilk dakikalarından itibaren
başlamış ve 1 saat inkübasyon sonunda aktivite %20 oranında azalmıştır. 70 oC’de
ve pH 6.0’da 1 saat inkübasyon sonucu en az aktivite kaybı CRK’nın
α-amilazında %20 olarak, en çok ise P/4’ün α-amilazında %32 olarak hesaplanmıştır.
a
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70
Süre (dakika)
Oransal Aktivite (%)
b
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70
Süre (dakika)Oransal Aktivite (%)
c
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70
Süre (dakika)
Oransal Aktivite (%)
d
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70
Süre (dakika)
Oransal Aktivite (%)
30
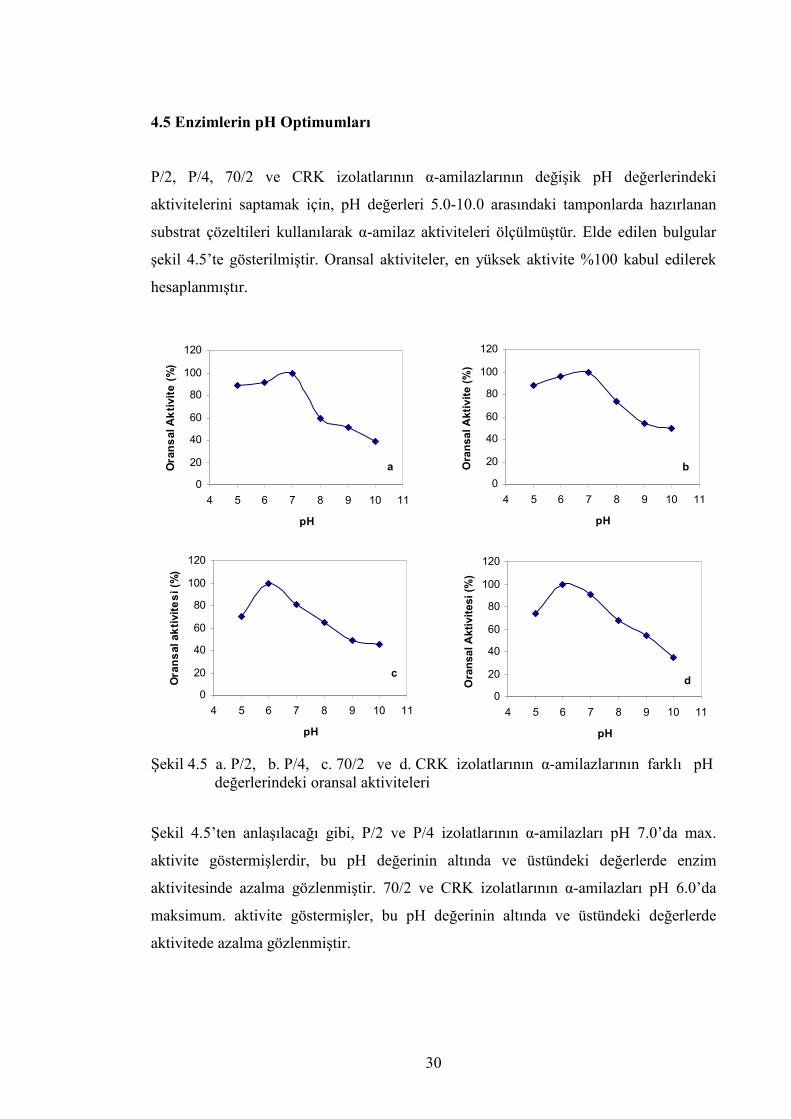
4.5 Enzimlerin pH Optimumları
P/2, P/4, 70/2 ve CRK izolatlarının α-amilazlarının değişik pH değerlerindeki
aktivitelerini saptamak için, pH değerleri 5.0-10.0 arasındaki tamponlarda hazırlanan
substrat çözeltileri kullanılarak α-amilaz aktiviteleri ölçülmüştür. Elde edilen bulgular
şekil 4.5’te gösterilmiştir. Oransal aktiviteler, en yüksek aktivite %100 kabul edilerek
hesaplanmıştır.
Şekil 4.5 a. P/2, b. P/4, c. 70/2 ve d. CRK izolatlarının α-amilazlarının farklı pH değerlerindeki oransal aktiviteleri Şekil 4.5’ten anlaşılacağı gibi, P/2 ve P/4 izolatlarının α-amilazları pH 7.0’da max.
aktivite göstermişlerdir, bu pH değerinin altında ve üstündeki değerlerde enzim
aktivitesinde azalma gözlenmiştir. 70/2 ve CRK izolatlarının α-amilazları pH 6.0’da
maksimum. aktivite göstermişler, bu pH değerinin altında ve üstündeki değerlerde
aktivitede azalma gözlenmiştir.
a
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11
pH
Oransal Aktivite (%)
b
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11
pH
Oransal Aktivite (%)
d
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11
pH
Oransal Aktivitesi (%)
c
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11
pH
Oransal aktivitesi (%)
31
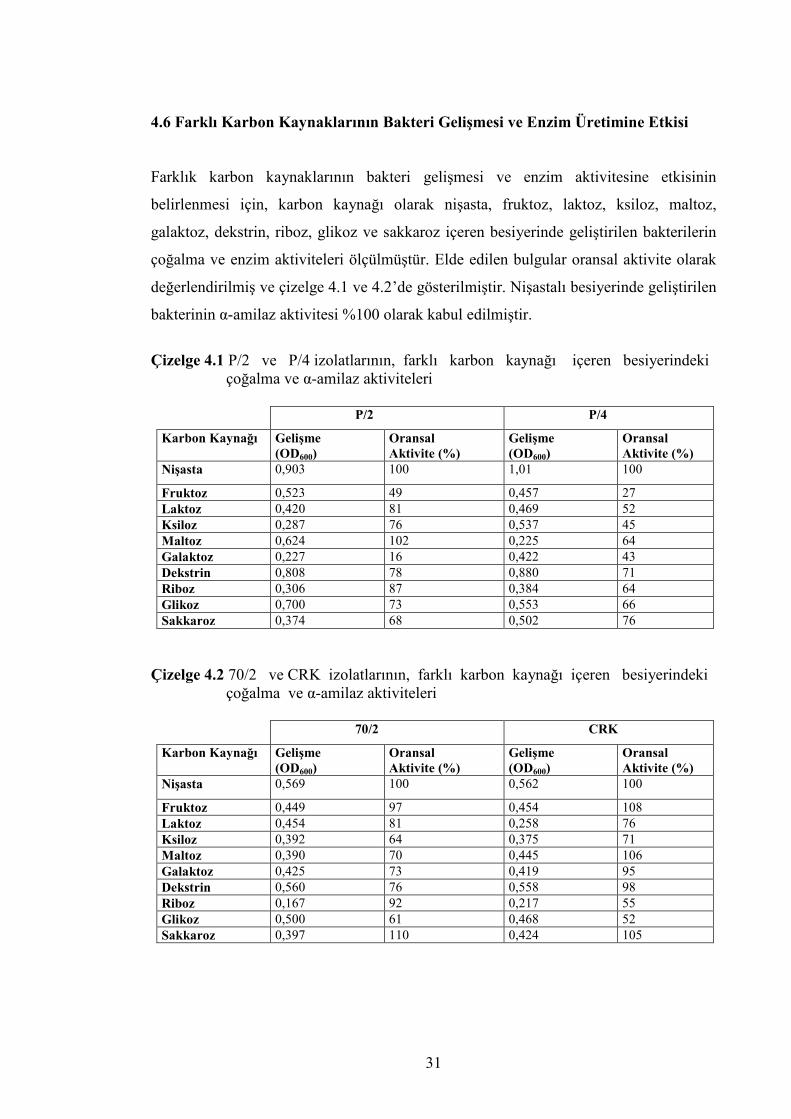
4.6 Farklı Karbon Kaynaklarının Bakteri Gelişmesi ve Enzim Üretimine Etkisi
Farklık karbon kaynaklarının bakteri gelişmesi ve enzim aktivitesine etkisinin
belirlenmesi için, karbon kaynağı olarak nişasta, fruktoz, laktoz, ksiloz, maltoz,
galaktoz, dekstrin, riboz, glikoz ve sakkaroz içeren besiyerinde geliştirilen bakterilerin
çoğalma ve enzim aktiviteleri ölçülmüştür. Elde edilen bulgular oransal aktivite olarak
değerlendirilmiş ve çizelge 4.1 ve 4.2’de gösterilmiştir. Nişastalı besiyerinde geliştirilen
bakterinin α-amilaz aktivitesi %100 olarak kabul edilmiştir.
Çizelge 4.1 P/2 ve P/4 izolatlarının, farklı karbon kaynağı içeren besiyerindeki
çoğalma ve α-amilaz aktiviteleri
P/2 P/4
Karbon Kaynağı Gelişme (OD600)
Oransal Aktivite (%)
Gelişme (OD600)
Oransal Aktivite (%)
Nişasta 0,903 100 1,01 100
Fruktoz 0,523 49 0,457 27 Laktoz 0,420 81 0,469 52 Ksiloz 0,287 76 0,537 45 Maltoz 0,624 102 0,225 64 Galaktoz 0,227 16 0,422 43 Dekstrin 0,808 78 0,880 71 Riboz 0,306 87 0,384 64 Glikoz 0,700 73 0,553 66 Sakkaroz 0,374 68 0,502 76
Çizelge 4.2 70/2 ve CRK izolatlarının, farklı karbon kaynağı içeren besiyerindeki
çoğalma ve α-amilaz aktiviteleri
70/2 CRK
Karbon Kaynağı Gelişme (OD600)
Oransal Aktivite (%)
Gelişme (OD600)
Oransal Aktivite (%)
Nişasta 0,569 100 0,562 100
Fruktoz 0,449 97 0,454 108 Laktoz 0,454 81 0,258 76 Ksiloz 0,392 64 0,375 71 Maltoz 0,390 70 0,445 106 Galaktoz 0,425 73 0,419 95 Dekstrin 0,560 76 0,558 98 Riboz 0,167 92 0,217 55 Glikoz 0,500 61 0,468 52 Sakkaroz 0,397 110 0,424 105

32
Çizelge 4.1’de görülgüğü gibi, P/2 izolatı, karbon kaynağı olarak, nişasta, dekstrin ve
glikoz içeren besiyerlerinde hızlı bir gelişme gösterirken laktoz, ksiloz, galaktoz, riboz
ve sakkaroz içeren besiyerlerinde daha yavaş bir gelişme göstermiştir. En hızlı gelişme,
nişasta içeren besiyerinde 0.903 değerinde olmasına karşın en fazla α-amilaz aktivitesi
maltoz içeren besiyerinde %102 olarak hesaplanmıştır. Glikoz ve dekstrin
mikroorganizma gelişimini olumlu yönde etkilerken enzim aktivitesini azaltmış, ksiloz,
laktoz ve riboz içeren besiyerlerinde düşük gelişmeye rağmen yüksek α-amilaz
aktivitesi gözlenmiştir. Ayrıca en düşük gelişme ve oransal α-amilaz aktivitesi
galaktozlu besiyerinde sırasıyla 0,227 ve %19 olarak hesaplanmıştır.
P/4 izolatı, karbon kaynağı olarak nişasta ve dekstrin içeren besiyerlerinde iyi, diğer
karbon kaynaklarını içeren besiyerlerinde daha yavaş bir gelişme göstermiştir. Nişasta,
dekstrin ve sakarozlu besiyerlerindeki α-amilaz aktivitesi diğer besiyerlerine göre daha
yüksek düzeyde olmuştur. En yüksek oransal enzim aktivite ve en yüksek gelişme
nişastalı besiyerinde sırasıyla %100 ve 1,01 olarak hesaplanmıştır. Buna karşın en
düşük gelişme maltozlu besiyerinde 0,225 ve en düşük oransal α-amilaz aktivitesi
fruktozlu besiyerinde %27 olarak hesaplanmıştır.
Çizelge 4.2’de görüldüğü gibi, 70/2 izolatı, karbon kaynağı olarak nişasta, laktoz, riboz
ve sakkaroz kullanılan besiyerlerinde diğerlerine göre daha yüksek α-amilaz aktivitesi
göstermiştir. En hızlı gelişme nişasta içeren besiyerinde 0.569 değerinde olmasına
karşın en yüksek oransal enzim aktivitesi sakkaroz kullanılan besiyerinde %110 olarak
hesaplanmıştır. Karbon kaynağı olarak riboz içeren besiyerinde 0.167 gibi çok düşük bir
gelişmeye karşın %92 değerinde oransal aktivite ölçülmüştür. Glikoz ve dekstrin
mikrobiyal gelişmeyi olumlu yönde etkilerken α-amilaz aktivitesine olumsuz etki
yapmışlardır. Ayrıca, en düşük oransal enzim aktivitesi glikoz içeren besiyerinde %61
olarak hesaplanmıştır.
CRK izolatı ise, karbon kaynağı olarak nişasta, fruktoz, maltoz, galaktoz, dekstrin ve
sakkaroz kullanılan besiyerlerinde yüksek α-amilaz aktivitesi göstermiştir. En yüksek
oransal enzim aktivitesi fruktoz ve maltoz kullanılan besiyerlerinde sırasıyla %108 ve
%106 olarak hesaplanmıştır. Laktoz, ksiloz ve riboz CRK izolatına hem gelişmeyi hem

33
de enzim sentezini azaltıcı yönde etki yapmış, glikoz, gelişmeyi artırırken α-amilaz
aktivitesini azaltıcı etki yapmış ve en düşük oransal enzim aktivitesi glikoz içeren
besiyerinde %52 olarak belirlenmiştir. Ayrıca, en yüksek gelişme oranı nişasta
kullanılan besiyerinde 0.562 olarak, en düşük gelişme ise riboz kullanılan besiyerinde
0.217 olarak ölçülmüştür.
4.7 Çeşitli Azotlu Bileşiklerin Bakteri Gelişmesi ve Enzim Aktivitesine Etkileri
Azot kaynaklarının bakteri gelişmesi ve enzim aktivitesine etkilerini belirlemek için,
azot kaynağı olarak; tripton, pepton, amonyum sülfat ve maya özütü kullanılan
besiyerlerinde geliştirilen bakterilerin mikrobiyal yoğunlukları ve α-amilaz aktiviteleri
ölçülmüştür. Elde edilen bulgular oransal aktivite olarak değerlendirilerek ve çizelge 4.3
ve 4.4’de gösterilmiştir.
Çizelge 4.3 P/2 ve P/4’ün farklı azot kaynağı içeren besiyerindeki gelişimleri ve α-amilaz aktiviteleri
P/2 P/4
Azot Kaynağı
Gelişme
(OD600)
Oransal
Aktivite (%)
Gelişme
(OD600)
Oransal
Aktivite (%)
Tripton 1,216 100 1,091 100
Pepton 1,100 95 1,015 81
Amonyum sülfat 0,369 69 0,609 75
Maya özütü 0,993 83 1,010 62
Çizelge 4.4 70/2 ve CRK’nın farklı azot kaynağı içeren besiyerindeki gelişimleri ve α-amilaz aktiviteleri
70/2 CRK
Azot Kaynağı
Gelişme
(OD600)
Oransal
Aktivite (%)
Gelişme
(OD600)
Oransal
Aktivite (%)
Tripton 0,768 100 0,793 100
Pepton 0,603 94 0,672 107
Amonyum sülfat 0,552 84 0,550 67
Maya özütü 0,688 11 0,655 97

34
P/2, P/4 ve 70/2 izolatları maksimum gelişmelerini sırasıyla 1.216, 1,091 ve 0.768
olarak ve oransal α-amilaz aktivitelerini %100 olarak tripton içeren besiyerinde
göstermişlerdir. Buna karşın CRK izolatı, en iyi gelişmeyi triptonlu besiyerinde 0,793
olarak, en yüksek α-amilaz aktivitesini ise peptonlu besiyerinde %100 olarak
göstermiştir. P/2 ve P/4 izolatları; pepton, tripton ve maya özütü içeren besiyerinde iyi
bir gelişme gösterirken amonyum sülfat içeren besiyerinde gelişmeleri önemli derecede
yavaşlamıştır. Amonyum sülfat, P/2 izolatının gelişmesini yavaşlattığı gibi α-amilaz
aktivitesini de %31 oranında azaltmıştır. Maya özütü, P/4’ün gelişimini olumlu yönde
etkilerken α-amilaz aktivitesini %38 oranında azaltmıştır. 70/2 izolatı, maya özütü
içeren besiyerinde iyi bir gelişme gösterirken, α-amilaz aktivitesi %59 oranında
azalmıştır. CRK izolatı ise, en yavaş gelişmeyi ve en düşük oransal enzim aktivitesini
sırasıyla 0.550 ve %67 olarak amonyum sülfat içeren besiyerinde göstermiştir.
4.8 Bazı Metallerin Enzim Aktivitesine Etkileri
Bazı metallerin test edilen bakteri izolatlarının α-amilaz aktivitelerine etkilerinin
belirlenmesi amacıyla enzim çözeltilerine Mg+2, Mn+2, Ca+2, ZN+2, Fe+2, Cu+2 Hg+2 ve
EDTA’nın çözeltileri ilave edilerek enzim aktiviteleri belirlenmiş ve sonuçlar oransal
aktivite olarak belirlenerek çizelge 4.5’de gösterilmiştir. Kontrol olarak kullanılan metal
içermeyen enzim çözeltisinin oransal α-amilaz aktivitesi %100 kabul edilmiştir.
Buna göre; P/2, P/4 ve 70/2 izolatlarının α-amilaz aktiviteleri Mn+2 ve Ca+2 varlığında
artarken, Mg+2, Zn+2, Fe+2, Cu+2, Hg+2 ve EDTA ilavesi ile azalmıştır. CRK izolatının
α-amilaz aktivitesi ise Mn+2 varlığında artarken diğer metallerin varlığında azalmıştır.
P/2 ve 70/2 izolatlarının α-amilazlarının maksimum aktiviteleri Ca+2 varlığında sırasıyla
%107 ve %110 olarak hesaplanırken P/4 ve CRK izolatlarının α-amilazlarının
maksimum aktiviteleri Mn+2 varlığında sırasıyla %107 ve %103 olarak hesaplanmıştır.
P/2 ve P/4 izolatlarının α-amilaz aktiviteleri Cu+2 varlığında çok fazla düşüş göstermiş
ve sırasıyla %5 ve %4 olarak belirlenmiştir. Ayrıca P/2 ve P/4 izolatlarının α-amilaz
aktiviteleri Hg+2 varlığında, 70/2 ve CRK izolatlarının α-amilaz aktiviteleri ise Cu+2 ve
Hg+2 varlığında tamamen durmuştur.

35
Çizelge 4.5 Metallerin P/2, P/4, 70/2 ve CRK α-amilazlarının aktivitesine etkileri
P/2 P/4 70/2 CRK
Metaller
Oransal Aktivite
(%)
Oransal Aktivite
(%)
Oransal Aktivite
(%)
Oransal Aktivite
(%)
Kontrol 100 100 100 100
Mg+2 98 88 73 75
Mn+2 103 107 105 103
Ca+2 107 106 110 95
Zn+2 60 93 72 52
Fe+2 61 72 82 78
Cu+2 5 4 0 0
Hg+2 0 0 0 0
EDTA 85 80 78 80

36
5. SONUÇ VE TARTIŞMA
Enzim teknolojisinin giderek gelişmesi, ürünlerin kullanım alanının çeşitliliği ve
ekonomik değerinin çok yüksek olması nedeniyle endüstriyel enzimlerle ilgili yapılan
çeşitli araştırmalar daha da önem kazanmaktadır. Termostabil amilazlar, termofilik ve
mezofilik bakterilerden elde edilebilirler ancak yüksek termostabil amilazlar termofil
bakterilerden sağlanırlar (Anonymous 1988).
Yüksek sıcaklıklarda (60-110 oC) gelişebilen mikroorganizmalar, termofiller, yüksek
sıcaklığa sahip, su ve karasal ortamlarda bulunmaktadırlar. 60-80 oC arasında
gelişebilen ekstrem termofiller genel olarak Bacillus, Clostridium,
Thermoanaerobacter, Thermus, Fervidobacterium, Thermotoga ve Aquifex cinslerine
aittirler (Haki and Rakshit 2003)
Bazı anaerobik termofiller enerjilerini sülfat indirgenmesiyle sağlarken, heterotrof
termofiller, organik bileşiklerin parçalanmasından sağlarlar. Buna göre, termofillerde
birçok enzimin termal stabilitesinin yüksek olduğu ve optimum gelişme sıcaklığında
enzimlerin maksimum aktiviteye sahip oldukları belirlenmiştir. Özellikle termofil
proteinlerin fizikokimyasal özelliklerinin, yapısının ve kararlılığının anlaşılması bunlara
olan ilginin ve akademik araştırmaların artmasına neden olmuştur (Sellek and
Chaundhuri 1999, Kumar and Nussinov 2000). Termofilik mikroorganizmaların
termostabil enzim kaynağı olarak kullanılmaya başlanmasından beri araştırmacılar,
nişasta üzerinde gelişebilen yeni termofilik anaerob bakteriler izole etmek için
çalışmalarını hızlandırmışlardır (Madi et al. 1987).
Çalışmada kullanılan dört bakteri de Thermoanaerobacter’ler için kullanılan DSMZ-
Medium 144 besiyerinde iyi bir gelişim göstermişlerdir. Şekil 4.1’de görüldüğü gibi P/2
ve P/4 izolatları yavaş fakat yoğun bir gelişme göstermişlerdir, sırasıyla 55. ve 48.
saatlerde logaritmik evreyi tamamlamış ve 1.05 ve 0,987 O.D değeri göstermişlerdir.
CRK ve 70/2 izolatları ise daha hızlı fakat düşük yoğunlukta gelişme göstermişlerdir,
yaklaşık 31. saatte logaritmik evreyi tamamlamış ve sırasıyla 0.750 ve 0.615 O.D
değerinde mikrobiyal yoğunluk değerine ulaşmışlardır.

37
Şekil 4.1’e göre, başlangıçta yaklaşık olarak 7.0 olan besiyeri pH’sının, P/2 ve P/4’ün
inkübasyonlarının 80. saatinde yaklaşık 4.5; 70/2 ve CRK’nın inkübasyonlarının 60.
saatinde ise yaklaşık 4.3 değerine kadar düştüğü belirlenmiştir.
Solfatarik alanlar, hem termofilik hem de asidik koşulları tercih eden
mikroorganizmalar için önemli kaynaklardır. Sulfolobus, Acidianus ve Thermoplasma
gibi termoasidofiller optimum 60-90 oC’de pH 5.0-7.0 arasında gelişirler ve anaerobik
üst tabakalarda bulunurlar. Thermoplasma tenax ve Methanothermus fervidus gibi
asidofilik ya da nötrofilik anaerobların daha derin katmanlardan izole edilebildiği
bildirilmiştir (Bertoldo and Antranikian 2002).
Şekil 4.2’de görüldüğü gibi bakterileri izolatlarının, α-amilaz aktivitelerinde
inkübasyonun 24. saatine kadar önemli bir artış gözlenmezken, bu saatten sonra üretim
artmıştır. P/2 ve CRK izolatları maksimum enzim aktivitesini, inkübasyonun 55.
saatinde sırasıyla 0.265 U/ml ve 0.324 U/ml olarak gerçekleştirmişlerdir. P/4 izolatı,
31.saatte 0.254 U/ml, 70/2 izolatı ise 72. saatte 0.278 U/ml olarak max. α-amilaz
aktivitesi gerçekleştirmişlerdir. P/2, 70/2 ve CRK izolatlarının maksimum enzim
aktivitelerini durağan evrede, P/4 izolatının ise logaritmik evrede gerçekleştirdiği
belirlenmiştir. Bakterilerin dördünde de ekstraselüler ya da intraselüler
α-glikozidaz aktivitesine rastlanamamıştır.
Yapılan bir çalışmada, bir arke olan Thermococcus hydrothermalis’in 2g/l maltoz + 2g/l
nişasta içeren besiyerinde hücre gelişimine bağlı olarak ekstraselüler
α-amilaz ürettiği bildirilmiştir. Bu bakterinin logaritmik gelişimini inkübasyonun 10.
saatinde tamamlaığı ve maksimum α-amilaz aktivitesini yaklaşık 10. saatte 0.270 U/ml
olarak gösterdiği saptanmıştır. Enzim aktivitesinin bu saatten sonra değişmediği,
bakterinin maksimum intraselüler α-glikozidaz aktivitesinin pH 7.5 ve 70 oC’de 5 saat
inkübasyon sonunda 15.0 U/l, ekstraselüler α-glikozidaz aktivitesinin ise 25 saat
inkübasyon sonunda 14.0 U/ml olduğu belirlenmiştir (Legin et al. 1998).
Madi et al. (1987) yaptıkları bir çalışmada termofil Clostridium sp. EM1 suşunda %0.5
nişasta içeren besiyerinde ekstraselüler α-amilaz aktivitesi belirlemişlerdir. Yaptıkları

38
çalışmada bakterinin logaritmik gelişimini inkübasyonun 12. saatinde tamamladığını ve
maksimum α-amilaz aktivitesinin yaklaşık olarak çoğalmanın 18. saatinde yani durağan
evrede 160 U/l olduğunu bildirmişlerdir.
C. acetobutylicum ATCC 824 suşu ile yapılan bir çalışmada %2 nişasta içeren
besiyerinde geliştirilen bakterinin amilolitik aktivitesinin bakteri gelişimi ile doğru
orantılı olduğu ve maksimum. amilolitik aktivite ile maksimum gelişmeyi inkübasyonun
33. saatinde gösterdiği belirtilmiştir (Annous and Blaschek 1990).
Legin et al. (1997) yaptıkları bir çalışmada deniz dibi hidrotermal kaynaklarından izole
ettikleri ve 60-80 oC’de gelişme gösteren arke bakterilerin amilolitik aktivitelerini
incelemişlerdir. %0.5 nişasta içeren besiyerinde 5.5 saatlik inkübasyon sonunda ölçülen
ekstraselüler ve intraselüler α-amilaz aktivitesinin sırasıyla U/l olarak, Thermococcus
fumicolans ST 557 için 8.64 ve 5.76; Thermococcus hydrothermalis AL 662 için 62.0
ve 8.6; Thermococcus sp. BI 533 suşu için 11.6 ve 1.4; Thermococcus sp. BI 552 suşu
için 20.6 ve 6.0; ST 553 izolatı için 44.5 ve 4.4; Thermococcus sp. AL 655 suşu için
81.2 ve 3.2 ve Themococcus sp. ST 701 suşu için 37.4 ve 3.0 olarak belirlenmiştir.
Termofil anaerob bakteri izolatı olan BI 534 ve Thermococcus BI 560 suşlarının sadece
ekstrasüler α-amilaz aktiviteleri U/ml cinsinden sırasıyla 10.0 ve 1.4 olarak
belirlenmiştir. Arkelerin sadece 5 tanesinde α-glikozidaz aktivitesi saptanabilmiş
ekstraselüler ve intraselüler α-glikozidaz aktivitesi U/ml cinsinden sırasıyla
Thermococcus fumicolans ST 557 için 0.6 ve 0.7 ve Thermococcus hydrothermalis AL
662 için 0.4 ve 3.9 olarak belirlenmiştir. Sadece intraselüler α-glikozidaz
aktivitesi ise U/ml cinsinden Thermococcus sp. BI 533 suşu için 1.4, BI 534 izolatı için
0.9 ve Thermococcus sp. ST 552 için 1.1 olarak belirlenmiştir. Diğer arke bakterilerde
α-glikozidaz aktivitesi belirlenememiştir.
Antranikian ve arkadaşları (1987) yaptıkları bir çalışmada Clostridium EMl suşunun pH
5.9 ve 60 oC’de intaselüler α-glikozidaz aktivitesini 7.0 U/ml ve ekstraselüler
α-glikozidaz aktivitesini de 9.0 U/ml olarak belirlemişlerdir.

39
Güvenç ve Üstünes (1987) yaptıkları bir çalışmada B. subtilis NCDO 1769 ve
B. amyloliquefaciens IAM 1521 suşlarının α-amilaz üretimini ve enzim üretiminin
kültür gelişimi ile ilişkisini incelemişlerdir. En yüksek enzim içeriğine B subtilis 48.
saat ve B. amyloliquefaciens 96. saat sonunda ulaşmıştır. En yüksek α-amilaz aktivtesi
değerleri B subtilis için 1.5 U/ml, B. amyloliquefaciens için 9.7 U/ml olarak
ölçülmüştür.
Bu çalışmada kullanılan mikroorganizmalar da dahil olmak üzere genel olarak termofil
mikroorganizmalardaki amilaz aktivitesinin mezofil olanlara göre daha düşük olduğu
belirlenmiştir. Bertoldo ve Antranikian (2002) birçok termofil mikroorganizmada
bulunan amilazların miktarının biyolojik uygulamalar için düşük düzeyde olduğunu ve
amilolitik genlerin termofillerden mezofillere transfer edildiğinde enzim sentezinin
önemli ölçüde arttığını bildirmişlerdir.
Bu çalışmaya ve yapılan diğer çalışmalara bakıldığında enzim üretiminin, genelde
mikroorganizma gelişmesi ile orantılı olarak arttığı fakat aralarında her zaman doğrusal
bir ilişki olmadığı, maksimum enzim aktivitesinin logaritmik ya da durağan evrede
olabileceği anlaşılmıştır.
Bu çalışmada, şekil 4.3’de görüldüğü gibi P/2 izolatının α-amilazının aktivitesinin,
50 oC’den 90 oC’ye kadar artan sıcaklıkla birlikte arttığı, 90 oC’de maksimum değere
ulaştığı ve 95 oC’de ise düşüş gösterdiği gözlenmiştir. P/4, 70/2 ve CRK izolatlarının
α-amilazlarının aktivitelerinin 70 oC’ye kadar artan sıcaklıkla birlikte arttığı bu
sıcaklıktan sonra ise azaldığı belirlenmiştir. 95 oC’de P/2, P/4, 70/2 ve CRK
izolatlarının α-amilazlarının oransal aktivitelerinin sırasıyla %94, %91.7, %43 ve %62
olduğu belirlenmiştir. Buna göre 70 oC’nin üzerindeki sıcaklıklarda en fazla aktivite
kaybının 70/2 izolatının α-amilazında, en az aktivite kaybının ise P/2 izolatının
α-amilazında olduğu belirlenmiştir.
Bu çalışmada, şekil 4.4’de görüldüğü gibi 70 oC ve pH 6.0’da 1 saat inkübasyon sonucu
P/2, P/4, 70/2 ve CRK izolatlarının α-amilazlarında sırasıyla %30, %32, %25 ve %20
oranlarında aktivite kaybı gözlenmiştir. P/2 izolatının α-amilazının 10 dakikaya kadar

40
stabilite gösterdiği 10. dakikadan sonra aktivitesinde düşüş meydana geldiği
belirlenmiştir. P/4 ve CRK izolatlarının α-amilazlarının aktivitesinde inkübasyonun
başlamasından itibaren düşüş gözlenmiştir. 70/2 izolatının α-amilazının aktivitesinde
yaklaşık 20 dakika boyunca önemli bir değişim olmadığı bu süreden sonra azalma
meydana geldiği belirlenmiştir
Koch et al. (1991) yaptıkları çalışmada Pyrococcus woesei α-amilazının 40-130 oC
arasında aktif olduğu ve optimum 100 oC’de ve pH 5.5’de aktivite gösterdiğini
bulmuşlardır. Yüksek derecede termostabilite gösteren bu enzimin 100 oC’de 5 saat
inkübasyondan sonra hala aktif olduğunu belirlemişlerdir.
Landerman et al. (1993a,b) yaptıkları çalışmada Pyrococcus furiosus’un α-amilaz
özelliklerini incelemişlerdir ve optimum 100 oC’de pH 6.5- 7.5’te aktivite gösterdiğini
belirlemişlerdir. Brown ve arkadaşları (1990) ise yaptıkları çalışmada Pyrococcus
furiosus α-amilazının yüksek derecede termostabil olduğu ve 98 oC’de 8 saatlik
inkübasyondan sonra bile aktivitesinin % 30’unu devam ettirdiğini belirlemişlerdir. 105 oC’de 2 saatlik inkübasyon sonunda ise aktivitenin sadece %10’unun devam ettiğini
bulmuşlardır.
Son yıllarda, 9 tane eksterm termofil arke suşunun ve 1 termofil bakterinin amilolitik
özellikleri incelenmiştir. Enzimatik aktivite genel olarak Thermoproteales
(Desulfurococcus mucosus ve Staphylothermus marinus), Thermococcales
( Thermococcus celer ve Thermococcus litoralis ) ve yeni izole edilmiş iki arke suşu
olan TYS ve TY’de incelenmiştir. Ayrıca Thermotogales cinsine ait yeni izole edilmiş
bir termofil bakteri ve daha önceden tanımlanmış olan Fervidobacterium pennavorans
‘da da amilolitik aktivite incelenmiştir. Enzimlerin genellikle 70-100 oC’de ve pH 5.5-
6.5’de optimum aktivite gösterdiği saptanmıştır (Canganella et al. 1994).
Yapılan bir çalışmada Clostridium sp. EM1 suşunun α-amilazının optimum 60 oC’de ve
pH 5.0’da aktivite gösterdiği belirlenmiştir. Enzimin, 70 oC’de aktivitesinin %25’ini,
80 oC’de ise %50’sini kaybettiği ve termal stabilitesinin substrat (nişasta) varlığında
oldukça arttığı belirlenmiştir. 75 oC’de 2 saat inkübasyon sonucu enzim aktivitesinin

41
nişasta yokluğunda %60 oranında, %0.5 nişasta varlığında %35 oranında, %1 nişasta
varlığında %25 oranında ve %2 nişasta varlığında %10 oranında azaldığı belirlenmiştir
(Madi et al. 1987).
Üstünes ve Güvenç (1987) yaptıkları çalışmada Bacillus amyloliquefaciens’den elde
edilen α-amilazın 70 oC’de ve pH 5.5’de max. aktivite gösterdiğini belirlemişlerdir.
Ayrıca enzimin bağıl aktivitesinin 40, 50, 60, 70, 80 ve 90 oC’de 30 dakika inkübasyon
sonunda sırasıyla %100, %89, %68, %51, %37 ve %26 olduğunu göstermişlerdir.
Yapılan bir çalışmada Bacillus licheniformis MY 10 suşunun α-amilazının özellikleri
incelenerek optimum 75-80 oC’de ve pH 5.0-6.5’da aktivite gösterdiği belirlenmiştir.
Oransal enzim aktivitesinin 70, 80 ve 90 oC’de 30 dakika inkübasyon sonucu sırasıyla
%100, %91.5 ve %59; 60 dakika inkübasyon sonucu ise sırasıyla %92, %68 ve %37.5
olduğu belirlenmiştir (Kumar et al. 1990).
B. stearothermophilus α-amilazının 55-70 oC ve pH 4.6- 5.1’de optimum aktivite
gösterdiği (Manning and Campbell 1961) belirlenmiştir. B. stearothermophilus ATCC
12980 suşunun α-amilazının 70-80 oC’de ve pH 5.0-6.0’da optimum aktivite gösterdiği
ve 70 oC’de 5 gün ya da 90 oC’de 45 dakika satabilite gösterdiği açıklanmıştır (Vihinen
and Mantsala 1989).
Thermoanaerobacter B6A’nın amilazının 40-90 oC gibi geniş sıcaklık aralığında aktif
olduğu ve optimum 75 oC’de aktivite gösterdiği, 70 oC’de 1 saat süreyle substratsız
inkübasyon sonunda enzimin hala etkin olduğu fakat 75 oC’de aktivitesini hızla
kaybettiği açıklanmıştır (Saha et al. 1990).
α-amilaz aktivitesi için sıcaklık optimumu geniş bir aralıktadır. En düşük sıcaklık
optimumu 25-30 oC olmak üzere F. oxysporum α-amilazına aitken, en yüksek sıcaklık
optimumu 100-130 oC olmak üzere arke bakterilerin α-amilazına aittir. α-amilaz
aktivitesi için pH optimumu da geniş bir aralık (2.0-12.0) göstermektedir.
Alicyclobacillus acidocaldarius α-amilazı optimum pH 3.0’da aktivite gösterir ve
asidik bir özelliğe sahiptir. Alkafilik bir Bacillus sp. suşundan elde edilen alkali

42
α-amilaz optimum pH 9.0-10.5’de aktivite gösterirken, ekstrem alkalifilik bir α-amilaz,
Bacillus sp. GM 8901 suşundan elde edilmiştir ve pH 11.0 –12.0’da optimum aktivite
gösterdiği belirlenmiştir (Gupta et al. 2003).
Bu çalışmada, P/2 ve P/4 izolatlarının α-amilazları, pH 7.0’da optimum aktivite
gösterdiklerinden nötral amilaz; 70/2 ve CRK izolatlarının α-amilazları, pH 6.0’da
optimum aktivite gösterdiklerinden (şekil 4.5) asidik amilaz oldukları belirlenmiştir.
Ayrıca enzimler yüksek sıcaklıkta aktivite gösterdikleri için termostabil amilaz olarak
değerlendirilmişlerdir.
Yapılan bu çalışmada ve diğer çalışmalarda termostabil α-amilazların hem mezofilik
hem de termofilik mikroorganizmalardan elde edilebilecekleri anlaşılmıştır. Haki ve
Rakshit (2003) yaptıkları çalışmada termostabil amilazların termofilik ve mezofilik
mikroorganizmalardan elde edilebileceğini fakat yüksek termostabiliteden termofil
mikroorganizmaların sorumlu olduğunu belirtmişlerdir. Termostabiliteyi etkileyen bir
çok faktör bulunmaktadır ve bu faktörler; kalsiyum, substrat ve diğer stabilizatörlerdir
(Vihinen and Mantsala 1989).
Bu çalışmada çeşitli karbon kaynaklarının bakteri gelişimi ve enzim üretimine olan
etkileri incelendiğinde (çizelge 4.1 ve çizelge 4.2 ), 4 bakterinin de nişastalı besiyerinde
maksimum gelişme gösterdikleri belirlenmiştir. P/2 izolatının, nişasta, fruktoz, maltoz,
dekstrin ve glikoz içeren besiyerlerinde hızlı bir gelişme gösterdiği; laktoz, ksiloz,
galaktoz, riboz ve sakkaroz içeren besiyerlerinde daha yavaş bir gelişme gösterdiği
belirlenmiştir. Glikoz ve maltozun gelişmeyi olumlu yönde etkilerken α-amilaz
sentezini azaltıcı etki yaptığı; ksiloz, laktoz ve ribozun ise gelişmeyi yavaşlattığı,
α-amilaz sentezini ise artırdığı saptanmıştır. P/4 izolatının nişasta ve dekstrin içeren
besiyerlerinde yüksek bir gelişme gösterdiği ve nişasta, dekstrin ve sakkaroz içeren
besiyerlerinde enzim sentezinin yüksek olduğu belirlenmiştir. 70/2 izolatının nişasta,
fruktoz, laktoz, riboz ve sakkaroz içeren besiyerlerinde yüksek α-amilaz sentezi
gerçekleştirdiği, ribozun gelişmeyi azalttığı; glikoz ve dekstrinin ise gelişmeyi artırdığı,
enzim aktivitesini azalttığı belirlenmiştir. CRK izolatının nişasta, fruktoz, maltoz,
galaktoz, dekstrin ve sakkaroz içeren besiyerlerinde yüksek oranda α-amilaz

43
sentezlediği belirlenmiştir. Laktoz, ksiloz ve ribozun gelişmeyi ve enzim sentezini
azalttığı; glikozun ise gelişmeyi artırdığı, enzim sentezini azalttığı saptanmıştır.
Bu çalışmada genel olarak 4 bakterinin de nişasta, dekstrin ve glikoz bulunan
besiyerlerinde yüksek yoğunlukta gelişme gösterdikleri; fruktoz, laktoz, ksiloz, riboz,
maltoz, galaktoz ve sakkaroz bulunan besiyerlerinde gelişmelerinin yavaşladığı
belirlenmiştir. α-amilaz aktivitesinin ise her bakteri için farklı karbon kaynakları
tarafından artırılıp azaltıldığı belirlenmiştir.
Antranikian et al. (1987) yaptıkları bir çalışmada Clostridium sp. EM1 suşunun %1
nişasta bulunan besiyerinde maksimum α-amilaz sentezi gerçekleştirdiğini
belirtmişlerdir. α-amilaz sentezini dekstrinin de önemli ölçüde artırdığını; maltoz ve
glikozun ise azalttığını belirlemişlerdir. Aynı bakteri ile yapılan başka bir çalışmada ise
α-amilaz üretiminin en fazla maltotrioz içeren besiyerinde olduğu fakat bakterinin en iyi
pullulan bulunan besiyerinde geliştiği belirtilmiştir. Ayrıca laktoz, fruktoz ve glikozun
α-amilaz sentezini azaltmalarına rağmen mikrobiyal gelişmeyi artırıcı yönde etki
yaptıkları belirlenmiştir (Madi et al. 1981).
P. furiosus’un α-amilaz özellikleri incelendiğinde glikojen, pullulan ve nişasta gibi
α-1,4 bağı içeren sakkaritlerin enzim sentezini önemli ölçüde teşvik ettiği; maltoz,
glikoz ve dekstrozun ise enzim sentezini baskıladıkları saptanmıştır (Brown et al. 1990).
Srivastova and Baruah (1986) yaptıkları bir çalışmada Bacillus stearothermophilus’un
termostabil α-amilazı üzerinde çalışmışlardır. Araştırmacılar, dekstrin, glikojen ve
çözünür nişasta gibi polisakkarit; rafinoz gibi trisakkarit ve selebiyoz, laktoz, maltoz ve
sakkaroz gibi monosakkaritlerin mikroorganizma gelişmesi ve α-amilaz sentezine
etkilerini incelemişlerdir. α-amilaz sentezinin bakterilerin, glikoz, maltoz ve sakkaroz
üzerinde gelişmesiyle baskılandığını; nişasta ve diğer polisakkaritlerde gelişmesiyle
teşvik edildiğini saptamışlardır. Farklı karbon kaynaklarının kullanımı ile meydana
gelen bakteri gelişiminin α-amilaz sentezini birebir etkilemediğini belirlemişlerdir.
Glikoz ve maltozun bakteri gelişimini artırırken α-amilaz sentezini önemli ölçüde
azalttıklarını belirtmişlerdir.

44
Ensley (1975), fruktozun karbon kaynağı olarak kullanıldığı zaman Clostridium
acetobutylicum’un hem gelişimini hem de α-amilaz sentezini azaltıcı etki yaptığını
saptamıştır. Ayrıca maltozun, Pseudomonas burtonii’nin α-amilaz sentezini teşvik
ettiğini belirlemiştir.
Kıran ve Çömlekçioğlu (2003), termofilik Bacillus sp. K-12 suşunun α-amilaz
üretimine nişastanın destekleyici etki yaptığını; dekstroz, laktoz ve sakkarozun ise
baskılayıcı etki yaptığını belirtmişlerdir. Bacillus sp. ile yapılan çeşitli çalışmalarda, De
Souza Teodoro ve Martins (2000), kültür ortamında bulunan glikozun α-amilaz
sentezini engellediğini; Bajpai ve Bajpai (1991), α-amilaz sentezini, sakkaroz, glikoz ve
fruktozun azalttığını, nişasta ve dekstrinin ise artırdığını; Albayrak ve arkadaşları
(1994), α-amilaz sentezini glikoz, fruktoz, sakkaroz ve maltozun azalttığını, dekstrin ve
pullulanın ise artırdığını saptamışlardır.
Yapılan bir çalışmada Endomycopsis fibuligera’nın α-amilaz sentezine glikoz, maltoz
ve sakkarozun baskılayıcı etki yaptıkları belirtilmiştir (Afunesyava et al. 1978).
Bu çalışmanın ve yapılan diğer çalışmaların sonuçlarına bakıldığında karbon
kaynaklarının mikrobiyal gelişmeye olan etkisi ile α-amilaz sentezine olan etkisi
arasında zorunlu bir ilişki olmadığı, birini azaltırken diğerini artırabildiği görülmektedir.
Genellikle glikozun mikroorganizma gelişimini artırdığı α-amilaz sentezini azalttığı;
ribozun ise mikroorganizma gelişimini azalttığı, enzim sentezini artırdığı belirlenmiştir.
Nişasta ve dekstrinin hem mikroorganizma gelişimini hem de α-amilaz sentezini
artırdığı; laktoz, ksiloz ve galaktozun ise hem mikroorganizma gelişimini hem de enzim
sentezini azalttığı saptanmıştır. Fruktoz ve sakkarozun bakteri gelişimini yavaşlattığı,
α-amilaz sentezine olan etkilerinin ise bakteriye göre değişiklik gösterdiği
belirlenmiştir.
Bu çalışmada azot kaynaklarının bakteri gelişimi ve enzim üretimine etkileri
incelendiğinde dört bakterinin de tripton içeren besiyerinde maksimum gelişme
gösterdikleri belirlenmiştir. Maksimum α-amilaz sentezini, P/2, P/4 ve 70/2 izolatlarının
tripton, CRK izolatının ise pepton bulunan besiyerinde gösterdikleri belirlenmiştir. Dört

45
bakterinin de tripton ve pepton içeren besiyerlerinde iyi bir gelişme ve α-amilaz sentezi
gerçekleştirdikleri gözlenmiştir. Bakterilerin amonyum sülfat bulunan besiyerinde hem
gelişimlerinin hemde α-amilaz sentezlerinin azaldığı; maya özütü içeren besiyerinde ise
gelişimlerinin iyi olmasına rağmen α-amilaz sentezlerinin azaldığı belirlenmiştir
(Çizelge 4.3 ve 4.4).
Kıran ve Çömlekçioğlu (2003), Bacillus sp. K-12’nin oransal α-amilaz aktivitesinin 48
saat inkübasyon sonunda azot kaynağı olarak maya özütü içeren besiyerinde %100, et
özütü içeren besiyerinde %81, pepton içeren besiyerinde %68 ve amonyum sülfat içeren
besiyerinde %10 olduğunu belirlemişlerdir.
Sarıkaya (1995), farklı azot kaynakları bulunan besiyerinde Bacillus
amyloliquefaciens’in α-amilaz aktivitesini test etmiştir. Çalışmada en yüksek oransal
aktivitenin maya özütü içeren ortamda olduğunu belirlenmiştir.
Goyal and arkadaşları (2005) Bacillus sp. 1-3’ün α-amilaz aktivitesinin azot kaynağı
olarak soya fasulyesi unu kullanılan besiyerinde maksimum olduğunu belirlemişlerdir.
Bacillus sp. WN11 ile yapılan bir çalışmada bakterinin karbon kaynağı olarak proteaz
pepton ve tripton kullanılan besiyerinde yüksek gelişme ve α-amilaz aktivitesi
gösterdiğini saptanmıştır (Mamo and Gessesse 1999).
Yapılan bir çalışmada B. subtilis’in maksimum α-amilaz üretimini azot kaynağı olarak
maya özütü bulunan besiyerinde gerçekleştirdiği bulunmuştur (Baysal et al. 2003).
Narang and Satyanarayana (2001), yeni izole edilmiş bir ekstrem termofil olan
Bacillus thermooleovorans ile yaptıkları bir çalışmada bakterinin α-amilaz üretiminin
tripton içeren besiyerinde maksimum düzeyde gerçekleştiğini belirtmişlerdir.
Et özütü, maya özütü, pepton ve soya fasulyesi unu gibi bir çok değişik organik nitrojen
kaynağı bakterilerde maksimum α-amilaz sentezini sağlayabilir ( Gupta et al. 2003).

46
Yapılan bu çalışma ve diğer çalışmalar dikkate alındığında, azot kaynağının bakteri
gelişimine ve α-amilaz sentezine olan etkisinin genelde orantılı olduğu fakat aralarında
zorunlu bir ilişki olmadığı belirlenmiştir. Amonyum sülfatın mikrobiyal gelişmeyi ve
α-amilaz sentezini önemli ölçüde baskıladığı; bu çalışmada tripton ve peptonun P/2, P/4,
70/2 ve CRK izolatlarının α-amilaz sentezini artırmalarına rağmen diğer çalışmalar da
dikkate alındığında tripton, pepton ve maya özütünün etkilerinin bakteriye göre
değiştiği belirlenmiştir.
Bu çalışmada bazı metallerin α-amilaz aktivitesine etkisi incelendiğinde, Mn+2 ve
Ca+2’nin aktivatör etki yaptıkları; Mg+2, Zn+2, Fe+2, Cu+2, Hg+2 ve EDTA’nın ise
inhibitör etki yaptıkları belirlenmiştir. Özellikle Cu+2’nin, P/2 ve P/4 izolatlarının α-
amilazlarının aktivitelerini sırasıyla %95, %96 oranında azalttığı, 70/2 ve CRK
izolatlarının α-amilazlarının aktivitelerini ise tamamen durdurduğu; Hg+2’nin ise dört
bakterinin de α-amilazlarının aktivitelerini tamamen dudurduğu belirlenmiştir. Ca+2’nin
aktiviteyi %3-7 oranında artırdığı, Mn+2’nin, CRK izolatının α-amilazının aktivitesini
%5 azalttığı, P/2, P/4 ve 70/2 izolatlarının α-amilazlarının aktivitelerini ise %7-10
artırdığı gözlenmiştir. Mg+2’nin P/2 izolatının α-amilazının aktivitesine önemli bir etki
yapmadığı; P/4, 70/2 ve CRK izolatlarının α-amilazlarının aktivitelerini %12-25
azalttığı belirlenmiştir. Bakterilerin α-amilazlarının aktivitelerini; Zn+2’nin %28-48,
Fe+2’nin %18-39, EDTA’nın ise %15-22 azalttıkları saptanmıştır (Çizelge 4.5).
Bacillus licheniformis MY 10 ile yapılan bir çalışmada enzim çözeltisinin çeşitli
metallerle 30 dakika inkübasyonu sonucunda, 50 mM NaCl varlığında α-amilaz
aktivitesinin %100 olduğu yani NaCl’nin enzim aktivitesine artırıcı ya da azaltıcı bir
şekilde etki etmediği bulunmuştur. 50 mM CaCl2, MgCl2 ve EDTA varlığında
aktivitenin sırasıyla %35, %20 ve %3 oranlarında azaldığı belirlenmiştir (Kumar et al.
1990).
P. woesei α-amilazının 120 oC’de Zn+2 varlığında %20 inhibe edildiği ve stabilite ya da
aktivite için Ca+2’ye ihtiyaç duymadığı belirtilmiştir (Bertodo and Antranikian 2002).
Ayrıca Cr+2, Cu+2, Fe+2 ve Zn+2’nin P. woesei α-amilazını inhibe ettikleri bildirilmiştir
(Ladermann et al. 1993a).

47
Antranikian et al. (1987), Clostridium sp. EM1’in α-amilazına Ca+2 ve Ni+2’nin
etkilerinin olmadığını, enzimi; Mg+2 ve Mo+2’nin %6 oranında, Fe+2’nin %77 oranında
ve EDTA’nın %20 oranında inhibe ettiğini belirtmişlerdir. Ayrıca Mn+2’nin aktiviteyi
%2 oranında artırdığını saptamışlardır.
Çeşitli metal iyonlarının Bacillus türlerinin α-amilaz enzimi üzerine farklı etkide
bulundukları belirtilmiştir. Srivastava (1987) B. stearothermophilus’un α-amilazına
Ca+2 ve Ba+2’nin aktivatör etki yaptıklarını, Ag+2 ve Fe+2’nin ise çok kuvvetli inhibitör
etki yaptıklarını belirtmiştir. Krishnan ve Chandra (1983) B. licheniformis’in bir
suşundan elde ettikleri α-amilaz enzimine Ca+2 ve Mg+2’nin aktivatör etki yaptığını;
Zn+2, Fe+2, Mn+2, Ag+2 ve Cu+2’nin ise önemli ölçüde inhibitör etti yaptığını
saptamışlardır.
Shaw et al. (1995), yaptıkları bir çalışmada ekstrem termofil bir bakteri olan Thermus
sp. ‘nin α-amilaz özelliklerini incelemişleridir. Enzimin 1, 5, 10 mM Co+2, Mg+2, Fe+2,
Cu+2 ve EDTA ilavesi ile inhibe olduğunu; Ca+2 ilavesinin enzim aktivitesine önemli bir
etkisinin olmadığını; Na+2 ilavesi ile aktivitenin arttığını belirlemişlerdir. Cu+2 ve Fe+2
ile meydana gelen inhibisyonun çok fazla olduğu ve enzim aktivitesinin sırasıyla %4 ve
%20 civarına düştüğü belirtilmiştir.
Albayrak (1994), Bacillus subtilis (5α-2)’in α-amilaz aktivitesinin 0.3 mM EDTA
konsantrasyonuna kadar arttığını ve daha yüksek konsantrasyonlarda azaldığını
saptamıştır.
Themoanaerobacter B6A’nın α-amilaz aktivitesi üzerine 10 mM çeşitli metal
çözeltilerinin etkilerinin incelendiğinde, enzimin; Ca+2 ve Mn+2 ile %100, Co+2 ve Mg+2
ile %78, Zn+2 ile %1 ve Fe+2 ile %10 oransal aktivite değeri gösterdiği belirlenmiştir.
Ayrıca α-amilaz aktivitesi üzerine EDTA’nın fazla etkili olmadığı 10 mM EDTA
konsantrasyonunda bile enzimin %89 aktivite gösterdiği saptanmıştır (Saha et al. 1990).
Brown et al. (1990), EDTA’nın P. furiosus α-amilaz aktivitesini önemli ölçüde
azalttığını belirlemişlerdir.

48
Deshpande and Cheryan (1984), metaller içinde özellikle Ca+2’nin, α-amilaz tarafından
nişastanın hidrolizi için gerekli bir metal olduğunu belirtmişlerdir.
Yapılan bu çalışma sonucunda ülkemizdeki çeşitli kaunaklardan izole edilerek
tanımlanan Thermoanaerobacter cinsine ait olan P/2, P/4, 70/2 ve CRK izolatlarının α-
amilaz enzim aktivitelerinin çeşitli faktörlerden etkikendiği ve optimum koşullar
sağlandığında kolay ve ekonomik proseslerle üretilebilecekleri belirlenmiştir. Ancak,
endüstriyel porseslerde kullanılmayan bu grup enzimlerin kolay ve ucuz yöntemlerle
üretildiğinde biyoteknolojik olarak kullanım olanağı bulabilecekleri düşünülmektedir.

49
KAYNAKLAR
Adams, M.W. 1993. Enzymes and proteins from organisms that grow near and above
100 degrees C. Annu Rev Microbiol, 47: 627–58.
Afunasyeva, V. R., Gridreva, T.V., Zaborna, O.E. and Bond, G.L. 1978. Glucose
catabolite repression of glucoamylase biosynthesis by the yeast Endomycropsis
fibiuligera. Priki. Biokhim Mikrobiol. 14:878-884.
Aiyer, P.V. 2005. Amylases and their applications. African Journal of Biotechnology, 4
(13); 1535-1529.
Albayrak, N., Dönmez, S. ve Balk, M. 1994. Toprak ve su örneklerinden amilolitik
Bakterilerin izolasyonu ve bazı enzimatik özelliklerinin belirlenmesi. Turkish
Journal. of Biology, 20, 47-54.
Annous, B.A. and Blaschek, H.P. 1990. Regulation and Localization of amylolytic
enzymes in Clostridium acetobutylicum ATTC 824. Applied and
Environmental Microbiology, p. 2559-2561.
Anonymous 1988. Handbook of amylases and related enzymes. Edited by the
Amylase Research Society of Japan, Permagan Press Oxford.
Antranikian, G., Herzberg, C. and Gottschalk, G. 1987. Production of thermostable
α-amylase, pullulanase and α-glucosidase in continuous culture by a new
Clostridium isolate. Applied and Environmental Microbiology, p. 1668-1673.
Bajpai, P. and Bajpai, P.K. 1991. Increased production of thermostable α-amylase
Enzyme by Bacillus sp. TCRDC-25A with maltodextrins. Applied
Biochemistry and Biotechnology, vol. 31, p. 159-164.
Banks, W. and Greenwood C.T. 1975. Starch and its components. Edinberg
University Press.
Baysal, Z., Uyar, F. and Aytekin, C. 2003. Production of α-amylase by thermotolerant
Bacillus subtilis in the presence of some carbon, nitrogen containing
compounds and surfactants. Annals of microbiology, vol. 53, pp. 323-328.
Becker, P., Abu-Reesh, I., Markossian, S., Antranikian, G. and Markl, H. 1997.
Determination of the kinetic parameters during continuous cultivation of the
lipase-producing thermophile Bacillus sp. IHI-91 on olive oil. Appl Microbiol
Biotechnol, 48:184–90.

50
Berkhout, F. 1976. The manufacture of maize starch. In: Starch Production Technology.
J. A. Radley (ed.) Applied Science Pub. Ltd., London. pp. 109-133.
Bertoldo, C. and Antranikian, G. 2002. Starch hidrolyzing enzymes from thermophilic
archaea and bacteria. Current Opinion in Chemical Biology, 6:151-160.
Brock, T.D. 1986. Introduction: an overview of the thermophiles, In T.D. Brock
(ed.), Thermophiles: general, molecular and applied microbiology. John Wiley
and Sons, Inc., p. 1-16, New York.
Brock, T.D., Brock, K.M., Belly, R.T. and Weiss, R.L. 1972. Sulfolobus: a new genus
of sulfur-oxidizing bacteria living at low pH and high temperature. Arch.
Microbiol. 84: 54-68.
Brown, S.H., Costantino, H.R. and Kelly, R.M. 1990. Characterization of amyloliytic
enzyme activities associated with the hyperthermophilic archaebacterium
Pyrococcus furiosus. Applied and Environmental Microbiology, p. 1985-1991.
Buonocore, V., Caporale, C., De Rosa, M. and Gambacorta, A. 1976. Stable, inducible
thermoacidophilic α- amylase from Bacillus acidocaldarius. J. Bacteriol. 128:
515-521.
Burggraf, S., Jannasch, H.W., Nicolaus, B. and Stetter, K.O. 1990. Archaeoglobus
Profundus sp. nov., represents a new species within the sulfate-reducing
archaebacterium. Syst. Appl. Microbiol. 13: 24-28.
Canganella, F., Andrade, C.M. and Antranikian, G. 1994. Characterization of
amylolytic and pullulytic activities from thermophilic archaea and from a new
Fervidobacterium species. Applied Microbiologyand Bacteriology in press.
Cowan, D.A. 1992. Biochemistry and molecular biology of the extremely thermophilic
archaeobacteria. In: Herbert, RA, Sharp, RJ, editors. Molecular biology and
biotechnology of extremophiles. London: Blackie and Son, pp. 1–43.
Crab, W. and Mitchinson, C. 1997. Enzymes involved in the processing of starch to
sugars. Trends Biotechnol. 15: 349-352.
Çoral, G ve Çolak, Ö. 2000. Bir Aspergillus niger izolatına ait glikoamilaz enziminin
izolasyonu ve karakterizasyonu. Turk J Biol, 24: 601–609.
Danson, M.J and Hough, D.W. 1998. Structure, function and stability of enzymes
from the Archaea. Trends Microbiol, 6:307–14.

51
De Rosa, M., Morana, A., Riccio, A., Gambacorta, A., Trincone, A. and Incani O. 1994.
Lipids of the archaea: a new tool for bioelectronics. Biosens. Bioelectr.
9, 669–675.
De Souza Teodoro, C.E. and Martins, M.L.L. 2000. Culture conditions for the
production of thermostable amylase by Bacillus sp. Braz. J.
Microbiol. vol.31 no.4.
Deshpande, S.S. and Cheryan, M. 1984. Effects of Phytic acid, divalent cations and
their interactions on amylase activity. J. Food Science, vol. 49, 516-519.
Demirjian, D.C., Francisco, M.V. and Constance, S.C. 2001. Enzymes from
extremophiles. Current Opinion in Chemical Biology, 5: 144-151.
Eichler, J. 2001. Biotechnological uses of archaeal ekstremozymes. Biotechnology
Advances, 19: 261- 278.
Ekşi, A. 1998. Meyve Suyu Durultma Tekniği. Gıda Teknolojisi Derneği Yayınları
No:9. 127 s. Ankara.
Ensley, B., McHugh, J.J. and Barton, L.L. 1975. Effect of carbon sources on formation
of ot-amylase and glucoamylase by Clostridium acetobutylicum. J. Gen. Appl.
Microbiol. 21:51-59.
Fitter, J., Herrmann, R., Hauss, T., Lechner, R.E. and Dencher, N.A. 2001. Dynamical
Properties of alpha-amylase in the folded and unfolded state: the role of
thermal equilibrium fluctuations for conformational entropy and protein
stabilization. Physica B: Condens. Matter. 301, 1–7.
Fogarty, M. 1983. Microbial enzymes and biotechnology. Applied Science Publishers,
England. Fontes, C., Hazlewood, G.P., Morag, E., Hall.
Gomes, J. and Steiner, W. 2004. Extremophiles and extremozymes. Food Technol.
Biotechnol. 42 (4) 223–235.
Goyal, N., Gupta, J.K. and Soni, S.K. 2005. A novel raw starch digesting thermostable
α-amylase from Bacillus sp. 1-3 and its use in the direct hydrolysis of raw
potato starch. Enzyme and Microbial Technology 37: 723-734.
Gupta, R., Gigras, P., Mohapatra, H., Goswami, V.K. and Chauhan, B. 2003.
Microbiyal α-amylases: A Biotechnological Perspective. Process
Biochemistry. 38: 1599-1616.

52
Gümüşel, F. 2002. Biyoteknoloji, genetik ve sağlık sektörü. Kocaeli Sanayii için
teknolojik uzgörü ortak projesi. s. 73-135.
Güvenç, U. ve Üstünes, H. 1987. B. subtilis ve B. amyloliquefaciens α-amilazlarının
oluşumu ve bazı fizikokimyasal özellikleri. E.Ü. Mühendislik Fakültesi, Seri:
B Gıda Mühendisliği, Cilt: 5, Sayı: 1.
Haki, G.D. and Rakshit, S.K. 2003. Developments in industrially important
thermostable enzymes: a review. Bioresource Technology 89: 17–34.
Herbert, R. and Sharp, R. 1992.Molecular biology and biotechnology of ekstremophiles.
Chapman and Hall, NY.
Hyun, H.H. and Zeikus, J.G. 1985. General biochemical characterization of
thermostable pullulanase and glucoamylase from Clostridium
thermohydrosulfuricum. Applied and Environmental Microbiology,
79(5): 1168-1173.
Jaenicke, R. and Bohm, G. 1998. The stability of proteins in extreme environments
Curr Opin Struct Biol, 8:738–48.
Kengen, S.W.M., Stams, A.J.M. and de Vos, W.M. 1996. Sugar metabolism of
hyperthermophiles. FEMS Microbiol. Rev. 18: 119-137.
Kıran, Ö.E. ve Çömlekçioğlu, U. 2003. Zeytinli Ilıcası (Kahramanmaraş)’ndan termofil
alkafilik amilolitik Bacillus sp. suşlarının izolasyonu ve amilaz üretme
yetenekleri üzerine azot kaynaklarının etkisi. KSÜ Fen ve Mühendislik Dergisi
6(2): 41-48..
Klingeberg, M., Galunsky, B., Sjoholm, C., Kasche, V. and Antranikian, G. 1995.
Purification and properties of highly thermostable, sodium dodecyl sulfate-
Resistant and stereospecific proteinase from the extremely thermophilic
Archaeon Thermococcus stetteri, Applied and Environmental Microbiology,
61(8): 3098.
Kocabaş, E.E. and Dizbay, M. 1999. Bacillus stearothermophilusvar. calidolactis
C953’ten α-galaktosidaz Üretimi. Tr. J. of Biology 23: 47-54.
Koch, R., Spreinat, A., Lemke, K. and Antranikian, G. 1991. Purification and properties
of a hyperthermoactive α-amylase from the archaebacterium Pyrococcus
woesei. Archives of Microbiology 155, 572-578.

53
Krahe, M., Antranikian, G. and Markl, H. 1996. Fermentation of extremophilic
microorganisms. FEMS Microbiol Rev, 18:271–85.
Krishnan, T. and Chandra, A.K. 1983. Purificationand characterization of α-amylase
From Bacillus licheniformis CUM C305. Appl. Envir. Microbiol. 46(2),
430-437.
Kumar, S. and Nussinov, R. 2001. How do thermophilic proteins deal with heat? Cell
Mol. Life Sci. (58) 1216-1233.
Kumar, S.U., Rehana, F. and Nand, K. 1990. Production of an extracellular
Thermostable calcium-inhibited α-amylase by Bacillus licheniformis MY 10.
Enzyme Microb. Technol., vol. 12.
Landermann, K.A., Davis, B.R., Krutzsch, H.C., Lewis, M.S., Griko, Y.V., Privalov,
P.L. and Anfinsen, C.B. 1993a. The purificationand characterization of an
extremely thermostable α-amylase fromthe hyperthermophilic archaebacterium
Pyrococcus furiosus. Journal of Biological Chemistry, 268, 24394-24401.
Landermann, K.A., Asada, K., Uemori, T., Mukai, H., Taguchi, Y., Kato, I. and
Anfinsen, C.B. 1993b. α-amylase from the hyperthermophilic archaebacterium
Pyrococcus furiosus. Journal of Biological Chemistry 268, 24402-24407.
Legin, E., Copinet, A. and Duchiron, F. 1998. Biotechnology Letters, Vol 20, No 4,
pp. 363-367.
Legin, E., Ladrat, C., Godfroy, A., Barbier, G. and Duchiron, F. 1997. Thermostable
Amylolytic enzymes of thermophilic microorganisms from deep-sea
hydrothermal vents. 1997. Life Sciences, 320, 893-898.
Lopez, G. 1999. DNA supercoiling and temperature adaptation: a clue to early
diversi.cation of life. J. Mol. Evol. 46, 439–452.
Lowe, S.E., Jain, M.K. and Zeikus, J.G. 1993. Biology, ecology, and biotechnological
applications of anaerobic bacteria adapted to environmental stresses in
temperature, pH, salinity, or substrates. Microbiol Rev. Jun;57(2):451–509.
Madi, E., Antranikian, G., Ohmiya, K. and Gottschalk, G. 1987. Thermostable
amylolitic enzymes from a new Clostridium ısolate. Applied and
Environmental Microbiology, p. 1661-1667.
Madigan, M. T. and Marrs, B. L. 1997. Extremophiles. Sci. Am. (276) 66–71.

54
Madsen, G.B. and Norman, B.E., Slott, S. 1973. A new heat stable bacterial amylase
and its use in high temperature starch liquefaction. Die Starke. 25: 304-308.
Mamo, G. and Gessesse, A. 1999. Effect of cultivation conditions on growth and
α-amylase production by a thermophilic Bacillus sp. Letters in Applied
Microbiology Volume 29, Page 61.
Manning, G.B. and Campbell, L.L. 1961. Thermostable a-amylase of Bacillus
stearothermophilus. J. Biol. Chem., 236: 2952- 2957.
Narang, S. and Satyanarayana, T. 2001. Thermostable α-amylase production by an
extreme thermophile Bacillus thermooleovorans. Letters in Applied
Microbiolog Volume 32 Page 31.
Niehaus, F., Bertoldo, C., Kahler, M. and Antranikian, G. 1999. Extremophiles as a
source of novel enzymes for industrial application. Appl Microbiol
Biotechnol, 51:711–29.
Ohlsson, E. 1930. The two components of malt diastase, especially with reference
to the mutarotation of the products formed in the hydrolysis of starch.
Physiol. Chem. 189: 17-63.
Pandey, A., Nigam, P., Soccol, C.R., Soccol, V.T., Singh, D. and Mohan, R. 2000.
Advances in microbial amylases. Biotechnol. Appl. Biochem. 31:135- 52.
Poonam, N. and Dalel, S. 1995. Enzyme and microbial systems involved in starch
processing. Enzyme Microb. Technol. 17: 770-778.
Rakshit, S.K. 1998. Utilization of starch industry wastes. In: Martin, A. (Ed.),
Bioconversion of Waste Materials to Industrial Products, second ed. Blackie
Academic and Professional.
Rao, M.B., Tanksale, A.M., Gathe, M.S. and Deshpande, V.V. 1998. Molecular and
biotechnological aspects of microbial proteases. Micrbiol. Mol. Biol. Rev.
62(3):597-635.
Rothschild, L. J. and ,Manicinelli, R. L. 2001. Life in extreme environments, Nature,
(409) 1092–1101.
Saboto, D., Nucci, R., Rossi, M., Gryczynski, I., Gryczyniski, Z. and Lakowicz, J. 1999.
The β-glycosidase from the hyperthermophilic archaeon Sulfolobus
solfataricus: enzyme activity and conformational dynamics at temperatures
above 100 oC. Biophys. Chem. 81, 23–31.

55
Sarıkaya, E. 1995. α-amilaz üreten bazı Bacillus suşlarının gelişme parametreleri, enzim
özellik ve üretim koşullarının optimizasyonu. Ankara Üniversitesi Fen
Bilimleri Enstitüsü Doktora tezi 2s.
Sarikaya, E., Higassa, T., Adachi, M. and Mikami, B. 2000.Comparison of degradation
abilities of α- and β-amylases on raw starch granules. Proc. Biochem. 35:
711-715.
Sellek, G.A. and Chaudhuri, J.B. 1999. Biocatalysis in organic media using enzymes
from extremophiles. Enzyme Microb Technol, 25: 471-482.
Shaw, J., Lin, F, Chen, S. and Chen, H. 1995. Purification and properties of an
extracellular a-amylase f rom Thermus sp. Bot. Bull. Acad. Sin. 36:
195-200.
Sindhu, G.S., Sharma, P., Chakrabarti, T. and Gupta, J.K.1997. Strain improvement for
the production of a thermostable α-amylase. Enzyme Microb.
Technol, 24:584-589.
Srivastava, R.A.K. 1987. Purification and chemical characterization of thermostable
amylases produced by Bacillus stearothermophilus. Enzyme Microb.
Technol. Vol. 9.
Srivastava, R.A.K. and Baruah, J.N. 1986. Culture conditions for production of
thermostable amylase by Bacillus stearothermophilus. Applied and
Environmental Microbiology, p. 179-184.
Sterner, R. and Lieble, W. 2001. Thermophilic adaptation of proteins. Crit. Rev.
Biochem Mol. (36) 39-106.
Stetter, K.O. 1996. Hyperthermophilic procaryotes. FEMS Microbiol. Rev. 18:149-158.
Suzuki, Y., Kishigami, T. and Abe, S. 1976. Production of extracellular α-glucosidase
by a thermophilic Bacillus species. Applied and Environmental Microbiology,
p. 807-812.
Underkofler, L.A., Danault, L.J. and Hou, E.F. 1965. Enzymes in starch industry Die
Starke 17: 179-184.
Üstünes, H. ve Güvenç, U. 1985. Bira ve alkol üretiminde kullanılan bazı enzimlerin
Mikrobiyal Yolla Eldesi. Ege Üniv. Müh. Fak. Dergisi, 3(1): 85-101.

56
Van der Maarel, M., Van der Veen, B., Uitdehaag, H., Leemhuis, H. and Dijkhuizen, L.
2002. Properties and applications of starchconverting enzymes of the
α-amylase family. J. Biotechnol. 94, 137–155.
Vihinen, M . and Mantsala, P. 1990. Characterization of a thermostable Bacillus
stearothermophilus α-amylase, Biotech. Appl. Biochem., 12: 427-435.
Wiegel, J. 1998. Anaerobic alkalithermophiles, a novel group of extremophiles.
Extremophiles 2: 257-267.
Wiseman, A. 1987. Handbook of enzyme biotechnology. Second Edition. Chapter 3.
The Application of Enzyme in Industry p. 274-373.
Woese, C.R., Kandler, O. and Wheelis, M.L. 1990. Towards a natural system of
organisms-proposal for the domain archaea from soil. Proc. Natl. Acad. Sci.
USA, 87: 4576-4579.
Worthington, C.C. 1988. Worthington enzyme manual, enzymes and related
biochemicals. Α-amylase, pp. 38-39, New Jersey.
Zeikus, J.G. and Wolfe, R.S. 1972. Methanobacterium thermoautotrophicum sp. nov.,
an anaerobic, autotrophic, extrem thermophile. J. Bacteriol. 109: 707-713.
Zilling, W., Holz, I., Janekovic, D., Klenk, H.P., Imsel, E., Trent, J., Wunderl, S., Forja,
V.H., Coutinho, R. and Ferreira, T. 1990. Hyperthermus butylicus, a
Hyperthermophilic sulfur-redusing archaebacterium that ferments peptides. J.
Bacteriol. 172:3959-3965.

57
ÖZGEÇMİŞ
Adı Soyadı : Gülay FUKARA
Doğum Yeri : Ankara
Doğum Tarihi : 23.09.1981
Medeni Hali : Bekar
Yabancı Dili : İngilizce
Eğitim Durumu
Lise : Yeşilöz Süper Lisesi, Ankara (1995-1999)
Lisans : Mersin Üniversitesi Mühendislik Fakültesi Gıda Mühendisliği
Bölümü (2000- 2004)
Yüksek Lisans : Ankara Üniversitesi Fen Bilimleri Enstitüsü Gıda Mühendisliği
Anabilim Dalı (Şubat 2005 - Mayıs 2007)





























