Embed Size (px)
Citation preview

1
Reduced Permeability to Rifampicin by Capsular Thickening as a Mechanism of 1
Antibiotic Persistence in Mycobacterium tuberculosis 2
3
4
Jees Sebastian, Sharmada Swaminath, Parthasarathi Ajitkumar* 5
Department of Microbiology and Cell Biology, Indian Institute of Science, Bangalore, 6
Karnataka, India. 7
8
Running title: Rifampicin persistence in Mtb by capsular thickening 9
10
*Address correspondence to P. Ajitkumar, [email protected]. 11
12
Key words: Mycobacterium tuberculosis; Rifampicin permeability; Persistence; Thick 13
outer layer; Polysaccharides 14
15
16
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

2
ABSTRACT 17
Persisters constitute a subpopulation of bacteria that can tolerate lethal concentrations 18
of antibiotics. Multiple mechanisms have been suggested for bacterial persistence 19
against antibiotics. With mycobacteria being no exception to this behaviour, we had 20
reported the de novo emergence of genetically antibiotic-resistant Mycobacterium 21
tuberculosis from persister cells upon prolonged exposure to microbicidal 22
concentrations of the anti-tuberculosis drugs, rifampicin and moxifloxacin. Here, we 23
present evidence for reduced permeability to rifampicin as a mechanism for 24
persistence of Mycobacterium tuberculosis in vitro. We observed that rifampicin 25
persistent M. tuberculosis cells developed a thick outer layer (TOL) capsule. The TOL 26
restricted the entry of fluorochrome-conjugated rifampicin, 5-carboxyfluorescein-27
rifampicin (5-FAM-rifampicin), which retained only 2.5% of its original bactericidal 28
activity, but high levels of permeability, on actively growing mid-log phase cells. Gentle 29
mechanical removal of TOL significantly enhanced 5-FAM-rifampicin entry into the 30
persister cells. The level of 5-FAM-rifampicin in the persister cells was not affected by 31
the pre-incubation of the cells with verapamil, a drug efflux pump inhibitor, ruling out 32
the involvement of efflux pumps in the reduced intracellular concentration of 5-FAM-33
rifampicin. GC-MS analysis of TOL showed the presence of ~7-fold, ~5-fold and ~2-34
fold higher levels of α-D-glucopyranoside, 1,2,5-linked-mannitol, and 3,4-linked 35
mannose, respectively, among ~2-fold higher levels of derivatives of several other 36
types of sugars such as arabinose and galactose. Taken together, the present study 37
reveals that rifampicin-persistent M. tuberculosis cells develop TOL that enables the 38
bacilli to restrict entry of rifampicin and thereby remain tolerant to the antibiotic in vitro. 39
40
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

3
INTRODUCTION 41
Mycobacterium tuberculosis, which is the causative agent of tuberculosis, is one of the 42
most successful human pathogens due to its ability to survive under diverse extreme 43
stress conditions. Like many other pathogenic bacteria, M. tuberculosis also exhibits 44
several survival strategies to overcome the lethality imposed by antibiotic exposure. 45
One of the unique features of M. tuberculosis that has enabled the bacilli to remain 46
refractory to many commonly used antibiotics is the distinct cell wall structure (1). The 47
complex cell wall of M. tuberculosis is well known for imposing limited permeability to 48
several host-derived antimicrobial biomolecules that makes the bacilli tolerant to many 49
toxic components from its normal habitats (2-5). Studies have been conducted on the 50
physiological significance of the changes that the cell wall undergo inside 51
macrophages (6), under extreme nutritional stress conditions (7) and hypoxia (8). 52
However, there is no information on whether the cell envelope has any contribution to 53
the persistence of the bacilli against antibiotics. 54
Persistence is one of the survival strategies of mycobacteria to remain tolerant 55
to lethal concentrations of antibiotics. Although several mechanisms that contribute to 56
mycobacterial persistence have been reported (9, 10), very little information is 57
available on the ultrastructural features of mycobacterial antibiotic persisters and their 58
contribution, if any, to antibiotic tolerance. Earlier studies in Escherichia coli and 59
Staphylococcus aureus have shown that exposure to a sub-lethal concentration of 60
antibiotics can cause the emergence of resisters through the generation of reactive 61
oxygen species and consequential mutagenesis (11). Subsequently, in our study, M. 62
tuberculosis persisters were found in the presence of lethal concentrations of 63
rifampicin and moxifloxacin, leading to the de novo emergence of antibiotic resisters 64
(12). These two studies raised the possibility that the bacilli might have some hitherto 65
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

4
unknown strategy to ensure sub-lethal concentration of antibiotics inside persister cells 66
even though they remain exposed to lethal concentrations of the antibiotics. In order 67
to verify this possibility, we examined whether the permeability of rifampicin into M. 68
tuberculosis persister cells was affected by any ultrastructural changes. The present 69
study reports the ultrastructural changes of rifampicin persistent M. tuberculosis cells 70
and their contribution to the restricted permeability of rifampicin, in comparison with 71
the entry of rifampicin into actively growing mid-log phase cells. 72
73
MATERIALS AND METHODS 74
75
Rifampicin treatment of Mycobacterium tuberculosis 76
Mycobacterium tuberculosis H37Ra (National JALMA Institute of Leprosy and Other 77
Mycobacterial Diseases, Agra, India) was used in all the experiments. Bacteria were 78
grown in Middlebrook 7H9 broth (BD) supplemented with 0.2% glycerol, 0.05% 79
Tween-80 and 10% ADS (albumin, dextrose, sodium chloride) until mid-log phase 80
(MLP, 0.6 at OD600 nm) at 37ºC under shaking at 170 rpm. Cultures were then exposed 81
to 1 µg/ml (10x MBC; 12) of freshly prepared rifampicin (Sigma) under the same 82
condition. Aliquots of the culture were withdrawn once in every 24 hrs and used for 83
serial dilution and plating on sterility checked Middlebrook 7H10 agar supplemented 84
with 10% ADS in the absence of rifampicin. Plates were sealed with parafilm and 85
incubated at 37ºC in CO2 incubator (5% CO2) for determining CFU. For experiments 86
using 5-FAM-rifampicin (Merck-Millipore), MLP or persistence phase cells were 87
exposed to 1.5 µg/ml (concentration equimolar to 10x MBC rifampicin) for the required 88
period and taken for flow cytometry analysis (see below). 89
90
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

5
Transmission electron microscopy 91
M. tuberculosis cells from different stages of rifampicin treatment were harvested and 92
processed for transmission electron microscopy as previously described (13). In brief, 93
the cells were prefixed in 1% (w/v) osmium tetroxide (solution in double-distilled water) 94
in 0.15 M cacodylate buffer (pH 7.2) for 1 hr at room temperature and washed once in 95
the same buffer. Further, cells were fixed using 2% (v/v) glutaraldehyde and 2% (w/v) 96
tannic acid in 0.15 M cacodylate buffer for 2 hrs at room temperature. Cells were 97
washed once in the same buffer and re-fixed in 1% osmium tetroxide overnight at 4ºC. 98
Samples were dehydrated by a series of ethanol washes (with 25%, 50%, 75% and 99
95% ethanol) (5 min incubation between every wash) and infiltrated with 50% LR white 100
resin in 50% ethanol for 24 hrs at 4°C. Dehydrated cells were collected by 101
centrifugation and the pellet was used for making blocks in gelatine capsules with 102
100% LR white resin. Blocks were solidified by incubating at 65°C for 2-3 days in a 103
dry bath. Ultrathin sections of 80 to 100 nm were prepared from the blocks using 104
ultramicrotome and collected on copper grids. Sections were stained with uranyl 105
acetate (0.5%) and lead citrate (0.04%) and observed under JEOL-100 CX II electron 106
microscope at 100 kV. 107
For ruthenium red staining, we followed the previously reported method (14). In 108
brief, the cells were harvested and washed once in Middlebrook 7H9 medium and 109
prefixed in the presence of paraformaldehyde and glutaraldehyde (2.5% w/v each), 110
CaCl2 and MgCl2 (5 mM each) and 0.05% (w/v) ruthenium red in 0.1 M cacodylate 111
buffer (pH 7.2) for 2 hrs at 4ºC. Cells were washed once in the same buffer and 112
incubated with 2.5% w/v glutaraldehyde and 0.05% (w/v) of ruthenium red in 113
cacodylate buffer. Cells were washed once and post-fixed in osmium tetroxide (OsO4) 114
and uranyl acetate (1% w/v of each) containing ruthenium red (0.05% w/v). Fixed cells 115
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

6
were dehydrated with a series of ethanol washes (with 25%, 50%, 75% and 95% 116
ethanol) (5 min incubation between every wash, ethanol contained 0.05% w/v of 117
ruthenium red) and infiltrated with 50% LR white resin in 50% ethanol for 24 hrs at 118
4°C. Cells were collected by centrifugation and the pellet was used for making blocks 119
in gelatine capsules with 100% LR White resin. Blocks were solidified by incubating at 120
65°C for 2-3 days in dry bath. Ultrathin sections of 80-100 nm were prepared from the 121
blocks using ultra-microtome and placed on copper grids. Sections were stained with 122
uranyl acetate (0.5%) and lead citrate (0.04%) and observed under JEOL-100 CX II 123
electron microscope at 100 kV. 124
125
Bacterial hydrophobicity assay 126
A modified protocol of a previously reported method (15, 16) was used. Cells from 127
MLP and rifampicin persistence phase were harvested by centrifugation and 128
resuspended in filter sterilised PUM buffer (100 mM K2HPO4.3H2O, 50 mM KH2PO4, 129
33.3 mM urea, 1 mM MgSO4.7H2O, in double-distilled autoclaved water, pH 7.1) to 130
get an approximate density of 0.7 at OD600 nm. Phase extraction was performed in 131
siliconised borosilicate tubes using one volume of PUM buffer containing cells against 132
three volumes of n-hexadecane and vortexed for 8-10 seconds and left at room 133
temperature for 15 min. The aqueous phase was collected into siliconised 134
microcentrifuge tubes using siliconised tips, mildly sonicated and used for serial 135
dilution and plating on sterility checked Middlebrook 7H10 agar plates containing 10% 136
ADS supplement. The difference in the CFU of the aqueous phase before and after 137
phase separation was used for calculating the percentage of cells with hydrophilic 138
outer layer. 139
140
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

7
Estimation of zeta potential of mycobacterial persister cells 141
For the determination of surface charge of M. tuberculosis cells, aliquots of the culture 142
were withdrawn on different days during rifampicin treatment and washed once with 143
Middlebrook 7H9 broth. Cells were resuspended in fresh medium and used for zeta 144
potential measurement using zeta sizer nano series (Nano-ZS90, Malvern 145
Instruments). For the estimation of the isoelectric point of the cells, the cells were 146
resuspended in solutions of varying pH from 2 to 10 in PPMS buffer (40 mM of 147
K2HPO4, 20 mM of KH2PO4, and 1.5 mM of MgSO4.7H2O per litre of Milli Q water, pH 148
adjusted using HCl or NaOH). Zeta potential values of the cells at different pH were 149
calculated and the graph was plotted, as described (17, 18). 150
151
GC-MS analysis of OL 152
Cells from MLP and persistence phase were collected and used for OL extraction as 153
previously reported (19) with minor modifications. Briefly, cells were washed once in 154
1x PBS and resuspended in 20 ml distilled water followed by incubation with 10 gm of 155
sterile 4 mm glass beads at 50 rpm for 15 min. The cell suspension was collected and 156
centrifuged at 12000 xg for 10 min. The supernatant was filtered through a 0.2 micron 157
filter and the filtrate was lyophilized. The sample was derivatised and subjected to GC-158
MS performed at Mass spectrometry glycomics facility at C-CAMP Bangalore. Data 159
analysis was also performed at the same facility. 160
161
Fluorescence microscopy 162
Cells from MLP and persistence phases were harvested and resuspended in 100 µl of 163
Middlebrook 7H9 broth. For the staining of cells, 5-FAM-rifampicin (1.5 µg/ml) or 164
calcofluor white (CFW) (1:1000 dilutions of 0.1% solution, Sigma) was added into the 165
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

8
cell suspension and incubated for 1 hr at 37ºC incubator shaker. Propidium iodide 166
(1:1000) was added (to discriminate dead cells) and layered over poly-L-lysine treated 167
multiwell slides 20 min in the dark. Wells were washed once with phosphate buffered 168
saline (PBS) and mounted with glycerol and cover slips and observed under 100X 169
objective in Zeiss AxioVision fluorescence microscope. Bead beating was performed 170
by incubating 20 ml culture with 10 grams of glass beads (4 mm) at 37ºC for 15 min 171
at 50 rpm, prior to the addition of 5-FAM-rifampicin and processed similarly as above. 172
173
Flow cytometry analysis 174
Cell suspension (500 µl) was exposed to 1.5 µg/ml of 5-FAM-rifampicin and incubated 175
at 37ºC incubator for 1 hr in the dark. Aliquots were collected at specified intervals and 176
cells were harvested by centrifugation at 12000 x g for 5 min at 4ºC and washed once 177
with ice-cold Middlebrook 7H9 broth and used for flow cytometry analysis. For bead 178
beating, 20 ml culture was incubated with 10 grams of glass beads (4 mm) at 37ºC for 179
15 min at 50 rpm prior to the addition of 5-FAM-rifampicin and processed similarly for 180
flow cytometry with 488 nm solid state laser and 527/32 nm emission filter (for GFP) 181
at low or medium flow rate. For relative permeability estimation of rifampicin persisters 182
for 5-FAM-rifampicin, cells from MLP, persistence phase and verapamil-pretreated (50 183
µg/ml) persisters were incubated with 1.5 µg/ml of 5-FAM-rifampicin and incubated for 184
1 hr at 37ºC in the dark and processed in the same way as described earlier. 185
For CFW staining and OL analysis, 500 µl aliquots of the cells before and after 186
bead beating were incubated with 1:1000 dilutions of CFW (0.1% solution, Sigma) for 187
1 hr at 37ºC. Cells were washed once with Middlebrook 7H9 broth and used for flow 188
cytometry analysis in BD FACSVerse flow cytometer. 189
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

9
About 10000 gated cells were analysed and used for the estimation of the median 190
fluorescence value of the 5-FAM-rifampicin stained cells while keeping the 191
autofluorescence at 2-log10 value. The photomultiplier tube (PMT) voltage settings 192
used for measuring the 5-FAM-rifampicin fluorescence of the cells were 208 (FSC), 193
333 (SSC). The instrument was calibrated using FACSuite cytometer set up and 194
tracking beads (CS&T, Becton Dickinson). Data were processed and analysed using 195
FACSuite software and the statistical significance between the time points was 196
calculated using paired t-test of GraphPad Prism version 5.0. 197
198
5-FAM-rifampicin bioassay 199
Rifampicin sensitive Staphylococcus aureus (ATCC 25923) was used for the bioassay 200
of 5-FAM-rifampicin (20). In brief, rifampicin (sigma) and 5-FAM-rifampicin were 201
dissolved in DMSO to make a stock solution of 2 mg/ml. Dilutions were made from the 202
stock solution and used for the agar diffusion assay. LB agar plates were made by 203
mixing 50 µl of the Staphylococcus aureus glycerol stock with 100 ml LB agar (warm 204
to the touch). Wells were made in the agar using a stainless steel puncture with 0.5 205
cm diameter. Known concentrations of rifampicin or 5-FAM-rifampicin solutions (50 µl) 206
were added into the well and incubated overnight at 37°C. The diameter of the zone 207
of inhibition was measured using Vernier caliper and used for plotting the standard 208
graph for rifampicin, from which and the zone of inhibition of 5-FAM-rifampicin, the 209
bioactivity of 5-FAM-rifampicin was calculated. 210
211
5-FAM-rifampicin permeability assay 212
To estimate the extent of 5-FAM-rifampicin entry into MLP cells and rifampicin 213
persisters, cultures were treated with a final concentration of 1.5 µg/ml of the drug 214
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

10
conjugate in 20 ml culture and incubated in a shaker at 37°C. Cultures for both (MLP 215
and persisters) were mildly beaten with 4 mm glass beads to remove the outer 216
capsular layer, as a control for the experiment. Aliquots were taken at every 15 min 217
intervals and quickly washed once with ice-cold Middlebrook 7H9 broth and used for 218
flow cytometry analysis using BD FACSVerse system. Samples before 5-FAM-219
rifampicin addition (0 min) were used as the control for autofluorescence and median 220
of fluorescence was kept at 102. 221
To construct a standard graph for 5-FAM-rifampicin entry, MLP cells were 222
incubated with two-fold increasing concentrations of 5-FAM-rifampicin in an incubator 223
shaker for 1 hr at 37°C. Cells were harvested and quickly washed once with ice-cold 224
Middlebrook 7H9 broth and used for flow cytometry analysis in BD FACSVerse 225
system. Autofluorescence median of the untreated cells was kept at 102 and the 226
normalised median values were used for plotting standard graph. Rifampicin 227
persisters, which were isolated from the 12th day of the treatment, were incubated with 228
5-FAM-rifampicin equivalent of 10x MBC rifampicin. These cells were also similarly 229
processed and readings were taken from 10000 cells. Normalised fluorescence values 230
from six independent persister cell samples were used for calculating the relative 231
concentration of 5-FAM-rifampicin in persister cells from the standard graph. 232
233
RESULTS AND DISCUSSION 234
Earlier studies from our laboratory had shown that actively growing MLP M. 235
tuberculosis cells exposed to 10x MBC concentration of rifampicin consistently 236
showed a triphasic response involving killing phase (0 to 10th day of exposure), 237
persistence phase (10th to 15th day of exposure), followed by regrowth phase (beyond 238
15th day of exposure) (12). As part of our earlier characterisation of the triphasic 239
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

11
response of M. tuberculosis cells to 10x MBC rifampicin, we had found that the 240
rifampicin concentration stays at ~5x MBC even on the 15th day of the prolonged 241
exposure that spanned for 20 days (12). Therefore, for the present ultrastructural 242
studies, the persistence phase cells from the 12th day of exposure to 10x MBC 243
rifampicin were used. 244
245
Rifampicin persister cells possess thick outer layer (TOL) 246
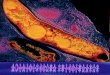
Transmission electron micrographs of rifampicin persister M. tuberculosis cells, 247
stained with tannic acid-lead citrate combination (13), showed a significantly thick, but 248
strikingly uneven, loosely bound and deeply stained outer layer (OL) (Fig. 1A). On the 249
contrary, the MLP cells (control) showed an evenly thin outer layer, which we called 250
normal outer layer (NOL) (Fig. 1B). The TOL thickness of persister cells ranged 251
between 25-130 nm (n = 27), while the MLP cells showed a more-or-less evenly thin 252
layer of NOL with an average thickness of ~20 nm (Fig. 1C). However, the thickness 253
of electron transparent layer (ETL) of the persister and the control cells were found 254
comparable and were morphologically like the already published data (Fig. 1D) (14, 255
21, 22). Since rifampicin exposure for the experimental sample was started from the 256
MLP cells, cultures from MLP stage for an equivalent number of days, but unexposed 257
to rifampicin did not show any TOL (Fig. S1). Thus, the ultrastructural difference 258
between the rifampicin persister cells and the control cells was confined to the outer 259
layer (OL) thickness. 260
Transmission electron microscopy of persister and MLP cells stained with 261
ruthenium red was performed to specifically stain polysaccharides for detection (14). 262
It showed the presence of polysaccharides as an uneven layer on the surface of 263
rifampicin persister cells unlike a very thin layer on the MLP control cells (n = 22) (Fig. 264
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

12
S2). The thin OL on the MLP also contained polysaccharides, as reported (19). The 265
presence of polysaccharides was further confirmed by staining with a polysaccharide 266
specific fluorophore, calcofluor white (CFW; 23, 24) (Fig. 1E, top panel). The persister 267
cells, which were gently bead beaten to remove OL, as described (19), and stained 268
with CFW, showed loss of CFW fluorescence (Fig. 1E, bottom panel). Flow cytometry 269
analysis of CFW stained native and bead beaten persister cells also showed loss of 270
CFW fluorescence. However, the MLP cells (Fig. 1F, H, J) did not show an appreciable 271
difference in CFW fluorescence between the native and the bead beaten samples, 272
compared to persister cells (Fig. 1G, I, J). Thus, by using multiple staining methods 273
with two different polysaccharide specific reagents confirmed the differential 274
polysaccharide content on the MLP and persister cells. 275
276
TOL imparts hydrophilicity to rifampicin persister cells’ surface 277
It was shown that the constituents of the OL of actively growing mycobacteria are 278
mostly polysaccharides and proteins with low lipid content (19). Therefore, due to the 279
thickening of the OL in persister cells, the possibility of a change in the hydrophilicity 280
of the surface of persister cells, as compared to that of the actively growing MLP cells, 281
was verified. For this purpose, cells from the rifampicin persistence phase and MLP 282
were subjected to phase separation between highly hydrophobic hexadecane and 283
aqueous buffer to measure cell-surface hydrophobicity, as described (15). The cells 284
retained in the aqueous phase after the phase extraction reflects the proportion of cells 285
that have a hydrophilic surface. We observed that an average 7% of the persister 286
population was hydrophilic and retained in the aqueous phase, while only 0.03% of 287
MLP cells being hydrophilic (Fig. 2A). This showed that the persister population 288
contained a relatively higher proportion of cells having hydrophilic surface. 289
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

13
290
Rifampicin persister M. tuberculosis cells possess negative surface charge 291
The presence of anionic polysaccharides in the TOL could impart a negative surface 292
charge to the persister cells and confer hydrophilicity. The cell surface negative charge 293
can be measured by the zeta potential (ZP) of intact cells (17, 18). The ZP of M. 294
tuberculosis cells was measured for MLP cells and the cells from the killing, 295
persistence and regrowth phases of rifampicin exposure, which were described before 296
(12). M. tuberculosis MLP cells showed a ZP value of -3.91 mV, with a gradual 297
increase in the negative surface charge over the course of rifampicin exposure (Fig. 298
2B). The gradual increase in the negative ZP value upon continued exposure to 299
rifampicin indicated accumulation of negatively charged molecules on the cell surface. 300
Cells from the killing phase showed a ZP of -19.26 mV indicating a remarkable 301
increase in their negative surface charge compared to that of the rifampicin-unexposed 302
MLP cells. The persistence phase cells showed a peak ZP value of -32.76 mV, 303
showing that the persister cells have significantly high negative surface charge density 304
compared to that of the cells from any other phase of rifampicin exposure. The high 305
negative ZP of persister cells indicated the accumulation of anionic polysaccharides 306
on the cell surface. The regrowth phase cells showed a reduction in the negative ZP 307
value indicating the loss of TOL once the cells gained rifampicin resistance and came 308
out of the persistence phase. In addition, we observed multiple peaks in the ZP 309
histogram of RG phase population, suggesting the possibility of heterogeneity in terms 310
of surface charge among the rifampicin-tolerant/resistant regrowing cells (Fig. 2C). 311
In order to determine ionic properties of the mycobacterial TOL, the isoelectric 312
point (pI) of M. tuberculosis cells was measured at different pH values (2, 4, 6, 8 and 313
10). The pH at which cells show zero ZP value was considered as the pI of the cells. 314
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

14
To estimate the isoelectric point (pI) of M. tuberculosis cells, at different pH values, we 315
measured the ZP of cells from various time points during rifampicin exposure. The 316
positive ZP of MLP cells at pH 2.0 and pH 4.0 were dropped to -2.58 at pH 6.0 317
indicating that the pI of MLP cells were between pH 4.0 and pH 6.0 (Fig. 2D). The 318
persistence phase cells showed a similar pI profile with a lower ZP value at high pH 319
(Fig. 2E). Although the cells in the killing phase also showed a similar pI, the extent of 320
negative ZP potential at high pH was considerably lesser than that of the persistence 321
phase cells (Fig. 2F). Thus, M. tuberculosis cells from MLP, killing and persistence 322
phases showed similar pI values with a varying surface charge at higher pH, probably 323
due to the high levels of anionic polysaccharide content on their surface. The 324
comparable pI values of the MLP cells and persisters indicated that the ionic properties 325
of the TOL might not have changed over the period of rifampicin exposure. This 326
alluded to the possibility that the nature of the molecules might be similar but their 327
relative composition might be different (see GCMS data described below), thereby 328
having similar pI. 329
330
Molecular analysis of TOL composition 331
A comparative molecular analysis using GC-MS was performed to find out the 332
polysaccharide composition of TOL on persistence phase cells and its difference from 333
that of the NOL on MLP cells. For this purpose, the NOL from MLP cells and TOL from 334
persister cells were gently extracted, as described (19) and used for mass 335
spectrometry. The percentage of relative abundance of the monosaccharides detected 336
are listed in Table 1. We observed ~5-fold increase in the levels of 1, 2, 5-mannitol 337
and ~6-7-fold increase in the α-D-glucopyranoside levels. This composition is 338
consistent with an earlier study that the NOL of actively growing M. tuberculosis (19). 339
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

15
The thickening of the OL has caused several fold increase in the levels of the 340
constituents that were present in the NOL of MLP cells (see Table 1). For instance, 341
the 6-7-fold higher levels of α-D-glucopyranoside, as a breakdown product in the GC-342
MS analysis, alluded to the possibility of the presence of trehalose, which is α-D-343
glucopyranosyl-α-D-glucopyranoside (glucose disaccharide), known to desiccate 344
bacterial cells against severe stress conditions (25-27). Similarly, glucan, which is a 345
component of the M. tuberculosis cell surface, is a polymer of glucose that is 346
expressed in vitro and in vivo (28). The presence of high levels of α-D-glucopyranoside 347
might be an indication of the presence of glucan as well. The increased levels of 348
arabinose and mannose probably signify the presence of arabinomannan, which is 349
one of the significant components of OL (1, 29, 30). The high levels of polysaccharides 350
in the OL are known to be used as a bacterial decoy for antimicrobial peptides, for 351
respiratory tract colonisation, pathogenesis, cellular invasion, antiphagocytosis (16, 352
30-32). The presence of high levels of polysaccharides in the TOL of rifampicin 353
persister M. tuberculosis cells brings up another role for the OL polysaccharides in 354
mycobacterial physiology. Although differences in terms of relative abundance of 355
polysaccharides were observed, the molecular composition of OL between the MLP 356
and the persister cells was comparable. 357
358
Persister cells restrict rifampicin permeability 359
Reduced cell wall permeability is known to be a factor contributing to antibiotic 360
resistance in Neisseria meningitidis and Staphylococcus aureus (33-35). In 361
mycobacteria also, the role of membrane permeability in rifampicin resistance in the 362
actively growing populations of Mycobacterium intracellulare has been reported (36). 363
Therefore, presuming that the TOL might restrict the entry of rifampicin, experiments 364
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

16
were carried out to understand the permeability characteristics of rifampicin persister 365
cells. For this purpose, 5-carboxyfluorescin (5-FAM) conjugated rifampicin (5-FAM-366
rifampicin) was used to monitor entry of the same into persister cells, in comparison 367
with its entry into MLP cells. 368
Conjugation of the small hydrophobic fluorophore, 5-FAM, to rifampicin 369
generated two possible isomers of 5-FAM-rifampicin depending on the ester bond 370
formed on the two aliphatic hydroxyl group on rifampicin molecule (Fig. 3A). 5-FAM 371
group was selected due to the nonpolar nature and smaller size of the fluorophore. 372
This was to avoid any kind of alteration in the polarity of the molecule and to minimise 373
the molecular size to maintain its entry into rifampicin-unexposed cells. Thus, it is like 374
rifampicin, which is nonpolar in nature and believed to passively diffuse through the 375
mycobacterial cell wall and accumulate inside the mycobacterial cell within 20 min of 376
exposure (37). Further, the conjugation of 5-FAM to rifampicin reduced the bactericidal 377
activity of the antibiotic to 2.5% of its original bioactivity, as calculated from the bio-378
assay (Fig. 3B; Fig. S3). An average zone of inhibition of 1.77 ± 0.035 cm was 379
obtained for 5-FAM-rifampicin at a concentration of 29.2 µg/ml, while 0.74 µg/ml of 380
rifampicin gave the same zone of inhibition, showing the drop in bioactivity of 5-FAM-381
rifampicin to 2.5% compared to that of rifampicin. Owing to these molecular properties, 382
use of 5-FAM-rifampicin avoided inflicting lethality on the cells which would have 383
otherwise affected the permeability assay. Radioactively labelled native rifampicin was 384
not used as it would have affected the viability of the cells. The concentration-385
dependent entry of 5-FAM-rifampicin into MLP cells confirmed its high level of 386
permeability (Fig. 3C, D). The extent of increase in the permeability showed a linear 387
correlation to the concentration of 5-FAM-rifampicin (Fig. 3E). This indicated that the 388
conjugation of 5-FAM to rifampicin did not affect its uptake by the actively growing 389
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

17
MLP cells. The permeability characteristics of 5-FAM-rifampicin into MLP cells 390
validated that the uptake of 5-FAM-rifampicin by the persister cells could be 391
considered as a measure of its extent of permeability into the persister cells as well, 392
in comparison to MLP cells. 393
MLP and persister cells were incubated with 1.5 µg/ml (concentration equivalent 394
to 10x MBC rifampicin used in our earlier study; 12) of 5-FAM-rifampicin and analysed 395
using fluorescence microscopy. The low levels of fluorescence in the persister cells, 396
as compared to that in the MLP cells, indicated restricted entry of 5-FAM-rifampicin 397
into persister cells (Fig. 4A). Fluorescence microscopy of persister cells, which were 398
gently bead beaten to remove TOL and incubated with 5-FAM-rifampicin, showed 399
enhanced levels of 5-FAM fluorescence indicating increased 5-FAM-rifampicin entry 400
into the persister cells (Fig. 4B). Further, we determined the relative time-dependent 401
entry of 5-FAM-rifampicin into MLP and persister cells, with or without bead beating. 402
For this purpose, the cells were incubated over a period of one hour with 1.5 µg/ml of 403
5-FAM-rifampicin and aliquots were withdrawn every 15 min followed by flow 404
cytometry analysis. Both the native and the bead beaten MLP cells showed a time-405
dependent steady increase in the 5-FAM-rifampicin fluorescence showing that the thin 406
OL of MLP cells did not play any role in the permeability of 5-FAM-rifampicin (Fig. 4C, 407
E, G). On the contrary, incubation of persister cells with 5-FAM-rifampicin did not show 408
any noticeable fluorescence inside the cells (Fig. 4D, F upper panels, H). This 409
indicated restricted permeability of the rifampicin conjugate into the cells. Whereas, 410
the bead beaten rifampicin persisters showed a significant time-dependent increase 411
in the fluorescence for at least upto first 30 min of incubation, followed by a level of 412
saturation, suggesting the increased permeability of 5-FAM-rifampicin (Fig. 4D, F 413
lower panels, H). Thus, the removal of the TOL by bead beating allowed permeability 414
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

18
of 5-FAM-rifampicin into the persister cells. These experiments confirmed the 415
substantial role of TOL to function as a ‘barrier’ to restrict permeability of rifampicin 416
into persister cells. 417
418
Verapamil did not affect the permeability of persisters to 5-FAM-rifampicin 419
Multidrug efflux pumps are known to contribute to antibiotic tolerance in M. 420
tuberculosis (38-40). It was reported that after incubation of rifampicin-exposed M. 421
tuberculosis cells in in vitro cultures and in infected macrophages with the efflux pump 422
inhibitor, verapamil, rifampicin levels inside the cells increased thereby enhancing 423
susceptibility (41, 42). This study showed that verapamil-sensitive efflux pump was 424
involved in the removal of rifampicin from the cells. With this background information, 425
it was of interest to find out whether efflux pumps were involved in the tolerance of M. 426
tuberculosis persister cells to rifampicin in vitro. 427
For this purpose, we exposed rifampicin persister cells to 50 µg/ml of verapamil 428
and used for 5-FAM-rifampicin permeability assay. We did not observe any difference 429
in terms of the 5-FAM-rifampicin fluorescence intensity of verapamil-treated and 430
untreated rifampicin persister cell samples (Fig. 5 A, B, C). It ruled out the possibility 431
of rifampicin efflux as a possible mechanism for the rifampicin-tolerance in persister 432
cells in vitro and further confirmed that the TOL plays a substantial role in the reduced 433
rifampicin permeability into M. tuberculosis persister cells. On the contrary, verapamil 434
was found to be an efflux inhibitor in M. tuberculosis infected mice, zebra fish and 435
macrophages (40, 43). This apparent contradiction with these works is probably due 436
to the difference in the physiological status of the cells exposed to verapamil. While 437
the cells were in the persistence phase in our study, the cells were in the infected 438
animal model and macrophages in the other studies (40, 43). 439
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

19
440
Discussion 441
M. tuberculosis is well known for its drug tolerance and natural resistance to many 442
antibiotics. The unique and complex cell wall features of mycobacteria imposing 443
limited permeability have been attributed to be the reason for its natural antibiotic 444
tolerance. Owing to the peripheral location, OL plays a significant role in macrophage 445
interaction, cell adhesion and pathogenicity of M. tuberculosis (44-46). Further, 446
metabolic and detoxifying enzymes, including penicillinase, urease, phosphatases and 447
superoxide dismutase, are present in the OL (30). The presence of immunomodulatory 448
factors in the OL of pathogenic M. tuberculosis plays a major role in the initial host 449
immune response (47). Previous reports have shown that the OL reduces bacterial 450
interaction with the macrophage in the absence of serum opsonins thereby possess 451
anti-phagocytic activity in M. tuberculosis (16) and in Gram-negative bacteria (31, 48). 452
Also, it has been reported that the capsular polysaccharides prevent the entry of 453
potentially harmful host-derived macromolecules like bactericidal peptides in Gram-454
negative bacteria (49-51). Thus, in addition to the contribution of OL to various 455
physiological aspects of mycobacteria (16, 29; 44-47), the present findings show yet 456
another role for OL in the rifampicin tolerance by persisters. 457
The strategy seemed to be to increase the levels of polysaccharide components 458
of OL significantly and thereby confer higher negative charge to restrict entry of 459
rifampicin. The negative surface charge density was found varying, probably 460
depending upon the extent of casing of the cells by the OL. The gradual increase in 461
the surface charge density of the persister cells might be due to the accumulation of 462
OL components over the cell surface during rifampicin exposure. The decrease in the 463
negative surface charge density during regrowth phase indicated the specific role of 464
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

20
OL thickening during persistence phase as a survival strategy during antibiotic 465
exposure which is no more present on (required for) the bacteria as they have gained 466
resistance and entered regrowth phase. Moreover, the multiple peaks of ZP values in 467
the persister and regrowth phase cells denote the presence of heterogeneous sub-468
populations with different surface charge density depending on the extent of OL 469
thickening of the cells. 470
It is possible that the persisters are getting benefited by the ‘barrier’ effect of TOL 471
in limiting the entry of antibiotics. Restricted antibiotic uptake through outer membrane 472
modification is a known mechanism of rifampicin resistance in the actively growing M. 473
intracellulare and N. meningitidis (33, 36). Previous study has reported that the use of 474
Tween 80 could improve the permeability of rifampicin in M. intracellulare in the growth 475
medium as it is known to reduce permeability barriers in mycobacteria (36) However, 476
in all our experiments we used 0.05% of Tween-80 containing 7H9 broth. Though it is 477
likely that the degradation and assimilation of Tween-80 by the mycobacterial cells 478
over the course of growth and drug exposure (52) could affect the effective 479
concentration of the Tween-80 and thereby altered permeability, the differential OL 480
thickness between the persisters and 12 day old rifampicin unexposed control cells 481
exclude this possibility (see Fig. S1). Since the surface charge of persister cells was 482
negative, the polar and charged nature of the OL need to be considered for its effect 483
on the permeability of a nonpolar antibiotic such as rifampicin. The higher negative 484
charge (polar nature) may be expected to reduce the permeability of a more non-polar 485
type of molecule, such as 5-FAM-rifampicin. However, an integrated study using 486
Wayne’s in vitro hypoxia model at pH 5.8 has shown that while many lipophilic drugs 487
(rifampicin, rifapentine, bedaquiline, clofazimine, nitrazoxamide) could reduce cfu of 488
hypoxic cells by ≥2-log10, many hydrophilic drugs (metronidazole, moxifloxacin, 489
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

21
pyrazinamide, ethambutol, isoniazid, meropenem) could not effectively reduce the cfu 490
of hypoxic cells (53). Therefore, the polar nature of the TOL cannot be taken as the 491
sole reason for the restricted entry of non-polar (lipophilic) rifampicin. A combination 492
of increased physical thickness and negative charge may be contributing to the 493
restricted entry of rifampicin into M. tuberculosis persister cells, thereby helping 494
survival and subsequent acquisition of drug resistance. 495
496
Acknowledgements 497
PA dedicates this work as a tribute to Prof. T. Ramakrishnan (late), who led the 498
pioneering and foundation-laying work on the biochemistry and molecular biology of 499
Mycobacterium tuberculosis at Indian Institute of Science, Bangalore. 500
501
Funding 502
The work was supported by funds from the DBT-IISc partnership programme and IISc. 503
Authors acknowledge DBT-supported FACS facility in the Biological Sciences 504
Division, and the infrastructure facilities supported by DST-FIST, UGC-CAS, ICMR-505
CAS, and IISc, in the MCB Dep't. JS and SS received SRF from DBT and CSIR, 506
respectively. 507
508
Conflicts of interest statement 509
None declared. 510
511
Author Contributions 512
PA, JS conceived/designed expts; JS, SS performed expts; PA, JS, SS analysed data; 513
PA contributed reagents, materials, and analysis tools; PA, JS wrote the manuscript. 514
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

22
515
References 516
1. Jankute M, Cox JA, Harrison J, Besra GS. 2015. Assembly of the mycobacterial 517
cell wall. Annu Rev Microbiol 69:405-423. 518
2. Lederer E, Adam A, Ciorbaru R, Petit JF, Wietzerbin J. 1975. Cell walls of 519
mycobacteria and related organisms; chemistry and immunostimulant 520
properties. Mol Cell Biochem 7:87-104. 521
3. Jarlier V, Nikaido H. 1994. Mycobacterial cell wall: structure and role in natural 522
resistance to antibiotics. FEMS Microbiol Lett 123:11-18. 523
4. Cook GM, Berney M, Gebhard S, Heinemann M, Cox RA, Danilchanka O, 524
Niederweis M. 2009. Physiology of mycobacteria. Adv Microb Physiol 55:81-525
182. 526
5. Nguyen L. 2016. Antibiotic resistance mechanisms in M. tuberculosis: an 527
update. Arch Toxicol 90:1585-1604. 528
6. Larrouy-Maumus G, Marino LB, Madduri AV, Ragan TJ, Hunt DM, Bassano L, 529
Gutierrez MG, Moody DB, Pavan FR, de Carvalho LP. 2016. Cell-envelope 530
remodeling as a determinant of phenotypic antibacterial tolerance in 531
Mycobacterium tuberculosis. ACS Infect Dis 2:352-360. 532
7. Bacon J, Alderwick LJ, Allnutt JA, Gabasova E, Watson R, Hatch KA, Clark SO, 533
Jeeves RE, Marriott A, Rayner E, Tolley H, Pearson G, Hall G, Besra GS, 534
Wernisch L, Williams A, Marsh PD. 2014. Non-replicating Mycobacterium 535
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

23
tuberculosis elicits a reduced infectivity profile with corresponding modifications 536
to the cell wall and extracellular matrix. PLoS One 6:e87329. 537
8. Cunningham AF, Spreadbury CL. 1998. Mycobacterial stationary phase 538
induced by low oxygen tension: cell wall thickening and localization of the 16-539
kilodalton alpha-crystallin homolog. J Bacteriol 180:801-808. 540
9. Höner zu Bentrup K, Russell DG. 2001. Mycobacterial persistence: adaptation 541
to a changing environment. Trends Microbiol 9:597-605. 542
10. Ehrt S, Schnappinger D, Rhee KY. 2018. Metabolic principles of persistence 543
and pathogenicity in Mycobacterium tuberculosis. Nat Rev Microbiol 16:496-544
507. 545
11. Kohanski MA, DePristo MA, Collins JJ. 2010. Sublethal antibiotic treatment 546
leads to multidrug resistance via radical-induced mutagenesis. Mol Cell 37:311-547
320. 548
12. Sebastian J, Swaminath S, Nair RR, Jakkala K, Pradhan A, Ajitkumar P. 2017. 549
De Novo emergence of genetically resistant mutants of Mycobacterium 550
tuberculosis from the persistence phase cells formed against antituberculosis 551
drugs in vitro. Antimicrob Agents Chemother 61:e01343-16. 552
13. Takade A, Takeya K, Taniguchi H, Mizuguchi Y. 1983. Electron microscopic 553
observations of cell division in Mycobacterium vaccae V1. J Gen Microbiol 554
129:2315-2320. 555
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

24
14. Rastogi N, Frehel C, David HL. 1986. Triple-layered structure of mycobacterial 556
cell wall: evidence for the existence of a polysaccharide-rich outer layer in 18 557
mycobacterial species. Curr Microbiol 13:237-242. 558
15. Rosenberg M, Gutnick D, Rosenberg E. 1980. Adherence of bacteria to 559
hydrocarbons: a simple method for measuring cell-surface hydrophobicity. 560
FEMS Microbiol Lett 9:29-33. 561
16. Stokes RW, Norris-Jones R, Brooks DE, Beveridge TJ, Doxsee D, Thorson LM. 562
2004. The glycan-rich outer layer of the cell wall of Mycobacterium tuberculosis 563
acts as an antiphagocytic capsule limiting the association of the bacterium with 564
macrophages. Infect Immun 72:5676-5686. 565
17. Wilson WW, Wade MM, Holman SC, Champlin FR. 2001. Status of methods 566
for assessing bacterial cell surface charge properties based on zeta potential 567
measurements. J Microbiol Methods 43:153-164. 568
18. Ayala-Torres C, Hernández N, Galeano A, Novoa-Aponte L, Soto CY. 2014. 569
Zeta potential as a measure of the surface charge of mycobacterial cells. Ann 570
Microbiol 64:1189-1195. 571
19. Ortalo-Magne A, Dupont MA, Lemassu A, Andersen AB, Gounon P, Daffé M. 572
1995. Molecular composition of the outermost capsular material of the tubercle 573
bacillus. Microbiology 141:1609-1620. 574
20. Dickinson JM, Aber VR, Allen BW, Ellard GA, Mitchison DA. 1974. Assay of 575
rifampicin in serum. J Clin Pathol 27:457-62. 576
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

25
21. Takade A, Umeda A, Matsuoka M, Yoshida S, Nakamura M, Amako K. 2003. 577
Comparative studies of the cell structures of Mycobacterium leprae and M. 578
tuberculosis using the electron microscopy freeze-substitution technique. 579
Microbiol Immunol 47:265-270. 580
22. Vijay S, Anand D, Ajitkumar P. 2012. Unveiling unique features of formation of 581
septal partition and constriction in mycobacteria—an ultrastructural study. J 582
Bacteriol 194:702-707. 583
23. Wood PJ. 1980. Specificity in the interaction of direct dyes with 584
polysaccharides. Carb Res 85:271-287. 585
24. Rasconi S, Jobard M, Jouve L, Sime-Ngando T. 2009. Use of calcofluor white 586
for detection, identification, and quantification of phytoplanktonic fungal 587
parasites. Appl Environ Microbiol 75:2545-2553. 588
25. Argüelles JC. 2000. Physiological roles of trehalose in bacteria and yeasts: a 589
comparative analysis. Arch Microbiol 174: 217-224. 590
26. Elbein AD, Pan YT, Pastuszak I, Carroll D. 2003. New insights on trehalose: a 591
multifunctional molecule. Glycobiology 13:17R-27R. 592
27. Chen Q, Haddad GG. 2004. Role of trehalose phosphate synthase and 593
trehalose during hypoxia: from flies to mammals. J Exp Biol 207:3125-3129. 594
28. Schwebach JR, Glatman-Freedman A, Gunther-Cummins L, Dai Z, Robbins 595
JB, Schneerson R, Casadevall A. 2002. Glucan is a component of the 596
Mycobacterium tuberculosis surface that is expressed in vitro and in vivo. Infect 597
Immun 70: 2566-2575. 598
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

26
29. Lemassu, A, Daffé M. 1994. Structural features of the exocellular 599
polysaccharides of Mycobacterium tuberculosis. Biochem J 297:351-357. 600
30. Daffé M, Etienne G. 1999. The capsule of Mycobacterium tuberculosis and its 601
implications for pathogenicity. Tuber Lung Dis 79:153-169. 602
31. Llobet E, Tomás JM, Bengoechea JA. 2008. Capsule polysaccharide is a 603
bacterial decoy for antimicrobial peptides. Microbiology 154:3877-3886. 604
32. Moxon ER, Kroll JS. 1990. The role of bacterial polysaccharide capsules as 605
virulence factors. Curr Top Microbiol Immunol 150:65-85. 606
33. Abadi FJ, Carter PE, Cash P, Pennington TH. 1996. Rifampin resistance in 607
Neisseria meningitidis due to alterations in membrane permeability. Antimicrob 608
Agents Chemother 40:646-651. 609
34. Cui L, Ma X, Sato K, Okuma K, Tenover FC, Mamizuka EM, Gemmell CG, Kim 610
MN, Ploy MC, El-Solh N, Ferraz V, Hiramatsu K. 2003. Cell wall thickening is a 611
common feature of vancomycin resistance in Staphylococcus aureus. J Clin 612
Microbiol 41:5-14. 613
35. Cui L, Iwamoto A, Lian JQ, Neoh HM, Maruyama T, Horikawa Y, Hiramatsu K. 614
2006. Novel mechanism of antibiotic resistance originating in vancomycin-615
intermediate Staphylococcus aureus. Antimicrob Agents Chemother 50:428-616
438. 617
36. Hui J, Gordon N, Kajioka R. 1977. Permeability barrier to rifampin in 618
mycobacteria. Antimicrob Agents Chemother 11:773-779. 619
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

27
37. Piddock LJ, Williams KJ, Ricci V. 2000. Accumulation of rifampicin by 620
Mycobacterium aurum, Mycobacterium smegmatis and Mycobacterium 621
tuberculosis. J Antimicrob Chemother 45:159-165. 622
38. Choudhuri BS, Bhakta S, Barik R, Basu J, Kundu M, Chakrabarti P. 2002. 623
Overexpression and functional characterisation of an ABC (ATP-binding 624
cassette) transporter encoded by the genes drrA and drrB of Mycobacterium 625
tuberculosis. Biochem J 367:279-285. 626
39. Pasca MR, Guglierame P, Arcesi F, Bellinzoni M, De Rossi E, Riccardi G. 2004. 627
Rv2686c-Rv2687c-Rv2688c, an ABC fluoroquinolone efflux pump in 628
Mycobacterium tuberculosis. Antimicrob Agents Chemother 48:3175-3178. 629
40. Gupta S, Tyagi S, Almeida DV, Maiga MC, Ammerman NC, Bishai WR. 2013. 630
Acceleration of tuberculosis treatment by adjunctive therapy with verapamil as 631
an efflux inhibitor. Am J Respir Crit Care Med 188:600-607. 632
41. Demitto Fde O, do Amaral RC, Maltempe FG, Siqueira VL, Scodro RB, Lopes 633
MA, Caleffi-Ferracioli KR, Canezin PH, Cardoso RF. 2015. In vitro activity of 634
rifampicin and verapamil combination in multidrug-resistant Mycobacterium 635
tuberculosis. PLoS One 10:e0116545. 636
42. Chen C, Gardete S, Jansen RS, Shetty A, Dick T, Rhee KY, Dartois V. 2018. 637
Verapamil targets membrane energetics in Mycobacterium tuberculosis. 638
Antimicrob Agents Chemother 62:e02107-e02117. 639
43. Adams KN, Takaki K, Connolly LE, Wiedenhoft H, Winglee K, Humbert O, 640
Edelstein PH, Cosma CL, Ramakrishnan L. 2011. Drug tolerance in replicating 641
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

28
mycobacteria mediated by a macrophage-induced efflux mechanism. Cell 642
145:39-53. 643
44. Armstrong JA, Hart PD. 1971. Response of cultured macrophages to 644
Mycobacterium tuberculosis, with observations on fusion of lysosomes with 645
phagosomes. J Exp Med 134:713-740. 646
45. Saito R, Tanaka A, Sugiyama K, Azuma I, Yamamura Y. 1976. Adjuvant effect 647
of cord factor, a mycobacterial lipid. Infect Immun 13:776-781. 648
46. Ragas A, Roussel L, Puzo G, Rivière M. 2007. The Mycobacterium tuberculosis 649
cell-surface glycoprotein apa as a potential adhesin to colonize target cells via 650
the innate immune system pulmonary C-type lectin surfactant protein A. J Biol 651
Chem 282:5133-5142. 652
47. Geurtsen J, Chedammi S, Mesters J, Cot M, Driessen NN, Sambou T, Kakutani 653
R, Ummels R, Maaskant J, Takata H, Baba O, Terashima T, Bovin N, 654
Vandenbroucke-Grauls CM, Nigou J, Puzo G, Lemassu A, Daffé M, Appelmelk 655
BJ. 2009. Identification of mycobacterial alpha-glucan as a novel ligand for DC-656
SIGN: involvement of mycobacterial capsular polysaccharides in host immune 657
modulation. J Immunol 183:5221-5231. 658
48. Domenico P, Salo RJ, Cross AS, Cunha BA. 1994. Polysaccharide capsule-659
mediated resistance to opsonophagocytosis in Klebsiella pneumoniae. Infect 660
Immun 62:4495-4499. 661
49. Costerton JW, Cheng KJ. 1975. The role of the bacterial cell envelope in 662
antibiotic resistance. J Antimicrob Chemother 1:363-377. 663
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

29
50. Slack MP, Nichols WW. 1982. Antibiotic penetration through bacterial capsules 664
and exopolysaccharides. J Antimicrob Chemother 10:368-372. 665
51. Campos MA, Vargas MA, Regueiro V, Llompart CM, Alberti S, Bengoechea JA. 666
2004. Capsule polysaccharide mediates bacterial resistance to antimicrobial 667
peptides. Infect Immun 72:7107-7114. 668
52. Schaefer WB, Lewis CW Jr. 1965. Effect of oleic acid on growth and cell 669
structure of mycobacteria. J Bacteriol 90:1438-1447. 670
53. Piccaro G, Poce G, Biava M, Giannoni F, Fattorini L. 2015. Activity of lipophilic 671
and hydrophilic drugs against dormant and replicating Mycobacterium 672
tuberculosis. J Antibiot (Tokyo) 68:711-714. 673
674
Figure Legends 675
FIG 1. Presence of TOL in rifampicin persister cells. Transmission electron micrograph 676
of M. tuberculosis cells showing differential thickening of OL in persister cell (A) and 677
in control MLP cells (B). Magnified images are shown right side, the yellow arrows 678
indicate OL. Thickness (in nm) of OL (C) and ETL (D) between persister and MLP 679
cells. (E) CFW staining of OL in persister cells before and after bead beating (BB). 680
Scale, 2 µm. Density plots of the flow cytometry analysed CFW stained MLP cells (F) 681
and persister cells (G) before and after BB. AF, autofluorescence. Histogram overlay 682
of the flow cytometry analysed CFW stained MLP cells (H) and persister cells (I) before 683
and after BB. (J) Bar graph showing normalised median fluorescence intensity of CFW 684
in MLP and persister cells before and after BB. One asterisk (*) indicates P value less 685
than or equal to 0.05 (P ≤ 0.05). Three asterisks (***) indicate P value lesser than 686
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

30
0.001 (P < 0.001). The statistical significance was calculated using two-tailed paired 687
t-test. 688
689
FIG 2. Physicochemical properties of rifampicin-exposed M. tuberculosis persister 690
cells. (A) Hexadecane assay showing the proportions of hydrophilic rifampicin 691
persister cells and MLP cells. (B) ZP of cells from different phases of rifampicin 692
exposed of M. tuberculosis cells and its corresponding (C) histogram overlay. ZP of 693
TOL as a function of pH showing the isoelectric point of the TOL between pH 4.0 and 694
pH 5.0 for: (D) MLP cells, (E) persister cells, and (F) cells from killing phase. One 695
asterisk (*) indicates P value less than or equal to 0.05 (P ≤ 0.05). Two asterisks (**) 696
indicate P value less than or equal to 0.01 (P ≤ 0.01). The statistical significance was 697
calculated using two-tailed paired t-test. 698
699
FIG. 3. Characterisation of 5-FAM-rifampicin permeability into MLP and rifampicin 700
persister cells. (A) The possible structural isomers of 5-FAM-rifampicin conjugates. 701
The carboxyl group of the 5-FAM can form an ester bond with any one of the hydroxyl 702
groups in the aliphatic ring of rifampicin, generating two possible isomers. Ester bond 703
is shown in red circle. (B) Bar graph showing the relative bactericidal activity of 704
rifampicin and 5-FAM-rifampicin using Staphylococcus aureus by agar diffusion assay. 705
(C) Density plots and (E) histogram overlay of MLP cells incubated with increasing 706
concentrations of 5-FAM-rifampicin by flow cytometry. (E) Standard graph showing the 707
5-FAM-rifampicin permeability into MLP cells at two-fold increasing concentrations. 708
709
FIG 4. Permeability of M. tuberculosis to 5-FAM-rifampicin. Fluorescence microscopy 710
images of M. tuberculosis rifampicin persister cells showing differential entry of 5-FAM-711
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

31
rifampicn in persisters (A) and MLP cells (B) before and after the removal of OL by 712
bead beating (BB). (Scale, 2 µm). Line graph from flow cytometry analysis for the time-713
dependent entry of 5-FAM-rifampicin into MLP (C) and rifampicin persisters (D) with 714
or without bead beating (BB) and its corresponding density plots (E) and histogram 715
overlay (F). (AF, Autofluorescence). 716
717
FIG 5. Estimation of the relative permeability of 5-FAM-rifampicin into rifampicin 718
persisters in presence of efflux pump inhibitor verapamil. (A) Density plots of 5-FAM-719
rifampicin entry in M. tuberculosis cells with or without verapamil exposure. (B) Bar 720
graph showing relative quantitation of the 5-FAM-rifampicin entry between MLP cells 721
and rifampicin persisters with and without verapamil (VM) treatment. (C) Histogram 722
overlay of the flow cytometry profile. 723
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

Table 1 Relative abundance of TOLa compounds in rifampicin persister cells with
respect to MLPb cells as detected by GC-MS
___________________________________________________________________
Compound Fold increase in Persister cells
(w.r.t. MLP cells)
___________________________________________________________________
α-D-Glucopyranoside 6.67
1, 2, 5-linked-Mannitol 4.97
3, 4-linked Mannose 2.22
Hexa-acetyl-Mannitol 1.84
1, 2, 3-Propanetriol 1.83
Methyl 1, 2, 3, 4-Tetrahydronaphthalene-2-Carboxylate 1.76
1, 3-Di-Iso-Propylnaphthalene 1.73
1, 2, 4-linked arabinitol 1.68
D- (1, 2-linked mannitol) 1.58
1, 3-Di-Iso-Propylnaphthalene 1.41
5-linked Galactonitrile 1.30
Galactose pyranoside 1.11
1, 7-Di-Iso-Propylnaphthalene 1.10
α- D-Mannopyranoside 1.08
___________________________________________________________________
aThick Outer Layer; bMid-log phase
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

FIG 1. Presence of TOL in rifampicin persister cells. Transmission electron micrograph of M.
tuberculosis cells showing differential thickening of OL in persister cell (A) and in control MLP cell
(B). Magnified images are shown right side, the yellow arrows indicate OL. Thickness (in nm) of
OL (C) and ETL (D) between persister and MLP cells. (E) CFW staining of OL in persister cells
before and after bead beating (BB). Scale, 2 µm. Density plots of the flow cytometry analysed
CFW stained MLP cells (F) and persister cells (G) before and after BB. AF, autofluorescence.
Histogram overlay of the flow cytometry analysed CFW stained MLP cells (H) and persister cells
(I) before and after BB. (J) Bar graph showing normalised median fluorescence intensity of CFW
in MLP and persister cells before and after BB. One asterisk (*) indicates P value less than or
equal to 0.05 (P ≤ 0.05). Three asterisks (***) indicate P value lesser than 0.001 (P < 0.001). The
statistical significance was calculated using two-tailed paired t-test.
0
50
100
150C
*
OL
Th
ickn
ess (
nm
)
Persisters Control
0
50
100
150
D
ET
L T
hic
kn
ess (
nm
)
Persisters Control
MLP AF MLP + CW
No
BB
BB
FPer AF Per + CW
G
H No BB BB
ML
P
Pers
iste
r
I No BB BB
Pers
iste
rC
on
tro
lA
B
JE
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

MLP
Killing
Per
RG
CBA
2 4 6 8 10
-30
-20
-10
0
10
pH
Zeta
Pote
nti
al
(mV
)
ED
2 4 6 8 10
-3
-2
-1
0
1
pH
Zeta
Pote
nti
al
(mV
)
F
2 4 6 8 10
-15
-10
-5
0
5
pH
Zeta
Po
ten
tial (m
V)
FIG 2. Physicochemical properties of rifampicin-exposed M. tuberculosis persister cells. (A)
Hexadecane assay showing the proportions of hydrophilic rifampicin persister cells and
MLP cells. (B) ZP of cells from different phases of rifampicin exposed of M. tuberculosis
cells and its corresponding histogram overlay (C). ZP of TOL as a function of pH showing
the isoelectric point of the TOL between pH 4.0 and pH 5.0 for: (D) MLP cells, (E) persister
cells, and (F) cells from killing phase. One asterisk (*) indicates P value less than or equal
to 0.05 (P ≤ 0.05). Two asterisks (**) indicate P value less than or equal to 0.01 (P ≤ 0.01).
The statistical significance was calculated using two-tailed paired t-test.
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

A
O
O
O
N
O
O
OH
O
O
OH OHCH3
OH
NN
N
O
O
O
CH3
NH
OCH3
CH3CH3
OH
OCH3
O
O
CH3
CH3
H
CH3
CH3
O
5-FAM-RIF Isomer 1
OH OHCH3
OH
NN
N
O
O
O
CH3
NH
OCH3
CH3CH3
O
OCH3
O
O
CH3
CH3
H
CH3
CH3
OH
O
O
O N
O
O
OH
O
O
5-FAM-RIF Isomer 2
0.0 0.5 1.0 1.5 2.0 2.50
500
1000
1500
2000
2500
Conc. of 5-FAM-RIF (g/ml)
5-F
AM
-RIF
(N
or.
A.U
)
E
B
D
Rifampicin 5-FAM-rifampicin0
5
10
60
80
100
% B
acteric
ida
l a
ctiv
ity
0.0156 µg/ml 0.03125 µg/ml 0.0625 µg/ml 0.125 µg/ml
0.25 µg/ml 0.5 µg/ml 1.0 µg/ml 2.0 µg/ml
Mid-log
C
FIG 3. Characterisation of 5-FAM-rifampicin permeability into MLP and rifampicin persister
cells. (A) The possible structural isomers of 5-FAM-rifampicin conjugates. The carboxyl group
of the 5-FAM can form an ester bond with any one of the hydroxyl groups in the aliphatic ring
of rifampicin, generating two possible isomers. Ester bond is shown in red circle. (B) Bar
graph showing the relative bactericidal activity of rifampicin and 5-FAM-rifampicin using
Staphylococcus aureus by agar diffusion assay. (C) Density plots and (E) histogram overlay of
MLP cells incubated with increasing concentrations of 5-FAM-rifampicin by flow cytometry. (E)
Standard graph showing the 5-FAM-rifampicin permeability into MLP cells at two-fold
increasing concentrations.
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

C
0 15 30 45 600
100
200
300 MLP
MLP-BB
Time (min)
5-F
AM
Med
ian
Flu
o.
(Nor.
) D
15 30 45 60
-100
0
100
200
300Per
Per-BB
Time (min)
5-F
AM
Med
ian
Flu
o. (N
or.)
MLP BBMLP
0 min
15 min
30 min
45 min
60 min
▬▬
▬
▬
▬
0 min
15 min
30 min
45 min
60 min
▬▬
▬
▬
▬
G Per Per BB
0 min
15 min
30 min
45 min
60 min
▬▬
▬
▬
▬
0 min
15 min
30 min
45 min
60 min
▬
▬
▬
▬
▬
H
Per AF Per+ 5-FAM-RIFFMLP AF MLP+ 5-FAM-RIF
No
BB
BB
E
DIC 5-FAM-RIF MERGEDP
ersi
ster
No B
B
ADIC 5-FAM-RIF MERGED
B
DIC 5-FAM-RIF MERGEDDIC 5-FAM-RIF MERGED
Per
sist
er
BB
ML
P
No B
B
ML
P
BB
FIG 4. Permeability of M. tuberculosis to 5-FAM-rifampicin. Fluorescence microscopy images
of M. tuberculosis rifampicin persister cells showing differential entry of 5-FAM-rifampicn in
persisters (A) and MLP cells (B) before and after the removal of OL by bead beating (BB).
(Scale, 2 µm). Line graph from flow cytometry analysis for the time-dependent entry of 5-
FAM-rifampicin into MLP (C) and rifampicin persisters (D) with or without bead beating (BB)
and its corresponding density plots (E) and histogram overlay (F). (AF, Autofluorescence).
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint

5-FAM-RIF
Pers
iste
r
Control
ML
PP
ers
iste
r+V
era
pam
ilA B
MLP Per Per+ VM0.0
0.5
1.0
1.5
Rel.
5-F
AM
-Con
c. (
g/m
l)
C
FIG 5. Estimation of the relative permeability of 5-FAM-rifampicin into rifampicin persisters in
presence of efflux pump inhibitor verapamil. (A) Density plots of 5-FAM-rifampicin entry in M.
tuberculosis cells with or without verapamil exposure. (B) Bar graph showing relative
quantitation of the 5-FAM-rifampicin entry between MLP cells and rifampicin persisters with
and without verapamil (VM) treatment. (C) Histogram overlay of the flow cytometry profile.
.CC-BY-NC-ND 4.0 International licenseunder acertified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available
The copyright holder for this preprint (which was notthis version posted May 1, 2019. ; https://doi.org/10.1101/624569doi: bioRxiv preprint