Embed Size (px)
Citation preview

Faculty of Veterinary Sciences Department of Veterinary Public Health & Food Safety
Laboratory for Hygiene and Technology
Antibiotic susceptibility in pig-
associated Escherichia coli and
Salmonella in Belgium:
extended spectrum β-lactamase
producers and colistin susceptibility
Jonathan Woollard
Student number: 01410461
Promoter: Prof. Dr. Kurt Houf
Scientific supervisor: Drs. Wauter Biasino
Master’s dissertation submitted to Ghent University to obtain the degree of Master of Science in
Biochemistry and Biotechnology. Major Microbial Biotechnology
Academic year: 2016 - 2017

I
Preface
With this dissertation an important chapter in my life comes to a close. I would not have reached this far in my life without the constant support of my parents, who have always put their children before themselves. For this I am forever grateful. I would also like to thank all my friends for being there for me when I needed advice or simply a break from writing. In particular I would like to thank all those who proofread this thesis.
I would like to thank Prof. Kurt Houf and Drs. Wauter Biasino for having offered me the chance to persue this thesis. Especially Wauter who also guided me throughout my entire thesis. Additionally, I would like to thank all the members of the lab for lending me their expertise.
Another special thanks goes to Dr. Inge Van Damme for letting me consult her whenever I had doubts or needed help.
Finally, I would like to thank all my fellow students who were also conducting research their thesis in the lab. You guys made my time spend on campus a truly fantastic experience!

II
Table of contents
Preface .................................................................................................. I
Table of contents ................................................................................. II
List of Figures ....................................................................................... V
List of Tables ....................................................................................... VI
List of Abbreviations .......................................................................... VII
Nederlandstalig Abstract ..................................................................... IX
English Abstract ................................................................................... X
1. Introduction ..................................................................................... 1
1.1. Antibiotic resistance ................................................................................... 1
1.1.1. History and spread. ........................................................................... 1
1.1.2. Antibiotic resistance crisis ................................................................. 4
1.1.2.1. Lack of new antibiotics .......................................................... 4
1.1.2.2. Overuse and misuse .............................................................. 4
1.1.3. Extended-spectrum β-lactamases and AmpC β-lactamases ................ 6
1.1.4. Colistin resistance ............................................................................. 9
1.2. Pigs and their importance to antibiotic resistance ..................................... 11
1.2.1. Salmonella as a foodborne pathogen ............................................... 11
1.2.2. E. coli as hygiene indicator and potential reservoir for resistance ..... 13
1.3. Methods for detection of antibiotic resistance .......................................... 14
1.4. Course of action against antibiotic resistance ............................................ 15
2. Aim of Research Project ................................................................. 17
3. Results ........................................................................................... 19
3.1. Study A - ESBL/AmpC characterisation of the isolates .............................. 19
3.1.1. Phenotyping of the isolates by means of disk diffusion ................... 19
3.1.2. Genetic profiling of the isolates for ESBL/AmpC genes .................... 19

III
3.2. Study A - Colistin susceptibility of ESBL/AmpC-producing E. coli and jjjjjjjjjjjjjSalmonella………………………………………………………………………………………....……22
3.2.1. Colistin susceptibly according to disk prediffusion ........................... 22
3.2.2. Genetic profiling of the isolates for plasmid-mediated jjjjjjjjjjjjjjcolistin resistance genes .................................................................. 22
3.3. Study A - Phenotypical testing by means of microbroth dilution ................ 23
3.4. Study A - Comparison of disk (pre)diffusion and microbroth dilution ......... 27
3.5. Study B - Occurrence of ESBL genes in different pig batches ...................... 29
4. Discussion ...................................................................................... 31
4.1. ESBL/AmpC characterisation of the isolates ............................................. 31
4.2. Study A - Colistin susceptibility of ESBL/AmpC-producing E. coli and jjjjjjjjjjjjjSalmonella ............................................................................................... 32
4.3. Study A - Phenotypical testing by means of microbroth dilution ............... 33
4.4. Study A - Comparison of disk (pre)diffusion and microbroth dilution ........ 34
4.5. Future prospects ...................................................................................... 35
5. Materials and Methods .................................................................. 37
5.1. Isolation of ESBL/AmpC-producing E. coli and Salmonella ......................... 37
5.2. Setting up antibiogram for ESBL/AmpC-producing E. coli and Salmonella . 38
5.2.1. Disk diffusion for ESBL/AmpC-producing E. coli and Salmonella…......38
5.2.2. Disk prediffusion for testing colistin susceptibility of ESBL/AmpC-jjjjjjjjjjjjjjproducing E. coli and Salmonella .................................................... 39
5.2.3. Microbroth dilution for ESBL/AmpC-producing E. coli and Salmonella …………………………………………………………………………………………………………………39
5.3. Detection of resistance genes .................................................................. 40
5.3.1. DNA extraction of ESBL/AmpC-producing E. coli and Salmonella ...... 40
5.3.2. PCR for the detection of plasmid-mediated resistance genes ........... 40
5.3.3. Gel electrophoresis for the detection of resistance genes ................ 41
5.4. Distribution of isolates ............................................................................. 41
5.4.1. Investigation of ESBL/AmpC-producing E. coli and Salmonella.......... 41
5.4.2. Investigation of the occurrence of ESBL genes in pig-associated jjjjjjjjjjjjjjESBL E. coli ...................................................................................... 43

IV
6. References ..................................................................................... 45
7. Addenda ......................................................................................... 54
7.1. Protocols ................................................................................................ 54
7.1.1. Isolation of ESBL/AmpC-producing E. coli ........................................ 54
7.1.2. Isolation of Salmonella .................................................................... 54
7.1.3. Disk diffusion for ESBL/AmpC-producing E. coli and Salmonella……… 54
7.1.4. Disk prediffusion for testing colistin susceptibility of ESBL/AmpC-jjjjjjjjjjjjjjproducing E. coli and Salmonella .................................................... 57
7.1.5. Microbroth dilution for ESBL/AmpC-producing E. coli and jjjjjjjjjjjjjjSalmonella ...................................................................................... 59
7.1.6. Detection of resistance genes .......................................................... 60
7.1.6.1. DNA extraction of ESBL/AmpC-producing E. coli ……………………….. and Salmonella .................................................................. 60
7.1.6.2. PCR for the detection of resistance genes ............................ 61
7.1.3.3. Gel electrophoresis for the detection of resistance genes .... 67
7.2. Composition of media .............................................................................. 68
7.2.1. XLD agar .......................................................................................... 68
7.2.2. TSB .................................................................................................. 68
7.2.3. PCA ................................................................................................. 69
7.2.4. TBX agar .......................................................................................... 69
7.2.5. MH II agar ....................................................................................... 69
7.2.6. MH II broth...................................................................................... 69
7.2. Preparation of media ................................................................................ 70
7.2.1. XLD agar (for 1L) .............................................................................. 70
7.2.2. TSB (for 1L) ...................................................................................... 70
7.2.3. PCA (for 1L) ..................................................................................... 70
7.2.4. TBX agar (for 1L) .............................................................................. 71
7.2.5. MH II agar (for 1L) ........................................................................... 72
7.2.6. MH II broth (for 1L) .......................................................................... 72

V
List of Figures Figure 1: Timeline of key events in antibiotic usage and resistance ............................................................. 1
Figure 2: Transfer of DNA between bacterial cells ........................................................................................ 2
Figure 3: Example of antibiotic resistant bacterial reservoirs and spread within a community ................... 3
Figurej4: Prescribed antimicrobial agents for humans and animals compared with the number of pigs
produced, Denmark (1994 – 2013) ............................................................................................................... 5
Figure 5: Basic structure of cephalosporins, carbapenems and cephamycin ............................................... 7
Figure 6: Chemical structure of colistin ....................................................................................................... 10
Figure 7 : Structure of Gram-negative bacteria .......................................................................................... 12
Figure 8: Summary of Study A and Study B ................................................................................................. 18
Figure 9: Resulting agarose gel of multiplex I PCR to determine the presence of ESBL genes in isolates .. 20
Figurej10: Resulting agarose gel of plasmid-mediated colistin resistance gene PCR to determine the
presence of plasmid-mediated colistin resistance genes in isolates .......................................................... 22
Figure 11: Distribution of ESBL E. coli and Salmonella isolates/batches as per slaughterhouse ................ 42
Figure 12: Distribution of the isolates/batches of the two Salmonella groups as per slaughterhouse ...... 42
Figurej13: General overview of the distribution of the three groups of isolates/batches as per
slaughterhouse ............................................................................................................................................ 43
Figure 14: Distribution of ESBL E. coli isolates taken from slaughterhouse B and slaughterhouse C ......... 43
Figure 15: Batch distribution of ESBL E. coli isolates ................................................................................... 44
Figure 16: Template for placing antibiotic disks ......................................................................................... 55
Figure 17: Template for placing colistin disk ............................................................................................... 57
Figure 18: Plate design microbroth dilution ................................................................................................ 60

VI
List of Tables
Table 1: Reasons for antibiotic use in food-producing animals individually or at group level and the most
frequently applied administration routes. .................................................................................................... 6
Table 2: Categorisation of the isolates according to their phenotype ........................................................ 19
Table 3: Genotypes of the isolates belonging to the different groups ....................................................... 21
Table 4: Resistance gene pattern of the different isolates ......................................................................... 21
Table 5: Number of isolates with reduced susceptibility to colistin ........................................................... 22
Table 6: Result of mcr-1/2 multiplex ........................................................................................................... 23
Table 7: Number of isolates demonstrating reduced susceptibility ........................................................... 24
Table 8: Patterns of reduced susceptibility ................................................................................................. 25
Table 9: Patterns of reduced susceptibility (continued) ............................................................................. 26
Table 10: Difference in antibiogram between disk diffusion and microbroth dilution............................... 27
Table 11: Difference between disk diffusion and microbroth dilution for CAZ .......................................... 27
Table 12: Difference between disk diffusion and microbroth dilution for FEP ........................................... 28
Table 13: Difference between disk diffusion and microbroth dilution for CTX .......................................... 28
Table 14: Difference between disk prediffusion and microbroth dilution for COL ..................................... 28
Table 15: Occurrence of ESBL genes in different pig batches at slaughter ................................................. 29
Table 16: ESBL resistance gene pattern of the ESBL E. coli isolates ............................................................ 30
Table 17: Differentiation between ESBL and AmpC phenotype ................................................................. 38
Table 18: Multiplex I PCR ............................................................................................................................. 62
Table 19: Multiplex II PCR ............................................................................................................................ 63
Table 20: Multiplex III PCR ........................................................................................................................... 64
Table 21: Simplex I PCR ............................................................................................................................... 65
Table 22: Colistin Multiplex PCR .................................................................................................................. 66
Table 23: PCR program for ESBL/AmpC gene amplification ........................................................................ 66
Table 24: PCR program for colistin plasmid-mediated gene amplification ................................................. 67

VII
List of Abbreviations
Abbreviation Description
AMI Amikacin AmpC AmpC β-lactamases A/S2 Ampicillin/sulbactam 2:1 ratio aw water activity AZT Aztreonam bp Basepair BPW Buffered Peptone Water CAZ Ceftazidime CAZ + C Ceftazidime + Clavulanic acid CFO Cefoxitine CIP Ciprofloxacin CLSI Clinical Laboratories Standards Institute cm centimeter CMY Cephamycinase COL Colistin CTX Cefotaxime CTX+C Cefotaxime + Clavulanic acid CTX-M Cefotaximase-Munich DOR Doripenem DOX Doxycycline ECOFFs Epidemiological Cut-Off Values E. coli e.g.
Escherichia coli Exemplī grātiā
ESBL Extended-spectrum β-lactamases EU European union EUCAST European Committee on Antimicrobial Susceptibility Testing FEP Cefepime FEP+C Cefepime + Clavulanic acid g g
gram gravitational acceleration
GEN Gentamicin h hour IMI Imipenem LEVO Levofloxacin MERO Meropenem MDR Multidrug Resistant mg milligram MH II Mueller Hinton II MIC Minimum Inhibitory Concentration

VIII
MIN Minocycline min minutes mL milliliter mM millimolar mm millimeter MSRV Modified Semi Solid Rappaport Vassiliadis medium NA Not applicable OXA Oxacillin-hydrolyzing PCA Plate Count Agar PCR Polymerase Chain Reaction pH Potential of Hydrogen pmol picomol POL Polymixin B P/T4 Piperacillin / Tazobactam constant 4 rpm revolutions per minute S. Salmonella sec seconds SHV Sulfhydryl Variable SXT Trimethoprim/Sulfamethoxazole TBE Tris-Borate-EDTA TBX Tryptone Bile Agar with X-Glucuronide TEM Temoniera TGC Tigecycline TIM2 Ticarcillin / clavulanic acid constant 2 TOB Tobramycin TSB Trypticase Soy Broth USA United States of America w:v weight by volume w:w weight by weight XLD Xylose Lysine Deoxycholate µg microgram µl microliter µM micromolar β Beta

IX
Nederlandstalig Abstract De toename van antibiotica resistente (AR) bacteriën is een wereldwijd, ernstig probleem. Het zou de globale economie tegen 2050 naar schatting 100 triljoen Amerikaanse dollars kosten en verantwoordelijk zou zijn voor het overlijden van 10 miljoen mensen per jaar wereldwijd. Op boerderijen kunnen AR-bacteriën zich ontwikkelen en verspreiden tussen dieren. Deze transmissie kan gebeuren via dierlijk vlees en/of groenten, alsook bijv. via meststoffen of water dat dierlijke uitwerpselen bevat. Een welbekend antimicrobieel resistentie mechanisme bestaat uit bacteriële stammen die breedspectrum beta (β)-lactamasen (ESBLn) en/of AmpC β-lactamasen (AmpC) produceren. ESBLn, gewoonlijk gedefinieerd als plamide-geëncodeerde enzymen, zijn terug te vinden in bijv. Enterobacteriaceae, die weerstand bieden aan een reeks ß-
lactam antibiotica maar zelden tegen carbapenems of cefamycines. Daar tegenover zijn AmpC β-lactamasen intrinsieke cefalosporinasen geëncodeëerd in het chromosomaal DNA van vele Gram-negatieve bacteriën, maar eveneens vastgesteld op plasmides. β-lactams vormen vaak de eerste-lijn therapie voor een brede waaier aan infecties die voorkomen in gemeenschappen en ziekenhuizen. Het gebruik van β-lactamasen beperkt zich niet tot de mens, integendeel, in de veeteelt is het gebruik hiervan zelfs groter dan in de menselijke geneeskunde. In 2015, waren in de veeteelt, de penicillines de meest gebruikte groep van antibiotica in België, goed voor 31% van het totale gebruik aan antibiotica. Er bestaan verschillende ESBL resistentiegenen waarvan de meest predominante gen-families TEM, SHV, OXA en CTX-M zijn. Voor AmpC werden reeds meer dan 15 verschillende plasmide-geëncodeerde enzymen (bijv. CMY, FOX) gerapporteerd, met CMY-2 als het meest voorkomende enzym. Plasmide-gemedieerde AmpC β-lactamases worden minder vaak aangetroffen dan ESBLs. Recentelijk is uit onderzoek gebleken dat plasmide-gemedieerde
antibiotica resistent zijn tegen colistine (COL), een antibioticum dat als laatste toevlucht wordt gebruikt. Door de opkomst van multidrug-resistente bacteriën, zoals ESBL-producerende bacteriële stammen, is er een toename van COL gebruik in de menselijke geneeskunde. Anderzijds is het gebruik van COL in de dierengeneeskunde reeds wijd verspreid. Volgens veldstudies wordt COL voornamelijk gebruikt bij varkens als eerste-keuze behandeling en preventie van neonatale diarree veroorzaakt door Escherichia coli (E. coli) en Salmonella bij biggen. Varkensvlees is het
meest geproduceerde vlees in de EU, met een geschatte productie van 23.0 miljoen ton in 2015. In België is de varkens veeteelt de tweede grootste gebruiker van antibiotica. Het doel van deze studie is dan ook om cefotaxime-resistente E. coli isolaten en Salmonella isolaten afkomstig van slachthuisvarkens te karakteriseren op de aanwezigheid van ESBL/AmpC productie en verminderde gevoeligheid voor COL. Het testen van deze gevoeligheid gebeurt door disk
(pre)diffusie en microbroth dilutie. De meeste E. coli isolaten (74%) vertoonden een ESBL phenotype, slechts een beperkt aantal vertoonden een AmpC phenotype (7%) en enkele vertoonden een gecombineerd ESBL en AmpC phenotype (16%). Anderzijds toonden geen van de Salmonella isolaten een ESBL/AmpC phenotype. Bovendien vertoonde geen van de gescreende isolaten een plasmide-gemedieerde COL resistentie gen (mcr-1/2). De conclusie van deze studie is dat er momenteel geen substantieel gevaar voor ESBL Salmonella bestaat bij Belgische varkens, hoewel er een reservoir van antibiotica resistentiegenen aanwezig is E. coli gevonden bij deze varkens.

X
English Abstract
Antibiotic resistant bacteria are roughly predicted to be the cause of death of ten million people globally each year and to cost the global economy 100 trillion US dollars by 2050. Therefore, the rising global antibiotic resistance is a severe problem. Food producing animals represent an important reservoir of resistant strains. In farms, drug-resistant bacteria can develop and spread between animals. These resistant bacteria may be transmitted from animals to humans via the consumption of contaminated meat or vegetables that are contaminated using fertilizer or water containing animal feces. A well-established antimicrobial resistance mechanism is that of bacterial strains producing extended-spectrum beta (β)-lactamases (ESBLs) and/or AmpC β-lactamases (AmpC). ESBLs are commonly defined as plasmid-encoded enzymes found in e.g.
Enterobacteriaceae conferring resistance to a range of ß-lactam antibiotics, but rarely against carbapenems or cephamycins. In contrast, AmpC β-lactamases are intrinsic cephalosporinases encoded in the chromosomal DNA of many Gram-negative bacteria but also observed on plasmids. β-lactams are often administered as a first-line therapy for a wide variety of infections occurring in communities and hospitals. The use of β-lactams is not only limited to humans, on the contrary, its use in animal husbandry is greater than in human medicine. In 2015, the most used group of antibiotics in animal husbandry in Belgium were the penicillins, accounting for 31% of the total use of antibiotics. A variety of ESBL resistance genes exist with the most predominate gene families being : TEM, SHV, OXA and CTX-M. As for AmpC, over 15 different plasmid-encoded enzymes (e.g. CMY, FOX) have been reported, with CMY-2 being the most commonly found enzyme. However, plasmid-mediated AmpC β-lactamases are less commonly found than ESBLs.
Recently, research has uncovered plasmid-mediated antibiotic resistance against colistin, a last resort antibiotic. Because of the emergence of multidrug resistant bacteria, such as ESBL-producing bacterial strains, usage of colistin in human medicine is increasing. However, colistin is already used widely in veterinary medicine. According to field studies, colistin use is primarily observed in pigs as first-choice treatment and prevention of neonatal diarrhoea caused by Escherichia coli (E. coli) and Salmonella in piglets. The primary meat product in the European
Union is pork, with an estimated production of 23.0 million tonnes in 2015. In Belgium, the second highest antimicrobial-consuming livestock production system is fattening pigs. Therefore, the aim of this study is to characterise cefotaxime resistant E. coli isolates and Salmonella isolates, originating from slaughterhouse pigs for the presence of ESBL/AmpC production and reduced susceptibility to colistin. Susceptibility was tested using disk (pre)diffusion and microbroth
dilution. Most of the E. coli isolates (74%) displayed an ESBL phenotype, only a limited number displayed an AmpC phenotype (7%) and few displayed a combined ESBL and AmpC phenotype (16%). On the other hand, none of the Salmonella isolates showed an ESBL/AmpC phenotype. Furthermore, none of the screened isolates were shown to carry a plasmid-mediated colistin resistance gene (mcr-1/2). This study concludes that there is no substantial threat regarding ESBL Salmonella in Belgian pork. However, a reservoir of antibiotic resistance genes remains within E. coli found in Belgian pigs.

1. Introduction
1
Figure 1: Timeline of key events in antibiotic usage and resistance
Dates are based upon early reports of resistance in the literature. In
the case of pan drug-resistant (PDR)- Acinetobacter and Pseudomonas,
the date is based upon reports of healthcare transmission or
outbreaks.
Time line showing introduction dates of antibiotics and the first
identified resistance against it.
Note: penicillin was in limited use prior to widespread population
usage in 1943
Figure from CDC. Antibiotic resistance threats in the United States,
2013. Current 114 (2013)
1. Introduction 1.1 Antibiotic resistance
1.1.1 History and spread
Antibiotics were discovered in the mid-20th century and are a cornerstone in today’s modern medicine, since complex surgeries such as organ transplantation and chemotherapy for cancer treatment rely on the availability of anti-infective drugs (World Health Organization, 2014; CDC, 2013; O’Neill, 2014). Therefore, the rising global antibiotic resistance is a severe problem. The complete prevention of development and spread of antibiotic resistance is impossible, as it also occurs as part of natural bacterial evolution (World Health Organization, 2014; CDC, 2013;
Arepyeva et al, 2017). However the usage of antibiotics puts a selective pressure on bacterial
populations, resulting in the selection and spread of resistant bacteria (CDC, 2013; Davies & Davies, 2010). As shown in figure 1, antibiotic resistance ensues with the use of antibiotics. There are two different ways in which bacteria acquire their resistance, either through spontaneous genetic mutation or through horizontal transfer of antibiotic resistance genes between bacterial strains which can be either distantly or closely related (CDC, 2013;
Frost et al, 2005; Davies & Davies, 2010; Read & Woods, 2014).

1. Introduction
2
To illustrate the different manners of horizontal transfer, in figure 2 the three ways of transfer of
DNA between a donor cell and recipient cell can be seen. Transduction is the first manner and consists of a phage taking up the ‘yellow’ marked host DNA and depositing it into the recipient cell to be integrated later on. The second manner is conjugation where the two cells are physically linked using pili for the transfer of plasmid(s) that may contain antibiotic resistance genes. Finally, the third manner is via transposons (pink) which integrate into new sites on the chromosome or plasmids by non-homologous recombination. Integrons (dark green) use similar mechanisms to exchange single gene cassettes (brown) (Frost et al, 2005).
Figure 2: Transfer of DNA between bacterial cells
Figure illustrating the different manners of horizontal DNA transfer between bacterial cells.
Source: (Frost et al, 2005)

1. Introduction
3
Once resistant populations are observed, they are hard to contain or eliminate, especially if the
antibiotic selection pressure remains. Common places for resistant populations are farms and hospitals due to the frequent use of antibiotics. (European Food Safety Authority & European Centre for Disease Prevention and Control, 2016; Mesa et al, 2006; Nordmann et al, 2009; Pitout et al, 2005). In farms, drug-resistant bacteria can develop and spread between animals. Transmission routes to humans include animal meats and vegetables and the use of e.g. fertilizer or water containing animal feces, through improper handling or cooking of these resulting products (CDC, 2013; World Health Organization, 2014; Leverstein-van Hall et al, 2011). Additionally, these individuals further spread drug-resistant bacteria within their community. Direct reservoir-to-human transmission is also possible through farmers, hospital staff, patients, delivery workers and visitors acting as potential carriers. Finally, through antibiotic use, humans can develop and spread drug-resistant bacteria (World Health Organization, 2014; CDC, 2013;
Davies & Davies, 2010). A simplified scheme can be seen in figure 3 and shows how antibiotic resistance spread is integrated into our society.
Figure 3: Example of antibiotic resistant bacterial reservoirs and spread within a community
A scheme illustrating how bacteria that have acquired resistance to antibiotics can spread and
persevere within a community.
Figure from CDC. Antibiotic resistance threats in the United States, 2013. Current 114 (2013)

1. Introduction
4
1.1.2 Antibiotic resistance crisis
1.1.2.1 Lack of new antibiotics
Antibiotic resistance is an increasing problem partly due to the fact that the pace at which novel antibiotics are being discovered or developed has slowed drastically, while antibiotic usage is rising. To illustrate this, over the past 30 years, mainly two classes of novel antibiotics have been on the market: the oxazolidinones (linezolid) and cyclic lipopeptides (daptomycin). In the meantime, resistance has been documented for both of these compounds (World Health Organization, 2014; Coates et al, 2011; Ventola, 2015a). Besides the obvious impact on human and animal health, there is also a large global economic factor (O’Neill, 2014). When first-line and second-line antibiotics fail, the use of more toxic and expensive antibiotics is required. Moreover,
failure of first/second-line antibiotics results in a prolonged hospital stay of patients with resistant infections (on average rising by 6.4 days up to 12.7 days), together with more doctors’ visits, and lengthier recuperations (CDC, 2013; Golkar et al, 2014). Antibiotic resistant bacteria are roughly predicted to be the cause of death of 10 million people globally each year and cost the global economy 100 trillion US dollars by 2050 (O’Neill, 2014).
At the moment pharmaceutical companies are neglecting the discovery or development of novel antibiotics. There are two major reasons for this: stringent regulatory requirements and a lack of monetary incentives for research (Fernandes & Martens, 2016; Williams & Bax, 2009). The latter is partly due to physicians who prefer to hold off on prescribing new antibiotics in fear of eliciting resistance against them. This results in reserving these antibiotics as a last resort. In addition to the prudence of prescription, antibiotics are also priced relatively low and their use is relatively
short. The potential profit of medicines used to treat, for example, chronic diseases or cancer is much higher since they are used immediately after emerging on the market, are higher priced and are used for longer periods of time. It is therefore no wonder that pharmaceutical companies opt to invest in these more profitable drugs (Gould & Bal, 2013; Piddock, 2012). As for the regulatory requirements, the size of antibiotic-related drug trials and the non-inferiority margins in phase III trials make for a difficult to reach goal not to forget the high cost. The problem with
non-inferiority tests is the comparison of novel antibiotics, effective against antibiotic resistant bacteria, to existing antibiotics which are ineffective against antibiotic resistant bacteria. The treatment group consists of patients with infections susceptible to both drugs. The tests are done this way since it would be unethical to give a patient suffering from an antibiotic resistant infection an ineffective antibiotic to compare results of two drugs. This results in a test where the
new drug is likely to be found inferior to the current drug, making such studies impractical. (Coates et al, 2011; Ventola, 2015a; Gould & Bal, 2013; Williams & Bax, 2009; Spellberg et al, 2011; Piddock, 2012).
1.1.2.2 Misuse
As previously mentioned, the use of antibiotics potentially acts as a stimulant for bacteria to develop resistance. In regards to this, antibiotics should be viewed as a limited natural resource, where individual misuse has an impact on the benefit of other users.
1. Introduction
5
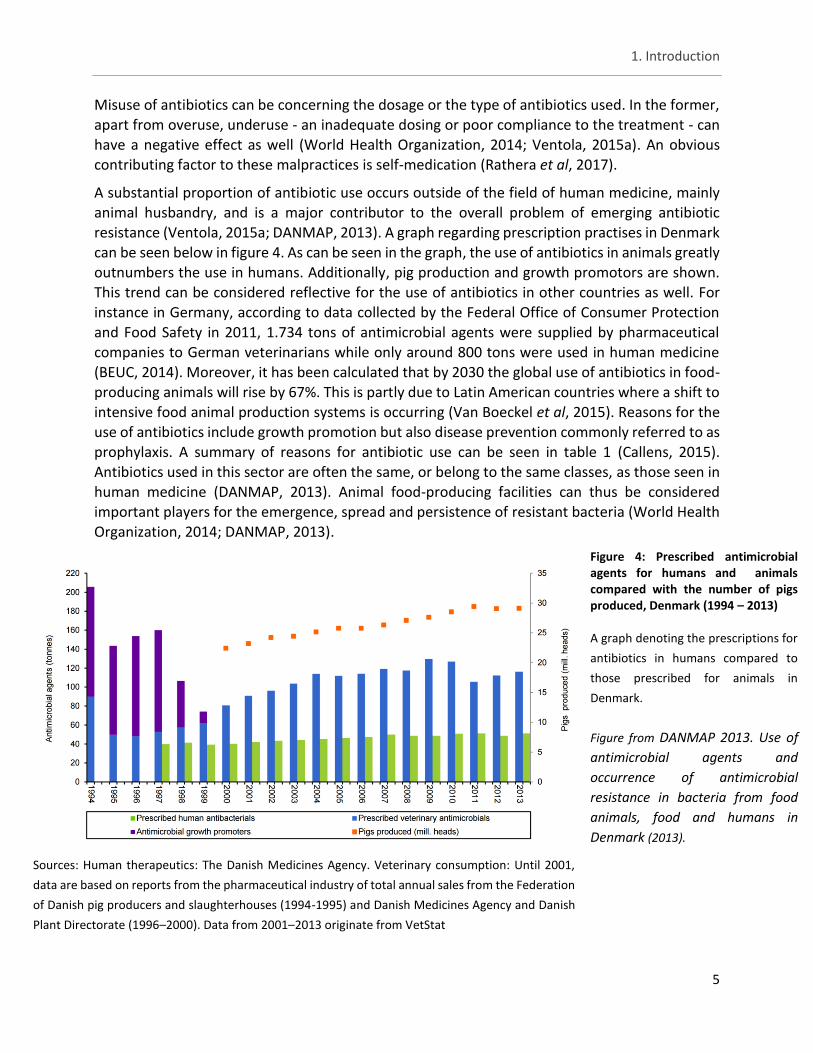
Figure 4: Prescribed antimicrobial agents for humans and animals compared with the number of pigs produced, Denmark (1994 – 2013)
A graph denoting the prescriptions for
antibiotics in humans compared to
those prescribed for animals in
Denmark.
Figure from DANMAP 2013. Use of
antimicrobial agents and
occurrence of antimicrobial
resistance in bacteria from food
animals, food and humans in
Denmark (2013).
Sources: Human therapeutics: The Danish Medicines Agency. Veterinary consumption: Until 2001,
data are based on reports from the pharmaceutical industry of total annual sales from the Federation
of Danish pig producers and slaughterhouses (1994-1995) and Danish Medicines Agency and Danish
Plant Directorate (1996–2000). Data from 2001–2013 originate from VetStat
Misuse of antibiotics can be concerning the dosage or the type of antibiotics used. In the former,
apart from overuse, underuse - an inadequate dosing or poor compliance to the treatment - can have a negative effect as well (World Health Organization, 2014; Ventola, 2015a). An obvious contributing factor to these malpractices is self-medication (Rathera et al, 2017).
A substantial proportion of antibiotic use occurs outside of the field of human medicine, mainly animal husbandry, and is a major contributor to the overall problem of emerging antibiotic resistance (Ventola, 2015a; DANMAP, 2013). A graph regarding prescription practises in Denmark can be seen below in figure 4. As can be seen in the graph, the use of antibiotics in animals greatly outnumbers the use in humans. Additionally, pig production and growth promotors are shown. This trend can be considered reflective for the use of antibiotics in other countries as well. For instance in Germany, according to data collected by the Federal Office of Consumer Protection
and Food Safety in 2011, 1.734 tons of antimicrobial agents were supplied by pharmaceutical companies to German veterinarians while only around 800 tons were used in human medicine
(BEUC, 2014). Moreover, it has been calculated that by 2030 the global use of antibiotics in food-producing animals will rise by 67%. This is partly due to Latin American countries where a shift to intensive food animal production systems is occurring (Van Boeckel et al, 2015). Reasons for the use of antibiotics include growth promotion but also disease prevention commonly referred to as prophylaxis. A summary of reasons for antibiotic use can be seen in table 1 (Callens, 2015). Antibiotics used in this sector are often the same, or belong to the same classes, as those seen in human medicine (DANMAP, 2013). Animal food-producing facilities can thus be considered important players for the emergence, spread and persistence of resistant bacteria (World Health Organization, 2014; DANMAP, 2013).
1. Introduction
6
1.1.3 Extended-spectrum β-lactamases and AmpC β-lactamases
A well-established antimicrobial resistance mechanism is that of bacterial strains producing extended-spectrum beta (β)-lactamases (ESBLs) and/or AmpC β-lactamases (AmpC). As a class, β-lactams inhibit bacterial cell wall synthesis which results in killing susceptible bacteria. On the other hand, ESBL- and/or AmpC-producing strains hydrolyse β-lactams, rendering them ineffective. ESBLs are commonly defined as plasmid-encoded enzymes found in e.g. Enterobacteriaceae conferring resistance to a range of ß-lactam antibiotics, including penicillins,
2nd-, 3rd- and 4th-generation cephalosporins and aztreonam (a monobactam) but rarely the carbapenems (e.g. imipenem) or the cephamycins (e.g. cefoxitin). Additionally, ESBL-producing bacteria are susceptible to β-lactamase inhibitors such as clavulanic acid (EFSA, 2011; Paterson & Bonomo, 2005). In contrast, AmpC β-lactamases are intrinsic cephalosporinases encoded in the chromosomal DNA of many Gram-negative bacteria but also observed on plasmids. They are not inhibited by clavulanic acid (Paterson & Bonomo, 2005; Jacoby, 2009). AmpC confers resistance to penicillins, 2nd- and 3rd-generation cephalosporins (e.g. cefotaxime), cephamycins (e.g.
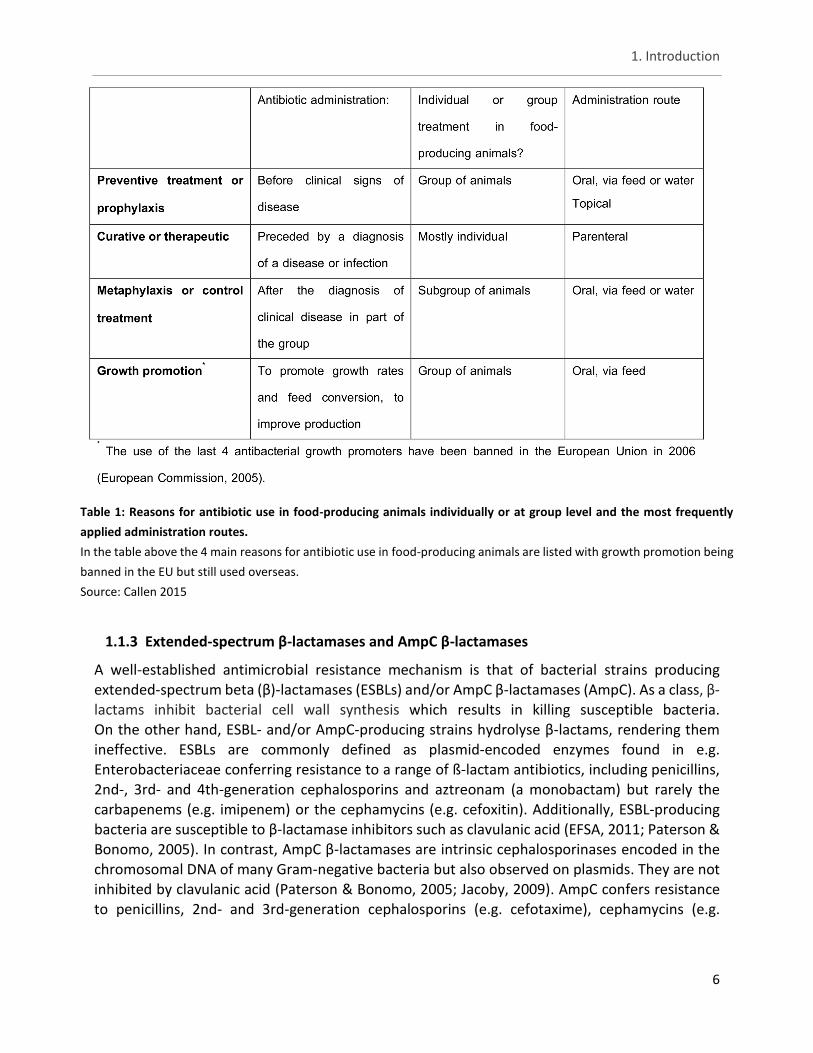
Table 1: Reasons for antibiotic use in food-producing animals individually or at group level and the most frequently
applied administration routes.
In the table above the 4 main reasons for antibiotic use in food-producing animals are listed with growth promotion being
banned in the EU but still used overseas.
Source: Callen 2015

1. Introduction
7
cefoxitin) including β-lactam/inhibitor combinations, but seldom to 4th-generation cephalosporin
(e.g. cefepime) and carbapenems (EFSA, 2011; Paterson & Bonomo, 2005; Asbel & Levison, 2000).
A similar structure is observed in the different classes of β-lactams. Cephalosporins, much like penicillins, possess a β-lactam ring which intervenes with the peptidoglycan cross-links within the bacterial cell wall. Cephalosporins differ from penicillins in that the penicillin 5-member thiazolidine ring is replaced by a 6-member dihydrothiazine ring. Cephalosporins are classified according to their antibacterial spectrum, a property depending on their side chain configurations. Early generations of cephalosporins showed a preferred activity against Gram-positive bacteria. In later generations, for example cefotaxime (3rd-generation), Gram-negative coverage was included (Wykoff et al, 2011). Much like cephalosporins, cephamycins exhibit a Gram-positive and Gram-negative coverage (Stapley et al, 1979; Brogden et al, 1979).
Furthermore, both share a cephem nucleus but cephamycins differ by possessing a 7-α-methoxyl group (fig. 5) which confers higher resistance to β-lactamases. In turn, carbapenems structurally
differ from other β-lactams by containing a methylene replacement for sulfur in the 5-membered α-ring structure (fig. 5). Carbapenems exhibit a wide range of activity, making them suitable against Gram-positive bacteria and Gram-negative bacteria (Asbel & Levison, 2000).
β-lactams are often administered as a first-line therapy for a wide variety of infections occurring in communities and hospitals. The most common cause of concern are community-acquired urinary tract infections caused by ESBL-producing Escherichia coli (E. coli). In a hospital setting, complications arise with respiratory tract and bloodstream infections caused by ESBL-producing
Klebsiella spp. (Pitout et al, 2005; Rodríguez-baño et al, 2004; Rodríguez-Baño, 2008). Furthermore, infections involving these resistant bacteria are associated with increased morbidity, mortality, increased length of stay and increased costs (Tumbarello et al, 2007; Ben-ami et al, 2009; Cosgrove, 2006). The use of β-lactams is not only limited to humans, on the contrary, its use in animal husbandry is greater than in human medicine (DANMAP, 2013; Li et al, 2007). In 2015, the most used group of antibiotics in animal husbandry in Belgium were the penicillins with 80.4 tons, accounting for 31% of the total use of antibiotics. In the same year 1.5 tons of cephalosporins were used which is only 0.58% of the total use of antibiotics (BelVetSAC,
Figure 5: Basic structure of cephalosporins, carbapenems and cephamycin
The different basic structures of some β-lactams. cephalosporins (1), cephamycins (2) and carbapenems (3)
Figure adapted from Asbel & Levison (2000) and Stapley (1979)

1. Introduction
8
2015). Consequently, ESBL-producing strains are also found in food-producing animals and in food
thus posing a public health concern (Lavilla et al, 2008). Next to ESBL-producing E. coli, other ESBL-producing bacteria are of interest as well, such as human pathogenic Salmonella (EFSA, 2011; Lavilla et al, 2008; Li et al, 2007; Bae et al, 2015).
Although there are a large number of genes which encode ESBL and AmpC, not all are equally prevalent among human and animal bacteria. The predominant ESBL families encountered are temoniera (TEM), sulfhydryl variable (SHV) and cefotaximase-munich (CTX-M). A widespread but not predominant ESBL gene family is oxacillin-hydrolyzing (OXA). The predominant AmpC-family is cephamycinase (CMY). The bacterial species most commonly identified with these genes are E. coli and non- typhoidal Salmonella (EFSA, 2011).
The TEM-derived β-lactamases are spread worldwide and responsible for 50–60% of the plasmid-
mediated ampicillin resistance in E. coli. The first TEM gene is capable of hydrolyzing penicillins and first-generation cephalosporins but unable to attack the oxyimino cephalosporins. Up until
now, more than 100 unique TEM variants have been described and increased activity against extended-spectrum cephalosporins is observed.
Much like TEM-1, SHV-1 confers resistance to broad-spectrum penicillins such as ampicillin, ticarcillin and piperacillin but not to the oxyimino substituted cephalosporins. The SHV gene is believed to have been derived from Klebsiella spp. So far, at least 40 variants have been described and most of them show enhanced resistance to cephalosporins (Paterson & Bonomo, 2005; Stürenburg & Mack, 2003). The rise in resistance against cephalosporins is especially noticed with cefotaxime and to a lesser extent against ceftazidime (Kliebe et al, 1985).
Unlike the two previous ESBL genes, where a few amino acid substitutions enabled an ancestral enzyme to expand its substrate spectrum, CTX-M has an intrinsic capability to hydrolyse extended-spectrum cephalosporins, preferentially cefotaxime. This capability was already present in the original chromosomal AmpC β-lactamase gene which was horizontally transferred and subsequently mutated (Poirel et al, 2002; Pe et al, 2001). Based on their amino acid sequences, they can be divided into five major groups: CTX-M group 1, 2, 8, 9, and 25 (Bonnet, 2004). An unusual characteristic of this gene family is that tazobactam exhibits an almost ten-fold stronger inhibitory activity on the produced enzymes compared to clavulanic acid (Stürenburg & Mack, 2003; Bush et al, 1993). It could be speculated that the CTX-M-type ESBLs are now actually the most frequent ESBL type worldwide (Paterson & Bonomo, 2005).
The last widespread family is the OXA-type enzymes. They are mainly found in Pseudomonas
aeruginosa, but also in Enterobacteriaceae. This group is considered to be phenotypic rather than genotypic due to an overall amino acid sequence correspondence of only 20–30% between the different oxacillinases (Stürenburg & Mack, 2003). Though OXA genes are found less than the other ESBL genes, some variants confer resistance to carbapenems, making them an important group since carbapenems (e.g. imipenem and meropenem) are the drugs of first choice in treating infections involving ESBL-producing strains. Besides the sporadic resistance against carbapenems, OXA enzymes confer resistance to penicillin, oxacillin and its activities are poorly inhibited by clavulanate (Evans & Amyes, 2014; Stürenburg & Mack, 2003; EFSA, 2011).

1. Introduction
9
It should be noted that the same organism may harbour ESBL genes from different ESBL families
and even AmpC-type β-lactamases, which may alter the antibiotic resistance phenotype (Yan et al, 2000).
Apart from carbapenems, the cephamycins (e.g. cefoxitin) are generally effective against Enterobacteriaceae producing TEM-, SHV- and CTX-M derived ESBLs. However this is not the case for AmpC producing strains. AmpC enzymes are phylogenetically very distinct from the ESBL families and confer resistance to penicillins, third-generation cephalosporins as well as to cephamycins. As for now, over 15 different plasmid-encoded enzymes (e.g. CMY, FOX, MOX, ACC, DHA, CIT) have been reported in several countries with CMY-2 being the most commonly found enzyme (Jacoby, 2009; Stürenburg & Mack, 2003; EFSA, 2011). However plasmid-mediated AmpC β-lactamases are less commonly found than ESBLs. The three-dimensional structures of AmpC
enzymes are very similar; they have a more open binding site to accommodate the bulkier side chains of cephalosporins (Jacoby, 2009).
Aside from carrying β-lactamase genes, most ESBL- and AmpC-producing strains also carry additional resistance genes against commonly used veterinary drugs such as sulphonamides (EFSA, 2011). In a study were 126 ESBL E. coli isolates were tested against 17 antibiotics (representing seven antibiotic groups) 92% showed resistance to at least one antibiotic and 57.9% were multidrug resistant (MDR; as defined in Magiorakos et al, 2012). The resistance profile of MDR strains ranged from 3 to 15 antibiotics (Ibrahim et al, 2016). For comparison, in a study concerning ESBL Salmonella isolates (44) 86% were MDR. The most frequent resistance profile was against beta-lactams, quinolones, and tetracyclines (Ziech et al, 2016). Other studies also show high percentages of ESBL isolates with additional resistance to other antibiotics (Smet et al,
2008). Therefore, generic antimicrobial use is a risk factor for ESBL/AmpC and it is not restricted specifically to the use of cephalosporins (EFSA, 2011; Jacoby, 2009). The transmission of ESBL/AmpC genes is mainly driven by integrons, insertion sequences, transposons and plasmids, some of which are homologous in isolates from both food producing-animals and humans (EFSA, 2011).
1.1.4 Colistin resistance
Recently, research has uncovered plasmid-mediated antibiotic resistance against colistin (polymixin E), a last resort antibiotic (Liu et al, 2016). Colistin belongs to the group of polymixins and is a cationic, multicomponent lipopeptide antibacterial agent (see Fig. 6) with narrow-spectrum activity against Gram-negative bacteria, including most species of the
Enterobacteriaceae family (Liu et al, 2016; Catry et al, 2015; Landman et al, 2008). Meaning that, much like β-lactams, its mode of action is disrupting bacterial cell walls. However, unlike β-lactams, the use of colistin for treatment of human infections was limited due to its systemic toxicity.

1. Introduction
10
Because of the emergence of multidrug resistant bacteria, such as ESBL-producing bacterial strains, usage of colistin in human medicine is increasing (Center for Disease Dynamics Economics & Policy, 2015). Whereas, colistin is already used widely in veterinary medicine. Main indications are infections caused by Enterobacteriaceae in pigs, poultry, bovines, sheep, goats and rabbits. According to field studies, colistin use is primarily observed in pigs as first-choice treatment and prevention of neonatal diarrhoea caused by E. coli and Salmonella in piglets (Callens et al, 2012; Timmerman et al, 2006; European Medicines Agency, 2013). In 2012, the total colistin consumption in medicated feed for pigs in Belgium was approximately 2500 kg active substance,
accounting for 4.5 % of the total medicated feed used (BelVet-SAC, 2013). In 2015, though, the use of colistin in Belgium has dropped about 51% when comparing to 2012; likely due to the use of zinc oxide in the treatment of post weaning diarrhoea in piglets rather than colistin (BelVetSAC, 2015). Additionally, colistin is largely used as a growth promotor in many countries (Hao et al, 2014). As mentioned above, nowadays, resistance to colistin has been found to occur plasmid-mediated and so far two distinct resistance genes are observed, namely mcr-1 and mcr-2. Both these genes are members of the phosphoethanolamine transferase enzyme family, which is responsible for adding a phosphoethanolamine to lipid A (Liu et al, 2016; Xavier et al, 2016). As of late, mcr-1 has at least been identified in 30 countries across five continents (Xavier et al, 2016; Skov & Monnet, 2016). Apart from resistance to colistin, some cases of E. coli with additional resistance mechanisms such as ESBL have been observed (Falgenhauer et al, 2016; Haenni et al, 2016b).
Figure 6: Chemical structure of colistin
Figure depicting the chemical structure of colistin with the
following abbreviations used: leucine (Leu), threonine (Thr),
phenyalanine (Phe), serine (Ser), isoleucine (Ile),
α,γ-diaminobutyric acid (Dab), 6-methyloctanoic acid
(MOA)
Source: (Kametani & Umezawa, 1980)

1. Introduction
11
1.2 Pigs and their importance to antibiotic resistance
The primary meat product in the European Union (EU) is pork, with an estimated production of 23.0 million tonnes in 2015 (Eurostat, 2016). In Belgium, the second highest antimicrobial-consuming livestock production system is fattening pigs, where on average over 200–250 per 1000 animals are treated daily with antimicrobials (Callens et al, 2012). As previously mentioned, the use of antibiotics in food-producing animals poses concerns to public health due to possible spread of antibiotic resistant bacteria (EFSA, 2011). Considering that in industrialized countries up to 10% of the population may annually suffer from foodborne diseases, having antibiotic resistant bacteria in food could be potentially disastrous (Kaferstein & Abdussalam, 1999).
1.2.1 Salmonella as a foodborne pathogen
One of the worldwide leading causes of foodborne illnesses is salmonellosis with an estimated 80.3 million cases occurring annually (Scallan et al, 2011; EFSA, 2009; Lahuerta et al, 2011; Majowicz et al, 2010). The genus Salmonella belongs to the Enterobacteriaceae family and is composed of facultative anaerobic, oxidase-negative, catalase-positive, Gram-negative, rod-shaped bacteria. Rods are typically 0.7 to 1.5 µm wide and 2 to 5 µm long. Most strains are motile and ferment glucose with production of both acid and gas. Currently two species of Salmonella exist i.e. Salmonella (S.) enterica and S. bongori. The former species is subdivided in 6 subspecies: subsp. enterica, subsp. salamae, subsp. arizonae, subsp. diarizonae, subsp. houtenae, subsp. indica. Salmonellae are further characterized into serotypes based on differences in epitopes of lipopolysaccharide, designated the O or somatic antigen, a major component of the outer membrane of Gram-negative bacteria. Lipopolysaccharide has three components: lipid A, core
polysaccharide, and an oligosaccharide side chain which confers serogroup specificity (Cox & Pavic, 2014). As of 2008, there were 2579 serotypes of Salmonella enterica, of which 58.9% belong to subspecies enterica (Cox & Pavic, 2014).
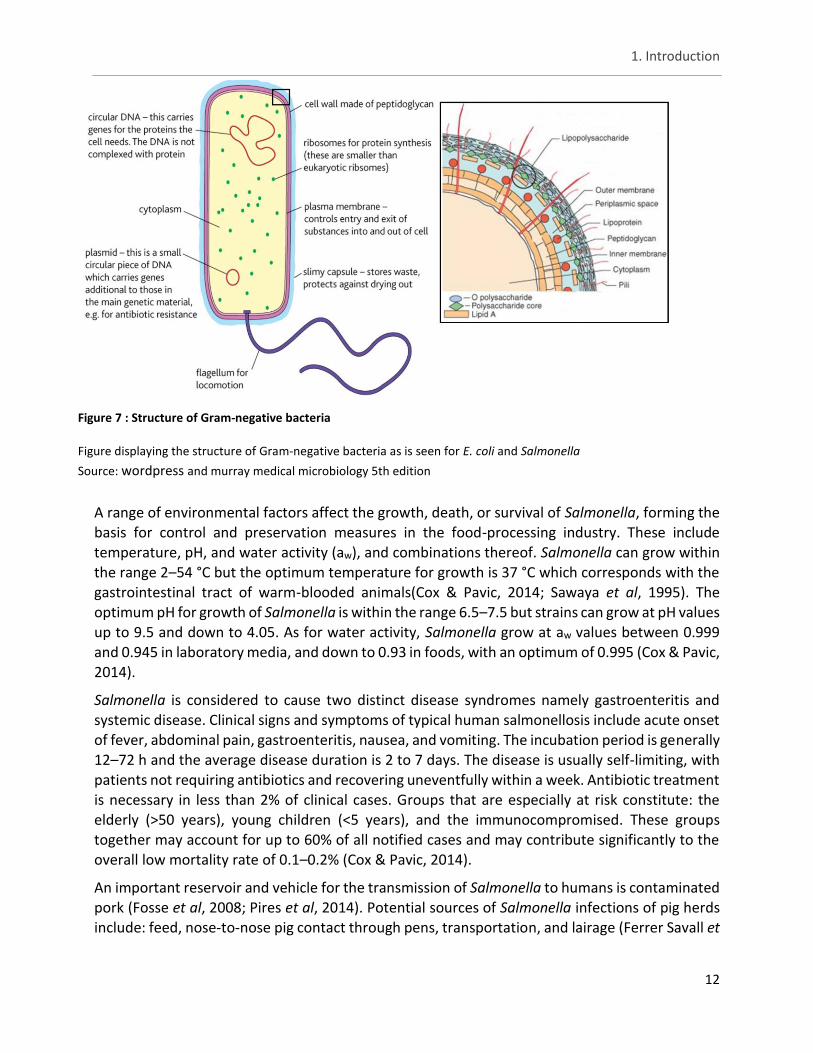
In figure 7, on the left, the entire rod shaped body of a Gram-negative bacteria can be seen with a single flagellum attached to it. In reality the bacteria has more than one attached to the body. The different features are marked and described on the figure, with the most important in regards to antibiotic resistance mechanisms being: the cell wall (further described on the right part of the figure), plasmid and the circular DNA. The plasmid is of importance since it can carry resistance genes and exchange these with other bacteria. In contrast, the circular DNA can also carry resistance genes but besides being able to transmit it through vertical transmission, the resistance is limited in spread and persistence. Finally, the cell wall is made up of different elements:
lipopolysaccharide is important for serotyping and consists of the three subunits (fig. 7). Lipoprotein is where colistin exudes its activity. Peptidoglycan keeps the cell intact, protects it from osmotic pressure and is the site with which β-lactam antibiotics interfere. Pili are one of the ways bacteria can exchange resistance genes with one another (Frost et al, 2005).
1. Introduction
12
Figure 7 : Structure of Gram-negative bacteria
Figure displaying the structure of Gram-negative bacteria as is seen for E. coli and Salmonella
Source: wordpress and murray medical microbiology 5th edition
A range of environmental factors affect the growth, death, or survival of Salmonella, forming the basis for control and preservation measures in the food-processing industry. These include
temperature, pH, and water activity (aw), and combinations thereof. Salmonella can grow within the range 2–54 °C but the optimum temperature for growth is 37 °C which corresponds with the gastrointestinal tract of warm-blooded animals(Cox & Pavic, 2014; Sawaya et al, 1995). The optimum pH for growth of Salmonella is within the range 6.5–7.5 but strains can grow at pH values up to 9.5 and down to 4.05. As for water activity, Salmonella grow at aw values between 0.999 and 0.945 in laboratory media, and down to 0.93 in foods, with an optimum of 0.995 (Cox & Pavic, 2014).
Salmonella is considered to cause two distinct disease syndromes namely gastroenteritis and systemic disease. Clinical signs and symptoms of typical human salmonellosis include acute onset of fever, abdominal pain, gastroenteritis, nausea, and vomiting. The incubation period is generally 12–72 h and the average disease duration is 2 to 7 days. The disease is usually self-limiting, with
patients not requiring antibiotics and recovering uneventfully within a week. Antibiotic treatment is necessary in less than 2% of clinical cases. Groups that are especially at risk constitute: the elderly (>50 years), young children (<5 years), and the immunocompromised. These groups together may account for up to 60% of all notified cases and may contribute significantly to the overall low mortality rate of 0.1–0.2% (Cox & Pavic, 2014).
An important reservoir and vehicle for the transmission of Salmonella to humans is contaminated pork (Fosse et al, 2008; Pires et al, 2014). Potential sources of Salmonella infections of pig herds include: feed, nose-to-nose pig contact through pens, transportation, and lairage (Ferrer Savall et

1. Introduction
13
al, 2016; Rostagno & Callaway, 2012; Mannion et al, 2008; De Busser et al, 2011).
Additionally, contamination can occur during the different stages of the slaughtering process, such as scalding, dehairing, and polishing. Since many pigs carry Salmonella asymptomatically at time of slaughter, these pathogens cannot be detected using routine ante and post mortem inspection, resulting in the spread to carcasses and meat during slaughter and processing of the meat (Van Damme et al, 2017; Rostagno & Callaway, 2012).
1.2.2 E. coli as hygiene indicator and potential reservoir for resistance
E. coli, commonly present in the intestinal tract of pigs, other animals and humans, can be used as an indicator of faecal contamination and possible presence of enteric pathogens (SØrum & Sunde, 2001). Studies have shown that increased levels of E. coli counts on pig carcasses correlate
to an increased probability of finding Salmonella on those pig carcasses and the sub sequential pork cuts (Bollerslev et al, 2017; Ghafir et al, 2008; Delhalle et al, 2008). Furthermore, E. coli could
possibly serve as indicator for antibiotic resistance. As these bacteria form a reservoir of mobile resistance genes that can be transferred to other bacteria including Salmonella, thereby playing a key role in the dissemination and persistence of resistance (Marshall & Levy, 2011). In addition, E. coli has an exceptional capability to acquire and spread resistance genes (Smith et al, 2007). Because of the reasons mentioned above, in addition to the fact that E. coli is easy to isolate and identify, it is internationally used as a hygiene indicator (Wray & Gnanou, 2000).
E. coli, just like Salmonella, is part of the Enterobacteriaceae family and a Gram-negative, facultative anaerobic, rod-shaped, coliform bacterial species with an optimum growth temperature of 37 °C (Tenaillon et al, 2010). However, unlike Salmonella, E. coli can be both a
commensal and a potential pathogen, depending on the strain (SØrum & Sunde, 2001).
In humans, there are six well-described intestinal pathogenic E. coli: : enteropathogenic E. coli, enterohaemorrhagic E. coli (Shiga toxin-producing E. coli ), enterotoxigenic E. coli, enteroaggregative E. coli, enteroinvasive E. coli and diffusely adherent E. coli. Infections with one of these pathogens can result in three general clinical syndromes namely enteric/diarrhoeal disease, urinary tract infections and sepsis/meningitis. Pathogenic E. coli exist in animals as well and give rise to similar diseases (Croxen et al, 2013; Kaper et al, 2004; Bélanger et al, 2011). Some major pathogenic E. coli in pigs are enterotoxigenic E. coli and Shiga toxin-producing E. coli. Enterotoxigenic E. coli are associated with neonatal diarrhoea in pigs as well as with post-weaning diarrhoea; whereas Shiga toxin-producing E. coli are responsible for oedema disease (Croxen et al, 2013). Shiga toxin cleaves ribosomal RNA, thereby disrupting protein synthesis and killing the
intoxicated epithelial or endothelial cells (Kaper et al, 2004).
Transfer of virulence genes and antibiotic resistance genes in E. coli in food-producing animals and on meat may pose a zoonotic risk and are a great concern for human health. In many recent studies, profiles of E. coli strains isolated from retail meat resembled, and in some cases matched, those of human extra-intestinal pathogenic E. coli clinical isolates (Bélanger et al, 2011; Leverstein-van Hall et al, 2011; Jakobsen et al, 2010; Ewers et al, 2009; Trobos et al, 2009).

1. Introduction
14
E. coli are exposed to antibiotics directly after oral treatment thus stimulating antibiotic
resistance. Interestingly besides exposure to antibiotics, stress also resulted in detecting more resistant E. coli in pigs (SØrum & Sunde, 2001).
Stress due to transport, overcrowding in holding pens and rough handling before slaughter but also heat or cold stress results in increased shedding of antibiotic resistant enteric bacteria into the environment due to increased intestinal motility (SØrum & Sunde, 2001; Molitoris et al, 1987; Moro et al, 2000, 1998).
1.3 Methods for detection of antibiotic resistance
Detection of antibiotic resistance/susceptibility can be done by means of disk diffusion or microbroth dilution or an E-test (EFSA, 2011). Disk diffusion is a method which screens antibiotic
susceptibility by noting specific zone diameters on agar plates. Just as the name suggests, a disk containing an antibiotic compound is placed onto an agar plate, inoculated with bacteria, where
the drug will diffuse through the agar. The concentration of the compound will be the highest next to the disk, and will decrease as distance from the disk increases. The zone diameters are the result of bacteria not growing enough to be visible (either through growth inhibition or killing), these are called inhibition zones (Patel et al, 2011). The MIC method consists of identifying the lowest concentration of an anti-microbial that will inhibit the visible growth of a microorganism after overnight incubation (Andrews, 2001). These tests are often accompanied by verification for the presence of resistance genes by means of a polymerase chain reaction (PCR) followed by agarose gel electrophoresis (Stürenburg & Mack, 2003; Smith, 1996; Paterson & Bonomo, 2005; EFSA, 2011; CLSI, 2013).
To identify ESBL suspected Enterobacteriaceae by MIC tests, optimum breakpoints or interpretive criteria need to be used. ESBL producers are resistant to cefotaxime, variably resistant to ceftazidime, susceptible to cefoxitin and inhibition occurs in the presence of clavulanic acid (EFSA, 2011). Confirmation of ESBLs is performed by testing for synergy with clavulanic acid by combination disks, ESBL E-tests or microbroth dilution including cefotaxime, and ceftazidime as single drugs, and in combination with clavulanic acid. AmpC producers are susceptible to cefepime and generally resistant to cefotaxime, ceftriaxone and cefoxitin (EFSA, 2011). Additionally AmpC β-lactamases are quite resistant to clavulanic acid (Monnaie & Frere, 1993). Strains producing enough AmpC β-lactamase will typically give a positive ESBL screening test but fail the confirmatory test involving clavulanate synergy (Bell et al, 2007). This phenotype is not, however, specific for an AmpC producer, since it can occur with certain complex TEM mutants,
OXA-type ESBLs, and carbapenemases and in strains with high levels of TEM-1 β-lactamase (Robin et al, 2006; Poirel et al, 2000; Jacoby, 2009; Bell et al, 2007). For these reasons there are presently no Clinical Laboratories Standards Institute (CLSI) or other approved criteria for AmpC detection (Doi & Paterson, 2007; Bhaskar & Mandal, 2016). However, cefoxitin resistance in Enterobacteriaceae combined with resistance to an oxyimino-cephalosporin, such as ceftazidime, suggests an AmpC phenotype (Livermore, 1995). In organisms which produce both ESBLs and AmpC, clavulanate may induce hyperproduction of AmpC, leading to hydrolysis of the 3rd-generation cephalosporin (e.g. cefotaxime, ceftazidime) masking any synergy arising from

1. Introduction
15
inhibition of the ESBL.The European Committee on Antimicrobial Susceptibility Testing (EUCAST)
epidemiological cut-off values (ECOFFs) are used to determine if an isolate belongs to the wild-type population or possesses one or more specific mechanisms that decrease its antibiotic susceptibility. For molecular identification of ESBL and/or AmpC genes, PCR and sequence analysis of the amplicons are used to screen which β-lactamase families are present (e.g. TEM, SHV, or CMY) (EFSA, 2011; Paterson & Bonomo, 2005).
Disk diffusion and microbroth dilution are generally accepted as giving comparative results (Mohd Nasir & Parasakthi, 2004; Skov et al, 2001). However, the disk diffusion technique was reported to be an unreliable method for evaluating susceptibility to polymyxins due to the poor diffusion of these agents in agar. Therefore, the use of microbroth dilution is recommended for testing colistin susceptibility (Landman et al, 2008; Galani et al, 2008; EUCAST, 2017). Recently, a study
by Boyen et al (2010) demonstrated good results in using disk prediffusion for testing colistin susceptibility in porcine E. coli strains.
1.4 Course of action against antibiotic resistance
To preserve the benefits of having effective antibiotics, different steps should be undertaken. An immediate solution would be improving sanitation and effective vaccines to reduce the prevalence of disease and thereby also reduce the need for antibiotics. This can be applied in animal production as well as in hospital settings. Improving hygiene at all steps of the food chain will play a short-term role in mediating the situation by lowering the overall bacterial count. However, as a sole measure it will not mitigate the selection at the source (World Health Organization, 2014; Bélanger et al, 2011; Center for Disease Dynamics Economics & Policy, 2015).
Prudent use of antibiotics is necessary as to reduce the selection pressure imposed by the use of such agents. This implies a decrease of the total antibiotic use in animal production. Stopping the use of antibiotics as growth promotor, as is already the case in the EU, would be important here. In addition, the use of antibiotics for disease prophylaxis should be kept to a minimum. Another way would be preserving activity of certain antibiotics (e.g. cephalosporins) by restricting their use, allowing use only under specific circumstances. Although co-resistance then becomes an important aspect to consider (Bartlett et al, 2013; Center for Disease Dynamics Economics & Policy, 2015).
Educating and informing health professionals, policymakers, and the public on sustainable antibiotic use will also play a critical role. For instance this might stop misuse (incorrect
prescribing practices), overuse and underuse (inadequate dosing) (Center for Disease Dynamics Economics & Policy, 2015; Ventola, 2015a, 2015b). Another way to reduce inappropriate antibiotic use is by improving diagnostic methods and tools. Furthermore, this will aid in identifying antibiotic resistant infections, making it possible to provide the appropriate health care (World Health Organization, 2014; Michael et al, 2014). The use of antibiotic stewardship programs which guide all prescribers in administering antibiotics correctly have shown to be effective.

1. Introduction
16
Multiple results such as improved patient care, shortened hospital stays, and reduced health care
facilities pharmacy costs are some examples (CDC, 2013; Embolism, 2008; Ventola, 2015b).
Finally alternative treatment methods such as bacteriophage therapy could be a potential solution for the antibiotic resistant bacteria. Not only can such a therapy be effective against antibiotic bacteria, it also brings less complications. Unlike antibiotics that target both pathogens and normal flora of patients; bacteriophages are very specific to their hosts and consequently minimize the chance of secondary infections. In addition, bacteriophages concentrate at the site of infection where they replicate and lyse the pathogens. In contrast, antibiotics travel throughout the body and do not specifically concentrate at the infection site. Unlike antibiotics, which are known to cause side effect, no side effects have been reported during or after phage application. Lastly, phage resistance is not nearly as worrisome as drug resistance. Since, like bacteria, phages
mutate meaning they can evolve to counter phage-resistant bacteria (Golkar et al, 2014; Ho, 2001; Matsuzaki et al, 2005).

2. Aim of the Research Project
17
2. Aim of the Research Project.
ESBL/AmpC-producing Enterobacteriaceae are already established as occurring worldwide (Ojer-Usoz et al, 2013). Community- and healthcare-associated infections with ESBL/AmpC β-lactamase–producing bacteria may result in delays in the initiation of timely and adequate antimicrobial therapy, increased morbidity and mortality, longer hospital stays, and higher costs (Pitout & Laupland, 2008; Schwaber & Carmeli, 2007; de Kraker et al, 2011; Liebana et al, 2013). Since the report of the plasmid-mediated mcr-1 and mcr-2 colistin resistance genes in 2015 and 2016, there have been reports of plasmid-mediated colistin resistant Enterobacteriaceae in many European countries such as Germany, The United Kingdom, Belgium and France as well as in other parts of the worlds, for example South Africa, the United States of America (USA), China, Brazil and Japan. Some of these reports concern mcr-1 positive bacteria isolated before 2015
(EFSA, 2011; Liu et al, 2016; Skov & Monnet, 2016; ECDC, 2016). Keeping in mind that colistin is a
last resort antibiotic, these findings are quite worrisome. There is limited knowledge about how far the extent of resistance has spread within the respective Enterobacteriaceae genera and even more about how fast the resistance spreads between the different genera. In addition, food could very well be a possible route for transmission of these bacteria (EFSA, 2011). ESBL/AmpC-producing and/or colistin resistant Enterobacteriaceae present a serious threat for our global health. As a result there is a need to contain the spread and to re-evaluate the situation.
Therefore the aim of this study is to characterise cefotaxime resistant E. coli isolates and Salmonella isolates, originating from slaughterhouse pigs for the presence of ESBL/AmpC production and reduced susceptibility to colistin. The isolates in the lab library were taken from
2014 till 2016. Susceptibility will be tested using disk (pre)diffusion and Minimum Inhibitory Concentration (MIC) by means of broth microdilution. Additionally the presence of resistance genes will be evaluated by PCR followed by agarose gel electrophoresis. The different genes responsible for ESBL/AmpC production will be examined to see which ones occur the most. Also, the possible co-occurrence and correlation of a resistant pathogen (Salmonella) and resistant E. coli will be investigated as to see if E. coli with reduced susceptibility can serve as an indicator for resistant Salmonella. Furthermore it will be investigated whether or not reduced susceptibility against the antibiotics of interest co-occurs with reduced susceptibility to other antibiotics. Finally, the different methods for testing antibiotic resistance/reduced susceptibility namely microbroth dilution and disk (pre)diffusion will be compared to each other.
The study was split up into two parts, with most of the aims being investigated in study A. In study
B, the specific aim was to study the occurrence of ESBL genes in pig-associated E. coli within pig batches at slaughter. A summary of this thesis can be seen in figure 8.

2. Aim of the Research Project
18
Figure 8: Summary of Study A and Study B
Figure depicting which parts of the thesis where done in function of study A and which parts where done in function of study B.
3. Results
19
3. Results Antibiotic susceptibility of pig-associated E. coli and Salmonella isolates taken from three slaughterhouses were evaluated by means of disk diffusion, disk prediffusion and microbroth dilution. Furthermore a PCR followed by agarose gel electrophoresis was performed to verify for the presence of ESBL/AmpC and/or colistin resistance genes. The source and number of E. coli and Salmonella isolates used can be seen in section 5.4.1.
3.1 Study A - ESBL/AmpC characterisation of the isolates
3.1.1 Phenotyping of the isolates by means of disk diffusion
A disk diffusion was performed on all isolates to determine whether or not they showed an ESBL and/or AmpC phenotype. However, for reasons mentioned in 1.3, the AmpC phenotype from this test is indicative rather than definitive. The results obtained from the disk diffusion are summarized in table 2. None of the Salmonella isolates showed an ESBL/AmpC phenotype. All but two E. coli isolates could be categorised as ESBL/AmpC. Most of the E. coli isolates (74%) displayed an ESBL phenotype, only a limited number displayed an AmpC phenotype (7%) and few displayed a combined ESBL and AmpC phenotype (16%).
3.1.2 Genetic profiling of the isolates for ESBL/AmpC genes
Genetic profiling was investigated by PCR followed by agarose gel electrophoresis. The PCRs were
split up in multiplex I: blaTEM, blaSHV and blaOXA, multiplex II:blaCTX-M1, blaCTX-M2 and blaCTX-M9, multiplex III: blaEBC, blaMOX, blaCIT, blaACC and blaFOX, simplex I: blaCMY-2. An example of a result of agarose gel electrophoresis can be viewed in figure 9. Likewise, the gels from the other multiplexes and isolates are similar to the example given in figures 9.
Number (%) of isolates categorised as
E. coli Salmonella found without ESBL/AmpC E. coli
Salmonella found together with ESBL/AmpC E. coli
ESBL strain 50 (74) 0 (0) 0 (0) AmpC strain 5 (7) 0 (0) 0 (0) ESBL + AmpC strain 11 (16) 0 (0) 0 (0) Atypical 2 (3) 0 (0) 0 (0)
Table 2: Categorisation of the isolates according to their phenotype
Table demonstrating the number (and percentages) of isolates that displayed one out of three phenotypes or was showed to
be atypical. The three phenotypes were ESBL, AmpC or a combination of the two. Isolates are presented according to the
group they belong to.

3. Results
20
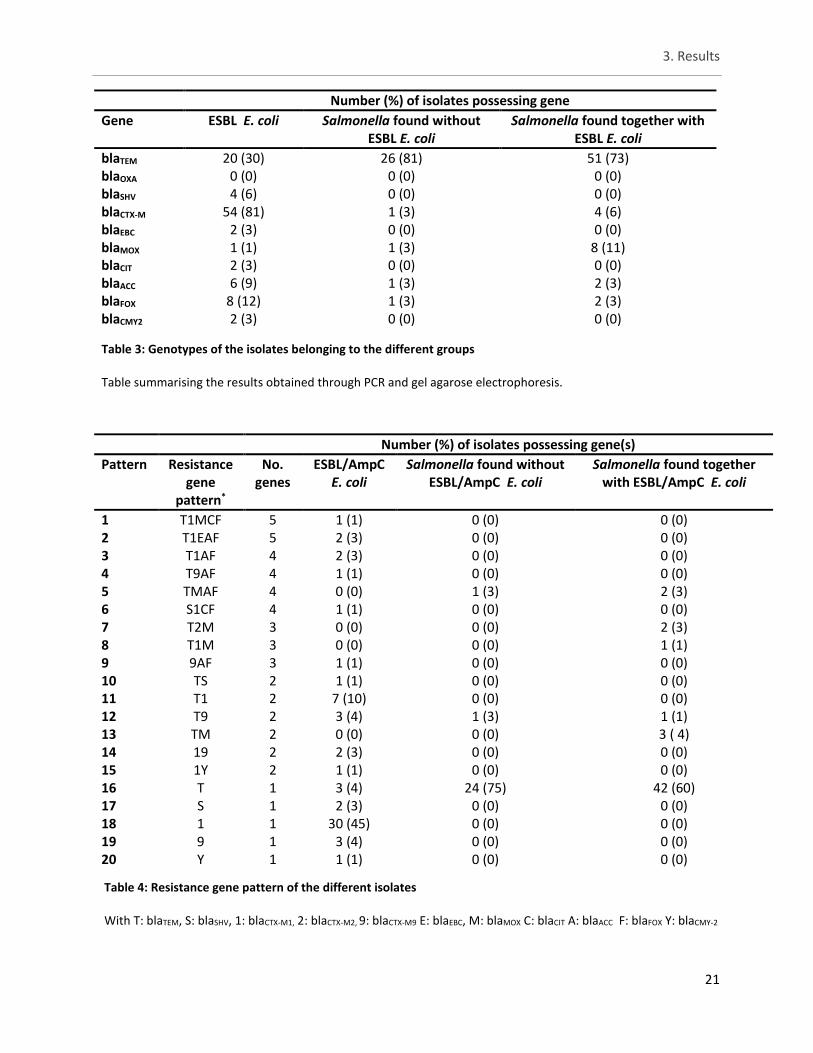
Both Salmonella groups showed a high amount (over 70%) of isolates possessing blaTEM gene. In contrast only 30% of ESBL/AmpC E. coli demonstrated this genotype, which is possible evidence that Salmonella found with ESBL/AmpC E. coli are not different from Salmonella found without the presence of ESBL/AmpC E. coli. For ESBL/AmpC E. coli, genes belonging to the blaCTX-M family were the most commonly occurring. In table 3, the summary of the genetic profiles. The different CTX-M genes were clustered together in table 3 but the distribution of the subtypes can be viewed in table 4. In table 4, the pattern of resistance genes are presented as to demonstrate the occurrence of multiple ESBL and/or AmpC genes in a single isolate. The total amount of ESBL/AmpC E. coli that exhibited more than one gene was 29%. Only three ESBL/AmpC E. coli isolates showed to possess five different genes all at once. In total 20 unique patterns were observed varying between five different genes to only possessing one. Especially for Salmonella, most of the time an isolate was found to only possess one resistance gene. For ESBL/AmpC E. coli three isolates (4%) were found with five different resistance genes all at once. Similarly, for both Salmonella groups 3% was found to possess four different resistance genes all at once.
Figure 9: Resulting agarose gel of multiplex I PCR to determine the presence of ESBL genes in isolates
Agarose gel of the multiplex I PCR (blaTEM, blaSHV, blaOXA) obtained after analysis with Quantity One®.
The ladder is marked on the left, a blanc and control bands can be seen on the far right. A ‘+’ sign represents an isolate that possessed the ESBL gene whereas a ‘-‘ sign signifies an isolates that was negative for the ESBL gene
3. Results
21
Number (%) of isolates possessing gene
Gene ESBL E. coli Salmonella found without ESBL E. coli
Salmonella found together with ESBL E. coli
blaTEM 20 (30) 26 (81) 51 (73) blaOXA 0 (0) 0 (0) 0 (0) blaSHV 4 (6) 0 (0) 0 (0) blaCTX-M 54 (81) 1 (3) 4 (6) blaEBC 2 (3) 0 (0) 0 (0) blaMOX 1 (1) 1 (3) 8 (11) blaCIT 2 (3) 0 (0) 0 (0) blaACC 6 (9) 1 (3) 2 (3) blaFOX 8 (12) 1 (3) 2 (3) blaCMY2 2 (3) 0 (0) 0 (0)
Number (%) of isolates possessing gene(s)
Pattern Resistance gene
pattern*
No. genes
ESBL/AmpC E. coli
Salmonella found without ESBL/AmpC E. coli
Salmonella found together with ESBL/AmpC E. coli
1 T1MCF 5 1 (1) 0 (0) 0 (0) 2 T1EAF 5 2 (3) 0 (0) 0 (0) 3 T1AF 4 2 (3) 0 (0) 0 (0) 4 T9AF 4 1 (1) 0 (0) 0 (0) 5 TMAF 4 0 (0) 1 (3) 2 (3) 6 S1CF 4 1 (1) 0 (0) 0 (0) 7 T2M 3 0 (0) 0 (0) 2 (3) 8 T1M 3 0 (0) 0 (0) 1 (1) 9 9AF 3 1 (1) 0 (0) 0 (0) 10 TS 2 1 (1) 0 (0) 0 (0) 11 T1 2 7 (10) 0 (0) 0 (0) 12 T9 2 3 (4) 1 (3) 1 (1) 13 TM 2 0 (0) 0 (0) 3 ( 4) 14 19 2 2 (3) 0 (0) 0 (0) 15 1Y 2 1 (1) 0 (0) 0 (0) 16 T 1 3 (4) 24 (75) 42 (60) 17 S 1 2 (3) 0 (0) 0 (0) 18 1 1 30 (45) 0 (0) 0 (0) 19 9 1 3 (4) 0 (0) 0 (0) 20 Y 1 1 (1) 0 (0) 0 (0)
Table 3: Genotypes of the isolates belonging to the different groups
Table summarising the results obtained through PCR and gel agarose electrophoresis.
Table 4: Resistance gene pattern of the different isolates
With T: blaTEM, S: blaSHV, 1: blaCTX-M1, 2: blaCTX-M2, 9: blaCTX-M9 E: blaEBC, M: blaMOX C: blaCIT A: blaACC F: blaFOX Y: blaCMY-2

3. Results
22
3.2 Study A - Colistin susceptibility of ESBL/AmpC-producing E. coli and Salmonella 3.2.1 Colistin susceptibly according to disk prediffusion
Only four isolates from the group of ‘Salmonella found without ESBL/AmpC E.coli ’ presented a reduced susceptibility to colistin. Admittedly, all these isolates were obtained from the same pig, with two isolates originating from the tonsils and two originating from rectum content. All other isolates exhibited susceptibility to the colistin. The results obtained from the disk prediffusion are shown in table 5.
3.2.2 Genetic profiling of the isolates for plasmid-mediated colistin resistance genes
In isolates showing reduced susceptibility to colistin, the presence of plasmid-mediated colistin
resistance genes in isolates indicative for the phenotype was investigated by a PCR multiplex for
mcr-1/2 followed by agarose gel electrophoresis. An example of a mcr-1/2 gel can be viewed in
figure 10.
Number (%) of isolates with reduced susceptibility
Antibiotic ESBL/AmpC E. coli Salmonella found without ESBL/AmpC E. coli
Salmonella found together with ESBL/AmpC E. coli
Colistin 0 (0) 4 (13) 0 (0)
Table 5: Number of isolates with reduced susceptibility to colistin
Table displaying the number (and percentages) of isolates with reduced susceptibility to colistin. Number of isolates are shown
according to their group.
Figure 10: Resulting agarose gel of plasmid-mediated colistin resistance gene PCR to determine the
presence of plasmid-mediated colistin resistance genes in isolates
Agarose gel to determine the presence of plasmid-mediated colistin genes (mcr-1/2) obtained after
analysis with Quantity One®.

3. Results
23
None of the screened isolates were shown to carry a plasmid-mediated colistin resistance gene
(mcr-1/2). This result is shown in table 6 below.
3.3 Study A - Phenotypical testing by means of microbroth dilution
For the remaining antibiotics the number of isolates with reduced susceptibility can be viewed in table 7. The Salmonella isolates from both groups showed reduced susceptibility to only a few antibiotics. The same four isolates with reduced susceptibility were observed here as well as one isolate belonging to ESBL E. coli and one isolate belonging to the Salmonella isolates found together with ESBL/AmpC E. coli. Importantly these additional two isolates did not come from the same batches so yet no correlation between the two groups is observed. For antibiotics not belonging to the β-lactams groups, reduced susceptibility against doxycycline (DOX) is observed in more than half of all the Salmonella isolates with 78% for ‘Salmonella found without ESBL/AmpC E.coli‘ and 56% for ‘Salmonella found together with ESBL/AmpC E.coli‘. In turn, for
trimethoprim/sulfamethoxazole 47% of the ‘Salmonella found without ESBL/AmpC E.coli’ showed reduced susceptibility and 56% of the ‘Salmonella isolates found together with ESBL/AmpC E.coli‘ showed reduced susceptibility. Finally for ciprofloxacin (CIP), only four isolates (6%) of ‘Salmonella found together with ESBL/AmpC E.coli ‘ showed reduced susceptibility.
For ESBL/AmpC E. coli various degrees of reduced susceptibility were observed although no reduced susceptibility against the three present carbapenems (meropenem, imipenem and doripenem) was shown, which is indicative that no isolates were carbapenemase producing. The most commonly observed reduced susceptibility in regards to ESBL/AmpC E. coli isolates was against trimethoprim/sulfamethoxazole, aztreonam and cefotaxime with an observed frequency higher than 80%. Additionally, frequent (approximate 70% occurring) reduced susceptibility
against cefepime and ceftazidime was observed. Apart from no reduced susceptibility to piperacillin/tazobactam constant 4, ESBL/AmpC E. coli showed various reduced susceptibility rates to DOX, gentamicin (GEN), tobramycin (TOB), CIP, levofloxacin (LEVO), ampicillin / sulbactam 2:1 ratio (A/S2) and ticarcillin / clavulanic acid constant 2 (TIM2) with an observed range between 10% - 46%. In conclusion, frequent reduced susceptibility against β-lactams was observed for the ESBL E. coli isolates.
Number (%) of isolates possessing gene
Gene ESBL E. coli Salmonella found without ESBL E. coli
Salmonella found together with ESBL E. coli
mcr-1/2 0 (0) 0 (0) 0 (0)
Table 6: Result of mcr-1/2 multiplex
Table summarising the results of the mcr-1/2 multiplex through PCR and gel agarose
electrophoresis.
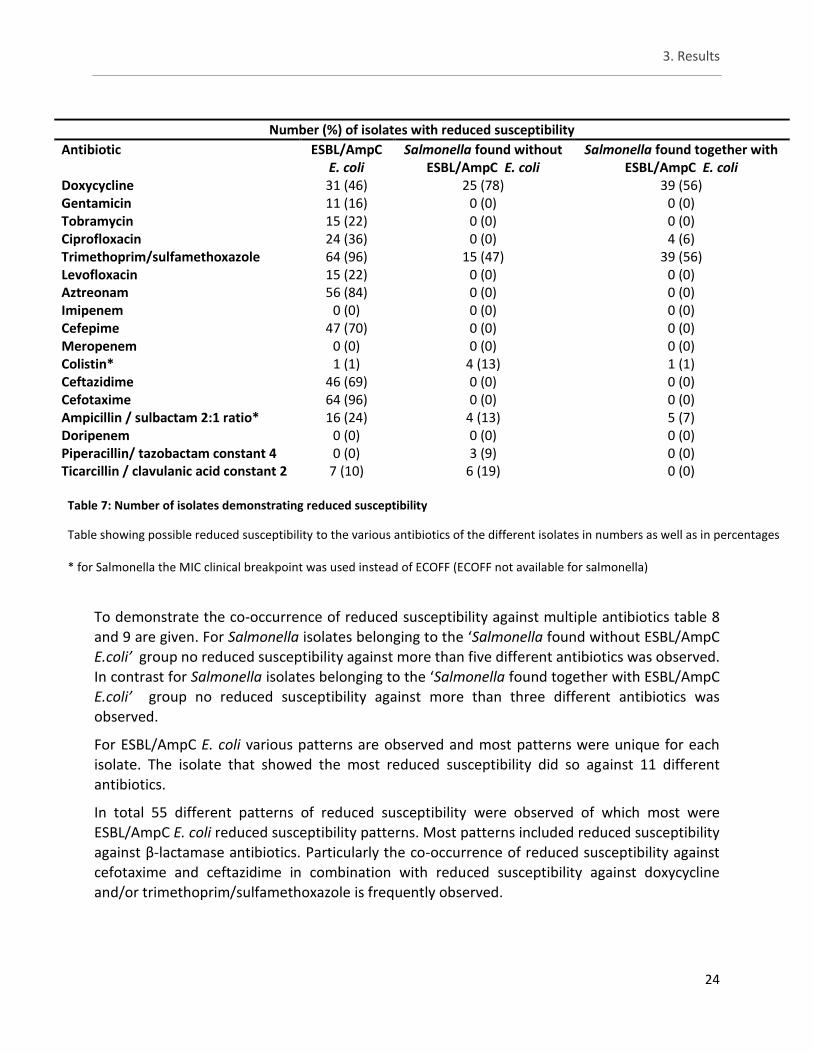
3. Results
24
To demonstrate the co-occurrence of reduced susceptibility against multiple antibiotics table 8 and 9 are given. For Salmonella isolates belonging to the ‘Salmonella found without ESBL/AmpC E.coli’ group no reduced susceptibility against more than five different antibiotics was observed. In contrast for Salmonella isolates belonging to the ‘Salmonella found together with ESBL/AmpC E.coli’ group no reduced susceptibility against more than three different antibiotics was
observed.
For ESBL/AmpC E. coli various patterns are observed and most patterns were unique for each isolate. The isolate that showed the most reduced susceptibility did so against 11 different
antibiotics.
In total 55 different patterns of reduced susceptibility were observed of which most were ESBL/AmpC E. coli reduced susceptibility patterns. Most patterns included reduced susceptibility against β-lactamase antibiotics. Particularly the co-occurrence of reduced susceptibility against cefotaxime and ceftazidime in combination with reduced susceptibility against doxycycline and/or trimethoprim/sulfamethoxazole is frequently observed.
Number (%) of isolates with reduced susceptibility
Antibiotic ESBL/AmpC E. coli
Salmonella found without ESBL/AmpC E. coli
Salmonella found together with ESBL/AmpC E. coli
Doxycycline 31 (46) 25 (78) 39 (56) Gentamicin 11 (16) 0 (0) 0 (0) Tobramycin 15 (22) 0 (0) 0 (0) Ciprofloxacin 24 (36) 0 (0) 4 (6) Trimethoprim/sulfamethoxazole 64 (96) 15 (47) 39 (56) Levofloxacin 15 (22) 0 (0) 0 (0) Aztreonam 56 (84) 0 (0) 0 (0) Imipenem 0 (0) 0 (0) 0 (0) Cefepime 47 (70) 0 (0) 0 (0) Meropenem 0 (0) 0 (0) 0 (0) Colistin* 1 (1) 4 (13) 1 (1) Ceftazidime 46 (69) 0 (0) 0 (0) Cefotaxime 64 (96) 0 (0) 0 (0) Ampicillin / sulbactam 2:1 ratio* 16 (24) 4 (13) 5 (7) Doripenem 0 (0) 0 (0) 0 (0) Piperacillin/ tazobactam constant 4 0 (0) 3 (9) 0 (0) Ticarcillin / clavulanic acid constant 2 7 (10) 6 (19) 0 (0)
Table 7: Number of isolates demonstrating reduced susceptibility
Table showing possible reduced susceptibility to the various antibiotics of the different isolates in numbers as well as in percentages * for Salmonella the MIC clinical breakpoint was used instead of ECOFF (ECOFF not available for salmonella)
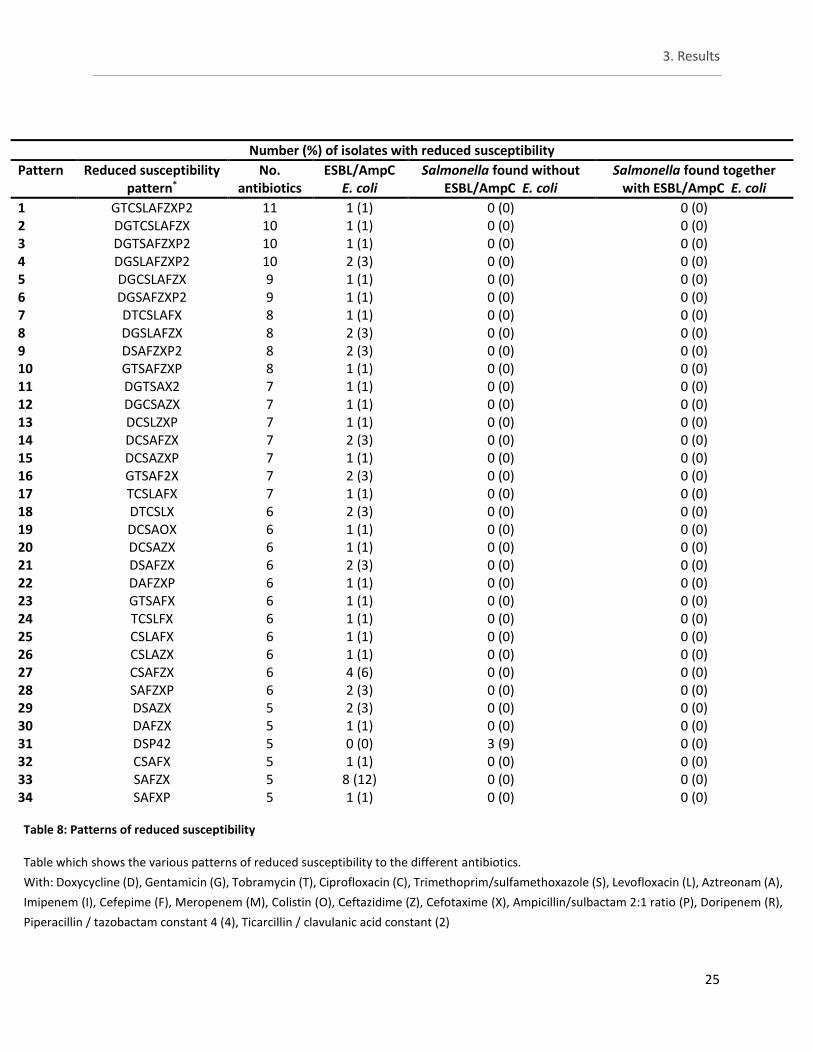
3. Results
25
Number (%) of isolates with reduced susceptibility
Pattern Reduced susceptibility pattern*
No. antibiotics
ESBL/AmpC E. coli
Salmonella found without ESBL/AmpC E. coli
Salmonella found together with ESBL/AmpC E. coli
1 GTCSLAFZXP2 11 1 (1) 0 (0) 0 (0) 2 DGTCSLAFZX 10 1 (1) 0 (0) 0 (0) 3 DGTSAFZXP2 10 1 (1) 0 (0) 0 (0) 4 DGSLAFZXP2 10 2 (3) 0 (0) 0 (0) 5 DGCSLAFZX 9 1 (1) 0 (0) 0 (0) 6 DGSAFZXP2 9 1 (1) 0 (0) 0 (0) 7 DTCSLAFX 8 1 (1) 0 (0) 0 (0) 8 DGSLAFZX 8 2 (3) 0 (0) 0 (0) 9 DSAFZXP2 8 2 (3) 0 (0) 0 (0) 10 GTSAFZXP 8 1 (1) 0 (0) 0 (0) 11 DGTSAX2 7 1 (1) 0 (0) 0 (0) 12 DGCSAZX 7 1 (1) 0 (0) 0 (0) 13 DCSLZXP 7 1 (1) 0 (0) 0 (0) 14 DCSAFZX 7 2 (3) 0 (0) 0 (0) 15 DCSAZXP 7 1 (1) 0 (0) 0 (0) 16 GTSAF2X 7 2 (3) 0 (0) 0 (0) 17 TCSLAFX 7 1 (1) 0 (0) 0 (0) 18 DTCSLX 6 2 (3) 0 (0) 0 (0) 19 DCSAOX 6 1 (1) 0 (0) 0 (0) 20 DCSAZX 6 1 (1) 0 (0) 0 (0) 21 DSAFZX 6 2 (3) 0 (0) 0 (0) 22 DAFZXP 6 1 (1) 0 (0) 0 (0) 23 GTSAFX 6 1 (1) 0 (0) 0 (0) 24 TCSLFX 6 1 (1) 0 (0) 0 (0) 25 CSLAFX 6 1 (1) 0 (0) 0 (0) 26 CSLAZX 6 1 (1) 0 (0) 0 (0) 27 CSAFZX 6 4 (6) 0 (0) 0 (0) 28 SAFZXP 6 2 (3) 0 (0) 0 (0) 29 DSAZX 5 2 (3) 0 (0) 0 (0) 30 DAFZX 5 1 (1) 0 (0) 0 (0) 31 DSP42 5 0 (0) 3 (9) 0 (0) 32 CSAFX 5 1 (1) 0 (0) 0 (0) 33 SAFZX 5 8 (12) 0 (0) 0 (0) 34 SAFXP 5 1 (1) 0 (0) 0 (0)
Table 8: Patterns of reduced susceptibility
Table which shows the various patterns of reduced susceptibility to the different antibiotics.
With: Doxycycline (D), Gentamicin (G), Tobramycin (T), Ciprofloxacin (C), Trimethoprim/sulfamethoxazole (S), Levofloxacin (L), Aztreonam (A),
Imipenem (I), Cefepime (F), Meropenem (M), Colistin (O), Ceftazidime (Z), Cefotaxime (X), Ampicillin/sulbactam 2:1 ratio (P), Doripenem (R),
Piperacillin / tazobactam constant 4 (4), Ticarcillin / clavulanic acid constant (2)
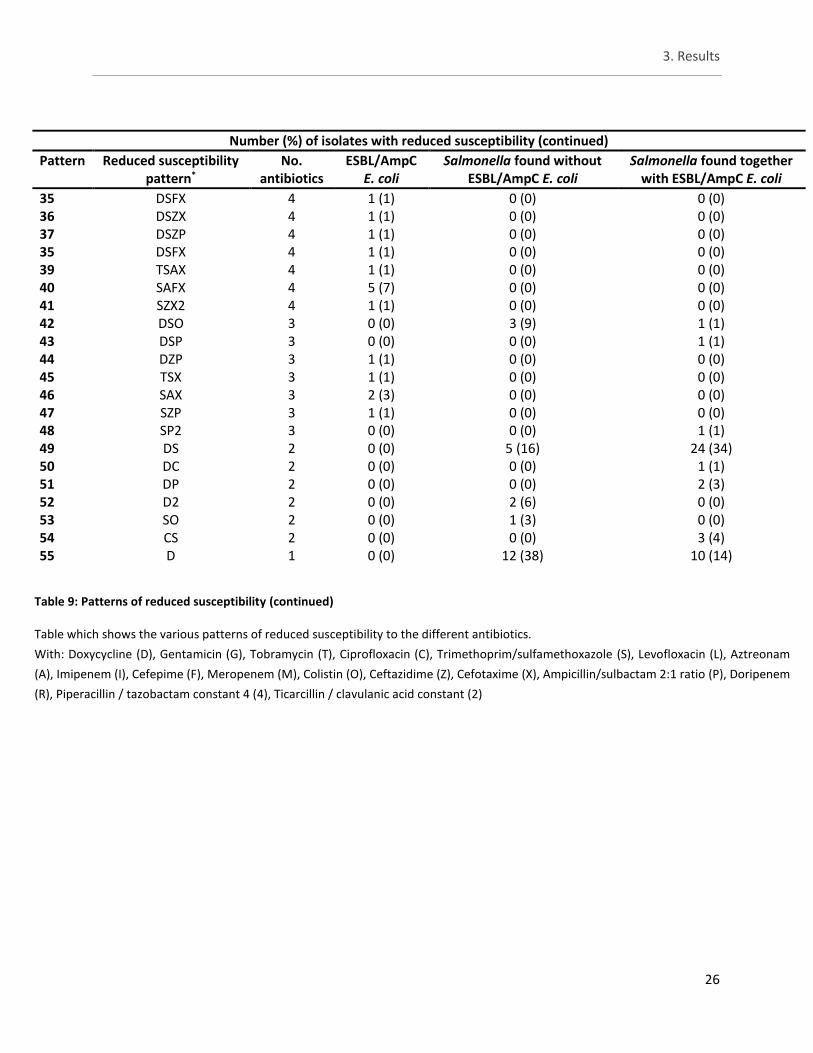
3. Results
26
Number (%) of isolates with reduced susceptibility (continued)
Pattern Reduced susceptibility pattern*
No. antibiotics
ESBL/AmpC E. coli
Salmonella found without ESBL/AmpC E. coli
Salmonella found together with ESBL/AmpC E. coli
35 DSFX 4 1 (1) 0 (0) 0 (0) 36 DSZX 4 1 (1) 0 (0) 0 (0) 37 DSZP 4 1 (1) 0 (0) 0 (0) 35 DSFX 4 1 (1) 0 (0) 0 (0) 39 TSAX 4 1 (1) 0 (0) 0 (0) 40 SAFX 4 5 (7) 0 (0) 0 (0) 41 SZX2 4 1 (1) 0 (0) 0 (0) 42 DSO 3 0 (0) 3 (9) 1 (1) 43 DSP 3 0 (0) 0 (0) 1 (1) 44 DZP 3 1 (1) 0 (0) 0 (0) 45 TSX 3 1 (1) 0 (0) 0 (0) 46 SAX 3 2 (3) 0 (0) 0 (0) 47 SZP 3 1 (1) 0 (0) 0 (0) 48 SP2 3 0 (0) 0 (0) 1 (1) 49 DS 2 0 (0) 5 (16) 24 (34) 50 DC 2 0 (0) 0 (0) 1 (1) 51 DP 2 0 (0) 0 (0) 2 (3) 52 D2 2 0 (0) 2 (6) 0 (0) 53 SO 2 0 (0) 1 (3) 0 (0) 54 CS 2 0 (0) 0 (0) 3 (4) 55 D 1 0 (0) 12 (38) 10 (14)
Table 9: Patterns of reduced susceptibility (continued)
Table which shows the various patterns of reduced susceptibility to the different antibiotics.
With: Doxycycline (D), Gentamicin (G), Tobramycin (T), Ciprofloxacin (C), Trimethoprim/sulfamethoxazole (S), Levofloxacin (L), Aztreonam
(A), Imipenem (I), Cefepime (F), Meropenem (M), Colistin (O), Ceftazidime (Z), Cefotaxime (X), Ampicillin/sulbactam 2:1 ratio (P), Doripenem
(R), Piperacillin / tazobactam constant 4 (4), Ticarcillin / clavulanic acid constant (2)
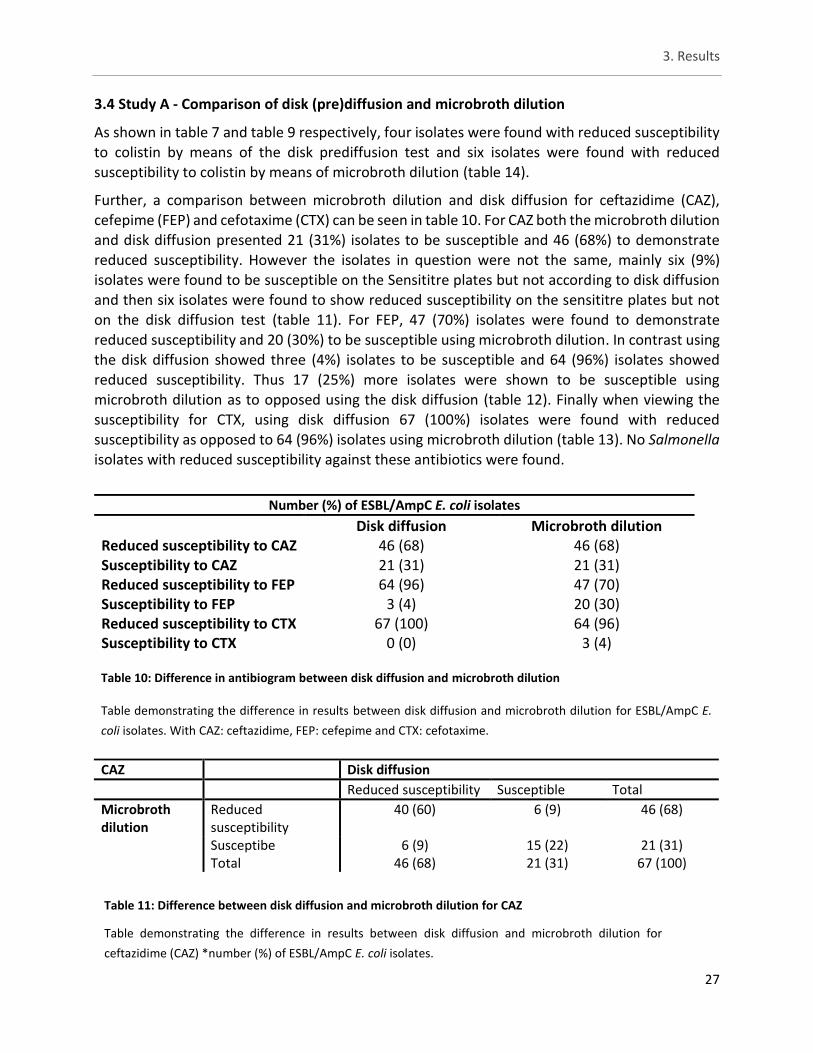
3. Results
27
3.4 Study A - Comparison of disk (pre)diffusion and microbroth dilution
As shown in table 7 and table 9 respectively, four isolates were found with reduced susceptibility to colistin by means of the disk prediffusion test and six isolates were found with reduced susceptibility to colistin by means of microbroth dilution (table 14).
Further, a comparison between microbroth dilution and disk diffusion for ceftazidime (CAZ), cefepime (FEP) and cefotaxime (CTX) can be seen in table 10. For CAZ both the microbroth dilution and disk diffusion presented 21 (31%) isolates to be susceptible and 46 (68%) to demonstrate reduced susceptibility. However the isolates in question were not the same, mainly six (9%) isolates were found to be susceptible on the Sensititre plates but not according to disk diffusion and then six isolates were found to show reduced susceptibility on the sensititre plates but not on the disk diffusion test (table 11). For FEP, 47 (70%) isolates were found to demonstrate
reduced susceptibility and 20 (30%) to be susceptible using microbroth dilution. In contrast using the disk diffusion showed three (4%) isolates to be susceptible and 64 (96%) isolates showed reduced susceptibility. Thus 17 (25%) more isolates were shown to be susceptible using microbroth dilution as to opposed using the disk diffusion (table 12). Finally when viewing the susceptibility for CTX, using disk diffusion 67 (100%) isolates were found with reduced susceptibility as opposed to 64 (96%) isolates using microbroth dilution (table 13). No Salmonella isolates with reduced susceptibility against these antibiotics were found.
CAZ Disk diffusion
Reduced susceptibility Susceptible Total
Microbroth dilution
Reduced susceptibility
40 (60) 6 (9) 46 (68)
Susceptibe 6 (9) 15 (22) 21 (31) Total 46 (68) 21 (31) 67 (100)
Number (%) of ESBL/AmpC E. coli isolates
Disk diffusion Microbroth dilution Reduced susceptibility to CAZ 46 (68) 46 (68) Susceptibility to CAZ 21 (31) 21 (31) Reduced susceptibility to FEP 64 (96) 47 (70) Susceptibility to FEP 3 (4) 20 (30) Reduced susceptibility to CTX 67 (100) 64 (96) Susceptibility to CTX 0 (0) 3 (4)
Table 10: Difference in antibiogram between disk diffusion and microbroth dilution
Table demonstrating the difference in results between disk diffusion and microbroth dilution for ESBL/AmpC E.
coli isolates. With CAZ: ceftazidime, FEP: cefepime and CTX: cefotaxime.
Table 11: Difference between disk diffusion and microbroth dilution for CAZ
Table demonstrating the difference in results between disk diffusion and microbroth dilution for
ceftazidime (CAZ) *number (%) of ESBL/AmpC E. coli isolates.
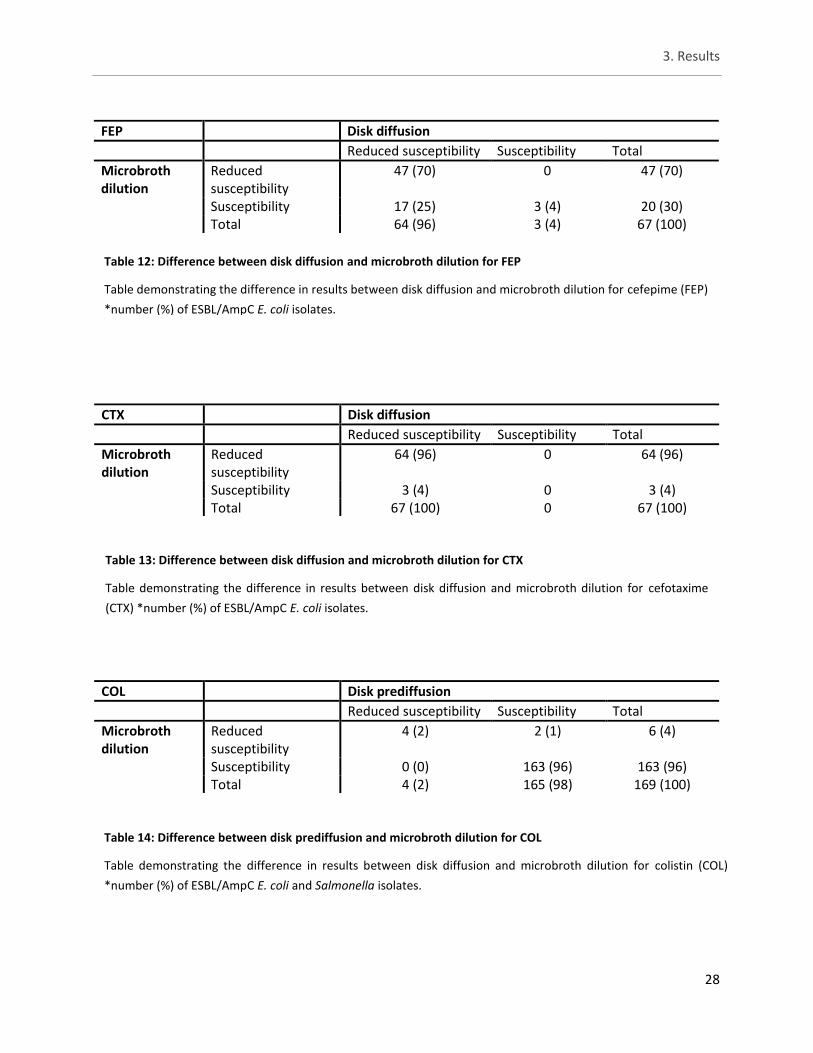
3. Results
28
FEP Disk diffusion
Reduced susceptibility Susceptibility Total
Microbroth dilution
Reduced susceptibility
47 (70) 0 47 (70)
Susceptibility 17 (25) 3 (4) 20 (30) Total 64 (96) 3 (4) 67 (100)
CTX Disk diffusion
Reduced susceptibility Susceptibility Total
Microbroth dilution
Reduced susceptibility
64 (96) 0 64 (96)
Susceptibility 3 (4) 0 3 (4) Total 67 (100) 0 67 (100)
COL Disk prediffusion
Reduced susceptibility Susceptibility Total
Microbroth dilution
Reduced susceptibility
4 (2) 2 (1) 6 (4)
Susceptibility 0 (0) 163 (96) 163 (96) Total 4 (2) 165 (98) 169 (100)
Table 12: Difference between disk diffusion and microbroth dilution for FEP
Table demonstrating the difference in results between disk diffusion and microbroth dilution for cefepime (FEP)
*number (%) of ESBL/AmpC E. coli isolates.
Table 13: Difference between disk diffusion and microbroth dilution for CTX
Table demonstrating the difference in results between disk diffusion and microbroth dilution for cefotaxime
(CTX) *number (%) of ESBL/AmpC E. coli isolates.
Table 14: Difference between disk prediffusion and microbroth dilution for COL
Table demonstrating the difference in results between disk diffusion and microbroth dilution for colistin (COL)
*number (%) of ESBL/AmpC E. coli and Salmonella isolates.
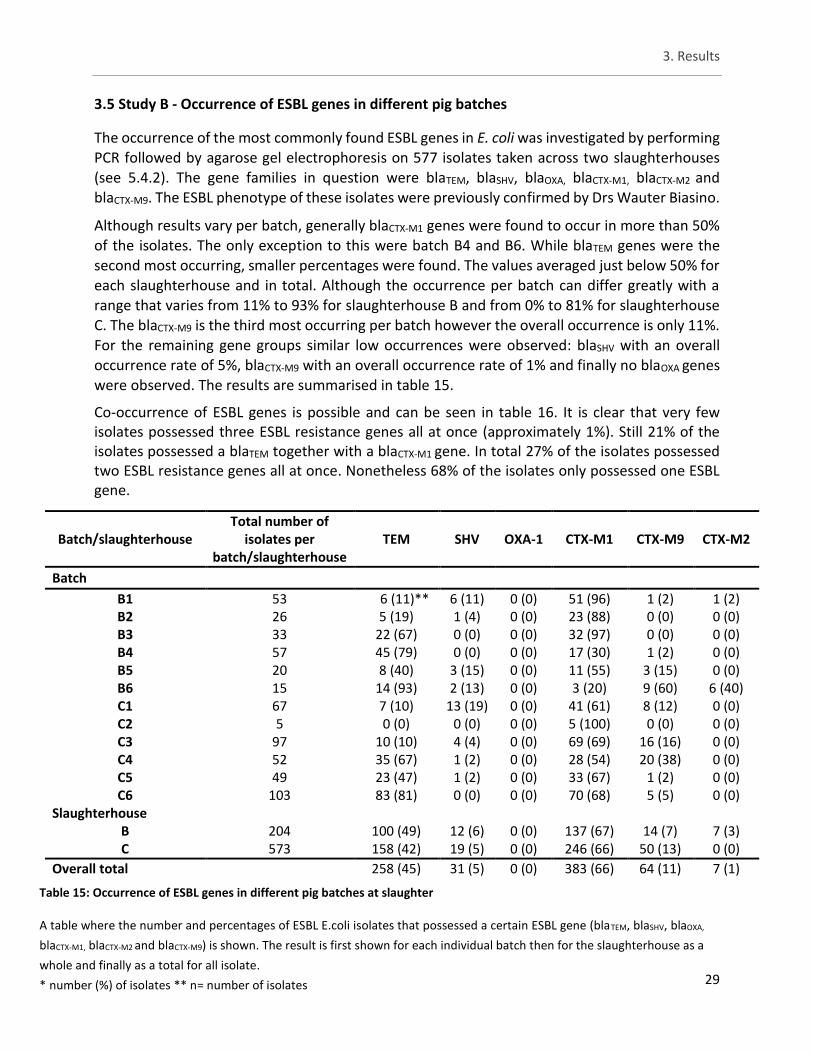
3. Results
29
3.5 Study B - Occurrence of ESBL genes in different pig batches
The occurrence of the most commonly found ESBL genes in E. coli was investigated by performing PCR followed by agarose gel electrophoresis on 577 isolates taken across two slaughterhouses (see 5.4.2). The gene families in question were blaTEM, blaSHV, blaOXA, blaCTX-M1, blaCTX-M2 and blaCTX-M9. The ESBL phenotype of these isolates were previously confirmed by Drs Wauter Biasino.
Although results vary per batch, generally blaCTX-M1 genes were found to occur in more than 50% of the isolates. The only exception to this were batch B4 and B6. While blaTEM genes were the second most occurring, smaller percentages were found. The values averaged just below 50% for each slaughterhouse and in total. Although the occurrence per batch can differ greatly with a range that varies from 11% to 93% for slaughterhouse B and from 0% to 81% for slaughterhouse C. The blaCTX-M9 is the third most occurring per batch however the overall occurrence is only 11%.
For the remaining gene groups similar low occurrences were observed: blaSHV with an overall occurrence rate of 5%, blaCTX-M9 with an overall occurrence rate of 1% and finally no blaOXA genes were observed. The results are summarised in table 15.
Co-occurrence of ESBL genes is possible and can be seen in table 16. It is clear that very few isolates possessed three ESBL resistance genes all at once (approximately 1%). Still 21% of the isolates possessed a blaTEM together with a blaCTX-M1 gene. In total 27% of the isolates possessed two ESBL resistance genes all at once. Nonetheless 68% of the isolates only possessed one ESBL gene.
Batch/slaughterhouse Total number of
isolates per batch/slaughterhouse
TEM SHV OXA-1 CTX-M1 CTX-M9 CTX-M2
Batch
B1 53 6 (11)** 6 (11) 0 (0) 51 (96) 1 (2) 1 (2) B2 26 5 (19) 1 (4) 0 (0) 23 (88) 0 (0) 0 (0) B3 33 22 (67) 0 (0) 0 (0) 32 (97) 0 (0) 0 (0) B4 57 45 (79) 0 (0) 0 (0) 17 (30) 1 (2) 0 (0) B5 20 8 (40) 3 (15) 0 (0) 11 (55) 3 (15) 0 (0) B6 15 14 (93) 2 (13) 0 (0) 3 (20) 9 (60) 6 (40) C1 67 7 (10) 13 (19) 0 (0) 41 (61) 8 (12) 0 (0) C2 5 0 (0) 0 (0) 0 (0) 5 (100) 0 (0) 0 (0) C3 97 10 (10) 4 (4) 0 (0) 69 (69) 16 (16) 0 (0) C4 52 35 (67) 1 (2) 0 (0) 28 (54) 20 (38) 0 (0) C5 49 23 (47) 1 (2) 0 (0) 33 (67) 1 (2) 0 (0) C6 103 83 (81) 0 (0) 0 (0) 70 (68) 5 (5) 0 (0)
Slaughterhouse B 204 100 (49) 12 (6) 0 (0) 137 (67) 14 (7) 7 (3) C 573 158 (42) 19 (5) 0 (0) 246 (66) 50 (13) 0 (0)
Overall total 258 (45) 31 (5) 0 (0) 383 (66) 64 (11) 7 (1)
Table 15: Occurrence of ESBL genes in different pig batches at slaughter
A table where the number and percentages of ESBL E.coli isolates that possessed a certain ESBL gene (blaTEM, blaSHV, blaOXA,
blaCTX-M1, blaCTX-M2 and blaCTX-M9) is shown. The result is first shown for each individual batch then for the slaughterhouse as a
whole and finally as a total for all isolate.
* number (%) of isolates ** n= number of isolates
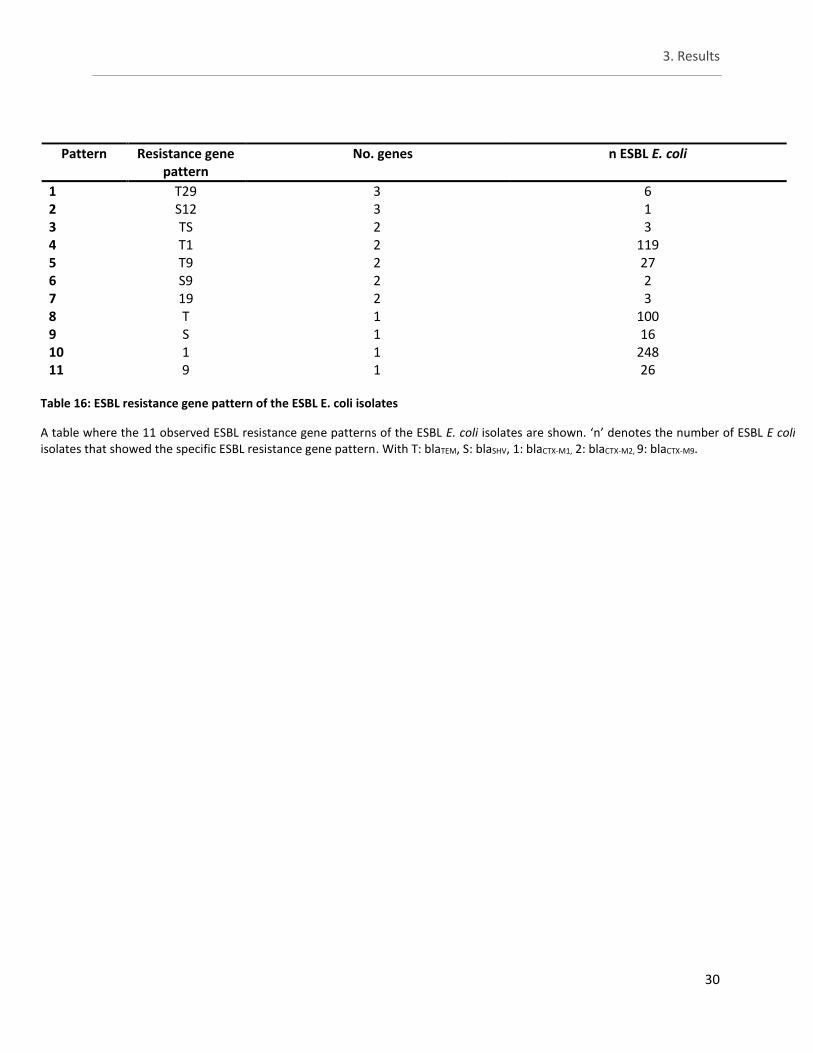
3. Results
30
Pattern Resistance gene pattern
No. genes n ESBL E. coli
1 T29 3 6 2 S12 3 1 3 TS 2 3 4 T1 2 119 5 T9 2 27 6 S9 2 2 7 19 2 3 8 T 1 100 9 S 1 16 10 1 1 248 11 9 1 26
Table 16: ESBL resistance gene pattern of the ESBL E. coli isolates
A table where the 11 observed ESBL resistance gene patterns of the ESBL E. coli isolates are shown. ‘n’ denotes the number of ESBL E coli isolates that showed the specific ESBL resistance gene pattern. With T: blaTEM, S: blaSHV, 1: blaCTX-M1, 2: blaCTX-M2, 9: blaCTX-M9.
and finally as a total for all isolate

4. Discussion
31
4. Discussion
4.1 ESBL/AmpC characterisation of the isolates
Selection of isolates was based on their ability to grow on TBX supplemented with cefotaxime which is already indicative for an ESBL/AmpC phenotype. Clearly in reality non-ESBL/AmpC producing E. coli are present and generally outnumber the ESBL/AmpC-producing E. coli population. To illustrate this, in a study conducted by Xu et al (2015) E. coli isolates taken from piglets with post-weaning diarrhoea were screened and of these isolates 43.2% (198/458) were ESBL producers (Xu et al, 2015). Furthermore in healthy pigs 12.6% (41/326) of E. coli isolates were found to be ESBL producing (Xu et al, 2015; Zheng et al, 2012).
There is a general lack of reduced susceptibility within the two Salmonella groups. Furthermore, almost no difference between the two Salmonella groups was observed. Undoubtedly, ESBL/AmpC E. coli isolates showed a variety of reduced susceptibility. The genotype and phenotype of the Salmonella isolates co-occurring with ESBL/AmpC E. coli differ from the ESBL/AmpC E. coli which adds further evidence that seemingly no reduced susceptibility is transferred between the groups. Although plasmid-mediated resistance transfer between E. coli and Salmonella has been demonstrated (Vaidya & Manzoor, 2009). Despite no Salmonella isolates presenting a ESBL phenotype, a majority was shown to possess resistance genes. Remarkably, these genes were different than those found in ESBL E. coli making transfer from E. coli unlikely. Regardless of the manner of acquisition, possible reasons for the lack of ESBL phenotype include: pseudogenes, low expression levels or a promotor region might be missing. In the literature discrepancy between phenotype and genotype is also observed albeit not quite large and reasons include intermediate resistance and loss of phenotype by an environment with low antibiotic usage (Davis et al, 2011; McDermott et al, 2016). While pseudogenes are observed within Salmonella no immediate antibiotic resistance genes were identified (Tariq et al, 2016; Holt et al, 2009).
The low occurrence of ESBL/AmpC Salmonella could be attributed to their low prevalence in pigs in Europe. According to an EFSA report in 2015 out of 443 isolates taken from pork: a prevalence of 0.9% was reported for ESBL Salmonella (two from Germany, one from Belgium and one from Italy), a prevalence of 0.7% was reported for AmpC Salmonella (two from Portugal and one from Italy) and finally a prevalence of 0.2% was reported for ESBL + AmpC Salmonella (one isolate form the Czech Republic) (EFSA, 2017). Because of this not much can be said about which ESBL/AmpC genes occur the most in Salmonella. But in comparison a study conducted in the Netherlands regarding ESBL Salmonella in poultry and human patients demonstrated that the majority (19/34) ESBL Salmonella carried a TEM gene, other genes found were CTX-M (6/34), ACC (9/34) and SHV (3/34) (Hasman et al, 2005).
For ESBL/AmpC E. coli there was a 85% agreement between phenotype and genotype. As for ESBL E. coli isolates that presented the ESBL phenotype but not a corresponding genotype, other ESBL genes such as PER-, GES-type also exist and might be the reason for the ESBL phenotypes that were not confirmed by the PCR (Shaikh et al, 2015; Paterson & Bonomo, 2005).

4. Discussion
32
The same can be said for AmpC genes where AmpC genes such as LAT and BIL were not included
in the PCR (Pérez-Pérez & Hanson, 2002).
In accordance with the literature the most common types of ESBL genes encountered (study A & study B) belonged either to the blaTEM or the blaCTX-M family (Stürenburg & Mack, 2003; Paterson & Bonomo, 2005). This was regardless of the slaughterhouse of origin. Since some isolates possess multiple ESBL resistance genes, reduced susceptibility to β-lactamases will be retained even if an event causes one of the genes to become faulty. This could be one of the ways how resistance persists. The occurrence of ESBL E. coli with multiple ESBL resistance gene is not uncommon. In a study by Seyedjavadi et al (2016), 45 (45%) out of 100 ESBL E. coli isolates carried more than one type of β-lactamase genes. Co-existence of blaCTX-M and blaTEM was detected in 22 isolates (22%), blaCTX-M and blaSHV in 12 isolates (12%), blaTEM and blaSHV in 8 isolates (8%) and blaCTX-M, blaSHV and
blaTEM in three isolates (3%)(Seyedjavadi et al, 2016). These findings are comparable to those found in this study. Furthermore, transcriptional response of a certain ESBL gene is dependent on
the type of antibiotic used (Maurya et al, 2017). Greater occurrence of the ESBL phenotype in comparison to the AmpC phenotype is similar to the finding of a German study also conducted in pigs and for E. coli. In this study consisting of 210 ESBL E. coli isolates: 89% showed an ESBL phenotype and 11% an AmpC phenotype, there was no mention of ESBL co-occurring with AmpC (Von Salviati et al, 2014).
4.2 Study A - Colistin susceptibility of ESBL/AmpC-producing E. coli and Salmonella
Though reduced susceptibility to colistin was observed in few pigs, this seemed to occur with no evident trend. Furthermore no evidence for colistin plasmid-mediated reduced susceptibility was
found in this study. Given that in a previous study conducted in Belgium, concerning 105 colistin resistant Salmonella isolates from various sources, only two isolates originating from pigs were found to possess colistin plasmid-mediated resistance (Garcia-Graells et al, 2016). In light of this, it could be said that our results are reflective of current literature. A possible explanation for chromosomal reduced susceptibility in Salmonella are missense mutations located in the pmrA and pmrB gene (encoding the regulator and sensor of a two-component regulatory system) (Sun et al, 2009). Although only very few isolates with reduced susceptibility to colistin were found in this study; a mutation rate to colistin resistance in Salmonella has been reported to be 0.6 X 106 per cell generation (Sun et al, 2009; Morales et al, 2012). As for colistin plasmid mediated resistance, in a study by Anjum et al (2016) plasmid transfer by conjugation from Salmonella to recipient Enterobacteriaceae was low, lying between 10-7 and 10-9 cfu/recipient
(Anjum et al, 2016). Co-occurrence of mcr-1 and ESBL/AmpC genes on a single plasmid has been demonstrated in Salmonella (Figueiredo et al, 2016; Yang et al, 2016; Doumith et al, 2016).
Likewise for E. coli co-occurrence of mcr-1 and ESBL genes on a single plasmid has been reported (Haenni et al, 2016a; Brauer et al, 2016). Chromosomal colistin resistance has also been reported in ESBL E. coli, not only mutations in the pmrAB gene but also in the PhoPQ gene can be responsible for colistin resistance in E. coli (Anjum et al, 2016; Nordmann et al, 2016).

4. Discussion
33
Although in this study only one E. coli with reduced susceptibility to colistin was found; a study in
Germany conducted in 2015 showed a 2.1% (out of 512 isolates) prevalence of colistin resistance in pigs at farm level however the same study did not find any colistin resistant E. coli in isolates originating from pig slaughterhouses (Irrgang et al, 2016). Similarly in France the prevalence of colistin resistant E. coli was reported to be at 0.5% (out of 200 strains) in 2011 (Perrin-Guyomard et al, 2016). Therefore the low prevalence of colistin resistant E. coli in pigs might be the reason why only one ESBL E. coli with reduced susceptibility to colistin was found in this study.
All Salmonella and E. coli isolates were investigated for the presence of ESBL/AmpC genes regardless of phenotype. Whereas, the screening for mcr-1/2 genes was only performed on isolates with a suggestive phenotype. The main reason for this is the low prevalence of plasmid-mediated colistin resistance in pigs(Garcia-Graells et al, 2016; Perrin-Guyomard et al, 2016;
Irrgang et al, 2016). Therefore it seemed more interesting to focus on the ESBL/AmpC gene profiles and the possible co-occurrence in E. coli and Salmonella.
4.3 Study A - Phenotypical testing by means of microbroth dilution
Co-occurrence of reduced susceptibility was observed and this notably for ESBL E. coli together with a reduced susceptibility to Trimethoprim/Sulfamethoxazole (SXT) and/or DOX. This type of co-occurrence is well documented to take place with rates against both being approximately 80% (Anago et al, 2015; Winokur et al, 2001; Lagacé-wiens et al, 2007).
The Salmonella isolates from both groups showed reduced susceptibility to only a few antibiotics, mostly against SXT and/or DOX. In terms of possible ß-lactamase production, the observed reduced susceptibility against a few penicillins (ampicillin/sulbactam 2:1 ratio,
piperacillin/tazobactam constant 4, ticarcillin/clavulanic acid constant 2) is of importance. However, this is not sufficiently to qualify an isolate as belonging to the ESBL/AmpC phenotype.
Because the number of isolates of both groups vary as well as the number of isolates per batch, no definitive statements will be made. Yet, it can be said that both Salmonella groups give comparable results or at least no correlation with ESBL/AmpC E. coli strains seem likely. The observed reduced susceptibility among the Salmonella isolates has already been reported to occur in the EU. An EFSA report regarding data from 2015 states that Salmonella isolates from pig meat show overall resistance levels of 44.7%, 48.5% and 49.1% to ampicillin, sulfamethoxazole and tetracyclines respectively (EFSA, 2017).
No reduced susceptibility against carbapenems were observed in this study. The most reasonable
explanation for this is that carbapenems are not registered for use in veterinary medicine, expect for use in specific circumstances in companion animals or horses when dealing with multidrug-resistant Enterobacteriaceae. Nevertheless this usage, at least in developed countries, remains rare (Poirel et al, 2014; EFSA, 2013).

4. Discussion
34
4.4 Study A - Comparison of disk (pre)diffusion and microbroth dilution
Microbroth dilution is a well-standardized reference method for antimicrobial susceptibility testing. A major advantage of using this method is that it allows convenient simultaneous testing of individual isolates with multiple antibiotics. Since few laboratories have the facilities required for preparation of broth microbroth dilution plates, several sources of commercially prepared antibiotic agent panels are available. The plates, consisting of wells containing prepared antibiotic dilutions, are provided either frozen or freeze-dried. Frozen plates must be stored at least at -22°C, whereas dried panels can be stored at room temperature. The plates can be inoculated with multichannel pipettors or using the multipoint inoculating devices accompanying the plates. This method allows results to be determined by visual examination or by use of semi-automated or automated instrumentation. Growth is best determined by comparison with that in the growth
control well and generally is indicated by turbidity throughout the well or by buttons, single or multiple, in the well bottom. Results for drugs with more than one skipped well should not be
reported (Patel et al, 2011).
The disk diffusion method of susceptibility testing allows categorization of most bacterial isolates as susceptible, intermediate, or resistant to a variety of antimicrobial agents (Patel et al, 2011). The resulting inhibition zones can be measured manually or by automated zone size reading devices (Nijs et al, 2003). The disk diffusion test has several advantages: (i) it is technically simple to perform and (ii) very reproducible, (iii) the reagents are relatively inexpensive, (iv) it does not require any special equipment, (v) it provides susceptibility category results that are easily understood by clinicians, and (vi) it is flexible regarding selection of antimicrobial agents for testing. Potential disadvantage of disk diffusion susceptibility testing is that it provides only a
qualitative result and a quantitative result indicating that the degree of susceptibility (MIC) may be needed in some cases. Additionally, a limitation of the disk diffusion test is the spectrum of organisms for which it has been standardized.
Finally, the disk prediffusion method for detecting reduced susceptibility to colistin, as described by Boyen et al, (2010) is performed in a similar manner as one would perform disk diffusion. However, it has not been validated to determine colistin resistance. The results in this study show a 33% difference between the two methods, albeit four isolates with disk prediffusion versus six isolates with microbroth dilution. Admittedly, the sample size in this study is insufficient for definitive statements. On the other hand, current literature suggest that disk prediffusion provides reliable information on colistin susceptibility. A study by Boyen et al, (2010) demonstrated a 96.8% (ot of 157 isolates) agreement between disk prediffusion and E-test with
agar dilution. Therefore it could be said that disk prediffusion could be used a reliable, cheap, alternative agar-based colistin susceptibility testing method to monitor the emergence of colistin resistance (Boyen et al, 2010).
When comparing these different types of tests, the following can be said: microbroth dilution plates contain a built in separate control (well not containing an antibiotic) while disk (pre)diffusion has zones where no antibiotic has diffused through the agar. Both microbroth dilution and the disk (pre)diffusion test required four days when taken the time period from

4. Discussion
35
growing isolates from stock till obtaining results. In this study both methods required some
preparatory work such as making medium but no significant labour difference can be stated both from this preparatory work as well as during performing the test. However when it comes to gathering the results in this study the ‘reading’ of microbroth dilution plates was quicker since the disk diffusion required manually measuring the different zone diameters for the different antibiotics. In contrast disk prediffusion could be measured faster since only one zone diameter had to be measured. Although, an advantage of microbroth dilution is that it can contain the antibiotics for ESBL/AmpC determination in addition to colistin while disk diffusion and disk prediffusion had to be performed on separate plates resulting in much more labour time. For both microbroth dilution and disk (pre)diffusion test it can be said that when manually collecting the results, the tests are liable to small subjective errors meaning that determining growth or measuring an inhibition zone might slightly deviate from the genuine result and results may vary
depending on the person determining growth or measuring the inhibition zone. Despite the added labour time, less costs are coupled to performing disk (pre)diffusion than performing
microbroth dilution mainly due to costs of commercial (custom) made microdilution plates. In conclusion the microbroth dilution shares most of the advantages described for disk diffusion in addition to allowing higher degree of simultaneous testing to various antibiotics but is a more expensive test.
Regarding the difference of the categorization of susceptible and reduced susceptible isolates by microbroth dilution and the disk (pre)diffusion test, it can be said that apart for the results of reduced susceptibility to FEP, the results were comparable. This is confirmed by a study performed by Skov et al, (2001) which stated good correlation between microbroth dilution and
disk diffusion (Skov et al, 2001).
4.5 Future prospects
Although plasmid-mediated resistances genes were of interest, the screening of ESBL/AmpC genes by means of PCR was performed with primers that could detect both plasmid-mediated genes as well as chromosomal genes. As plasmid-mediated antibiotic resistance can be passed on between bacterial strains which can be either distantly or closely related (CDC, 2013; Frost et al, 2005), a plasmid purification should be performed followed by screening for the presence of resistance genes. Of course even in the absence of plasmid-mediated resistance, the danger of horizontal transfer of resistance still exist by means of either transduction via bacteriophages or transposition via integrons (Frost et al, 2005).
Besides the confirmation of plasmid-mediated resistance genes also the transfer of resistance between E. coli and Salmonella and other pathogenic Enterobacteriaceae such as Yersinia should be investigated as this could be indicative for future problems concerning food safety. Admittedly no direct evidence of transfer between ESBL E. coli and Salmonella was discovered in this study. Nevertheless this should be thoroughly investigated due to the importance regarding food safety. This should be combined with plasmid extraction for confirmation of plasmid-mediated resistance, partly to exclude other resistance mechanisms such as mutation.

4. Discussion
36
This study concludes that there is no substantial threat regarding ESBL Salmonella in Belgian pork.
Furthermore ESBL E. coli does not seem like an indicator for the presence of Salmonella with reduced susceptibility, since no Salmonella were found with reduced susceptibility despite of co-occurrence with ESBL E. coli. However, a reservoir of antibiotic resistance genes remains within E. coli found in Belgian pigs.
The efficiency of penicillins, cephalosporins and colistin need to be protected. This could be achieved through further epidemiological surveillance and investigation combined with resistance characteristics research which will result in improved appropriate control measures for limiting the spread of resistant bacteria (PAHO/WHO, 2016; ECDC, 2016; BelVetSAC, 2015).
Alternatives to antibiotics should be investigated such as the use of zinc oxide in the treatment of post weaning diarrhoea in piglets opposed to colistin has reduced the use of colistin in Belgium
(BelVetSAC, 2015). Other alternatives in the future may include silver nanoparticles and capsaicin as they have been shown to inhibit the growth of MDR-ESBL producing pathogen E. coli. Since
these alternatives might also be effective against antibiotic resistant bacteria, research involving in vivo study with combination therapy is suggested (Kar et al, 2016).
Another example of alternatives to antibiotics are feeding supplements. Studies performed in Ireland involving such feeding supplements to lower Salmonella carriage and seroprevalence were carried out. One study involved strategic feeding of sodium butyrate to finishing pigs for a relatively short period of time (< 30 days) immediately prior to slaughter was effective in reducing Salmonella shedding and seroprevalence in one of two trials. Lack of efficacy in the second trial could be explained by concomitant infection with Lawsonia intracellularis (Walia et al, 2016). Likewise formic acid, citric acid and essential oils, have the potential to prevent increased
Salmonella shedding at certain time points and seroprevalence (Walia et al, 2017).
Despite showing an effect all these food additives did not reduce intestinal carriage, nor did they reduce seroprevalence to below the cut-off used for the high Salmonella risk category in Ireland (50%) (Walia et al, 2016, 2017). Nonetheless the potential of these agents to lower use of antibiotic warrant further research and, possibly, a right combination of these supplements might prove to be the solution.
Apart from alternatives, prevention or more correctly reduction of (resistant) food pathogens should be attempted. For example, as transport increases the Salmonella isolation rate; more stringent cleaning procedures of transport trucks, including the use of a disinfectant, should be warranted. This especially when transporting pigs from high-risk herds so as to further reduce the
potential for contamination during the slaughter process. Additionally, to reduce the risk for cross-contamination between pigs from different herds, trucks should not be permitted to transport pigs from high- and low-risk herds on the same day (Mannion et al, 2008).
In conclusion, the effectiveness of the different types of antibiotics needs to be protected mainly through epidemiological surveillance and a reduction in the use of antibiotics in veterinarian healthcare primarily by reducing prophylactic use. Nonetheless, correct use in human medicine is also warranted. Alternatives to antibiotics might prove to be useful by reducing the selective burden of bacteria towards antibiotics.

5. Materials and Methods
37
5. Materials and Methods
Salmonella and ESBL/AmpC-producing E. coli strains have been previously isolated from tonsils and rectal content of slaughterhouse pigs. A large part of the ESBL/AmpC-producing E. coli strains had already been subjected to phenotypic antibiotic resistance confirmation tests. These strains were investigated for the presence of ESBL resistance genes using PCR followed by agarose gel electrophoresis. The other part of ESBL/AmpC-producing E. coli strains had to be tested for ESBL/AmpC production but were previously isolated using Tryptone Bile Agar with X-Glucuronide (TBX; Bio-Rad, Marnes-la-Coquette, France) medium supplemented with cefotaxime (1 mg/mL). These strains were further investigated during this study together with Salmonella strains isolated from the same pigs.
5.1 Isolation of ESBL/AmpC-producing E. coli and Salmonella
The isolation of ESBL/AmpC-producing E. coli and Salmonella from tonsils and rectal content of
slaughterhouse pig were for the largest part already performed in function of research carried out by Dr. Inge Van Damme and Drs. Wauter Biasino. The isolates date from 2010 till 2016. The procedure used in this study was similar as described by Van Damme et al. (2017). The slaughterhouses were located in Flanders (northern part of Belgium), where over 90% of Belgian pig production is located (Van Damme et al, 2017). The intestinal tract of the pigs were collected immediately after evisceration and the rectum was closed with threads, followed by excision of the rectum and transfer into a plastic bag. The tonsils were aseptically excised after removal of the pluck, after which the tonsils were put in a sterile plastic bag. All samples were transported to the laboratory under refrigerated conditions and processed within 2 h after sampling. The
tonsillar tissue and rectal content were homogenized in Buffered Peptone Water (BPW; Bio-Rad, Marnes-la-Coquette, France) (1:10 w:w) using a stomacher apparatus (Colworth Stomacher 400, Seward Ltd, London, UK).
For isolation of ESBL/AmpC-producing E. coli 1 mL of the homogenate is taken and plated on TBX, supplemented with cefotaxime (1 mg/mL). The plates were then incubated aerobically at 44 °C for 24 h.
The isolation of Salmonella was based on ISO 6579:2002/Amd 1:2007 (Annex D). A pre-enrichment was carried out by placing the homogenate at 37 °C for 18 h. After pre-enrichment, 0.1 mL of the broth dispersed in three drops) was transferred onto Modified Semi Solid Rappaport Vassiliadis medium (MSRV; LabM, Lancashire, UK) and incubated at 41.5°C for 24 h. For MSRV
plates showing a migration zone, a subcultivation of the migration zone was done on an Xylose Lysine Deoxycholate (XLD) agar (Bio-Rad, Marnes-la-Coquette, France) plate, which was incubated at 37 °C for 24 h. One or two suspected colonies per plate were tested and confirmed biochemically (triple sugar iron, indole reaction and lysine decarboxylase test).
After this both ESBL/AmpC-producing E. coli and Salmonella were grown in Trypticase Soy Broth (TSB; Bio-Rad, Marnes-la-Coquette, France) before being stored in glycerol at -20 °C.
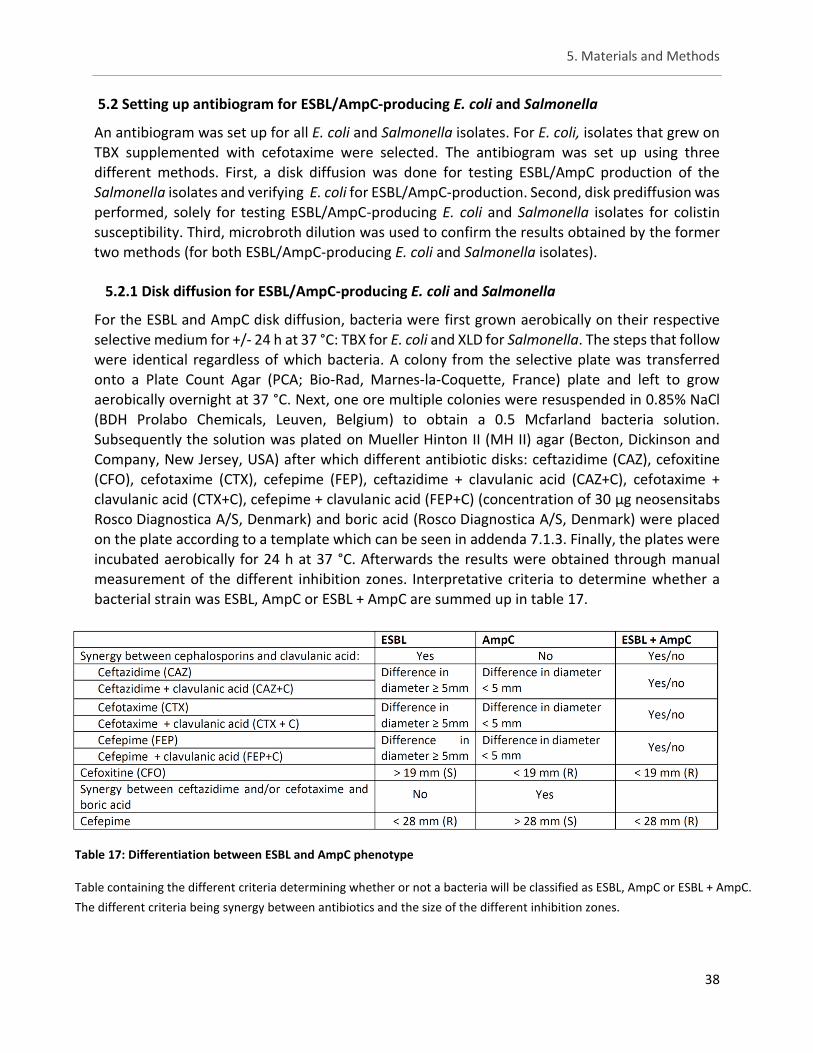
5. Materials and Methods
38
5.2 Setting up antibiogram for ESBL/AmpC-producing E. coli and Salmonella
An antibiogram was set up for all E. coli and Salmonella isolates. For E. coli, isolates that grew on TBX supplemented with cefotaxime were selected. The antibiogram was set up using three different methods. First, a disk diffusion was done for testing ESBL/AmpC production of the Salmonella isolates and verifying E. coli for ESBL/AmpC-production. Second, disk prediffusion was performed, solely for testing ESBL/AmpC-producing E. coli and Salmonella isolates for colistin susceptibility. Third, microbroth dilution was used to confirm the results obtained by the former two methods (for both ESBL/AmpC-producing E. coli and Salmonella isolates).
5.2.1 Disk diffusion for ESBL/AmpC-producing E. coli and Salmonella
For the ESBL and AmpC disk diffusion, bacteria were first grown aerobically on their respective
selective medium for +/- 24 h at 37 °C: TBX for E. coli and XLD for Salmonella. The steps that follow were identical regardless of which bacteria. A colony from the selective plate was transferred onto a Plate Count Agar (PCA; Bio-Rad, Marnes-la-Coquette, France) plate and left to grow aerobically overnight at 37 °C. Next, one ore multiple colonies were resuspended in 0.85% NaCl (BDH Prolabo Chemicals, Leuven, Belgium) to obtain a 0.5 Mcfarland bacteria solution. Subsequently the solution was plated on Mueller Hinton II (MH II) agar (Becton, Dickinson and Company, New Jersey, USA) after which different antibiotic disks: ceftazidime (CAZ), cefoxitine (CFO), cefotaxime (CTX), cefepime (FEP), ceftazidime + clavulanic acid (CAZ+C), cefotaxime + clavulanic acid (CTX+C), cefepime + clavulanic acid (FEP+C) (concentration of 30 µg neosensitabs Rosco Diagnostica A/S, Denmark) and boric acid (Rosco Diagnostica A/S, Denmark) were placed on the plate according to a template which can be seen in addenda 7.1.3. Finally, the plates were
incubated aerobically for 24 h at 37 °C. Afterwards the results were obtained through manual measurement of the different inhibition zones. Interpretative criteria to determine whether a bacterial strain was ESBL, AmpC or ESBL + AmpC are summed up in table 17.
Table 17: Differentiation between ESBL and AmpC phenotype
Table containing the different criteria determining whether or not a bacteria will be classified as ESBL, AmpC or ESBL + AmpC.
The different criteria being synergy between antibiotics and the size of the different inhibition zones.

5. Materials and Methods
39
5.2.2 Disk prediffusion for testing colistin susceptibility of ESBL/AmpC-producing E. coli and Salmonella
A 2 h + 18 h disk prediffusion test was performed with Neosensitabs tablets (10 µg, Rosco Diagnostica A/S, Denmark) according to CLSI-guide-lines (CLSI, 2008). The protocol was taken from Boyen et al. (2010). The colistin containing tablets (10 mg, Rosco, Denmark) were placed on uninoculated MH II agar plates. After 2 h incubation at 22 °C (room temperature) the disks were removed. The plates were further kept at 22 °C for 18 h. Subsequently, the plates were inoculated with the different strains using a 0.5 McFarland inoculum as described for the disk diffusion test and thereafter the plates were incubated aerobically overnight at 35 °C. Interpretative criteria to determine clinical resistance were based upon clinical breakpoints as described by the manufacturer and Boyen et al. (2010) (sensitive ≥ 20mm; intermediate 17–19 mm; resistant ≤16
mm). Growth inhibition zone diameters were measured manually.
5.2.3 Microbroth dilution for ESBL/AmpC-producing E. coli and Salmonella
Microbroth dilution was performed mainly to evaluate colistin susceptibility as recommend by EUCAST (EUCAST, 2017) but could also be used to evaluate ESBL and/or AmpC production. SensititreTM 96-well plates (plate code: GNX3F, Thermo Fisher Scientific, USA) were used and contained the following antibiotics (all at different concentration , see plate design in addenda): amikacin (AMI), tigecycline (TGC), doxycycline (DOX), minocycline (MIN), gentamicin (GEN), tobramycin (TOB), ciprofloxacin (CIP), trimethoprim/sulfamethoxazole (SXT), levofloxacin (LEVO), aztreonam (AZT), imipenem (IMI), cefepime (FEP), meropenem (MERO), colistin (COL), polymixin B (POL), ceftazidime (CAZ), cefotaxime (CTX), ampicillin/sulbactam 2:1 ratio (A/S2), doripenem
(DOR), piperacillin / tazobactam constant 4 (P/T4), ticarcillin / clavulanic acid constant 2 (TIM2) and three wells were left as positive control.
Microbroth dilution was performed as follows, a 0.5 McFarland solution was obtained by following the same steps as described for the disk diffusion test. Subsequently, MH II broth (Becton, Dickinson and Company, New Jersey, USA) was inoculated with 10 µL of the 0.5 McFarland solution. A volume of 50 µL MH II broth was added to each well and the 96-well plates
were incubated aerobically for approximately 18 h at 37 °C. Finally, the MIC was determined as the lowest concentration that inhibited visible growth. A strain was considered to possess reduced susceptibility by using the ECOFF values for E.coli or Salmonella for the different antibiotics used. The data was taken from the EUCAST MIC distribution website, last accessed 27/03/2017 (http://www.eucast.org). The antibiotic susceptibilities that were of interest in this
study were: AZT, IMI, CEF, MERO, COL, POL, CAZ, CTX, A/S2, DOR, P/T4 and TIM2. Nevertheless, all values were documented and kept for potential future studies.

5. Materials and Methods
40
5.3 Detection of resistance genes
PCR was used to verify for the presence of resistance genes responsible for either ESBL or AmpC production or colistin resistance. During this study, PCR reactions were aimed to detect plasmid-mediated resistance genes. The detection of resistance genes can be split into three parts: DNA extraction, performing the PCR and running gel electrophoresis. The PCR reactions for detection of ESBL/AmpC genes were based on the protocol described by Dallenne et al. (2010) and the PCR reaction for detection of plasmid-mediated colistin genes were based on the protocol described by Cavaco et al. (2016).
5.3.1 DNA extraction of ESBL/AmpC-producing E. coli and Salmonella
Salmonella or ESBL/AmpC-producing E. coli isolates were taken from the glycerol stock and first grown aerobically on their respective selective medium for +/- 24 h at 37 °C: TBX for E. coli and XLD for Salmonella. The steps that follow were identical regardless of which bacteria. A colony from the selective plate was transferred onto a PCA plate and left to grow aerobically overnight at 37 °C.
The DNA extraction process was identical for Salmonella and ESBL/AmpC-producing E. coli. Crude DNA was prepared by suspending one colony, taken from the PCA plate, in 100 µL sterile distilled water; followed by a heat shock for 10 min at 95 °C and centrifugation for 2 min at 15 294 g (Eppendorf centrifuge 5417C, Eppendorf, Hamburg, Germany). The samples were stored at -20 °C and later used for PCR targeting resistance genes.
5.3.2 PCR for the detection of resistance genes
Four multiplex PCRs were used in this study: a blaTEM/blaSHV/blaOXA-1 multiplex PCR for ESBL genes; a blaCTX-M multiplex PCR including phylogenetic groups 1, 2 and 9; a plasmid-mediated AmpC β-lactamase gene multiplex PCR including six groups (ACC, MOX, FOX, DHA, CIT and EBC) based on percentage of similarity (as previously described by Pérez-Pérez & Hanson, 2002); a plasmid-mediated colistin gene multiplex consisting of mcr-1 & mcr-2 (Sigma-Aldrich, Steinheim, Germany); also one simplex PCR blaCMY-2 for plasmid-mediated AmpC β-lactamase gene. Specifics for each multiplex, primers, composition of the used mastermix and PCR program can be seen in the tables in addendum 7.1.6.2. Extracted DNA of each isolate was subjected to each multiplex and simplex PCR except for the colistin multiplex (see 3.2.2.). The following control strains were used: Salmonella possessing TEM and OXA (ref TIAC1281) , Klebsiella pneumoniae possessing SHV (ref ATCC700603), E. coli possessing CTX-M1 (ref TIAC 809), Salmonella possessing CTX-M2 (ref
2652), Salmonella possessing CTX-M9 (ref 2015_1130), E. coli possessing CIT and CMY-2 (ref TIAC 811 and TIAC 1973), E. coli possessing ACC and CMY-2 (ref CP156). All control strains originated from ‘Belgian Scientific Institute for Public Health (WIV/ISP) (Dr. Nadine Bottledoorn, Dr. Cristina Garcia-Graells).

5. Materials and Methods
41
5.3.3 Gel electrophoresis for the detection of resistance genes
Horizontal gel electrophoresis was performed with 1.5% (w:v) ultra-pure agarose (Invitrogen, Paisley, Scotland) gel (buffered with Tris-Borate-EDTA (TBE; Sigma-Aldrich, Steinheim, Germany). The agarose gel matrix was loaded with 10 µL sample, a 100-bp DNA marker (Bioline, USA) and one slot was left as blank control. Afterwards the gel was subjected to a steady electric field of 120V and left to run for 1-1.5 h (Powerpac 3000, Bio-Rad, Marnes- la-Coquette, France). Amplicons were stained with ethidium bromide (0.5 µg/mL) (Sigma-Aldrich, Steinheim, Germany) for 30 – 45min followed by visualizing and photographing under UV exposure. Next, the photos were processed using Quantity One® 1-D analysis software (Bio-Rad, Marnes-la-Coquette, France). Finally amplicon sizes were determined by comparison to the 100-bp DNA marker and the control strains.
5.4 Distribution of isolates
Below graphs showing how many isolates were used for each experiment.
5.4.1 Investigation of ESBL/AmpC-producing E. coli and Salmonella
Antibiotic susceptibility of pig-associated E. coli and Salmonella isolates taken from three slaughterhouses were evaluated by means of disk diffusion, disk prediffusion and microbroth dilution. Furthermore a PCR followed by agarose gel electrophoresis was performed to verify for the presence of ESBL/AmpC and/or colistin resistance genes. The source and number of E. coli and Salmonella isolates used can be seen in figure 11. Only E. coli isolates that showed growth on TBX supplemented with CTX were used (indicative for ESBL/AmpC phenotype). Although isolates
originate from either tonsil and/or rectum content no distinction was made due to the number of isolates not being comparable. Because Salmonella that co-occurred with ESBL/AmpC E. coli could have given a different phenotype/ genotype than the Salmonella that occurred without ESBL/AmpC E. coli, two groups of Salmonella were created. These two groups are shown in figure 12. Since comparisons were made between the two groups of Salmonella isolates and the ESBL/AmpC E. coli isolates, a general overview graph can be seen in figure 13. Just like in figure 11 the number of isolates per slaughterhouse and the number of batches per slaughterhouse can be seen in figure 12 and figure 13. As can be seen in figures 11 through 13, the distributions of isolates among the different slaughterhouses were unequal which made for difficult comparisons of the results between the different slaughterhouses. Additionally although isolates were taken from different batches, the number of isolates per batch varied from one to ten isolates. Finally
the number of ESBL E. coli and Salmonella found with ESBL E. coli isolates were approximately 70 whereas the number of Salmonella isolates found without the presence of ESBL E. coli was only 32. For these reasons the results were analysed as function of the isolates as a whole group and not split into categories such as batches or slaughterhouse. Yet the different batches eliminate possible biased results that could have arised if only one large batch should have been chosen.
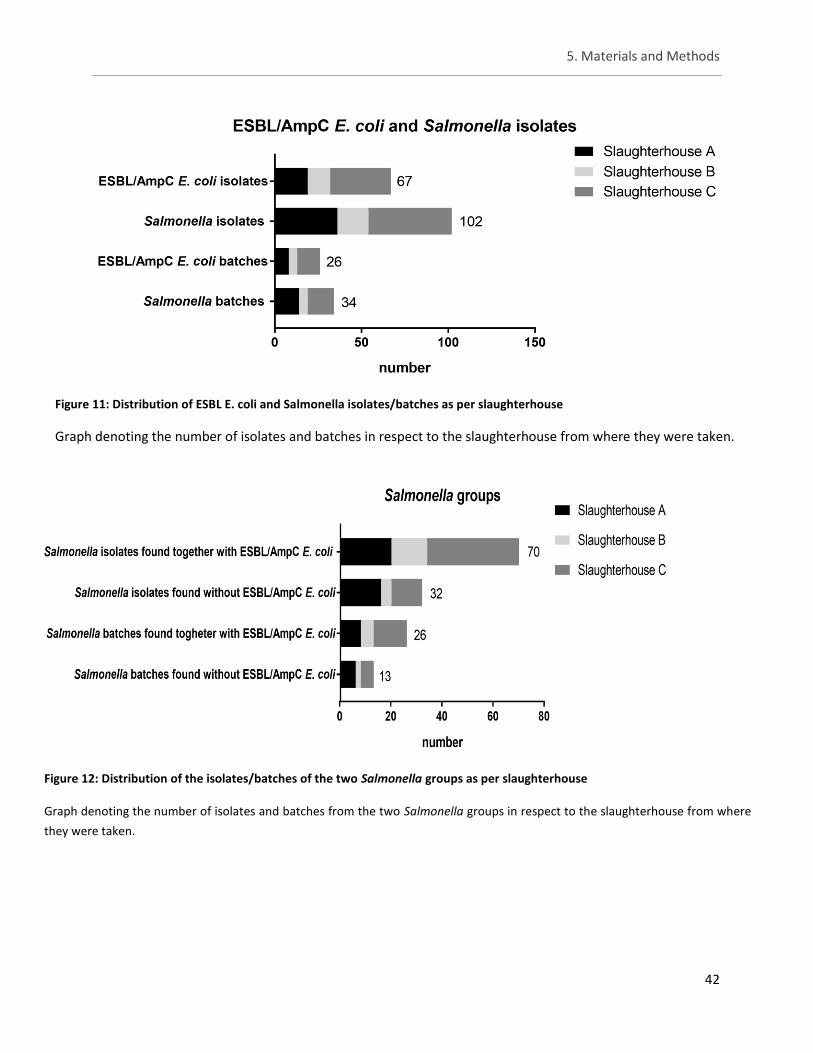
5. Materials and Methods
42
Figure 12: Distribution of the isolates/batches of the two Salmonella groups as per slaughterhouse
Graph denoting the number of isolates and batches from the two Salmonella groups in respect to the slaughterhouse from where
they were taken.
Figure 11: Distribution of ESBL E. coli and Salmonella isolates/batches as per slaughterhouse
Graph denoting the number of isolates and batches in respect to the slaughterhouse from where they were taken.
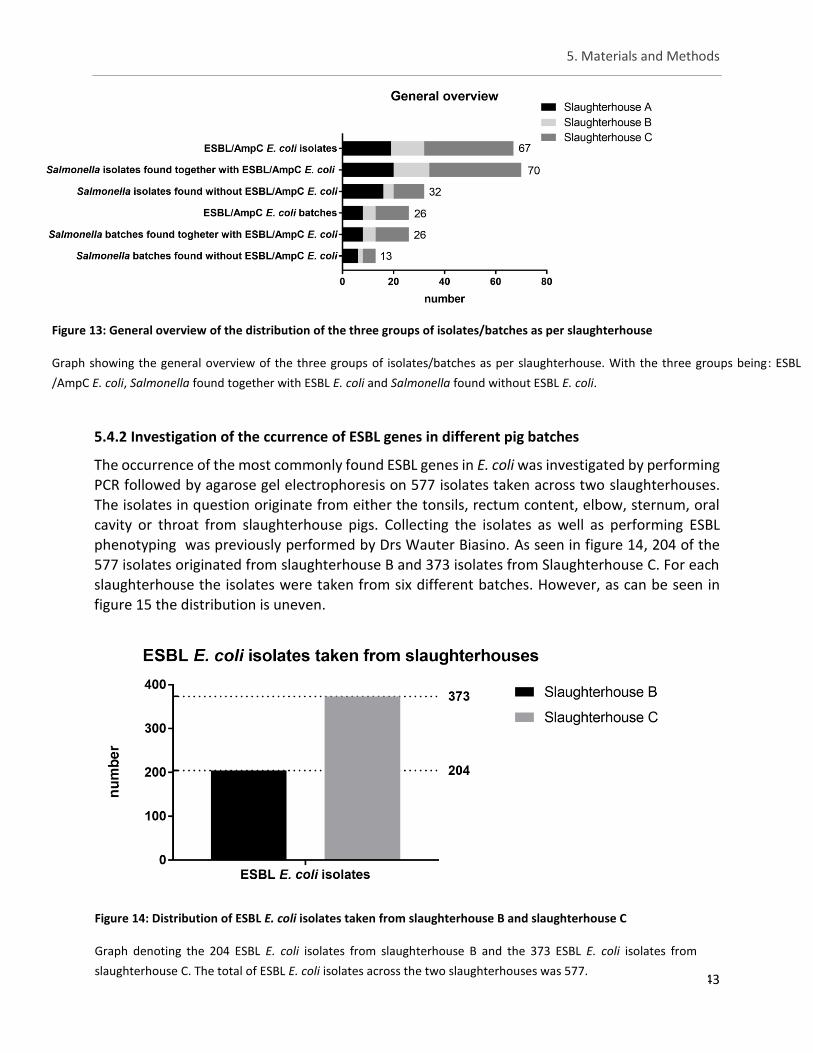
5. Materials and Methods
43
5.4.2 Investigation of the ccurrence of ESBL genes in different pig batches
The occurrence of the most commonly found ESBL genes in E. coli was investigated by performing PCR followed by agarose gel electrophoresis on 577 isolates taken across two slaughterhouses. The isolates in question originate from either the tonsils, rectum content, elbow, sternum, oral cavity or throat from slaughterhouse pigs. Collecting the isolates as well as performing ESBL phenotyping was previously performed by Drs Wauter Biasino. As seen in figure 14, 204 of the
577 isolates originated from slaughterhouse B and 373 isolates from Slaughterhouse C. For each slaughterhouse the isolates were taken from six different batches. However, as can be seen in figure 15 the distribution is uneven.
Figure 14: Distribution of ESBL E. coli isolates taken from slaughterhouse B and slaughterhouse C
Graph denoting the 204 ESBL E. coli isolates from slaughterhouse B and the 373 ESBL E. coli isolates from
slaughterhouse C. The total of ESBL E. coli isolates across the two slaughterhouses was 577.
Figure 13: General overview of the distribution of the three groups of isolates/batches as per slaughterhouse
Graph showing the general overview of the three groups of isolates/batches as per slaughterhouse. With the three groups being: ESBL
/AmpC E. coli, Salmonella found together with ESBL E. coli and Salmonella found without ESBL E. coli.
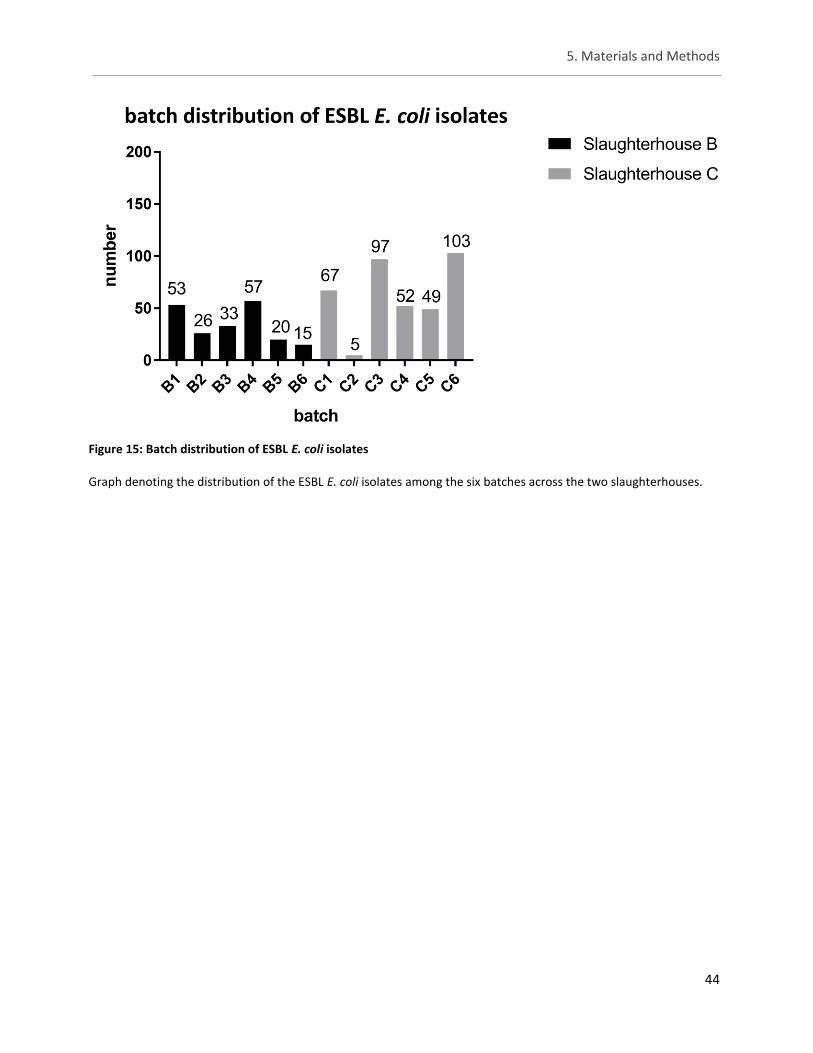
5. Materials and Methods
44
Figure 15: Batch distribution of ESBL E. coli isolates
Graph denoting the distribution of the ESBL E. coli isolates among the six batches across the two slaughterhouses.

6 References
45
6. References
Anago E, Ayi-Fanou L, Akpovi CD, Hounkpe WB, Agassounon-Djikpo Tchibozo M, Bankole HS & Sanni A (2015) Antibiotic resistance and genotype of beta-lactamase producing Escherichia coli in nosocomial infections in Cotonou, Benin. Ann. Clin. Microbiol. Antimicrob. 14: 5
Andrews J (2001) Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 48: 5–16
Anjum MF, Duggett NA, AbuOun M, Randall L, Nunez-Garcia J, Ellis RJ, Rogers J, Horton R, Brena C, Williamson S, Martelli F, Davies R & Teale C (2016) Colistin resistance in Salmonella and Escherichia coli isolates from a pig farm in Great Britain. J. Antimicrob. Chemother. 71: 2306–2313
Arepyeva MA, Kolbin AS, Sidorenko SV, Lawson R, Kurylev AA, Balykina YE, Mukhina NV & Spiridonova AA (2017) A mathematical model for predicting the development of bacterial resistance based on the relationship between the level of antimicrobial resistance and the volume of antibiotic consumption. J. Glob. Antimicrob. Resist.
Asbel LE & Levison ME (2000) Cephalosporins, carbapenems, and monobactams. Infect. Dis. Clin. 14: 435–447
Bae D, Cheng CM & Khan AA (2015) Characterization of extended-spectrum β-lactamase (ESBL) producing non-typhoidal Salmonella (NTS) from imported food products. Int. J. Food Microbiol. 214: 12–17
Bartlett JG, Gilbert DN & Spellberg B (2013) Seven ways to preserve the Miracle of antibiotics. Clin. Infect. Dis. 56: 1445–1450
Bélanger L, Garenaux A, Harel J, Boulianne M, Nadeau E & Dozois CM (2011) Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. Coli. Immunol. Med. Microbiol. 62: 1–10
Bell JM, Chitsaz M, Turnidge JD, Barton M, Walters LJ & Jones RN (2007) Prevalence and significance of a negative extended-spectrum β-lactamase (ESBL) confirmation test result after a positive ESBL screening test result for isolates of Escherichia coli and Klebsiella pneumoniae: Results from the SENTRY asia-pacific surveilla. J. Clin. Microbiol. 45: 1478–1482
BelVet-SAC (2013) Belgian Veterinary Surveillance of Antimicrobial Consumption National consumption report. : 1–43
BelVetSAC (2015) Belgian Veterinary Surveillance of Antibacterial Consumption. : 1–40
Ben-ami R, Arslan H, Pitout JDD, Quentin C, Calbo ES, Azap K, Arpin C, Pascual A, Livermore DM, Garau J & Carmeli Y (2009) A Multinational Survey of Risk Factors for Infection with Extended-Spectrum β-Lactamase–Producing Enterobacteriaceae in Nonhospitalized Patients. Clin. Infect. Dis. 49: 2015–2016
BEUC (2014) Antibiotic use in livestock : Time to act. Eur. Consum. Organ. 32:
Bhaskar M & Mandal J (2016) Use of a modified disc diffusion test to detect AmpC beta lactamase production in Escherichia coli from cases of urinary tract infections. Int. J. Pharma Bio Sci. Microbiol. 7: 579–583
Bollerslev AM, Nauta M, Hald T, Hansen TB & Aabo S (2017) A risk-based approach for evaluation of hygiene performance at pig slaughter. Food Control 75: 116–125
Bonnet R (2004) Growing group of extended spectrum: the CTX-M enzymes. Antimicrob Agent Chemother 48: 1–14
Boyen F, Vangroenweghe F, Butaye P, De Graef E, Castryck F, Heylen P, Vanrobaeys M & Haesebrouck F (2010) Disk prediffusion is a reliable method for testing colistin susceptibility in porcine E. coli strains. Vet. Microbiol. 144: 359–362
Brauer A, Telling K, Laht M, Kalmus P, Lutsar I, Remm M, Kisand V & Tenson T (2016) Plasmid with colistin resistance gene mcr-1 in extended-spectrum-β-Lactamase-producing Escherichia coli strains isolated from pig slurry in Estonia. Antimicrob. Agents Chemother. 60: 6933–6936

6 References
46
Brogden RN, Heel RC, Speight TM & Avery GS (1979) Cefuroxime: A Review of its Antibacterial Activity, Pharmacological Properties and Therapeutic Use. Drugs 17: 233–266
Bush K, Macalintal C, Rasmussen BA, Lee VJ & Yang Y (1993) Kinetic interactions of tazobactam with β-lactamases from all major structural classes. Antimicrob. Agents Chemother. 37: 851–858
Callens B (2015) Antimicrobial use and resistance in Belgian pig production.
Callens B, Persoons D, Maes D, Laanen M, Postma M, Boyen F, Haesebrouck F, Butaye P, Catry B & Dewulf J (2012) Prophylactic and metaphylactic antimicrobial use in Belgian fattening pig herds. Prev. Vet. Med. 106: 53–62
Catry B, Cavaleri M, Baptiste K, Grave K, Grein K, Holm A, Jukes H, Liebana E, Navas AL, Mackay D, Magiorakos AP, Romo MAM, Moulin G, Madero CM, Pomba MCMF, Powell M, Pyorala S, Rantala M, Ruzauskas M, Sanders P, et al (2015) Use of colistin-containing products within the European Union and European Economic Area (EU/EEA): development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 46: 297–306
Cavaco L, Mordhorst H & Hendriksen R (2016) Laboratory protocol: PCR for plasmid-mediated colistin resistance genes, mcr-1 and mcr-2 (multiplex). DTU food Natl. food Inst. 2: 1–15
CDC (2013) Antibiotic resistance threats in the United States, 2013
Center for Disease Dynamics Economics & Policy (2015) The State of the world’s antibiotics 2015
CLSI (2008) Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals ; Approved Standard — Second Edition Serving the World ’ s Medical Science Community Through Voluntary Consensus. 28:
CLSI (2013) Performance Standards for Antimicrobial Susceptibility Testing, M100-S23
Coates AR, Halls G & Hu Y (2011) Novel classes of antibiotics or more of the same? Br. J. Pharmacol. 163: 184–194
Cosgrove SE (2006) The relationship between antimicrobial resistance and patient outcomes: mortality, length of hospital stay, and health care costs. Clin Infect Dis 42: S82–S89
Cox JM & Pavic A (2014) Salmonella. Encycl. Food Microbiol. 3: 322–331
Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M & Finlay BB (2013) Recent Advances in Understanding Enteric Pathogenic Escherichia coli. Clin. Microbiol. Rev. 26: 822–880
Dallenne C, da Costa A, Decré D, Favier C & Arlet G (2010) Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 65: 490–495
DANMAP (2013) DANMAP 2013. Use of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from food animals, food and humans in Denmark
Davies J & Davies D (2010) Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 74: 417–433
Davis MA, Besser TE, Orfe LH, Baker KNK, Lanier AS, Broschat SL, New D & Call DR (2011) Genotypic-phenotypic discrepancies between antibiotic resistance characteristics of Escherichia coli isolates from calves in management settings with high and low antibiotic use. Appl. Environ. Microbiol. 77: 3293–3299
De Busser E V., Maes D, Houf K, Dewulf J, Imberechts H, Bertrand S & De Zutter L (2011) Detection and characterization of Salmonella in lairage, on pig carcasses and intestines in five slaughterhouses. Int. J. Food Microbiol. 145: 279–286
De Kraker MEA, Wolkewitz M, Davey PG, Koller W, Berger J, Nagler J, Icket C, Kalenic S, Horvatic J, Seifert H, Kaasch A, Paniara O, Argyropoulou A, Bompola M, Smyth E, Skally M, Raglio A, Dumpis U, Kelmere AM, Borg M, et al (2011) Burden of antimicrobial resistance in European hospitals: Excess mortality and length of hospital stay associated with bloodstream infections due to Escherichia coli resistant to third-generation cephalosporins. J.

6 References
47
Antimicrob. Chemother. 66: 398–407
Delhalle L, De Sadeleer L, Bollaerts K, Farnir F, Saegerman C, Korsak N, Dewulf J, De Zutter L & Daube G (2008) Risk factors for Salmonella and hygiene indicators in the 10 largest Belgian pig slaughterhouses. J. Food Prot. 71: 1320–1329
Doi Y & Paterson DL (2007) Detection of plasmid-mediated class C β-lactamases. Int. J. Infect. Dis. 11: 191–197
Doumith M, Godbole G, Ashton P, Larkin L, Dallman T, Day M, Day M, Muller-Pebody B, Ellington MJ, de Pinna E, Johnson AP, Hopkins KL & Woodford N (2016) Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J. Antimicrob. Chemother. 71: 2300–2305
ECDC (2016) Plasmid-mediated colistin resistance in Enterobacteriaceae
EFSA (2009) The Community Summary Report on Food-borne Outbreaks in the European Union in 2007
EFSA (2011) Scientific Opinion on the public health risks of bacterial strains producing extended-spectrum β -lactamases and / or AmpC β -lactamases in food and food producing strains. EFSA J. 9: 1–95
EFSA (2013) Scientific Opinion on Carbapenem resistance in food animal ecosystems. EFSA J. 11: 3501
EFSA (2017) The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2015. EFSA J. 15:
Embolism P (2008) Surgeon General ’ s Perspectives. 123: 420–421
EUCAST (2017) European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters
European Food Safety Authority & European Centre for Disease Prevention and Control (2016) The European Union Summary Report on antimicrobial resistance in Antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in the European Union in 2014. EFSA J. 14: 4380
European Medicines Agency (2013) Use of colistin products in animals within the European Union: development of resistance and possible impact on human and animal health. 44: 25
Eurostat (2016) The EU in the world 2016
Evans BA & Amyes SGB (2014) OXA β-lactamases. Clin. Microbiol. Rev. 27: 241–263
Ewers C, Anta E, Diehl I, Philipp H & Wieler LH (2009) Intestine and Environment of the Chicken as Reservoirs for Extraintestinal Pathogenic Escherichia coli Strains with Zoonotic Potential. Appl. Environ. Microbiol. 75: 184–192
Falgenhauer L, Waezsada SE, Yao Y, Imirzalioglu C, Käsbohrer A, Roesler U, Michael GB, Schwarz S, Werner G, Kreienbrock L & Chakraborty T (2016) Colistin resistance gene mcr-1 in extended-spectrum β-lactamase-producing and carbapenemase-producing Gram-negative bacteria in Germany. Lancet Infect. Dis. 16: 282–283
Fernandes P & Martens E (2016) Antibiotics in Late Clinical Development. Biochem. Pharmacol.
Ferrer Savall J, Bidot C, Leblanc-Maridor M, Belloc C & Touzeau S (2016) Modelling Salmonella transmission among pigs from farm to slaughterhouse: Interplay between management variability and epidemiological uncertainty. Int. J. Food Microbiol. 229: 33–43
Figueiredo R, Card RM, Nunez J, Pomba C, Mendonça N, Anjum MF & Da Silva GJ (2016) Detection of an mcr-1-encoding plasmid mediating colistin resistance in Salmonella enterica from retail meat in Portugal. J. Antimicrob. Chemother. 71: 2338–2340
Fosse J, Seegers H & Magras C (2008) Foodborne zoonoses due to meat: A quantitative approach for a comparative risk assessment applied to pig slaughtering in Europe. Vet. Res. 39: 1–16

6 References
48
Frost LS, Leplae R, Summers AO, Toussaint A & Edmonton A (2005) Mobile genetic elements: The agents of open source evolution. Nature 3: 722–732
Galani I, Kontopidou F, Souli M, Rekatsina PD, Koratzanis E, Deliolanis J & Giamarellou H (2008) Colistin susceptibility testing by Etest and disk diffusion methods. Int. J. Antimicrob. Agents 31: 434–439
Garcia-Graells C, De Keersmaecker S, Pochet B, Dierick K & Botteldoorn N (2016) Detection of plasmid mediated colistin resistance (mcr-1 gene) in Salmonella spp. isolated from food in Belgium. In Proceedings of the Twenty-first Conference on Food Microbiology, September 15th & 16th, 2016, Brussels p 113.
Ghafir Y, China B, Dierick K, De Zutter L & Daube G (2008) Hygiene indicator microorganisms for selected pathogens on beef, pork, and poultry meats in Belgium. J. Food Prot. 71: 35–45
Golkar Z, Bagasra O & Gene Pace D (2014) Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries. 8: 129–136
Gould IM & Bal AM (2013) New antibiotic agents in the pipeline and how they can help overcome microbial resistance. Virulence 4: 185–91
Haenni M, Métayer V, Gay E & Madec JY (2016a) Increasing trends in mcr-1 prevalence among extended-spectrum-β-lactamase-producing Escherichia coli isolates from French calves despite decreasing exposure to colistin. Antimicrob. Agents Chemother. 60: 6433–6434
Haenni M, Poirel L, Kieffer N, Châtre P, Saras E, Métayer V, Dumoulin R, Nordmann P & Madec JY (2016b) Co-occurrence of extended spectrum β lactamase and MCR-1 encoding genes on plasmids. Lancet Infect. Dis. 16: 281–282
Hao H, Cheng G, Iqbal Z, Ai X, Hussain HI, Huang L, Dai M, Wang Y, Liu Z & Yuan Z (2014) Benefits and risks of antimicrobial use in food-producing animals. Front. Microbiol. 5: 1–11
Hasman H, Mevius D, Veldman K, Olesen I & Aarestrup FM (2005) β-Lactamases among extended-spectrum β-lactamase (ESBL)-resistant Salmonella from poultry, poultry products and human patients in The Netherlands. J. Antimicrob. Chemother. 56: 115–121
Ho K (2001) Bacteriophage Therapy for Bacterial Infections: Rekindling a Memory from the Pre-Antibiotics Era. Perspect. Biol. Med. 44: 1–16
Holt KE, Thomson NR, Wain J, Langridge GC, Hasan R, Bhutta ZA, Quail MA, Norbertczak H, Walker D, Simmonds M, White B, Bason N, Mungall K, Dougan G & Parkhill J (2009) Pseudogene accumulation in the evolutionary histories of Salmonella enterica serovars Paratyphi A and Typhi. BMC Genomics 10: 36
Ibrahim DR, Dodd CER, Stekel DJ, Ramsden SJ & Hobman JL (2016) Multidrug resistant, extended spectrum β-lactamase (ESBL)-producing Escherichia coli isolated from a dairy farm. FEMS Microbiol. Ecol. 92: 1–13
Irrgang A, Roschanski N, Tenhagen BA, Grobbel M, Skladnikiewicz-Ziemer T, Thomas K, Roesler U & Käsbohrer A (2016) Prevalence of mcr-1 in E. coli from livestock and food in Germany, 2010-2015. PLoS One 11: 2010–2015
Jacoby GA (2009) AmpC β-Lactamases. Clin. Microbiol. Rev. 22: 161–182
Jakobsen L, Spangholm DJ, Pedersen K, Jensen LB, Emborg H, Agersø Y, Aarestrup FM, Hammerum AM & Frimodt-møller N (2010) International Journal of Food Microbiology Broiler chickens , broiler chicken meat , pigs and pork as sources of ExPEC related virulence genes and resistance in Escherichia coli isolates from community-dwelling humans and UTI patients. Int. J. Food Microbiol. 142: 264–272
Kaferstein F & Abdussalam M (1999) Food safety in the 21st century. Bull. World Health Organ. 77: 347–351
Kametani T & Umezawa O (1980) Structure-Activity Relationship of Colistins. Chem. Pharm. Bull. 28: 369–375
Kaper JB, Nataro JP & Mobley HLT (2004) Pathogenic Escherichia coli. Nat. Rev. 2: 123–140

6 References
49
Kar D, Bandyopadhyay S, Dimri U, Mondal DB, Nanda PK, Das AK, Batabyal S, Dandapat P & Bandyopadhyay S (2016) Antibacterial effect of silver nanoparticles and capsaicin against MDR-ESBL producing Escherichia coli: An in vitro study. Asian Pacific J. Trop. Dis. 6: 807–810
Kliebe C, Nies BA, Meyer JF, Tolxdorff-Neutzling RM & Wiedemann B (1985) Evolution of plasmid-coded resistance to broad-spectrum cephalosporins. Antimicrob. Agents Chemother. 28: 302–307
Lagacé-wiens PRS, Nichol KA, Nicolle LE, Decorby MR, Mccracken M, Alfa MJ, Mulvey MR, Zhanel GG, Lagacé-wiens PRS, Nichol KA & Nicolle LE (2007) ESBL genotypes in fluoroquinolone-resistant and fluoroquinolone-susceptible ESBL-producing Escherichia coli urinary isolates in Manitoba. Can. J. Infect. Dis. Med. Microbiol. 18: 133–137
Lahuerta a, Westrell T, Takkinen J, Boelaert F, Rizzi V, Helwigh B, Borck B, Korsgaard H, Ammon A & Makela P (2011) Zoonoses in the European Union: origin, distribution and dynamics - the EFSA-ECDC summary report 2009. Euro Surveill. 16: 1–4
Landman D, Georgescu C, Martin DA & Quale J (2008) Polymyxins revisited. Clin. Microbiol. Rev. 21: 449–465
Lavilla S, González-López JJ, Miró E, Domínguez A, Llagostera M, Bartolomé RM, Mirelis B, Navarro F & Prats G (2008) Dissemination of extended-spectrum β-lactamase-producing bacteria: The food-borne outbreak lesson. J. Antimicrob. Chemother. 61: 1244–1251
Leverstein-van Hall MA, Dierikx CM, Cohen Stuart J, Voets GM, van den Munckhof MP, van Essen-Zandbergen A, Platteel T, Fluit AC, van de Sande-Bruinsma N, Scharinga J, Bonten MJM & Mevius DJ (2011) Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17: 873–880
Li XZ, Mehrotra M, Ghimire S & Adewoye L (2007) β-Lactam resistance and β-lactamases in bacteria of animal origin. Vet. Microbiol. 121: 197–214
Liebana E, Carattoli A, Coque TM, Hasman H, Magiorakos AP, Mevius D, Peixe L, Poirel L, Schuepbach-Regula G, Torneke K, Torren-Edo J, Torres C & Threlfall J (2013) Public health risks of enterobacterial isolates producing extended-spectrum β-lactamases or AmpC β-lactamases in food and food-producing animals: An EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 56: 1030–1037
Liu YY, Wang Y, Walsh TR, Yi LX, Zhang R, Spencer J, Doi Y, Tian G, Dong B, Huang X, Yu LF, Gu D, Ren H, Chen X, Lv L, He D, Zhou H, Liang Z, Liu JH & Shen J (2016) Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 16: 161–168
Livermore DM (1995) beta-Lactamases in laboratory and clinical resistance. Clin. Microbiol. Rev. 8: 557–84
Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olsson-Liljequist B, Paterson DL, Rice LB, Stelling J, Struelens MJ, Vatopoulos A, Weber JT & Monnet DL (2012) Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18: 268–281
Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O’Brien SJ, Jones TF, Fazil A & Hoekstra RM (2010) The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 50: 882–889
Mannion C, Egan J, Lynch BP, Fanning S & Leonard N (2008) An investigation into the efficacy of washing trucks following the transportation of pigs--a Salmonella perspective. Foodborne Pathog. Dis. 5: 261–271
Marshall BM & Levy SB (2011) Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 24: 718–733
Matsuzaki S, Rashel M, Uchiyama J, Sakurai S, Ujihara T, Kuroda M, Ikeuchi M, Tani T, Fujieda M, Wakiguchi H & Imai S (2005) Bacteriophage therapy: A revitalized therapy against bacterial infectious diseases. J. Infect. Chemother.

6 References
50
11: 211–219
Maurya AP, Dhar Chanda D, Bora D, Das Talukdar A, Chakravarty A & Bhattacharjee A (2017) Transcriptional Response of Multiple ESBL Genes Within Escherichia coli Under Oxyimino-Cephalosporin Stress. Microb. drug Resist. 23: 133–138
McDermott PF, Tyson GH, Kabera C, Chen Y, Li C, Folster JP, Ayers SL, Lam C, Tate HP & Zhao S (2016) Whole-genome sequencing for detecting antimicrobial resistance in nontyphoidal Salmonella. Antimicrob. Agents Chemother. 60: 5515–5520
Mesa RJ, Blanc V, Blanch AR, Cortés P, González JJ, Lavilla S, Miró E, Muniesa M, Saco M, Tórtola MT, Mirelis B, Coll P, Llagostera M, Prats G & Navarro F (2006) Extended-spectrum β-lactamase-producing Enterobacteriaceae in different environments (humans, food, animal farms and sewage). J. Antimicrob. Chemother. 58: 211–215
Michael CA, Dominey-Howes D & Labbate M (2014) The antimicrobial resistance crisis: causes, consequences, and management. Front. public Heal. 2: 145
Mohd Nasir MD & Parasakthi N (2004) Comparison of susceptibility test methods to detect penicillin susceptibility in Streptococcus pneumoniae isolates. Malays. J. Pathol. 26: 29–33
Molitoris E, Fagerberg DJ, Quarles CL & Krichevsky MI (1987) Changes in Antimicrobial Resistance in Fecal Bacteria Associated With Pig Transit and Holding Times At Slaughter Plants. Appl. Environ. Microbiol. 53: 1307–1310
Monnaie D & Frere JM (1993) Interaction of clavulanate with class C β-lactamases. FEBS Lett. 334: 269–271
Morales AS, Fragoso de Araújo J, de Moura Gomes VT, Reis Costa AT, Prazeres Rodrigues D dos, Porfida Ferreira TS, de Lima Filsner PHN, Felizardo MR & Micke Moreno A (2012) Colistin Resistance in Escherichia coli and Salmonella enterica Strains Isolated from Swine in Brazil. Sci. World J. 2012: 1–4
Moro MH, Beran GW, Griffith RW & Hoffman LJ (2000) Effects of heat stress on the antimicrobial drug resistance of Escherichia coli of the intestinal flora of swine. J. Appl. Microbiol. 88: 836–844
Moro MH, Beran GW, Hoffman LJ & Griffith RW (1998) Effects of cold stress on the antimicrobial drug resistance of Escherichia coli of the intestinal flora of swine. Lett. Appl. Microbiol. 27: 251–254
Nijs A, Cartuyvels R, Mewis A, Peeters V, Rummens JL & Magerman K (2003) Comparison of efficacy and cost-effectiveness of BIOMIC VIDEO and Vitek antimicrobial susceptibility test systems for use in the clinical microbiology laboratory. J. Clin. Microbiol. 41: 3627–3630
Nordmann P, Cuzon G & Naas T (2009) The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect. Dis. 9: 228–236
Nordmann P, Jayol A & Poirel L (2016) Rapid detection of polymyxin resistance in Enterobacteriaceae. Emerg. Infect. Dis. 22: 1038–1043
O’Neill J (2014) Antimicrobial Resistance : Tackling a crisis for the health and wealth of nations. Rev. Antimicrob. Resist.: 1–16
Ojer-Usoz E, González D, Vitas AI, Leiva J, García-Jalón I, Febles-Casquero A & Escolano M de la S (2013) Prevalence of extended-spectrum β-lactamase-producing Enterobacteriaceae in meat products sold in Navarra, Spain. Meat Sci. 93: 316–321
PAHO/WHO (2016) Epidemiological Alert: Enterobacteriacea with plasmid-mediated transferable colistin resistance, public health implications in the Americas
Patel JB, Tenover FC, Turnidge JD & Jorgensen JH (2011) Susceptibility Test Methods: Dilution and Disk Diffusion Methods. In Manual of Clinical Microbiology pp 1122–1143.
Paterson DL & Bonomo RA (2005) Extended-Spectrum beta-Lactamases : a Clinical Update. Clin. Microbiol. Rev. 18: 657–686

6 References
51
Pe C, Coque TM, Baquero F, Oliver A & Microbiologı S De (2001) Nucleotide Sequence and Characterization of a Novel Cefotaxime-Hydrolyzing β-Lactamase (CTX-M-10) Isolated in Spain. Antimicrob. Agents Chemother. 45: 616–620
Pérez-Pérez FJ & Hanson ND (2002) Detection of plasmid-mediated AmpC beta-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 40: 2153–2162
Perrin-Guyomard A, Bruneau M, Houée P, Deleurme K, Legrandois P, Poirier C, Soumet C & Sanders P (2016) Prevalence of mcr-1 in commensal Escherichia coli from French livestock, 2007 to 2014. Euro Surveill. 21: 1–3
Piddock LJ V (2012) The crisis of no new antibiotics-what is the way forward? Lancet Infect. Dis. 12: 249–253
Pires SM, Vieira AR, Hald T & Cole D (2014) Source Attribution of Human Salmonellosis: An Overview of Methods and Estimates. Foodborne Pathog. Dis. 0: 1–10
Pitout JD & Laupland KB (2008) Extended-spectrum β-lactamase-producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect. Dis. 8: 159–166
Pitout JDD, Nordmann P, Laupland KB & Poirel L (2005) Emergence of Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs) in the community. J. Antimicrob. Chemother. 56: 52–59
Poirel L, Kämpfer P & Nordmann P (2002) Chromosome-encoded ambler class a β-lactamase of Kluyvera georgiana, a probable progenitor of a subgroup of CTX-M extended-spectrum β-lactamases. Antimicrob. Agents Chemother. 46: 4038–4040
Poirel L, Naas T, Nicolas D, Collet L, Bellais S, Cavallo JD & Nordmann P (2000) Characterization of VIM-2, a carbapenem-hydrolyzing metallo-β-lactamase and its plasmid- and integron-borne gene from a Pseudomonas aeruginosa clinical isolate in France. Antimicrob. Agents Chemother. 44: 891–897
Poirel L, Stephan R, Perreten V & Nordmann P (2014) The carbapenemase threat in the animal world: the wrong culprit. J. Antimicrob. Chemother. 69: 791–795
Rathera I, Kim B-C, Bajpai VK & Parka Y-H (2017) Self-medication and antibiotic resistance: Crisis, current challenges, and prevention. Saudi J. Biol. Sci.
Read AF & Woods RJ (2014) Antibiotic resistance management. Evol. Med. Public Heal. 2014: 147–147
Robin F, Delmas J, Archambaud M, Schweitzer C, Chanal C & Bonnet R (2006) CMT-type β-lactamase TEM-125, an emerging problem for extended-spectrum β-lactamase detection. Antimicrob. Agents Chemother. 50: 2403–2408
Rodríguez-Baño J (2008) Community Infections Caused by Extended-Spectrum. Arch. Intern. Med. 168: 1897–1902
Rodríguez-baño J, Navarro MD, Martínez-martínez L, Muniain M a, Perea J, Pérez-cano R, Pascual A, Romero L & Perea EJ (2004) Epidemiology and Clinical Features of Infections Caused by Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in Nonhospitalized Patients. J. Clin. Microbiol. 42: 1089–1094
Rostagno MH & Callaway TR (2012) Pre-harvest risk factors for Salmonella enterica in pork production. Food Res. Int. 45: 634–640
Sawaya WN, Elnawawy AS, Abu-Ruwaida AS, Khalafawi S & Dashti B (1995) Influence of modified atmosphere packaging on shelf-life of chicken carcasses under refrigerated storage conditions. J. Food Saf. 15: 35–51
Scallan E, Hoekstra RM, Angulo FJ, Tauxe R V., Widdowson MA, Roy SL, Jones JL & Griffin PM (2011) Foodborne illness acquired in the United States-Major pathogens. Emerg. Infect. Dis. 17: 7–15
Schwaber MJ & Carmeli Y (2007) Mortality and delay in effective therapy associated with extended-spectrum β-lactamase production in Enterobacteriaceae bacteraemia: A systematic review and meta-analysis. J. Antimicrob. Chemother. 60: 913–920
Seyedjavadi SS, Goudarzi M & Sabzehali F (2016) Relation between blaTEM, blaSHV and blaCTX-M genes and acute

6 References
52
urinary tract infections. J. Acute Dis. 5: 71–76
Shaikh S, Fatima J, Shakil S, Rizvi SMD & Kamal MA (2015) Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 22: 90–101
Skov R, Frimodt-Møller N & Espersen F (2001) Correlation of MIC methods and tentative interpretive criteria for disk diffusion susceptibility testing using NCCLS methodology for fusidic acid. Diagn. Microbiol. Infect. Dis. 40: 111–116
Skov RL & Monnet DL (2016) Plasmid-mediated colistin resistance (mcr-1 gene): three months later, the story unfolds. Euro Surveill. 21: 1–6
Smet A, Martel A, Persoons D, Dewulf J, Heyndrickx M, Catry B, Herman L, Haesebrouck F & Butaye P (2008) Diversity of extended-spectrum β-lactamases and class C β-lactamases among cloacal Escherichia coli isolates in Belgian broiler farms. Antimicrob. Agents Chemother. 52: 1238–1243
Smith DR (1996) Agarose Gel Electrophoresis. Methods in Molecular Biology. 58: 17–21
Smith JL, Drum DJ V, Dai Y, Kim JM, Sanchez S, Maurer JJ, Hofacre CL & Lee MD (2007) Impact of antimicrobial usage on antimicrobial resistance in commensal Escherichia coli strains colonizing broiler chickens. Appl. Environ. Microbiol. 73: 1404–1414
SØrum H & Sunde M (2001) Resistance to antibiotics in the normal flora of animals. Vet. Res. 32: 227–241
Spellberg B, Blaser M, Guidos RJ, Boucher HW, Bradley JS, Eisenstein BI, Gerding D, Lynfield R, Reller LB, Rex J, Schwartz D, Septimus E, Tenover FC & Gilbert DN (2011) Combating antimicrobial resistance: Policy recommendations to save lives. Clin. Infect. Dis. 52: 397–428
Stapley EO, Birnbaum J, Miller K, Wallick H, Hendlin D & Boyd Woodruff H (1979) Cefoxitin and cephamycins: Microbiological studies. Rev. Infect. Dis. 1: 73–87
Stürenburg E & Mack D (2003) Extended-spectrum β-lactamases: Implications for the clinical microbiology laboratory, therapy, and infection control. J. Infect. 47: 273–295
Sun S, Negrea A, Rhen M & Andersson DI (2009) Genetic analysis of colistin resistance in Salmonella enterica serovar typhimurium. Antimicrob. Agents Chemother. 53: 2298–2305
Tariq F, Khalid Q, Sehgal SA, Mannan S & Jamil F (2016) Possible Roles of the Pseudogenes of Salmonella typhimurium. Zool. Soc. Pakistan 48: 1805–1810
Tenaillon O, Skurnik D, Picard B & Denamur E (2010) The population genetics of commensal Escherichia coli. Nat.Rev.Microbiol. 8: 207–217
Timmerman T, Dewulf J, Catry B, Feyen B, Opsomer G, Kruif A de & Maes D (2006) Quantification and evaluation of antimicrobial drug use in group treatments for fattening pigs in Belgium. Prev. Vet. Med. 74: 251–263
Trobos M, Lester CH, Olsen JE, Frimodt-møller N & Hammerum AM (2009) Natural transfer of sulphonamide and ampicillin resistance between Escherichia coli residing in the human intestine. J. Antimicrob. Chemother. 63: 80–86
Tumbarello M, Sanguinetti M, Montuori E, Trecarichi EM, Posteraro B, Fiori B, Citton R, D’Inzeo T, Fadda G, Cauda R & Spanu T (2007) Predictors of mortality in patients with bloodstream infections caused by extended-spectrum-β-lactamase-producing Enterobacteriaceae: Importance of inadequate initial antimicrobial treatment. Antimicrob. Agents Chemother. 51: 1987–1994
Vaidya VK & Manzoor MA (2009) Horizontal transfer of antibiotic resistance in the marine environment. Nat. Environ. Pollut. Technol. 8: 451–458
Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, Teillant A & Laxminarayan R (2015) Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. U. S. A.: 1–6

6 References
53
Van Damme I, Mattheus W, Bertrand S & De Zutter L (2017) Quantification of hygiene indicators and Salmonella in the tonsils, oral cavity and rectal content samples of pigs during slaughter. Food Microbiol.: 1–9
Ventola CL (2015a) The antibiotic resistance crisis: part 1: causes and threats. Pharm. Ther. 40: 277–83
Ventola CL (2015b) The Antibiotic Resistance Crisis Part 2 : Management Strategies and New Agents. Pharm. Ther. 40: 344–352
Von Salviati C, Friese A, Roschanski N, Laube H, Guerra B, Kasbohrer A, Kreienbrock L & Roesler U (2014) Extended-spectrum beta-lactamases (ESBL)/AmpC beta-lactamases-producing Escherichia coli in German fattening pig farms: a longitudinal study. Berl. Munch. Tierarztl. Wochenschr. 127: 412–419
Walia K, Argüello H, Lynch H, Leonard FC, Grant J, Yearsley D, Kelly S, Duffy G, Gardiner GE & Lawlor PG (2016) Effect of feeding sodium butyrate in the late finishing period on Salmonella carriage, seroprevalence, and growth of finishing pigs. Prev. Vet. Med. 131: 79–86
Walia K, Argüello H, Lynch H, Leonard FC, Grant J, Yearsley D, Kelly S, Duffy G, Gardiner GE & Lawlor PG (2017) Effect of strategic administration of an encapsulated blend of formic acid, citric acid, and essential oils on Salmonella carriage, seroprevalence, and growth of finishing pigs. Prev. Vet. Med. 137: 28–35
Williams KJ & Bax RP (2009) Challenges in developing new antibacterial drugs. Curr. Opin. Investig. Drugs 10: 157–163
Winokur P., Canton R, Casellas J-M & Legakis N (2001) Variations in the prevalence of strains expressing an extended-spectrum β-lactamase phenotype and characterization of isolates from Europe, the Americas, and the Western Pacific region. Clin. Infect. Dis. 32: 94–103
World Health Organization (2014) The evolving threat of antimicrobial resistance: Options for action. WHO Publ.: 1–119
Wray C & Gnanou JC (2000) Antibiotic resistance monitoring in bacteria of animal origin: Analysis of national monitoring programmes. Int. J. Antimicrob. Agents 14: 291–294
Wykoff CC, Flynn HW & Han DP (2011) Allergy to povidoneiodine and cephalosporins: The clinical dilemma in ophthalmic use. Am. J. Ophthalmol. 151: 4–6
Xavier BB, Lammens C, Ruhal R, Kumar-Singh S, Butaye P, Goossens H & Malhotra-Kumar S (2016) Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2 , in Escherichia coli , Belgium, June 2016. Euro Surveill. 21: 1–6
Xu G, An W, Wang H & Zhang X (2015) Prevalence and characteristics of extended-spectrum β-lactamase genes in Escherichia coli isolated from piglets with post-weaning diarrhea in Heilongjiang province, China. Front. Microbiol. 6: 1–9
Yan J-J, Ko W-C, Tsai S-H, Wu H-M, Jin Y-T & Wu J-J (2000) Dissemination of CTX-M-3 and CMY-2 beta -Lactamases among Clinical Isolates of Escherichia coli in Southern Taiwan. J. Clin. Microbiol. 38: 4320–4325
Yang YQ, Zhang AY, Ma SZ, Kong LH, Li YX, Liu JX, Davis MA, Guo XY, Liu BH, Lei CW & Wang HN (2016) Co-occurrence of mcr-1 and ESBL on a single plasmid in Salmonella enterica. J. Antimicrob. Chemother. 71: 2336–2338
Zheng H, Zeng Z, Chen S, Liu Y, Yao Q, Deng Y, Chen X, Lv L, Zhuo C, Chen Z & Liu JH (2012) Prevalence and characterisation of CTX-M β-lactamases amongst Escherichia coli isolates from healthy food animals in China. Int. J. Antimicrob. Agents 39: 305–310
Ziech RE, Lampugnani C, Perin AP, Sereno MJ, Sfaciotte RAP, Viana C, Soares VM, de Almeida Nogueira Pinto JP & dos Santos Bersot L (2016) Multidrug resistance and ESBL-producing Salmonella spp. isolated from broiler processing plants. Brazilian J. Microbiol. 47: 191–195

7. Addenda
54
7. Addenda
7.1 Protocols
Isolates were kept as a glycerol stock at -20 °C and DNA lysates were kept in sterile water also at -20 °C.
7.1.1 Isolation of ESBL/AmpC-producing E. coli and Salmonella
- Using a stomacher apparatus homogenise the tonsillar tissue and rectal content.
- Take 1 mL of the homogenate and plate on TBX supplemented with cefotaxime.
- Incubate overnight aerobically at 44 °C.
7.1.2 Isolation of ESBL/AmpC-producing Salmonella
- Using a stomacher apparatus homogenise the tonsillar tissue and rectal content.
- Pre-enrich the homogenate by incubating it aerobically at 37 °C for 18 h.
- Transfer 0.1 mL of the homogenate (dispersed in 3 drops) into MSRV plate.
- Incubate aerobically at 41,5 °C for 24 h.
- If a migration zone is observed sub cultivate on a XLD agar plate.
- Incubate aerobically at 37 °C for 24 h.
- Pick up a black colony for biochemical tests ( triple sugar ion, indole reaction and lysine decarboxylase test).
o Not self-performed so no further description is given.
7.1.3 Disk diffusion for ESBL/AmpC-producing E. coli and Salmonella
All procedures are to be performed while wearing gloves and working sterile. Leave plates to dry in the flow before use.
Day 1
- Grow bacteria from stock on selective plate (TBX for E. coli and XLD for Salmonella) aerobically for 24 h at 37 °C.
The steps that follow are identical regardless of which bacteria.
Day 2
- Transfer a colony from the selective plate onto a PCA plate, leave to grow aerobically overnight at 37 °C.

7. Addenda
55
Day 3
- Take four to five colonies from the PCA plate and resuspend these in a sterile 0.85% NaCl solution till a 0.5 Mcfarland concentration is obtained.
o Try to pick up identical colonies.
o Adjust the density by either adding more bacteria or adding more 0.85% NaCl.
- Using a sterile swab, plate 0.5 Mcfarland bacteria solution homogeneously on MH II agar plates.
o Preferably inoculate the plates within 15 min and definitely within 60 min of making the suspension.
o Squeeze excess suspension from the swab by pressing against the inside of the tube as to prevent a to high inoculum on the plates.
o Plates have to be at room temperature (drying the plates for approximately 10 min should be enough).
o Spread the inoculum as homogeneously as possible across the entire plate by inoculating in three directions.
- Place the antibiotic disks correctly on the MH II plates (see figure 16 below) using sterile tweezers. Respect the location and distance between the disks.
o Place the antibiotic disks on the MH II plate within 15 min after inoculation
o Place the antibiotic disks at room temperature before opening as to prevent condensation on the disks
o Make sure the disks are well places (gently press on them with the tweezers after placement)
Figure 16: Template for placing antibiotic disks
Figure showing were to place the antibiotic disks for
ESBL/AmpC disk diffusion.

7. Addenda
56
o The different antibiotic disks are: ceftazidime (CAZ), cefoxitine (CFO), cefotaxime (CTX), cefepime (FEP), ceftazidime + clavulanic acid (CAZ+C), cefotaxime + clavulanic acid (CTX+C), cefepime + clavulanic acid (FEP+C) (concentration of 30µg) and boric acid.
- Incubate the MH II plates aerobically for 24 h at 37 °C.
o Incubate the plates within 15 min after the antibiotic disks are placed.
o Don’t stack the plates too high as this can affect the temperature distribution which may in turn influence the inhibition zones.
Day 4
- Verify the plates for growth
o Growth must be homogenously across the whole surface of the plate; inhibition zones must be uniform circles.
o Culture must be pure.
o If individual colonies are visible than the inoculum wasn’t concentrated enough or a contaminant is present.
- Measure the inhibition zones using an electronic calliper and note the diameters in mm
o Start measuring from the point of total inhibition ( according to the naked eye from a distance of approximately 30 cm).
o Measure the zones from the back of the plates.
o If individual colonies are visible within a inhibition zone: sub cultivate and test for purity.
- Interpretative criteria to determine whether a bacterial strain was ESBL, AmpC or ESBL + AmpC are summed up in table 17.

7. Addenda
57
7.1.4 Disk prediffusion for testing colistin susceptibility of ESBL/AmpC-producing E. coli and
Salmonella All procedures are to be performed while wearing gloves and working sterile. Leave plates to dry in the flow before use. Day 1
- Grow bacteria from stock on selective plate (TBX for E. coli and XLD for Salmonella) aerobically for 24 h at 37 °C.
The steps that follow are identical regardless of which bacteria.
Day 2
- Transfer a colony from the selective plate onto a PCA plate, leave to grow aerobically
overnight at 37 °C.
- Place a 10 µg colistin antibiotic disk on MH II plate using sterile tweezers as shown in figure 17.
- Leave to incubate for 2 h aerobically for 2 h at 22 °C.
- Remove the disk and further incubate for an additional 18 h at 22 °C.
Day 3
- Take four to five colonies from the PCA plate and resuspend these in a sterile 0.85% NaCl solution till a 0.5 Mcfarland concentration is obtained.
o Try to pick up identical colonies.
o Adjust the density by either adding more bacteria or adding more 0.85% NaCl.
- Using a sterile swab, plate 0.5 Mcfarland bacteria solution homogeneously on MH II agar plates.
Figure 17: Template for placing colistin disk
Figure showing where the colistin disk is placed.

7. Addenda
58
o Preferably inoculate the plates within 15 min and definitely within 60 min of
making the suspension.
o Squeeze excess suspension from the swab by pressing against the inside of the tube as to prevent a to high inoculum on the plates.
o Spread the inoculum as homogeneously as possible across the entire plate by inoculating in 3 directions.
- Incubate the MH II plates aerobically overnight at 35 °C.
o Don’t stack the plates too high as this can affect the temperature distribution which may in turn influence the inhibition zones.
Day 4
- Verify the plates for growth.
o Growth must be homogenously across the whole surface of the plate; inhibition zones must be uniform circles.
o Culture must be pure.
o If individual colonies are visible than the inoculum wasn’t concentrated enough or a contaminant is present.
- Measure the inhibition zone using an electronic calliper and note the diameters in mm
o Start measuring from the point of total inhibition ( according to the naked eye from
a distance of approximately 30 cm).
o Measure the zone from the back of the plates.
o If individual colonies are visible within the inhibition zone: sub cultivate and test for purity.
- Interpretative criteria are as followed: sensitive ≥ 20mm; intermediate 17–19 mm; resistant ≤16 mm.

7. Addenda
59
7.1.5 Microbroth dilution for ESBL/AmpC-producing E. coli and Salmonella
All procedures are to be performed while wearing gloves and working sterile. Leave agar plates to dry in the flow before use.
Day 1
- Grow bacteria from stock on selective plate (TBX for E. coli and XLD for Salmonella) aerobically for 24 h at 37 °C.
The steps that follow are identical regardless of which bacteria.
Day 2
- Transfer a colony from the selective plate onto a PCA plate, leave to grow aerobically
overnight at 37 °C.
Day 3
- Take four to five colonies from the PCA plate and resuspend these in a sterile 0.85% NaCl solution till a 0.5 Mcfarland concentration is obtained.
o Try to pick up identical colonies.
o Adjust the density by either adding more bacteria or adding more 0.85% NaCl.
- Inoculate MH II broth with 10 µL of the 0.5 McFarland solution.
- Open the SensititreTM 96-well plates containing the different antibiotics (see 5.2.3).
o For plate design see figure 18 below.
- With a multichannel pipet add 50 µL MH II broth to each well.
o Watch for air bubbles and half-filled tips.
- Seal the 96-well plates with the included foil.
- Incubate aerobically for approximately 18 h at 37 °C.
o Do not stack more than three plates on top of each other. Day 4
- Verify each well for growth by comparing with the positive control.
o Positive wells will show a button and/or turbidity throughout the well.
- A strain is considered to have reduced susceptibility by using the ECOFF values for E.coli or Salmonella for the different antibiotics used.
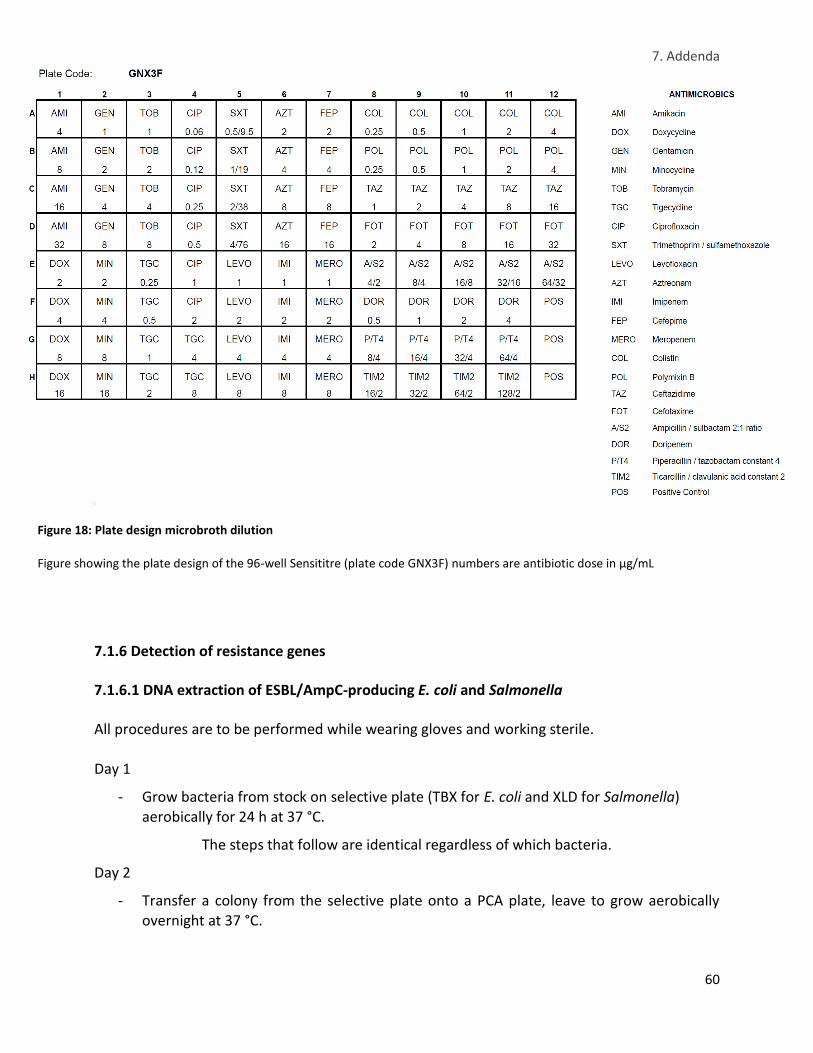
7. Addenda
60
j
7.1.6 Detection of resistance genes 7.1.6.1 DNA extraction of ESBL/AmpC-producing E. coli and Salmonella All procedures are to be performed while wearing gloves and working sterile.
Day 1
- Grow bacteria from stock on selective plate (TBX for E. coli and XLD for Salmonella) aerobically for 24 h at 37 °C.
The steps that follow are identical regardless of which bacteria.
Day 2
- Transfer a colony from the selective plate onto a PCA plate, leave to grow aerobically overnight at 37 °C.
Figure 18: Plate design microbroth dilution
Figure showing the plate design of the 96-well Sensititre (plate code GNX3F) numbers are antibiotic dose in µg/mL

7. Addenda
61
Day 3
- Suspend 1 colony from the PCA plate in 100 µL sterile distilled water.
- Lysate at 95 °C during 10 min.
- Leave to cool for 2 min.
- Centrifugate for 2 min at 12.000 rpm.
o Samples are to be stored quickly at -20 °C or used immediately.
7.1.6.2 PCR for the detection of resistance genes This procedure is to be performed in the molecular lab and gloves are to be worn and rinsed with bleach as to not contaminate the sample with foreign DNA.
- Thaw the products needed to make a PCR mastermix except for the Taq polymerase.
- If necessary thaw the samples.
- Vortex and short spin everything (after thawing).
- Prepare the mastermix.
o Make enough for your samples + blanc + controls + three extra to ensure to have enough.
o Only 96 PCR tubes fit in the machine at one time.
o Always start with the largest volume (water).
o Add Taq polymerase last.
- Distribute 48 µL of mastermix in PCR tubes and add 2 µL of sample (DNA).
four different Multiplexes are available and 1 Simplex: Multiplex I and II for ESBL genes, Multiplex III for AmpC gene, Simplex I for CMY-2 gene (AmpC) and colistin Multiplex for mcr-1 and mcr-2. Primers are diluted to make a 200 pmol/µL or 100 pmol/µL stock solution and then further diluted to a 10 pmol/µL work solution. How to prepare the different Multiplexes is summarized in the tables here below (table 18 through 22). DNA is added in a separate cabin.
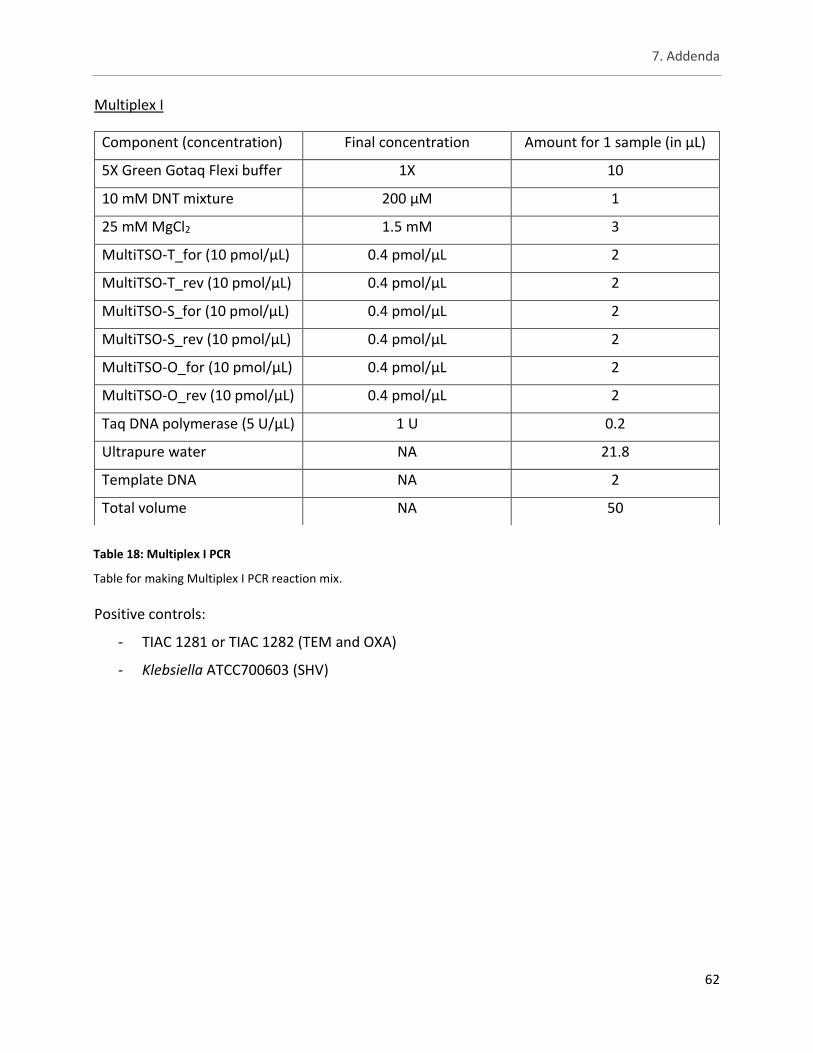
7. Addenda
62
Multiplex I
Positive controls:
- TIAC 1281 or TIAC 1282 (TEM and OXA)
- Klebsiella ATCC700603 (SHV)
Component (concentration) Final concentration Amount for 1 sample (in µL)
5X Green Gotaq Flexi buffer 1X 10
10 mM DNT mixture 200 µM 1
25 mM MgCl2 1.5 mM 3
MultiTSO-T_for (10 pmol/µL) 0.4 pmol/µL 2
MultiTSO-T_rev (10 pmol/µL) 0.4 pmol/µL 2
MultiTSO-S_for (10 pmol/µL) 0.4 pmol/µL 2
MultiTSO-S_rev (10 pmol/µL) 0.4 pmol/µL 2
MultiTSO-O_for (10 pmol/µL) 0.4 pmol/µL 2
MultiTSO-O_rev (10 pmol/µL) 0.4 pmol/µL 2
Taq DNA polymerase (5 U/µL) 1 U 0.2
Ultrapure water NA 21.8
Template DNA NA 2
Total volume NA 50
Table 18: Multiplex I PCR
Table for making Multiplex I PCR reaction mix.
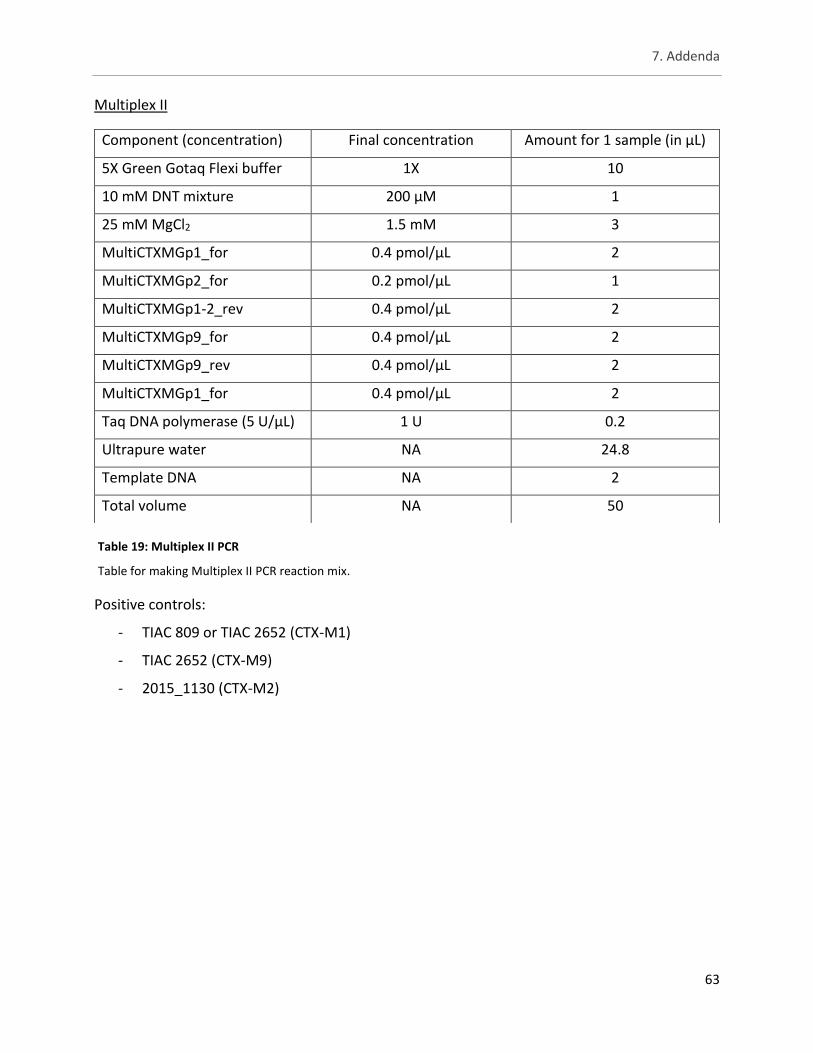
7. Addenda
63
Multiplex II
Positive controls:
- TIAC 809 or TIAC 2652 (CTX-M1)
- TIAC 2652 (CTX-M9)
- 2015_1130 (CTX-M2)
Component (concentration) Final concentration Amount for 1 sample (in µL)
5X Green Gotaq Flexi buffer 1X 10
10 mM DNT mixture 200 µM 1
25 mM MgCl2 1.5 mM 3
MultiCTXMGp1_for 0.4 pmol/µL 2
MultiCTXMGp2_for 0.2 pmol/µL 1
MultiCTXMGp1-2_rev 0.4 pmol/µL 2
MultiCTXMGp9_for 0.4 pmol/µL 2
MultiCTXMGp9_rev 0.4 pmol/µL 2
MultiCTXMGp1_for 0.4 pmol/µL 2
Taq DNA polymerase (5 U/µL) 1 U 0.2
Ultrapure water NA 24.8
Template DNA NA 2
Total volume NA 50
Table 19: Multiplex II PCR
Table for making Multiplex II PCR reaction mix.
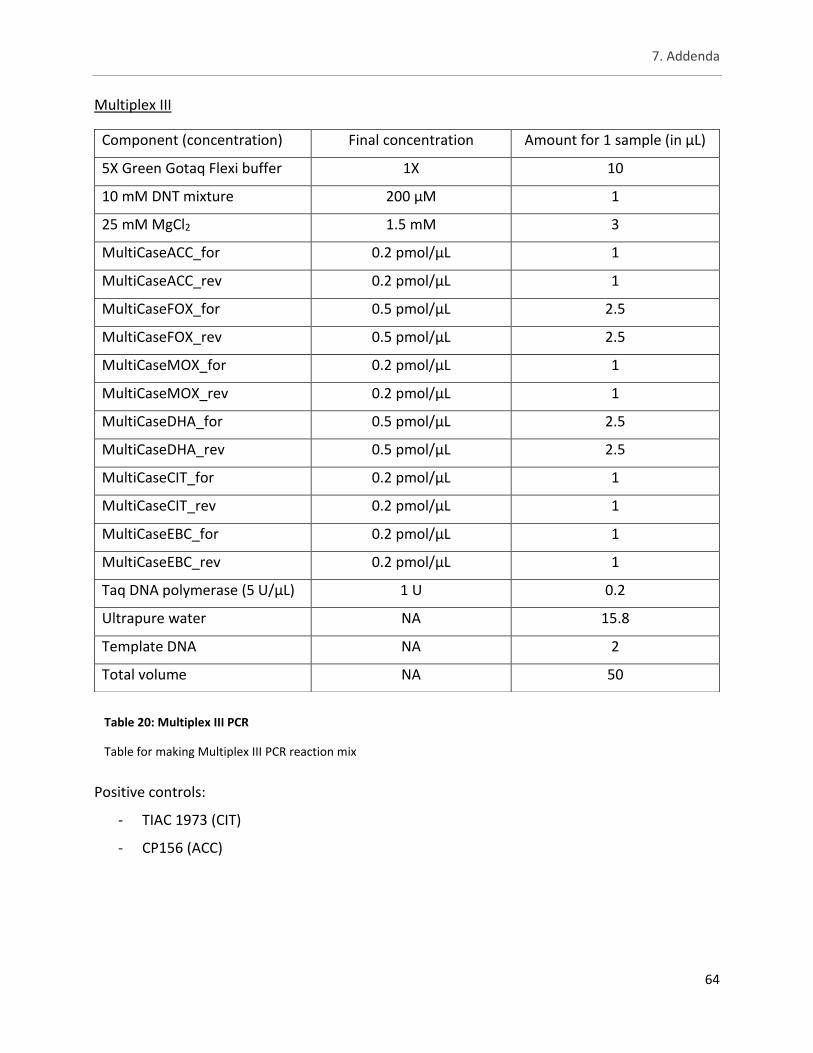
7. Addenda
64
Multiplex III
Positive controls:
- TIAC 1973 (CIT)
- CP156 (ACC)
Component (concentration) Final concentration Amount for 1 sample (in µL)
5X Green Gotaq Flexi buffer 1X 10
10 mM DNT mixture 200 µM 1
25 mM MgCl2 1.5 mM 3
MultiCaseACC_for 0.2 pmol/µL 1
MultiCaseACC_rev 0.2 pmol/µL 1
MultiCaseFOX_for 0.5 pmol/µL 2.5
MultiCaseFOX_rev 0.5 pmol/µL 2.5
MultiCaseMOX_for 0.2 pmol/µL 1
MultiCaseMOX_rev 0.2 pmol/µL 1
MultiCaseDHA_for 0.5 pmol/µL 2.5
MultiCaseDHA_rev 0.5 pmol/µL 2.5
MultiCaseCIT_for 0.2 pmol/µL 1
MultiCaseCIT_rev 0.2 pmol/µL 1
MultiCaseEBC_for 0.2 pmol/µL 1
MultiCaseEBC_rev 0.2 pmol/µL 1
Taq DNA polymerase (5 U/µL) 1 U 0.2
Ultrapure water NA 15.8
Template DNA NA 2
Total volume NA 50
Table 20: Multiplex III PCR
Table for making Multiplex III PCR reaction mix
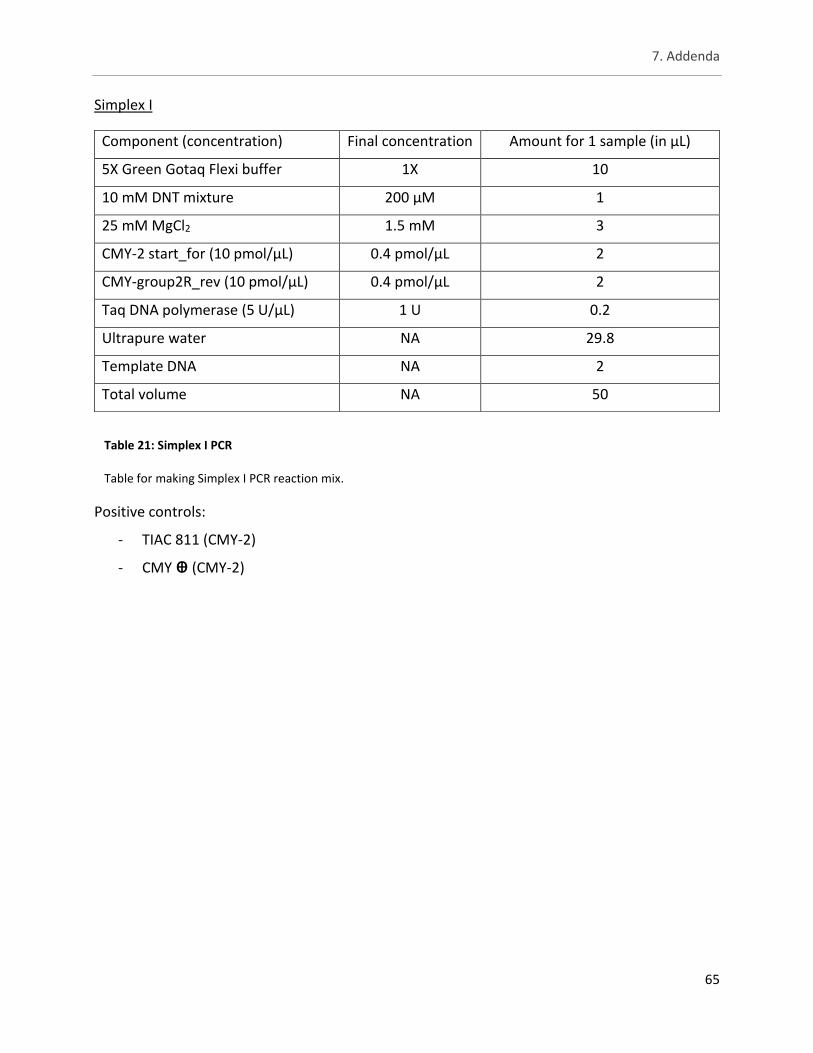
7. Addenda
65
Simplex I
Positive controls:
- TIAC 811 (CMY-2)
- CMY ꚛ (CMY-2)
Component (concentration) Final concentration Amount for 1 sample (in µL)
5X Green Gotaq Flexi buffer 1X 10
10 mM DNT mixture 200 µM 1
25 mM MgCl2 1.5 mM 3
CMY-2 start_for (10 pmol/µL) 0.4 pmol/µL 2
CMY-group2R_rev (10 pmol/µL) 0.4 pmol/µL 2
Taq DNA polymerase (5 U/µL) 1 U 0.2
Ultrapure water NA 29.8
Template DNA NA 2
Total volume NA 50
Table 21: Simplex I PCR
Table for making Simplex I PCR reaction mix.
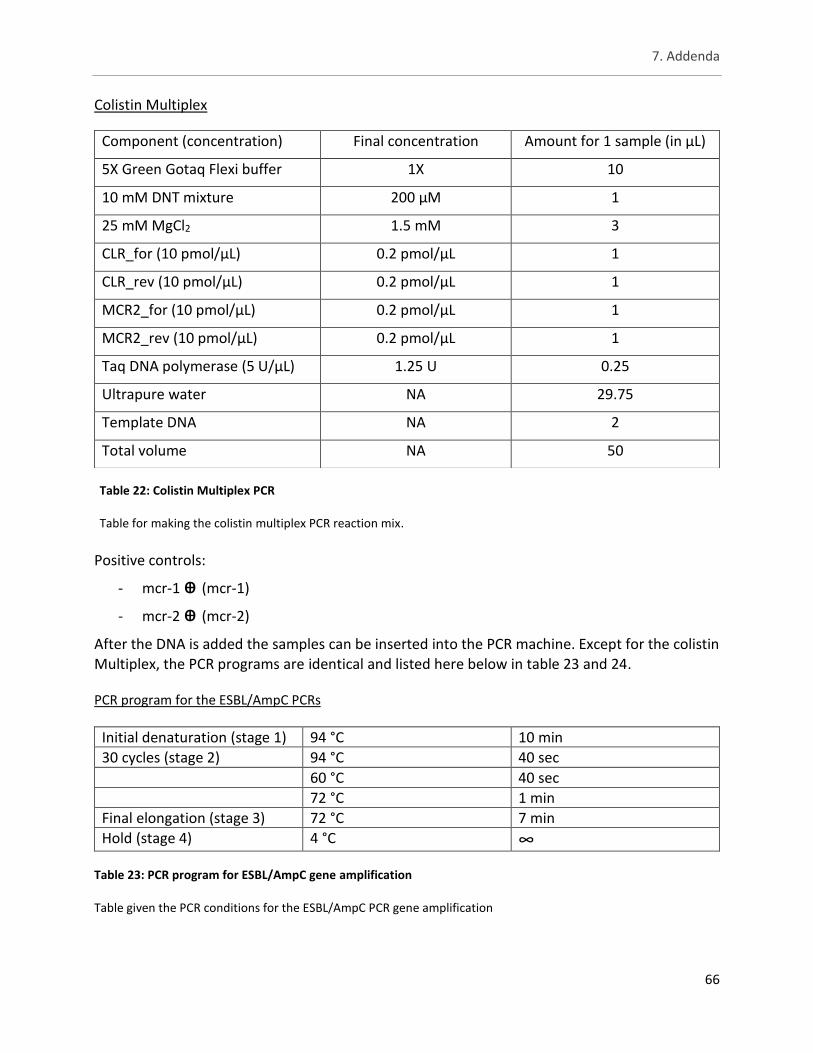
7. Addenda
66
Colistin Multiplex
Positive controls:
- mcr-1 ꚛ (mcr-1)
- mcr-2 ꚛ (mcr-2)
After the DNA is added the samples can be inserted into the PCR machine. Except for the colistin Multiplex, the PCR programs are identical and listed here below in table 23 and 24. PCR program for the ESBL/AmpC PCRs
Initial denaturation (stage 1) 94 °C 10 min
30 cycles (stage 2) 94 °C 40 sec
60 °C 40 sec
72 °C 1 min
Final elongation (stage 3) 72 °C 7 min
Hold (stage 4) 4 °C ∞
Table 23: PCR program for ESBL/AmpC gene amplification
Table given the PCR conditions for the ESBL/AmpC PCR gene amplification
Component (concentration) Final concentration Amount for 1 sample (in µL)
5X Green Gotaq Flexi buffer 1X 10
10 mM DNT mixture 200 µM 1
25 mM MgCl2 1.5 mM 3
CLR_for (10 pmol/µL) 0.2 pmol/µL 1
CLR_rev (10 pmol/µL) 0.2 pmol/µL 1
MCR2_for (10 pmol/µL) 0.2 pmol/µL 1
MCR2_rev (10 pmol/µL) 0.2 pmol/µL 1
Taq DNA polymerase (5 U/µL) 1.25 U 0.25
Ultrapure water NA 29.75
Template DNA NA 2
Total volume NA 50
Table 22: Colistin Multiplex PCR
Table for making the colistin multiplex PCR reaction mix.

7. Addenda
67
PCR program for the colistin PCRs
Initial denaturation (stage 1) 94 °C 15 min
25 cycles (stage 2) 94 °C 30 sec
58 °C 90 sec
72 °C 60 sec
Final elongation (stage 3) 72 °C 10 min
Hold (stage 4) 4 °C ∞
- Immediately perform gel electrophoresis or store the PCR samples in the fridge at 4 °C.
7.1.6.3 Gel electrophoresis for the detection of resistance genes
Double gloves, overshoes and an apron have to be worn when working in the ethidiumbromide zone. 1.5 % agarose gel are used. Be careful not to expose your eyes directly to UV light.
- Weight 3.15 g agarose (for a 15 x 20 tray, 0.75 cm). - Add 210 mL TBE buffer.
- Microwave for approximately 2 min at 800 Watt. o Liquid must be clear and all the agarose dissolved. o Leave the cap loose. o Gel can be made in advanced and kept warm in a 50 °C water bath.
- Poor the gel in the gel tray and add combs (3 rows of 20 wells). o Make sure the gelholder is tight. o Leave enough space between the combs. o Be careful for air bubbles, push them to the side if formed.
- Let the gel become stiff (30 min). - Remove combs.
o careful not to break the wells. - Load 10 µL of ladder in the first row.
o here a 100 bp ladder. - Load 10 µL sample DNA from the PCR reaction on the gel. - Load 10 µL control DNA from the PCR reaction in the last wells. - Carefully place gel in tank with TBE buffer.
o Make sure gel is completely under, if needed add some more buffer. o Alternatively the gel can be loaded before placing in the tank
- Place the lit on the tank and place the electrodes: black (-) up top and red (+) at the bottom.
Table 24: PCR program for colistin plasmid-mediated gene amplification
Table given the PCR conditions for the colistin plasmid-mediated PCR gene amplification

7. Addenda
68
- Let the gel run for about an hour on 120V.
o Do not let the gel run too long or all the bands will be concentrated at the end of the gel.
- Remove the gel from the tank and place it in the ethidiumbromide area. o Wear protective gear in the ethidiumbromide area!
- Immerse the gel in the EtBr bath for staining. o 30 to 45 min.
- Remove the gel from the bath and place it into the UV transilluminator. o Close the cover before turning the UV light on.
- Take picture using the camera. - Throw gel in yellow bin. - Process image using Quantity one software.
7.2 Composition of media
7.2.1 XLD agar
Yeast extract 3.0 g/liter
L-Lysine HCl 5.0 g/liter
Xylose 3.75 g/liter
Lactose 7.5 g/liter
Sucrose 7.5 g/liter
Sodium desoxycholate 1.0 g/liter
Sodium chloride 5.0 g/liter
Sodium thiosulphate 6.8 g/liter
Ferric ammonium citrate 0.8 g/liter
Phenol red 0.08 Agar 12.5 g/liter
pH 7.4 ± 0.2 at 25°C
7.2.2 TSB
Pancreatic digest of casein 17.0 g/liter
Enzymatic digest of soya bean 3.0 g/liter
Sodium chloride 5.0 g/liter
Dipotassium hydrogen phosphate 2.5 g/liter
Glucose 2.5 g/liter
pH 7.3 ± 0.2 at 25°C

7. Addenda
69
7.2.3 PCA
Tryptone 5.0 g/liter
Yeast extract 2.5 g/liter
Glucose 1.0 g/liter
Agar 9.0 g/liter
pH 7.0 ± 0.2 at 25°C
7.2.4 TBX agar
Tryptone 20.0 g/liter
Bile Salts No. 3 1.5 g/liter
Agar 15.0 g/liter
X-glucuronide 0.075 g/liter
pH 7.2 ± 0.2 @ 25 °C
7.2.5 MH II agar
Beef Extract 2.0 g/liter
Acid hydrolysate of Casein 17.5 g/liter
Starch 1.5 g/liter
Agar 17.0 g/liter
pH 7.3 ± 0.1 @ 25 °C
7.2.5 MH II broth
Beef Extract 3.0 g/liter
Acid hydrolysate of Casein 17.5 g/liter
Starch 1.5 g/liter
Agar 17.0 g/liter
pH 7.3 ± 0.2 @ 25 °C

7. Addenda
70
7.3 Preparation of media
Plates are poured in a laminar flow. Media is poured in a tube for pH testing without compromising the sterility of the Erlenmeyer.
7.3.1 XLD agar (for 1 L)
- Weight 53 g of the XLD agar medium in an Erlenmeyer.
- Suspend this in 1 liter of distilled water.
- Mix well using magnetic stirrer.
- Heat with frequent agitation until the medium boils.
o DO NOT OVERHEAT.
o Mixture will be ready when colour starts becoming clearer.
o Once heated treat mixture as sterile.
- Transfer to a water bath at 50°C.
o Supplements can be added as this point.
o Agar can be kept in this bath if immediate pouring is not possible.
- Verify pH level.
- Pour into sterile Petri dishes.
o Careful to not let agar cool off too much.
- Let the plates dry as to solidify the agar.
- Take a blanc to test for sterility and put rest in the fridge at 4 °C.
7.3.2 TSB (for 1 L)
- Weight 30 g TSB in an Erlenmeyer.
- Add 30g to 1 litre of distilled water.
- mix well using magnetic stirrer.
- Distribute into final containers.
- Sterilize by autoclaving at 121°C for 15 minutes.
- Verify pH.
- Store in fridge at 4 °C.
7.3.3 PCA (for 1 L)
- Weight 17.5 g PCA medium in an Erlenmeyer.
- Add 1 L of distilled water to the Erlenmeyer.
- mix well using magnetic stirrer.

7. Addenda
71
- Heat with frequent agitation until the medium boils.
o DO NOT OVERHEAT.
o Mixture will be ready when colour becomes clear.
- Sterilize by autoclaving at 121 °C for 15 minutes.
- Transfer to a water bath at 50°C.
o Supplements can be added as this point.
o Agar can be kept in this bath if immediate pouring is not possible.
- Verify pH level.
- Pour into sterile Petri dishes.
o Careful to not let agar cool off too much.
- Let the plates dry as to solidify the agar.
- Take a blanc to test for sterility and put rest in the fridge at 4 °C.
7.3.4 TBX agar (for 1 L)
- Weight 36.6 g PCA medium in an Erlenmeyer.
- Add 1 L of distilled water to the Erlenmeyer.
- mix well using magnetic stirrer.
- Heat with frequent agitation until the medium boils.
o DO NOT OVERHEAT.
o Mixture will be ready when colour becomes clear.
- Sterilize by autoclaving at 121 °C for 15 minutes.
- Transfer to a water bath at 50°C.
o Supplements can be added as this point.
o Agar can be kept in this bath if immediate pouring is not possible.
- Verify pH level.
- Pour into sterile Petri dishes.
o Careful to not let agar cool off too much.
- Let the plates dry as to solidify the agar.
- Take a blanc to test for sterility and put rest in the fridge at 4 °C.

7. Addenda
72
7.3.5 MH II agar (for 1 L)
- Weight 38 g MH II agar in an Erlenmeyer.
- Add 1 L of distilled water to the Erlenmeyer.
- mix well using magnetic stirrer.
- Heat with frequent agitation until the medium boils.
o DO NOT OVERHEAT.
o Mixture will be ready when colour becomes clear.
- Sterilize by autoclaving at 121 °C for 15 minutes.
- Transfer to a water bath at 50°C.
o Supplements can be added as this point.
o Agar can be kept in this bath if immediate pouring is not possible.
- Verify pH level.
- With a pipet pour 22 mL into sterile Petri dishes.
o Careful to not let agar cool off too much.
- Let the plates dry as to solidify the agar.
- Take a blanc to test for sterility and put rest in the fridge at 4 °C.
7.3.6 MH II broth (for 1 L)
- Weight 22 g MH II broth in an Erlenmeyer.
- Add 1 L of distilled water to the Erlenmeyer.
- mix well using magnetic stirrer.
- Heat with frequent agitation until the medium boils.
o DO NOT OVERHEAT.
o Mixture will be ready when colour becomes clear.
- Divide broth into tubes (10 mL)
- Sterilize by autoclaving at 121 °C for 10 minutes.
- Verify pH level.
- Take a blanc to test for sterility and put rest in the fridge at 4 °C.





























