Embed Size (px)
Citation preview

Butanol Production from Lignocellulosic Feedstocks by Acetone-Butanol-Ethanol Fermentation with Integrated Product Recovery
Dissertation
Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduation School of The Ohio State University
By
Congcong Lu, M.S.
Graduate Program in Chemical and Biomolecular Engineering
The Ohio State University
2011
Dissertation Committee:
Professor Shang-Tian Yang, Advisor
Professor Jeffrey Chalmers
Professor Andre Palmer
1

Copyright by
Congcong Lu
2011
2

ii
Abstract
n-Butanol has been attracting research attention as a liquid biofuel recently, in
addition to its current application as an industrial chemical and solvent. With the concerns
of diminishing fossil reserves, environmental issues caused by greenhouse gas emission,
and unstable supply and price spike of crude oil, renewed interests have returned to
pursue biobutanol production through acetone-butanol-ethanol (ABE) fermentation as
opposed to petrochemically-derived butanol. However, the conventional ABE
fermentation suffers from many limitations, including low butanol titer, high cost of
traditional food-based raw materials, end-product inhibition and high butanol recovery
cost by distillation, which negatively impacts the process efficiency and economics.
Fortunately, these hurdles are being overcome by technological advances on ABE
fermentation in the past few decades.
Research on genetic modifications and chemical mutation of solventogenic
Clostridia has focused on obtaining mutant strains with enhanced butanol producing
ability. Adequate research success in utilizing renewable and sustainable lignocellulosic
biomass has identified a novel group of cost-effective feedstocks for ABE fermentation in
replacement of the traditional costly starch and sugar-based substrates. Novel fed-batch

iii
and continuous fermentation processes with cell immobilization and cell recycle have
been developed for more efficient substrate conversion and butanol production. When
further integrated with alternative energy-efficient butanol recovery techniques, such as
gas stripping and pervaporation, the integrated ABE fermentation process can achieve
high overall butanol production, reactor productivity, sugar conversion, and simplified
downstream separation.
Therefore, the overall goal of this project was to develop a process to produce
butanol through ABE fermentation by hyper-butanol-producing mutants using
lignocellulosic biomass, and integrate online product recovery to achieve enhanced
overall butanol production and process efficiency. Corn fiber, cassava bagasse, wood pulp
and sugarcane bagasse were investigated as potential feedstocks for butanol production
from ABE fermentation, and gas stripping as the online butanol recovery technique was
evaluated and integrated with ABE fermentation. In batch fermentations with the mutant
strain JB200, which was derived from C. beijerinckii ATCC 55025, immobilized in a
fibrous bed bioreactor, 12.7 g/L and 15.4 g/L ABE were obtained using corn fiber
hydrolysate and cassava bagasse hydrolysate, respectively. For wood pulp hydrolysate
and sugarcane bagasse hydrolysate, which contained significant amounts of inhibitors
from acid pretreatment, C. beijerinckii CC101 (an adaptant derived from NCIMB 8052)
and its recombinant mutant strain CC101-SV6, were able to produce 11.35 g/L and 9.44
g/L ABE in free-cell batch fermentations, respectively. ABE production from wood pulp

iv
hydrolysate was further enhanced to 17.73 g/L in a gas stripping integrated ABE batch
fermentation process, with a higher ABE yield of 0.44 g/g compared with 0.39 g/g from
non-integrated control study. Concentrated cassava bagasse hydrolysate containing 584.4
g/L glucose was utilized by the mutant strain JB 200 in an integrated fed-batch ABE
fermentation process, and 90.3 g/L ABE were produced with a productivity of 0.53 g/L. h,
which was further improved to 108.5 g/L with nutrient supplementation.
This project demonstrated that butanol can be produced from various lignocellulosic
feedstocks, from agricultural biowastes to woody biomass residues, with a high yield and
at a high titer using selected mutant strains of C. beijerinckii. By employing mutant
strains of solventogenic Clostridia bacteria, different fermentation modes, and gas
stripping as online product recovery, an integrated process was developed for the
production of n-butanol that can potentially replace petroleum-based butanol.

v
Dedication
Dedicated to my parents

vi
Acknowledgements
First of all, I would like to thank my advisor, Dr. Shang-Tian Yang, for his guidance,
encouragement, patience, and full support during my entire graduate study. I am sincerely
thankful and grateful for all his help academically and financially throughout my Ph.D.
study. I have never met a person of his graciousness and admirable personality. It has
always been a great honor to have him as my advisor both in academia and in life. He set
up an example to look up to as an excellent scientific researcher and a fantastic leader,
and I have truly learned and benefited a lot from him. For this, I will eternally be grateful.
I would also like to thank Dr. Jeffrey Chalmers and Dr. Andre Palmer for taking time
to be on my committee, as well as their valuable recommendations and advice to my
research project.
I would like to acknowledge Dr. Jingbo Zhao for teaching me all the hands-on
techniques and knowledge essential to operating anaerobic ABE fermentation at the
beginning of my Ph.D. study, and Dr. Chuang Xue for his help on setting up the gas
stripping apparatus. I would also like to thank all the previous and current laboratory
members in our research group, especially Dr. Wei-lun Chang, Dr. Mingrui Yu,
Ching-suei Hsu, Baohua Zhang and Zhongqiang Wang for their helpful suggestions,

vii
support and encouragement.
In addition, I would like to specially thank for all the help and valuable suggestions
from Vennie Tee at ButylFuelTM LLC, and the lignocellulosic hydrolysates kindly
provided by ButylFuel. I would also like to thank Dr. Dong Wei from South China
University of Technology for providing cassava bagasse, and Saju Varghese for
constructing the plasmid for the mutant strain of C. beijerinckii CC101-SV6.
Financial supports from the Ohio Department of Development Third Frontier
Advanced Energy Program and Ohio State University Graduate School fellowship
are deeply appreciated.
Finally, I would like to thank my parents, Mr. Yi Lu and Mrs. Yue Tan, my
grandparents, my relatives and all my friends for their faith and support in me.

viii
Vita
June 2003………………………………………Yantai No.2 senior high
2003 – 2007…………………………………… B.S. Materials Science and Engineering,
Donghua University
2007 – 2008…………………………………….Graduate Fellowship, The Ohio State
University
2008 – 2010……………………………………. Graduate Research Associate,
Department of Chemical and
Biomolecular Engineering, The Ohio
State University
2010 – present…………………………………..Graduate Fellowship, The Ohio State
University
Fields of Study
Major Field: Chemical and Biomolecular Engineering

ix
Table of Contents
Abstract……………………………………………………………………………………ii
Dedication…………………………………………………………………………………v
Acknowledgements……………………………………………………………………….vi
Vita………………………………………………………………………………………viii
Table of Contents…………………………………………………………………………ix
List of Tables………………………………………………………………......…….…xvii
List of Figures………………………........……………………………………..………..xx
Chapter 1: Introduction…………………………………………………………………....1
1.1 Project goals and specific tasks……………………………………………….5
1.2 Significance and major impacts……………………………………………….7
1.3 References…………………………………………………………….............8
Chapter 2: Literature Review…………………………………………………………….14
2.1 Acetone-Butanol-Ethanol (ABE) fermentation………………………..…….14
2.1.1 Microorganisms and strain improvements…………………….......16
2.1.2 Traditional substrates and renewable lignocellulosic feedstocks….20
2.1.3 Developments in fermentation process………………………...….23

x
2.2 Pretreatment and detoxification of lignocellulosic feedstocks………………28
2.2.1 Pretreatment of lignocellulose……………………………………..28
2.2.2 Detoxification of lignocellulosic hydrolysate……………………..34
2.3 Product recovery and separation technologies………………………………38
2.3.1 Gas stripping……………………………………………………….40
2.3.2 Pervaporation………………………………………………………44
2.3.3 Liquid-liquid extraction……………………………………………50
2.3.4 Adsorption…………………………………………………………55
2.4 Integrated ABE fermentation process with online product recovery………..58
2.5 References…………………………………………………………………...59
Chapter 3: Butanol Production from Corn Fiber Hydrolysate by Clostridium beijerinckii
in a Fibrous Bed Bioreactor………………………………………………….96
3.1 Introduction………………………………………………………………….97
3.2 Materials and methods……………………………………………………….99
3.2.1 Hydrolysis of corn fiber…………………………………………...99
3.2.2 Detoxification ………………………………………………...….100
3.2.3 Culture and media ……………………………………………….100
3.2.4 Fermentation and cell immobilization in fibrous bed bioreactor...101
3.2.5 Analytical methods……………………………………………….103
3.3 Results and discussion……………………………………………………...104

xi
3.3.1 ABE fermentation in glucose, xylose, and glucose/xylose mixture
medium………………………………………………………………....104
3.3.2 ABE fermentation in undetoxified CFH-based medium…………106
3.3.3 ABE fermentation in boiling and activated carbon detoxified
CFH-based medium…………………………………………………….109
3.4 Conclusion………………………………………………………………….112
3.5 References………………………………………………………………….114
Chapter 4: Evaluation of Butanol Recovery by Gas Stripping from Model solution and
Fermentation Broth…………………………………………………………124
4.1 Introduction……………………………………………………………...…125
4.2 Materials and methods……………………………………………………...121
4.2.1 Experimental setup and process design…………………………..128
4.2.2 Analytical methods……………………………………………….129
4.3 Results and discussion……………………………………………………...130
4.3.1 Effect of cooling temperature…………………………………….130
4.3.2 Effect of gas flow rate……………………………………………133
4.3.3 Effect of cells and components from fermentation broth………...135
4.3.4 Selectivity of acetone, butanol, and ethanol……………………...137
4.4 Conclusion………………………………………………………………….140
4.5 References………………………………………………………………….140

xii
Chapter 5: Fed-batch Fermentation for Butanol Production from Cassava Bagasse
Hydrolysate in a Fibrous Bed Bioreactor with Continuous Gas Stripping..153
5.1 Introduction………………………………………………………………...154
5.2 Materials and methods……………………………………………………...156
5.2.1 Enzymatic hydrolysis of cassava bagasse………………………..156
5.2.2 Strain and medium preparation…………………………………..158
5.2.3 Experimental setup……………………………………………….159
5.2.4 Cell immobilization in fibrous bed bioreactor…………………...160
5.2.5 Batch and integrated fed-batch fermentations……………………160
5.2.6 Analytical methods……………………………………………….162
5.3 Results and Discussion……………………………………………………..163
5.3.1 Batch fermentation kinetics………………………………………163
5.3.2 Fed-batch fermentation with simultaneous product removal by gas
stripping………………………………………………………………...165
5.4 Conclusion………………………………………………………………….171
5.5 References………………………………………………………………….172
Chapter 6: Biobutanol Production from Renewable Wood Pulp Hydrolysate in an
Integrated Process: Evaluation of Detoxifications and Alternative Nitrogen
Sources…………………………………………………………………….183
6.1 Introduction………………………………………………………………...184

xiii
6.2 Materials and methods……………………………………………………...187
6.2.1 Strain and inoculum preparation…………………………………187
6.2.2 Wood pulp hydrolysate…………………………………………...188
6.2.3 Detoxification procedures………………………………………..188
6.2.4 Preparation of alternative nitrogen sources………………………189
6.2.5 Production medium preparation and fermentation……………….191
6.2.6 Analytical methods……………………………………………….193
6.3 Results and discussion……………………………………………………...195
6.3.1 Effect of dilution and detoxifications on WPH compositions and
ABE fermentation…….…………………………………………195
6.3.2 Investigation of potential economic nitrogen source…………….201
6.3.3 ABE production on the bioreactor integrated with gas stripping as
product recovery…………………………………………………205
6.4 Conclusion………………………………………………………………….209
6.5 References………………………………………………………………….209
Chapter 7: Enhanced Biological Butanol Production and Acid Assimilation in ABE
Fermentation using a Recombinant Mutant of Clostridium beijerinckii….223
7.1 Introduction………………………………………………………………...224
7.2 Materials and methods………………………………………….…………..228
7.2.1 Plasmid construction……………………………………………..228

xiv
7.2.2 Strain and inoculum preparation…………………………………228
7.2.3 Production medium preparation and fermentation...……………..229
7.2.4 Analytical methods…………………………………………….....231
7.3 Results and discussion……………………………………………………...232
7.3.1 ABE production from glucose-P2 medium with parental and mutant
strains……………………………………………………………...232
7.3.2 Effect of cysteine addition on ABE production..............................235
7.3.3 Effect of butyric acid addition on ABE production……...……….238
7.3.4 ABE production from renewable lignocellulosic substrates…...240
7.4 Conclusion………………………………………………………………….243
7.5 References………………………………………………………………….244
Chapter 8: Conclusions and Recommendations..............................................................257
8.1 Conclusions...................................................................................................257
8.1.1 Butanol production by engineered mutant strains..........................257
8.1.2 Butanol production from lignocellulosic feedstocks......................258
8.1.3 Gas stripping as an alternative butanol recovery technique...........259
8.1.4 Enhanced butanol production in the integrated fermentation process
with online product recovery.........................................................260
8.2 Recommendations.........................................................................................261
8.2.1 Improvement on the fermentability of the lignocellulosic

xv
hydrolysates.................................................................................261
8.2.2 Optimization of butanol recovery by gas stripping and investigation
on alternative recovery techniques................................................262
8.2.3 Process development on ABE fermentation...................................263
Bibliography.....................................................................................................................265
Appendix A: Analytical Procedures.................................................................................289
A.1 Gas chromatograph.......................................................................................289
A.2 High performance liquid chromatograph......................................................290
Appendix B: Evaluation of Liquid-liquid Extraction Using Ionic Liquid for Butanol
Recovery..................................................................................................306
B.1 Materials and methods..................................................................................306
B.2 Results and discussion..................................................................................307
B.3 References.....................................................................................................310
Appendix C: pSV6 Plasmid Construction and Transformation into Clostridium
beijerinckii CC101.................................................................................317
C.1 PCR amplification of the truncated sol operon from C. beijerinckii
CC101..........................................................................................................317
C.2 Cloning of the t-SOL into pCR2.1 vector.....................................................317
C.3 Cloning of the FRT-Hyg-FRT into pMTL-thl-adhE2...................................318
C.4 Cloning of the t-SOL(ald + ctfA + ctfB) into pSV4 vector...........................319

xvi
C.5 Transformation of pSV6 plasmid into E. coli CAC434................................320
C.6 Conjugation of pSV6 harboring E. coli CAC434 cells into C. beijerinckii
CC101 by filter mating................................................................................321

List of Tables
Table 2.1 Important fuel properties of butanol, ethanol, methanol and gasoline………...81
Table 2.2 Summary of various solventogenic Clostridia with their substrates, products,
fermentation pH and temperature……………………………………………..82
Table 2.3 ABE production by solventogenic Clostridia from traditional substrates and
lignocellulosic biomass………………………………………………………..83
Table 2.4 Compositions of different lignocellulosic biomass and their current use……..84
Table 2.5 Advances in fermentation process with cell immobilization and cell recycle...85
Table 2.6 Comparison of leading pretreatment methods for improving the digestibility of
lignocellulosic materials………………………………………………………86
Table 2.7 Major fermentation inhibitors present in the hydrolysates generated from
lignocellulose degradation.................................................................................87
Table 2.8 Alternative separation techniques for butanol recovery from ABE
fermentation………………………………………………………………….88
Table 2.9 Solvent selectivities and operating conditions for butanol recovery in the gas
stripping processes...........................................................................................89
Table 2.10 Comparison of membrane performances for butanol recovery in the
xvii

xviii
pervaporation processes ..................................................................................90
Table 2.11 Solvents evaluation as extractants for butanol recovery by liquid-liquid
extraction and their toxicity towards Clostridium beijerinckii........................91
Table 2.12 Performances and capacities of different adsorbent materials for butanol
recovery by adsorption.....................................................................................92
Table 2.13 Integrated processes for enhanced ABE production from various substrates
and strains........................................................................................................93
Table 3.1 ABE fermentation by C. beijerinckii JB 200 using glucose, xylose,
glucose-xylose mixture and CFH....………………………………………...117
Table 3.2 Butanol production from different lignocellulosic hydrolysates……………..118
Table 5.1 Composition of different cassava bagasse hydrolysate used in this study…...175
Table 5.2 ABE production from cassava bagasse hydrolysate in batch fermentation and
integrated fed-batch fermentation by C. beijerinckii JB 200………………...176
Table 5.3 Summary of performance of each cycle in the integrated fed-batch
fermentations………………………………………………………………177
Table 6.1 Compositional analysis of the original WPH, detoxified WPHs, CSL, CPH,
SMH and molasses used in this study..............................................................214
Table 6.2 Comparison and summary of ABE production using different WPHs in batch
and integrated batch studies by Clostridium beijerinckii CC101....................215
Table 6.3 Evaluation of alternative nitrogen sources and ammonia acetate on ABE

xix
production using 70% resin and evaporation detoxified WPH by Clostridium
beijerinckii CC101....................................................................................….216
Table 7.1 Comparison and summary of the performance of parental strain and mutant
strain under all the scenarios evaluated in this study..........................….......250
Table B.1 Distribution coefficient of butanol, acetone, and butyric acid from different
extractants in model solution and fermentation broth....................................312
Table B.2 Selectivity of butanol over acetone and butyric acid from different extractants
in model solution and fermentation broth........................................................313

xx
List of Figures
Figure 1.1 Overview of project goal and major tasks carried out in this study………….13
Figure 2.1 Metabolic pathway of Clostridium acetobutylicum from glucose to acids and
solvents during acidogensis and solventogensis……………………………..94
Figure 2.2 Alternative butanol recovery processes: A. Gas stripping, B. Pervaporation, C.
Liquid-liquid extraction, D. Adsorption……………………………………..95
Figure 3.1 ABE production from glucose, xylose and glucose-xylose mixture by C.
beijerinckii JB 200. (A) Glucose, (B) Xylose, (C) Glucose-xylose
mixture…………………………………………………………………….119
Figure 3.2 ABE production from diluted undetoxified CFH by C. beijerinckii JB 200. (A)
Sugar utilization, (B) Solvents and acids production………………………121
Figure 3.3 ABE production from diluted detoxified CFH by C. beijerinckii JB 200. (A)
Sugar utilization, (B) Solvents and acids production……………………….122
Figure 3.4 ABE production from undiluted detoxified CFH with C. beijerinckii JB 200.
(A) Sugar utilization, (B) Solvents and acids production…………………..123
Figure 4.1 Schematic diagram of the gas stripping process…………………………….144
Figure 4.2 Effect of cooling temperature on gas stripping performance with model

xxi
solution. (a) Concentration of solvents in the feed vs. time, (b)
Acetone/butanol removal rate vs. acetone/butanol concentration, (c)
Concentration of solvents in the condensate vs. in the feed………………145
Figure 4.3 Effect of gas flow rate on gas stripping performance with model solution. (a)
Concentration of solvents in the feed vs. time, (b) Acetone/butanol removal
rate vs. acetone/butanol concentration, (c) Concentration of solvents in the
condensate vs. in the feed…………………………………………………..147
Figure 4.4 Effect of cells and other fermentation components on gas stripping
performance with model solution and fermentation broth. (a) Concentration
of solvents in the feed vs. time, (b) Acetone/butanol removal rate vs.
acetone/butanol concentration, (c) Concentration of solvents in the
condensate vs. in the feed…………………………………………………149
Figure 4.5 Selectivity of solvents under the conditions evaluated in this study. (a) Butanol
selectivity vs. concentration, (b) Acetone selectivity vs. concentration, (c)
Ethanol selectivity vs. concentration……………………………………….151
Figure 5.1 Experimental setup of FBB-connected fermentor with gas stripping as online
butanol recovery…………………………………………………………….178
Figure 5.2 ABE Batch fermentation from different carbon sources by C. beijerinckii JB
200. (A) Glucose, (B) Cassava bagasse hydrolysate, (C) Cassava bagasse
hydrolysate supplemented with additional glucose………………………...179

xxii
Figure 5.3 Fed-batch fermentation integrated with gas stripping using CCBH by C.
beijerinckii JB 200. (A) Glucose and products in the reactor vs. time, (B)
Composition of the condensate recovered in each cycle…………………181
Figure 5.4 Fed-batch fermentation integrated with gas stripping using CCBH by C.
beijerinckii JB 200 with periodical nutrient supplementation. (A) Glucose and
products in the reactor vs. time, (B) Composition of the condensate recovered
in each cycle…………………………………………………………...……182
Figure 6.1 Effect of dilution and different detoxification methods on ABE fermentation
by Clostridium beijerinckii CC101 using WPH. (a) Butanol and total ABE
production, (b) Butanol yield, ABE yield, and sugar conversion…………..217
Figure 6.2 Effect of alternative nitrogen sources and ammonia acetate on ABE production
using 70% resin and evaporation detoxified WPH by Clostridium beijerinckii
CC101. (a) Butanol production, (b) Butanol yield…..............................218
Figure 6.3 ABE production from 70% WPH on the bioreactor integrated with gas
stripping by Clostridium beijerinckii CC101. (a) Solvents and acids
production, (b) Sugar consumption, (c) Composition of the recovered
condensate at different time intervals…..................................................…219
Figure 6.4 ABE production from resin and evaporation detoxified WPH on the bioreactor
integrated with gas stripping by Clostridium beijerinckii CC101. (a) Solvents
and acids production, (b) Sugar consumption, (c) Composition of the

xxiii
recovered condensate at different time intervals….......................................221
Figure 7.1 Fermentation kinetics of C.beijerinckii CC101 and C. beijerinckii CC101-SV6
in synthetic glucose-P2 medium. (a) ABE production of C. beijerinckii CC101,
(b) ABE production of C. beijerinckii CC101-SV6.....................…..............251
Figure 7.2 Comparison of butanol production and acids assimilation from C. beijerinckii
CC101 and C. beijerinckii CC101-SV6 in synthetic glucose- P2 medium. (a)
Acid production, (b) Butanol production…………..........................……….252
Figure 7.3 Fermentation kinetics of C. beijerinckii CC101 and C. beijerinckii
CC101-SV6 in synthetic glucose-P2 medium with cysteine. (a) ABE
production of C. beijerinckii CC101, (b) ABE production of C. beijerinckii
CC101-SV6…..............................................................................253
Figure 7.4 Comparison of butanol production and acids assimilation from C. beijerinckii
CC101 and C. beijerinckii CC101-SV6 in glucose-P2 medium with cysteine.
(a) Acid production. (b) Butanol production.......................….......................254
Figure 7.5 Effect of cysteine and butyrate on ABE production with C. beijerinckii CC101
and C. beijerinckii CC101-SV6. (a) Effect on butanol production, (b) Effect on
residual acids..............................................…..................................................255
Figure 7.6 ABE production from sugarcane bagasse hydrolysate. (a) Fermentation
kinetics of C. beijerinckii CC101, (b) Fermentation kinetics of C. beijerinckii
CC101-SV6…..............................................................................................256

xxiv
Figure A.1 GC chromatogram of the standard sample containing acetone, butanol, ethanol,
acetic acid and butyric acid using external standard and internal standard
methods. (A) External standard method (1g/L each), (B) Internal standard
method (0.5 g/L each)....................................................................................292
Figure A.2 GC chromatogram of 10-fold diluted ABE fermentation sample containing 8.2
g/L acetone, 16.6 g/L butanol, 2.7 g/L ethanol, 5.1 g/L acetic acid and 2.9 g/L
butyric acid (External standard method)........................................................293
Figure A.3 GC chromatogram of 20-fold diluted ABE fermentation sample containing
2.54 g/L acetone, 8.15 g/L butanol, 0.21 g/L ethanol, 0.96 g/L acetic acid and
1.66 g/L butyric acid (Internal standard method)........................................294
Figure A.4 GC chromatogram of 100-fold diluted condensate sample from gas stripping
process containing 26.1 g/L acetone, 166.6 g/L butanol, 4.8 g/L ethanol, 0.46
g/L acetic acid, and 1.0 g/L butyric acid (External standard method).........295
Figure A.5 GC chromatogram of 200-fold diluted condensate sample recovered from gas
stripping process containing 30.9 g/L acetone, 137.3 g/L butanol, 2.1 g/L
ethanol, 0.7 g/L acetic acid and 0.8 g/L butyric acid (Internal standard
method)........................................................................................................296
Figure A.6 HPLC chromatogram of the standard sample containing glucose, xylose,
arabinose, acetic acid, lactic acid, butyric acid and butanol (2 g/L each)....297
Figure A.7 HPLC chromatogram of the 10-fold diluted corn fiber hydrolysate containing

xxv
27.0 g/L glucose, 22.7 g/L xylose, 11.2 g/L arabinose, 2.6 g/L acetic acid...298
Figure A.8 HPLC chromatogram of the 10-fold diluted cassava bagasse hydrolysate
containing 42.1 g/L glucose, 1.80 g/L xylose, 0.22 g/L lactic acid and 0.48
g/L acetic acid..............................................................................................299
Figure A.9 HPLC chromatogram of the 10-fold diluted wood pulp hydrolysate containing
9.1 g/L glucose, 39.7 g/L xylose, 2.1 g/L arabinose and 0.06 g/L acetic
acid.................................................................................................................300
Figure A.10 HPLC chromatogram of the 20-fold diluted sugarcane bagasse hydrolysate
containing 60.5 g/L glucose, 30.0 g/L xylose, 1.8 g/L arabinose, 0.2 g/L
lactic acid and 6.2 g/L acetic acid................................................................301
Figure A.11 HPLC chromatogram of 10-fold diluted ABE fermentation sample using
glucose and xylose as substrates containing 5.8 g/L glucose, 14.2 g/L xylose,
1.5 g/L acetic acid, 1.9 g/L butyric acid and 5.4 g/L butanol.....................302
Figure A.12 HPLC chromatogram of 10-fold diluted ABE fermentation sample using
corn fiber hydrolysate as substrate containing 19.8 g/L glucose, 11.5 g/L
xylose, 3.5 g/L arabinose, 1.0 g/L lactic acid, 5.7 g/L acetic acid, 5.8 g/L
butyric acid and 3.8 g/L butanol................................................................303
Figure A.13 HPLC chromatogram of 10-fold diluted ABE fermentation sample using
cassava bagasse hydrolysate as substrate containing 23.9 g/L glucose, 0.61
g/L xylose, 0.5 g/L lactic acid, 5.5 g/L acetic acid, 4.7 g/L butyric acid and

xxvi
7.3 g/L butanol...........................................................................................304
Figure A.14 HPLC chromatogram of 10-fold diluted ABE fermentation sample using
wood pulp hydrolysate as substrate containing 2.4 g/L glucose, 13.5 g/L
xylose, 2.1 g/L arabinose, 1.5 g/L lactic acid, 1.6 g/L acetic acid, 0.7 g/L
butyric acid and 4.5 g/L butanol................................................................305
Figure B.1 Concentration of acetone, butanol, ethanol, acetic acid, and butyric acid in the
aqueous solution before and after the liquid-liquid extraction. (A) Model
solution, (B) Fermentation broth....................................................................314
Figure B.2 Comparison of butanol and acetone distribution coefficient of different
extractants in model solution and fermentation broth. (A) Dbutanol, (B)
Dacetone..........................................................................................................315
Figure B.3 Comparison of butanol/acetone selectivity of different extractants in model
solution and fermentation broth...................................................................316
Figure C.1 Truncated sol operon (2.9 kb) with ald, ctfA and ctfB genes.........................322
Figure C.2 Constructed pMTL-thl-adhE2 plasmid..........................................................323
Figure C.3 Constructed pSV6 plasmid............................................................................324

1
Chapter 1: Introduction
n-Butanol is a four-carbon primary alcohol, and is currently mainly used as a solvent,
chemical intermediate, and extractant in cosmetics and pharmaceutical industries as well
as production of butyl acrylate and methacrylate (Dürre, 1998; 2007; Garćia et al., 2011;
Lee et al., 2008). In recent years, butanol has been attracting research attention as an
alternative biofuel to bioethanol. Compared to ethanol, butanol is considered as the next
generation biofuel due to many advantages it offers, such as higher energy content and
lower volatility (Dürre, 2007; Lee et al., 2008; Nigam and Singh, 2011). Butanol can be
used directly or blended with gasoline and diesel as fuel additives in the current
automobile engine without any modification or substitution. In addition, butanol is
compatible with the current transportation pipeline for gasoline (Dürre, 2007; Lee et al.,
2008).
Acetone-butanol-ethanol (ABE) fermentation was an important industrial process
during the early 1900s, and was first reported for butanol production by Louis Pasteur in
1861 (Gabriel, 1928; Gabriel and Crawford, 1930). However, butanol production by ABE
fermentation declined rapidly during the 1950’s due to the rise of cheaper petrochemical
synthesis and increased cost of fermentation raw materials (Dürre, 2007; Kumar and

2
Gayen, 2011). Even today, butanol is predominately produced through petrochemical
synthesis via Oxo process, which relies on crude oil supply. With the growing concerns of
environmental issues, depleting fossil resources and increasing crude oil price, renewed
interest has returned to fermentative butanol production, not only as a chemical but also
as an alternative biofuel (Ezeji et al., 2004a; 2007; Kumar and Gayen, 2011; Lee et al.,
2008). To overcome the limitations of conventional ABE fermentation such as low titer
and high substrate cost, areas under research and development include utilization of
renewable and low-cost feedstocks, development in novel fermentation processes,
alternative product recovery technologies, and metabolic engineering of
solvent-producing microorganisms (Chernova et al., 2010; Ezeji et al., 2010; Huang et al.,
2010; Qureshi and Ezeji, 2008; Vane, 2008).
Solventogenic Clostridia species, which are commonly used in ABE fermentation,
produce acetone, butanol, ethanol, acetic acid, butyric acid, hydrogen, and carbon dioxide
as the main products. A very distinct feature for solventogenic Clostridia is the biphasic
fermentation (Fond et al., 1985; Girbal and Soucaille, 1998), with a metabolic shift from
acidogensis to solventogensis. The typical acetone/butanol/ethanol ratio is 3:6:1 with
10-13 g/L butanol and 15-18 g/L ABE production in a conventional ABE fermentation
(Dürre, 1998; Ezeji et al., 2004a; Qureshi and Ezeji, 2008). The low butanol titer is the
biggest limitation affecting the competiveness of ABE fermentation, which is due to end
product butanol inhibition on cells at a concentration as low as 5-10 g/L (Qureshi and

3
Ezeji, 2008). Hyper-butanol-producing mutant strains obtained using chemical mutagen
and metabolic engineering have been reported with enhanced butanol production and
tolerance compared to parental strains, such as C. beijerinckii BA101 and recombinant C.
acetobutylicum ATCC 824 (Mermelstein et al., 1993; Qureshi and Blaschek, 2001). In
addition, asporogenous strain C. beijerinckii ATCC 55025 has also been suggested as a
stable butanol producing strain as opposed to other sporulating strains (Jain et al., 1993).
Substrate cost constitutes at least 50% of the total production cost in ABE
fermentation, and the process economics and feasibility largely depends on the
availability of cost-effective raw materials (Dürre, 2007; Garćia et al., 2011; Qureshi and
Ezeji, 2008). Lignocellulosic biomass has been recently suggested as renewable and
low-cost raw material for ABE fermentation, substituting the increasingly costly
traditional substrates such as cane molasses and corn (Qureshi and Ezeji, 2008). Many
pretreatment and hydrolysis methods, such as dilute acid and alkaline pretreatment, have
been extensively studied in order to utilize the sugars stored in lignocellulose (Kumar et
al., 2009). Detoxifications, such as overliming and activated charcoal, have also been
proposed to remove the inhibitory compounds present in the resulting hydrolysate,
improving the efficiency of the sequential fermentation process (Martinez et al., 2001;
Mussatto and Roberto, 2004). Lignocellulosic corn fiber, wheat straw and switchgrass,
have been investigated and identified as alternative substrates for butanol production via
ABE fermentation by solventogenic Clostridia (Qureshi et al., 2007; 2008; 2010).

4
In addition to conventional batch fermentation, fed-batch and continuous
fermentation techniques have been developed to utilize concentrated substrates and
eliminate downtime, reducing the reactor size and capital cost with enhanced reactor
productivity (Ezeji et al., 2004b; 2005). Cell immobilization with bonechar, brick, and
cotton towels as supporting materials has also been applied in ABE fermentation to
achieve high cell density and reactor productivity (Huang et al., 2004; Qureshi and
Maddox, 1988; Qureshi et al., 2000). In addition, cell recycle by membrane filtration has
been suggested to retain a high cell density in the reactor as an alternative to cell
immobilization (Afschar et al., 1985; Yang and Tsao, 1995). Fibrous bed bioreactor (FBB)
was patented as a superior cell-immobilization system with cotton towels as supporting
material, achieving constant cell-renewal and high viable cell density (Yang, 1996). FBB
has been applied in continuous butanol fermentation and demonstrated enhanced reactor
productivity (Huang et al., 2004).
Many butanol recovery techniques, including gas stripping, pervaporation,
liquid-liquid extraction and adsorption, have been proposed as alternatives to the
conventional distillation process, which is cost-intensive due to the low butanol titer in
the broth (Ezeji et al., 2004b; 2005; Qureshi and Blaschek, 1999; Roffler et al., 1987;
Yang and Tsao, 1995). These alternative recovery techniques provide feasible solutions to
not only energy-saving separation, but also online butanol recovery, neither of which can
be realized using distillation. Among the alternative recovery techniques, gas stripping

5
was suggested as the most effective one for online butanol recovery in ABE fermentation
(Zheng et al., 2009). Gas stripping does not require membrane assistance like
pervaporation, and doesn’t foul or exhibit decreased efficiency over time like
pervaporation, liquid-liquid extraction and adsorption techniques (Vane, 2008). When
integrating gas stripping with ABE fermentation for online product recovery, 500 g/L
glucose were utilized with 232.8 g/L ABE production in fed-batch fermentation,
compared to 45.4 g/L glucose consumed and 17.6 g/L ABE obtained in the control batch
fermentation (Ezeji et al., 2004b).
1.1 Project goal and specific tasks
The overall project goal is to develop a process to produce butanol via ABE
fermentation from various lignocellulosic feedstocks using hyper-butanol-producing
mutant strains, and to integrate online product recovery with the fermentation process for
enhanced butanol production, reactor productivity and sugar conversion. Figure 1.1
provides an overview of this study. The specific objectives and major tasks are described
below.
Task 1: Enhanced butanol production using hyper-butanol-producing mutants
Two mutant strains were employed in this project in order to obtain superior butanol
production. Mutant strain JB200 of asporogenous C. beijerinckii ATCC 55025 was

6
obtained and isolated using adaptation and evolutionary engineering, whereas mutant C.
beijerinckii CC101-SV6 was obtained by overexpressing the solvent-producing genes on
the sol operon of C. acetobutylicum. These mutants exhibited stable and high butanol
production from glucose and xylose, as well as lignocellulosic substrates, including corn
fiber, cassava bagasse and sugarcane bagasse. The results using these mutants are
presented in Chapters 3, 5 and 7.
Task 2: Butanol production from lignocellulosic biomass
Corn fiber, cassava bagasse, wood pulp and sugarcane bagasse were investigated
and utilized as lignocellulosic substrates for ABE fermentation (Chapters 3, 5, 6, and 7).
Fibrous bed bioreactor was employed as an immobilized-cell system for batch and
fed-batch operations using corn fiber and cassava bagasse as substrates for enhanced
butanol production (Chapters 3 and 5). Several detoxification procedures, including
overliming and adsorption with activated carbon and ion exchange resins, were
performed on these hydrolysates, and the effects of these procedures on butanol
production were compared and evaluated using wood pulp hydrolysate (Chapter 6).
Task 3: Evaluation of gas stripping as product recovery technique
Gas stripping was employed as an online product recovery technique with ABE
fermentation. Several operating parameters, including gas flowrate, condensation
temperature, and presence of cells were studied and the results are presented in Chapter 4.
The effects of these factors on butanol stripping rate and butanol selectivity were

7
evaluated in order to optimize the gas stripping process and gain knowledge that is
necessary for the later integrated process.
Task 4: Integration of ABE fermentation with online product recovery
Integrated ABE fermentation process with gas stripping as online product recovery
using cassava bagasse hydrolysate and wood pulp hydrolysate was studied and the results
are discussed in Chapter 5 and Chapter 6, respectively. Free-cell batch fermentation was
employed in the study described in Chapter 6, whereas immobilized-cell fed-batch
fermentation was employed in Chapter 5. Enhanced butanol production was obtained in
both studies, compared with control non-integrated process.
1.2 Significance and major impacts
n-Butanol is an important industrial chemical and solvent currently sold at $2002.62
~ 2018.02/t or $6.14 – 6.19 per gallon (http://price.alibaba.com, 1-butanol, retrieved on
5/2/2011). The annual worldwide market for butanol as a chemical is estimated at 350
million gallons with 220 million gallons in the US domestic market, projecting a $2.14
billion butanol market worldwide. Currently, gasoline is sold at $4.15 per gallon with
ascending trend in the US and at much higher prices in European and Asian countries due
to instability within the Middle Eastern countries which directly influences crude oil
supply. Current research on biofuels is supported by the US government, which calls for
36 billion gallons of annual production of biofuels by 2022 based on the Energy

8
Independence and Security Act (EISA 2007) passed in 2007. Second-generation biofuels
based on lignocellulosic feedstocks are especially promising (Festel, 2008). Therefore, it
is of great interest to research on biobutanol production as a potential biofuel to replace
gasoline using domestically produced lignocellulosic feedstocks. It was estimated that
based on yield of 3.89 ton/acre and 78.1 million acres of corn production nationwide,
8.27 billion gallons of butanol can be obtained from the bioconversion of corn stover,
replacing 7.55 billion gallons of gasoline every year (Swana et al., 2011). Currently,
biobutanol can be blended with US gasoline up to 11.5% (v/v), and it is very promising to
replace a large portion of gasoline currently used in the US in the near future (Nigam and
Singh, 2011).
In summary, biobutanol production from lignocellulosic biomass is crucial in
developing energy independence and sustainable fuel security of the country, preserving
and prolonging the life of fossil reserves and minimizing environmental impacts. This
project studied the biobutanol production from various lignocellulosic feedstocks,
including corn fiber, cassava bagasse, wood pulp and sugarcane bagasse, and
demonstrated the feasibility and advantages of the integrated process for enhanced
process efficiency. Further scaling up for commercial applications is currently undertaken
by industrial collaborators.
1.3 References

9
Afschar, A.S., H. Biebl, K. Schaller, and K. Schugerl (1985). Production of acetone and butanol by Clostridium acetobutylicum in continuous culture with cell recycle. Appl. Microbiol. Biotechnol., 22, 394-398.
Chernova, N.I., T.P. Korobkova, and S.V. Kiseleva (2010). Use of biomass for producing liquid fuel: Current state and innovations. Thermal Eng., 57, 937-945.
Dürre, P. (1998). New insights and novel developments in clostridial acetone/ butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Dürre, P. (2007). Biobutanol: An attractive biofuel. Biotechnol. J., 2, 1525-1534.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004a). Butanol fermentation research: upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004b). Acetone-butanol-ethanol production from concentrated substrate: reduction in substrate inhibition by fed-batch technique and product inhibition by gas stripping. Appl. Microbiol. Biotechnol., 63, 653-658.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2005). Process for continuous solvent production. United States Patent Application Publication, US patent 20050089979A1.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007). Bioproduction of butanol from biomass: from genes to bioreactors. Current Opinion in Biotechnol., 18, 220-227.
Ezeji, T.C., C. Milne, N.D. Price, H.P. Blaschek (2010). Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol., 85, 1697-1712.
Festel, G.W. (2008). Review : Biofuels – Economic aspects. Chem. Eng. Technol., 31, 715-720.
Fond, O., G. Matta Ammouri, H. Petitdemange, and J.M. Engasser (1985). The role of acids on the production of acetone and butanol by Clostridium acetobutylicum. Appl. Microbiol. Biotechnol., 22, 195-200.
Gabriel, C.L. (1928). Butanol fermentation process. Ind. Eng. Chem., 20, 1063-1067.
Gabriel, C.L. and F.M. Crawford (1930). Development of the butyl-acetonic fermentation

10
industry. Ind. Eng. Chem., 22, 1163-1165.
Garćia, V., J. Päkkilä, H. Ojamo, E. Muurinen, R.L. Keiski (2011). Challenges in biobutanol production: How to improve the efficiency. Renew. Sustain. Ener. Reviews, 15, 964-980.
Girbal, L. and P. Soucaille (1998). Regulation of solvent production in Clostridium acetobutylicum. Trends Biotechnol., 16, 11-16.
Huang, W.C., D.E. Ramey, and S.T. Yang (2004). Continuous production of butanol by Clostridium acetobutylicum immobilized in a fibrous bed bioreactor. Appl. Biochem. Biotechnol., 113-116, 887-898.
Huang, H., H. Liu, Y.R. Gan (2010). Genetic modification of critical enzymes and involved genes in butanol biosynthesis from biomass. Biotech. Adv., 28, 651-657.
Jain, M.K., D. Beacom, and R. Datta (1993). Mutant strain of C. acetobutylicum and process for making butanol. United States Patent, US Patent 5192673.
Kumar, P., D.M. Barrett, M.J. Delwiche and P. Stroeve (2009). Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem., 48, 3713-3729.
Kumar, M. and K. Gayen (2011). Developments in biobutanol production: New insights. Appl. Ener., 88, 1999-2012.
Lee, S.T., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentive butanol production by Clostridia. Biotechnol. Bioeng., 101,209-228.
Martinez, A., M.E. Rodriguez, M.L. Wells, S.W. York, J.F. Preston and L.O. Ingram (2001). Detoxification of dilute acid hydrolysates of lignocellulose with lime. Biotechnol. Prog., 17, 287-293.
Mussatto, S.I. and I.C. Roberto (2004). Alternatives for detoxification of diluted-acid lignocellulosic hydrolysates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Nigam, P.S. and A. Singh (2011). Production of liquid biofuels from renewable resources. Prog. Ener. Combust. Sci., 37, 52-68.

11
Qureshi, N. and I.S. Maddox (1988). Reactor Design for the ABE fermentation using cells of Clostridium acetobutylicum immobilized by adsorption onto bonechar. Bioprocess Eng., 3, 69-72.
Qureshi, N. and H.P. Blaschek (1999). Production of acetone butanol ethanol (ABE) by a hyper-producing mutant strain of Clostridium beijerinckii BA101 and recovery by pervaporation. Biotechnol. Prog., 15, 594-602.
Qureshi, N. and H.P. Blaschek (2001). Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 27, 287-291.
Qureshi, N., J. Schripsema, J. Lienhardt and H.P. Blaschek (2000). Continuous solvent production by Clostridium beijerinckii BA 101 immobilized by adsorption onto brick. J. Microbiol. Biotechnol., 16, 377-382.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst. Eng., 30, 419-427.
Qureshi, N. and T.C. Ezeji (2008). Butanol, ‘a superior biofuel’ production from agricultural residues (renewable biomass): recent progress in technology. Biofuels, Bioprod. Bioref., 2, 319-330.
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta and H.P. Blaschek (2008). Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.
Qureshi, N., B.C. Saha, R.E. Hector, B. Dien, S. Hughes, S. Liu, L. Iten, M.J. Bowman, G. Sarath, M.A. Cotta (2010). Production of butanol (a biofuel) from agricultural residues: Part II-Use of corn stover and switchgrass hydrolysates. Biomass Bioenergy, 34, 566-571.
Roffler, S.R., H.W. Blanch, and C.R. Wilke (1987). In-situ recovery of butanol during fermentation, part 1: batch extractive fermentation. Bioprocess Eng., 2, 1-12.
Swana, J., Y. Yang, M. Behnam, R. Thompson (2011). An analysis of net energy production and feedstock availability for biobutanol and bioethanol. Bioresour. Technol., 102, 2112-2117.
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols

12
from fermentation broths. Biofuls, Bioprod. Bioref., 2, 553-588.
Yang, S.T. (1996). Extractive fermentation using convoluted fibrous bed bioreactor. United States Patent, US patent 5563069.
Yang, X. and G.T. Tsao (1995). Enhanced acetone-butanol fermentation using repeated fed-batch operation coupled with cell recycle by membrane and simultaneous removal of inhibitory products by adsorption. Biotechnol. Bioeng., 47, 444-450.
Zheng, Y.N., L.Z. Li, M. Xian, Y.J. Ma, J.M. Yang, X. Xu, D.Z. He (2009). Problems with the microbial production of butanol. J. Ind. Microbiol. Biotechnol., 36, 1127-1138.

Task 1 Butanol production
using hyper-butanol-
producing mutants
C. beijerinckii ATCC
55025 mutant JB200
(Chapters 3, 5)
C. beijerinckii mutant
CC101-SV6
(Chapter 7)
Task 2 Butanol production
from lignocellulosic
biomass
Corn fiber (Chapter 3)
Cassava bagasse
(Chapter 5)
Wood pulp (Chapter 6)
Sugarcane bagasse
(Chapter 7)
Task 3 Evaluation of gas
stripping as product
recovery technique
(Chapter 4)
Task 4 Integration of ABE
fermentation with
online product recovery
(Chapters 5, 6)
Project Goal To develop a process for butanol production
from lignocellulosic biomass in ABE fermentation
integrated with online product recovery
Figure 1.1 Overview of project goal and major tasks carried out in this study
13

14
Chapter 2: Literature Review
2.1 Acetone-Butanol-Ethanol (ABE) fermentation
Biological butanol production via fermentation was first reported in 1861 by Louis
Pasteur, who first discovered and isolated a butyric acid producing strain and later on
observed butanol production along with butyric acid (Dürre, 1998; Garćia et al., 2011;
Jones and Woods, 1986). In the following years, many scientists including Albert Fitz
and Martinus Beijerinck continued the work of butanol-producing microorganisms and
isolated several additional strains such as Bacillus butylicus, Granilobacter butylicus and
Granulobacter saccharobutyricum (Dürre and Bahl, 1996; Dürre, 1998; Garćia et al.,
2011). In 1926, McCoy et al. first used the name of Clostridium acetobutylicum in their
paper and this name was officially recognized and accepted as the butanol producing
microorganism (McCoy et al., 1926). Weizmann, along with a British company Strange
& Graham Ltd., later on isolated a strain that showed good acetone and butanol
producing ability, and developed and patented a process based on this strain to produce
butanol (Dürre, 1998; Gabriel, 1928; Gabriel and Crawford, 1930; Jones and Woods,
1986; Kumar and Gayen, 2011). This process played an important role in World War I,

15
and since 1920 acetone and butanol have became major fermentation products for their
excellent properties as solvents (Beesch, 1952; Dürre, 2007; Ennis et al., 1986a; Garćia et
al., 2011). Many countries, including USA, England, China, Australia, and Canada, built
biological butanol plants employing ABE fermentation between 1920 and 1980, but they
all eventually came to a standstill due to the rise of cheaper petrochemical synthesis of
butanol from crude oils and the high cost of fermentation raw materials (Ezeji et al.,
2004a; 2007a; Garćia et al., 2011; Kumar and Gayen, 2011).
However, a revisit on ABE fermentation over the past few decades has made
significant advances and breakthroughs in the bioproduction of butanol from various
alternative feedstocks (Ezeji et al., 2004a; 2010; Demain, 2009; Dürre, 1998; 2007; Lee
et al., 2008; Ni and Sun, 2009; Nigam and Singh, 2011; Qureshi and Ezeji, 2008; Swana
et al., 2011; Weber et al., 2010; Zheng et al., 2009). Recently, with the depleting fossil
fuel reserves and surging crude oil price, biological production of butanol as a superior
biofuel candidate has become a hot research topic. Compared to ethanol, butanol is a
superior fuel candidate, and the characteristics of butanol are very similar to gasoline. A
comparison of some properties among butanol, ethanol, methanol and gasoline is
summarized in Table 2.1. Butanol has a higher energy content and lower volatility than
ethanol and methanol. Most importantly, butanol can be directly used as an alternative to
gasoline or fuel additive in the current internal combustion engine without any

16
modification. Therefore, butanol could become the next generation liquid biofuel in the
near future (Nigam and Singh, 2011).
2.1.1 Microorganisms and strain improvements
Butanol (and acetone, ethanol and isopropanol) is naturally produced by genus
Clostridia bacteria (Jones and Woods, 1986; Kumar and Gayen, 2011; Lee et al., 2008).
Clostridia are rod-shaped, spore-forming, gram-positive and obligate anaerobic bacteria.
Due to some special genes and various enzymes produced in Clostridia, they are mostly
used as the solvent-producing bacteria in ABE fermentation (Cornillot et al., 1997; Dürre,
1998; Ezeji et al., 2007a). Butanol-producing Clostridia include a variety of species,
including acetobutylicum, beijerinckii, saccaroperbutylacetonicum,
saccharoacetobutylicum, aurantibutyricum, pasteurianum, sporogenes, and
tetanomorphum (Kumar and Gayen, 2011). Among these species, C. acetobutylicum, C.
beijerinckii, C. saccharoacetobutylicum, and C. saccaroperbutylacetonicum are the
primary producers with good butanol production and yields (Lee et al., 2008). The
substrate utilization ability among naturally solventogenic Clostridia is very different
from each other, as well as their optimal pH, temperature, and product profiles. Most of
the species can ferment pentose and hexose sugars, as well as starch, while some strains
also possess the ability to utilize syngas and glycerol as the carbon source. Table 2.2
compares and summarizes substrates utilized by various solventogenic Clostridia species,

17
along with their main fermentation products. Most of the species produce butanol as the
main product, although some also produce significant amounts of 1,3-propanediol and
isopropanol.
C. acetobutylicum was the main species employed in industrial ABE fermentation
until more detailed taxonomy was developed and some strains of C. acetobutylicum were
re-classified as C. beijerinckii based on the product type (Dürre, 1998). Many different
strains of these two species have been extensively studied, including C. acetobutylicum
ATCC 824, P262, P260 and DSM 1731, and C. beijerinckii ATCC 55025, NCIMB 8052,
and BA101 (Bahl et al., 1982; Huang et al., 2004; Maddox et al., 1995; Parekh et al.,
1998; Qureshi et al., 2006; Soni et al., 1987). These strains all showed good butanol
production between 10 g/L to 20 g/L. Some of these popular strains have been compared
in a study by Gutierrez et al. (1998) using potato as the substrate, and strong
solventogenic abilities were reported. Among all the afore-mentioned strains, ATCC
55025 is the only asporogenous strain, while the rest of them all produce endospores
under severe environmental stresses. It is generally accepted that sporulation happens
when solvents are produced and endospores function as a defense against the harsh
environment, which however also results in unstable solvent production in ABE
fermentation. The asporogenous feature ensured solvent-producing stability of ATCC
55025, which also had higher butyrate uptake and butanol tolerance (Jain et al., 1993).

18
A very distinctive feature of Clostridia is the biphasic fermentation (Ezeji et al., 2010;
Kumar and Gayen, 2011; Lee et al., 2008; Jones and Woods, 1986). During the first
phase, which is known as acidogensis, acids (acetate and butyrate) and carbon dioxide are
produced as the main products during the exponential growth phase, lowering the pH of
the medium. Then, through a series of regulations, signals and change in gene expression,
the second phase, which is known as the solventogensis, is triggered and acids are
reassimilated and converted to solvents (acetone, butanol and ethanol) (Gottschalk and
Morris, 1981). A detailed metabolic pathway with genes and enzymes for reactions
during acidogenesis and solventogenesis is shown in Figure 2.1.
Butanol, a severe fermentation inhibitor to Clostridia, changes the phospholipid and
fatty acid composition in the cell membrane, alters the membrane structure and
compromises the fluidity of the membrane. It also adversely affects the solute transport,
membrane permeability, and maintenance of intracellular pH and ATP level (Ezeji et al.,
2010; Kumar and Gayen, 2011). Fermentation is severely inhibited when butanol
concentration reaches above 1% and stopped at 2% for most of microorganisms
(Knoshaug and Zhang, 2009). Many strain improvement strategies including mutation
and genetic engineering have been proposed and conducted to enhance the microbial
butanol tolerance, butanol production and yields (Ezeji et al., 2010; Dürre, 2007; Harris
et al., 2002; Jones and Woods, 1986; Lee et al., 2008; Nair et al., 1999; Thormann and
Dürre, 2001; Thormann et al., 2002). The mutant SA-1 of C. acetobutylicum ATCC 824

19
and mutant C. beijerinckii BA101 of C. beijerinckii NCIMB 8052 were two
representative mutants successfully obtained by using chemical mutagens (Formanek et
al., 1997; Lin and Blaschek, 1983; Qureshi and Blaschek, 2001b). SA-1 was obtained
through a serial culture transfer into medium containing increasing amounts of butanol
(mutagen), whereas BA101 was obtained using N-methyl-N9-nitro-N-nitrosoguanidine
(mutagen) along with selective enrichment on glucose analog 2-deoxyglucose. C.
beijerinckii BA101 was reported to be capable of producing up to 2% butanol with very
efficient acids conversion to solvents (Formanek et al., 1997), while the butanol tolerance
of C. acetobutylicum SA-1 was reported to be 121% higher than that of its parental strain
(Lin and Blaschek, 1983).
As for metabolic engineering of Clostridia, only five genes (buk, pta, adhE, solR,
and spo0A) have been knocked out in C. acetobutylicum due to lack of efficient
knock-out methods to date, which hindered the genetic engineering of Clostridia (Zheng
et al., 2009). Spo0A has been identified as a positive regulator that enhanced solvent
production. Harris et al. (2002) reported that inactivation of Spo0A resulted in 1.0 g/L
butanol, whereas overexpression of Spo0A resulted in a 10.2 g/L butanol in C.
acetobutylicum ATCC 824. Besides Spo0A, ctfA, ctfB, adc and aad have also been
reported as solvent producing genes, and overexpression of these genes in C.
acetobutylicum ATCC 824 resulted in a 37% and 90% increase in butanol (13.2 g/L) and
acetone (8.6 g/L) production, respectively. More recently, a mutant of C. tyrobutyricum

20
overexpressing adhE2 gene was found to produce 10 g/L butanol from glucose and 16
g/L from manitol (Yu et al., 2011).
Besides Clostridia, E. coli, P. putida and B. subtilis have also been engineered as
hosts to produce butanol by introducing the butanol-producing genes from C.
acetobutylicum (Atsumi et al., 2007; 2008; Inui et al., 2008; Nielsen et al., 2009; Shen
and Liao, 2008). More detailed information on gene up-regulators, down-regulators, hosts
and pathways can be found in several recent review articles (Dürre, 2008; Ezeji et al.,
2010; Lee et al., 2008; Kumar and Gayen, 2011; Zheng et al., 2009).
2.1.2 Traditional substrates and renewable lignocellulosic feedstocks
Substrate cost is a very important factor impacting on the economics of butanol
production via fermentation. Traditionally, corn, molasses and glucose were the major
substrates utilized in commercial ABE fermentation in the early 20th century (Dürre, 1998;
Ezeji et al., 2004a; 2007a; Jones and Woods, 1986; Qureshi and Ezeji, 2008). In a typical
batch fermentation, 20-25 g/L ABE can be obtained within 36-72 h followed by
distillation as butanol recovery. Butanol production using traditional substrates such as
glucose and corn starch is listed in Table 2.3.
With the increasing demand of food supply worldwide, utilization of food-based
substrates has become cost-intensive and controversial (Garćia et al., 2011; Kumar and
Gayen, 2011; Nigam and Singh, 2011). With the concern of sustainability and

21
cost-effectiveness in mind, research motives have been driven in the direction to search
for inexpensive and non-food based substrates for butanol production via ABE
fermentation. Fortunately, Clostridia can utilize a variety of carbohydrates, including
glucose, xylose, arabinose, fructose, mannose, sucrose, lactose, cellobiose, starch,
glycerol and dextrin, but not trehalose, rhamnose and melibiose (Ezeji et al., 2004a;
2007a; Jones and Woods, 1986; Kumar and Gayen, 2011; Qureshi and Ezeji, 2008). This
feature of Clostridia effectively broadens the substrate pool, and makes it possible to
utilize lignocellulosic biomass feedstocks. It has been estimated that the net energy
generated from corn-to-butanol is 6.53 MJ/L, which could be significantly improved to
15.90 MJ/L if lignocellulosic biomass is used instead of corn (Swana et al., 2011).
Therefore, it is of great interest to research on biobutanol production through ABE
fermentation using domestically produced lignocellulosic feedstocks as potential
substrates. Based on current crop harvest yield and 0.42 g/g butanol yield from ABE
fermentation using life cycle analysis, 8.27 billion gallons of butanol can be obtained
from bioconversion of renewable and sustainable lignocellulosic biomass, such as corn
stover and switchgrass, replacing 7.55 billion gallons of gasoline every year (Swana et al.,
2011).
Lignocellulosic biomass consists of a variety of agro-industrial residues (e.g. corn
fiber, corn stover, wheat straw, barley straw and sugarcane bagasse), energy crops (e.g.
switchgrass), forestry products (wood chips), and municipal solid wastes (Howard et al.,

22
2003; Kumar et al., 2009; Reddy and Yang, 2005; Saha, 2003). Every year, around
2×1011 tons of lignocellulosic biomass are produced (Reddy and Yang, 2005),
representing the most abundant renewable sugar source. Lignocellulose consists of
mainly cellulose (35-50%), hemicellulose (25-35%) and lignin (10-25%), and a small
amount of protein, ash and some extractives (Kumar et al., 2009; Jorgensen et al., 2007).
The composition and current use of some common lignocellulosic feedstocks are
summarized in Table 2.4.
Lignocellulose is the largest reservoir of solar energy stored in the form of carbon
source on earth, representing a potential group of feedstocks suitable for many
bioconversion processes. As shown in Table 2.4, most of the lignocellulosic biomass is
considered as waste materials from industrial processing and sold at low prices for animal
feed or burnt as a source of energy. It is especially appealing that lignocellulosic
feedstocks are renewable and available in abundance. Many processes have been studied
and reported for the bioconversion of lignocellulosic biomass into various value-added
products, such as enzymes, biofuels, and chemicals (Duff and Murray, 1996; Kim et al.,
1999; Malherbe and Cloete, 2003; Olsson and Hahn-Hagerdal, 1996; Rabinovich et al.,
2002; Roberto et al., 1995; Sun and Cheng, 2002). Several lignocellulosic materials such
as corn fiber, dried distiller grains and solubles, wheat straw, and switchgrass have been
reported and successfully applied in ABE fermentation as substrates to produce butanol
(Table 2.3). In general, cellulose and hemicellulose present in the lignocellulosic

23
feedstocks are not directly accessible to the microorganisms because solventogenic
Clostridia do not posses enzymes that can breakdown these materials. In order to utilize
the lignocellulosic biomass, the sugars stored in the form of hemicellulose and cellulose
must first be released. Therefore, lignocellulose has to be pretreated and hydrolyzed to
release all the sugars that can be utilized by the microorganisms in the subsequent
fermentation process. Due to the lignin protection and crystalline cellulose microfibrils,
lignocellulosic materials are usually very resistant to enzymatic hydrolysis (Howard et al.,
2003 Jorgensen et al., 2007; Yat et al., 2008). In addition, under the extreme conditions
employed in pretreatment processes, many toxic compounds that are severe fermentation
inhibitors are inevitably generated (Hendriks and Zeeman, 2009; Moiser et al., 2005;
Mussatto and Roberto, 2004a). Detoxification of lignocellulosic hydrolysate is preferred
in order to obtain better butanol production in the subsequent fermentation process.
Details on pretreatment, hydrolysis, and detoxification will be elaborated in Section 2.2.
2.1.3 Developments in fermentation process
Conventional ABE fermentation is usually operated with free cells in the batch mode.
Due to end product toxicity, free-cell batch fermentation suffers from low cell density
and low reactor productivity (Dürre, 1998; Ezeji et al., 2004a; 2007a; 2010; Maddox,
1989; Qureshi and Ezeji, 2008). As a result, the butanol yield in traditional ABE
fermentation is low, typically around 20% and rarely exceeds 25%, with a cell density of

24
~3-4 g/L. Due to the low cell density and severe product inhibition, the reactor
productivity is usually around 0.25-0.4 g/L·h, rarely over 0.5-0.6 g/L. h.
In order to achieve high cell density in the bioreactor, cell immobilization and cell
recycle have been applied (Ezeji et al., 2007a; 2010; Maddox, 1989). In cell
immobilization, cells are fixed on a support through adsorption or entrapment, whereas in
cell recycle, cells are retained in the reactor usually by using a membrane. Cell
immobilization by adsorption allows cell renewal, which can maintain a highly viable
cell density in the reactor. It is also ideal for cell mutation and evolution over an extended
period of time under harsh environment (Huang et al., 2002; 2004; Silva and Yang, 1995;
Yang, 1996). With the assistance of a membrane, cells in the reactor can be retained
and recycled, preventing any loss that may happen in the immobilized cell reactors. Both
cell immobilization and cell recycle can significantly increase the reactor productivity
due to the increased cell density per reactor volume and the elimination of reactor
downtime (Qureshi and Ezeji, 2010). Novel reactor designs based on cell immobilization
and cell recycle have been studied, and many materials including sponge, brick and corn
stalk have been suggested as potential support materials for cell immobilization. Table
2.5 summarizes immobilized cell fermentations using different materials and membrane-
assisted cell recycle fermentations for enhanced reactor productivity. Reactor
productivity as high as 15.8 g/L. h was achieved using brick as support material in a

25
continuous ABE fermentation, and average productivity using cell immobilization and
cell recycle was between 4.0-6.0 g/L. h.
Among all the materials listed in Table 2.5 for cell immobilization, cotton towel is
the most commonly available and inexpensive material. Yang (1996) elaborated in his
patent on this spiral-wound cell immobilization system with stainless steel mash and
cotton towel, which offers large contact surface area and good mass transfer. The spaces
between fibrous matrices provide large void volume to allow the fermentation gases and
particles to easily pass through, avoiding pressure build-up and reducing the clogging
problems. Constant cell renewal is realized by reversible adsorption, maintaining high
cell density with viable cells. Enhanced reactor productivity and final product
concentration were reported in several processes employing this fibrous system (Wu and
Yang, 2003; Zhu and Yang, 2003).
In addition to cell immobilization and cell recycle, fed-batch and continuous
fermentation technologies have been applied to overcome some drawbacks such as low
butanol titer and productivity associated with batch fermentation process. Fed-batch
fermentation is a technology of adding highly concentrated substrates into reactor at
intervals to maintain a desirable substrate concentration to avoid substrate inhibition
(Ezeji et al., 2004a; 2010). Fed-batch fermentation usually starts with a substrate level
equivalent of a batch process. As the substrate is being utilized by the cells, a small
volume of highly concentrated substrate is added to replace the consumed substrate,

26
resulting in higher final product concentration and reactor productivity. Due to the
accumulation of end product, which can cause inhibition on the cells, fed-batch
fermentation is feasible only when coupled with online product recovery. Fed-batch
technology can significantly improve the reactor productivity and reduce the reactor
volume, lowering the capital cost and thus improving the process economics (Dürre, 1998;
Ezeji et al., 2007a; Kumar and Gayen, 2011; Lee et al., 2008). It was reported (Ezeji et al.,
2004b) that 500 g/L glucose was utilized in fed-batch fermentation coupled with gas
stripping as product recovery, resulting in 232.8 g/L ABE with a productivity of 1.16 g/L.
h. In the control batch reactor, 45.4 g/L glucose was consumed, with a 17.6 g/L ABE
production. In continuous fermentation, fresh medium is continuously fed into reactor at
the same rate of product stream flowing out the reactor, keeping a constant volume in the
reactor (Ezeji et al., 2004a). Due to the dilution by fresh medium, end product inhibition
is prevented and dead cells and toxic metabolites are removed in continuous fermentation,
leading to a longer fermentation life. Continuous fermentation can be operated with free
cells, or operated with cell immobilization or cell recycle in order to achieve higher cell
density (see Table 2.5). Continuous fermentation can achieve high solvent productivity,
but at the expense of lower product concentration due to dilution. A productivity of 12.4
g/ L. h was reported in a continuous fermentation process with cell immobilization, with a
maximum ABE concentration of 8.8 g/L (Qureshi et al., 2004). Continuous fermentation

27
can eliminate the downtime and simplify the downstream process, lowering the process
cost and increasing efficiency (Ezeji et al., 2005b).
Besides fed-batch and continuous fermentation technologies for butanol production,
simultaneous saccharification and fermentation (SSF) has been recently proposed as
another feasible technology for ABE fermentation (Qureshi et al., 2008c; 2008d). Usually,
separate hydrolysis and fermentation (SHF) process was employed when using
lignocellulosic biomass as substrate for fermentation. The advantage of SHF is that the
hydrolysis process and fermentation process can be operated under their optimal
conditions (usually pH 5.0 and 50oC for enzymatic hydrolysis, and fermentation
temperature 30-37oC) (Hahn-Hagerdal et al., 2006). However, as the end product of the
hydrolysis, sugars inhibit the enzyme activity and lower the enzyme efficiency.
Simultaneous saccharification fermentation can solve this problem by integrating the two
processes together, with the enzymes, pretreated lignocellulose and microorganism all
present in the same reactor. A compromised condition, usually pH 5.0 and 37oC, is used
in SSF (Taherzadeh and Karimi, 2007). Enzyme converts the cellulose and hemicellulose
into sugars, and enzyme inhibition by sugars is relieved due to the simultaneous
utilization of the released sugars by the microorganism. SSF is commonly employed in
ethanol fermentation from lignocellulosic biomass, lowering the process energy
requirement and improving the enzyme efficiency and ethanol production
(Hahn-Hagerdal et al, 2006). Using wheat straw as the substrate, 13.12 g/L ABE were

28
produced from SHF by C. beijerinckii P260, whereas similar ABE production of 11.93
g/L was obtained from SSF, indicating that SSF is also a feasible option for ABE
fermentation using lignocellulosic biomass (Qureshi et al., 2008c).
2.2 Pretreatment and detoxification of lignocellulosic feedstocks
2.2.1 Pretreatment of lignocellulose
Lignocellulosic biomass mainly contains lignin, hemicellulose and cellulose. Lignin,
a highly cross-linked polymer complex comprising of phenolic alcohol monomers,
imparts structural support for plant cell wall. Lignin links and forms a rigid physical seal
around hemicellulose and cellulose to prevent solvent permeability and microbial attack
(Perez et al., 2002). Hemicellulose is composed of hetero-polysaccharide backbone
(mostly formed by xylose, arabinose, galactose and mannose) with short branches linked
also by β-(1-4)-glycosidic bonds. Hemicellulose acts like filler between lignin and
cellulose microfibrils (Saha, 2003; Reddy and Yang, 2005). Cellulose is the main
structural components in the plant cell wall, and is usually packed into tight microfibrils
due to the hydrogen bond linkage of cellulose long chain (Kumar et al., 2009). In plant
biomass, cellulose is usually in the crystalline form with a small portion in amorphous
form (Perez et al., 2002), which determines the hard-to-breakdown nature of cellulose by
both acid and enzyme hydrolysis. In order to efficiently convert cellulose to fermentable
sugars, lignin and hemicellulose must be removed. The goal of the pretreatment is to

29
remove lignin and hemicellulose, reduce the crystallinity of cellulose, and increase the
porosity of the lignocellulosic biomass. A comparison of different pretreatment methods
is presented in Table 2.6.
2.2.1.1 Physical/mechanical pretreatments
Physical pretreatment, also known as mechanical pretreatment, employs machinery
chipping, grinding, or milling to reduce the size of biomass and the cellulose crystallinity
improving easy acid/enzyme access. Depending on the requirements, biomass can first be
sent through a chipping machine to obtain particles at sizes of 10-30 mm; and if fine
powder is preferred, the biomass can be further sent for grinding or milling to reduce the
size to 0.2-2 mm (Sun and Cheng, 2002). In general, the smaller the particle size, the
easier for the microorganism or enzyme to digest. Smaller size also helps to disrupt the
crystalline structure of cellulose better. However, higher cost is usually associated with
finer particle size (Cadoche and Lopez, 1989).
2.2.1.2 Thermal pretreatment
Steam explosion employs high temperature steam (160-270oC) at high pressure
(0.69-4.83MPa) to treat the lignocellulosic biomass for a few seconds to minutes before
the biomass is suddenly exposed to atmospheric pressure, during which the biomass
undergoes an explosive decompression due to the sudden pressure drop (Duff and Murray,

30
1996; Kumar et al., 2009; Sun and Chang, 2002). It was reported that steam explosion
can greatly increase the enzymatic hydrolysis efficiency and reducing sugar yield from
many different lignocellulosic biomass such as corn stover, wheat straw, and wheat fiber
(Cara et al., 2007; Palmarola-Adrados et al., 2004 ;Varga et al., 2004; Zhang et al., 2008).
Steam explosion has been applied to and is recognized as one of the most effective
pretreatment methods for lignocellulosic materials, particularly agricultural residues and
hardwood (Kumar et al., 2009; Sun and Cheng, 2002). Advantages of steam explosion
mainly include reducing the biomass size, effective removal of lignin and hemicellulose
without dilution of the resulting sugars and lower energy cost compared to mechanical
milling (Cara et al., 2007; Kumar et al., 2009; Zhang et al., 2008).
Liquid hot water pretreatment is an alternative thermal treatment to steam
explosion. Water is kept at liquid state at very high temperature (200-230oC) to treat
lignocellulosic biomass for about 15 minutes (Hendriks and Zeeman, 2009; Kumar et al.,
2009; Mosier et al., 2005). Liquid hot water pretreatment is supposed to solubilize part of
the biomass instead of converting it to monomeric sugars, thus avoiding the formation of
fermentation inhibitors due to the degradation of the sugars (Hendriks and Zeeman, 2009).
It was reported that 40-60% of the biomass can be dissolved during the liquid hot water
pretreatment, with 4-22% cellulose, 35-60% lignin and all of the hemicellulose removed
(Kumar et al., 2009; Mosier et al., 2005). Liquid hot water pretreatment is usually applied
to biomass in three ways: co-current, counter-current and flow through. Compared to

31
steam explosion, liquid hot water pretreatment reduces the solubilized hemicellulose and
lignin concentration due to the large amount of water input, thus reducing the possibility
of hemicellulose and lignin’s further degradation into furfural, HMF and phenolic
compounds (Hendriks and Zeeman, 2009).
2.2.1.3 Ammonia fiber explosion (AFEX)
Ammonia fiber explosion is another pretreatment method that is very similar to
steam explosion. It also employs high temperature (lower compared to steam explosion,
80-100oC), high pressure, and the sudden pressure release in the end as steam explosion.
The major difference is that lignocellulosic biomass is exposed to liquid ammonia under
the treatment instead of steam (Mes-Hartree et al., 1988; Teymouri et al., 2004). The
goal of AFEX pretreatment is to use ammonia vapor to penetrate through the tight fibrous
structure of biomass, destroy the lignin protection and disrupt the cellulose crystallinity,
making the following enzymatic hydrolysis of hemicellulose and cellulose efficient
(Kumar et al., 2009). It was also reported that during AFEX pretreatment, no
fermentation inhibitors were produced, thus promoting the following bioconversion
process (Mes-Hartree et al., 1988). However, due to the heavy loading of liquid ammonia,
high ammonia price, and difficulties in recycling the ammonia, AFEX pretreatment
process is very costly and not suitable for commercialization (Mosier et al., 2005).

32
2.2.1.4 Chemical pretreatments
Acid pretreatment can be divided into dilute acid and concentrated acid
pretreatment (Jorgensen et al., 2007; Malherbe and Cloete, 2003; Sun and Cheng, 2002).
The goal of acid pretreatment is to partially or completely hydrolyze hemicellulose, break
down the lignin structure and disrupt the cellulose crystallinity for further enzymatic
digestion to release fermentable sugars (Perez et al., 2002; Sun and Cheng, 2002).
Generally, concentrated acid (H2SO4 and HCl) pretreatment is considered to be too
corrosive and dangerous to operate. In addition, a large amount of base is required for
neutralization, resulting in high salt concentration in the hydrolysate highly inhibitory to
the fermentation (Jorgensen et al., 2007; Kumar et al., 2009; Malherbe and Cloete, 2003;
Perez et al., 2002). Therefore, dilute acid pretreatment is much more commonly used
compared to the concentrated acid pretreatment.
Dilute H2SO4 and HCl are commonly used in dilute acid pretreatment of biomass
with concentration ranging from 0.5% to 5%(v/v), or 0.05 to 1 N depending on the
biomass type or process time (Mussatto and Roberto, 2004a; Qureshi et al., 2007; Sun
and Cheng, 2002; Zhu et al., 2002). Dilute acid treatment is effect in removing
hemicellulose, with almost all the hemicellulose hydrolyzed and recovered as the
dissolved sugars such as xylose, glatactose, arabinose etc in the hydrolysate (Kumar et al.,
2009). The removal of hemicellulose exposes the cellulose to enzyme attack, increasing
the enzymatic digestibility and sugar yield in the residue solid left after the acid

33
pretreatment. Various agro-industrial residues, including corn fiber, corn cob, corn stover,
whey straw, whey bran, sugarcane bagasse, and cassava bagasse, have been studied under
different acid concentrations and residence times in search for an optimal condition (Kim
et al., 1999; Lu et al., 2007; Nuttha et al., 2009; Pandey et al., 2000a; Pandey et al., 2000b;
Qureshi et al., 2007; Zhu et al., 2002). A variety of degradation products (phenolic
compounds, furan derivatives, etc.) usually come with acid pretreatment (Saha, 2003).
Balancing the sugar yield, acid concentration and pretreatment time can control the
inhibitors present in the hydrolysate (Kumar et al., 2009), alleviating the stress on the
following fermentation process.
Alkaline pretreatment with strong bases like sodium hydroxide, potassium
hydroxide, calcium hydroxide, and ammonia hydroxide is also widely used. Compared to
acid pretreatment, alkaline pretreatment uses relatively mild conditions, such as room or
slightly elevated temperature and atmospheric pressure (Kumar et al., 2009; Mosier et al.,
2005). As a result of this mild condition, the duration of alkaline pretreatment usually
takes hours to days instead of few minutes. Elevated temperature can significantly reduce
the reaction time; therefore, 80-120oC is often used in alkaline pretreatment (Chang et al.,
1997; Chang et al., 1998; Chang et al., 2001). Among all the common strong bases, lime
is mostly chosen due to the competitive low price and renewability (Mosier et al.,
2005).Various feedstock have been treated with alkaline, such as bagasse, wheat straw,
corn stover, switchgrass, wood chips and more (Chang et al., 1997; Chang et al., 1998;

34
Chang et al., 2001; MacDonald et al., 1983). The main goal of the alkaline pretreatment
is to remove the lignin from biomass (Chang et al., 2001; Kumar et al., 2009; Mosier et
al., 2005), while hemicellulose is also partially dissolved leaving cellulose accessible to
enzymes. It was also reported that in the presence of an oxidizing agent such as oxygen,
the removal of lignin is greatly enhanced while cellulose in the biomass is not affected
(Chang et al., 2001).
2.2.2 Detoxification of lignocellulosic hydrolysate
Various byproducts, also known as inhibitors in the latter bioconversion process, are
generated during the pretreatment process. The major byproducts include furan
derivatives (furfural and 5-hydroxymethylfurfural (HMF), sugar degradation), phenolic
compounds (syringaldehyde, vanillin, syringic acid, vanillic acid, p-coumaric acid, ferulic
acid, lignin degradation), and weak acid (acetic acid, lignocellulose structure degradation)
(Ezeji et al., 2007b; Mussatto and Roberto, 2004a; Olsson and Hahn-Hagerdal, 1996).
Table 2.7 shows some major fermentation inhibitors present in the hydrolysate generated
during the pretreatment process due to lignocellulose degradation.
Pentose and hexose are released during the hydrolysis of lignocellulosic biomass,
and then further degraded into furfural and HMF, respectively (Mussatto and Roberto,
2004a). Furfural and HMF are generally recognized as the major inhibitors to the
microorganisms. Phenolic, aromatic compounds and aldehydes are degradation products

35
generated from lignin. These compounds, especially the low molecular weight ones, are
very toxic to the fermentation microorganisms, even when their concentrations are low
(Ezeji et al., 2007b; Mussatto and Roberto, 2004a; Parajo et al., 1998). Acetic acid is
derived from the acetyl side-groups of hemicellulose, and is considered as a product of
lignocellulosic structure degradation. The inhibitory effect of acetic acid is usually not as
severe as furan derivatives or phenolic compounds. At low concentrations, several reports
showed that acetic acid actually enhanced the solvent production and prevented the
culture degeneration (Chen and Blaschek, 1999; Felipe et al., 1995).
When using lignocellulosic hydrolysate all of the above mentioned substances can
cause some degrees of inhibition in the fermentation process. Due to the presence of
various inhibitors, the lag phase is prolonged, sugar utilization is reduced, and the product
formation (concentration, yield, productivity) is significantly hindered (Hendriks and
Zeeman, 2009; Mussatto and Roberto, 2004a; Palmqvist et al., 1999; Palmqvist and
Hahn-Hagerdal, 2000a; 2000b). The inhibitory concentration of each compound can not
be strictly determined due to the diversity of microorganism (Palmqvist and
Hahn-Hagerdal, 2000a). Moreover, it was reported that while an individual compound
may not cause inhibition, when in the presence with other compounds a signigifant
“synergistic effect” (Mussatto and Roberto, 2004a; Palmqvist et al., 1999; Palmqvist and
Hahn-Hagerdal, 2000b) may exhibit. Detoxification is usually needed to re-condition the
lignocellulosic hydrolysates to a suitable substrate for microorganisms to digest.

36
2.2.2.1 Physical detoxification
Physical detoxification usually uses vacuum evaporation technique to remove the
volatile toxic substances, such as furfural and acetic acid. Usually the furfural can be
efficiently removed by this method, and the sugar is concentrated after water evaporates.
The down side of this pretreatment is that non-volatile substances accumulate and stay in
the concentrated hydrolysate (Klinke et al., 2004; Mussatto and Roberto, 2004a).
2.2.2.2 Chemical detoxification
In general, chemical detoxification includes using pH adjustment to precipitate and
remove toxic substances, and adsorption with activated charcoal or ion-exchange resins
(Klinke et al., 2004; Martinez et al., 2001; Mussatto and Roberto, 2001; Mussatto and
Roberto, 2004b; Nilvebrant et al., 2001; Qureshi et al., 2008a). Since some inhibitors are
unstable at a certain pH, pH adjustment with Ca(OH)2 (lime) is the most commonly used
detoxification method for a variety of lignocellulose hydrolysates. Generally, lime is
added to adjust the pH to 9-10, and then acid (H2SO4 or HCl) is added to readjust pH to
5.5-6.5 (Ezeji et al., 2007b; Ezeji and Blaschek, 2008; Martinez et al., 2001; Palmqvist
and Hahn-Hagerdal, 2000a; Qureshi et al., 2008a). It was reported that overlime
detoxification reduced over 51% of furans, 41% of phenolic compounds, and only 8.7%
of sugars (Martinez et al., 2001).

37
Activated charcoal and ion-exchange resins can be used to adsorb the toxic
chemicals to detoxify the hydrolysate. It was reported that using activated carbon alone
with rice straw hydrolysate, 27% phenolic compounds were removed and similar yield
and productivity were achieved compared to control, suggesting successful removal of
inhibitors (Mussatto and Roberto, 2001). Later Mussatto et al. (2004) reported that pH
was an important factor in the activated carbon detoxification. Adjusting the initial pH of
rice straw hydrolysate from 0.4 to 2.0 followed by addition of 2.5% activated carbon,
72.9 % HMF, 89.3% furfural and 34.3% lignin degradation products were removed with
sugar loss less than 11.5%. Increasing the operating temperature for activated carbon
adsorption also helped to remove more inhibitors (Mussatto and Roberto, 2004a; 2004b).
Ion-exchange resins are usually effective but pricy (Mussatto and Roberto, 2004a;
Nilvebrant et al., 2001). Three different resins (anion, cation, and hydrophobic) were
studied by Nilvebrant et al. (2001). Anion resin AG 1-X8 (OH-) performed the best under
pH 10, followed by XAD-8 hydrophobic resin, and then the cation resin AG 50W-X8.
Sometimes several detoxification methods are combined to achieve a better inhibitor
removal rate (Converti et al., 2000; Ezeji and Blaschek, 2008; Qureshi et al., 2008a). A
three-step detoxification, first with overlime, second with heat and gas stripping, and
third with activated carbon was reported by Converti et al. (2000). Overliming and
activated carbon removed 95% of the lignin degradation products, and acetic acid and
furfural were removed by boiling. Qureshi et al. (2008) also reported to treat corn fiber

38
hydrolysate first with modified overliming method and then with XAD-4 resin, which
resulted in significant ABE production (9.3 g/L) compared to 1.7 g/L in untreated corn
fiber hydrolysate.
2.3 Product recovery and separation technologies
No matter whether it is to produce fuel-grade ethanol or butanol, multi-column
distillation followed by molecular sieve adsorption has always been the standard
operation procedure in the industrial process. Distillation offers a wide range of
advantages, such as high alcohol recovery, multi-stage operation, being easy to scale-up,
and relatively energy-efficient when alcohol concentration in the feed stream is high.
There are also many less-attractive facts about recovering alcohol using distillation, such
as energy-intensive for low alcohol concentration feed, high-temperature operation which
is lethal to microorganisms, and necessity for an additional dehydration step in order to
reach the fuel-grade specification Vane, 2008). Because the ethanol concentration is
usually high at the end of the process (~10%), distillation is favorable for ethanol
recovery.
Butanol recovery is the most energy-intensive and costly step in the whole
biobutanol production process (Ezeji et al., 2004a; 2007a). In ABE fermentation, the
butanol final concentration is usually 1-2% in the fermentation broth. Recovering butanol
using distillation is thus extremely energy-intensive and costly. Unlike ethanol, butanol

39
has a low vapor pressure and high boiling point (118 oC), which pose further challenges
in distillation and require more energy. Alternative separation technologies that are
energy-efficient and suitable to recover low concentration alcohol in the fermentation
broth are in demand. Over the years many relatively economic and feasible techniques,
including gas stripping, liquid-liquid extraction, adsorption, pervaporation and
perstraction, have been developed to recovery solvents from the fermentation broth.
These technologies are more energy-efficient than the traditional distillation approach in
terms of lowering the process cost. Table 2.8 summarizes and compares the pros and cons
of these alternative butanol recovery methods.
There are usually two alcohol recovery approaches from the fermentation broth,
“end-of-pipe” and “slip-stream” as referred by Vane (2008). The end-of-pipe approach
refers to the alcohol recovery after the fermentation is completed, and the
alcohol-depleted broth is sent to the next step for processing. This approach is usually
employed in ethanol recovery from fermentation due to the high end product
concentration present in the feed stream. Slip-stream approach refers to alcohol recovery
while the fermentation is still on-going in the bioreactor, and the alcohol-depleted stream
is returned to or never leaves the bioreactor. This process is also known as the integrated
process, meaning that the separation technology is integrated with fermentation and the
desired product can be in-situ recovered simultaneously. The slip-stream approach is
mostly seen in butanol recovery due to the severe end product inhibition on

40
microorganisms caused by butanol. By employing the slip-stream approach, the butanol
inhibition is relived and the butanol-free broth is recycled back into the bioreactor,
increasing the volumetric productivity of the reactor (Vane, 2008). It is clear that due to
the high temperature employed in the distillation process, distillation can only be used in
end-of-pipe approach, while the alternative separation technologies such as gas stripping
and pervaporation can be used in slip-stream approach to increase the reactor productivity
and overall butanol concentration.
2.3.1 Gas stripping
Gas stripping is an easy-to-operate technique to recover butanol from fermentation
broth. Figure 2.2A shows a schematic diagram of a typical gas stripping process. Gas
stripping can either be integrated with fermentation in the bioreactor, or performed in an
individual stripping column. Therefore, the gas stripper shown in Figure 2.2A can either
be a bioreactor or a separate stripping column. In ABE fermentation, either nitrogen or
fermentation gases (H2 and CO2) can be used as stripping gases (Ezeji et al., 2004a) to
ensure the anaerobic condition. In the integrated scenario, stripping gas is introduced to
the fermentation broth in the bioreactor and captures the volatile solvents in the broth,
and the gas containing solvents is subsequently passed through a condenser where the
solvents are condensed and enriched in the condensate stream. In the separate gas stripper
scenario, feed stream (broth) is sent to the stripper where the solvents are captured by

41
stripping gas, and the feed low in solvents is then recycled back to the bioreactor. Gas
flow can also be operated in either single-pass mode or recycle mode. In the single-pass
mode, once gas passes condenser it is released into open air, which may result in solvent
loss depending on the efficiency of the condenser. In the recycle mode, gas free of
solvents after condensing is recycled back into the stripper/bioreactor to capture more
solvents, and the process is a closed loop which prevents any solvent loss.
Gas stripping offers many advantages as an integrated product recovery technology
with fermentation, including utilization of fermentation gases as stripping gas, ability to
operate under fermentation temperature and flexibility with or without solids removal
from fermentation broth (Vane, 2008). The principle behind this technology is the solvent
to water ratio in the inert gas at equilibrium, which is strongly governed by temperature.
The partial pressure of any component i in the gas phase can be expressed at following:
Pi = yiPtotal = xiγiPisat
Pi is the partial pressure, yi and xi are the mole fractions in the gas and liquid phase,
respectively, whereas Ptotal is the total pressure of gas phase and Psat is the saturated vapor
pressure of component i at the current temperature. Psat is strongly determined by
temperature, increasing temperature increases the Psat, which further affects the Pi in the
gas phase (Vane, 2008). It can be seen that increasing temperature favors higher vapor
phase concentration; unfortunately, this principle applies to both volatile solvents and
water. Many other factors also affect the performance of gas stripping, such as bubble

42
size, mass transfer coefficient, interfacial contact area and contact time, cooling
temperature and gas flow rate. Ezeji et al. (2005a) studied the effect of bubble size and
gas flow rate on butanol removal, and reported that a bubble size between 0.5 to 5 mm
had no effect on butanol removal rate under the condition tested, whereas increasing the
flow rate from 43 cm3/s to 80 cm3/s resulted in a 2.51-fold increase in gas-stripping rate
constant. They reported that in a 2.0 L reactor the gas bubbles had sufficient contact time
to be 95% saturated with butanol within 0.14 s, and smaller bubbles (< 0.5 mm) were not
necessary. Ezeji et al. (2005a) also mentioned that further reducing the bubble size had no
impact on increasing the solvent stripping rate, but actually reduced the reactor
productivity. Ezeji et al. (2003) studied butanol removal using model solution and real
fermentation broth, and they reported that gas stripping was highly selective towards
butanol over acetone, and the presence of cells in the fermentation broth adversely
affected butanol removal. No acids were taken out by gas stripping during the process,
indicating that gas stripping was only selective towards volatile solvents (Ezeji et al.,
2003). It has also been reported that gas stripping did not harm cells or remove any
nutrients from the broth when integrated with fermentation (Qureshi and Blaschek,
2001a).
Gas stripping has been successfully demonstrated and applied in many fermentation
processes and improved overall butanol production and productivity (Ezeji et al., 2003;
2004b; 2005b; 2007c). Table 2.9 summarizes performance and solvent selectivities of gas

43
stripping processes for butanol recovery under various operating conditions. With
simultaneous product removal, concentrated substrate can be utilized by microorganism
in an integrated fermentation process, which would otherwise cause substrate inhibition.
It was reported (Ezeji et al., 2003) that 161.7 g/L glucose was utilized and 75.9 g/L ABE
were obtained in a batch process integrated with gas stripping, whereas only 17.7 g/L
ABE were produced and 45.4 g/L glucose was consumed in the control non-integrated
batch process. If operated in fed-batch mode, highly concentrated substrate can be
utilized and substrate inhibition can be avoided. Ezeji et al. (2004b) reported that 500 g/L
glucose was periodically added into the reactor to replenish depleted sugar, and in total
500 g glucose were utilized by the bacteria in a 1.0 L reactor in fed-batch fermentation
with gas stripping as the product removal technique. 232.8 g/L ABE were obtained from
this integrated fed-batch process with an enhanced productivity of 1.16 g/L. Besides
batch and fed-batch fermentations, a continuous ABE fermentation was reported to utilize
1163 g/L glucose, resulting in a total 460 g/L ABE production with 0.91 g/L·h
productivity with solvents recovered by gas stripping.
Currently, gas stripping has not been commercially used for ethanol or butanol
recovery. The alcohol-rich condensate from gas stripping requires at least one additional
step for alcohol dehydration in order to meet the fuel-grade specifications. Vane (2008)
suggested that in case of butanol recovery by gas stripping, phase separation is a feasible
choice due to the high alcohol (butanol) concentration in the condensate. Besides process

44
design and unit fabrication, he also suggested that improvement on mass and energy
integration schemes for gas stripping is needed in order to make this process
economically feasible and attractive.
2.3.2 Pervaporation
Pervaporation is a membrane-based separation technique. Liquid feed containing
volatile species flows on one side of the membrane, while the other side of the membrane
is under vacuum. Components of the liquid stream, depending on the chemical properties,
penetrate and diffuse through the membrane and evaporate into permeate vapor under
vacuum. The permeate vapor is then condensed in cooling trap as condensed permeate
(Vane, 2005; 2008; Thongsukmak and Sirkar, 2007). A schematic diagram of
pervaporation is depicted in Figure 2.2B.
Pervaporation is a selective separation process based on the membranes employed in
the module. Components in the liquid feed have different chemical and physical
properties; some components have similar properties to the selective membrane material,
and can diffuse through the membrane and enrich in the permeate side, while others stay
on the other side of the membrane. The concentration of solvents on the permeate side is
a function of feed concentration, and depends on the composition and selectivity of the
membrane used (Ezeji et al., 2004a; Vane, 2005; 2008). When the selected components
diffuse to and enrich in the permeate side, the concentration of these components is

45
reduced in the liquid feed, and the retentate leaving the module is low in concentration of
the selected components, completing the separation process. Due to the selective nature
of the membrane and diffusion rates of different components, the concentration ratio of
one component in permeate to feed can range from single digit to over a 1000 (Vane,
2005). If the membrane is hydrophobic, the permeate side will enrich in organic
compounds relative to water. If the membrane is hydrophilic, the feed liquid will be
dehydrate as water permeates through the membrane, which is the primary commercial
(Jonquieres et al., 2002) use of pervaporation process. The chemical activity difference
(concentration gradient) on the feed side and the permeate side is the driving force for a
component to transport across the membrane, and the flux is inversely proportional to the
overall resistance and proportional to the concentration gradient (Vane, 2005). The
resistance to transport across the membrane includes diffusion in the stagnant feed liquid
to the membrane, diffusion through the membrane, and diffusion in the permeate vapor.
It has been concluded that the primary factors affecting the separation by pervaporation
are membrane materials and feed species, whereas feed temperature, composition and
permeate pressure are only secondary factors (Vane, 2005).
In the case of butanol separation from water by pervaporation, a hydrophobic
membrane is needed in order to get butanol-rich condensate on the permeate side. It has
been widely studied and reported that pervaporation has been employed in butanol
recovery from water or fermentation broth (Geng and Park, 1994; Fadeev et al., 2000;

46
Fadeev et al., 2001; Jonquieres and Fane, 1997; Qureshi and Blaschek, 1999a; 1999b;
Qureshi et al., 1999; 2001a). Table 2.10 presents various membranes that have been
applied in the pervaporation process for butanol recovery and their performances.
Currently, the poly(dimethyl siloxane) membrane, which is also known as PDMS or
silicone rubber membrane, is the benchmark of hydrophobic membrane commonly used
in alcohol/water separation by pervaporation (Vane, 2005; 2008). PDMS membrane
offers a separation factor of 4.4-10.8 for ethanol/water system, and 40-60 for
butanol/water separation (Vane, 2005). Many factors affect the performance of
pervaporation with PDMS membranes, such as operating temperature, feed concentration,
thickness of the membrane, and PDMS source and fabrication procedure. There have
been research efforts trying to improve on the performance of pervaporation using PDMS.
Recently, Li et al. (2010) reported using a tri-layer PDMS composite membrane
(PDMS/PE/brass support) to recover butanol by pervaporation, and a separation factor of
34 was obtained. Other polymeric materials have been studied as potential membranes
for pervaporation, including polypropylene (PP) and PTFE, and low separation factors
ranging from 3-9.5 for butanol/water separation were reported (Qureshi et al., 1992;
Vrana et al., 1993). Besides PDMS, poly[1-(trimethylsilyl)-1-propyne], also known as
PTMSP, is another polymeric/organic membrane that offers good alcohol/water
separation factor. PTMSP has high free volume in the membrane, offering more void
spaces for higher permeability than PDMS (Volkov et al., 1997; 2004). It has been

47
reported that the butanol/water separation factor in PTMSP can reach as high as 70
(Fadeev et al., 2001). However, due to the high free volume, which attracts foulants
inside the membrane, the performance of PTMSP is not as stable as PDMS, and the flux
and selectivity of PTMSP gradually decrease over time (Fadeev et al., 2003; Schmidt et
al., 1997).
In addition to polymeric membranes, inorganic zeolite materials, such as silicalite
and Ge-ZSM-5, have also been studied as hydrophobic membranes in pervaporation
applications (Li et al., 2003; Sano et al., 1994). These inorganic materials are usually
supported by a solid frame, such as stainless steel, in order to act as a membrane. Li et al.
(2003) studied the stainless steel supported Ge-ZSM-5 membrane on ethanol, methanol,
butanol, and 2-propanol separation through pervaporation. The ethanol/water separation
factor was reported at 47, which was at least 4-5 fold higher than PDMS, but the
butanol/water separation factor was lower than PDMS, only at 19. Silicalite also
delivered excellent separation factor for ethanol/water separation, with 60 reported by
Sano et al. (1994) and an average of 40 widely reported in the literature (Vane, 2005).
However, the biggest downside associated with the inorganic membrane is the fabrication
cost. Therefore, it has also been proposed that silicalite can be dispersed in PDMS to
fabricate a mixed matrix membrane to incorporate the advantages of both zeolite and
PDMS. A wide range of butanol/water separation factors of 50-111 (Huang and Meagher,
2001), 55-209 (Qureshi and Blaschek, 1999a), 70-97 (Qureshi et al., 2001a), and 100-108

48
(Qureshi et al., 1999) have been reported in the literature using ABE model solution or
real fermentation broth. Compared with typical 40-60 separation factor for butanol/water
separation in PDMS membrane, the addition of these inorganic silicalite improved the
performance of PDMS. The fabrication process of the mixed matrix silicalite/PDMS
membrane is similar as PDMS, and the cost is expected to be close to PDMS, yet
significantly lower than the inorganic membrane (Vane, 2005).
In addition to the afore-mentioned membranes, which are solid membranes,
supported liquid membrane has also been studied in pervaporation process to recover
alcohol from dilute aqueous solutions (Izak et al., 2008; Matsumura and Kataoka, 1987;
Thongsukmak and Sirkar, 2007). Oleyl alcohol is the common material employed in
supported liquid membrane, and a high 180 butanol/water separation factor was reported
using a porous PP supported oleyl alcohol liquid membrane in the pervaporation process
(Matsumura and Kataoka, 1987). The general requirement for liquid membrane is that the
organic solvent is biocompatible with the microorganisms in the fermentation, which
would otherwise be toxic to the culture and hinder the fermentation and decrease the life
of liquid membrane. In general, the solvent concentration in the permeate from the
liquid-membrane based pervaporation is higher than that from the polymeric and ceramic
membrane based pervaporation process (Thongsukmak and Sirkar, 2007). A major
problem associated with the liquid membrane is that the liquid leaks into the fermentation
broth over time and the liquid membrane has to be regenerated. Thongsukmak and Sirkar

49
(2007) employed a novel nanoporous coating (fluorosilicone) on the polypropylene
hollow fiber as the support material to prevent and minimize the migration of liquid
membrane into the fermentation broth, and used trioctylamine (TOA) as a liquid
membrane for butanol recovery through pervaporation. A butanol selectivity of 108-141
was reported in this porous PP hollow fiber supported TOA liquid membrane using
model butanol solution, and a selectivity of 71-104 was reported using ABE mixture
model solution. Pervaporation via a supported ionic liquid membrane integrated with
ABE fermentation enhanced the solvent productivity to 2.34 g/L h (Izak et al., 2008).
Tetrapropylammonium tetracyano-borate was the ionic liquid used in the study, which
was supported by PDMS as a supported liquid membrane. The butanol enrichment factor
was reported to be 11.23 in this study.
The employment of a membrane in pervaporation makes it very efficient and highly
selective, even a low concentration species in the feed mixture can be enriched
significantly through pervaporation if using a suitable membrane (Vane, 2005). However,
due to the employment of the membrane, the performance of pervaporation is very
sensitive and can be affected by many factors when integrated with on-going
fermentation process. Fouling is the most common problem with any membrane-based
separation technology (Fadeev et al, 2000; Qureshi and Blaschek, 1999c). Vane (2005)
summarized a list of factors that impede the performance of pervaporation by
fermentation broth, including dead cells, suspended solids, cell metabolites, sugars,

50
organic acids and fatty acids. Dead cells and suspended solids are the mostly likely to
accumulate in the pervaporation membrane module, clog the pores and block the flow
path. Organic acids are the second group that impacts on the pervaporation performance,
and competitive sorption with alcohol has been proposed as a potential mechanism. Since
acids are in the undissociated form at pH lower than their pKa value, increasing the pH to
4-6 can significantly reduce the impact of acids on the membranes.
Pervaporation is an emerging membrane-based technology with high selectivity to
efficiently recover alcohol from dilute aqueous solutions. Many obstacles are still to
overcome in order to develop a process suitable for commercial application for butanol
recovery, such as membrane fouling and high fabrication cost. Membranes that are highly
permeable to alcohol with good alcohol/water separation factor are desired for
pervaporation, and the stability of membrane over extended period of time is required.
Vane (2008) suggested that a silicone rubber (PDMS) membrane coupled with efficient
vapor condensation and dehydration system is a good choice for butanol recovery.
2.3.3 Liquid-liquid extraction
Liquid-liquid extraction is another alternative separation technique proposed for
recovering butanol from dilute aqueous solution. Extractant liquid is placed in contact
with fermentation broth, and solvents transport from fermentation broth into the
extractant phase due to the solubility difference, thus being separated from the aqueous

51
solution (Ezeji et al., 2004a; Vane, 2008). This broth/extractant contact can be either
done in a direct way, i.e. mixing, or an indirect way, i.e. using a membrane to separate the
two phases. The latter procedure is often referred as perstraction (Ezeji et al., 2007a;
Vane, 2008). The employment of membrane in perstraction to separate the two phases is
to avoid problems usually associated with traditional liquid-liquid extraction, including
toxicity to microorganisms, emulsion, loss of extractant, and transfer of cells from broth
to extractant phase (Ezeji et al., 2007a). After the extractant is enriched with alcohols,
these alcohols must be removed and recovered in a regeneration unit in order to get the
desired product and recycle the extractant back into the process. Common extractant
regeneration methods include: distillation, vacuum evaporation, and pervaporation (Ezeji
et al., 2004a; Vane, 2008).The schematic design of a liquid-liquid extraction process is
depicted in Figure 2.2C.
There are many requirements a solvent must meet in order to be considered as a
suitable extractant to recover butanol via liquid-liquid extraction. Ennis et al. (1986),
Maddox (1989), and Vane (2008) discussed and summarized these factors in details.
1. High selectivity of alcohol to water (separation factor)
2. High distribution coefficient, which reduces the volume of extractant needed to
recover the same amount of alcohol
3. Immiscible, non-emulsifying, clear phase separation from aqueous solution

52
4. Non-toxic to microorganisms, non-reactive with fermentation components, and
non-flammable to ensure safety when operating
5. Inexpensive to use and easily available
Table 2.11 shows the performance and toxicity of some solvents evaluated for
butanol recovery by liquid-liquid extraction processes. Thirty-one commonly used
solvents were evaluated as extractants in an extractive ABE fermentation process by C.
acetobutylicum (Barton and Daugulis, 1992), and some of the good candidates reported
were poly(propylene glycol) (PPG) 1200, PPG 2000, oleyl alcohol, isophytol, eutanol G
and triethyl citrate, based on butanol partition coefficient and biocompatiblity. It was
reported in their work (Barton and Daugulis, 1992) that an extractive ABE fermentation
using PPG 1200 resulted in 58.6 g/L acetone and butanol, which was 3-fold higher than
the production in the control study. Besides poly(propylene glycol), n-decanol,
dibutyl-phthalate, and oleyl alcohol have also been reported as suitable extractants to
recover butanol with high partition coefficients and low toxicity (Eckert and Schügerl,
1987; Wayman and Parekh, 1987; Roffler et al., 1988). Oleyl alcohol is the most often
used and investigated extractant in butanol recovery (Ezeji et al., 2004a; Roffler et al.,
1987a; 1987b). It was reported (Qureshi and Maddox, 1995) that oleyl alcohol was the
most effective candidate in extracting butanol and the least in reducing the productivity.
Roffler et al. (1987a) studied six solvents/solvent mixtures as extractants in extractive
ABE fermentation, including kerosene, tetradecanol, oleyl alcohol, dodecanol, benzyl

53
benzoate, and reported that oleyl alcohol or oleyl alcohol and benzyl benzoate mixture
resulted in the best result in batch fermentation. Glucose consumption was improved
from 80 g/L to over 100 g/L, with a 60% increase in volumetric butanol productivity.
19.7 g/L and 19.3 g/L butanol was produced in oleyl alcohol and oleyl alcohol with
benzyl benzoate extractive fermentations, respectively, compared with 14.6 g/L butanol
obtained in control batch fermentation. In an extractive fed-batch ABE fermentation,
oleyl alcohol was mixed with broth at a ration of 1, 1.5, and 2.3, and the final butanol
production achieved in each process was 32 g/L, 45 g/L and 63 g/L, respectively (Roffler
et al., 1987b). This indicated that with a high extractant/broth ratio, more butanol was
recovered in the extractant phase and end product inhibition was relieved on
microorganisms, resulting in higher total butanol production. In each scenario,
fermentation stopped when butanol concentration reached 30-35 g/L in the extractant
phase, indicating that the saturation point of butanol in oleyl alcohol is about this
concentration. Oleyl alcohol has also been suggested (Evans and Wang, 1988) as a
co-extractant to mix with other extractants that have high partition coefficient yet toxic to
the cells to obtain an extractant mixture with overall high partition coefficient and
relatively low toxicity.
The most commonly studied traditional extractants are usually long-chain alcohols,
alkanes, esters, fatty acids and oils (Vane, 2008). Some novel materials such as ionic
liquid or biodiesel has also been suggested as potential extractant for butanol recovery via

54
liquid-liquid extraction. Ionic liquid (IL) is a group of salts that exist in the liquid form at
low temperature (<100oC) or room temperature, and is considered as a green and safe
solvent due to its thermally and chemically stable properties (Earle and Seddon, 2000;
Fadeev and Meagher, 2001; Hagiwara and Ito, 2000; Huddleston et al., 1998; Seddon,
1997; Toh et al., 2006; Zhao et al., 2005). The miscibility and hydrophobicity of ILs can
be adjusted by manipulating the structure of anions and cations. It was reported that
anions determine the water miscibility of ILs, whereas cations have more influence on the
hydrophobicity of ILs (Zhao et al., 2005). ILs have been used as extractants in many
areas, such as metal ions (Wei et al., 2003), carbohydrates (Liu et al., 2005), organic
acids (Matsumoto et al., 2004), and biofuels (Fadeev and Meagher, 2001). [PF6]- based
ILs are usually water-immiscible, and 1-butyl-3- methyl-1H-imidazol-3-ium
([BMIM][PF6]) has been identified as a suitable extractant for butanol recovery (Fadeev
and Meagher, 2001). In addition to ILs, biodiesel is another exotic extractant proposed
for butanol recovery (Adhami et al., 2009; Li et al., 2010). Biodiesel can be utilized as
diesel fuel; with butanol added into biodiesel via extraction the fuel properties of
biodiesel can be enhanced. Li et al. (2010) reported that biodiesel preferably extracted
butanol with a partition coefficient of 1.23, and the fuel properties of ABE-enriched
biodiesel were significantly improved, with the cetane number increasing from 48 to 54,
and cold filter plugging point decreasing from 5.8 to 0.2 oC.

55
The alcohol-rich product recovered by liquid-liquid extraction usually requires
additional steps for dehydration and purification, and the concentration of alcohol in the
extractant strongly depends on the selectivity of the extractant. The regeneration step is
the most energy-intensive procedure in liquid-liquid extraction, and it was suggested that
a butanol/water separation factor of 30-50 is desirable to significantly reduce the
theoretical energy demand (Vane, 2008).
2.3.4 Adsorption
Besides the above-mentioned techniques, adsorption is another alternative separation
process for butanol recovery. Butanol is first adsorbed by adsorbent materials in a packed
column from dilute solution during the loading cycle, and then desorbed by heating the
adsorbent to obtain a concentrated butanol solution during the regeneration cycle (Vane,
2008). The adsorption process is similar to liquid-liquid extraction. Adsorbent needs to be
regenerated to recover the adsorbed butanol and for continued reuse, and high separation
factor and distribution coefficient are two key parameters in selecting proper adsorbent
materials. A typical adsorption process is illustrated in Figure 2.2D.
The most commonly used adsorbent material for alcohol recovery is hydrophobic
zeolites, especially silicalite-1 (Groot et al., 1992; Holtzapple and Brown, 1994;
Milestone and Bibby, 1981; Oudshoorn et al., 2009; Qureshi et al., 2005). Other materials
such as resin, activated carbon, and polyvinylpyridine have also been suggested and

56
studied as adsorbent materials for alcohol recovery (Groot and Luyben, 1986; Nielsen
and Prather, 2009; Qureshi et al., 2005). Table 2.12 lists the butanol adsorption capacities
and performances of different adsorbent materials for butanol recovery by adsorption
processes. Milestone and Bibby (1981) reported that using silicalite as adsorbent in
butanol recovery from a dilute 0.5% solution, a 98% concentrated butanol was obtained
by heating the adsorbent to 150 oC after preliminary drying at 40 oC to remove bulk water.
Oudshoorn et al. (2009) evaluated three zeolites of different structures and SiO2/Al2O3
ratios, and reported that the zeolite with the lowest SiO2/Al2O3 ratio had the highest
capacity for butanol adsorption (high distribution coefficient). Zeolite with ZSM-5
structure and high SiO2/Al2O3 ratio showed excellent affinity to butanol even when
butanol concentration was low, indicating that the affinity to butanol was associated with
the hydrophobicity of the zeolite. It was mentioned that the presence of cells did not
affect the butanol adsorption behavior of all the zeolites investigated. Nielsen and Prather
(2009) investigated and compared the performance of several commercially available
resins in butanol recovery by adsorption. They identified two resins of
poly(styrene-co-divinylbenzene) that showed the best n-butanol affinity, and concluded
that the butanol partition coefficient of resins was determined by the specific surface
areas. Due to the high specific loadings of resin (266- 403 g-butanol/kg-resin), butanol
was recovered by vacuum evaporation at 100 oC with 78-85% recovery efficiency, and
this process was predicted to be economically favorable. Qureshi et al. (2005) extensively

57
studied various adsorbent materials, including silicalite, resins, bone charcoal, activated
charcoal, bonopore, and polyvinylpyridine, providing a systematic comparison of butanol
adsorption efficiency using these materials. Bone charcoal and activated carbon were
reported to have the highest butanol adsorption capacity using ABE model solution, but
the adsorbed butanol on these materials could not be completely recovered during the
desorption process. Silicalite was suggested to be the most appealing adsorbent,
concentrating butanol to 810 g/L from a 5 g/L dilute feed solution with a complete
butanol recovery in the desorption process. A comparison of energy input for ABE
separation using alternative recovery technologies was illustrated, and adsorption was
projected to be the most energy-efficient, followed by liquid-liquid extraction,
pervaporation, gas stripping, and steam stripping distillation (Qureshi et al., 2005).
Adsorbent fouling by cells and adsorption of other fermentation components, such as
nutrients, substrates and acids, have been the major concerns of applying adsorption
technology with fermentation to recover alcohols (Vane, 2008). In order to avoid fouling
by cells, it was suggested that a membrane-assisted cell recycle or cell removal by
centrifuge could be considered in the integrated fermentation process with adsorption
(Nielsen et al., 1988; Yang and Tsao, 1995).

58
2.4 Integrated ABE fermentation process with online product recovery
Butanol is the most desired product from ABE fermentation and yet the most toxic
product to the culture. Severe butanol inhibition exists in ABE fermentation which results
in low final butanol concentration, low yield and low productivity. All these limitations
hamper the economic application of deriving biobutanol from ABE fermentation.
In a typical batch ABE fermentation, only 15-18 g/L total ABE can be obtained
through a period of 40-60 h until the fermentation stops due to inhibition (Woods, 1995).
The in-situ recovery of butanol is therefore crucial in improving the reactor performance.
Simultaneous butanol recovery can relieve the product inhibition and leads to a more
complete conversion of the carbon source. It allows the usage of a concentrated feed and
extends the fermentation period (Dürre, 1998; Ezeji et al., 2004a; Groot et al., 1990).
Moreover, online butanol recovery also simplifies the downstream separation process,
which lowers the energy consumption and brings down the whole process cost. In the
past, distillation was widely employed to recover butanol, and it was proved to be costly
due to the low butanol concentration in the broth (Ezeji et al., 2004a). In recent years,
new advances in butanol recovery techniques such as liquid-liquid extraction,
pervaporation and gas stripping are integrated with fermentation in an effort to develop a
commercial process for biobutanol synthesis (Ezeji et al., 2003; 2005a; Groot et al., 1990;
Izak et al., 2008; Qureshi and Maddox, 1995; Vane, 2005). These integrated fermentation
processes have been shown to be superior in aspects of sugar consumption, ABE final

59
concentration and ABE productivity. Some of the reported research on integrated ABE
process are summarized and compared in Table 2.13.
From Table 2.13, it is clear that online butanol recovery can increase the final ABE
concentration and the reactor productivity significantly. Since butanol is timely removed
from the fermentation broth, the butanol left in the reactor never exceeds the inhibitory
level, which results in a higher sugar utilization rate. Compared to batch control
fermentation, online butanol recovery allows the usage of highly concentrated feed
solution in fed-batch and continuous processes in an extended period of time, which
further lead to high ABE production. Some of the integrated processes also exhibit the
potential of commercializing the ABE fermentation at the industry scale. It was reported
(Ezeji et al., 2005b) that gas stripping integrated with continuous fermentation utilized
1163 g/L glucose and produced 460 g/L ABE in total. This illustrated that integrating
simultaneous product recovery with ABE fermentation is promising in enhancing the
fermentation efficiency and process economics in an effort of eventually developing a
commercial process.
2.5 References
Afschar, A.S., H. Biebl, K. Schaller, and K. Schugerl (1985). Production of acetone and butanol by Clostridium acetobutylicum in continuous culture with cell recycle. Appl. Microbiol. Biotechnol., 22, 394-398.
Ahn, J.H., B.I. Sang, and Y. Um (2011). Butanol production from thin stillage using Clostridium pasteurianum. Bioresour. Technol., 102, 4934-4937

60
Atsumi, S., A.F. Cann, M.R. Connor, C.R. Shen, K.M. Smith, M.P. Brynildsen, K.J. Chou, T.Hanai, J.C. Liao (2007). Metabolic engineering of Escherichia coli for 1-butnaol production. Metab. Eng., 10, 305-311.
Bahl, H., W. Andersch, K. Braun, and G. Gottschalk (1982). Effect of pH and butyrate concentration on the production of acetone and butanol by Clostridium acetobutylicum grown in continuous culture. European J. Appl. Microbiol. Biotechnol., 14, 17-20.
Badr, H.R., R. Toledo, and M.K. Hamdy (2001). Continuous acetone-ethanol-butanol fermentation by immobilized cells of Clostridium acetobutylicum. Biomass and Bioenergy, 20, 119-132.
Barton, W.E. and A.J. Daugulis (1992). Evaluation of solvents for extractive butanol fermentation with Clostridium acetobutylicum and the use of poly(propylene glycol) 1200. Appl. Microbiol. Biotechnol., 36, 632-639.
Beesch, S.C. (1952). Acetone-butanol fermentation of sugars. Ind. Eng. Chem., 44, 1677-1682.
Biebi, H (2001). Fermentation of glycerol by Clostridium pasteurianum – batch and continuous culture studies. J. Ind. Microbiol. Biotechnol., 27,18-26.
Boddeker, K.W., G. Bengtson and H. Pingel ( 1990). Pervaporation of isomeric butanols. J. Memb. Sci., 54, 1-12.
Bruant, G., M.J. Levesque, C. Peter, S.R. Guiot, L. Masson (2010). Genomic analysis of carbon monoxide utilization and butanol production by Clostridium carboxidivorans strain P7. PLoS ONE, 5 (9), article number e13033.
Cadoche, L. and G.D. Lopez (1989). Assessment of size reduction as a preliminary step in the production of ethanol from lignocellulosic wastes. Biol. Waste., 30, 153-157.
Cara, C., M. Moya, I. Ballesteros, M.J. Negro, A. Gonzalez and E. Ruiz (2007). Influence of solid loading on enzymatic hydrolysis of steam exploded or liquid hot water pretreated olive tree biomass. Process Biochemistry, 42, 1003-1009.
Chang, V.S., B. Burr and M.T. Holtzapple (1997). Lime pretreatment of switchgrass. Appl. Biochem. Biotechnol., 63, 3–19.

61
Chang, V.S., M. Nagwani and M.T. Holtzapple (1998). Lime pretreatment of crop residues bagasse and wheat straw. Appl. Biochem. Biotechnol., 74, 135–159.
Chang, V.S., M. Nagwani, C.H. Kim and M.T. Holtzapple (2001). Oxidative lime pretreatment of high-lignin biomass. Appl. Biochem. Biotechnol., 94, 1–28.
Chen, C.K. and H.P. Blaschek (1999). Acetate enhances solvent production and prevents degeneration in Clostridium beijerinckii BA 101. Appl. Microbiol. Biotechnol., 52, 1701-73.
Cho, D.H., Y.J. Lee, Y. Um, B.I. Sang, Y.H. Kim (2009). Detoxification of model phenolic compounds in lignocellulosic hydrolysates with peroxidase for butanol production from Clostridium beijerinckii. Appl. Microbiol. Biotechnol., 83, 1035-1043.
Converti, A., J.M. Dominguez, P. Perego, S.S. Silva and M. Zilli (2000). Wood hydrolysis and hydrolysate detoxification for subsequent xylitol production. Chem. Eng. Technol., 23, 1013-1020.
Cornillot, E., R. Nair, E.T. Papoutsakis, P. Soucaille (1997). The genes for butanol and acetone formation in Clostridium acetobutylicum ATCC 824 reside on a large plasmid whose loss leads to degeneration of the strain. J. Bacteriol., 179, 5442-5447.
Delgenes, J., R. Moletta and J.M. Navarro (1996). Effects of lignocellulose degradation products on ethanol fermentations of glucose and xylose by Saccharomyces cerevisiae, Zymomonas mobilis, Pichia stipitis and Candida shehatae. Enzyme Microb. Technol., 19, 220-225.
Demain, A.L. (2009). Biosolutions to the energy problem. J. Ind. Microbiol. Biotechnol., 36, 319-332.
Dien, B.S., R.B. Hespell, L.O. Ingram and R.J. Bothast (1997). Conversion of corn milling fibrous co-products into ethanol by recombinant Escherichia coli strains K011 and SL40. World J. Microbiol Biotechnol., 13, 619-625.
Duff, S.J.B. and W.D. Murray (1996). Bioconversion of forest products industry waste cellulosics to fuel ethanol: a review. Bioresour. Technol., 55, 1-33.

62
Dürre, P. and H. Bahl (1996). Microbial production of acetone/butanol/isopropanol. In: Roehr M (ed) Biotechnoloy, 2nd edition, vol 6.VCH, Weinheim, pp 229-268.
Dürre, P. (1998). New insights and novel developments in clostridial acetone/ butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Dürre, P. (2007). Biobutanol: An attractive biofuel. Biotechnol. J., 2, 1525-1534.
Earle, M.J. and K.R. Seddon (2000). Ionic liquids. Green solvents for the future. Pure Appl. Chem., 72, 1391-1398.
Eckert, G. and K. Schügerl (1987). Continuous acetone-butanol production with direct product removal. Appl. Microbiol. Biotechnol., 27, 221-228.
Ennis, B.M., N.A. Gutierrez, I.S. Maddox (1986a). The actone-butanol-ethanol fermentation: a current assessment. Process Biochem., 21, 131-147.
Ennis, B., C.T. Marshall, I.S. Maddox, A.H.J. Paterson (1986b). Continuous product recovery by in-situ gas stripping/condensation during solvent production from whey permeate using Clostridium acetobutylicum. Biotechnol. Lett., 8, 725-730.
Ennis, B.M., N. Qureshi and I.S. Maddox (1987). In-line toxic product removal during solvent production by continuous fermentation using immobilized Clostridium acetobutylicum. Enzyme Microbial. Technol., 9, 672-675.
Evans, P.J. and H.Y. Wang (1988). Response of Clostridum acetobutylicum to the presence of mixed extractants. Appl. Environ. Microbiol., 54, 175-192.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2003). Production of butanol by Clostridium beijerinckii BA101 and in-situ recovery by gas stripping. J. Microbiol. Biotechnol., 19, 595-603.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004a). Butanol fermentation research: upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004b). Acetone-butanol-ethanol production from concentrated substrate: reduction in substrate inhibition by fed-batch technique and product inhibition by gas stripping. Appl. Microbiol. Biotechnol., 63, 653-658.

63
Ezeji, T.C., P.M. Karcher, N. Qureshi, H.P. Blaschek (2005a). Improving performance of a gas stripping-based recovery system to remove butanol from Clostridium beijerinckii fermentation. Bioprocess Biosyst. Eng., 27, 207-214.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2005b). Process for continuous solvent production. United States Patent Application Publication, US patent 20050089979A1.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007a). Bioproduction of butanol from biomass: from genes to bioreactors. Current Opinion in Biotechnol., 18, 220-227.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007b). Butanol production from agricultural residues: impact of degradation products on Clostridium beijerinckii growth and butanol fermentation. Biotechnol. Bioeng., 97, 1460-1469.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007c). Production of acetone butanol (AB) from liquefied corn starch, a commercial substrate, using Clostridium beijerinckii coupled with product recovery by gas stripping. J Ind. Microbiol. Biotechnol., 34, 771-777.
Ezeji, T.C. and H.P. Blaschek (2008). Fermentation of dried distillers’ grains and solubles (DDGS) hydrolysates to solvents and value-added products by solventogenic clostridia. Bioresour. Technol., 99, 5232-5242.
Ezeji, T.C., C. Milne, N.D. Price, H.P. Blaschek (2010). Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol., 85, 1697-1712.
Fadeev, A.G., M.M. Meagher, S.S. Kelley, V.V. Volkov (2000). Fouling of poly-[1-(trimethylsilyl)-1-propyne] membranes in pervaporative recovery of butanol from aqueous solutions and ABE fermentation broth. J. Membr. Sci., 173, 133-144.
Fadeev, A.G. and M.M. Meagher (2001). Opportunities for ionic liquids in recovery of biofuels. Chem. Commun., 295-296.
Fadeev, A.G., YaA Selinskaya, S.S. Kelley, M.M. Meagher, E.G. Litvinova, V.S. Khotimsky, V.V. Volkov (2001). Extraction of butanol from aqueous solutions by pervaporation through poly(1-trimethylsilyl-1-propyne). J. Membr. Sci., 186,205-217.

64
Fadeev, A.G., S.S. Kelley, J.D. McMillan, YaA Selinskaya, V.S. Khotimsky, V.V. Volkov (2003). Effect of yeast fermentation by-products on poly(1-trimethylsilyl-1-propyne) pervaporative performance. J. Membr. Sci., 214,229-238.
Felipe, M.G.A., M.V. Veira, M. Vitolo, I.M. Mancilha, I.C. Roberto and S.S. Silva (1995). Effect of acetic acid on xylose fermentation to xylitol by Candida guilliermondii. J. Basic Microb., 35, 171-177.
Formanek, J., R. Mackie, H.P. Blaschek (1997). Enhanced butanol production by Clostridium beijerinckii BA101 grown in semidefined P2 medium containing 6 percent maltodextrin or glucose. Appl. Environ. Microbiol., 63, 2306-2310.
Frick, C. and K. Schugerl (1986). Continuous acetone-butanol production with free and immobilized Clostridium acetobutylicum. Applied Microbiol. Biotechonol., 25, 186-193.
Gabriel, C.L. (1928). Butanol fermentation process. Ind. Eng. Chem., 20, 1063-1067.
Gabriel, C.L. and F.M. Crawford (1930). Development of the butyl-acetonic fermentation industry. Ind. Eng. Chem., 22, 1163-1165.
Gapes, J.R., D. Nimcevic, and A. Friedl (1996). Long-term continuous cultivation of Clostridium beijerinckii in a two-stage chemostat with on-line solvent removal. Appl. Environ. Microbiol., 62, 3210-3219.
Garćia, V., J. Päkkilä, H. Ojamo, E. Muurinen, R.L. Keiski (2011). Challenges in biobutanol production: How to improve the efficiency. Renew. Sustain. Ener. Reviews, 15, 964-980.
Geng, Q and C-H Park (1994). Pervaporative butanol fermentation by Clostridium acetobutylicum B18. Biotech. Bioeng., 43, 978-986.
George, H.A., J.L. Johnson, W.E.C. Moore, L.V. Holdeman, J.S. Chen (1983). Acetone, isopropanol, and butanol production by Clostridium beijerinckii (syn. Clostridium butylicum) and Clostridium aurantibutyricum. Appl. Environ Microbiol., 45, 1160-1163.

65
Gottschalk, J.C. and J.G. Morris (1981). The induction of acetone and butanol production in cultures Clostridium acetobutylicum by elevated concentrations of acetate and butyrate. FEMS Microb. Letters, 12, 385-389.
Groot, W.J. , C.E. Van der Oever and N.W.F. Kossen (1984). Pervaporation for simultaneous product recovery in the butanol/isopropanol batch fermentation. Biotechnol. Lett., 6, 709-714.
Groot, W.J. and K. Ch. A. M. Luyben (1986). In situ product recovery by adsorption in the butanol/isopropanol batch fermentation. Appl. Microbiol. Biotechnol., 25, 29-31.
Groot, W.J., R.G.J.M. van der Lans, and K. Ch. A.M. Luyben (1989). Batch and continuous butanol fermentations with free cells: integration with product recovery by gas-stripping. Appl. Microbiol. Biotechnol., 32, 305-308.
Groot, W.J., H.S. Soedjak, P.B. Donck, R.G.J.M. van der Lans, K. Ch. A.M. Luyben, J.M.K. Timmer (1990). Butanol recovery from fermentations by liquid-liquid extraction and membrane solvent extraction. Bioprocess Eng., 5, 203-216.
Groot, W.J., R.G.J.M. van der Lans, and K. Ch. A.M. Luyben (1992). Technologies for butanol recovery integrated with fermentations. Proc. Biochem., 27, 61-65.
Gutierrez, N.A., I.S. Maddox, K.C. Schuster, H. Swoboda, J.R. Gapes (1998). Strain comparison and medium preparation for the acetone-butanol-ethanol (ABE) ferment process using substrate potato. Bioresour. Technol., 66, 263-265.
Hagiwara, R. and Y. Ito (2000). Room temperature ionic liquids of alkylimidazolium cations and fluoroanions. J. Fluor. Chem., 105, 221-227.
Hahn-hagerdal, B., M. Galbe, M.F. Gorwa-Grauslund, G. Liden and G. Zacchi (2006). Bio-ethanol—the fuel of tomorrow from the residues of today. TRENDS in Biotechnol., 12, 549-556.
Harris, L.M., N.E. Welker, E.T. Papousakis (2002). Northern, morphological, and fermentation analysis of spo0A inactivation and over-expression in Clostridium acetobutylicum ATCC824. J. Bacteriol., 184, 3586-3597.
Hendriks A.T.W.M. and G. Zeeman (2009). Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresour. Technol., 100, 10-18.

66
Hickey, P.J., F.P. Juricic and C.S. Slater (1992). The effect of process parameters on the pervaporation of alcohols through organophilic membranes. Separation Sci. Technol., 27, 843-861.
Hipolito, C.N., E. Crabbe, C.M. Badillo, O.C. Zarrabal, M.A. M. Mora, G.P. Flores, M.A.H. Cortazar, A. Ishizaki (2008). Bioconversion of industrial wastewater from palm oil processing to butanol by Clostridium saccharoperbutylacetonicum N1-4 (ATCC 13564). J. Cleaner Prod., 16, 632-638.
Holtzapple, M.T. and R.F. Brown (1994). Conceptual design for a process to recover volatile solutes from aqueous solutions using silicalite. Sep. Technol., 4, 213-229.
Howard, R.L., E. Abotsi, E.L. Jansen van Rensburg and S. Howard (2003). Lignocellulosic biotechnology: issues of bioconversion and enzyme production. African J. Biotechnol., 2(12), 602-619.
Huang J. and M.M. Meagher (2001). Pervaporative recovery of n-butanol from aqueous solutions and ABE fermentation broth using thin-film silicalite-filled silicone composite membranes. J. Membr. Sci., 192, 231-242.
Huang, Y.L., Z. Wu, L. Zhang, C.M. Cheung and S.T. Yang (2002). Production of carboxylic acids from hydrolyzed corn meal by immobilized cell fermentation in a fibrous-bed bioreactor. Bioresource Technol., 82, 51-59.
Huang, W.C., D.E. Ramey, and S.T. Yang (2004). Continuous production of butanol by Clostridium acetobutylicum immobilized in a fibrous bed bioreactor. Appl. Biochem. Biotechnol., 113-116, 887-898.
Huddleston, J.G., H.D. Willauer, R.P. Swatloski, A.E. Visser, R.D. Rogers (1998). Room temperature ionic liquids as novel media for “clean” liquid-liquid extraction. Chem Commun., 1765-1766.
Inui, M., M. Suda, S. Kimura, K. Yasuda, H. Suzuki, H. Toda, S. Yamamoto, S. Okino, N. Suzuki, H. Yukawa (2008). Expression of Clostridium acetobutylicum butanol synthetic genes in Escherichia coli. Appl. Microbiol. Biotechnol., 77, 1305-1316.
Izak, P., K. Schwarz, W. Ruth, H. Bahl, U. Kragl (2008). Increased productivity of Clostridium acetobutylicum fermentation of acetone, butanol, and ethanol by pervaporation through supported ionic liquid membrane. Appl. Microbiol. Biotechnol., 78, 597-602.

67
Jain, M.K., D. Beacom, and R. Datta (1993). Mutant strain of C. acetobutylicum and process for making butanol. United States Patent, US Patent 5192673.
Jesse, T.W., T.C. Ezeji, N. Qureshi, H.P. Blaschek (2002). Production of butanol from starch-based waste packing peanuts and agricultural waste. J. Ind. Microbiol. Biotechnol., 29, 117-123.
Jones, D.T. and D. Woods (1986). Acetone-butanol fermentation revisted. Microbiol. Reviews, 50, 484-524.
Jonquieres, A. and A. Fane (1997). Filled and unfilled composite GFT PDMS membranes for the recovery of butanol from dilute aqueous solutions: influence of alcohol polarity. J. Membr. Sci., 125, 245-255.
Jonquieres, A., R. Clement, P. Lochon, J. Neel, M. Dresch, B. Chertien (2002). Industrial state-of-art of pervaporation and vapour permeation in the western countries. J. Membr. Sci., 206, 87-117.
Jorgensen, H., J. B. Kristensen and C. Felby (2007). Enzymatic conversion of lignocellulose into fermentable sugars: Challenges and opportunities. Biofuels. Bioprod. Bioref., 1, 119–134.
Kim S.Y., D.K. Oh and J.H. Kim (1999). Evaluation of xylitol production from corn cob hemicellulose hydrolysate by Candida parapsilosis. Biotechonol. Lett., 21, 891-895.
Klinke, H.B., A.B. Thomsen and B.K. Ahring (2004). Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pre-treatment of biomass. Appl. Microbiol. Technol., 66, 10-26.
Knoshaug, E.P. and M. Zhang (2009). Butanol tolerance in a selection of microorganisms. Appl. Biochem. Biotechnol., 153, 13-20.
Kumar, P., D.M. Barrett, M.J. Delwiche and P. Stroeve (2009). Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem., 48, 3713-3729.
Kumar, M. and K. Gayen (2011). Developments in biobutanol production: New insights. Appl. Ener., 88, 1999-2012.
Larrayzo M.A. and L. Puigjaner (1987). Study of butanol extraction through

68
pervaporation in acetobutylic fermentation. Biotechnol. Bioeng., 30, 692-696.
Lee, S.T., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentive butanol production by Clostridia. Biotechnol. Bioeng., 101,209-228.
Li, S., V.A. Tuan, J.L. Falconer, R.D. Noble (2003). Properties and separation performance of Ge-ZSM-5 membranes. Micropor. Mesopor. Mater., 58, 137-154.
Li, S-Y, R. Srivastava and R.S. Parnas (2010). Separation of 1-butanol by pervaporation using novel tri-layer PDMS composite membrane. J. Membr. Sci., 363, 287-294.
Lin, Y.L. and H.P. Blaschek (1983). Butanol production by a butanol-tolerant strain of Clostridium acetobutylicum in extruded corn broth. Appl. Environ. Microbiol., 45, 966-973.
Liou, J.S.C., D.L. Balkwill, G.R. Drake, R.S. Tanner (2005). Clostridium carboxidivorans sp. nov., a solvent-producing Clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen Clostridium scatologenes strain SL1 as Clostridium drakei sp. nov. Int. J. Syst. Evol. Microbiol., 55, 2085-2091.
Liu, Q., M.H.A. Janssen, F. van Rantwijk, R.A. Sheldon (2005). Room temperature ionic liquids that dissolve carbohydrates in high concentrations. Green Chemistry, 7, 39-42.
Liu, Z., Y.Ying, F. Li, C. Ma, P. Xu (2010). Butanol production by Clostridium beijerinckii ATCC 55025 from wheat bran. J. Ind. Microbiol. Biotechnol., 37, 495-501.
Lu, X.B., Y.M. Zhang, J. Yang and Y. Liang (2007). Enzymatic hydrolysis of corn stover after pretreatment with dilute sulfuric acid. Chem. Eng. Technol., 30, 938-944.
MacDonald, D. G.; N.N. Bakhshi, J.F. Mathews, A. Roychowdhury, P. Bajpai and M. Moo-Young (1983). Alkali treatment of corn stover to improve sugar production by enzymatic hydrolysis. Biotechnol. Bioeng., 25, 2067–2076.
Maddox, I.S. (1982). Use of silicalite for the adsorption of n-butanol from fermentation liquors. Biotechnol. Lett., 4, 759-760.

69
Maddox, I.S.(1989). The actone-butanol-ethanol fermentation: recent progress in technology. Genet. Eng. Rev., 7, 189-220.
Maddox, I.S., N. Qureshi and K. Roberts-Thomson (1995). Production of acetone-butanol-ethanol from concentrated substrates using Clostridium acetobutylicum in an integrated fermentation-product removal process. Process Biochemistry, 30, 209-215.
Madihah, M.S., A.B. Ariff, K.M. Sahaid, A.A. Suraini, M.I.A. Karim (2001). Direct fermentation of gelatinized sago starch to acetone-butanol-ethanol by Clostridium acetobutylicum. World J. Microbiol. Biotechnol., 17, 567-576.
Malherbe, S. and T.E. Cloete (2003). Lignocellulose biodegradation: fundamentals and applications: A review. Environ. Sci. Biotechnol., 1, 105-114.
Martinez, A., M.E. Rodriguez, M.L. Wells, S.W. York, J.F. Preston and L.O. Ingram (2001). Detoxification of dilute acid hydrolysates of lignocellulose with lime. Biotechnol. Prog., 17, 287-293.
Matsumoto, M., K. Mochiduki, K. Fukunishi, K. Kondo (2004). Extraction of organic acids using imidazolium-based ionic liquids and their toxicity to Lactobacillus rhamnosus. Separ. Purif. Technol., 40, 97-101.
Matsumura, M. and H. Kataoka (1987). Separation of dilute aqueous butanol and acetone solutions by pervaporation through liquid membranes. Biotechnol. Bioeng., 30, 1991-1992.
McCoy, E., E.B. Fred, W.H. Peterson, E.G. Hastings (1926). A cultural study of the acetone butyl alcohol organisms. J. Infect. Dis., 39, 457-483.
Meagher, M.M., N. Qureshi and R.W. Hutkins (1998). Silicalite membrane and method for the selective recovery and concentration of acetone and butanol from model ABE solutions and fermentation broth. U.S. Patent 5,755,967.
Mermelstein, L.D., E.T. Papoutsakis, D.J. Petersen, G.N. Bennett (1993). Metabolic engineering of Clostridium acetobutylicum ATCC 824 for increase solvent production by enhancement of acetone formation enzyme-activities using a synthetic acetone operon. Biotechnol. Bioeng., 42, 1053-1060.

70
Mes-Hartree, M., B. E. Dale and W.K. Craig (1988). Comparison of steam and ammonia pretreatment for enzymatic hydrolysis of cellulose. Appl. Microbiol. Biotechnol., 29, 462–468.
Milestone, N.B. and D.M. Bibby (1981). Concentration of alcohols by adsorption on silicalite. J. Chem. Technol. Biotechnol., 31, 732-736.
Mitchell, W.J. (1996). Carbohydrate uptake and utilization by Clostridium beijerinckii NCIMB 8052. Anaerobe, 2, 379-384.
Mosier, N.S., C. Wyman, B.Dale, R.Elander, Y.Y. Lee, M. Holtzapple and M.R. Ladisch (2005). Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96, 673-668.
Mussatto, S.I. and I.C. Roberto (2001). Hydrolysate detoxification with activated charcoal for xylitol production by Candida guilliermondii. Biotechnol. Lett., 23, 1681-1684.
Mussatto, S.I. and I.C. Roberto (2004a). Alternatives for detoxification of diluted-acid lignocellulosic hydrolysates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Mussatto, S.I. and I.C. Roberto (2004b). Optimal experimental condition for hemicellulosic hydrolysate treatment with activated charcoal for xylitol production. Biotechnol. Prog., 20, 134-139.
Mussatto, S.I., J.C. Santos and I.C. Roberto (2004). Effect of pH and activated charcoal adsorption on hemicellulosic hydrolysate detoxification for xylitol production. J. Chem. Technol. Biotechnol., 79, 590-596.
Nair, R.V., E.M. Green, D.E. Watson, G.N. Bennett, E.T. Papouskis (1999). Regulation of the sol locus genes for butanol and acetone formation in Clostridium acetobutylicum ATCC824 by a putative transcriptional regressor. J. Bacteriol., 181, 319-330.
Napoli, F., G. Olivieri, M.E. Russo, A. Marzocchella, P. Salatino (2010). Butanol production by Clostridium acetobutylicum in a continuous packed bed reactor. J. Ind. Microbiol. Biotechnol., 37, 603-608.

71
Ni, Y. and Z. Sun (2009). Recent progress on industrial fermentative production of acetone-butanol-ethanol by Clostridium acetobutylicum in China. Appl. Microbiol. Biotechnol., 83, 415-423.
Nielsen, L., M. Larsson, O. Holst, B. Mattiasson (1988). Adsorbents for extractive bioconversion applied to the acetone-butanol fermentation. Appl. Microbiol. Biotechnol., 28, 335-339.
Nielsen, D.R. and K.J. Prather (2009). In situ product recovery of n-butanol using polymeric resins. Biotechnol. Bioeng., 102,811-821.
Nielsen, D.R., E. Leonard, S.H. Yoon, H.C. Tseng, C. Yuan, K.L.J. Parther (2009). Engineering alternative butanol production platforms in heterologous bacteria. Metab. Eng., 11, 262-273.
Nigam, P.S. and A. Singh (2011). Production of liquid biofuels from renewable resources. Prog. Ener. Combust. Sci., 37, 52-68.
Nilvebrant, N.O., A. Reimann, S. Larsson and L.J. Jonsson (2001). Detoxification of lignocellulose hydrolysates with ion exchange resins. Appl. Biochem. Biotechnol., 91, 35-49.
Nuttha, T., S. Navankasattusas and S.T. Yang (2009). Production of lactic acid and ethanol by Rhizopus oryzae integrated with cassava pulp hydrolysis. Bioprocess. Biosyst. Eng., DOI 10.1007/s00449-009-0341-x.
Olsson, L and B. Hahn-Hagerdal (1996). Fermentation of lignocellulosic hydrolysates for ethanol production. Enzyme Microb. Technol., 18, 312-331.
Oudshoorn, A., L.A.M. van der Wielen, and A.J.J. Straathof (2009). Adsorption equilibria of bio-based butanol solutions using zeolite. Biochem. Eng. J., 48, 99-103.
Palmarola-Adrados, B., M. Galbe and G. Zacchi (2004). Combined steam pretreatment and enzymatic hydrolysis of starch-free wheat fibers. Appl. Biochem. Biotechnol., 113, 989-1002.
Palmqvist, E., H. Grage, N.Q. Meinander and B. Hahn-Hagerdal (1999). Main and interaction effects of acetic acid, furfural and p-hydroxybenzoic acid on growth and ethanol productivity of yeasts. Biotechnol. Bioeng., 63, 46-55.

72
Palmqvist, E. and B. Hahn-Hagerdal (2000a). Fermentation of lignocellulosic hydrolysates. I: inhibition and detoxification. Bioresour. Technol., 74, 17-24.
Palmqvis, E. and B. Hahn-Hagerdal (2000b). Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol., 74, 25-33.
Pandey, A., C.R. Soccol, P. Nigam and V.T. Soccol (2000a). Biotechnological potential of agro-industrial residues. I: sugarcane bagasse. Bioresour. Technol., 74, 69-80.
Pandey, A., C.R. Soccol, P. Nigam, V.T. Soccol, L.P.S. Vandenberghe and R. Mohan (2000b). Biotechnological potential of agro-industrial residues. II: cassava bagasse. Bioresour. Technol., 74, 81-87.
Park, C., M. Okos and P. Wankat (1989). Acetone-butanol-ethanol (ABE) fermentation in an immobilized cell tickle bed reactor. Biotechnol. Bioeng., 34, 18-29.
Parekh, M., J. Formanek and H.P. Blaschek (1998). Development of a cost-effective glucose-corn steep medium for production of butanol by Clostridium beijerinckii. J. Industrial Microbiol. Biotechnol., 21, 187-191.
Parekh, M., J. Formanek, and H.P. Blaschek (1999). Pilot-scale production of butanol by Clostridium beijerinckii BA 101 using a low-cost fermentation medium used on corn steep water. Appl. Microbiol. Biotechnol., 51, 152-157.
Perez, J., J.M. Dorado, T.D. Rubia and J. Martinez (2002). Biodegradation and biological treatment of cellulose, hemicellulose and lignin: An overview. Int. Microbiol., 5, 53-63.
Pierrot, P., M. Fick, and J.M. Engasser (1986). Continuous acetone-butanol fermentation with high productivity by cell ultrafiltration and recycling. Biotechnol. Lett., 8, 253-256.
Qureshi, N., I.S. Maddox, and A. Friedl (1992). Application of continuous substrate feeding to the ABE fermentation: relief of product inhibition using extraction, perstraction, stripping and pervaporation. Biotechnol. Prog., 8, 382-390.
Qureshi, N. and H.P. Blaschek (1999a). Butanol recovery from model solution/ fermentation broth by pervaporation: evaluation of membranes performance. Biomass and Bioenergy, 17, 175-184.

73
Qureshi, N. and H.P. Blaschek (1999b). Production of acetone butanol ethanol (ABE) by a hyper-producing mutant strain of Clostridium beijerinckii BA101 and recovery by pervaporation. Biotechnol. Prog., 15, 594-602.
Qureshi, N. and H.P. Blaschek (1999c). Fouling studies of a pervaporation membrane with commercial fermentation media and fermentation broth of hydrper-butanol-producing Clostridium beijerinckii BA101. Sep. Sci. Technol., 34, 2803-2815.
Qureshi N. and H.P. Blaschek (2000). Butanol production using Clostridium beijerinckii BA101 hyper-butanol producing mutant strain and recovery by pervaporation. Appl. Biochem. Biotechnol., 84, 225-235.
Qureshi, N. and H.P. Blaschek (2001a). Recovery of butanol from fermentation broth by gas stripping. Renewable Energy, 22, 557-564.
Qureshi, N. and H.P. Blaschek (2001b). Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 27, 287-291.
Qureshi, N. and I.S. Maddox (1988). Reactor Design for the ABE fermentation using cells of Clostridium acetobutylicum immobilized by adsorption onto bonechar. Bioprocess Eng., 3, 69-72.
Qureshi, N. and I.S. Maddox (1991). Integration of continuous production and recovery of solvents from whey permeate: use of immobilized cells of Clostridium acetobutylicum in a fluidized bed reactor coupled with gas stripping. Bioproc. Eng., 6, 63-69.
Qureshi, N. and I.S. Maddox (1995). Continuous production of acetone-butanol- ethanol using immobilized cells of Clostridium acetobutylicum and integration with product removal by liquid-liquid extraction. J. Ferment. Bioeng., 80, 185-189.
Qureshi, N. and I.S. Maddox (2005). Reduction in butanol inhibition by perstraction: Utilization of concentrated lactose/whey permeate by Clostridium acetobutylicum to enhance butanol fermentation economics. Trans IChemE, Part C, Food and Bioproducts Processing, 83, 43-52.

74
Qureshi, N., M.M. Meagher and R.W. Hutkins (1999). Recovery of butanol from model solutions and fermentation broth using silicalite/silicone membrane. J. Membr. Sci., 158, 115-125.
Qureshi, N., J. Schripsema, J. Lienhardt and H.P. Blaschek (2000). Continuous solvent production by Clostridium beijerinckii BA 101 immobilized by adsorption onto brick. J. Microbiol. Biotechnol., 16, 377-382.
Qureshi, N., M.M. Meagher, Jicai Huang, R.W. Hutkins (2001a). Acetone butanol ethanol (ABE) recovery by pervaporation using silicate-silicone composite membrane from fed-batch reactor of Clostridium acetobutylicum. J. Membrane Sci., 187, 93-102.
Qureshi, N., A. Lolas and H.P. Blaschek (2001b). Soy molasses as fermentation substrate for production of butanol using Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 26, 290-295.
Qureshi, N., L.L. Lai, and H.P. Blaschek (2004). Scale-up of a high productivity continuous biofilm reactor to produce butanol by adsorbed cells of Clostridium beijerinckii. Food Bioprod. Process, 82, 164-173.
Qureshi, N., S. Hughes, I.S. Maddox, M.A. Cotta (2005). Energy-efficient recovery of butanol from model solutions and fermentation broth by adsorption. Bioprocess Biosyst. Eng., 27, 215-222.
Qureshi, N., X. Li, S. Hughes, B.C. Saha and M.A. Cotta (2006). Butanol production from corn fiber xylan using Clostridim acetobutylicum. Biotechnol. Prog., 22, 673-680.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst. Eng., 30, 419-427.
Qureshi, N. and T.C. Ezeji (2008). Butanol, ‘a superior biofuel’ production from agricultural residues (renewable biomass): recent progress in technology. Biofuels, Bioprod. Bioref., 2, 319-330.
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta and H.P. Blaschek (2008a). Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.

75
Qureshi, N., B.C. Saha, R.E. Hector, M.A. Cotta (2008b). Removal of fermentation inhibitors from alkaline peroxide pretreated and enzymatically hydrolyzed wheat straw: production of butanol from hydrolysate using Clostridium beijerinckii in batch reactors. Biomass Bioenergy, 32, 1353-1358.
Qureshi, N., B.C. Saha, R. E. Hector, S.R. Hughes, M.A. Cotta (2008c). Butanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part I- Batch fermentation. Biomass Bioener., 32, 168-175.
Qureshi, N., B.C. Saha and M.A. Cotta (2008d). Butanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part II- Fed-batch fermentation. Biomass Bioener., 32, 176-183.
Qureshi, N., B.C. Saha, B. Dien, R.E. Hector, M.A. Cotta (2010a). Production of butanol (a biofuel) from agricultural residues: Part I-Use of barley straw hydrolysate. Biomass Bioenergy, 34, 559-565.
Qureshi, N., B.C. Saha, R.E. Hector, B. Dien, S. Hughes, S. Liu, L. Iten, M.J. Bowman, G. Sarath, M.A. Cotta (2010b). Production of butanol (a biofuel) from agricultural residues: Part II-Use of corn stover and switchgrass hydrolysates. Biomass Bioenergy, 34, 566-571.
Rabinovich, M.L., M.S. Melnik and A.V. Bolobova (2002). Microbial cellulases: A review. Appl. Biochem. Microbiol., 38, 305-321.
Rajagopalan, S., R.P. Datar, and R.S. Lewis (2002). Formation of ethanol from carbon monoxide via a new microbial catalyst. Biomass and bioenergy, 23, 487-493.
Reddy, N. and Y. Yang (2005). Biofibers from agricultural byproducts for industrial applications. TRENDS in Biotechnol., 23, 22-27.
Roberto, I.C., M.G.A. Felipe, I.M. Mancilha, M. Vitola, S. Sato and S.S. Silva (1995). Xylitol production by Candida guilliermondii as an approach for the utilization of agroindustrial residues. Bioresour. Technol., 51, 255-257.
Roffler, S.R., H.W. Blanch, and C.R. Wilke (1987a). In-situ recovery of butanol during fermentation, part 1: batch extractive fermentation. Bioprocess Eng., 2, 1-12.

76
Roffler, S.R., H.W. Blanch, and C.R. Wilke (1987b). In-situ recovery of butanol during fermentation, part 2: fed-batch extractive fermentation. Bioprocess Eng., 2, 181-190.
Roffler, S.R., H.W. Blanch, C.R. Wilke (1988). In situ extractive fermentation of acetone and butanol. Biotechnol. Bioeng., 31, 135-143.
Saha, B.C. (2003). Hemicellulose bioconversion. J Ind Microbiol Biotechnol., 30, 279-291.
Sano, T., H. Yanagishita, Y. Kiyouzumi, F. Mizukami, H. Haraya (1994). Sepation of ethanol/water mixture by silicalite membrane on pervaporation. J. Membr. Sci., 95, 221-228.
Schlote, D. and G. Gottschalk (1986). Effect of cell recycle on continuous butanol-acetone fermentation with Clostridium acetobutylicum under phosphate limitation. Appl. Microbiol. Biotechnol., 24, 1-5.
Schmidt, S.L., M.D. Myers, S.S. Kelley, J.D. McMillan, N. Padukone (1997). Evaluation of PTMSP membranes in achieving enhanced ethanol removal from fermentations by pervaporation. Appl. Biochem. Biotechnol., 63-65, 469-482.
Seddon, K.R. (1997). Review: Ionic liquids for clean technology. J. Chem. Tech. Biotechnol., 68, 351-356.
Servinsky, M.D., J.T. Kiel, N.F. Dupuy, C.J. Sund (2010). Transcriptional analysis of different carbohydrate utilization by Clostridium acetobutylicum. Microbiology, 156, 3478-3491.
Shen, C.R. and J.C. Liao (2008). Metabolic engineering of Escherichia coli for 1-butanol and 1-propanol production via keto-acid pathways. Metab. Eng., 10, 312-320.
Silva, E.M. and S.T. Yang (1995). Kinetics and stability of a fibrous-bed bioreactor for continuous production of lactic from unsupplemented acid whey. J. Biotechnol., 41, 59-70.
Somrutai, W., M. Takagi and T. Yoshida (1996). Acetone-butanol fermentation by Clostridium aurantibutyricum ATCC 17777 from a model medium for palm oil mill effluent. J. Ferment. Bioeng., 81, 543-547.

77
Soni, B.K., P. Soucaille and G.Goma (1987). Continuous acetone-butanol fermentation: a global approach for the improvement in the solvent productivity in synthetic medium. Appl. Microbiol. Biotechnol., 25, 317-321.
Sriroth, K., R. Chollakup, S. Chotineeranat, K. Piyachomkwan and C.G. Oates (2000). Processing of cassava waste for improved biomass utilization. Bioresour. Technol., 71, 63-69.
Sun, Y. and J. Cheng (2002). Hydrolysis of lignocellulosic material from ethanol production: A review. Bioresour. Technol., 83, 1-11.
Swana, J., Y. Yang, M. Behnam, R. Thompson (2011). An analysis of net energy production and feedstock availability for biobutanol and bioethanol. Bioresour. Technol., 102, 2112-2117.
Teymouri, F., L.L. Perez, H. Alizadeh and B.E. Dale (2004). Ammonia fiber explosion treatment of corn stover. Appl. Biochem. Biotechnol., 113, 951–963.
Taherzadeh, M.J. and K. Karimi (2007). Enzyme-based hydrolysis processes for ethanol from lignocellulosic materials: a review. Bioresour., 2, 707-738.
Thang, V.H., K. Kanda, and G. Kobayashi (2010). Production of acetone-butanol- ethanol (ABE) in direct fermentation of cassava by Clostridium saccharoperbutylacetonicum N1-4. Appl. Biochem. Biotechnol., 161, 157-170.
Thongsukmak, A. and K.K. Sirkar (2007). Pervaporation membranes highly selective for solvents present in fermentation broth. J. Memb.Sci., 302, 45-48.
Thormann, K. and P. Dürre (2001). Orf5/SolR: a transcriptional repressor of the sol operon of Clostridium acetobutylicum? J. Ind. Microbiol. Biotechnol., 27, 307-313.
Thormann, K., L. Feustel, K. Lorenz, S. Nakotte, P. Dürre (2002). Control of butanol formation in Clostridium acetobutylicum by transcritional activation. J. Bacteriol., 184, 1966-1973.
Toh, S.L.I., J. McFarlane, C. Tsouris, D.W. DePaoli, H. Luo, S. Dai (2006). Room-temperature ionic liquids in liquid-liquid extraction: effects of solubility in aqueous solutions on surface properties. Solvent Extraction and Ion Exchange, 24, 33-56.

78
Vane, L.M. (2005). Review: A review of Pervaporation for product recovery from biomass fermentation processes. J. Chem. Technol. Biotechnol., 80, 603-629.
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuls, Bioprod. Bioref., 2, 553-588.
Varga, E., K. Reczey and G. Zacchi (2004). Optimization of steam pretreatment of corn stover to enhance enzymatic digestibility. Appl. Biochem. Biotechnol., 113, 509-523.
Volkov, V.V., V.S. Khotimsky, E.G. Litvinova, A.G. Fadeev, YaA Selinskaya, N.A. Plate, J.D. McMillan, S.S. Kelley (1997). Development of novel poly[-1-(trimethylsilyl)-1-propyne] based materials for pervaporation separation in biofuel production. Polym. Mater. Sci. Eng., 77, 339-340.
Volkov, V.V., A.G. Fadeev, V.S. Khotimsky, E.G. Litvinova, YaA Selinskaya, J.D. McMillan, S.S. Kelley (2004). Effects of synthesis conditions on the pervaporation properties of poly[-1-(trimethylsilyl)-1-propyne] useful for membrane bioreactors. J. Appl. Polym. Sci., 91, 2271-2277.
Vrana, D.L., M.M. Meagher, R.W. Hutkins, B. Duffield (1993). Pervaporation of model acetone-butanol-ethanol fermentation product solutions using Polytetrafluoroethylene membranes. Sep. Sci. Technol., 28, 2167-2178.
Wang, B., T. Ezeji, Z. Shi, H. Feng, H.P. Blaschek (2009). Pretreatment and conversion of distiller’s dried grains with solubles for acetone-butanol-ethanol (ABE) production. Transactions of ASABE, 52, 885-892.
Wayman, M. and R. Parekh (1987). Production of acetone-butanol by extractive fermentation using dibutylphthalate as extractant. J. Ferment. Technol., 65, 295-300.
Weber, C., A. Farwick, F. Benisch, D. Brat, H. Dietz, T. Subtil, E. Boles (2010). Trends and challenges in the microbial production of lignocellulosic bioalcohol fuels. Appl. Microbiol. Biotecnol., 87, 1303-1315.
Wei, G.T., Z. Yang and C.J. Chen (2003). Room temperature ionic liquid as a novel medium for liquid/liquid extraction of metal ions. Analytica Chimica Acta, 488, 183-192.
Woods, D.R. (1995). The genetic engineering of microbial solvent production. Trends Biotechnol., 13, 259-264.

79
Wu, Z. and S.T. Yang (2003). Extractive fermentation for butyric acid production from glucose by Clostridium tyrobutyricum. Biotechnol. Bioeng., 82, 93-102.
Yang, X., G.J. Tsai and G.T. Tsao (1994). Enhancement of in situ adsorption on the acetone-butanol fermentation by Clostridium acetobutylicum. Separation Techonol., 4, 81-92.
Yang, X. and G.T. Tsao (1995). Enhanced acetone-butanol fermentation using repeated fed-batch operation coupled with cell recycle by membrane and simultaneous removal of inhibitory products by adsorption. Biotechnol. Bioeng., 47, 444-450.
Yang, S.T. (1996). Extractive fermentation using convoluted fibrous bed bioreactor. United States Patent, US patent 5563069.
Yat, S.C., A. Berger and D.R. Shonnard (2008). Kinetic characterization of dilute surface acid hydrolysis of timber varieties and switchgrass. Bioresour. Technol., 99, 3855-3863.
Yu, M., Y. Zhang, I. Tang, S.T. Yang (2011). Metabolic engineering of Clostridium tyrobutyricum for n-butanol production. Metab. Eng., in press, doi:10.1016/j.ymben.2011.04.002.
Zautsen, R.R.M., F. Maugeri-Filho, C.E. Vaz-Rossell, A.J.J. Straathof, L.A.M. van der Wielen, J.A.M. de Bont (2009). Liquid-liquid extraction of fermentation inhibiting compounds in lignocellulose hydrolysate. Biotechnol. Bioeng., 102, 1354-1360.
Zhang, L., D. Li, L. Wang, T. Wang, L. Zhang, X.D. Chen and Z. Mao (2008). Effect of steam explosion on biodegradation of lignin in wheat straw. Bioresour. Technol., 99, 8512-8515.
Zhang, Y., Y. Ma, F. Yang, C. Zhang (2009). Continous acetone-butanol-ethanol production by corn stalk immobilized cells. J. Ind. Microbiol. Biotechnol., 36, 1117-1121.
Zhao, H., S. Xia and P. Ma (2005). Review: use of ionic liquids as “green” solvents for extractions. J. Chem. Technol. Biotechnol., 80, 1089-1096.
Zheng, Y.N., L.Z. Li, M. Xian, Y.J. Ma, J.M. Yang, X. Xu, D.Z. He (2009). Problems with the microbial production of butanol. J. Ind. Microbiol. Biotechnol., 36, 1127-1138.

80
Zhu, Y. and S.T. Yang (2003). Adaption of Clostridium tyrobutyricum for enhanced tolerance to butyric acid in a fibrous bed bioreactor. Biotechnol. Progress, 19, 365-372.
Zhu, Y., Z. Wu, and S.T. Yang (2002). Butyric acid production from acid hydrolysate of corn fibre by Clostridium tyrobutyricum in a fibrous bed bioreactor. Process Biochem., 38, 657-666.

81
Fuel properties n-Butanol Ethanol Methanol Gasoline
Energy density (MJ/L) 29.2 19.6 16 32
Heat of vaporization (MJ/kg) 0.43 0.92 1.2 0.36
Research octane number 96 107 106 91-99
Motor octane number 78 89 92 81-89
Air to fuel ratio 11.2 9.0 6.4 14.6
Specific energy (MJ/kg air) 3.2 3.0 3.1 2.9
Table 2.1 Important fuel properties of butanol, ethanol, methanol and gasoline

82
Species / strain Substrates Products pH Temp. (oC) References Glucose, xylose, arabinose, cellobiose, mannose, galactose
Ezeji and Blaschek, 2008
Starch Madihah et al., 2001 Lactose Qureshi and Maddox, 2005
C. acetobutylicum P262 ATCC 824
Sucrose, fructose, lactose, maltose, cellobiose
Acetone, butanol, ethanol, acetate, butyrate, H2, CO2
5.5 – 6.5 35 ± 1
Servinsky et al., 2010
Syngas (H2, CO, CO2) 5.0 - 6.06.2
Bruant et al., 2010 Liou et al., 2005
C. carboxidivorans P7
H2, CO
Acetate, ethanol, butyrate, butanol 5.7-5.8
37 ± 1 Rajagopalan et al., 2002
Glucose, starch, maltose 6.2 Thang et al., 2010 C. saccharoperbutylacetonicum N1-4
Molasses, starch Acetone, butanol, ethanol, acetate, butyrate, H2, CO2
5.6-5.9 30 Hipolito et al., 2008
C. saccharobutylicum 262
Glucose, xylose, arabinose, cellobiose, mannose, galactose
Acetone, butanol, ethanol, acetate, butyrate, H2, CO2
5.5 – 6.5 35 Ezeji and Blaschek, 2008
C. butylicum NRRL 592
Glucose, xylose, arabinose, cellobiose, mannose, galactose
Acetone, butanol, ethanol, acetate, butyrate, H2, CO2
5.5 – 6.5 35 Ezeji and Blaschek, 2008
Glucose, xylose, arabinose, cellobiose, mannose, galactose,
5.5 – 6.5 35 Ezeji and Blaschek, 2008 Ezeji et al., 2007b
Starch 6.8 36 Jesse et al., 2002 Sucrose, fructose 6.8 35 Qureshi et al., 2001b Maltodextrin 6.5 33-35 Formanek et al., 1997
C. beijerinckii BA101 NCIMB 8052
Glucitol (sorbitol), mannitol
Acetone, butanol, ethanol, acetate, butyrate, H2, CO2
N/A 37 Mitchell, 1996
Glucose, , xylan, starch, pectin, arabinose, xylose, galactose, mannose
5.5 – 6.8 37 Somrutai et al., 1996 C. aurantibutyricum
ATCC 17777 NCIB 10659 Glucose
Acetone, butanol, isopropanol, acetate, butyrate 6.8 35 George et al., 1983
5.0 – 7.0 37 Ahn et al., 2011 Glycerol 4.5 – 7.5 35 Biebl, 2001
C. pasteurianum DSM 525
Butanol, ethanol, 1,3-propanediol, acetate, butyrate, lactate 7.0 35 Taconi et al., 2009
Table 2.2 Summary of various solventogenic Clostridia with their substrates, products, fermentation pH and temperature
82
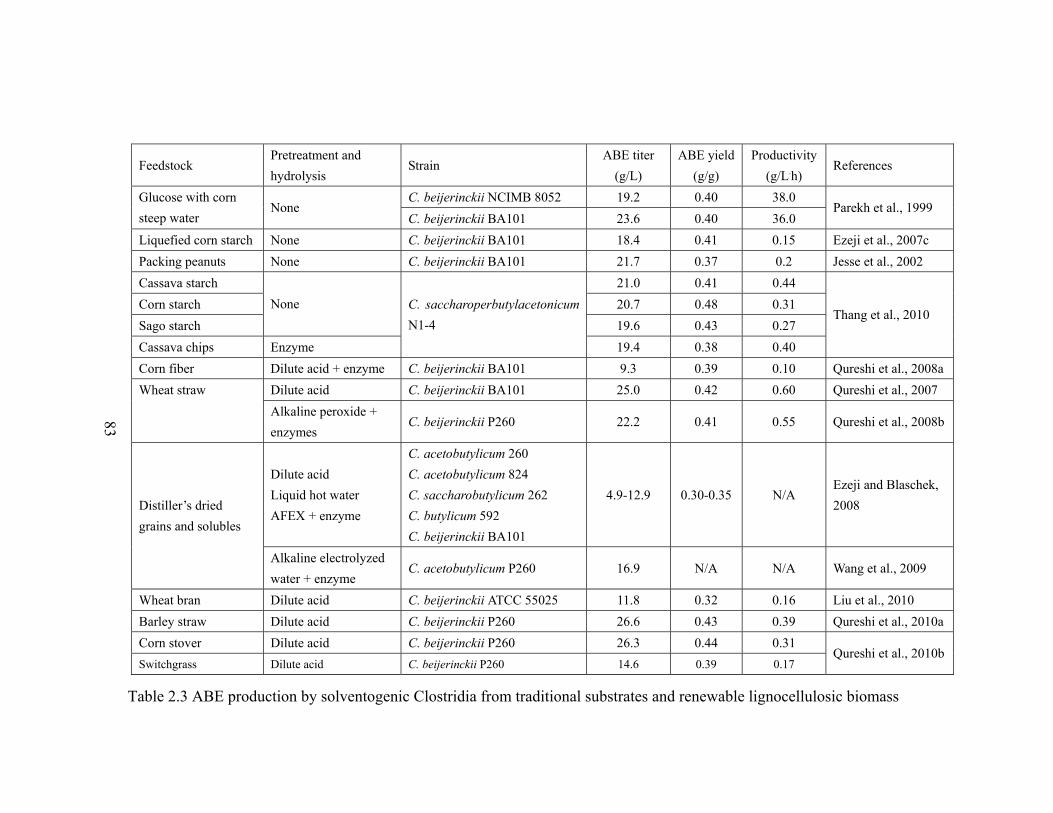
83
Feedstock Pretreatment and
hydrolysis Strain
ABE titer
(g/L)
ABE yield
(g/g)
Productivity
(g/L.h) References
C. beijerinckii NCIMB 8052 19.2 0.40 38.0 Glucose with corn
steep water None
C. beijerinckii BA101 23.6 0.40 36.0 Parekh et al., 1999
Liquefied corn starch None C. beijerinckii BA101 18.4 0.41 0.15 Ezeji et al., 2007c
Packing peanuts None C. beijerinckii BA101 21.7 0.37 0.2 Jesse et al., 2002
Cassava starch 21.0 0.41 0.44
Corn starch 20.7 0.48 0.31
Sago starch
None
19.6 0.43 0.27
Cassava chips Enzyme
C. saccharoperbutylacetonicum
N1-4
19.4 0.38 0.40
Thang et al., 2010
Corn fiber Dilute acid + enzyme C. beijerinckii BA101 9.3 0.39 0.10 Qureshi et al., 2008a
Dilute acid C. beijerinckii BA101 25.0 0.42 0.60 Qureshi et al., 2007 Wheat straw
Alkaline peroxide +
enzymes C. beijerinckii P260 22.2 0.41 0.55 Qureshi et al., 2008b
Dilute acid
Liquid hot water
AFEX + enzyme
C. acetobutylicum 260
C. acetobutylicum 824
C. saccharobutylicum 262
C. butylicum 592
C. beijerinckii BA101
4.9-12.9 0.30-0.35 N/A Ezeji and Blaschek,
2008 Distiller’s dried
grains and solubles
Alkaline electrolyzed
water + enzyme C. acetobutylicum P260 16.9 N/A N/A Wang et al., 2009
Wheat bran Dilute acid C. beijerinckii ATCC 55025 11.8 0.32 0.16 Liu et al., 2010
Barley straw Dilute acid C. beijerinckii P260 26.6 0.43 0.39 Qureshi et al., 2010a
Corn stover Dilute acid C. beijerinckii P260 26.3 0.44 0.31
Switchgrass Dilute acid C. beijerinckii P260 14.6 0.39 0.17 Qureshi et al., 2010b
Table 2.3 ABE production by solventogenic Clostridia from traditional substrates and renewable lignocellulosic biomass
83
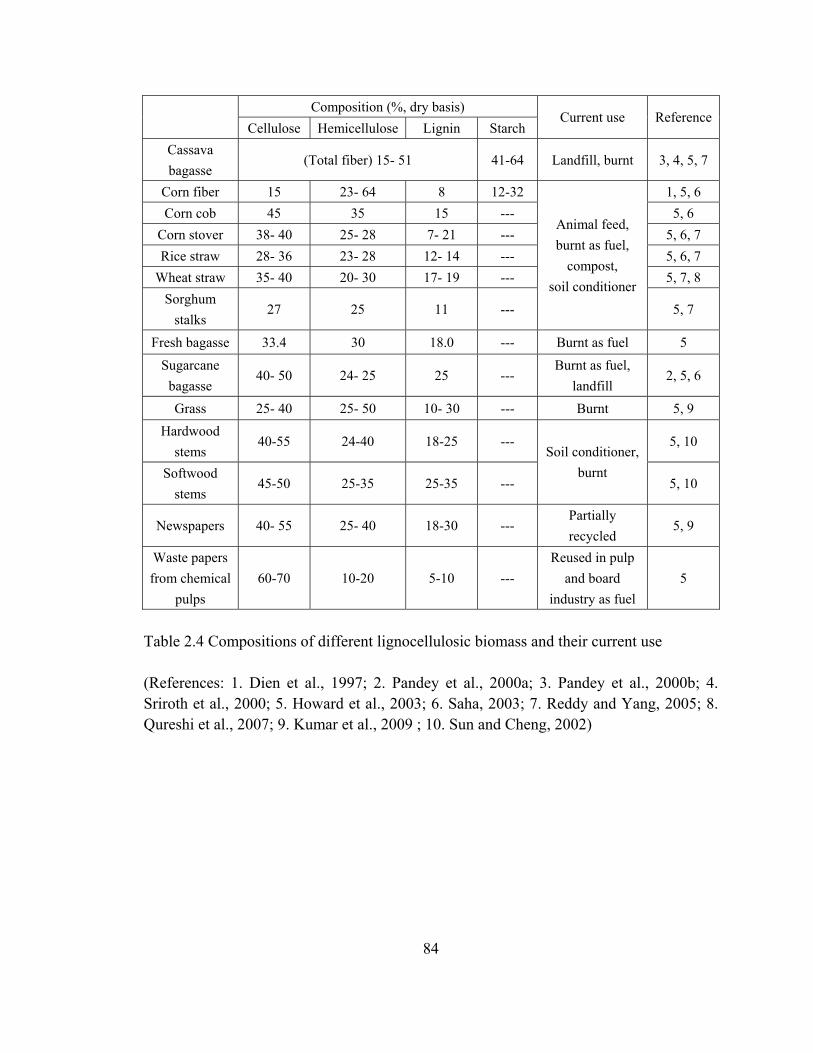
84
Composition (%, dry basis)
Cellulose Hemicellulose Lignin StarchCurrent use Reference
Cassava
bagasse (Total fiber) 15- 51 41-64 Landfill, burnt 3, 4, 5, 7
Corn fiber 15 23- 64 8 12-32 1, 5, 6
Corn cob 45 35 15 --- 5, 6
Corn stover 38- 40 25- 28 7- 21 --- 5, 6, 7
Rice straw 28- 36 23- 28 12- 14 --- 5, 6, 7
Wheat straw 35- 40 20- 30 17- 19 --- 5, 7, 8
Sorghum
stalks 27 25 11 ---
Animal feed,
burnt as fuel,
compost,
soil conditioner
5, 7
Fresh bagasse 33.4 30 18.0 --- Burnt as fuel 5
Sugarcane
bagasse 40- 50 24- 25 25 ---
Burnt as fuel,
landfill 2, 5, 6
Grass 25- 40 25- 50 10- 30 --- Burnt 5, 9
Hardwood
stems 40-55 24-40 18-25 --- 5, 10
Softwood
stems 45-50 25-35 25-35 ---
Soil conditioner,
burnt 5, 10
Newspapers 40- 55 25- 40 18-30 --- Partially
recycled 5, 9
Waste papers
from chemical
pulps
60-70 10-20 5-10 ---
Reused in pulp
and board
industry as fuel
5
Table 2.4 Compositions of different lignocellulosic biomass and their current use (References: 1. Dien et al., 1997; 2. Pandey et al., 2000a; 3. Pandey et al., 2000b; 4. Sriroth et al., 2000; 5. Howard et al., 2003; 6. Saha, 2003; 7. Reddy and Yang, 2005; 8. Qureshi et al., 2007; 9. Kumar et al., 2009 ; 10. Sun and Cheng, 2002)
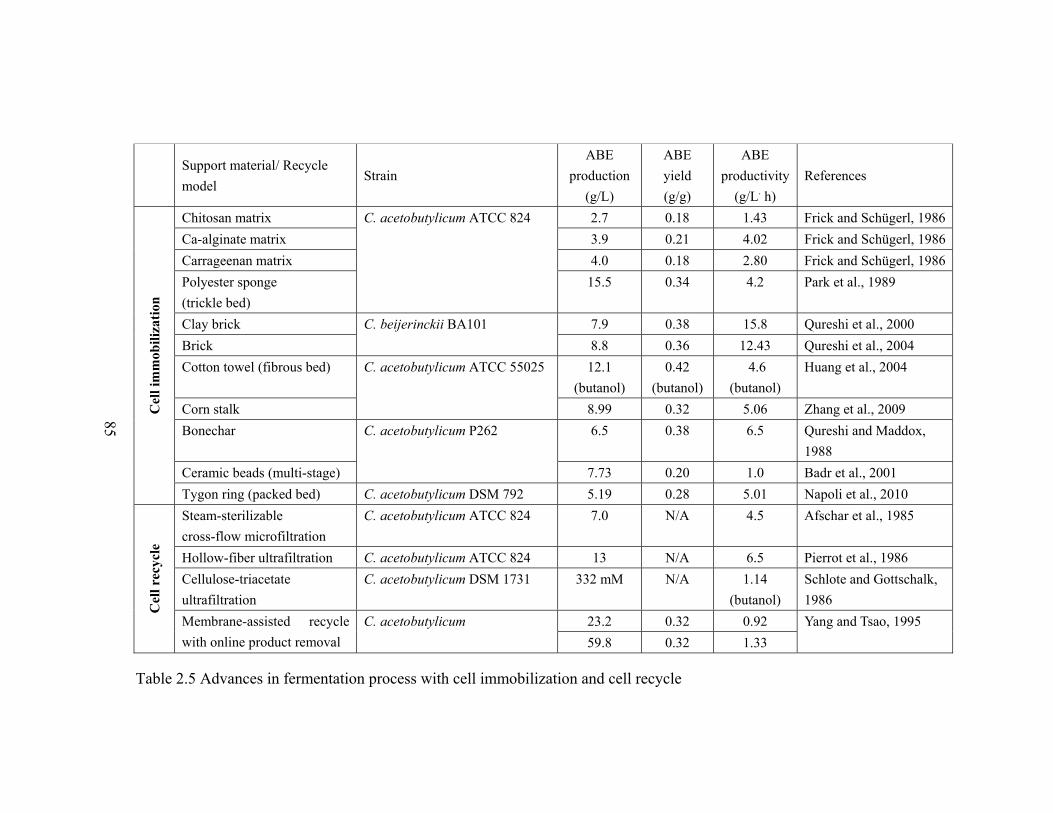
85
Table 2.5 Advances in fermentation process with cell immobilization and cell recycle
Support material/ Recycle
model Strain
ABE
production
(g/L)
ABE
yield
(g/g)
ABE
productivity
(g/L. h)
References
Chitosan matrix 2.7 0.18 1.43 Frick and Schügerl, 1986
Ca-alginate matrix 3.9 0.21 4.02 Frick and Schügerl, 1986
Carrageenan matrix 4.0 0.18 2.80 Frick and Schügerl, 1986
Polyester sponge
(trickle bed)
C. acetobutylicum ATCC 824
15.5 0.34 4.2 Park et al., 1989
Clay brick 7.9 0.38 15.8 Qureshi et al., 2000
Brick
C. beijerinckii BA101
8.8 0.36 12.43 Qureshi et al., 2004
Cotton towel (fibrous bed) 12.1
(butanol)
0.42
(butanol)
4.6
(butanol)
Huang et al., 2004
Corn stalk
C. acetobutylicum ATCC 55025
8.99 0.32 5.06 Zhang et al., 2009
Bonechar 6.5 0.38 6.5 Qureshi and Maddox,
1988
Ceramic beads (multi-stage)
C. acetobutylicum P262
7.73 0.20 1.0 Badr et al., 2001
Cel
l im
mob
iliza
tion
Tygon ring (packed bed) C. acetobutylicum DSM 792 5.19 0.28 5.01 Napoli et al., 2010
Steam-sterilizable
cross-flow microfiltration
C. acetobutylicum ATCC 824 7.0 N/A 4.5 Afschar et al., 1985
Hollow-fiber ultrafiltration C. acetobutylicum ATCC 824 13 N/A 6.5 Pierrot et al., 1986
Cellulose-triacetate
ultrafiltration
C. acetobutylicum DSM 1731 332 mM N/A 1.14
(butanol)
Schlote and Gottschalk,
1986
23.2 0.32 0.92
Cel
l rec
ycle
Membrane-assisted recycle
with online product removal
C. acetobutylicum
59.8 0.32 1.33
Yang and Tsao, 1995
85

Method Advantages Disadvantages
Mechanical Reduces particle size, cellulose
crystallinity, increase accessible areas
High power consumption, high cost
Steam
explosion
Causes hemicellulose degradation
and lignin transformation;
cost-effective
Destruction of a portion of the xylan
fraction; incomplete disruption of the
lignin- carbohydrate matrix; generation
of inhibitory compounds
Liquid hot
water
Avoids lignin and hemicellulose
degradation, reduces inhibitor
formation
Lower concentration due to large water
input
Ammonia fiber
explosion
Increases accessible surface area,
removes some lignin and
hemicellulose; does not produce
inhibitors
Inefficient for biomass with high lignin
content; high-cost, large demand of
ammonia; difficult to recycle ammonia
Acid Hydrolyzes hemicellulose to xylose
and other sugars; alters lignin
structure
High cost; equipment corrosion;
formation of toxic substances
Alkaline removes hemicellulose and lignin;
increases accessible surface area
long residence times required;
irrecoverable salts formed and
incorporated into biomass
Table 2.6 Comparison of leading pretreatment methods for improving the digestibility of lignocellulosic materials (References: Hendriks and Zeeman, 2009; Kumar et al., 2009; Mosier et al., 2005).
86

87
Categories Fermentation inhibitors, source of origin Sugar degradation products Furfural (from xylose)
5-hydroxymethyl furfural (HMF) (from hexose) Formic acid (from furfural and HMF) Levulinic acid (from HMF)
Lignin degradation products Vanillin, vanillic acid (from guaiacylpropane units) Syringaldehyde, syringic acid (from syringyl propane units) Hydroquinone (1,4-di-hydroxybenzene), 4-hydroxybenzoic acid Catechol (1,2-di-hydroxybenzene) p-Coumaric acid Ferulic acid Glucuronic acid Coniferyl aldehyde
Lignocellulose structure degradation product
Acetic acid (from the acetyl groups present in the hemicellulose)
Table 2.7 Major fermentation inhibitors present in the hydrolysates generated from lignocellulose degradation (References: Ezeji et al., 2007b; Cho et al., 2009; Mussatto and Roberto, 2004a; Palmqvis and Hahn-Hagerdal, 2000b; Zautsen et al., 2009)

88
Method Principle Advantages Disadvantages
Adsorption Adherence of solvents to
silicalite resin, clay,
activated carbon, or other
adsorptive materials
Easy to operate, low
energy requirement
High cost, low efficiency,
low selectivity, low
adsorbent capacity
Gas stripping Volatile solvents being
stripped out by gases and
then condensed
Easy to operate, no harm
to the culture, strips only
the volatiles, no fouling
Low selectivity
Liquid-liquid
extraction
Using the soluble
differences of solvents in
extractants and aqueous
phase for separation
High selectivity, efficient High cost, forming
emulsion, toxic to the
culture
Perstraction Membrane-based
extraction, separating the
fermentation broth from
the extractive solvents
High selectivity, low
toxic to the culture
compared to liquid-liquid
extraction
High cost, emulsion and
fouling problems
Pervaporation Using membrane to
selectively let the
vaporous solvents pass
trough, permeate side is
under vacuum
High selectivity,
relatively high mass flux
Membrane fouling
problem, more energy
required, high cost, not
easy to operate
Table 2.8 Alternative separation techniques for butanol recovery from ABE fermentation (References: Durre, 1998; Ezeji et al., 2004a; Ezeji et al., 2007a)

Conditions Stripping
Temp. (oC)
Condensation
Temp. (oC)
Stripping gas and gas
recycle rate*
Selectivity References
Integrated with batch reactor 34 -60 N2, 2.7 L/min ABE 23.4 Ennis et al., 1986b
Separate stripper, continuous
fermentation
30 -5 to -40 10 L L-1 min ABE 4.0 Groot et al., 1989
Separate stripper, continuous
fermentation
65 – 67 3 – 4 N2, 2.5 L/min ABE 30.5 Qureshi and Maddox,
1991
Integrated with fed-batch reactor 35 0 – 3 H2 and CO2,
3 – 3.2 L L-1 min
ABE 6 – 23 Qureshi et al., 1992
Integrated with batch reactor 34 -0.8 H2 and CO2,
1.5 – 3.3 L L-1 min
ABE 9.5 – 13 Maddox et al., 1995
Model solution 35 -2 N2, 4.6 L/min Butanol 10.3 – 13.8
Acetone 4.1 – 6.4
Ethanol 4.9 – 7.9
Integrated with batch reactor 33 – 35 -2 H2 and CO2, 3 L/min Butanol 6.7 – 13.2
Acetone 4.7 – 10.5
Ethanol 4.7 – 9.3
Ezeji et al., 2003
Integrated with fed-batch reactor 33 – 35 -2 H2 and CO2, 6 L/min Butanol 10.3 – 22.1 Ezeji et al., 2004b
89
Table 2.9 Solvent selectivities and operating conditions for butanol recovery in the gas stripping processes * L/min: liter gas per minute; L L-1 min: liter gas per liter broth per minute.
89

90
Membrane Membrane
thickness (μm)
Total flux
(gm-2h-1)
Selectivity Temp. (oC)
Feed, condensate
Feed CBuOH
(g/L)
References
Poly (dimethyl siloxane) (PDMS) 25 282 – 1000 15-35 50, -198 5 – 7 Hickey et al., 1992
PDMS 50 70 37 50, -198 10 Boddeker et al., 1990
PDMS 190 300 26.8 40, cold trap 10 – 50 Jonquieres and Fane, 1997
Zeolite filled PDMS 210 100 – 230 36-45 40, cold trap 10 – 50 Jonquieres and Fane, 1997
Silicalite filled PDMS 306 90 – 237 55 – 105 78, -198 7 – 78 Qureshi and Blascheck, 1999
Polytetrafluoroethylene (PTFE) 25-40 35 – 2100 2.7 – 4.8 30 – 55, dry ice 3 – 30 Vrana et al., 1993
Poly (methoxy siloxane) (PMS) N/A 150 – 400 10 – 15 50, -198 10 – 70 Hickey et al., 1992
Polyurethane (PU) 50 7 – 88 9 50, -198 10 Boddeker et al., 1990
Polyether block amide (PEBA) 50 60 – 800 20 50, -198 10 – 52.5 Boddeker et al., 1990
Polypropylene (PP) N/A 1400 – 1600 6.3 36, 5 3.5 – 14 Gapes et al., 1996
Silicone 1000 4.42 – 11.5 46 – 58 37, -30 14 – 17.5 Larrayoz and Puigjaner, 1987
Silicone 400 12.9 – 19.5 45 – 47 37, -60 4.3 – 17 Groot et al., 1984
Silicone (thin film) 50 52.8 42 30, cold trap 10 Huang and Meagher, 2001
Silicalite filled silicone 19 62.8 – 607 85.9 – 111.3 30 – 70, cold trap 10 Huang and Meagher, 2001
Zeolite (Ge-ZSM-5) 30 9.6 19 30, N/A 50 Li et al., 2003
Liquid membrane (oleyl alcohol) 25 25 – 450 180 30, -20 to -100 2.5 – 37.5 Matsumura and Kataoka, 1987
Liquid membrane (trioctylamine) N/A 8.3 – 10.7 71 – 104 54, -198 16.4 – 19.7 Thongsukmak and Sirkar,
2007
Table 2.10 Comparison of membrane performances for butanol recovery in the pervaporation processes
90

91
Solvent Toxicity Distribution coefficient
Butanol selectivity
Hexane N-T 0.5 2700 Heptane N-T 0.5 3300 Octane N-T 0.3 4100 Decane N-T 0.3 4300 Dodecane N-T 0.3 2900 Gasoline N-T 0.3 ND Hexanol T 12 160 Heptanol T 11 180 Octanol T 10 130 Oleyl alcohol N-T 3.6 ND Decanol T 8 200 Dodecanol T 6 140 Corn oil N-T 0.7 440 Olive oil N-T 0.7 470 Sesame oil N-T 0.3 220 Butyl acetate T ~ 3 ND Hexyl acetate N-T 3.6 5 Dibutyl phthalate N-T 1.4 3 Dibutyl adipate T 2.5 3 Dibutyl maleate T 2.0 3 Tributyl citrate N-T 2.4 2 Tributyrin N-T ND ND Ethyl oenanthate N-T 2.0 4 Methyl laurate N-T 1.8 7 Ethyl laurate N-T 1.7 7 Isopropyl myristate N-T 1.4 7 Isophytol N-T 3.2 ND Methyl oleate N-T 1.3 6 Ethyl oleate N-T 1.3 6 Ethyl stearate N-T 0.8 7 Butyl stearate N-T 1.2 ND Oleic acid N-T 3.9 6
ND: not determined; N-T: non toxic; T: toxic
Table 2.11 Solvents evaluation as extractants for butanol recovery by liquid-liquid extraction and their toxicity towards Clostridium beijerinckii (References: Barton and Daugulis, 1992; Groot et al., 1990)

92
Adsorbent Feed CBuOH
(g/L)
Butanol
adsorption
capacity (mg/g)
Adsorbent
loading
(g/L)
References
Activated carbon 15.0 252 10 Groot and Luyben,
1986
Silicalite 21.5 97 40 Milestone and
Bibby, 1981
Silicalite 10.0 48 200 Meagher et al.,
1998
Silicalite 11.7 – 16.8 64 – 85 168 Maddox, 1982
Silicalite 8.3 63.5 85
XAD-16 9.2 75 85
Ennis et al., 1987
XAD-2 16.5 78 10
XAD-4 14.4 100 10
XAD-8 15.5 66 10
Groot and Luyben,
1986
Amberlite XAD-4 4.0 – 20.0 27 – 83 100 – 200
Amberlite XAD-7 4.0 – 20.0 22 – 69 100 – 200
Bonopore 4.0 – 20.0 23 – 74 100 – 200
Bonopore, nitrated 4.0 – 20.0 13 – 55 100 – 200
Nielsen et al., 1988
Polyvinylpyridine 14.9 68 100 Yang et al., 1994
Zeolite (CBV811) 4.8 – 9.0 98 – 117 7 – 25 Oudshoorn et al.,
2009
Poly(styrene-co-DVB) 5.0 22.3 – 56.3 100
Poly(methacrylate) 5.0 34.7 100
Poly(butrylene phthalate) 5.0 7.4 100
Nielsen and
Prather, 2009
Table 2.12 Performances and capacities of different adsorbent materials for butanol recovery by adsorption

93
Recovery
technique
Substrate Strain Fermentation
mode
ABE
(g/L)
ABE yield
(g/g)
Productivity
(g/L. h)
Ref.
Whey permeate C. acetobutylicum P262 Batch 70.0 0.35 0.32 1
Whey permeate C. acetobutylicum P262 Continuous 69.1 0.38 0.26 2
Glucose C. beijerinckii BA101 Batch 79.5 0.47 0.60 3
Glucose C. beijerinckii BA101 Fed-batch 232 0.47 1.16 4
Wheat straw C. beijerinckii P260 Batch 47.6 0.37 0.36 5
Liquefied corn starch C. beijerinckii BA101 Batch 23.9 0.43 0.31 6
Gas stripping
Saccharified liquefied corn
starch
Batch
Fed-batch
26.5
81.3
0.41
0.36
0.40
0.59
Whey permeate C. acetobutylicum P262 Continuous 42.0 0.34 0.14 2
Glucose C. beijerinckii BA101 Fed-batch 165 0.43 0.98 7
Glucose C. acetobutylicum ATCC 824 Fed-batch 155 0.35 0.18 8
Pervaporation
Glucose C. beijerinckii NRRL B592 Continuous 13.1 0.28 1.72 9
Glucose C. acetobutylicum ATCC 824 Batch 22.5–34.3 0.26–0.33 N/A 10
Glucose C. acetobutylicum ATCC 824 Fed-batch 50.5–96.5 0.33–0.36 1.4–2.3 11
Liquid-liquid
extraction
Whey permeate C. acetobutylicum P262 Continuous 23.8 0.35 0.14 2
Whey permeate + lactose C. acetobutylicum P262 Batch 136.6 0.44 0.21 12Perstraction
Whey permeate C. acetobutylicum P262 Continuous 57.8 0.37 0.24 2
Glucose C. acetobutylicum Batch 23.2 0.32 0.92 13
Fed-batch 59.8 0.32 1.33
Adsorption
Repeated
fed-batch
387.3 0.32 1.69
Table 2.13 Integrated processes for enhanced ABE production from various substrates and strains (References: 1. Maddox et al.,1995; 2. Qureshi et al., 1992; 3. Ezeji et al., 2003; 4. Ezeji et al., 2004b; 5. Qureshi et al., 2007; 6. Ezeji et al., 2007c; 7. Qureshi and Blaschek, 2000; 8. Qureshi et al., 2001a; 9. Gapes et al., 1996; 10. Roffler et al., 1987a; 11. Roffler et al., 1987b; 12. Qureshi and Maddox, 2005; 13. Yang and Tsao, 1995)
93

Figure 2.1 Metabolic pathway of Clostridium acetobutylicum from glucose to acids and solvents during acidogensis and solventogensis (Lee et al., 2008).
94
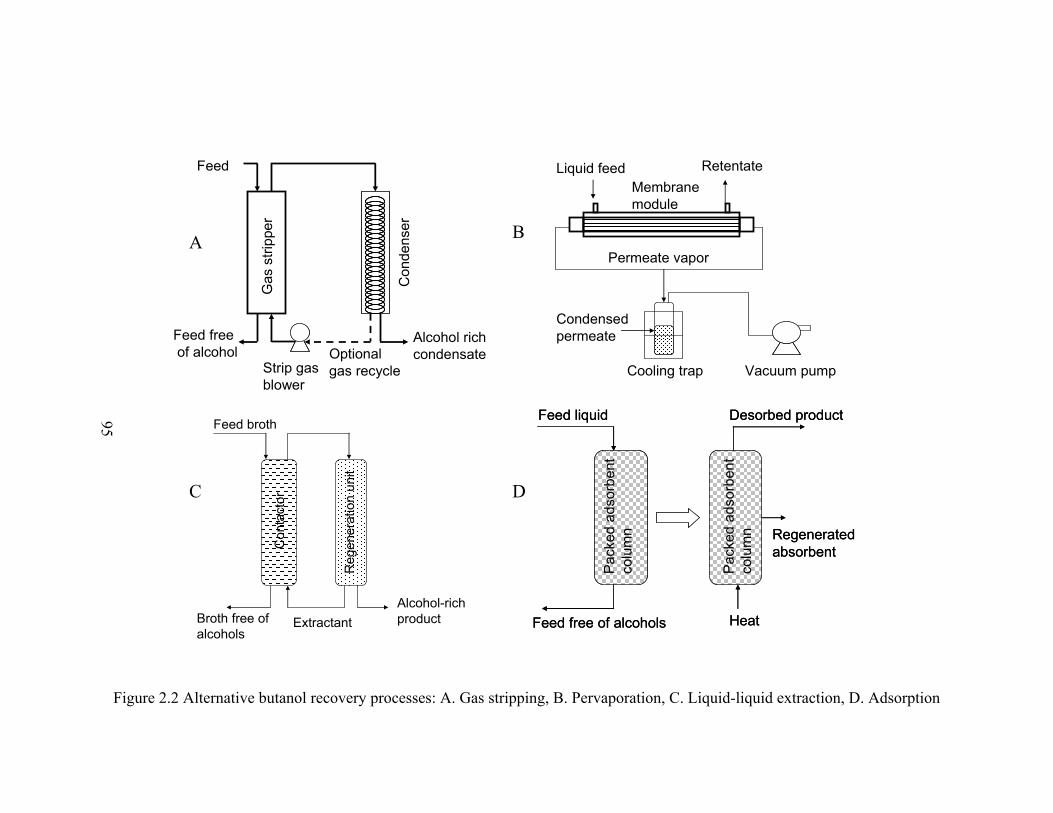
95
Feed liquid
Pac
ked
adso
rben
tco
lum
n
Pac
ked
adso
rben
tco
lum
n
Heat
Desorbed product
Feed free of alcohols
Regeneratedabsorbent
Feed liquid
Pac
ked
adso
rben
tco
lum
n
Pac
ked
adso
rben
tco
lum
n
Heat
Desorbed product
Feed free of alcohols
Regeneratedabsorbent
Figure 2.2 Alternative butanol recovery processes: A. Gas stripping, B. Pervaporation, C. Liquid-liquid extraction, D. Adsorption
Gas
str
ippe
r
Feed
Feed freeof alcohol
Strip gas blower
Alcohol rich condensate
Con
dens
er
Optionalgas recycle
Liquid feed Retentate
Cooling trap Vacuum pump
Permeate vapor
Condensed permeate
Membranemodule
Feed broth
Broth free ofalcohols
Con
tact
or
Reg
ene
ratio
n un
it
Extractant
Alcohol-richproduct
95
A
C D
B

96
Chapter 3: Butanol Production from Corn Fiber Hydrolysate by Clostridium
beijerinckii in a Fibrous Bed Bioreactor
Abstract
Sulfuric acid and enzyme hydrolyzed corn fiber was studied as a potential feedstock
for acetone-butanol-ethanol (ABE) fermentation with a hyper butanol producing mutant
Clostridium beijerinckii JB 200 in a fibrous bed bioreactor. Fermentation kinetics in
glucose, xylose, and glucose/xylose media were studied first to evaluate the hexose and
pentose sugar utilization by the mutant cells. Corn fiber hydrolysate (CFH) containing
71.6 g/L (39.4 g/L glucose, 23.3 g/L xylose and 8.9 g/L arabinose) total sugar was
obtained after dilute acid and enzyme hydrolysis. CFH severely inhibited ABE
production (1.9 g/L) due to the inhibitors generated during the severe pretreatment
process. Boiling and activated carbon were investigated as a detoxification method for
CFH in this study. Using detoxified CFH, 8.8 g/L ABE was produced with 38.3 g/L
reducing sugar left at the end of fermentation. 12.7 g/L ABE was produced when the
detoxified CFH was further diluted and all the reducing sugars were depleted within 65 h.
These results suggested that boiling and activated carbon was effective in removing

97
inhibitors from CFH, and further diluting the CFH reduced the inhibition to a negligible
level.
3.1 Introduction
Butanol is an important industrial chemical and solvent used in many fields, such as
the food and cosmetics industries. Recently, butanol has been attracting more attention as
a superior transportation fuel. Compared to ethanol, butanol has many advantages such as
high energy density, low vapor pressure and a similar air/fuel ratio to gasoline (Dürre,
2007). In addition, butanol is compatible with current pipeline for transportation, and can
be used directly as a gasoline replacement or fuel additives (Lee et al., 2008). Currently,
butanol is predominantly produced through petrochemical routes (Dürre, 1998). In recent
years, the surging crude oil price and increasing concern for environmental issues have
renewed interest in biological butanol production through acetone-butanol-ethanol (ABE)
fermentation (Dürre, 1998; Ezeji et al., 2004; 2007a; Lee et al., 2008).
Solventogenic Clostridia bacteria are commonly employed in ABE fermentation to
produce butanol, and sugar or starch based substrates such as molasses and liquefied corn
starch are used as commercial substrates in this process (Ezeji et al., 2007b; Qureshi and
Blaschek, 2001). However, these substrates are also food sources, and the limited
quantity is not able to meet the large global butanol demand. It was estimated that
substrate cost has a significant impact on final butanol price, accounting for over 56% of

98
the production cost (Qureshi and Blaschek, 2000; 2001). Therefore it is of interest to
search for other potential low-cost feedstocks for ABE fermentation. Clostridia are
known to be able to ferment various hexose and pentose sugars (Ezeji et al., 2004); this
feature makes it possible to utilize lignocellulosic feedstocks as potential substrates,
replacing costly starch and sugar. Lignocellulosic biomass is the most abundant
renewable carbon source on earth and it includes many low-value bio-wastes such as
agro-industrial residues and forestry residues (Howard et al., 2003; Saha, 2003; Sun and
Cheng, 2002). Lignocellulose mainly contains lignin, carbohydrate (hemicellulose and
cellulose), ash, protein, and some extractives (Kumar et al., 2009; Mosier et al., 2005).
Hemicellulose and cellulose are sugar polymers, and can be converted into various
pentose and hexose sugar such as xylose, arabinose and glucose. Corn is the staple
product in many agro-based states in the United States, and many byproducts such as
corn fiber are produced in large quantities from the corn refinery industry. Currently, corn
fiber is considered as waste product in the milling process, and is either sold as a
low-value animal feed or directly disposed into landfills. Corn fiber contains
approximately 5-10% lignin, 20-30% cellulose, 25-35% hemicellulose, and 7-25%
residue starch depending on region and milling processes (Saha, 2003; Mosier et al., 2005;
Zhu et al., 2002). The rich carbohydrate content in corn fiber makes it very appealing as a
carbon source for ABE fermentation. Using corn fiber as the feedstock for ABE
production not only adds value to the corn refinery industry, but also improves the overall

99
process economics of fermentative butanol production.
The objective of this study was to investigate the feasibility of using corn fiber for
butanol production with a mutant strain of Clostridium beijerinckii in a fibrous bed
bioreactor (FBB). Corn fiber was studied as a potential carbon source, and dilute acid and
enzymatic hydrolysis were investigated for sugar yield in corn fiber hydrolysate (CFH).
ABE fermentation using non-detoxified CFH was investigated, and activated carbon and
boiling were employed as detoxification to further improve the fermentability of the
CFH.
3.2 Materials and methods
3.2.1 Hydrolysis of corn fiber
Corn fiber obtained from Cargill’s Corn Milling Division (IA, USA) was dried at
60oC for 12 hours to remove the moisture content before experiments. For acid hydrolysis,
10 g dried corn fiber was mixed with 90 ml 0.1 N hydrochloric acid, corresponding to a
10% solid loading. The mixture was then sent to autoclave at 121 oC and 15 psig for 45
min for acid hydrolysis of corn fiber. After acid hydrolysis, hemicellulose fraction of the
corn fiber was converted to primarily pentose sugars, leaving cellulose behind in the
insoluble residue.
After acid hydrolysis, the mixture was then neutralized with NaOH to pH 5.0 under
an aseptic environment. Cellulase (Accellerase 1500, endoglucanase activity: 2200-2800

100
CMC U/g, beta-glucosidase activity: 525-775 pNPG U/g, Genencor, NY, USA) was then
added at a loading of 0.1 ml/g dry CF to hydrolyze the remaining cellulose. The
enzymatic hydrolysis was operated at 50oC, pH 5.0, and 200 rpm for 24 hours. The
remaining large insolubles were first removed by cheese cloth, and the liquid fraction was
centrifuged at 7000 rpm for 10 minutes to remove smaller particles. The clear liquid, corn
fiber hydrolysate (CFH) was used in the fermentation studies as carbon source.
3.2.2 Detoxification
Activated carbon and boiling was employed as detoxification method in this study to
improve the fermentability of the CFH. CFH was first heated until boiling for 10 min at
200 rpm to remove volatile inhibitors, such as acetic acid, furfural and HMF. It was
reported that temperature, pH, concentration and contact time were all operation
parameters in activated carbon detoxification. After boiling, 2% (w/w) activated carbon
was added into CFH. The temperature was maintained at 80oC and the mixture was
stirred at 400 rpm for 30 minutes. Activated carbon was then removed by centrifuging at
7000 rpm for 10 minutes. The clear CFH was then used as detoxified CFH for future
fermentation studies. Samples were taken after detoxification for sugar analysis.
3.2.3 Culture and media
A hyper butanol producing mutant, Clostridium beijerinckii JB 200, was isolated

101
through adaptation and evaluation engineering under stressful butanol environment in
FBB from the parental strain Clostridium beijerinckii ATCC 55025 (obtained from ATCC
deposit) by Dr. Jingbo Zhao in our research lab at The Ohio State University. This
asporogenous mutant was used as the solventogenic bacterium in this study for all ABE
fermentations. C. beijerinckii JB 200 was stored in a 15% glycerol-P2 stock solution in
the -80oC freezer. The culture was inoculated into 100 ml tryptone-yeast extract-glucose
growth medium in a rubber-capped serum bottle and incubated anaerobically for 12-15 h
at 37oC.
P2 medium was used in this study. P2 medium contained carbon source (glucose,
xylose, or CFH), yeast extract (1 g/L), buffer (0.5 g/L KH2PO4 and 0.5 g/L K2HPO4),
2.2g/L ammonium acetate, vitamin (0.001 g/L para-amino-benzoic acid (PABA),
0.001g/L thiamin and 10-5 g/L biotin), and mineral salts (0.2 g/L MgSO4·7H20, 0.01 g/L
MnSO4· H20, 0.01 g/L FeSO4· 7H20, 0.01 g/L NaCl). Carbon source, nitrogen source
(yeast extract and ammonium acetate) and buffer were autoclaved at 121oC and 15 psig
for 30 minutes for sterilization. Minerals and vitamins were prepared at 100-fold and
1000-fold concentration, and were filter through 0.2 μm sterile membrane (25mm 0.2μm
syringe filter, Fisherbrand, NJ, USA) for sterilization.
3.2.4 Fermentation and cell immobilization in fibrous bed bioreactor
All fermentations were carried out in a 5L FBB-connected stirred-tank fermentor

102
(B.E. Marubishi, Co., Ltd., Model MD-300). The fibrous bed bioreactor was made of a
glass column packed with spiral wound cotton towels and stainless steel wire cloth. The
working volume of the FBB was about 400ml. Detailed construction of the FBB can be
referred to Silva and Yang (1995) or Yang (1996). The fermentor and the FBB were
autoclaved for 45 minutes for sterilization separately, and the FBB was then aseptically
connected to the fermentor. The fermentation system contained 2L medium, which was
sparged with nitrogen for 1-2 h until it was oxygen-free. All fermentation was maintained
at 37oC, agitated at 150 rpm, and pH controlled at 5.0 by adding ammonia. The actively
growing cells were inoculated into P2 glucose medium at 5% (v/v), and 24-48 hours were
allowed for growth until the OD600 reached ~5.0. Cell immobilization was then carried
out by circulating the broth into the FBB, allowing cells to attach to the fibrous matrix.
After 36-48 h, the cell density in the broth no longer decreased and most of the cells were
immobilized onto the FBB. The medium was drained and replace with a fresh P2 glucose
medium to allow the immobilized cells in the FBB to continue to grow without adding
fresh cells. The cell immobilization in FBB was repeated for several batches using P2
glucose medium until a stable and high cell density in FBB was achieved. Once cell
immobilization was done, no more cells were inoculated into the fermentor and the cells
immobilized in the FBB were used as the seed culture for each of the following
fermentation process. The fermentation broth in the fermentor was replaced with a
different fresh medium (glucose medium, xylose medium or CFH-based medium) to start

103
a new batch but the immobilized cells in the bioreactor were allowed to continuously
grow batch after batch. Samples were taken at intervals for analysis of cell density, sugar
consumption and ABE production.
3.2.5 Analytical methods
Cell density was measured as optical density by a spectrophotometer (Sequoia-turner,
model 340, Mountain View, CA, USA) at 600 nm wavelength. The OD 600nm was then
converted to cell density by a calibration curve (1 unit of O.D. 600nm corresponded to
0.5899 g/L). The concentrations of glucose, xylose and arabinose were measured by a
high performance liquid chromatography (HPLC) with an organic acid column (Bio-Rad
HPX-87, ion exclusion organic acid column, 300 mm × 7.8mm). Samples were
centrifuged at 13.2 g for 5 min in microcentrifuge tubes and diluted 10 times with
distilled water prior to analysis on HPLC. HPLC was run at 45 oC using 0.01N H2SO4 as
the eluent at a flow rate of 0.6 ml/min. 15μL sample was injected by an automatic injector
(SIL-10Ai) and the running time was set at 36 min. A refractive index (RI) detector
(Shimadzu RID-10A) was set at the range of 200 to detect the organic compounds in the
sample. The HPLC column was installed in a column oven (CTO-10A) with temperature
control at 45 oC. Peak height was used to calculate concentration of sugars in the sample
based on the peak height of standard sample.
The fermentation products, acetone, butanol, ethanol, acetic acid, and butyric acid

104
were measured by a Shimadzu GC-2014 gas chromatograph (GC) (Shimadzu, Columbia,
MD, USA) equipped with a flame ionization detector (FID) and a 30.0 m fused silica
column (0.25m film thickness and 0.25 mm ID, Stabilwax-DA). The gas chromatograph
was operated at an injection temperature of 200 oC with 1 μL of the acidified sample
injected by the AOC-20i Shimadzu auto injector. Column temperature was held at 80 oC
for 3 min, raised to 150 oC at a rate of 30 oC/min, and held at 150 oC for 3.7 min.
3.3. Results and discussion
3.3.1 ABE fermentation in glucose, xylose and glucose/xylose mixture medium
36.4 g/L ABE, of which 22.2 g/L was butanol, was produced by C. beijerinckii JB
200 using 88.5 g/L glucose in ABE batch fermentation, compared with 13.5 g/L butanol
produced by the parental strain C. beijerinckii ATCC 55025 (fermentation kinetics not
shown in this paper). A 64.4% increase in butanol production indicated that this
hyper-butanol-producing mutant was superior in butanol tolerance and butanol
production than the parental strain. FBB was proven to be a powerful tool for cell
adaptation and evolution towards high butanol production. The fibrous matrix provided
excellent support for cell attachment, achieving high cell density in FBB. The viability of
the cells was also guaranteed since FBB allowed constant cell-renewal through dynamic
adsorption and desorption (Yang, 1996). Cells gradually adapted to tolerate high butanol
concentration and mutated to produce more butanol. Therefore, FBB and the mutant

105
strain were both well-suited for the purpose of this study.
Xylan, stored in the form of hemicellulose, is the main component in lignocellulosic
biomass. Xylose, which is the building block of xylan in hemicellulose, represents the
most abundant pentose sugar in lignocellulosic hydrolysate (Saha, 2003). Before using
CFH-based medium, control batch fermentations were carried out using glucose, xylose
and glucose-xylose mixture as carbon sources to evaluate the mutant’s ability for utilizing
hexose and pentose sugars. The initial sugar concentration in all the control studies was
prepared at about 45 g/L. 15.5, 16.9 and 16.7 g/L ABE were produced from glucose,
xylose, and glucose/xylose medium, respectively (Figure 3.1). ABE yield was 0.35, 0.41,
and 0.39 g/g, respectively (Table 3.1). Sugars in all control experiments were quickly
consumed, suggesting that all fermentations stopped due to lack of carbon source at the
end. The final butanol concentration was not yet at the inhibitory level for the bacteria,
and if given sufficient carbon source, more butanol could be produced. The ABE yield in
glucose-based medium was lower compared to xylose-based medium, which was
probably due to more biomass production in the presence of glucose. More of the carbon
source was directed to cell biomass formation in the beginning of glucose fermentation.
Once cell density reached a certain level in the broth, the carbon source was then directed
to ABE production. Glucose was consumed faster by the mutant cells compared to xylose,
which also explained the higher ABE productivity in glucose-based medium (Table 3.1.)
It should be noted that the mutant cells utilized both hexose and pentose sugar efficiently,

106
and produced similar amount of ABE in all sugar studies (Figure 3.1). When both glucose
and xylose were present (Figure 3.1c), glucose was quickly consumed within 40 h,
whereas xylose concentration decreased slightly in the beginning. Once glucose was
depleted, the mutant instantly switched to pentose utilization pathway, and began to
quickly utilize xylose and convert it into more ABE. This result showed that the mutant
can utilize both hexose and pentose sugar simultaneously, but at different rates when both
sugars were present. Glucose was favored with a faster rate, while xylose was consumed
at a slower rate.
3.3.2 ABE fermentation in undetoxified CFH-based medium
Sulfuric acid and enzyme hydrolyzed CFH contained 39.4 g/L glucose, 23.3g/L
xylose, 8.94 g/L arabinose and 3.25 g/L acetic acid. 22.6g/L glucose was released from
hemicellulose and residue starch in corn fiber after sulfuric acid hydrolysis, whereas the
additional 16.8 g/L glucose was released from cellulose by enzymatic hydrolysis. Xylose
and arabinose were obtained during the sulfuric acid hydrolysis from the hemicellulose
fraction, while cellulose remained insoluble in the residue solids and later digested by the
enzyme. The CFH result obtained after acid hydrolysis was very similar to our group’s
previous study (Zhu et al., 2002). Compared to another study (Qureshi et al., 2008a), the
glucose concentration was higher in our CFH. This is probably due to the residue starch
present in the CFH, the content of which can be very different depending on milling

107
process and geographic regions. An initial 71.6 g/L total sugar was present in the CFH,
which makes CFH suitable as a carbon source for the fermentation studies.
It was anticipated that various fermentation inhibitors were present in the CFH, such
as furan derivatives (furfural and HMF), phenolic compounds and acids (Palmqvist and
Hahn-Hagerdal, 2000a; 2000b; Mussatto and Roberto, 20004a). It was reported that using
undetoxified CFH as carbon source for ABE production, the cell growth was poor and
ABE production was less than 1.6 g/L (Qureshi et al., 2008a). Similar results were also
observed during our study using undetoxified CFH. Only 1.9 g/L total ABE was produced,
of which 1.3 g/L was butanol (data not shown). Most of the sugars remained in the broth
after the fermentation stopped due to the severe inhibition. It was reported that even
though the fermentation inhibitors were present at low concentrations (Ezeji et al., 2007c),
most of the microorganisms were still very sensitive to these toxic compounds. Diluting
the CFH by half still resulted in a reasonable amount of sugars in the CFH, and reduced
the inhibitor concentration by half, which could be very significant in improving the
fermentability of the CFH. Therefore, the CFH was diluted by half before used for
fermentation to investigate if the inhibition effect could be alleviated. The results are
shown in Figure 3.2. 8.1 g/L ABE was produced from the diluted undetoxified CFH
medium, of which 5.2 g/L was butanol. The ABE yield and butanol yield were 0.32 g/g
and 0.2 g/g respectively. After about 55 h, the fermentation slowed down and eventually
stopped due to inhibition. At the end of the fermentation, 7.22 g/L xylose, and 3.16 g/L

108
arabinose were measured in the broth. This result indicated that sugar concentration was
not the primary limitation in this fermentation, and glucose was preferably consumed by
the mutant strain over the other two pentose sugars. Acetic acid and butyric acid
increased throughout the fermentation, and 5.6 g/L acetic acid and 7.1g/L butyric acid
were measured at the end of fermentation. The high acid accumulation indicated that the
metabolic shift from acidogensis to solventogensis was unsuccessful (Ezeji et al., 2007c;
Ezeji and Blaschek, 2008), and these acids could not be converted into acetone and
butanol.
Compared to the control batch fermentation (Table 3.1), the ABE yield and butanol
yield were both lower in the diluted undetoxified CFH fermentation, suggesting that part
of the carbon source was used to provide extra energy for the mutant to survive in the
toxic and inhibitory environment. By diluting the CFH by half, the concentration of all
the possible inhibitors was reduced by half, which could be very significant in improving
the fermentability of the CFH (Ezeji et al., 2007c). A 326% increase in ABE production
was achieved compared to using undiluted and undetoxified CFH fermentation,
indicating that the inhibition was significantly relieved after dilution by half. However,
glucose was not completely utilized by the mutant, and the fermentation stopped due to
inhibition before xylose and arabinose could be utilized. This suggested that
detoxification of the CFH was needed in order to improve the fermentability of the CFH
and ABE production.

109
3.3.3 ABE fermentation in boiling and activated carbon detoxified CFH-based medium
CFH was detoxified with boiling and activated carbon before fermentations.
Activated carbon is used to adsorb the toxic inhibitors to detoxify the CFH, and is
economic compared to ion-exchange resins (Mussatto and Roberto, 2004a; 2004b;
Mussatto et al., 2004). It was also reported that increasing the operating temperature can
facilitate inhibitor removal in the activated carbon detoxification, such as phenolic
compounds (Mussatto and Roberto, 2004a). In addition, boiling and increasing
temperature can significantly influence the removal of volatile inhibitors, such as furfural,
HMF and acetic acid (Coverti et al., 2000). It was reported that overliming and activated
carbon removed 95% of the lignin degradation production, and acetic acid and furfural
were removed by boiling and striping. After CFH was detoxified, 64.8 g/L reducing sugar
was present (36.4 g/L glucose, 20.2 g/L xylose, and 8.2 g/L arabinose), corresponding to
a 9.5% reducing sugar loss compared to non-detoxified CFH. Detoxified CFH was
diluted by half to investigate the efficiency of detoxification compared to previous study
(section 3.3.2) using diluted undetoxified CFH. Undiluted detoxified CFH was also
studied as carbon source to evaluate the feasibility and efficiency of using CFH for ABE
fermentation.
12.7 g/L total ABE was produced in diluted detoxified CFH medium, of which 7.88
g/L was butanol (Figure 3.3). All reducing sugars were depleted at the end of the
fermentation, and fermentation stopped due to lack of carbon source after 66 h. No

110
inhibition was observed in this fermentation. ABE yield and butanol yield were 0.41 g/g
and 0.25 g/g, respectively, which was higher than the yields obtained using diluted
undetoxified CFH medium and comparable to those obtained in control studies (Table
3.1). Compared to the results in diluted non-detoxified CFH medium, ABE production
was increased by 56.8 %. It suggested that after CFH was detoxified and diluted by half,
most of the fermentation inhibitors were removed or below the inhibitory level to the
mutant cells. Fermentation kinetics obtained in the diluted detoxified CFH was very
similar to the control study using glucose/xylose mixture (Table 3.1). Glucose was first
consumed, followed by arabinose and xylose. The concentration of butyric acid decreased
and remained low after 26 h, suggesting successful conversion to butanol. At the end of
the fermentation, only 1.7 g/L butyric acid and 4.2 g/L acetic acid were measured. In
undiluted detoxified CFH study, 8.79 g/L ABE was produced, of which 5.58 g/L was
butanol (Figure 3.4). At the end of fermentation, 38.3 g/L reducing sugars (16.4 g/L
glucose, 17.3 g/L xylose, and 4.6 g/L arabinose) were measured. The butyric acid
concentration was high in the early stage of the fermentation (highest at 6.7 g/L), and
decreased after 40 h and remained at 5.0 g/L in the end of the fermentation. The acetic
acid concentration increased throughout the fermentation, and 6.1 g/L acetic acid was
measured at the end. ABE yield and butanol yield were 0.33 g/g and 0.21 g/g,
respectively (Table 3.1).
Comparing the results of detoxified and undetoxified CFH fermentation, it is evident

111
that boiling and activated carbon adsorption removed some of the inhibitors successfully
and improved the fermentability of the CFH significantly. ABE production was increased
from 1.9 g/L to 8.79 g/L, which was a 363% increase. Comparing the results of detoxified
and diluted detoxified CFH fermentation, it suggests that some inhibitors were still
present in the CFH after detoxification. The inhibition lowered the ABE production and
yield, and eventually stopped the fermentation before all the sugars could be utilized.
Diluting the detoxified CFH by half further reduced the concentration of the inhibitors by
half, at which level that was no longer toxic to the mutant cells. In addition, similar ABE
production was obtained in the diluted undetoxified CFH medium and undilted detoxified
medium, suggesting that boiling and activated carbon removed at least 50% of the
fermentation inhibitors (Table 3.1). Based on the results of detoxified CFH and diluted
detoxified CFH, it suggested that a 75% removal of inhibitors is essential in successfully
using CFH for ABE production without inhibition.
There have been other reported ABE production from different lignocellulosic
feedstocks, and they are summarized and compared with results in this work in Table 3.2.
It should be noted that results obtained using different feedstocks can not be compared
equally due to the large variation of composition difference in each lignocellulosic
biomass. In addition, different feedstocks require different pretreatment methods, which
are usually crucial in determining the inhibitor formation in the lignocellulosic
hydrolysate (Kumar et al., 2009). It was suggested that the generation of fermentation

112
inhibitors were feedstock and pretreatment specific (Qureshi et al., 2008a). More ABE
was produced from undetoxified whey straw hydrolysate compared to detoxified CFH
(Table 3.2), suggesting that whey straw hydrolysate may be less toxic to the C.
beijerinckii P260. Different pretreatment methods on the same feedstock, such as
distillers dried grains and solubles, also resulted in large variation in ABE production
(Table 3.2). Compared to similar work done with corn fiber, more ABE was produced in
this work from diluted detoxified CFH with higher yield and productivity. It should also
be noted that within 5g/L ABE production difference, both work using CFH produced
less ABE compared to other studies such as whey straw, suggesting that CFH is more
toxic to Clostridia and challenging to detoxify to a satisfactory level.
3.4. Conclusion
Sulfuric acid and enzyme hydrolyzed corn fiber was studied in this paper as a
potential feedstock for fermentative ABE production. A high sugar concentration (71.6
g/L total sugar) CFH was obtained in this work, which makes corn fiber a suitable carbon
source for the fermentation. Fermentation kinetics with glucose, xylose, and
glucose/xylose mixture suggested that the mutant can utilize both hexose and pentose
sugar efficiently. Similar ABE production (15.5 – 16.9 g/L) and yield (0.35 – 0.41 g/g)
were obtained in this study. Due to the fermentation inhibitors present in the CFH,
undetoxified CFH was very toxic to the mutant cells (1.9 g/L ABE). It was found that by

113
diluting the CFH by half, the inhibition effect was greatly alleviated and the
fermentability of the CFH was significantly improved. 8.1 g/L ABE was produced with a
0.32 g/g yield in the diluted undetoxified CFH-based medium within 68 h, which was a
326% increase in ABE production compared to undiluted and undetoxified CFH medium.
10.4 g/L reducing sugar was left at the end of the fermentation, indicating that
fermentation stopped due to inhibition instead of depletion of carbon source. After the
CFH was detoxified with boiling and activated carbon, 8.8 g/L ABE was produced with
0.33 g/g yield. Due to similar ABE production, yield and productivity from diluted
undetoxified CFH and undiluted detoxified CFH, it was evident that boiling and activated
carbon removed at least 50% of the inhibitors from CFH. After the detoxified CFH was
further diluted by half, 12.7 g/L ABE with a yield 0.41 g/g was obtained. All reducing
sugars were quickly depleted, and fermentation stopped due to lack of carbon source.
This indicated that after CFH was detoxified and diluted, all the fermentation inhibitors in
the CFH were below the toxic level to the mutant cells. It indicated that about 75%
removal of fermentation inhibitors is essential in successfully using CFH as a substrate
for ABE fermentation without inhibition with the mutant C. beijerinckii JB 200.
Compared to other studies (Table 3.2) using different lignocellulosic biomass, corn fiber
hydrolysate seems to be more challenging in detoxification and more toxic to the
Clostridia bacteria during the fermentation.

114
3.5 References
Converti, A., J.M. Dominguez, P. Perego, S.S. Silva and M. Zilli (2000). Wood hydrolysis and hydrolysate detoxification for subsequent xylitol production. Chem. Eng. Technol., 23, 1013-1020.
Durre, P. (1998). New insights and novel developments in clostridial acetone/ butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Durre, P. (2007). Biobutanol: An attractive biofuel. Biotechnol. J., 2, 1525-1534.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004). Butanol fermentation research: upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007a). Bioproduction of butanol from biomass: from genes to bioreactors. Current Opinion in Biotechnol., 18, 220-227.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007b). Butanol production from agricultural residues: impact of degradation products on Clostridium beijerinckii growth and butanol fermentation. Biotechnol. Bioeng., 97, 1460-1469.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007c). Production of acetone butanol (AB) from liquefied corn starch, a commercial substrate, using Clostridium beijerinckii coupled with product recovery by gas stripping. J Ind. Microbiol. Biotechnol., 34, 771-777.
Ezeji, T.C. and H.P. Blaschek (2008). Fermentation of dried distillers’ grains and solubles (DDGS) hydrolysates to solvents and value-added products by solventogenic clostridia. Bioresour. Technol., 99, 5232-5242.
Howard, R.L., E. Abotsi, E.L. Jansen van Rensburg and S. Howard (2003). Lignocellulosic biotechnology: issues of bioconversion and enzyme production. African J. Biotechnol., 2(12), 602-619.
Kumar, P., D.M. Barrett, M.J. Delwiche and P. Stroeve (2009). Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem., 48, 3713-3729.
Lee, S.Y., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim and K.S. Jung (2008). Fermentative butanol production by Clostridia. Biotechnol. Bioeng., 101, 209-228.

115
Mosier, N.S., C. Wyman, B.Dale, R.Elander, Y.Y. Lee, M. Holtzapple and M.R. Ladisch (2005). Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96, 673-668.
Mussatto, S.I. and I.C. Roberto (2004a). Alternatives for detoxification of diluted-acid lignocellulosic hydrolysates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Mussatto, S.I. and I.C. Roberto (2004b). Optimal experimental condition for hemicellulosic hydrolysate treatment with activated charcoal for xylitol production. Biotechnol. Prog., 20, 134-139.
Mussatto, S.T., J.C. Santos and I.C. Roberto (2004). Effect of pH and activated charcoal adsorption on hemicellulosic hydrolysate detoxification for xylitol production. J. Chem. Technol. Biotechnol., 79, 590-596.
Palmqvist, E. and B. Hahn-Hagerdal (2000a). Fermentation of lignocellulosic hydrolysates. I: inhibition and detoxification. Bioresour. Technol., 74, 17-24.
Palmqvis, E. and B. Hahn-Hagerdal (2000b). Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol., 74, 25-33.
Qureshi, N. and H.P. Blaschek (2000). Economics of butanol fermentation using hyper-butanol producing Clostridium beijerinckii BA 101. Trans. IChem E., 78, 139-144.
Qureshi, N. and H.P. Blaschek (2001). ABE production from corn: a recent economic evaluation. J. Ind. Microbio. Biotechnol., 27, 292-297.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst Eng., 30, 419-427.
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta and H.P. Blaschek (2008a). Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.
Qureshi, N., B.C. Saha, R.E. Hector, and M.A. Cotta (2008b). Removal of fermentation inhibitors from alkaline peroxide pretreated and enzymatically hydrolyzed wheat straw: Production of butanol from hydrolysate using Clostridium beijerinckii in

116
batch reactors. Biomass Bioenergy, 32, 1353-1358.
Saha, B.C. (2003). Hemicellulose bioconversion. J Ind Microbiol Biotechnol., 30, 279-291.
Silva, E.M. and S.T. Yang (1995). Kinetics and stability of a fibrous-bed bioreactor for continuous production of lactic from unsupplemented acid whey. J. Biotechnol., 41, 59-70.
Sun, Y. and J. Cheng (2002). Hydrolysis of lignocellulosic material from ethanol production: A review. Bioresour. Technol., 83, 1-11.
Wang, B., T. Ezeji, Z. Shi, H. Feng, and H.P. Blaschek (2009). Pretreatment and conversion of distiller’s dried grains with solubles for acetone-butanol-ethanol (ABE) production. Transactions of the ASABE, 52, 885-892.
Yang, S.T. (1996). Extractive fermentation using convoluted fibrous bed bioreactor. US Patent No. 5,563,069.
Zhu, Y., Z. Wu, and S.T. Yang (2002). Butyric acid production from acid hydrolysate of corn fibre by Clostridium tyrobutyricum in a fibrous bed bioreactor. Process Biochem., 38, 657-666.

117
Butanol ABE
Substrate
(g/L)
Inhibition+
level Titer (g/L)
Yield (g/g)
Productivity(g/L. h)
Titer (g/L)
Yield (g/g)
Productivity (g/L. h)
Glucose 100 N/A 22.2 0.25 0.32 36.4 0.41 0.52 Glucose 43.8 N/A 9.8 0.22 0.2 15.5 0.35 0.32 Xylose 40.8 N/A 10.4 0.25 0.14 16.9 0.41 0.23
Glu+Xyl Glu 21.4Xyl 20.8
N/A 10.5 0.25 0.14 16.7 0.39 0.23
Undetoxified CFH Glu 39.4Xyl 23.3Ara 8.9
100% 1.3 0.15 --- 1.9 0.22 ---
Diluted undetoxified CFH
Glu 19.2Xyl 11.7Ara 4.5
50% 5.2 0.20 0.08 8.1 0.32 0.12
Detoxified CFH Glu 36.4Xyl 20.2Ara 8.4
~50%* 5.6 0.21 0.09 8.8 0.33 0.15
Diluted detoxified CFH
Glu 17.9Xyl 9.9 Ara 4.0
~25%* 7.9 0.25 0.12 12.7 0.41 0.19
Table 3.1 ABE fermentation by C. beijerinckii JB 200 using glucose, xylose, glucose-xylose mixture and CFH.
+: inhibition level is defined as 100% for undetoxified, and 50% if diluted by half; ---: not calculated due to poor growth; *: derived based on similar results in this study
117

Feedstock Pretreatment and
hydrolysis
Inhibitor
removal
Culture ABE
production
(g/L)
ABE
Yield
(g/g)
ABE
productivity
(g/L.h)
References
Corn fiber Dilute acid+
enzyme
Overliming +
XAD-4 resin
C. beijerinckii
BA101
9.3 0.39 0.1 Qureshi et al.,
2008a
Wheat straw Dilute sulfuric acid None C. beijerinckii
BA101
25.0 0.42 0.6 Qureshi et al.,
2007
Wheat straw Alkaline peroxide +
enzymes
Electrodialysis C. beijerinckii
P260
22.17 0.41 0.55 Qureshi et al.,
2008b
Distillers dried
grains and
solubles
Dilute acid/
Liquid hot water/
AFEX + enzyme
Overliming 5 different
species of
Clostridia
bacteria
4.9–12.1
10.5– 12.9
7.9–11.6
0.3–0.35
0.31–0.34
0.32–0.34
N/A Ezeji and
Blaschek,
2008
Distillers dried
grains and
solubles
Alkaline
electrolyzed water
+ enzyme
None C.
acetobutylicum
P260
16.9
(30% solid
loading)
N/A N/A Wang et al.,
2009
Corn fiber Dilute acid+
enzyme
Boiling+
activated
carbon
+dilution
C. beijerinckii
JB 200
12.7 0.41 0.19 This work
Corn fiber Dilute acid+
enzyme
Boiling+
Activated
carbon
C. beijerinckii
JB 200
8.8 0.33 0.15 This work
Table 3.2 Butanol production from different lignocellulosic hydrolysates
118
118

0
5
10
15
20
25
30
35
40
45
50
0 10 20 30 40 50
Time (h)
Glu
cose
(g
/L)
0
2
4
6
8
10
12
14
16
18
So
lve
nts
, aci
ds,
ce
ll (g
/L)
Glucose
Acetone
Butanol
Ethanol
Acetic acid
Butyric acid
Cells
Total ABE
A
0
5
10
15
20
25
30
35
40
45
0 10 20 30 40 50 60 70 80 90
Time (h)
Xyl
ose
(g
/L)
0
2
4
6
8
10
12
14
16
18
So
lve
nts
, aci
ds,
ce
ll (g
/L)
Xylose
Acetone
Butanol
Ethanol
Acetic acid
Butyric acid
Cells
Total ABE
B
Figure 3.1 ABE production from glucose, xylose, and glucose-xylose mixture by C. beijerinckii JB 200. (A) Glucose, (B) Xylose, (C) Glucose-xylose mixture.
119

Figure 3.1 continued
0
2
4
6
8
10
12
14
16
18
20
22
0 10 20 30 40 50 60 70 80
Time (h)
Glu
cose
,xyl
ose
(g
/L)
0
2
4
6
8
10
12
14
16
18
So
lve
nts
, aci
ds,
ce
ll (g
/L)
Glucose
Xylose
Acetone
Butanol
Ethanol
Total ABE
Acetic Acid
Butyric Acid
Cells
C
120

0
2
46
8
10
12
14
1618
20
22
0 10 20 30 40 50 60 70
Fermentation Time (h)
Su
ga
rs (
g/L
)
Glucose Xylose Arabinose
A
0123456789
101112
0 10 20 30 40 50 60 70
Fermentation Time (h)
Pro
du
cts
(g/L
)
Acetone Butanol Ethanol
Total ABE Acetic Acid Butyric Acid
B
Figure 3.2 ABE production from diluted undetoxified CFH by C. beijerinckii JB 200. (A) Sugar utilization, (B) Solvents and acids production.
121

0
2
4
6
8
1012
14
16
18
20
22
0 10 20 30 40 50 60 70
Fermentation Time (h)
Su
ga
rs(g
/L)
Glucose Xylose Arabinose
A
0
2
4
6
8
10
12
14
16
0 10 20 30 40 50 60 70
Fermentation Time (h)
Pro
du
cts
(g/L
)
Acetone Butanol Ethanol
Total ABE Acetic Acid Butyric Acid
B
Figure 3.3 ABE production from diluted detoxified CFH by C. beijerinckii JB 200. (A) Sugar utilization, (B) Solvents and acids production.
122

0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60 70
Time (h)
Su
ga
rs(g
/L)
Glucose Xylose Arabinose
A
0
2
4
6
8
10
12
14
0 10 20 30 40 50 60 70
Time (h)
Pro
du
cts
(g
/L)
Acetone Butanol Ethanol
Total ABE Acetic Acid Butyric Acid
B
Figure 3.4 ABE production from undiluted detoxified CFH with C. beijerinckii JB 200. (A) Sugar utilization, (B) Solvents and acids production.
123

124
Chapter 4: Evaluation of Butanol Recovery by Gas Stripping
from Model solution and Fermentation Broth
Abstract
Condensation temperature, gas flow rate, and cells and components from
fermentation broth were all known factors that affect the performance of gas stripping. In
this study, these factors were investigated for their effects on solvent removal rate,
condensate composition, and solvent selectivities in a gas stripping process using both
model solution and fermentation broth obtained from Clostridium beijerinckii with P2
medium. Low condensation temperature and high gas flow rate was found to have
positive effect on butanol removal rate, but negative effect on condensate concentration
due to excess removal of water. The presence of cells did not have a significant effect on
butanol removal rate, but adversely affected the butanol selectivity. No clear relationship
was observed between butanol selectivity and concentration, whereas acetone and ethanol
selectivity stayed in a narrow range regardless of concentration.

125
4.1 Introduction
Biological butanol production via acetone-butanol-ethanol (ABE) fermentation has
been the research focus in recent years since butanol has been widely recognized as a
superior biofuel to ethanol (Jones and Woods, 1986; Kumar and Gayen, 2011; Lee et al.,
2008). The fuel properties of butanol, including energy density, air-fuel ratio, research
octane number and motor octane number, are all superior to ethanol and very similar to
gasoline (Lee et al., 2008). Even though the primary application of butanol is currently
industrial solvent, butanol received great attention when David Ramey drove his car
fueled solely by butanol across the country in 2005, which promoted butanol as an
alternative biofuel (Dürre, 2007). Due to depleting fossil fuel resources and high prices of
crude oil, butanol production through ABE fermentation has been favored with renewed
research interests over the petrochemical synthesis. Many breakthroughs and advances
have been achieved in the past few decades with efforts to improve on ABE fermentation,
from upstream strain improvement, process design, to downstream butanol recovery
(Dürre, 1998; Ezeji et al., 2004a; 2007a; 2010; Qureshi and Ezeji, 2008; Zheng et al,
2009).
Butanol is very toxic and inhibitory to microorganisms in fermentation when present
at concentrations as low as 5-10 g/L (Qureshi and Ezeji, 2008). The typical ABE
production from batch fermentation is usually between 15-18 g/L within 40-60 h (Dürre,
1998). Even with a hyper-butanol-producing mutant Clostridium beijerinckii BA101

126
developed recently, only 25-33 g/L solvents were produced in batch process with
traditional substrates such as glucose, starch and molasses (Ezeji et al., 2004a; Qureshi
and Blaschek, 2001a). Compared with typical concentration of 10% in ethanol production
by yeast, this low concentration of ABE (1-2%) significantly increases the cost of butanol
recovery by distillation due to the dilute concentration and large process stream volume
(Vane, 2008). This limitation negatively impacts economic butanol production from ABE
fermentation relative to petrochemical synthesis. Many separation technologies have been
developed in recent years as an alternative to distillation for butanol recovery from dilute
solutions, including gas stripping, pervaporation, liquid-liquid extraction and adsorption
(Ezeji et al., 2004a; Vane, 2008; Zheng et al., 2009). These alternative separation
technologies can be integrated with fermentation to simultaneously recover butanol,
relieving end product inhibition and resulting in enhanced overall butanol production and
reactor productivity. When coupling with integrated product recovery, ABE fermentation
can be operated for an extended period of time utilizing highly concentrated substrate
(fed-batch fermentation), eliminating downtime and reducing the process stream volume
(Ezeji et al., 2004b). Gas stripping and pervaporation have been proposed as the two most
promising separation technologies to be integrated with ABE fermentation for butanol
recovery. Gas stripping appears to be more energy-efficient and hassle-free, as
pervaporation requires membrane assistance (Vane, 2008). Stripping gas is bubbled in to
fermentation broth and captures volatile solvents in the broth, and the gas stream

127
containing solvents is sequentially passed through a condenser where the solvents are
condensed and enriched in the condensate stream. Gas stripping offers many advantages
as an integrated product recovery technology with fermentation, including utilization of
fermentation gases as stripping gas and the ability to operate under fermentation
temperature with optional solids removal from fermentation broth (Dürre, 1998; Ezji et
al., 2004a; Lee et al., 2008; Zheng et al., 2009) Gas stripping has been successfully
applied in various ABE fermentation processes, and enhanced butanol production and
reactor productivity have been reported (Ezeji et al., 2003; 2004b; 2005a; 2005b; 2007b;
Maddox et al., 1995; Qureshi and Blaschek, 2001b; Qureshi et al., 1992; 2007). Ezeji et
al. (2003) reported 75.9 g/L ABE were obtained in gas stripping integrated batch
fermentation utilizing 161.7 g/L glucose, while only 17.7 g/L ABE was obtained in the
control study without gas stripping using 44.6 g/L glucose. Highly concentrated substrate
(500 g/L glucose) was consumed in fed-batch fermentation with online butanol removal
by gas stripping, and 232.8 g/L ABE was produced with a productivity of 1.16 g/L. h,
compared with 17.6 g/L ABE in control study with a productivity of 0.29 g/L. h (Ezeji et
al., 2004b). In a continuous fermentation integrated with gas stripping, 1163 g/L glucose
was utilized and 460 g/L ABE were produced, with a productivity of 0.91 g/L. h (Ezeji et
al., 2005a).
Many factors affect the efficiency and performance of gas stripping, including gas
flow rate, contact time, surface area, temperature, presence of cells and components from

128
fermentation broth. Ezeji et al. (2005b) studied the effect of gas recycle rate and bubble
size on butanol recovery by gas stripping, and recommended a 0.5-5.0 mm bubble size to
be used in gas stripping process. In this study, the effect of cooling temperature, gas flow
rate, and presence of cells on gas stripping performance was evaluated with model
solutions and fermentation broth for process optimization. These studies will help
researchers to better understand the gas stripping process, and optimize the design to
integrate with ABE fermentation to obtain optimal butanol production.
4.2 Materials and methods
4.2.1 Experimental setup and process design
Figure 4.1 illustrates the experimental setup of gas stripping process in this study.
All gas stripping experiments were conducted in a cylindrical glass column (stripper, i.d.
50mm×300mm) integrated with water jacket at controlled fermentation temperature (36
oC) through a water bath (Fisher Scientific, Model 910, PA, USA). A model solution
containing 8 g/L acetone, 16 g/L butanol, 3g/L ethanol, 2 g/L acetic acid, and 2 g/L
butyric acid was used in this study, as well as fermentation broth obtained from using
synthetic P2 medium with a mutant strain of Clostridium beijerinckii ATCC 55025
(obtained and named C. beijerinckii JB200 in our research lab at the Ohio State
University). The fermentation broth contained ~9 g/L acetone, 18 g/L butanol, 1 g/L
ethanol, 5 g/L acetic acid, and 4 g/L butyric acid, with a cell concentration of ~ 4 g/L. The

129
working volume of the stripping was 500 ml, and 250 ml model solution or fermentation
broth was used in the stripper to leave some free headspaces for bubbling. Air was used
as stripping gas to simulate the fermentation gases (H2 and CO2) that would otherwise be
used in the integrated process. Air was bubbled into the stripper using a peristaltic pump,
capturing volatile components. Gas flow rate was measured and calculated by a timed
water-displacement method. The ABE vapor was then condensed in a coil condenser
(Pyrex, Graham condenser, 300 mm jacket, Fisher Scientific) using commercial coolant
(50% (v/v) ethylene glycol) circulated through the condenser by an isotemp refrigerated
circulator (Fisher Scientific, Model 910, PA, USA). The condensate was collected at the
bottom of the condenser using a 125 ml conical flask. Air, free of solvents, was then
recycled back through the peristaltic pump into the stripper to recover more solvents. The
entire process was a closed loop, preventing any loss into open air. Water loss due to
condensation during the stripping was not compensated. Samples were taken from the
stripper and condensate at intervals for analysis. Every time sample was taken, the
condensate was completely emptied in the conical flask.
4.2.2 Analytical methods
The fermentation products, acetone, butanol, ethanol, acetic acid, and butyric acid,
were measured with a Shimadzu GC-2014 gas chromatograph (GC) (Shimadzu,
Columbia, MD, USA) equipped with a flame ionization detector (FID) and a 30.0 m

130
fused silica column (0.25m film thickness and 0.25 mm ID, Stabilwax-DA). To reduce
the injection mechanic error margin, internal standard method was used to analyze the
concentration of products in the samples. Isobutanol and isobutyric acid were used as
internal standards for the solvent products and acid products present in the samples. An
internal standard buffer solution containing 0.5 g/L isobutanol, 0.1 g/L isobutyric acid,
and 1% phosphoric acid was used to dilute each sample 20 times for acidification and
calibration prior to analysis on GC. The gas chromatograph was operated at an injection
temperature of 200 oC with 1 μL of the acidified sample injected by the AOC-20i
Shimadzu auto injector. Column temperature was held at 80 oC for 3 min, raised to 150
oC at a rate of 30 oC/min, and held at 150 oC for 3.7 min.
Selectivity is calculated as α = [y/(1-y)]/[x/(1-x)], where x and y represent the
weight fractions of components in model solution/fermentation broth and condensate,
respectively.
4.3 Results and discussion
4.3.1 Effect of cooling temperature
The effect of cooling temperature on gas stripping was evaluated with model
solution containing ABE, acetic acid and butyric acid. 0 oC and below 0 oC (-5 oC) were
studied in this paper of their effect on solvent stripping rate and condensate concentration.
A flow rate of 1.25 L/min was used to strip solvents in the stripper. The results are shown

131
in Figure 4.1. The initial butanol concentration in the model solution was 18.1 g/L, and
was reduced to 4.7 g/L and 6.3 g/L after 8 hrs under -5 oC and 0 oC cooling conditions,
respectively. At the same time, the acetone concentration was reduced from initially 8.2
g/L to 5.5 g/L and 5.9 g/L under -5 oC and 0 oC cooling conditions, respectively. The
ethanol concentration was least affected by gas stripping under both temperature
scenarios, remaining almost unchanged throughout the process (decreased from 2.9 g/L to
2.2 g/L and 2.3 g/L, respectively, under -5 oC and 0 oC). The acid concentrations were not
affected by gas stripping, remaining unchanged during the process. However, the acids
were detected in the condensate at a concentration level below 1 g/L for both acetic acid
and butyric acid, whereas the ethanol concentration was detected between 7-10 g/L in the
condensate. This indicated that even though the concentration change seemed to be
similar for ethanol and acids, gas stripping was still highly selective towards solvent
instead of acids. It has been reported that acids were not removed from the fermentor
when gas stripping was integrated with an on-going ABE fermentation process (Ezeji et
al., 2003; 2004b). Our study supports the same statement since acids level remained
unchanged in the model solution in the stripper, albeit a very small amount of acids were
detected in the condensate.
The low cooling temperature increased the condenser efficiency, condensed and
recovered most of the solvent in the vapor phase, resulting in gas containing less residual
solvents to be recycled back into stripper for more complete solvent recovery. This is

132
illustrated in Figure 4.2a and 2b. The solvent concentration decreased more rapidly in the
stripper when using -5 oC in the condenser, and higher solvent removal rate was obtained
at -5 oC than at 0 oC under the same solvent concentration. Complete condensation of
alcohols from gas is not practical, since it will lead to complete condensation of water
(Vane, 2008). If condensation of a higher fraction of butanol is desired, a lower
temperature is needed which will also result in a higher percentage of water condensation.
The concentration of solvents in the condensate was concentration dependent, as shown
in Figure 4.4c. When the solvent concentration in the feed was high, the concentration of
recovered solvent in the condensate was high as well. It is also shown in Figure 4.4c that
at the same feed concentration, the cooling temperature of 0 oC resulted in higher solvent
concentration than at -5 oC, which was the result of excessive water condensation under
lower cooling temperature. In order to avoid water condensation and obtain highly
concentrated solvent in the condensate, fractional condensation with multi-stage has been
proposed to first knock out bulk water and then recover the solvent at a higher
concentration in the condensate instead of using single stage condensation process
(Taylor et al., 2000). The concentration in the condensate for butanol, acetone, and
ethanol, was in the range of 46.8-165.7 g/L, 19.6-36.8 g/L, and 7.16-11.3 g/L,
respectively, depending on the concentration of each solvent present in the feed (Figure
4.2c).
The butanol saturation point in the water is about 8%; when present at higher

133
concentration, butanol will be separated from water by natural phase separation. This was
observed in our study as well. When the butanol concentration reached over 90 g/L in the
condensate, two phase separation was clearly observed with butanol and acetone on the
top organic phase. This usually happened when the butanol concentration in the feed was
over 8 g/L. As the butanol concentration continued to decrease in the feed, more water
was taken out by stripping gas as opposed to butanol, resulting in more water
condensation and low butanol concentration in the condensate.
4.3.2 Effect of gas flow rate
Two gas flow rates, 1.0 L/min and 1.25 L/min were studied using 250 ml model
solution in the stripper. Feed temperature in the stripper was at 36 oC, and cooling
temperature was controlled at 0 oC. The results are shown in Figure 4.3. Butanol was
rapidly reduced from 17.8 g/L to 6.5 g/L and 4.7 g/L within 10 hrs, respectively, with the
flow rate of 1.0 L/min and 1.25 L/min (Figure 4.3a). Butanol removal rate was found to
be concentration dependent, and was within the range of 0.5-3.0 g/L. h, depending on the
feed concentration. As shown in Figure 4.3b, higher flow rate resulted in higher butanol
and acetone removal rate under the same feed concentration. This was because at high
flow rate, more stripping gas was bubbled into stripper and the gas-liquid contact areas
were increased within the same unit of time. Ezeji et al. (2005b) defined stripping rate as
the following:

134
Rs = KsaCs
Rs was the stripping rate, Cs was the solvent concentration in the aqueous phase, and
Ksa was the stripping rate constant (“a” being the interfacial area). Stripping rate was
determined by both concentration and stripping rate constant. In order to increase Ksa,
either small bubbles with the same gas flow rate or increased gas flow rates could be used.
Ezeji et al. (2005b) tested two bubble deliver systems at two different flow rates, and
obtained higher Ksa at higher flow rate regardless of bubble sizes. In addition to the
increased gas volume by high flow rate, the added turbulence also attributed to the
increased Ksa, resulting in a 2.51-fold increase in Ksa with only 1.86-fold increase in gas
flow rate. Therefore, the relation between gas flow rate and butanol removal rate was not
linear due to the positive impact of turbulence in the flow. As shown in Figure 4.3b, a
25% increase in flow rate resulted in higher butanol and acetone removal rate, especially
at low feed concentrations. With a butanol concentration increase from 10 g/L to 15 g/L
in the feed, butanol removal rate could be increased from 0.8 g/L h to 2.4 g/L h with 1.0
L/min gas flow rate. High flow rate not only increased butanol and acetone removal, but
also facilitated water removal. When more solvents were transferred into gas phase, more
water was taken out by stripping gas as well due to the enhanced mass transfer as a result
of increased interfacial contact area. This was confirmed and shown in Figure 4.3c. The
concentration of solvents in the condensate was lower with high flow rate than obtained
with low flow rate. When solvent concentration in the feed was low, this effect was not as

135
obvious as water overweighed solvents although the solvent removal rate was higher at
high flow rate.
4.3.3 Effect of cells and components from fermentation broth
Fermentation broth contains cell bodies, which can increase the viscosity of the
solution and change how the solution behaves. In addition to cells, other components
such as proteins, sugars, and salts all affect the properties of broth and can impact on gas
stripping performance as compared with simply model solution. In order to understand if
gas stripping pattern will be affected, fermentation broth obtained from C. beijerinckii
JB200 with glucose-P2 medium was evaluated in this study. Fermentation broths with
and without cells (removed by centrifugation) were both studied in order to investigate
the effect of cell presence and other fermentation components. Due to the presence of
cells and proteins, which affected the surface properties of the broth, antifoam was added
when necessary to prevent excess bubbling. The cooling temperature was kept at 0 oC,
and gas flow rate was maintained at 1.25 L/min with 250 ml broth in the stripper. The
results are shown in Figure 4.4.
Butanol concentration in the stripper was rapidly reduced from the initial 18.5 g/L to
5.0 g/L and 5.5 g/L, respectively, for broths with cells and without cells. The fermentation
broth without cells was a light yellowish clear liquid, and the collected condensate in this
study was a clear colorless solution for broth with or without cells, indicating that gas

136
stripping did not take cells or any other components in the broth. Figure 4.4a shows that
the concentration change of acetone, butanol and ethanol in the feed did not have any
significant difference due to the presence of cells. When compared with results from
model solution (Figure 4.4b), butanol removal rate was not significantly affected by the
presence of cells or other components present in the broth. This result was different from
reported in the literature (Ezeji et al., 2003). ABE solution with 11 g/L cells was studied
and compared with ABE model solution without cells, and the presence of cells was
found to have an adverse impact on butanol removal rate at high butanol concentration
(above 7.5 g/L)(Ezeji et al., 2003). This was probably due to the difference of cell
concentration present in the broth. In our study, the broth was obtained from C.
beijerinckii JB 200, which was a mutant strain of C. beijerinckii ATCC 55025. The
maximum cell concentration in the course of batch fermentation was around 4 g/L, which
corresponded to an 8 – 9 optical density at 600nm UV ray. This cell concentration was
much lower than used by Ezeji et al. (2003), which was an 11 g/L cells. The high cell
concentration can greatly affect the behavior of broth due to the increased viscosity,
resulting in different outcome in stripping performance.
Even though the presence of cell did not seem to affect the butanol removal rate in
this study, acetone removal rate was different with the fermentation broth and model
solution. As shown in Figure 4.2b, 3b and 4b, acetone removal rate was concentration
dependent in model solutions, whereas acetone removal rate remained almost constant

137
(0.5 g/L. h) regardless of acetone concentration in fermentation broth with or without
cells. The concentration of acetone and butanol in the condensate was also concentration
dependent with fermentation broth, and found to be higher than obtained with model
solution at the same concentration (Figure 4.4c). This effect was very obvious for butanol
concentration in the condensate, especially at high butanol concentration in the feed.
Acetone concentration remained between 30 – 40 g/L in the condensate, regardless of
acetone concentration in the feed solution or type of solutions used. Butanol
concentration varied dramatically in the condensate, from 50 g/L to 250 g/L depending on
the feed concentration. Even at the same feed concentration (15 g/L), butanol
concentration in the condensate was found to be at 248.7 g/L, 203.6 g/L and 155.5 g/L,
for fermentation broth without cells, fermentation broth and model solution, respectively.
This indicated that fermentation broth was in favor of butanol concentration in the
condensate compared with model solution. The fermentation broth contained ~4 g/L
acetic acid and butyric acid, and 0.5-1.0 g/L acetic acid and butyric acid were detected in
the condensate.
4.3.4 Selectivity of acetone, butanol, and ethanol
In this study, five conditions were evaluated including temperature, gas flow rate,
and the presence of cells and other fermentation components. Model solution at T = 0 oC,
flow rate = 1.0 L/min, model solution at T = 0 oC, flow rate = 1.25 L/min, model solution

138
at T = -5 oC, flow rate = 1.25 L/min, fermentation broth with cells at T = 0 oC, flow rate =
1.25 L/min, and fermentation broth without cells at T = 0 oC, flow rate = 1.25 L/min,
were defined as condition 1-5, respectively. The acetone, butanol and ethanol selectivity
under these conditions are shown in Figure 4.5. From condition 1 to 5, butanol selectivity
was in the range of 12.0-14.5, 9.0-14.5, 9.6-18.0, 7.4-17.6, and 10.6-21.4, respectively.
Comparing results from condition 1 to 3 with model solution, cooling temperature had a
more significant effect on increasing butanol selectivity than gas flow rate (Figure 4.5a).
When butanol concentration was between 5-12 g/L, butanol selectivity at T = -5 oC was
15.7-18.0, compared with 12.1-14.5 at T = 0 oC under the same flow rate of 1.25 L/min.
Comparing the results in condition 4 and 5 with fermentation broth, the presence of cells
adversely affected the butanol selectivity. Ezeji et al. (2003) also reported that in the
presence of 11 g/L cells, butanol selectivity was significantly reduced as compared with
model solution free of cells. Fermentation broth without cells resulted in the best butanol
selectivity at high concentration (10-15 g/L), indicating that the fermentation components
had positive effect on butanol selectivity compared with simple model solution. When the
butanol concentration was low (less than 10 g/L), butanol selectivity was mostly in the
range of 11- 15 at T = 0 oC regardless of other conditions, with only a few outliers.
Acetone selectivity was within a narrow range of 3.0-4.5 regardless of the conditions
evaluated. Compared with butanol selectivity, which was on average at 11-15, gas
stripping was highly selective towards butanol over acetone. Acetone selectivity was not

139
greatly affected by the concentration, remaining almost constant under various
concentrations as shown in Figure 4.5b. Ethanol selectivity was in the same narrow range
as acetone, mostly in between 3.0 to 4.0 as shown in Figure 4.5c. The ethanol
concentration in the fermentation broth was around 0.5 g/L, which was lower than the
ethanol concentration in the model solution. Similar ethanol selectivity was obtained in
broth and model solution, regardless of ethanol concentration. This indicated that butanol
selectivity was more susceptible to the conditions employed, whereas acetone and ethanol
selectivity remained almost unchanged under most of the conditions evaluated in this
study. Except that all the butanol selectivity data scattered within a defined range, no
other clear trend was observed under the conditions evaluated. Ezeji et al. (2003) reported
that butanol and ethanol selectivity followed no clear trend, whereas acetone selectivity
followed a straight line in their study. The reported butanol selectivity was 13.83-10.26,
which was similar to our study. The acetone and ethanol selectivity was reported to be in
a wider range in their study, 4.12-6.42 and 4.9-7.9, respectively. Cooling temperature
appeared to have the most positive impact, as the selectivity of acetone, butanol and
ethanol were all higher under -5 oC than under 0 oC (Figure 4.5). Condensation
temperature from -60 oC to 4 oC have been employed in gas stripping at different ABE
fermentation processes, and butanol selectivity from 6 to 30.5 were reported (Ennis et al.,
1986; Groot et al., 1989; Maddox et al., 1995; Qureshi and Maddox, 1991; Qureshi et al.,
1992).

140
4.4 Conclusion
The effect of condensation temperature, gas flow rate, and presence of cells and
other fermentation components on gas stripping were evaluated in this study. Low
condensation temperature was found to have a positive effect on butanol removal rate and
a negative effect on condensate concentration. High gas flow rate not only facilitated
butanol removal, but also increased water removal, which negatively impacted
condensate concentration. Butanol and acetone removal rates of 0.5 – 3.5 g/L. h and 0.1 –
0.5 g/L. h were obtained in this study under various conditions evaluated. Condensate
concentration was found to be concentration dependent, and high butanol concentration
in the condensate was obtained with fermentation broth without cells, followed by broth
with cells and model solution, indicating that the fermentation components had a positive
effect on butanol condensate concentration. Butanol selectivity of 7.4 – 21.4 was obtained
in this study under various conditions tested, with an average range of 11.0 – 15.0. The
presence of cells had no significant effect on butanol removal rate, but adversely
impacted butanol selectivity. Acetone and ethanol selectivity was 3.0 – 4.5 and 3.0 – 4.0,
respectively, indicating that gas stripping was highly selective towards butanol removal.
4.5 References
Dürre, P. (1998). New insights and novel developments in clostridial acetone/ butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Dürre, P. (2007). Biobutanol: An attractive biofuel. Biotechnol. J., 2, 1525-1534.

141
Ennis, B., C.T. Marshall, I.S. Maddox, A.H.J. Paterson (1986). Continuous product recovery by in-situ gas stripping/condensation during solvent production from whey permeate using Clostridium acetobutylicum. Biotechnol. Lett., 8, 725-730.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2003). Production of butanol by Clostridium beijerinckii BA101 and in-situ recovery by gas stripping. J. Microbiol. Biotechnol., 19, 595-603.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004a). Butanol fermentation research: upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004b). Acetone-butanol-ethanol production from concentrated substrate: reduction in substrate inhibition by fed-batch technique and product inhibition by gas stripping. Appl. Microbiol. Biotechnol., 63, 653-658.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2005a). Process for continuous solvent production. United States Patent Application Publication, US patent 20050089979A1.
Ezeji, T.C., P.M. Karcher, N. Qureshi, H.P. Blaschek (2005b). Improving performance of a gas stripping-based recovery system to remove butanol from Clostridium beijerinckii fermentation. Bioprocess Biosyst. Eng., 27, 207-214.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007a). Bioproduction of butanol from biomass: from genes to bioreactors. Current Opinion in Biotechnol., 18, 220-227.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007b). Production of acetone butanol (AB) from liquefied corn starch, a commercial substrate, using Clostridium beijerinckii coupled with product recovery by gas stripping. J Ind. Microbiol. Biotechnol., 34, 771-777.
Ezeji, T.C., C. Milne, N.D. Price, H.P. Blaschek (2010). Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol., 85, 1697-1712.
Groot, W.J., R.G.J.M. van der Lans, K.Ch.A.M. Luyben (1989). Batch and continuous butanol fermentations with free cells: integration with product recovery by gas- stripping. Appl. Microbiol. Biotechnol., 32, 305-308.
Jones, D.T. and D. Woods (1986). Acetone-butanol fermentation revisted. Microbiol.

142
Reviews, 50, 484-524.
Kumar, M. and K. Gayen (2011). Developments in biobutanol production: New insights. Appl. Ener., 88, 1999-2012.
Lee, S.T., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentive butanol production by Clostridia. Biotechnol. Bioeng., 101,209-228.
Maddox, I.S., N. Qureshi and K. Roberts-Thomson (1995). Production of acetone-butanol-ethanol from concentrated substrates using Clostridium acetobutylicum in an integrated fermentation-product removal process. Process Biochemistry, 30, 209-215.
Qureshi, N. and I.S. Maddox (1991). Integration of continuous production and recovery of solvents from whey permeate: use of immobilized cells of Clostridium acetobutylicum in a fluidized bed reactor coupled with gas stripping. Bioproc. Eng., 6, 63-69.
Qureshi, N., I.S. Maddox, A. Friedl (1992). Application of continuous substrate feeding to the ABE fermentation: relief of product inhibition using extraction, perstraction, stripping and pervaporation. Biotechnol. Prog., 8, 382-390.
Qureshi, N. and H.P. Blaschek (2001a). Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 27, 287-291.
Qureshi, N. and H.P. Blaschek (2001b). Recovery of butanol from fermentation broth by gas stripping. Renewable Energy, 22, 557-564.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst. Eng., 30, 419-427.
Qureshi, N. and T.C. Ezeji (2008). Butanol, ‘a superior biofuel’ production from agricultural residues (renewable biomass): recent progress in technology. Biofuels, Bioprod. Bioref., 2, 319-330.
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuls, Bioprod. Bioref., 2, 553-588.
Taylor, F., M.J. Kurantz, N. Goldberg, A.J. McAloon, J.C. Craig, Jr. (2000). Dry-grind

143
process for fuel ethanol by continuous fermentation and stripping. Biotechnol. Prog., 16, 541-547.
Zheng, Y.N., L.Z. Li, M. Xian, Y.J. Ma, J.M. Yang, X. Xu, D.Z. He (2009). Problems with the microbial production of butanol. J. Ind. Microbiol. Biotechnol., 36, 1127-1138.

Waterbath
Str
ippe
r
Con
den
ser
Gas recycle
Gas recycle
Peristaltic pump
Model solutionor fermentation broth
Coo
lant
circ
ula
tion
Condensate
Figure 4.1 Schematic diagram of the gas stripping process
144
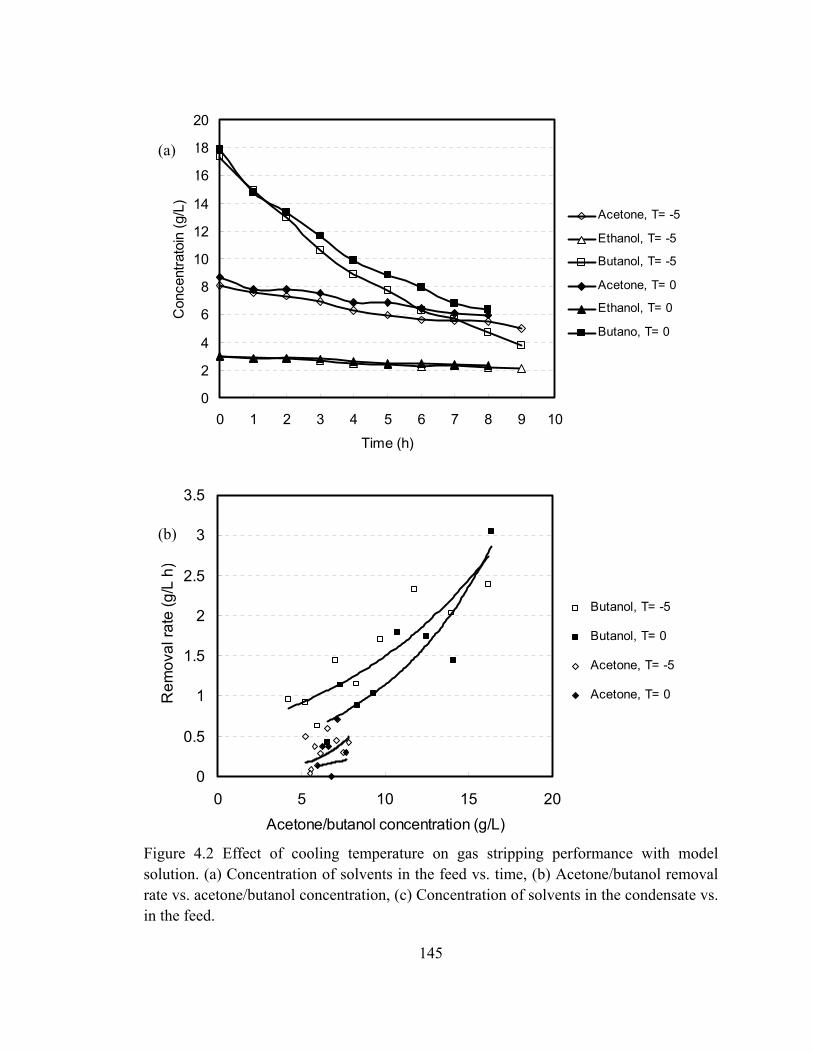
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8 9 10
Time (h)
Con
cent
rato
in (
g/L) Acetone, T= -5
Ethanol, T= -5
Butanol, T= -5
Acetone, T= 0
Ethanol, T= 0
Butano, T= 0
0
0.5
1
1.5
2
2.5
3
3.5
0 5 10 15 20
Acetone/butanol concentration (g/L)
Re
mo
val r
ate
(g
/L h
)
Butanol, T= -5
Butanol, T= 0
Acetone, T= -5
Acetone, T= 0
(a)
(b)
Figure 4.2 Effect of cooling temperature on gas stripping performance with model solution. (a) Concentration of solvents in the feed vs. time, (b) Acetone/butanol removal rate vs. acetone/butanol concentration, (c) Concentration of solvents in the condensate vs. in the feed.
145

Figure 4.2 continued
0
20
40
60
80
100
120
140
160
180
200
0 5 10 15 20
Concentration in feed (g/L)
Co
nce
ntr
atio
n in
co
nd
en
sate
(g
/L)
Butanol, T= 0
Butanol, T= -5
Acetone, T= 0
Acetone, T= -5
Ethanol, T= 0
Ethanol, T= -5
(c)
146

0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8 9 10 11
Time (h)
Co
nce
ntr
atio
n (
g/L
)
Acetone, 1 L/min
Ethano, 1 L/min
Butanol,1 L/min"
Acetone, 1.25 L/min
Ethanol, 1.25 L/min
Butanol, 1.25 L/min
(a)
0
0.5
1
1.5
2
2.5
3
3.5
0 5 10 15 20
Concentration (g/L)
Re
mo
val r
ate
(g
/L h
)
Butanol, 1 L/min
Acetone, 1 L/min
Butanol, 1.25 L/min
Acetone, 1.25 L/min
(b)
Figure 4.3 Effect of gas flow rate on gas stripping performance with model solution. (a) Concentration of solvents in the feed vs. time, (b) Acetone/butanol removal rate vs. acetone/butanol concentration, (c) Concentration of solvents in the condensate vs. in the feed.
147

Figure 4.3 continued
0
20
40
60
80
100
120
140
160
180
200
0 5 10 15 20
Feed concentration (g/L)
Co
nd
ens
ate
co
nce
ntr
atio
n (g
/L)
Butanol, 1 L/min
Acetone, 1 L/min
Butanol, 1.25 L/min
Acetone, 1.25 L/min
(c)
148
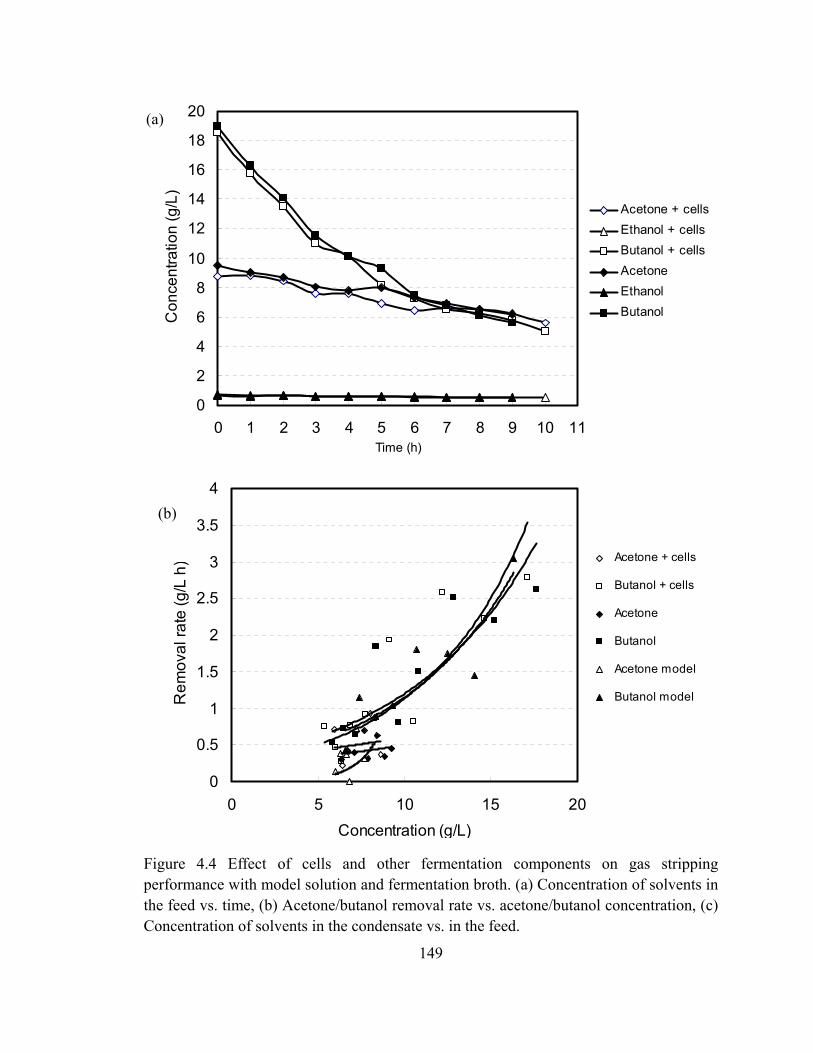
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8 9 10 11Time (h)
Co
nce
ntr
atio
n (
g/L
)
Acetone + cells
Ethanol + cells
Butanol + cells
Acetone
Ethanol
Butanol
(a)
0
0.5
1
1.5
2
2.5
3
3.5
4
0 5 10 15 20
Concentration (g/L)
Re
mo
val r
ate
(g
/L h
) Acetone + cells
Butanol + cells
Acetone
Butanol
Acetone model
Butanol model
(b)
Figure 4.4 Effect of cells and other fermentation components on gas stripping performance with model solution and fermentation broth. (a) Concentration of solvents in the feed vs. time, (b) Acetone/butanol removal rate vs. acetone/butanol concentration, (c) Concentration of solvents in the condensate vs. in the feed.
149

Figure 4.4 continued
0
50
100
150
200
250
300
0 5 10 15 20
Feed concentration (g/L)
Co
nd
en
sate
co
nce
ntr
atio
n (
g/L
)
Butanol + cells
Acetone + cells
Butanol
Acetone
Butanol model
Acetone model
(c)
150

5
7
9
11
13
15
17
19
21
23
0 5 10 15 20
Butanol concentration (g/L)
Bu
tan
ol s
ele
ctiv
ityModel solution, T=0, 1 L/min
Model solution, T=0, 1.25 L/min
Model solution,T=-5,1.25 L/min
Broth, T=0, 1.25 L/min
Broth w/o cells, T=0, 1.25 L/min
(a)
2
2.5
3
3.5
4
4.5
5
2 4 6 8 10
Acetone concentration (g/L)
Ace
ton
e s
ele
ctiv
ity
Model solution, T=0, 1 L/min
Model solution, T=0, 1.25 L/min
Model solution,T=-5,1.25 L/min
Broth, T=0, 1.25 L/min
Broth w/o cells, T=0, 1.25 L/min
(b)
Figure 4.5 Selectivity of solvents under the conditions evaluated in this study. (a) Butanol selectivity vs. concentration, (b) Acetone selectivity vs. concentration, (c) Ethanol selectivity vs. concentration.
151

Figure 4.5 continued
2
2.5
3
3.5
4
4.5
5
0 0.5 1 1.5 2 2.5 3 3.5
Ethanol concentration (g/L)
Eth
an
ol s
ele
ctiv
ity
Model solution, T=0, 1 L/min
Model solution, T=0, 1.25 L/min
Model solution,T=-5,1.25 L/min
Broth, T=0, 1.25 L/min
Broth w/o cells, T=0, 1.25 L/min
(c)
152

153
Chapter 5: Fed-batch Fermentation for Butanol Production from Cassava Bagasse
Hydrolysate in a Fibrous Bed Bioreactor with Continuous Gas Stripping
Abstract
The enzymatic hydrolysate of cassava bagasse as a potential economic feedstock for
acetone-butanol-ethanol (ABE) fermentation was studied with a hyper-butanol-producing
Clostridium beijerinckii strain in a fibrous bed bioreactor. About 33.9 g/L ABE were
produced from cassava bagasse hydrolysate (CBH) and glucose in batch fermentation.
Concentrated CBH containing 584.4 g/L glucose was used in fed-batch fermentation with
gas stripping for continuous butanol recovery. Nutrient supplementation was investigated
to evaluate its effect on the long-term operational stability of the fed-batch fermentation.
With periodical nutrient supplementation, 108.5 g/L ABE, of which 76.44 g/L was
butanol, was produced over 283 h with an average sugar consumption rate of 1.28 g/L·h.
The overall ABE and butanol yields were 0.35±0.03 g/g and 0.23±0.01 g/g, respectively,
whereas the overall ABE and butanol productivities were 0.47±0.06 g/ L·h and 0.32±0.03
g/L·h, respectively. The productivities in the fed-batch fermentation process using CBH
were slightly lower than those from the batch system with glucose as the substrate

154
probably due to the accumulation of cell metabolites and inhibitors present in the highly
concentrated CBH. With gas stripping, concentrated butanol of ~100 g/L was produced in
the process, allowing for more energy-efficient purification of butanol in subsequent
distillation.
5.1 Introduction
Acetone-butanol-ethanol fermentation, which is also known as ABE fermentation, is
one of the oldest fermentations in human history which can be traced to 1861 (Jones and
Woods, 1986). Butanol is not only an important industrial solvent, but also a superior
biofuel candidate to ethanol. Compared to ethanol, butanol has a higher energy content,
lower vapor pressure, and a similar air-to-fuel ratio to gasoline. More importantly,
butanol is compatible with the current automobile engine design and the transportation
pipeline, making butanol a perfect candidate to replace gasoline. With unstable crude oil
supplies and prices in the world market, producing green butanol from biomass through
improved ABE fermentation and novel butanol recovery techniques is the future (Ezeji et
al., 2004a; 2007a; Dürre, 1998; Lee et al., 2008).
Clostridia bacteria commonly used in ABE fermentation are able to ferment a variety
of sugars, including glucose, xylose, arabinose, cellobiose, and mannose (Ezeji and
Blascheck, 2008). Corn starch and molasses are considered uneconomical for biofuels
production due to the limited supply and high costs of these food-based substrates, which

155
may comprise more than 50% of the product cost. Therefore, alternative lignocellulosic
feedstocks, including agricultural residues, forestry wastes and energy crops, will be the
main feedstocks for the biorefinery industry (Kumar et al., 2009). ABE fermentation
using lignocellulosic feedstocks, including corn fiber, wheat straw and dried distillers’
grains and solubles, have been studied (Ezeji and Blaschek, 2008; Qureshi et al., 2007;
Qureshi et al., 2008). In this work, cassava bagasse was studied as a potential feedstock
for biobutanol production. Cassava is an important food source in many countries and
regions, including China, Thailand, and Latin America (Pandey et al., 2000). Cassava
bagasse, the fibrous residue from industrial processing of cassava for starch extraction, is
generated in large quantities in these countries and treated as solid waste because bagasse
can be used only as low-value animal feed or must be disposed into landfills (Pandey et
al., 2000). Biological conversion of cassava bagasse has been previously reported for
organic acids and aroma compounds production (Bramorski et al., 1998; Carta et al.,
1999; Thongchul et al., 2009), but never reported for butanol production. Utilizing
cassava bagasse for butanol production not only lowers substrate costs, but also adds
value to the cassava processing industry, reducing environmental pollution caused by
bagasse disposal.
Besides using renewable lignocellulosic substrates, several advanced fermentation
technologies have been developed to improve the ABE fermentation. Fed-batch
fermentation offers a number of advantages compared to other fermentation modes,

156
including the utilization of highly concentrated substrate, which can reduce the reactor
volume and wastewater generated in the process (Ezeji et al., 2004a; 2004b; 2005; 2007a;
Qureshi and Blaschek, 2001a). However, due to end product (butanol) inhibition,
fed-batch fermentation can not be effectively operated without simultaneous product
removal (Ezeji et al., 2004b; 2005). Among all the product recovery techniques, gas
stripping is an economic and favorable choice due to its operation simplicity. Gas
stripping selectively removes volatile substances such as butanol and acetone, and does
not strip nutrients out or harm the cells in the fermentation (Durre, 1998). Gas stripping
has been successfully applied in ABE fermentation to increase butanol production by
relieving the product inhibition (Ezeji et al., 2003; 2007b; Qureshi et al., 2007).
The goal of this study was to evaluate the feasibility of producing butanol from
enzymatic hydrolysate of cassava bagasse as an alternative carbon source in ABE
fermentation. The batch fermentation kinetics with cassava bagasse hydrolysate (CBH)
was first investigated, and then a fed-batch fermentation process using highly
concentrated CBH was studied with simultaneous product removal by gas stripping. The
results showed the integrated fermentation-gas stripping process was effective in
converting sugars, mainly glucose, in concentrated CBH to butanol.
5.2 Materials and methods
5.2.1 Enzymatic hydrolysis of cassava bagasse

157
Cassava bagasse, obtained from a cassava-processing factory in Guangdong, China,
was dried and mechanically milled to fine powder (about 50 to 100 μm in diameter).
Before enzymatic hydrolysis, 10 g dried cassava bagasse powder were mixed with 90 ml
water (corresponding to a 10% (w/w) solid loading) and autoclaved at 121oC and 15 psig
for 30 min. Then, commercial glucoamylase (Distillase L-400, activity: 350 GAU/g,
specific gravity: 1.13 to 1.15 g/ml, Genencor, NY, USA) was added at a 0.06% (w/w)
loading (kg glucoamylase per kg cassava bagasse on a dry solids basis) to hydrolyze the
cooked starch content into glucose. This enzymatic hydrolysis process was operated at 65
oC, pH 4.2 and 200 rpm for 24 h. Then cellulase (Accellerase 1500, endoglucanase
activity: 2200-2800 CMC U/g, -glucosidase activity: 525-775 pNPG U/g, Genencor, NY,
USA) was added into the mixture at a loading of 0.1 ml/g dry cassava bagasse to
hydrolyze the remaining cellulose content into more glucose. This process was operated
at 50oC, pH 5.0, and 200 rpm for 24 hours. Sterile HCl and NaOH solutions were used for
all pH adjustments during these two hydrolysis processes. After the hydrolysis process
was complete, the mixture was centrifuged at 7000 rpm for 10 min to remove the
insolubles, and the clear liquid, cassava bagasse hydrolysate (CBH), was used as the
carbon source in ABE fermentation. Concentrated CBH (>500 g/L sugars) was prepared
by evaporation using a rotary evaporator under vacuum at 80oC and used in fed-batch
fermentation. The compositions of CBH and concentrated CBH are shown in Table 5.1.

158
5.2.2 Strain and medium preparation
A hyper butanol producing mutant strain Clostridium beijerinckii JB 200 derived
from ATCC 55025 through mutagenesis and adaptation in a fibrous bed bioreactor was
used in this study. The stock culture of this mutant strain was stored in a 15% glycerol-P2
stock solution in a -80oC freezer. To prepare the seed inoculum for fermentation studies, 2
ml of the glycerol stock culture was inoculated into 100 ml of tryptone-yeast
extract-glucose growth medium in a rubber-capped serum bottle, and incubated
anaerobically at 37oC for 12-15 h until cells were highly active.
Unless otherwise noted, ABE fermentation was studied using the P2 medium
containing carbon source (glucose or CBH), yeast extract (1 g/L), phosphate buffer (0.5
g/L KH2PO4 and 0.5 g/L K2HPO4), ammonium acetate (2.2 g/L), vitamins (1 mg/L
para-amino-benzoic acid (PABA), 1 mg/L thiamin and 0.01 mg/L biotin), and mineral
salts (0.2 g/L MgSO4·7H20, 0.01 g/L MnSO4· H20, 0.01 g/L FeSO4·7H20, 0.01 g/L NaCl).
For batch fermentation with CBH as the carbon source on the bioreactor, 955 ml of
CBH were mixed with nitrogen source (1 g yeast extract and 2 g ammonium acetate in 20
ml H2O), phosphate buffer (in 20 ml H2O), minerals (200-fold concentrated, 5 ml) and
vitamins (1000-fold concentrated, 1 ml) to make 1.0 L CBH-based P2 medium. CBH,
nitrogen source and buffer were autoclaved separately to avoid chemical reactions at
121oC and 15 psig for 30 minutes for sterilization. Minerals and vitamins were prepared
at 200-fold and 1000-fold concentration, respectively, and were filter-sterilized through

159
sterile membrane filters (25 mm, 0.2 μm syringe filter, Fisher, NJ, USA) to avoid
oxidation and destruction of vitamins. After minerals and vitamins were aseptically added
into fermentation medium, the medium was nitrogen purged for 1 h through a sterile 0.2
μm vent filter unit (Millex, Millipore Corp., MA, USA) to ensure an anaerobic condition
in the system.
5.2.3 Experimental setup
Figure 5.1 illustrates the experimental setup used for integrated fed-batch
fermentation with gas stripping, which consisted of three parts: a fibrous bed bioreactor
(FBB) for immobilized-cell fermentation, a 1.0 L three-necked spinner flask (Bellco)
with temperature and pH control, and a Pyrex Graham coil condenser (Fisher Scientific,
water jacket 300mm) for vapor condensation. The FBB was made of a glass column
packed with spiral wound cotton towel and stainless steel wire cloth with a working
volume of ~400 ml. Detailed description of the FBB construction can be found elsewhere
(Silva and Yang, 1995; Yang, 1996). The spinner flask, FBB and condenser were
autoclaved separately for 45 min, and aseptically connected after sterilization. The whole
system was sparged with nitrogen to ensure an oxygen-free environment. For batch
fermentation without gas stripping, the system was operated without connecting to the
condenser.

160
5.2.4 Cell immobilization in fibrous bed bioreactor
Before inoculation, the spinner flask containing 1 L P2 medium and the FBB were
sparged with nitrogen for 1-2 h until oxygen-free. Unless otherwise noted, the
fermentation system was maintained at 37oC, the spinner flask was agitated at 150 rpm,
and the medium pH was controlled at 5.0 by adding 6 N NaOH. Actively growing cells
(12-16 h) were inoculated into the spinner flask containing the P2 glucose medium at 5%
(v/v), and 24-36 h were allowed for growth until the OD600 reached over 6.0. Cell
immobilization was then carried out by circulating the broth into the FBB, allowing cells
to attach to the fibrous matrix through adsorption. After 24-36 h circulation, the cell
density in the broth no longer decreased and most of the cells were immobilized onto the
FBB. The old broth was then drained and replaced with a fresh P2 glucose medium to
allow the cells in the FBB to continue to grow. Again the old medium was changed and
the process was repeated several times until a stable and high cell density in the FBB was
achieved. Once cell immobilization was complete, batch and fed-batch fermentation
studies were carried out with cells in the FBB. The viability of the immobilized cells was
ensured through active adsorption and desorption, and immobilization helped cells to
adapt and survive in a stressful environment (Yang, 1996).
5.2.5 Batch and integrated fed-batch fermentations
For batch operation, fresh medium was put into the fermentor using a peristaltic

161
pump to start each experiment. When one batch was done, the reactor was completely
drained and fresh medium was put in again using the cells immobilized in FBB as seed
culture. For fed-batch operation, cassava bagasse hydrolysate with additional glucose
was used to initiate the fermentation. Fermentation was allowed to proceed for ~30 h
until the butanol concentration reached 3-4 g/L. Gas stripping was then initiated by
recycling fermentation gas (H2 and CO2, 1.0 L/min) through the fermentor using a
peristaltic pump (Masterflex, L/S, standard drive, Cole-Parmer, IL, USA) at 1.25 L/min.
The gas stream containing volatile substances was then cooled in the coil condenser at
1oC using a refrigerated circulator (Fisher Scientific, Isotemp Refrigerated Circulator,
Model 910, PA, USA). The condensate was collected at the bottom of the condenser
using a conical flask (cooling trap). The gas stripping cycle was a closed loop to prevent
any loss into open air. Samples were taken from the reactor at intervals for analysis of
sugar consumption and ABE production. Recovered stream (condensate) was collected at
intervals (end of each fermentation cycle) to evaluate the product yield, titer and
productivity of each cycle. Based on the results of sugar consumption, concentrated CBH
was added into the fermentor at intervals to increase sugar level and to compensate for
the water loss due to gas stripping. Other than the study of the effect of nutrient addition,
nutrient was only provided at the beginning of the fermentation (P2 formula). For the
study of the effect of nutrient addition on fermentation performance, yeast extract was
periodically added to the reactor. No additional minerals, vitamins, or buffer were added.

162
Concentrated CBH and nutrient solution were autoclaved for 30 min for sterilization and
then purged with nitrogen to oxygen-free before adding to the reactor.
5.2.6 Analytical methods
The sugars present in the CBH and fermentation broth were measured by a high
performance liquid chromatography (HPLC) with an organic acid column (Bio-Rad
HPX-87, ion exclusion organic acid column, 300 mm × 7.8mm). Samples were
centrifuged at 13.2 g for 5 min in microcentrifuge tubes and diluted 10 times with
distilled water prior to analysis on HPLC. HPLC was run at 45 oC using 0.01N H2SO4 as
the eluent at a flow rate of 0.6 ml/min. 15 μL sample was injected by an automatic
injector (SIL-10Ai) and the running time was set at 36 min. A refractive index (RI)
detector (Shimadzu RID-10A) was set at the range of 200 to detect the organic
compounds in the sample. The HPLC column was installed in a column oven (CTO-10A)
with temperature control at 45 oC. Peak height was used to calculate concentration of
sugars in the sample based on the peak height of standard sample. The glucose
concentration was also measured with a glucose analyzer (YSI 2700 Select, Yellow
Spring, OH).
The fermentation products, acetone, butanol, ethanol, acetic acid, and butyric acid were
measured with a Shimadzu GC-2014 gas chromatograph (GC) (Shimadzu, Columbia,
MD, USA) equipped with a flame ionization detector (FID) and a 30.0 m fused silica

163
column (0.25m film thickness and 0.25 mm ID, Stabilwax-DA). The gas chromatograph
was operated at an injection temperature of 200 oC with 1 μL of the acidified sample
injected by the AOC-20i Shimadzu auto injector. Column temperature was held at 80 oC
for 3 min, raised to 150 oC at a rate of 30 oC/min, and held at 150 oC for 3.7 min.
5.3. Results and discussion
5.3.1 Batch fermentation kinetics
Batch fermentation using glucose-based P2 medium was used as a control in this
study. From Figure 5.2A, 88.4 g/L initial glucose was used within 54 h, and 20.4 g/L
butanol (total ABE of 33.8 g/L) was produced in the broth. The butanol yield was 0.23
g/g, and the productivity was 0.38 g/L. h. The total ABE yield was 0.38 g/g, and the
productivity was 0.625 g/L. h. The Acetone-Butanol ratio was at ~1:2, which was typical
for Clostridia bacteria. At the end of fermentation, there was 2.58 g/L acetic acid and
1.50 g/L butyric acid left, and the low acids level indicated that the shift from acidogensis
to solventogensis was successful. Most of the Clostridia bacteria typically produced
~10-13 g/L butanol in batch system (Durre, 1998), and this mutant doubled the butanol
production. It was reported before that Clostridium beijerinckii BA101 was a
hyper-butanol producing mutant, yielding 19 g/L butanol and 29 g/L total ABE in the
batch system (Qureshi and Blaschek, 2001b; Ezeji et al., 2004b). It is evident that this
mutant has a hyper-butanol producing ability, and this feature makes it very promising in

164
economically producing butanol through ABE fermentation. The mutant strain was
obtained in our lab through repeated adaption and evolution in the FBB, and it proved
that FBB was very useful in enhancing the cells’ tolerance towards stressful environment
and selecting mutated strain. FBB provided a support for cell-immobilization, and kept
the active cells highly viable to enhance the reactor productivity through cell-renewal.
Batch fermentations using CBH was studied to evaluate the feasibility of using CBH
as an alternative carbon source for biological butanol production. After the two-step
enzymatic hydrolysis, the resulting CBH contained 44.8 g/L glucose, 1.63 g/L xylose,
0.055 g/L arabinose, 0.353 g/L lactic acid, and 0.457 g/L acetic acid (Table 5.1). The total
sugar yield was 0.46 g/g cassava bagasse based on a 10% solid loading. The starch and
cellulose components of the bagasse were converted into fermentable sugars after
enzymatic hydrolysis. The remaining 54% of the bagasse could not be hydrolyzed into
sugars by hemicellulose enzymes or acid hydrolysis (data not shown), which probably
accounted for moisture and insolubles. It was reported (Pandey et al., 2000) that the
compositions of cassava bagasse from different regions varied dramatically, with starch
ranging from 40-60% and fibers from 14-50%. The decent amount of glucose present in
the CBH makes it suitable as an alternative carbon source for fermentation process. ABE
production using CBH is shown in Figure 5.2B. Glucose was rapidly depleted within 40 h,
and the butanol reached 9.7 g/L. Like its parental strain C. beijerinckii ATCC 55025, the
mutant prefers glucose over xylose, and depletes glucose first before using xylose (strain

165
study, data not shown in this work). Due to the trivial amount of xylose and arabinose,
the mutant did not utilize these sugars after glucose was depleted. Fermentation stopped
due to lack of carbon source, and no inhibition was observed using the enzymatic
hydrolysate. Because of the mild condition used in the enzymatic hydrolysis of cassava
bagasse and no acid or alkaline pretreatment was employed, it is most likely that the
resulting hydrolysate was not inhibitory to the fermentation process; however, this
conclusion can not be reached until further investigation. Therefore, CBH was
supplemented with additional glucose to make a comparable initial glucose concentration
as control to further evaluate the ABE production. The results are shown in Figure 5.2C.
20.3 g/L butanol (total ABE of 33.8 g/L) was produced from CBH with additional
glucose, and glucose was used up in this process as well. The fermentation kinetics was
similar to the control. No inhibition caused by the hydrolysate was observed, and it
proved that the cassava bagasse hydrolysate can function successfully as an alternative
carbon source for ABE production. This result suggested that common fermentation
inhibitors from hydrolysate such as furfural, HMF and phenolic compounds may not be
generated, or generated at a non-inhibitory concentration, during the enzymatic process
under the mild conditions.
5.3.2 Fed-batch fermentation with simultaneous product removal by gas stripping
Concentrated cassava bagasse hydrolysate (CCBH) obtained through evaporation

166
was used as feed substrate for fed-batch operation. Fermentation kinetics in the reactor is
shown in Figure 5.3. CCBH contained 584.4 g/L glucose, 14.72 g/L xylose, 0.61 g/L
arabinose, 1.89 g/L acetic acid, and 1.38 g/L lactic acid (Table 5.1). The concentration of
acids in the CCBH was a higher than the original CBH (0.35 g/L acetic acid and 0.48 g/L
lactic acid), but lower than expected by concentration. This result indicated that
evaporation helped to remove volatile acids from hydrolysate, which was reported as a
physical method before as an alternative detoxification for hydrolysate (Mussatto and
Roberto, 2004). It was reported that furfural, which is a fermentation inhibitor derived
from sugar degradation, was also volatile and can be removed by 90% through rotary
evaporation (Larsson et al., 1999). However, other non-volatile compounds, such as
extractives and phenolic compounds derived from lignin degradation could accumulate in
the resulting concentrated hydrolysate and cause a more severe degree of inhibition on
the fermentation (Mussatto and Roberto, 2004).
Compared with batch process, fed-batch fermentation was extended to 169 h, and the
total ABE production was 90.3 g/L, of which 59.8 g/L was butanol (Table 5.2). Average
ABE and butanol yield was 0.37 g/g and 0.25 g/g, respectively. Highly concentrated
substrate (CCBH containing 584.4 g/L glucose) was utilized in this process without any
substrate inhibition, and 244.6 g/L glucose was consumed with an average utilization rate
of 1.45 g/L. The peak butanol concentration in the reactor was at 40 h (16.45 g/L), and
was maintained at below 10 g/L afterwards throughout the fermentation. From 30 h to 40

167
h, the increasing butanol concentration in this period indicated that the butanol
production rate was faster than the butanol stripping rate when stripping was first
initiated. Gradually, a dynamic equilibrium between removal and production rate was
reached, and butanol level stayed relatively stable in the reactor afterwards. This
indicated that 1.25 L/min gas flow rate was sufficient to keep butanol level below 10 g/L
for a 1.0 L fermentation system. Further increasing the gas flow rate would lower the
butanol level in the reactor, which adversely affects the butanol level in the recovered
stream. From Figure 5.3B, butanol concentration in the recovered stream ranged from 10
to 16%, and at this level it led to a very distinct butanol/aqueous phase separation based
on an 8% butanol saturation point in water. This phase separation would significantly
simplify the butanol purification process and make the butanol recovery energy-saving
and economic. It is worth noticing (Figure 5.3A) that after the initiation of product
removal, the ratio of acetone and butanol in the reactor became ~ 1:1 instead of 1:2
(typical ratio is ABE 3:6:1). In addition, the butanol concentration was about 5-fold more
than the acetone concentration in the recovered stream (Figure 5.3B), and the butanol to
acetone ratio was in the range of 2.82 to 4.58:1 in the condensate throughout the
fermentation. This explained the ratio change in the reactor, and demonstrated that the
gas stripping was highly selective towards butanol compared to acetone.
Fermentation results of each individual cycle are summarized and compared in Table
5.3. Butanol yield was lower during the first cycle, which was probably due to sugar

168
usage on cell growth at the beginning. When the cells entered the stationary phase and
utilized carbon source mainly for ABE production during this phase, butanol yield
increased to 0.27 g/g (second cycle), and stayed stable at 0.25 g/g afterwards (third and
fourth cycle). However, butanol productivity, along with glucose utilization rate
decreased gradually with time (Table 5.3), indicating that the fermentation has been
slowed down significantly. There are many possible explanations for this phenomenon,
and lack of nutrient seemed to be a very reasonable one. It was reported that ABE
fermentation failed due to exhaustion of nutrients, and addition of nutrients in the reactor
led to an increase in glucose consumption and cell concentration (Ezeji et al., 2003;
2004b). The average glucose utilization rate was slower than from batch fermentation,
which was probably due to slow cell activity caused by exhaustion of nutrients.
In order to investigate the effect of nutrient on fed-batch fermentation, extra yeast
extract (0.5 g) was added along with concentrated substrate (equivalent to 0.5 g/L in the
medium) after 85 h for each cycle. 1g/L initial yeast extract (P2 formula) was used to
start the fermentation at the beginning like others. No buffer solution was supplemented
to the fermentation additionally because the pH was maintained carefully at 5.0
throughout the fermentation. No vitamins or minerals were additionally supplemented.
The fermentation results in the reactor are shown in Figure 5.4.
The ABE produced in this process was 108.5 g/L, of which 76.4 g/L was butanol.
336.6 g/L glucose was utilized, and the average consumption rate was 1.28 g/L. h. The

169
duration of this fermentation was extended to 263 h. The average ABE and butanol yield
was 0.32 g/g and 0.23 g/g, respectively (Table 5.2). From Table 5.3, glucose consumption
rate was fast for the first two cycles (average from 1.95 to 3.16 g/L. h), and slowed down
at the end of the third cycle before nutrient supplementation (1.38 g/L. h). Similar trends
were also observed on butanol and ABE productivity. This result was similar to our
observation from the fed-batch fermentation without nutrient supplementation. With
nutrient supplementation, glucose consumption rate increased from 0.82 g/L h to 1.17 g/L
h during the fourth and fifth cycle, indicating nutrient supplementation played a role in
reviving the cells. During the sixth cycle, glucose was depleted, indicating that the
fermentation can still carry on for a longer operation time given more carbon source.
Butanol productivity was maintained stably at 0.2- 0.23 g/L. h during the last three cycles
with nutrient supplement. Due to the extended fermentation time, dead cells and
non-active cells coexisted with actively solvent-producing cells in the fermentation broth
(Qureshi et al., 1988). Supplementation of nutrient provided an opportunity to generate
fresh new cells to replace the non-productive old cells, keeping the fermentation going
and maintaining reactor productivity. However, it should be noted that nutrient
supplementation can only rejuvenate the cell’s viability and solvent productivity to some
extent, accumulation of cellular metabolites and other toxic substances from CCBH will
have an adverse effect over time on the cells. The complexity of hydrolysate posed a
challenge in accurately analyzing the exact compounds. Even though some possible

170
inhibitors may not be at an inhibitory level in the CBH, the amount of non-volatile
inhibitors could be significantly increased after concentration. With the repeated feeding
of the CCBH, these inhibitors may accumulate in the broth and reach an inhibitory level,
and eventually slow down the fermentation process or even cease it.
The butanol to acetone ratio in the recovered stream was in the range of 3.18 to
5.13:1, which was similar to what we observed earlier. More butanol was recovered from
the first three cycles (about 150 g/L in the condensate), and less butanol was recovered
from the last three cycles (about 115 g/L in the condensate) (Figure 5.4B). The acetone
concentration in the condensate remained fairly stable at around 30 g/L. The change of
butanol concentration in the recovered stream was consistent with the trend of glucose
utilization rate and reactor productivity. When glucose was utilized quickly, butanol
productivity was fast and more butanol was recovered by stripping. When butanol
productivity was slower, more water was stripped due to lower butanol concentration in
the reactor (around 5 g/L, Figure 5.3A).
It was also interesting to notice that only 26.9 g/L acetone was produced, and the
acetone-butanol (AB) ratio became 1: 2.84 since 76.4 g/L butanol was produced. This
ratio was dramatically different from the typical 1:2 AB ratio. Due to the select removal
of butanol and acetone by gas stripping, the acetone-butanol ratio strayed far away from
typical 2:1 in the reactor. We propose that this phenomenon caused an imbalance in the
reactor, and the cells started to produce more butanol and less acetone to compensate,

171
which led to more overall butanol production and a lower total AB ratio. Another reason
was probably due to unknown factors in the CBH. There may be some proteins or
compounds in the CBH that affect the cells, directing more flux towards the butanol
production. It was reported (Ezeji et al., 2007c) that the acetone butanol ratio changed
from 1:2.5 to 1:1 in a Ca(OH)2 detoxified corn fiber hydrolysate fermentation. Even
though the trend of ratio change is different from this work, it was still uncharacteristic of
the C. beijerinckii culture. It was mentioned (Ezeji et al., 2007c) that some substrates can
change the acetone butanol ratio, but the real reason behind the change remained unclear
and unverified. Residual inhibitors in the corn fiber hydrolysate were proposed as a
possible cause for this unusual ratio change by Ezeji et al. (2007c).
5.4. Conclusion
In summary, this work demonstrated an integrated fermentation process utilizing
cassava bagasse hydrolysate for butanol production. Enzymatic hydrolysis of cassava
bagasse is a mild yet efficient process to yield fermentable sugars with no inhibition on
sequential fermentation process. The super-butanol-producing mutant utilized a highly
concentrated hydrolysate containing 584.4 g/L glucose in fed-batch fermentation, and
produced 108.5 g/L ABE with simultaneous butanol recovery by gas stripping.
Accumulation of dead cells and cellular metabolites impeded butanol production in
long-term operation, and supplementing nutrients revitalized cells and kept sugar

172
utilization rate at a stable level.
5.5 References
Bramorski, A., C.R. Soccol, P. Christen, S. Revah (1998). Fruity aroma production by Ceratocystis fimbriata in solid cultures from agro-industrial wastes. Rev. Microbiol., 29, 208-212.
Carta, F.S., C.R. Soccol, L.P. Ramos, J.D. Fontana (1999). Production of fumaric acid by fermentation of enzymatic hydrolysates derived from cassava bagasse. Bioresour. Technol., 68, 23-28.
Dürre, P. (1998). New insights and novel developments in clostridial acetone/butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2003). Production of acetone, butanol and ethanol by Clostridium beijerinckii BA 101 and in situ recovery by gas stripping. World J. Microbiol. Biotechnol., 19, 595-603.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004a). Butanol fermentation research: Upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004b). Acetone butanol ethanol (ABE) production from concentrated substrate: reduction in substrate inhibition by fed-batch technique and product inhibition by gas stripping. Appl. Microbiol. Biotechnol., 63, 653-658.
Ezeji, T.C., P.M. Karcher, N. Qureshi, H.P. Blaschek (2005). Improving performance of a gas stripping-based recovery system to remove butanol from Clostridium beijerinckii fermentation. Bioproc. Biosyst. Eng., 27, 207-214.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2007a). Bioproduction of butanol from biomass: from genes to bioreactors. Curr. Opin. Biotechnol., 18, 220-227.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2007b). Production of acetone butanol (AB) from liquefied corn starch, a commercial substrate, using Clostridium beijerinckii coupled with product recovery by gas stripping. J. Ind. Microbiol. Biotechnol., 34, 771-777.

173
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2007c). Butanol production from agricultural residues: Impact of degradation products on Clostridium beijerinckii growth and butanol fermentation. Biotechnol. Bioeng., 97, 1460-1469.
Ezeji, T.C. and H.P. Blaschek (2008). Fermentation of dried distillers’ grains and solubles (DDGS) hydrolysates to solvents and value-added products by solventogenic clostridia. Bioresour. Technol., 99, 5232-5242.
Jones, D.T. and D.R. Woods (1986). Acetone-butanol fermentation revisited. Microbiol. Rev., 50, 484-524.
Larsson, S., A. Reimann, N. Nilvebrant, J.J. Jönsson (1999). Comparison of different methods for the detoxification of lignocellulose hydrolysates of spruce. Appl. Biochem. Biotechnol., 77-79, 91-103.
Lee, S.Y., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentative butanol production by Clostridia. Biotechnol. Bioeng., 101, 209-228.
Mussatto, S.I. and I.C. Roberto (2004). Alternatives for detoxification of diluted-acid lignocellulosic hydrolyzates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Pandey, A., C.R. Soccol, P. Nigam, V.T. Soccol, L.P.S. Vandenberghe, R. Mohan (2000). Biotechnological potential of agro-industrial residues. II: cassava bagasse. Bioresour. Technol., 74, 81-87.
Qureshi, N., A.H.J. Paterson, and I.S. Maddox (1988). Model for continuous production of solvents from whey permeate in a packed bed reactor using cells of Clostridium acetobutylicum immobilized by adsorption onto bonechar. Appl. Microbiol. Biotechnol., 29, 323-328.
Qureshi, N. and H.P. Blaschek (2001a). Recovery of butanol from fermentation broth by gas stripping. Renew. Ener., 22, 557-564.
Qureshi, N. and H.P. Blaschek (2001b). Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 27, 287-291.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioproc. Biosyst Eng., 30, 419-427.

174
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta, H.P. Blaschek (2008). Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.
Silva, E.M. and S.T. Yang (1995). Kinetics and stability of a fibrous-bed bioreactor for continuous production of lactic from unsupplemented acid whey. J. Biotechnol., 41, 59-70.
Thongchul, N., S. Navankasattusas, and S.T. Yang (2009). Production of lactic acid and ethanol by Rhizopus oryzae integrated with cassava pulp hydrolysis. Bioproc. Biosyst. Eng., 33, 407-416.
Yang, S.T. (1996). Extractive fermentation using convoluted fibrous bed bioreactor. US Patent No. 5,563,069.
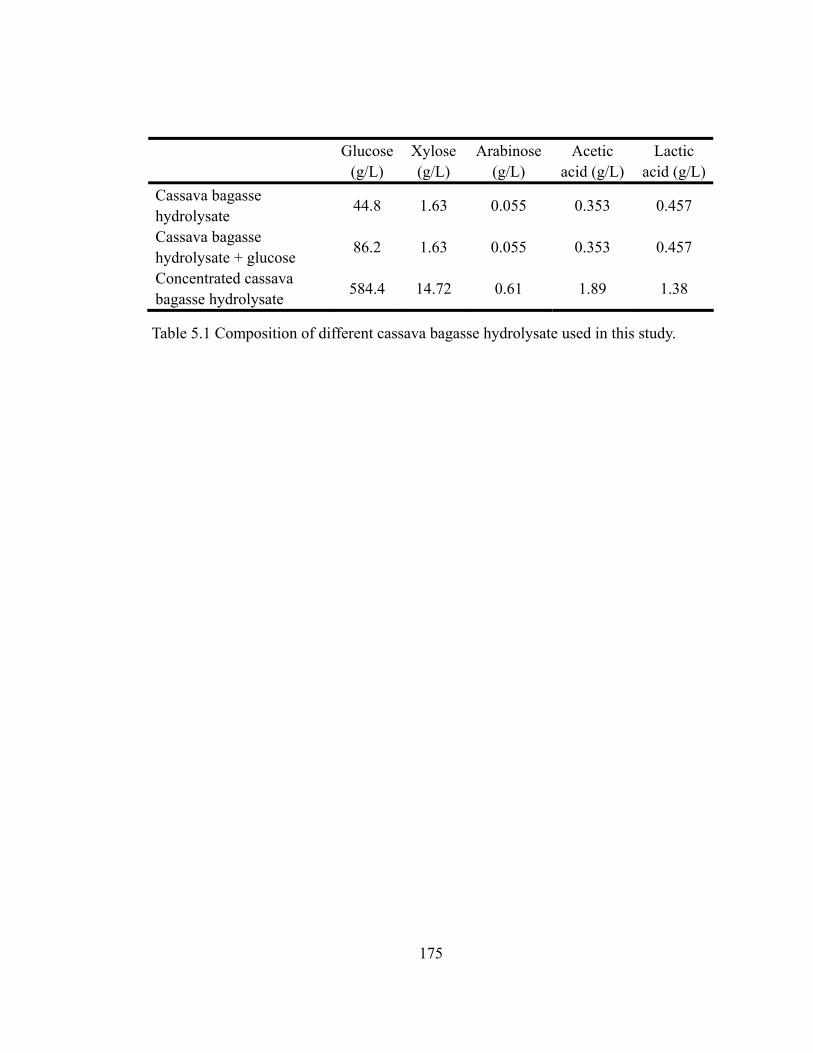
175
Glucose
(g/L) Xylose (g/L)
Arabinose (g/L)
Acetic acid (g/L)
Lactic acid (g/L)
Cassava bagasse hydrolysate
44.8 1.63 0.055 0.353 0.457
Cassava bagasse hydrolysate + glucose
86.2 1.63 0.055 0.353 0.457
Concentrated cassava bagasse hydrolysate
584.4 14.72 0.61 1.89 1.38
Table 5.1 Composition of different cassava bagasse hydrolysate used in this study.

Control
(glucose)CBEHbatch
CBEH + glucose batch
Integrated fed-batchw/o nutrient addition
Integrated fed-batchw/ nutrient addition
Acetone (g/L) 11.16 4.33 10.23 25.72 26.98 Butanol (g/L) 20.39 9.71 20.32 59.81 76.44 Ethanol (g/L) 2.22 1.37 3.32 4.78 5.09 Total ABE (g/L) 33.77 15.41 33.87 90.31 108.50 Butanol yield (g/g) 0.23 0.22 0.24 0.25 0.23 Ave. Butanol productivity (g/L. h) 0.38 0.24 0.37 0.35 0.29 ABE yield (g/g) 0.38 0.34 0.39 0.37 0.32 Ave. ABE productivity (g/L. h) 0.63 0.39 0.62 0.53 0.41 Acetic acid (g/L) 2.58 3.68 3.21 4.29 5.24 Butyric acid (g/L) 1.50 2.68 1.84 3.82 1.97 Total Acids (g/L) 4.08 6.36 5.06 8.11 7.21 Glucose utilized (g/L) 88.4 44.8 86.2 244.6 336.9 Glucose in the feed (g/L) 88.4 44.8 86.2 584.4 584.4 Ave. Glucose utilization rate (g/L . h) 1.64 1.12 1.57 1.45 1.28 Fermentation time (h) 54 40 55 169 263
176
Table 5.2 ABE production from cassava bagasse hydrolysate in batch fermentation and integrated fed-batch fermentation by C. beijerinckii JB 200.
176

Integrated fed-batch without nutrient supplement Integrated fed-batch with periodical nutrient supplement
Cycle 1 2 3 4 1 2 3 4 5 6
Fermentation Time (h) 40 36 37 53 38 17 30 58.5 46 67
Butanol (g/L) 16.45 15.09 15.68 12.59 17.13 13.53 8.84 11.64 9.96 15.47
ABE (g/L) 28.33 20.61 23.44 17.93 27.68 22.5 10.79 16.65 11.05 19.96
Butanol yield (g/g) 0.21 0.27 0.25 0.25 0.23 0.25 0.21 0.24 0.19 0.23
ABE yield (g/g) 0.36 0.37 0.38 0.36 0.37 0.42 0.26 0.35 0.21 0.3
Butanol productivity (g/L· h) 0.41 0.42 0.42 0.24 0.45 0.8 0.29 0.2 0.22 0.23
ABE productivity (g/L· h) 0.71 0.57 0.63 0.34 0.73 1.32 0.36 0.28 0.24 0.3
Glucose consumption (g/L) 79.0 55.1 62.2 50.3 74.2 53.8 41.3 47.7 53.6 66.3
Glucose utilization rate (g/L· h) 1.98 1.53 1.68 0.95 1.95 3.16 1.38 0.82 1.17 0.99
Table 5.3 Summary of performance of each cycle in the integrated fed-batch fermentations
177
177

sampling
Feed ordrain
FBB circulation
pH controller
Base
Gas circulation
FB
B
Co
nd
ense
r
Recoveredstream
sampling
Feed ordrain
FBB circulation
pH controller
Base
Gas circulation
FB
B
Co
nd
ense
r
Recoveredstream
Figure 5.1 Experimental setup of FBB-connected fermentor with gas stripping as online butanol recovery
178
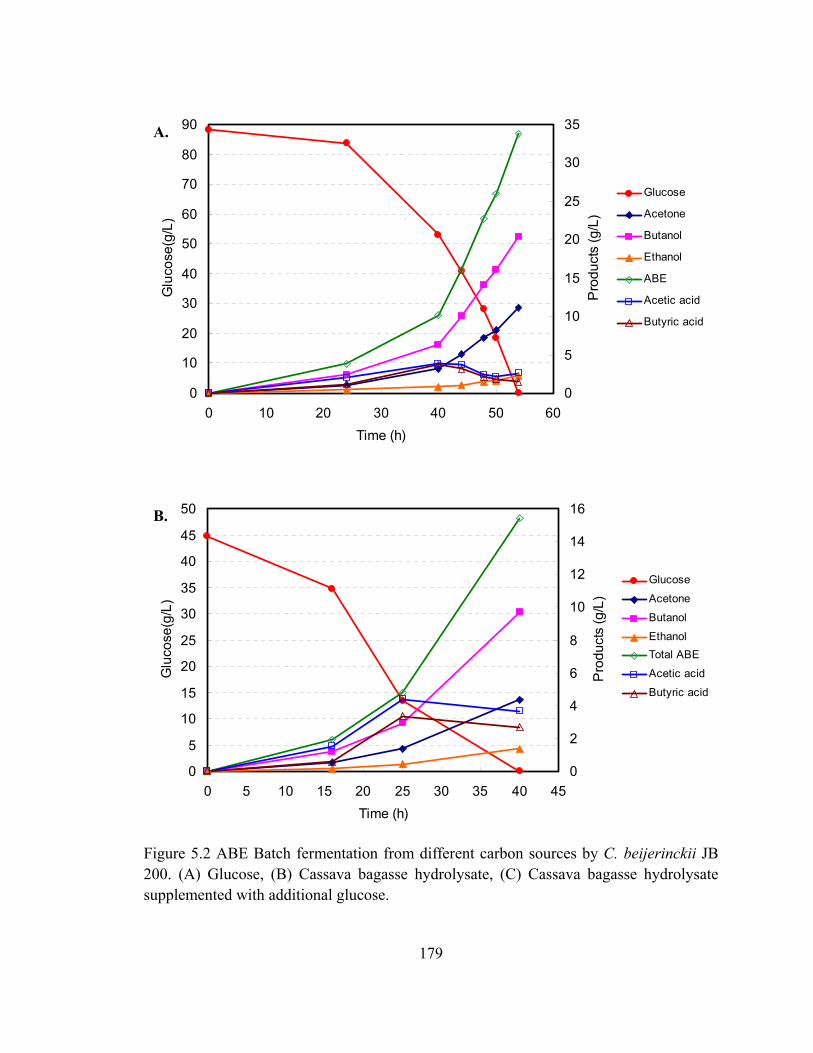
0
10
20
30
40
50
60
70
80
90
0 10 20 30 40 50 60
Time (h)
Glu
cose
(g/L
)
0
5
10
15
20
25
30
35
Pro
du
cts
(g/L
)
Glucose
Acetone
Butanol
Ethanol
ABE
Acetic acid
Butyric acid
A.
0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25 30 35 40 45
Time (h)
Glu
cose
(g/L
)
0
2
4
6
8
10
12
14
16
Pro
du
cts
(g/L
)
Glucose
Acetone
Butanol
Ethanol
Total ABE
Acetic acid
Butyric acid
B.
Figure 5.2 ABE Batch fermentation from different carbon sources by C. beijerinckii JB 200. (A) Glucose, (B) Cassava bagasse hydrolysate, (C) Cassava bagasse hydrolysate supplemented with additional glucose.
179

Figure 5.2 continued
0
10
20
30
40
50
60
70
80
90
0 10 20 30 40 50 60
Time (h)
Glu
cose
(g
/L)
0
5
10
15
20
25
30
35
Pro
du
cts
(g/L
)
Glucose
Acetone
Butanol
Ethanol
Total ABE
Acetic acid
Butyric acid
C.
180

FBB-connected fermentor
0
10
20
30
40
50
60
70
80
90
0 20 40 60 80 100 120 140 160
Time (h)
Glu
cose
(g/L
)
0
5
10
15
20
25
30
Pro
du
cts
(g/L
)
Glucose Acetone ButanolEthanol ABE Acetic AcidButyric Acid
Gas stripping initiated
1
2 34
FBB-connected fermentor
0
10
20
30
40
50
60
70
80
90
0 20 40 60 80 100 120 140 160
Time (h)
Glu
cose
(g/L
)
0
5
10
15
20
25
30
Pro
du
cts
(g/L
)
Glucose Acetone ButanolEthanol ABE Acetic AcidButyric Acid
Gas stripping initiated
1
2 34
A.
0
50
100
150
200
250
Pro
du
cts
(g/L
)
50 77 115 169
Time (h)
Condensate
Acetone
Butanol
Ethanol
ABE
Acetic Acid
Butyric Acid
Total Acids
1 2 3 4
0
50
100
150
200
250
Pro
du
cts
(g/L
)
50 77 115 169
Time (h)
Condensate
Acetone
Butanol
Ethanol
ABE
Acetic Acid
Butyric Acid
Total Acids
1 2 3 4
B.
Figure 5.3 Fed-batch fermentation integrated with gas stripping using CCBH by C. beijerinckii JB 200. (A) Glucose and products in the reactor vs. time, (B) Composition of the condensate recovered in each cycle.
181

FBB-connected fermentor
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280
Time (h)
Glu
cose
(g/L
)
0
5
10
15
20
25
30
Pro
du
cts
(g/L
)
Glucose Acetone ButanolEthanol ABE Acetic acidButyric acid
Gas stripping initiated
1
4
3
25
6
:supplementing nutrient
FBB-connected fermentor
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280
Time (h)
Glu
cose
(g/L
)
0
5
10
15
20
25
30
Pro
du
cts
(g/L
)
Glucose Acetone ButanolEthanol ABE Acetic acidButyric acid
Gas stripping initiated
1
4
3
25
6
:supplementing nutrient
A.
0
50
100
150
200
Pro
du
cts
(g/L
)
38 56.5 87 148.5 195 263
Time (h)
Condensate
Acetone
Butanol
Ethanol
ABE
Acetic acid
Butyric acid
Total Acid
1 32 4 5 6
0
50
100
150
200
Pro
du
cts
(g/L
)
38 56.5 87 148.5 195 263
Time (h)
Condensate
Acetone
Butanol
Ethanol
ABE
Acetic acid
Butyric acid
Total Acid
1 32 4 5 6
B.
Figure 5.4 Fed-batch fermentation integrated with gas stripping using CCBH by C. beijerinckii JB 200 with periodical nutrient supplementation. (A) Glucose and products in the reactor vs. time, (B) Composition of the condensate recovered in each cycle.
182

183
Chapter 6: Biobutanol Production from Renewable Wood Pulp Hydrolysate in an
Integrated Process: Evaluation of Detoxification and Alternative Nitrogen Sources
Abstract
Wood pulp hydrolysate was examined as a potential substrate for butanol
fermentation in this study. Due to the inhibitors present in the hydrolysate, several
dilution levels and detoxification treatments, including overliming, activated charcoal,
and resin and evaporation, were evaluated for their effectiveness in relieving the
inhibition on fermentation. 6.73 g/L total solvents were obtained in the non-treated and
non-diluted wood pulp hydrolysate, whereas 11.35 g/L total solvents were achieved in the
resin and evaporation treated hydrolysate. Coupled with gas stripping, the total solvent
production was further enhanced to 17.73 g/L with simultaneous butanol recovery. Four
alternative nitrogen sources, corn steep liquor, molasses, soybean meal and cottonseed
protein, were evaluated for their effect on butanol fermentation to replace yeast extract in
search of a cost-effective medium formula. 3.34-8.99 g/L solvents were produced from
wood pulp hydrolysate when using these nitrogen sources alone. When supplemented
with ammonia acetate, the solvent production was significantly enhanced, and 8.2-11.21
g/L solvents were obtained under the same condition.

184
6.1 Introduction
Biofuel butanol has been the research focus for the past few decades in search of a
suitable and sustainable fuel alternative to fossil fuel (Ezeji et al., 2004a; 2007; Jones and
Woods, 1986; Kumar and Gayen, 2011; Lee et al., 2008). Quickly depleting oil reserves
is a major concern during this fuel-dependent time and demands for renewable alternative
fuels have become crucial to increase the fuel independency of a country. Biobutanol has
demonstrated its superiority to bioethanol in terms of energy density, engine
compatibility and safety, and has become the center of research as the next generation
biofuel since 2005 (Kumar and Gayen, 2011; Nigam and Singh, 2011; Qureshi and Ezeji,
2008). Historically, butanol was produced through Acetone-Butanol-Ethanol (ABE)
fermentation in the early 20th century, and thrived during the 1930s and 1940s (Jones and
Woods, 1986). Traditional substrates such as corn, glucose and cane molasses, were
primarily used for ABE fermentation, and butanol produced in this manner was
considered economic and feasible (Qureshi and Ezeji, 2008). However, due to the
increasing demand of food supply worldwide and the debate of “food vs. fuel”, ABE
fermentation from these food sources became cost-intensive and gradually phased out
during 1970s (Ezeji et al., 2004a; Nigam and Singh, 2011). In the past few decades,
renewed interest in ABE fermentation has returned with advances in many aspects, in
search for a solution to serious environmental concerns caused by the extensive abuse of
fossil fuel and the rapidly depleting petroleum fuel supply (Ezeji et al., 2010; Nigam and

185
Singh, 2011; Zheng et al., 2009).
Raw material has always been the most cost-intensive part of ABE fermentation,
which greatly influences the final butanol price and rationales the process economics and
feasibility (Durre, 1998; Gapes, 2000; Qureshi and Blaschek, 2000; 2001). Biobutanol
from food-based substrates such as corn or molasses, is considered as first-generation
biofuel, whereas biobutanol from inedible biomass is believed to be biofuel of the second
generation and attracts the most research efforts (Nigam and Singh, 2011; Weber et al.,
2010). Lignocellulosic biomass represents the largest renewable carbon source on earth,
including agricultural residues, energy crops, forestry woody residues and municipal
wastes (Kumar et al., 2009). Pretreatment procedures such as acid/alkaline hydrolysis and
enzyme digestion are required to break down cellulose and hemicellulose to release
sugars that are accessible to microorganisms in the sequential fermentation process
(Hendriks and Zeeman, 2009; Mosier et al., 2005; Saha, 2003). Successful utilization of
the lignocellulosic hydrolysate usually requires detoxification as many fermentation
inhibitors are produced under the extreme conditions during pretreatment (Mussatto and
Roberto, 2004a). Many agricultural residues and energy crops, such as barley straw,
wheat straw, corn fiber, corn stover switchgrass, wheat bran and distillers’ dried grains
and soluble, have been successfully investigated as substrates for ABE fermentation with
10-20 g/L ABE production (Ezeji and Blaschek, 2008; Liu et al., 2010; Qureshi et al.,
2007; 2008a; 2008b; 2010a; 2010b). However, another category of lignocellulosic

186
biomass, woody residues, has not been studied for butanol production. Green liquor,
which is mostly dissolved hemicellulose after cellulose extraction from wood chips in the
paper pulping industry, is a great resource of carbohydrates for butanol production.
Among some byproducts from industrial processing of agricultural crops, only corn steep
liquor (Parekh et al., 1998; Parekh et al., 1999) has been studied as a nutrient source for
ABE fermentation. Other potential nutrients such as soybean meal and cottonseed
proteins, haven’t been evaluated for butanol production in ABE fermentation as
cost-effective nitrogen sources. Aside from the raw material, the biggest limiting factor in
ABE fermentation has always been the butanol inhibition and toxicity (Ezeji et al., 2010).
Many recovery techniques have been reported to relieve butanol inhibition and enhance
the efficiency of ABE fermentation, such as pervaporation, liquid-liquid extraction and
gas stripping (Durre, 1998; Ezeji et al., 2004a; Lee et al., 2008; Vane, 2008). Gas
stripping is a very effective technique with low energy requirement to integrate with the
fermentation process, and has been demonstrated to work successfully for ABE recovery
(Ezeji et al., 2003; 2004b; Qureshi et al., 2007).
In this work, wood pulp was investigated as a new category of substrate as opposed
to agricultural residues and energy crops to broaden the feedstock pool of ABE
fermentation by Clostridium beijerinckii. Gas stripping was employed to study its effect
on butanol production and sugar conversion. Several byproducts from the processing
industry including cottonseed protein, soybean meal, corn steep liquor and molasses,

187
were evaluated as alternative nitrogen sources for Clostridium beijerinckii in ABE
fermentation. Our aim was to evaluate the feasibility of using wood pulp hydrolysate to
produce butanol and investigate alternative cost-effective nitrogen sources in order to
develop an economic and efficient process to produce renewable and green butanol.
6.2 Materials and methods
6.2.1 Strain and inoculum preparation
C. beijerinckii NCIMB 8052 was obtained from ATCC deposit (ATCC number
51743). After adaption and evolution engineering at our research lab, a mutant strain
from C. beijerinckii NCIMB 8052 was obtained, namely C. beijerinckii CC101, which
was used as the working culture for ABE fermentations in this study. Spores of C.
beijerinckii CC101 were routinely stored in the refrigerator at 4oC in the clostridia
medium. Spores (2 ml) were heat-shocked at 80 oC for 3 min and transferred to 50 ml
RCM growth medium (Difco Reinforced Clostridia Medium, Becton, Dickinson and
Company, MD, USA). 3.8 g of solid RCM powder was dissolved in 100 ml distill water
to prepare 100 ml liquid RCM in a 125 ml serum bottle. The medium was
nitrogen-purged for 8 min to remove oxygen. The serum bottle was tightly capped by a
rubber stopper and aluminum seal. The mixture was autoclaved at 121 oC for 30 min
followed by cooling to 37 oC. The heat-shocked spores were incubated at 37 oC for 12-16
hrs until cells were highly active. The active culture (5% inoculum) was used as seed

188
culture for all fermentation studies, both the serum bottle and bioreactor studies.
6.2.2 Wood pulp hydrolysate
Wood pulp hydrolysate (WPH) was generously provided by ButylFuel LLC.
(Columbus, OH). Wood chips were first cooked, washed and extracted of cellulose in
alkaline for paper pulping. The dissolved hemicellulose, along with lignin, was left in the
green liquor, and the hemicellulose was further hydrolyzed by acid to release all the
monosaccharide that was used as carbon source in the fermentations. The pH of the wood
pulp hydrolysate was adjusted to ~2-3 to avoid contamination during transfer and storage.
The resulting wood pulp hydrolysate was sent to our lab for evaluation as a potential
renewable feedstock for ABE fermentation.
6.2.3 Detoxification procedures
6.2.3.1 Overliming detoxification
The pH of WPH was first adjusted to 10.0 with Ca(OH)2. The mixture was heated to
90 oC and stirred at 100 rpm for 30 min. The precipitates were removed by centrifuge at
7000 g for 10 min followed by cooling to the room temperature. The pH of the mixture
was then adjusted back to 6.5 by adding concentrated H2SO4. The precipitates were again
removed by centrifuge at 7000 g for 10 min.

189
6.2.3.2 Activated carbon/charcoal detoxification
2% activated carbon was added into WPH (2 g activated carbon per 100 ml WPH) at
pH 2.0. The mixture was heated to 90 oC and stirred at 150 rpm for 30 min. Activated
carbon was then removed by vacuum filtration and the resulting WPH was ready for
evaluation as detoxified WPH.
6.2.3.3 Resin and evaporation detoxification
One batch of wood pulp hydrolysate sent to our research lab was detoxified using
resin and evaporation detoxification techniques, which was provided by ButylFuel LLC.
6.2.4 Preparation of alternative nutrient sources
6.2.4.1 Corn steep liquor (CSL) and molasses
Corn steep liquor was obtained from Cargill, Iowa, USA. It was highly concentrated
and very viscous in the form of a paste. 10 g CSL was mixed and dissolved in 40 ml
distill water to prepare CSL stock solution (equivalent of 250 g/L). After dissolving, the
remaining solid was removed by vacuum filtration. 100 ml clear CSL solution was
nitrogen-purged for 8 min to remove oxygen in a 125 ml serum bottle, which was later
tightly capped with a rubber stopper and aluminum seal. The clear solution was
autoclaved at 121 oC for 30 min for sterilization.
Molasses syrup was obtained from a local market made of sugarcane. The molasses

190
syrup was also highly concentrated and very viscous. Due to the similar physical form of
molasses and corn steep liquor, molasses stock solution was prepared in the exact same
way as the CSL solution. Commercial molasses was a source for trace amounts of
vitamins and significant amounts of several minerals, including calcium, magnesium,
potassium and iron. Most of these minerals were required for ABE production by
Clostridium beijerinckii, and were included in the P2 formula which was commonly used
as a semi-defined clostridia production medium formula.
6.2.4.2 Cottonseed protein hydrolysate and soybean meal hydrolysate
Cottonseed protein (CP) was obtained from Cargill, Iowa, USA. CP is a protein-rich
byproduct from oil extraction in the cottonseed processing industry, so it is insoluble in
aqueous solution. In order to utilize the protein and other nutrient components in CP, CP
was hydrolyzed with 0.3N HCl (10% solid loading (w/v), equivalent of 100 g biomass/L)
at 121oC for 30 min. The pH of the resulting hydrolysate was neutralized with solid
NaOH to 7.0, and the remaining solids were removed by centrifuge. This cottonseed
protein hydrolysate (CPH) was nitrogen purged for 8 min to remove oxygen and then
autoclaved for 30 min at 121 oC for sterilization in a 125 ml serum bottle.
Soybean meal (SM) was obtained from United Soybean Board (USB), Chesterfield,
MO, USA. Like CP, SM is insoluble in aqueous solution. Both SM and CP are commonly
used as animal feed, and they are both rich in protein. Therefore, SM was pretreated in

191
the same way as CP and the resulting soybean meal hydrolysate (SMH) was obtained.
The above-mentioned CSL, molasses, CPH, SMH were all evaluated as potential
nutrient sources to substitute for yeast extract in ABE fermentation in this study.
6.2.5 Production medium preparation and fermentation
Unless otherwise noted, P2 formula was used in the production medium in this study.
P2 medium contained carbon source (glucose, or WPH), yeast extract (2 g/L), buffer (0.5
g/L KH2PO4 and 0.5 g/L K2HPO4), 2.2 g/L ammonium acetate, vitamins (0.001 g/L
para-amino-benzoic acid (PABA), 0.001g/L thiamin and 10-5 g/L biotin), and mineral
salts (0.2 g/L MgSO4·7H20, 0.01 g/L MnSO4· H20, 0.01 g/L FeSO4· 7H20, 0.01 g/L NaCl).
For the fermentations performed in serum bottle study, the volume of media was 50 ml.
For the fermentations performed on the bioreactor, the volume of media was 1.0 L.
Carbon source and P2 stock solution (yeast extract, ammonium acetate and buffer,
10-fold concentrated) were autoclaved separately at 121oC and 15 psig for 30 minutes for
sterilization to avoid chemical reactions between sugar and nitrogen sources. Minerals
and vitamins were prepared at 200-fold and 1000-fold concentration separately, and were
filtered through 0.2 μm sterile membrane (25mm 0.2μm syringe filter, Fisherbrand, NJ,
USA) for sterilization. Based on concentration-fold and medium volume, proper amount
of P2 stock solution was aseptically transferred into serum bottle/bioreactor containing
carbon source, followed by addition of minerals and vitamins. The medium in the serum

192
bottle study was nitrogen purged for 8 min prior to sterilization, whereas the medium in
the bioreactor was aseptically nitrogen purged for 2 hr after sterilization using a 0.2μm
vent filter unit (Millex, Millipore corp., MA, USA) to reach anaerobia. The bioreactor
used in this study was Marubishi MD-300 (B.E. Marubishi Co. Ltd., 5.0-Liter) integrated
with a pH controller (Cole-Parmer, IL, USA).
When using the diluted WPH medium, X% WPH medium meant that X% (v/v) of
the total volume was pure WPH, with the rest compensated with distilled water thus
accomplishing dilution purpose. When using CSL and molasses as the sole nutrient
source, 2 ml CSL or molasses stock solution was added into the serum bottle containing
carbon source, making a total of 50 ml production medium with the equivalent of 10 g/L
CSL or molasses in the medium. When using both CSL and molasses as the combined
nutrient source, only 1 ml of each stock solution was added into a serum bottle containing
total 50 ml medium. When using CPH or SMH as nutrient source, 5 ml of each solution
was added into the serum bottle to make 50 ml production medium. Prior to fermentation,
the pH of all media was adjusted to 6.5 using 6N NaOH.
Actively grown C. beijerinckii CC101 cells were inoculated into fermentation media
at 5% inoculum. All fermentation was performed at 37 oC with no agitation. Samples
were taken periodically for analysis of sugar consumption and ABE production. 0.2%
CaCO3 was employed in the serum bottle studies to ensure the pH staying above 5.0. pH
of the fermentation on the bioreactor was carefully monitored and maintained at 5.0 by

193
periodically adding 6N NaOH. Gas stripping was employed as a product recovery
technique to online recover butanol produced in the bioreactor, and was initiated by
circulating fermentation gases (H2 and CO2) into the fermentation broth by a peristaltic
pump (Masterflex L/S, standard drive, Cole-parmer, IL, USA) at 1.25 L/min. The
stripping gases containing ABE vapors from the broth sequentially passed through a coil
condenser (Pyrex Brand Graham Condenser, 300 mm water jacket, Fisher Scientific, NJ,
USA), where the ABE vapors were condensed and collected as ABE condensate in a 125
ml conical flask cooling trap. The condensation temperature was controlled at 1 oC by a
refrigerated circulator using 50% (v/v) ethylene (Isotemp Refrigerated circulator Model
910, Fisher Scientific, PA, USA). Gas streams free or less of ABE vapors were circulated
back into the bioreactor to remove more volatile solvents, and the gas recycle was in a
closed loop to prevent any loss into open air. Gas stripping device (condenser and tubings)
was autoclaved for sterilization and nitrogen purged before being aseptically connected to
the fermentor.
6.2.6 Analytic methods
The compositions of WPH, CSL, molasses, SMH and CPH were analyzed by a high
performance liquid chromatography (HPLC) with an organic acid column (Bio-Rad
HPX-87, ion exclusion organic acid column, 300 mm × 7.8mm). Samples were
centrifuged at 13.2 g for 5 min in microcentrifuge tubes and diluted 10 times with

194
distilled water prior to analysis on HPLC. HPLC was run at 45 oC using 0.01N H2SO4 as
the eluent at a flow rate of 0.6 ml/min. 15μL sample was injected by an automatic injector
(SIL-10Ai) and the running time was set at 36 min. A refractive index (RI) detector
(Shimadzu RID-10A) was set at the range of 200 to detect the organic compounds in the
sample. The HPLC column was installed in a column oven (CTO-10A) with temperature
control at 45 oC. Peak height was used to calculate concentration of sugars in the sample
based on the peak height of standard sample.
Glucose concentration was measured by a glucose and lactate analyzer, YSI
biochemistry analyzer (2700 Select). The fermentation products, acetone, butanol,
ethanol, acetic acid, and butyric acid, were measured with a Shimadzu GC-2014 gas
chromatograph (GC) (Shimadzu, Columbia, MD, USA) equipped with a flame ionization
detector (FID) and a 30.0 m fused silica column (0.25m film thickness and 0.25 mm ID,
Stabilwax-DA). To reduce the injection mechanic error margin, internal standard method
was used to analyze the concentration of products in the samples. Isobutanol and
isobutyric acid were used as internal standards for the solvent products and acid products
present in the samples. An internal standard buffer solution containing 0.5 g/L isobutanol,
0.1 g/L isobutyric acid, and 1% phosphoric acid was used to dilute each sample 20 times
for acidification and calibration prior to analysis on GC. The gas chromatograph was
operated at an injection temperature of 200 oC with 1 μL of the acidified sample injected
by the AOC-20i Shimadzu auto injector. Column temperature was held at 80 oC for 3 min,

195
raised to 150 oC at a rate of 30 oC/min, and held at 150 oC for 3.7 min.
6.3 Results and discussion
6.3.1 Effect of dilution and detoxifications on WPH compositions and ABE
fermentation
Lignocellulosic feedstocks require pretreatment such as acid or alkaline hydrolysis
in order to release the fermentable sugars accessible to the bacteria. During this process,
many degradation products from sugars (furfural and hydroxymethylfurfural (HMF)),
lignin (phenolic compounds) and acetyl groups attached to hemicellulose backbone
(acetic acid) are formed inevitably (Mussatto and Roberto, 2004a). These chemicals are
highly inhibitory and hinder the ABE production. Therefore, it was anticipated that the
WPH was inhibitory to ABE fermentation. Dilution, overliming, activated carbon and
resin and evaporation detoxification methods were studied to evaluate their effects on
improving ABE production.
The compositions of the WPH used in this study are summarized in Table 6.1. It
contained a total sugar of 65.54 g/L, which were mostly xylose and some glucose. Acetic
acid, a byproduct during the acid hydrolysis of hemicellulose, was present at a tolerable
concentration of ~2 g/L. A small amount of formic acid was also detected in the WPH at
a concentration below 1 g/L. A 15.6% total sugar loss was observed when using
overliming detoxification. Martinez et al. (Martinez et al., 2001) reported a 8.7 ± 4.5% of

196
sugar reduction when performing overliming at 60 oC on sugarcane bagasse hydrolysate.
They also reported that acetic acid level was not affected by overliming, while furan and
phenolic compounds were reduced by 51 ± 9% and 41 ± 6 %, respectively. The reported
acetic acid and sugar level changes coincided with results obtained in this study. When
activated carbon detoxification was performed on WPH, the sugar composition remained
almost unchanged, indicating that this method did not result in a significant sugar loss.
The composition of resin and evaporation detoxified WPH was different from the results
using the other two methods, with a lower glucose but higher xylose concentration. This
was probably due to slight compositional difference between each batch of raw materials,
which resulted in a sugar difference in the resulting WPH. Activated carbon detoxified
and overlimed WPH were from the same batch, while resin and evaporation detoxified
was from another batch. It is hard to discuss the effect of resin on sugar loss due to lack
of control. However, it was noticeable that the acetic acid was not detectable in the resin
detoxified WPH, indicating that resin was very successful in removing the acetic acid.
96% acetic acid removal was reported using activated carbon treatment on wood
hemicellulose hydrolysate (Larsson et al., 1999).The acetic acid concentration in the
activated carbon detoxified and overlimed WPH was slightly lower compared to the
untreated WPH, which was probably due to the elevated temperature (90 oC) under which
the detoxification was performed. It has been reported (Mussatto and Roberto, 2004a)
that under elevated temperature, volatile inhibitors such as furfural and acetic acid were

197
reduced at some level.
ABE production with C. beijerinckii CC101 using diluted and detoxified WPH is
illustrated in Figure 6.1 and summarized in Table 6.2. Glucose/xylose P2 medium was
used as a control in this study. 13.67 g/L ABE was produced, of which 10.59 g/L was
butanol. Butanol yield was 0.32 g/g, and ABE yield was 0.42 g/g (Table 6.2). When using
untreated WPH as carbon source, only 6.73 g/L ABE was produced, of which 4.48 g/L
was butanol (Table 6.2). Compared with control study, ABE yield was reduced from 0.42
g/g to 0.29 g/g, with only 0.19 g/g butanol yield. This indicated that the ABE
fermentation suffered a several inhibition, and more carbon source was directed to
generate energy to maintain the cell metabolism, resulting in a very low yield of products.
When using a 50% diluted WPH, the inhibition was reduced as evidenced by an increased
ABE production of 8.14 g/L. The ABE yield was also improved to a 0.38 g/g, with a
butanol yield of 0.31 g/g. It was very clear that with the increasing amount of WPH in the
medium (Figure 6.1a, Table 6.2, from 50%, 60%, 70% to untreated WPH) the ABE
production was decreased accordingly, from 8.14 g/L to 6.73 g/L. The ABE yield and
butanol yield was also decreased with the increasing inhibition caused by the WPH
(Figure 6.1b).
Overliming was the least effective detoxification method for WPH among the three
tested in this study. From Figure 6.1 and Table 6.2, 5.83 g/L ABE was produced from
overlimed WPH, of which 4.41 g/L was butanol. ABE yield was 0.28 g/g, of which

198
butanol yield was 0.21 g/g. This result was very similar to the one using untreated WPH,
and no significant improvement was observed (Table 6.2). This indicated that overliming
may not be very effective in detoxifying WPH to improve the ABE production. It has
been reported (Qureshi et al., 2010a) that overliming was very successful in removing the
inhibitors from barley straw hydrolysate, resulting in 26.64 g/L ABE as compared to only
7.09 g/L ABE using untreated barley straw hydrolysate. This is probably due to the
chemical and compositional difference between feedstocks, and one type of
detoxification method does not apply to all kinds of hydrolysates. Activated carbon
detoxified WPH resulted in 8.98 g/L ABE, of which 6.27 g/L was butanol. This was a
40% increase in butanol production, and 33% increase in total ABE production,
compared with the results using untreated WPH. ABE and butanol yield in activated
carbon detoxified WPH was similar to untreated WPH, and was low compared with
glucose/xylose P2 control study (Table 6.2). This indicated that activated carbon
treatment successfully removed a fraction of inhibitors from WPH and relieved
fermentation stresses to an extent. It has been previously reported (Dominguez et al.,
1996; Mussatto and Roberto, 2004b; Rodrigues et al., 2001) that activated charcoal
treatment reduced the toxicity level caused by color and lignin degradation products.
Mussato and Roberto (2004b) reported that the adsorption onto activated charcoal was
affected by many factors, among which pH and temperature were the most important.
Activated carbon treatment was performed in this study under pH 2.0, 90 oC, 150 rpm for

199
30 min. Acetic acid (pKa = 4.75) was in the undissociated form under low pH condition,
and the removal was favored in this form by activated charcoal (Rodrigues et al., 2001).
It was also suggested that elevated temperature facilitated that packing density of
phenolic molecules in the pores of activated charcoal (Ravi et al., 1998). Mussatto and
Roberto (2004b) reported that the optimal condition obtained for activated charcoal
treatment on rice straw hydrolysate for xylitol production was at pH 2.0, 45 oC, 150 rpm
for 60 min, removing 48.9% color and 25.8% lignin degradation products. This also
supported our theory that activated charcoal treatment can only remove inhibitors to
certain extent, but not completely. From Figure 6.1a, it was shown that resin and
evaporation detoxification yielded the best ABE production from WPH with C.
beijerinckii CC101 in this study. 11.35 g/L ABE was obtained, of which 9.14 g/L was
butanol. ABE yield was as high as 0.39 g/g, of which butanol yield was 0.31 g/g (Table
6.2). This result was comparable to the control study using glucose/xylose P2 medium, in
terms of ABE production and yield. This concluded that resin and evaporation was very
successful in improving the fermentability of the WPH, and almost all of the inhibition
factors were removed from the WPH which resulted in an 81% increase in ABE
production compared with untreated WPH. Resin is a very effective treatment in
detoxifying lignocellulosic hydrolysate, even though the cost is high compared to
overliming and activated carbon treatment. Larsson et al. (1999) reported that among
several detoxification treatments on spruce hemicellulose hydrolysate, anion-exchange

200
resin was the most effective one, removing 96% acetic acids, 73% furan, 70% HMF and
91% phenolic compounds. Carvalho et al. (2006) also supported the statement that
adsorbent ion-exchange resin was very efficient in removing HMF, phenolics and acetic
acid. They recommended combining several detoxification procedures together to obtain
the maximum efficiency, and 82.5% acetic acid, 100% furfural and HMF, and 94%
phenolics were removed first with vacuum evaporation followed by adsorption onto
activated charcoal and resin.
When inhibition was severe in the ABE fermentation, lots of residual sugars were
observed at the end of fermentation. Sugar conversion rate was as low as 43% in the
untreated WPH, and fermentation stopped before these sugars can be turned into butanol.
This wastes carbon source, and dilution and detoxification are needed to achieve higher
sugar conversion for process efficiency. As shown in Table 6.2, 50% dilution increased
the sugar conversion from 42.7 % to 68.7 %, and resin and evaporation detoxification
increased the sugar utilization rate to 65.6 %. In the control study, over 86% of the sugars
were converted. This indicated that the bacteria can only utilize a fraction of sugars
before butanol started to inhibit the fermentation. It is recommended that WPH is first
detoxified and then diluted to proper concentration to find an optimal condition with both
high sugar conversion and high butanol production for the best process efficiency and
economics.

201
6.3.2 Investigation of potential economic nitrogen source
Production cost is always the primary drive for process development and
commercialization concern. Besides economic raw materials, cheap and effective
nitrogen source that is essential for microorganism’s growth is also indispensable. Four
potential nitrogen sources, corn steep liquor, molasses, cottonseed protein, and soybean
meal, were evaluated in this study in search of potential economic nitrogen source to
replace yeast extract. These sources are all by products from industrial processing of
agricultural products such as corn, cottonseed, and soybean, and are usually sold as
low-value animal feed. They are rich in protein, amino acids, minerals and trace elements,
and the complex nature makes them an excellent source for providing necessary nutrients
to the microorganism. The compositional analysis of each alternative nitrogen source
(stock solution prepared using the method listed in Section 6.2.4) is summarized in Table
6.1. CSL stock solution contained a small amount of glucose and xylose, and due to the
residual hemicellulose content in the soybean meal and cottonseed protein, both
hydrolysates also contained a small amount of glucose and xylose. 8.93 g/L acetic acid
was detected in CSL, where as only 0.87 g/L and 0.49 g/L acetic acid was detected in
CPH and SMH. Molasses stock solution contained sucrose, glucose and fructose, which
were all residual sugars from sugarcane juice extraction. Only 4% (v/v) of CSL, molasses,
and 10% (v/v) SMH and CPH, respectively, were added into fermentation media to
provide the nitrogen source in each study, so the sugar level of the fermentation medium

202
was not greatly affected. In this study, not only the effect of each individual alternative
nitrogen source was evaluated, but the effect of ammonia acetate was also studied.
Acetate was an ingredient in the optimized P2 formula, and it has been previously studied
and reported to enhance solvent production and prevent degeneration of the C.beijerinckii
and C. acetobutylicum (Chen and Blaschek, 1999; Gu et al., 2009).
The results are shown in Figure 6.2 and Table 6.3. 70% resin and evaporation
detoxified WPH was used as carbon source, and P2 formula was employed in the control
study. When solely using alternative nitrogen source in the medium, CSL resulted in the
best butanol production, (7.46 g/L), followed by CSL with molasses (6.37 g/L), CPH
(5.33 g/L), SMH (4.63 g/L), and molasses (2.49 g/L). This indicated that all the
alternative nitrogen sources evaluated in this study can provide nutrients to the cells on
some level, and CSL was the richest among all the candidates. Molasses was comprised
of mostly sugars and some minerals, but lack of sufficient nitrogen source for the cell
growth resulted in poor butanol production. CSL is the by-product of the corn
wet-milling industry, and has been previously reported to provide nutrients to produce
ethanol (Kadam and Newman, 1997), butanol (Parekh et al., 1998; Parekh et al., 1999),
and acetic acid (Bock et al., 1997). Soybean meal and cottonseed protein are generally
considered as animal feed, and have never been reported as potential nutrients for
bacteria or yeast to produce butanol or ethanol. From this study, it was shown that SM
and CP provided nutrients to C. beijerinckii CC101, and resulted in a decent amount of

203
butanol (4-5 g/L) in the fermentation with no other supplement (Table 6.3). This indicated
that SM and CP were rich in protein, minerals and salts that were necessary for the
growth of Clostridium beijerinckii. Cell can solely rely on the nutrients present in these
two alternative sources to grow and produce butanol. However, the butanol production
from these alternative nitrogen sources alone was lower than the control using P2 formula.
This indicated that some ingredient present in P2 formula was lacking in these alternative
sources. It is shown in Figure 6.2a that the supplementation of ammonia acetate had a
significant effect on butanol production. When ammonia acetate was supplemented, CSL
with molasses medium resulted in 9.17 g/L butanol, which was very similar as the 9.51
g/L butanol obtained from control using P2 formula. When ammonia acetate was added,
butanol production was improved from 7.46 g/L to 8.63 g/L in CSL medium (Table 6.3).
No additional P2 minerals or vitamins were employed in this study. The enhancement on
butanol production by ammonia acetate was most clearly demonstrated in the molasses
fermentation study. When ammonia acetate was added, the butanol production was
boosted from 2.49 g/L to 5.72 g/L, corresponding to a 130% increase. Butanol production
was also increased in CPH and SMH study, from 5.33g/L to 7.42 g/L, and 4.63 g/L to 6.0
g/L, respectively (Table 6.3). Ammonia acetate not only provided additional NH4+ that
can be utilized by the cells as a nitrogen source, but also provided acetate that was
important to ABE fermentation. It has been reported that ammonia acetate sufficiently
provided the essential nutrient present in corn but lacking in cassava for butanol

204
fermentation, and proposed that acetate was associated with important enzyme expression
during acidogensis and solventogensis for successful butanol production (Gu et al., 2009).
It has also been reported that acetate can stabilize the solvent production, ensure the
expression of sol operon, and prevent the degeneration of C. beijerinckii (Chen and
Blaschek, 1999). Comparing CSL and CSL with molasses results, more butanol was
produced with the presence of molasses, indicating that molasses supplemented
additional nutrients such as minerals and salts lacking in CSL, resulting in better butanol
production. From Figure 6.2a, CSL was the best alternative nitrogen source for butanol
fermentation by C. beijerinckii CC101, and soybean meal and cottonseed protein
represented another two alternative candidates suitable for butanol fermentation. Butanol
yields from the alternative nitrogen sources were very similar to each other, generally
around 0.3 g/g, except from the molasses medium, which was around 0.25 g/g with or
without the presence of ammonia acetate (Figure 6.2b). This was probably due to the
poor cell growth in the molasses medium as a result of lacking sufficient nitrogen source.
It was observed that in the SMH medium, butanol yield was higher when ammonia
acetate was not present, which was different from the rest of the studies. The reason for
this is unclear at the moment. From our results (Figure 6.2), it seemed that ammonia
acetate did not have a significant effect on butanol yield, only on boosting butanol
production.

205
6.3.3 ABE production on the bioreactor integrated with gas stripping as product
recovery
In order to study the scale-up of ABE production from WPH, we also examined the
fermentation on the 1.0 L bioreactor. To further improve the sugar conversion and butanol
production, gas stripping was employed as a simultaneous product recovery technique to
integrate with the fermentation to relieve butanol inhibition. Gas stripping has been
reported to only remove volatile solvents, and not harm cells or remove any nutrients
when integrated with the ABE fermentation (Durre, 1998; Ezeji et al., 2004a). From the
serum bottle study, it showed that the sugars present in the WPH could not be completely
utilized by the bacteria due to inhibition. Therefore, diluted 70% WPH was used on the
reactor to reduce sugar level and hydrolysate inhibition on the cells. A total of 12.89 g/L
ABE was produced, of which 9.38 g/L was butanol (Table 6.2, Figure 6.3). ABE yield
was 0.39 g/g and butanol yield was 0.29 g/g. A significant increase of 62.3% in butanol
production was observed in this study, as compared with 5.78 g/L butanol obtained in the
serum bottle study under the same condition. This indicated that butanol was a primary
inhibitor in addition to the inhibitors from WPH during the ABE fermentation. With
in-situ removal of butanol by stripping, the overall butanol production was significantly
improved and the process efficiency was enhanced. Gas stripping was initiated at 24 h
using the fermentation gas (H2 and CO2). Fermentation continued for 75 h, and butanol
concentration was kept below 6g/L throughout the course of fermentation by gas

206
stripping which was below the toxic level to bacteria. As shown in Figure 6.3a and 6.3c,
gas stripping was highly efficient in removing butanol and the recovered butanol was
present at a much higher concentration in the condensate. It was also noticed that acetic
acid and butyric acid were not removed by gas stripping (Figure 6.3c). Only solvents
were recovered by stripping gases, and the butanol was collected at a much higher
concentration than acetone and ethanol, indicating that gas stripping was highly selective
towards butanol. At the beginning of stripping, more butanol was being produced in the
fermentation so more butanol was recovered in the condensate. With the progress of
fermentation, cells gradually lost activity and started to produce butanol at a slower rate,
which resulted in a lower butanol concentration in the condensate. 14 ml condensate was
collected at 35 h (from 24 h to 35 h), 30 ml was collected at 50 h (from 35 h to 50 h), and
45 ml was collected at 75 h (from 50 h to 75). Towards the end of fermentation, more
water was taken from the broth as opposed to the beginning. At the end of fermentation,
9.19 g/L sugars were left, and sugar utilization was improved from 58.2 % in the serum to
78.0 % (Table 6.2). C. beijerinckii CC101 utilized both glucose and xylose at the same
time, and no differentiation between the hexose and pentose was observed. Arabinose
level did not change much throughout the fermentation, indicating that C. beijerinckii
CC101 preferred the other two sugars over arabinose.
Figure 6.4 shows the results of using resin and evaporation detoxified WPH on the
bioreactor with gas stripping. 17.73 g/L ABE was produced, of which 13.46 g/L was

207
butanol. ABE yield was 0.44 g/g and butanol yield was 0.32 g/g (Table 6.2). Gas
stripping was initiated at 36h, and butanol concentration was kept at ~6 g/L throughout
the fermentation course. Initially 54.73 g/L sugar was present, and 13.8 g/L residual sugar
was left in the end, corresponding to a 75% sugar conversion. The fermentation
progressed for 86 h, and no increase in solvent production was observed after 70 h,
indicating that the fermentation was complete at 70 h. Compared with 9.14 g/L butanol
produced from serum bottle study under the same condition, a 47.2% increase in butanol
production was achieved with the help of gas stripping in this case. More sugar was
utilized, increasing the consumption rate from 65.6 % in serum bottle to 74.6 % in this
case. When the toxicity of inhibitors present in the hydrolysate combined with butanol
inhibition, cells suffered a great stress and poor butanol production was resulted
(untreated WPH data in Table 6.2). With butanol removed from the broth, one major
inhibition factor was taken out, leading to better and more successful ABE production.
Even though butanol concentration was kept below toxic level in both studies (70% WPH
and resin detoxified WPH), fermentation stopped before all the sugars could be utilized
and converted to solvents. This indicated that butanol inhibition was not the only factor
that hindered the fermentation, some unknown factors still existed, which was possibly
the residual inhibitors from WPH, accumulation of cell metabolites and dead cells (Ezeji
et al., 2004b). Both studies showed that gas stripping was very successful in improving
the overall solvent production and sugar utilization, and the recovered solvents were

208
concentrated in the condensate stream for easier downstream separation. Gas stripping
has been reported in many literatures to improve the overall solvent production and sugar
conversion, both in batch (Ezeji et al., 2003; Qureshi et al., 2007) and fed-batch
fermentation (Ezeji et al., 2004b; Qureshi et al., 2008c). Qureshi et al. (2008c) showed
that 47.6 g/L total ABE was produced from wheat straw hydrolysate supplemented with
glucose (128.3 g/L) when gas stripping was employed to simultaneously remove butanol.
In this experiment, 3 ml, 30 ml, 22 ml, and 23 ml condensate was collected at 44 h,
60.5 h, 70 h and 86 h. It was shown that at 60.5 h and 70 h, the butanol concentration in
the condensate was around 100 g/L (Figure 6.4c). Butanol saturation point in aqueous
solution was around 8%, which meant that at 100 g/L butanol separated from aqueous
phase and formed a second layer of organic phase. This phenomenon was observed in this
experiment. The upper layer of organic phase was a mixture of acetone and butanol, with
a small amount of dissolved water. The lower layer of aqueous phase was the saturated
solution of acetone and butanol. Ethanol was detected in the condensate at a
concentration around 1 g/L, which was extremely low compared with acetone and
butanol. No acids were detected in the condensate, indicating that gas stripping was
highly selective towards volatile solvents, especially butanol. In the condensate, the
butanol concentration is usually 4-5 folds higher than acetone, as shown in Figure 6.3c
and 6.4c. The phase separation was very valuable for an energy-saving butanol separation
and purification process.

209
6.4 Conclusion
In summary, butanol was successfully produced from wood pulp hydrolysate by C.
beijerinckii CC101 in this study, presenting another renewable substrate for economic
butanol production. Resin and evaporation detoxification was found to be very effective
in removing most of the inhibitors from the hydrolysate, resulting in 11.35 g/L total
solvents in the batch fermentation. With simultaneous butanol recovery by gas stripping,
total solvents were enhanced to 17.73 g/L in the integrated batch fermentation. Besides
corn steep liquor, cottonseed protein and soybean meal represented another two good
candidates as alternative nitrogen sources for butanol fermentation. When ammonia
acetate was supplemented to the medium with alternative nitrogen sources, butanol
production was significantly improved, indicating that ammonia acetate was very
important for successful butanol production. 6-9 g/L butanol was produced from the
hydrolysate using alternative nitrogen sources supplemented with ammonia acetate,
making it very promising to replace the complex and expensive yeast extract.
6.5 References
Bock, S.A., S.L. Fox, and W.R. Gibbons (1997). Development of a low-cost, industrially suitable medium for the production of acetic acid from Clostridium thermoaceticum. Biotechnol. Appl. Bioc., 25, 117-125.
Chen, C.K. and H.P. Blaschek (1999). Acetate enhances solvent production and prevents degeneration in Clostridium beijerinckii BA101. Appl. Microbiol. Biotechnol., 52, 170-173.

210
Dominguez, J.M., C.S. Gong, and G.T. Tsao (1996). Pretreatment of sugar cane bagasse hemicellulose hydrolysate for xylitol production by yeast. Appl. Biochem. Biotech., 57-8, 49-56.
Durre, P. (1998). New insights and novel developments in clostridial acetone/butanol/isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2003). Production of acetone, butanol and ethanol by Clostridium beijerinckii BA101 and in situ recovery by gas stripping. World J. Microbiol. Biotechnol., 19, 595-603.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004a). Butanol fermentation research: Upstream and downstream manipulations. Chem. Rec., 4, 305-314.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004b). Acetone butanol ethanol (ABE) production from concentrated substrate: reduction in substrate inhibition by fed-batch technique and product inhibition by gas stripping. Appl. Microbiol. Biotechnol., 63, 653-658.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2007). Bioproduction of butanol from biomass: from genes to bioreactors. Curr. Opin. Biotech., 18, 220-227.
Ezeji, T. and H.P. Blaschek (2008). Fermentation of dried distillers' grains and solubles (DDGS) hydrolysates to solvents and value-added products by solventogenic clostridia. Bioresour. Technol., 99, 5232-5242.
Ezeji, T., C. Milne, N.D. Price, H.P. Blaschek (2010). Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol., 85, 1697-1712.
Gapes, J.R. (2000). The economics of acetone-butanol fermentation: Theoretical and market considerations. J. Mol. Microbiol. Biotech., 2, 27-32.
Gu, Y., S.Y. Hu, J. Chen, L.J. Shao, H.Q. He, Y.L. Yang, S. Yang, W.H. Jiang (2009). Ammonium acetate enhances solvent production by Clostridium acetobutylicum EA 2018 using cassava as a fermentation medium. J. Ind. Microbiol. Biotechnol., 36, 1225-1232.
Hendriks, A.T.W.M. and G. Zeeman (2009). Pretreatments to enhance the digestibility of

211
lignocellulosic biomass. Bioresour. Technol., 100, 10-18.
Jones, D.T. and D.R. Woods (1986). Acetone-Butanol Fermentation Revisited. Microbiol. Rev., 50, 484-524.
Kadam, K.L. and M.M. Newman (1997). Development of a low-cost fermentation medium for ethanol production from biomass. Appl. Microbiol. Biotechnol., 47, 625-629.
Kumar, M. and K. Gayen (2011). Developments in biobutanol production: New insights. Appl. Ener., 88, 1999-2012.
Kumar, P., D.M. Barrett, M.J. Delwiche, P. Stroeve (2009). Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel Production. Ind. Eng. Chem. Res., 48, 3713-3729.
Larsson, S., A. Reimann, N.O. Nilvebrant, L.J. Jonsson (1999). Comparison of different methods for the detoxification of lignocellulose hydrolysate of spruce. Appl. Biochem. Biotech., 77-79, 91-103.
Lee, S.Y., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentative butanol production by clostridia. Biotechnol. Bioeng., 101, 209-228.
Liu, Z.Y., Y. Ying, F.L. Li, C.Q. Ma, P. Xu (2010). Butanol production by Clostridium beijerinckii ATCC 55025 from wheat bran. J. Ind. Microbiol. Biotechnol., 37, 495-501.
Martinez, A., M.E. Rodriguez, M.L. Wells, S.W. York, J.F. Preston, L.O. Ingram (2001). Detoxification of dilute acid hydrolysates of lignocellulose with lime. Biotechnol. Progr., 17, 287-293.
Mosier, N., C. Wyman, B. Dale, R. Elander, Y.Y. Lee, M. Holtzapple, M. Ladisch (2005). Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96, 673-686.
Mussatto, S.I. and I.C. Roberto (2004a). Alternatives for detoxification of diluted-acid lignocellulosic hydrolyzates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Mussatto, S.I. and I.C. Roberto (2004b). Optimal experimental condition for

212
hemicellulosic hydrolyzate treatment with activated charcoal for xylitol production. Biotechnol. Progr., 20, 134-139.
Nigam, P.S. and A. Singh (2011). Production of liquid biofuels from renewable resources. Progress in Energy and Combustion Science, 37, 52-68.
Parekh, M., J. Formanek, and H.P. Blaschek (1998). Development of a cost-effective glucose corn steep medium for production of butanol by Clostridium beijerinckii. J.l Ind. Microbiol. Biotechnol., 21, 187-191.
Parekh, M., J. Formanek, and H.P. Blaschek (1999). Pilot-scale production of butanol by Clostridium beijerinckii BA101 using a low-cost fermentation medium based on corn steep water. Appl. Microbiol. Biotechnol., 51, 152-157.
Qureshi, N. and H.P. Blaschek (2000). Economics of butanol fermentation using hyper-butanol producing Clostridium beijerinckii BA101. Food Bioprod. Process, 78, 139-144.
Qureshi, N. and H.P. Blaschek (2001). ABE production from corn: a recent economic evaluation. J. Ind. Microbiol. Biotechnol., 27, 292-297.
Qureshi, N. and T.C. Ezeji (2008). Butanol, 'a superior biofuel' production from agricultural residues (renewable biomass): recent progress in technology. Biofuel Bioprod. Bior., 2, 319-330.
Qureshi, N., B.C. Saha, and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioproc. Biosyst. Eng., 30, 419-427.
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta, H.P. Blaschek (2008a). Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.
Qureshi, N., B.C. Sahaa, R.E. Hector, S.R. Hughes, M.A. Cotta (2008b). Butanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part I - Batch fermentation. Biomass Bioenerg., 32, 168-175.
Qureshi, N., B.C. Saha, and M.A. Cotta (2008c). Butanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part II - Fed-batch fermentation. Biomass Bioenerg., 32, 176-183.

213
Qureshi, N., B.C. Saha, B. Dien, R.E. Hector, M.A. Cotta (2010a). Production of butanol (a biofuel) from agricultural residues: Part I - Use of barley straw hydrolysate. Biomass Bioenerg., 34, 559-565.
Qureshi, N., B.C. Saha, R.E. Hector, B. Dien, S. Hughes, S. Liu, L. Iten, M.J. Bowman, G. Sarath, M.A. Cotta (2010b). Production of butanol (a biofuel) from agricultural residues: Part II - Use of corn stover and switchgrass hydrolysates. Biomass Bioenerg., 34, 566-571.
Ravi, V.P., R.V. Jasra, and T.S.G. Bhat (1998). Adsorption of phenol, cresol isomers and benzyl alcohol from aqueous solution on activated carbon at 278, 298 and 323 K. J. Chem. Technol. Biot., 71, 173-179.
Rodrigues, R.C.L.B., M.G.A. Felipe, J.B.A.E. Silva, M. Vitolo, P.V. Gomez (2001). The influence of pH, temperature and hydrolyzate concentration on the removal of volatile and nonvolatile compounds from sugarcane bagasse hemicellulosic hydrolyzate treated with activated charcoal before or after vacuum evaporation. Braz. J. Chem. Eng., 18, 299-311.
Saha, B.C. (2003). Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol., 30, 279-291.
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuel Bioprod. Bior., 2, 553-588.
Weber, C., A. Farwick, F. Benisch, D. Brat, H. Dietz, T. Subtil, E. Boles (2010). Trends and challenges in the microbial production of lignocellulosic bioalcohol fuels. Appl. Microbiol. Biotechnol., 87, 1303-1315.
Zheng, Y.N., L.Z. Li, M. Xian, Y.J. Ma, J.M. Yang, X. Xu, D.Z. He (2009). Problems with the microbial production of butanol. J. Ind. Microbiol. Biotechnol., 36, 1127-1138.

214
Table 6.1 Compositional analysis of the original WPH, detoxified WPHs, CSL, CPH, SMH and molasses used in this study
Glucose Xylose Arabinose Total sugar Acetic acid Formic acid
Wood pulp hydrolysate 20.84 42.74 1.97 65.54 2.95 0.27 Activated carbon detoxified wood pulp hydrolysate
20.15 41.09 1.21 62.45 2.17 0.19
Overlimed wood pulp hydrolysate 17.80 35.71 1.83 55.34 2.46 0.21 Resin and evaporation detoxified wood pulp hydrolysate
11.80 47.18 2.60 61.58 0 0.20
Corn steep liquor 8.99 2.06 0.00 11.05 8.93 0 Cottonseed protein hydrolysate 2.69 2.86 4.20 9.75 0.87 0 Soybean meal hydrolysate 5.83 2.50 1.95 10.29 0.49 0
Glucose Sucrose Fructose Total sugar Acetic acid Formic acid
Molasses 23.86 79.06 30.42 133.34 0 0
214

215
Table 6.2 Comparison and summary of ABE production using different WPHs in batch and integrated batch studies by Clostridium beijerinckii CC101.
Conditions Substrate Initial Sugar (g/L)
Final Sugar (g/L)
Total Sugar conversion
Butanol (g/L)
Total ABE (g/L)
Butanol yield (g/g)
ABE yield (g/g)
50% WPH 31.08 9.74 68.7% 6.53 8.14 0.31 0.38
60% WPH 33.23 11.47 65.5% 5.79 7.83 0.27 0.36
70% WPH 39.84 16.64 58.2% 5.78 7.61 0.25 0.33
Non-treated WPH 54.31 31.14 42.7% 4.48 6.73 0.19 0.29
Overlimed WPH 48.26 27.14 43.8% 4.41 5.83 0.21 0.28
Activated carbon treated WPH
49.50 17.75 64.1% 6.27 8.98 0.20 0.28
Resin and evaporation treated
WPH 44.44 15.27 65.6% 9.14 11.35 0.31 0.39
Serum bottle
Control (glucose/xylose)
38.03 5.47 85.6% 10.59 13.67 0.32 0.42
70% WPH 41.84 9.19 78.0% 9.38 12.89 0.29 0.39 1.0 L bioreactor with gas stripping
resin and evaporation treated
WPH 54.37 13.80 74.6% 13.46 17.73 0.32 0.44
215

Table 6.3 Evaluation of alternative nitrogen sources and ammonia acetate on ABE production using 70%
w/o ammonia acetate w ammonia acetate
Butanol (g/L)
Total ABE (g/L)
Butanol yield (g/g)
ABE yield (g/g)
Butanol (g/L)
Total ABE (g/L)
Butanol yield (g/g)
ABE yield (g/g)
CSL 7.46 8.99 0.35 0.42 8.63 11.21 0.33 0.43 Molasses 2.49 3.34 0.25 0.33 5.72 8.20 0.26 0.37 CSL+Molasses 6.37 7.92 0.32 0.40 9.17 10.57 0.34 0.42 CPH 5.33 6.27 0.29 0.36 7.42 10.53 0.28 0.40 SMH 4.64 6.05 0.34 0.41 5.99 8.64 0.27 0.39 Control P2 N/A N/A N/A N/A 9.51 11.40 0.34 0.41
resin and evaporation detoxified WPH by Clostridium beijerinckii CC101
216
216

217
16(a)
0
2
4
6
8
10
12
14
50% 60% 70% Non-treated Overlimed Activatedcarbon
Resin andevaporation
Control
Co
nce
ntra
tion
(g
/L)
Butanol
ABE
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
50% 60% 70% Non-treated Overlimed Activatedcarbon
Resin andevaporation
Control
Butanol yield (g/g)
ABE yield (g/g)
Sugar conversion (%)
(b)
Figure 6.1 Effect of dilution and different detoxification methods on ABE fermentation by Clostridium beijerinckii CC101 using WPH. (a) Butanol and total ABE production, (b) Butanol yield, ABE yield, and sugar conversion.

0
1
2
3
4
5
6
7
8
9
10
CSL Molasses CSL+M Cottonseedprotein
Soybeanmeal
Control
Bu
tan
ol (
g/L
)
w/o NH4AC
w/ NH4AC
(a)
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
CSL Molasses CSL+M Cottonseedprotein
Soybeanmeal
Control
Bu
tan
ol y
ield
(g
/g)
w/o NH4AC
w/ NH4AC
(b)
Figure 6.2 Effect of alternative nitrogen sources and ammonia acetate on ABE production using 70% resin and evaporation detoxified WPH by Clostridium beijerinckii CC101. (a) Butanol production, (b) Butanol yield.
218
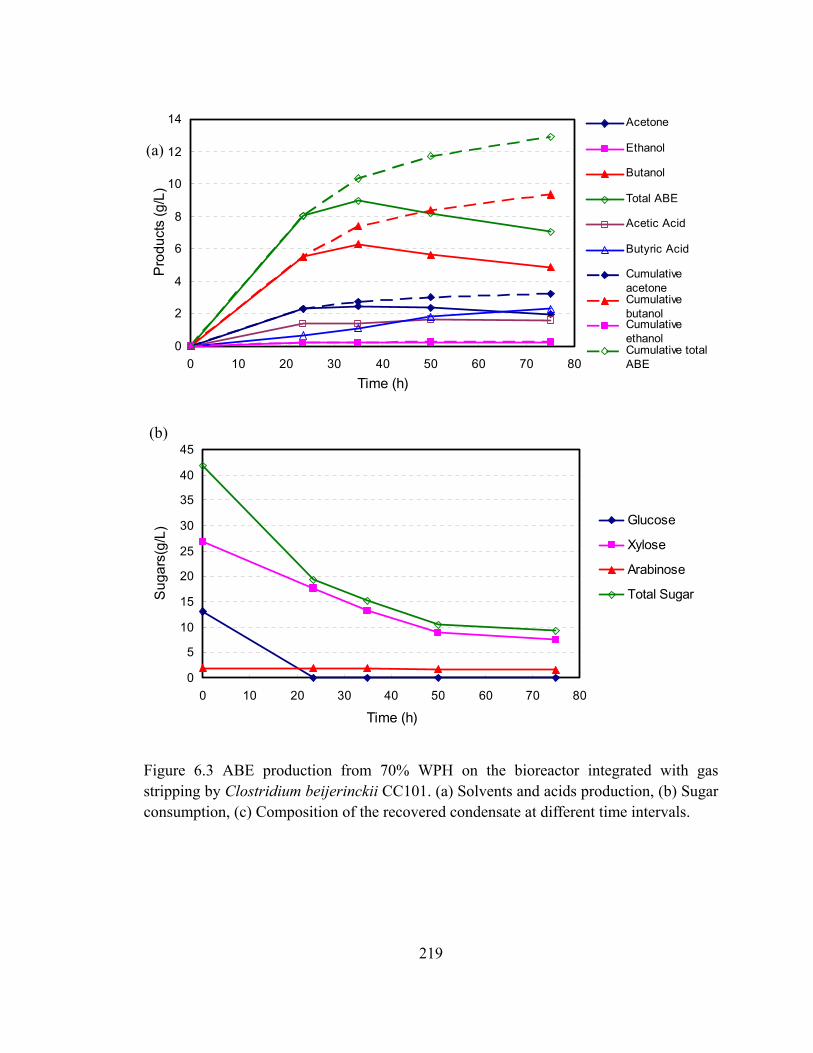
0
2
4
6
8
10
219
12
14
0 10 20 30 40 50 60 70 80
Time (h)
Pro
du
cts
(g/L
)
Acetone
(a) Ethanol
Butanol
Total ABE
Acetic Acid
Butyric Acid
CumulativeacetoneCumulativebutanolCumulativeethanolCumulative totalABE
0
5
10
15
20
25
30
35
40
45
0 10 20 30 40 50 60 70 80
Time (h)
Su
ga
rs(g
/L) Glucose
Xylose
Arabinose
Total Sugar
(b)
Figure 6.3 ABE production from 70% WPH on the bioreactor integrated with gas stripping by Clostridium beijerinckii CC101. (a) Solvents and acids production, (b) Sugar consumption, (c) Composition of the recovered condensate at different time intervals.

Figure 6.3 continued
Condensate
0
10
20
30
40
50
60
70
80
90
35 50 75Time (h)
Con
cent
ratio
n (g
/L)
Acetone
Butanol
Ethanol
(c)
220

0
2
4
6
8
10
12
14
16
18
20
0 10 20 30 40 50 60 70 80 90 100
Time (h)
Pro
du
cts
(g
/L)
Acetone
Ethanol
Butanol
Total ABE
Acetic Acid
Butyric Acid
CumulativeAcetoneCumulativeButanolCumulativeEthanolCumulative ABE
(a)
0
10
20
30
40
50
60
0 10 20 30 40 50 60 70 80 90 100
Time (h)
Su
ga
rs(g
/L)
Glucose
Xylose
Arabinose
Total sugars
(b)
Figure 6.4 ABE production from resin and evaporation detoxified WPH on the bioreactor integrated with gas stripping by Clostridium beijerinckii CC101. (a) Solvents and acids production, (b) Sugar consumption, (c) Composition of the recovered condensate at different time intervals.
221

Figure 6.4 continued
Condensate
0
20
40
60
80
100
120
44 60.5 70 86Time (h)
Co
nce
ntr
atio
n (
g/L
) Acetone
Butanol
Ethanol
(c)
222

223
Chapter 7: Enhanced Biological Butanol Production and Acid Assimilation in ABE
Fermentation using a Recombinant Mutant of Clostridium beijerinckii
Abstract
pSV6 plasmid overexpressing ald, adhE2, ctfA and ctfB genes from sol operon in
Clostridia was constructed in this study. pSV6 plasmid was transformed into C.
beijerinckii CC101 to obtain the recombinant mutant of C. beijerinckii CC101-SV6.
Butanol production and acid assimilation were compared between the parental strain and
the mutant strain. The mutant was found to convert acids produced into solvents more
efficiently than the parental strain under all scenarios evaluated in this study. The effect
of cysteine and butyrate on ABE production was investigated, and the parental strain was
found to be more sensitive to both cysteine and butyrate than the mutant. Cysteine was
found to have a negative impact on butanol production in both parental and mutant strain
study, and led to butyric acid over accumulation in the parental strain study. Significant
acid accumulation and less solvent production were observed with parental strain at the
end of fermentation if butyrate was added at the beginning, whereas most of the butyrate
was converted into butanol by the mutant under the same condition. Sugarcane bagasse

224
hydrolysate was investigated as an alternative substrate for ABE fermentation, and severe
inhibition was observed for the parental strain. The mutant demonstrated robustness and
tolerance in the sugarcane bagasse hydrolysate, with a 9.44 g/L ABE production
compared with 2.57 g/L obtained with parental strain. With the overexpression of
solvent-producing related genes from sol operon, the mutant C. beijerinckii CC101-SV6
stably produced 9.44 – 13.78 g/L solvents under all conditions evaluated, and was less
sensitive to environmental factors than the parental C. beijerinckii CC101.
7.1 Introduction
Butanol is a four carbon chain alcohol, and is currently used as a solvent, chemical
intermediate, and extractant in many areas such as cosmetic and pharmaceuticals (Garćia
et al., 2011). Recently, butanol has attracted more research attention as an alternative
biofuel. Compared to ethanol, butanol is considered as the second generation biofuel due
to the advantages it offers, including high energy content, high research and motor octane
number, low volatility, and low vapor pressure (Dürre, 2007; Lee et al., 2008; Nigam and
Singh, 2011). The fuel properties of butanol are very similar to those of gasoline, and
butanol can be used directly or blended with gasoline and diesel to fuel the automobiles
without any engine modification. In addition, butanol is compatible with the current
transportation pipeline for gasoline (Dürre, 2007; Lee et al., 2008). Currently, butanol is
predominately produced through petrochemical synthesis via Oxo process, which relies

225
on crude oil supply. The alternative route for butanol production is through biological
conversion by acetone-butanol-ethanol (ABE) fermentation. Due to the growing concerns
of environmental issues, depleting fossil resources, and increasing crude oil price, ABE
fermentation has been a popular research topic for economically feasible butanol
production to compete with petrochemical synthetic route (Dürre, 1998; Ezeji et al., 2004;
2007a; Kumar and Gayen, 2011; Lee et al., 2008; Naik et al., 2010).
Many limitations are associated with traditional ABE fermentation, including low
yield, low production, sluggish fermentation, high substrate cost and expensive product
recovery (Ezeji et al., 2010), which significantly impede the economical competitiveness
of biological butanol production through ABE fermentation. Areas under research and
development to improve the efficiency of ABE fermentation are utilization of renewable
and low-cost substrates, development of fermentation processes and alternative product
recovery technologies, and metabolic engineering of solvent-producing microorganisms
(Chernova et al., 2010; Cho et al., 2009; Huang et al., 2010; Ni and Sun, 2009; Pfromm et
al., 2010; Qureshi and Ezeji, 2008; Vane, 2005; 2008). Butanol production from
renewable and sustainable lignocellulosic biomass has been studied in recent years to
replace the traditional substrates such as corn starch and molasses, and many feedstocks,
including corn fiber, corn stover, switchgrass and wheat straw, have been reported as
potential substrates with 9.3-21.4 g/L ABE production (Qureshi et al., 2007; 2008;
2010b). The upper butanol production limit is often stated as 13-15 g/L in conventional

226
ABE batch fermentation with free cells, which is believed to be the butanol-tolerant level
of naturally butanol-producing microorganisms (Dürre, 1998). Strain mutation by
chemical mutagen and subsequent selection has been reported to obtain mutants with
enhanced butanol producing ability (Formanek et al., 1997; Lin and Blaschek, 1983). C.
Beijerinckii BA101 was reported as a hyper-butanol-producing mutant strain from
parental strain of C. Beijerinckii NCIMB 8052 by using
N-methyl-N9-nitro-N-nitrosoguandine mutagen and subsequent selection on glucose
analog 2-deoxyglucose, which had the ability to produce two-folds higher butanol than
parental strain up to 33 g/L total solvents (Formanek et al., 1997; Qureshi and Blaschek,
2001). Many metabolic engineering studies to improve the solvent production, yield, and
butanol selectivity have been also carried out since the entire genomes of C.
acetobutylicum ATCC 824 and C. beijerinckii NCIMB 8052 have been sequenced, which
are two primary naturally butanol-producing bacteria with good butanol titer and yield
(Ezeji et al., 2007a; Lee et al., 2008; Kumar and Gayen, 2011; Papoutsakis, 2008;
Paredes et al., 2004; 2005; Shi and Blaschek, 2008). Some transcriptional regulating
genes, aad (alcohol/aldehyde dehydrogenase), adc (acetoacetate decarboxylase), ctfA and
ctfB (CoA transferase), which formed sol operon in Clostridia, were reported responsible
for the encoding of enzymes for acetone and butanol production, and destruction of these
genes in C. acetobutylicum ATCC 824 led to failure in solvent production (Cornillot et al.,
1997; Nair et al., 1999). ctfA and ctfB were responsible for converting acetate into

227
acetyl-CoA and butyrate to butyryl-CoA, whereas adc and adhE were responsible for
acetone and butanol formation from acetoactate and butyraldehyde (Garćia et al., 2011;
Lee et al., 2008). Overexpressing of adc and ctfAB in C. acetobutylicum led to earlier
induction of acetone formation, with enhanced acetone (95%), butanol (37%), and
ethanol (90%) production (Mermelstein et al., 1993). adhE (aldehyde dehydrogenase),
which was responsible for acetaldehyde and butyraldehyde formation, has been reported
to restore butanol production in C. acetobutylicum M5, which lacked the mega-plasmid
pSOL1 carrying all four genes from sol operon (Nair and Papoutsakis, 1994).
In this study, plasmid pSV6 overexpressing the adhE2 (alcohol/aldehyde
dehydrogenase), ald (aldehyde dehydrogenase), ctfA, and ctfB (CoA transferase) genes
was constructed and inserted into a mutant strain of C. beijerinckii NCIMB 8052, namely
C. beijerinckii CC101, and a recombinant mutant C. beijerinckii CC101-SV6 was
obtained. ABE production from the parental strain and the mutant strain was evaluated
and compared in synthetic glucose-P2 medium, and the effect of gene overexpression on
butanol formation and acid assimilation was explored. Butyrate was investigated as a
co-substrate of its effect on inducing solventogensis and overall butanol production with
both strains. In search for alternative feedstocks for economical ABE production,
sugarcane bagasse hydrolysate (SBH) was evaluated as a potential renewable substrate in
this study. The information provided in this work helped to understand the effect of
solvent-producing genes in Clostridium beijerinckii on butanol production in ABE

228
fermentation.
7.2 Materials and methods
7.2.1 Plasmid construction
Plasmid pSV6 construction was done by Saju Varghese in our research lab at the
Ohio State University. The overexpression of adhE2 gene was under the control of
thiolase promoter, whereas the overexpression of ald, ctfA and ctfB genes were under the
control of fac promoter. Hygromycin B was used as the selection marker in this plasmid.
The detailed description of the pSV6 plasmid is given in Appendix C. This plasmid was
transformed into C. beijerinckii CC101 to obtain mutant C. beijerinckii CC101-SV6.
7.2.2 Strain and inoculum preparation
C. beijerinckii NCIMB 8052 was obtained from ATCC (ATCC number 51743).
After adaption and evolution engineering at our research lab, a mutant strain from C.
beijerinckii NCIMB 8052 was obtained, namely C. beijerinckii CC101, which was used
as the parental strain in this study to obtain the recombinant mutant C. beijerinckii
CC101-SV6. For the inoculum procedure, both parental and mutant strains were treated
the same. Spores of parental and mutant strains were routinely stored in the refrigerator at
4oC in the Clostridia medium. Spores (2 ml) were heat-shocked at 80 oC for 3 min and
transferred to 50 ml RCM growth medium (Difco Reinforced Clostridia Medium, Becton,

229
Dickinson and Company, MD, USA). For growth medium preparation, 3.8 g of solid
RCM powder was dissolved in 100 ml distilled water to prepare 100 ml liquid RCM in a
125 ml serum bottle. The medium was nitrogen-purged for 8 min to remove oxygen. The
serum bottle was tightly capped by rubber stopper and aluminum seal. The mixture was
then autoclaved at 121 oC for 30 min for sterilization followed by cooling to 37 oC. For
the mutant strain, antibiotics hygromycin B (Hygromycin B in PBS, 50 mg/mL,
Invitrogen, USA) was added at 0.4 μl/100 ml medium to ensure the selection of the
mutant strain. The heat-shocked spores were incubated at 37 oC for 12-16 hrs until cells
were highly active. The active culture was used as seed culture for the sequential
fermentation studies with parental and mutant strain, and 5% inoculum was used in all
fermentation studies.
7.2.3 Production medium preparation and fermentation
P2 formula was used in production medium in all ABE fermentation studies.
Sugarcane bagasse hydrolysate (SBH) was kindly provided by ButylFuel LLC.
(Columbus, OH). SBH contained mostly glucose and xylose, and was used as a carbon
source for ABE production. P2 medium contained carbon source (glucose or SBH), yeast
extract (2 g/L), buffer (0.5 g/L KH2PO4 and 0.5 g/L K2HPO4), 2.2 g/L ammonium acetate,
vitamins (0.001 g/L para-amino-benzoic acid (PABA), 0.001g/L thiamin and 10-5 g/L
biotin), and mineral salts (0.2 g/L MgSO4·7H20, 0.01 g/L MnSO4· H20, 0.01 g/L FeSO4·

230
7H20, 0.01 g/L NaCl). The pH of SBH was adjusted to 6.5 using NaOH before use. All
fermentations were carried out in serum bottles containing 50 ml production medium. In
the study to investigate the effect of butyrate, 3 g/L sodium butyrate was added into the
P2-production medium. All the fermentations were carried out in serum bottles
containing 50 ml medium. Carbon source and concentrated P2 stock solution (containing
yeast extract, ammonium acetate, buffer and butyrate (for butyrate study), 10-fold
concentrated) were autoclaved separately at 121oC and 15 psig for 30 minutes for
sterilization to avoid reaction between nitrogen source and carbon source. All minerals
were prepared in a concentration mineral stock solution at 200-fold concentration and
autoclaved at 121oC and 15 psig for 30 minutes for sterilization. The vitamins were
prepared at 1000-fold concentration in vitamin stock solution, and were filtered through
0.2 μm sterile membrane (25mm 0.2μm syringe filter, Fisherbrand, NJ, USA) for
sterilization to avoid denaturalization under autoclave. 0.2% CaCO3 was used in the
serum bottle to ensure the pH staying above 5.0 throughout the fermentation course. All
solutions were nitrogen purged to oxygen free to ensure the anaerobia. Proper amounts of
concentrated P2 stock solution, mineral solution and vitamin solution were aseptically
transferred into a serum bottle containing carbon source by syringe to make the final P2
production medium of the above-mentioned formula. The initial pH of all production
medium was between 6.2 and 6.5.
Actively grown C. beijerinckii CC101 and C. beijerinckii CC101-SV6 cells were

231
inoculated into fermentation media at 5% inoculum. All fermentation was performed at
37 oC with no agitation in the incubator. Due to the addition of CaCO3, no pH adjustment
was made during the fermentation. Samples were taken periodically for analysis of sugar
consumption and ABE production.
7.2.4 Analytical methods
The concentration of sugars (glucose and xylose) was analyzed by a high
performance liquid chromatography (HPLC) with an organic acid column (Bio-Rad
HPX-87, ion exclusion organic acid column, 300 mm × 7.8mm). Samples were
centrifuged at 13.2 g for 5 min in microcentrifuge tubes and diluted 10 times with
distilled water prior to analysis on HPLC. HPLC was run at 45 oC using 0.01N H2SO4 as
the eluent at a flow rate of 0.6 ml/min. 15μL sample was injected by an automatic injector
(SIL-10Ai) and the running time was set at 36 min. A refractive index (RI) detector
(Shimadzu RID-10A) was set at the range of 200 to detect the organic compounds in the
sample. The HPLC column was installed in a column oven (CTO-10A) with temperature
control at 45 oC. Peak height was used to calculate concentration of sugars in the sample
based on the peak height of the standard sample.
Glucose concentration was also measured by a glucose and lactate analyzer, YSI
biochemistry analyzer (2700 Select). The fermentation products, acetone, butanol,
ethanol, acetic acid, and butyric acid, were measured with a Shimadzu GC-2014 gas

232
chromatograph (GC) (Shimadzu, Columbia, MD, USA) equipped with a flame ionization
detector (FID) and a 30.0 m fused silica column (0.25m film thickness and 0.25 mm ID,
Stabilwax-DA). To reduce the injection mechanical error margin, internal standard
method was used to analyze the concentration of products in the samples. Isobutanol and
isobutyric acid were used as internal standards for the solvent products and acid products
present in the samples. An internal standard buffer solution containing 0.5 g/L isobutanol,
0.1 g/L isobutyric acid, and 1% phosphoric acid was used to dilute each sample 20 times
for acidification and calibration prior to analysis on GC. The gas chromatograph was
operated at an injection temperature of 200 oC with 1 μL of the acidified sample injected
by the AOC-20i Shimadzu auto injector. Column temperature was held at 80 oC for 3 min,
raised to 150 oC at a rate of 30 oC/min, and held at 150 oC for 3.7 min.
7.3 Results and discussion
7.3.1 ABE production from glucose-P2 medium with parental and mutant strains
Mutant strain C. beijerinckii CC101-SV6 and parental strain C. beijerinckii CC101
were investigated for butanol production with synthetic glucose-P2 medium. The results
are shown in Figure 7.1. Parental strain C. beijerinckii CC101 produced 9.8 g/L butanol
within 68 h, compared with 11.3 g/L butanol obtained from mutant C. beijerinckii
CC101-SV6. The initial glucose concentration present in the medium was 36.0 and 36.6
g/L, and 11.5 g/L and 6.3 g/L residual glucose were observed for C. beijerinckii CC101

233
and CC101-SV6 at the end of fermentation, respectively. This indicated that the mutant
strain utilized more glucose than parental strain, and converted additional glucose into
15% more butanol production than parental strain. At the end of fermentation, 3.4 g/L
total residual acids (1.6 g/L acetic acid and 1.8 g/L butyric acid) were observed from C.
beijerinckii CC101, whereas 1.5 g/L total acids (0.7 g/L acetic acid and 0.8 g/L butyric
acid) were left in the broth with C. beijerinckii CC101-SV6. This result showed that with
the overexpression of solventogenic genes of sol operon, the mutant strain assimilated
acids better than parental strain, and efficiently converted acetic acid and butyric acid into
solvents. ctfA and ctfB genes encoded for enzymes of CoA trasnferase, which was
responsible for converting acetate to acetyl-CoA and butyrate to butyryl-CoA in the
metabolic pathway of Clostridia (Garćia et al., 2011).
A comparison of butanol and acid production from parental strain and mutant strain
is shown in Figure 7.2. During the acidogensis stage, acetic acid and butyric acid were
produced by both strains, and total acid levels were 3.6 g/L and 3.4 g/L at 18 h for
parental and mutant strain, respectively. After the bacteria entered solventogensis stage,
acids were quickly assimilated by C. beijerinckii CC101-SV6 and converted into solvent,
whereas the acid level only decreased slightly for C. beijerinckii CC101, as shown in
Figure 7.2b. Butanol production was associated with acid production and assimilation,
and after 18 h, butanol production from the mutant strain was always higher than parental
strain due to successful conversion of acids into solvents, as is evidently shown and

234
compared in Figure 7.2a. As shown in Table 7.1, butanol yield was very similar between
the two strains, which were 0.39 g/g and 0.38 g/g for parental and mutant strain,
respectively. Total ABE of 11.3 g/L were obtained from parental strain, compared with
13.78 g/L ABE produced from mutant strain.
It was noticed that the butanol/acetone ratio was significantly higher for both
parental and mutant strain in this study. Only 1-2 g/L acetone was produced with around
10g/L butanol production in this study with synthetic glucose-P2 medium, corresponding
to ~ 5:1 butanol/acetone ratio. The typical butanol/acetone ratio for clostridia species was
2:1. C. beijerinckii NCIMB 8052 was reported to have a typical butanol/acetone ratio of
2:1 in a 20.0-liter pilot scale study using glucose as substrate, while the mutant strain C.
beijerinckii BA101 had an enhanced butanol/acetone ratio of 3:1 (Parekh et al., 1999).
The minerals used in the medium were reported to influence the butanol production and
butanol/acetone ratio, and a high butanol/acetone ratio of 4.0 was previous reported in a
glucose-corn steep water medium supplemented with several minerals by C. beijerinckii
BA101 (Parekh et al., 1998). The butanol/acetone ratio in their study was between 1.5 to
3.9 under other scenarios with different nutrient and minerals supplementation (Parekh et
al., 1998). The parental strain used in this study, C. beijerinckii CC101, was a mutant
strain obtained from C. beijerinckii NCIMB 8052 after adaption and selection. C.
beijerinckii CC101 had higher selectivity in butanol production than acetone formation as
shown in this study, but the exact reason for this phenomenon was unclear at the moment.

235
Mermelstein et al. (1993) previously overexpressed the genes of adc, ctfA and ctfB
from the sol operon in C. acetobutylicum ATCC 824, and acetone and butanol production
were enhanced by 90% and 37%, respectively. The butanol/ABE ratio reached over 0.80
in this study for both parental and mutant strain. A similar butanol/ABE ratio (4:5) was
reported by Jiang et al. (2009) by inactivating the gene encoded for acetoacetate
decarboxylase. 14 g/L butanol was obtained with only 0.3 g/L acetone production under
pH controlled conditions.
7.3.2 Effect of cysteine addition on ABE production
Cysteine is an amino acid with reducing power. Butanol is a reducing product in
ABE synthesis by Clostridia bacteria from pyruvate, and the intracellular ATP, NAD+,
and NADH level all affect the biosynthesis of butanol in the solventogenic clostridia
(Ezeji et al., 2010). Adding cysteine into the fermentation medium can lower the
redox-potential of the system, increasing the possibility for the formation of reducing
products such as butanol. Therefore, the effect of cysteine on butanol production from
ABE fermentation was investigated in this study, and the results are shown in Figure 7.3
and Table 7.1. 6.18 g/L butanol was obtained from glucose-P2medium containing 0.5 g/L
cysteine with parental strain, whereas 9.38 g/L butanol was obtained from the mutant
strain under the same condition. Compared with 9.84 g/L butanol and 11.28 g/L butanol
obtained from glucose-P2 control fermentations, the addition of cysteine adversely

236
affected the butanol production, corresponding to 37.2 % and 16.8 % reduction in butanol
production for parental strain and mutant strain, respectively. It was noticed that the
addition of cysteine had less effect on the mutant strain than the parental strain, as
evidenced by both butanol production and acid assimilation (Figure 7.4). Comparing
results from glucose-P2 and glucose-P2 with cysteine (Figure 2 and Figure 7.4), acid
assimilation from the parental strain and the mutant strain was very similar under both
conditions, and the mutant strain was more efficient in converting the acids into solvents.
Total acids accumulated to a maximum 2.82 g/L at 18 h in the mutant study with cysteine
present in the medium, and decreased to 1.69 g/L at the end of fermentation, whereas
total acids steadily increased throughout the fermentation course with 4.15 g/L left at the
end in the parental strain study with cysteine present (Figure 7.4). In the glucose-P2
medium, the acids level reached a maximum of 3.96 g/L at 28 h, and slightly decreased to
3.39 g/L at the end in the parental strain study (Figure 7.2). This indicated that the
parental strain could not efficiently convert the acids in both conditions, and the addition
of cysteine negatively impacted acid assimilation in the parental strain study, whereas the
acid conversion in the mutant strain study was not affected by this external factor and
remained efficient regardless of the redox potential of the system. The addition of
cysteine did not improve the butanol production as expected, which was probably due to
the disruption of the sensitive balance of NAD+/NADH inside the cells. Lowering the
redox potential of the system may favor the butyric acid formation pathway of

237
Clostridium beijerinckii since NADH can only be oxidized in this pathway and
regenerate NAD+ (Ezeji et al., 2010). This phenomenon was observed in the parental
strain study with cysteine present in the medium. The residual butyric acid with cysteine
present in the medium was 2.60 g/L at the end of fermentation, compared to 1.65 g/L left
in the medium without cysteine (Figure 7.1a and Figure 7.3a). The residual butyric acid
level in the mutant strain studies was similar, which was 0.84 g/L and 0.73 g/L with or
without cysteine addition, respectively. This was probably due to the better acid
assimilation mechanism in the mutant with the overexpression of acid converting genes.
Based on the results of the parental strain, over accumulation of butyric acid was toxic
and inhibitory to Clostridium beijerinckii if the butyric acid could not be rapidly
converted to butanol. Even though the formation of butyric acid was important in
maintaining the redox equilibrium between NADH and NAD+, the presence of acetic acid
and a small amount of ABE was necessary to induce the solventogensis from acidogensis
(Shi and Blaschek, 2008). Although it was found in this study that addition of reducing
agent cysteine did not improve butanol production and led to an accumulation of butyric
acid in the medium, other reducing agents such as viologen and carbon monoxide have
been reported to be effective in facilitating butanol production by altering electron flow
and favoring butyrate uptake (Meyer et al., 1986; Rao and Mutharasan, 1987; Tashiro et
al., 2007).

238
7.3.3 Effect of butyric acid addition on ABE production
Butyric acid is an intermediate product in the pathway of solvent-producing
Clostridia bacteria, which is produced during acidogensis and converted into butanol
during solventogensis (Lee et al., 2008). It has been reported in the literature that feeding
butyric acid with glucose as a co-substrate in the medium facilitated butanol production
by reducing acidogensis and inducing solventogensis early (Bahl et al., 1982; Geng et al.,
1995; Huang et al., 2004; Tashiro et al.,2004). Therefore, butyric acid was evaluated in
this study of its effect on ABE production from C. beijerinckii CC101 and C. beijerinckii
CC101-SV6. 3 g/L sodium butyrate was added in the synthetic glucose-P2 medium. The
results are shown in Figure 7.5 and Table 7.1. Figure 7.5 also included the results
obtained from cysteine addition, so that both the effect of cysteine and butyrate on ABE
production could be compared with control glucose-P2 medium.
Parental strain produced 8.1 g/L butanol with a yield of 0.27 g/g in the P2-butyrate
medium, whereas mutant strain produced 9.3 g/L butanol with a yield of 0.33 g/g under
the same condition. Compared with results from glucose-P2 medium, the butanol
production from P2-butyrate medium was lower, which was probably due to the
inhibition caused by initial butyrate in the medium. The butanol yield for both strains was
lower in the P2-butyrate medium than P2 medium, which was a sign of inhibition. It
seemed that C. beijerinckii CC101 was very sensitive to butyrate; the butanol production
and yield was affected even at concentration as low as 3 g/L. This was evidenced by acid

239
accumulation at the end of fermentation. From Figure 7.5b, the residual acids in C.
beijerinckii CC101 and C. beijerinckii CC101-SV6 were 5.8 g/L and 1.4 g/L, respectively,
in the P2-butyrate medium. Compared with P2 medium, a significant accumulation of
acids was observed with parental strain C. beijerinckii CC101, indicating that the initial
butyrate present in the medium could not be utilized and converted to butanol by the
parental strain. The residual acids in the P2-butyrate medium with mutant strain were
similar to the results in P2 medium, which meant that the uptake of butyric acid was
successful and the mutant was able to efficiently convert the acid into butanol. This was
attributed to the overexpression of ctfB, ald and adhE2 genes from sol operon, which first
converted butyrate into butyryl-CoA then to butyraldehyde. Butyric acid was reported as
an important inducer for genes expression to produce enzymes associated with butanol
synthesis in C. acetobutylicum at a concentration of 13-18 mM (Terracciano and Kashket,
1986). It has been reported that intracellular metabolic precursors, butyryl-CoA and
butyrylphosphate, played an important role during the phase shift from acidogensis and
solventogensis (Harris et al., 2000).The shift from acidogensis to solventogensis was a
very complicated process, which was regulated by many genes and triggered by a series
of signals. The expression of the relating genes was the key to ensure the successful shift
between two phases and solvent production, which could be affected by many internal
and external factors. If the shift between the two phases failed, it would lead to
accumulation of acids and poor solvent production, and eventually the fermentation

240
would be stopped due to acid inhibition. The promoter used on the pSV6 plasmid in the
mutant was constitutive, which meant that the expression of these solvent-producing
genes was not controlled by those complex factors any longer. This feature helped to
ensure the stable solvent production in the mutant strain, and made the solvent production
process less sensitive to environmental factors. It also facilitated the assimilation of acids,
naturally produced or manually provided, and converted these acids into solvents,
improving solvent yield.
7.3.4 ABE production from renewable lignocellulosic substrates
Lignocellulosic feedstocks have been studied extensively for butanol production as
renewable and sustainable green substrates, such as wheat straw, corn fiber, corn stover
and barley straw (Ezeji and Blaschek, 2008; Qureshi and Ezeji, 2008; Qureshi et al., 2007;
2008; 2010a; 2010b). However, the inhibition from lignocellulosic hydrolysate on the
solvent-producing microorganisms was severe due to the degradation products generated
during pretreatment process to convert hemicellulose and cellulose into fermentable
sugars, resulting in poor solvent production and cell growth (Ezeji et al., 2007b). In this
study, sugarcane bagasse hydrolysate (SBH) was investigated as a potential
lignocellulosic substrate for butanol production with parental strain C. beijerinckii CC101
and mutant strain C. beijerinckii CC101-SV6. The sugarcane bagasse received contained
69.0 g/L glucose, 32. 7g/L xylose, 0.48 g/L lactic acid, 6.8 g/L acetic acid, 0.3 g/LHMF

241
and 0.9 g/L furfural. Due to the high sugar concentration present in the SBH, it was
diluted to proper sugar level similar to that used in synthetic P2 medium to avoid
substrate inhibition and relieve inhibition of SBH on the bacteria. The results are shown
in Figure 7.6 and Table 7.1.
2.05 g/L and 7.63 g/L butanol were obtained from parental and mutant strain in the
SBH medium, respectively. Initially, 18.8 g/L glucose and 10.3 g/L xylose were present
in the medium (29.1 g/L total sugar available). At the end of fermentation, 19.9 g/L
residual sugar was left in the SBH medium with parental strain, whereas only 3.1 g/L
residual sugar was observed with the mutant strain, indicating that most of the sugars
from SBH were utilized by the mutant. The sugar conversion was 31.3% and 89.5% for
parental strain and mutant strain, respectively. ABE fermentation was severely inhibited
with the parental C. beijerinckii CC101, as was evidenced by both butanol production
and sugar utilization. This was probably due to SBH inhibition. Some major fermentation
inhibitors from lignocellulosic hydrolysate were furfural, HMF and phenolic compounds,
which were sugar and lignin degradation products during pretreatment (Hendriks and
Zeeman, 2009; Moiser et al., 2005; Mussatto and Roberto, 2004). These compounds have
been reported to affect and inhibit ABE production from C. beijerinckii BA101 (Ezeji et
al., 2007b), which was also a mutant from C. beijerinckii NCIMB 8052. Due to the acetic
acid present in the original SBH, an initial 1.8 g/L acetic acid was present in the medium
for both parental and mutant strains. At the early stage of fermentation, acetic acid level

242
increased to 5.8 g/L and 5.9 g/L in parental strain and mutant strain fermentations during
acidogensis. This high level of acetic acid combined with inhibition from SBH could
adversely impact on C. beijerinckii CC101 in ABE fermentation. From 20 h to 30 h, there
was a sharp decrease in acetic acid level in both fermentations, indicating a shift from
acidogensis to solventogensis (Figure 7.6a and 7.6b). However, the acids level in the
parental strain study was still high during the solventogensis, with an average of 2.8 g/L
acetic acid and 2.3 g/L butyric acid throughout the fermentation course. The total acid
level in the mutant medium was kept under 2 g/L, with an average 1.0 g/L acetic acid and
0.5 g/L butyric acid in the solventogensis stage. The butyric acid level in the parental
strain fermentation was 4-folds higher than that from mutant fermentation, indicating that
butyric acid could not be converted to butanol due to inhibition. This showed that the
mutant strain had the ability to efficiently assimilate and convert the acids into solvents,
whereas the parental strain was inhibited by acid accumulation and inhibition of SBH and
failed to convert acids into solvents. A total 9.44 g/L ABE was obtained from SBH using
mutant strain C. beijerinckii CC101-SV6, compared with 2.57 g/L ABE from parental
strain C. beijerinckii CC101. This result showed that the mutant strain was more robust
and inhibition-tolerant than the parental strain in the toxic lignocellulosic hydrolysate
medium. Acid production was associated with cell growth during the exponential phase
for Clostridia, whereas solvent production, which was secondary products, was regulated
and controlled by many genes and conditions. Comparing the results obtained with

243
parental and mutant strains in glucose-P2 medium and SBH medium, the mutant strain
was less affected by environmental factors and was capable of stably producing solvents,
demonstrating superiority to parental strain under both non-stressed and stressed
conditions.
7.4 Conclusion
In this study, a recombinant C. beijerinckii CC101-SV6 mutant overexpressing ald,
adhE2, ctfA and ctfB genes from sol operon, was evaluated and compared with parental
strain C. beijerinckii CC101. The mutant demonstrated superiority in butanol production
and acids assimilation under all conditions evaluated. The mutant can efficiently convert
the acids produced into corresponding solvents, resulting in higher solvent production
than parental strain under all scenarios investigated in this study. Reducing agent cysteine
was shown to have a more negative effect on butanol formation in the parental strain
study than in the mutant strain study, and led to butyric acid over-accumulation in the
parental strain which lacked efficient acid conversion mechanism compared to the mutant.
The parental strain C. beijerinckii CC101 was found to be very sensitive to butyrate, and
initial addition of butyrate to the medium adversely affected the butanol production from
parental strain with significant acids accumulation at the end of fermentation. On the
contrast, the mutant C. beijerinckii CC101-SV6 was less sensitive and converted initial
butyrate into more butanol under the same condition. The mutant was more robust and

244
inhibition-tolerant than the parental strain in the sugarcane bagasse hydrolysate, which
was toxic and inhibitory to the parental strain due to presence of fermentation inhibitors.
Stable solvent production of 13.78 and 9.44 g/L were obtained with the mutant strain
using glucose and sugarcane bagasse, respectively, whereas only 2.57 g/L solvent was
produced by parental strain with sugarcane bagasse hydrolysate compared with 11.3 g/L
solvents production with glucose.
7.5 References
Chernova, N.I., T.P. Korobkova, and S.V. Kiseleva (2010). Use of biomass for producing liquid fuel: Current state and innovations. Thermal Eng., 57, 937-945.
Cho, D.H., Y.H. Lee, Y. Um, B.I. Sang, Y.H. Kim (2009). Detoxification of model phenolic compounds in lignocellulosic hydrolysates with peroxidase for butanol production from Clostridium beijerinckii. Appl. Microbiol Biotechnol., 83, 1035-1043.
Cornillot, E., R.V. Nair, E.T. Papoutsakis, P. Soucaille (1997). The gene for butanol and acetone formation in Clostridium acetobutylicum ATCC 824 reside on a large plasmid whose loss leads to degeneration of the strain. J. Bacteriol., 179, 5442-5447.
Dürre, P. (1998). New insights and novel developments in clostridial acetone/butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Dürre, P. (2007). Biobutanol: An attractive biofuel. Biotechnol. J., 2, 1525-1534.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004). Butanol fermentation research: upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007a). Bioproduction of butanol from biomass: from genes to bioreactors. Current Opinion in Biotechnol., 18, 220-227.

245
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007b). Butanol production from agricultural residues: impact of degradation products on Clostridium beijerinckii growth and butanol fermentation. Biotechnol. Bioeng., 97, 1460-1469.
Ezeji, T.C. and H.P. Blaschek (2008). Fermentation of dried distillers’ grains and solubles (DDGS) hydrolysates to solvents and value-added products by solventogenic clostridia. Bioresour. Technol., 99, 5232-5242.
Ezeji, T.C., C. Milne, N.D. Price, H.P. Blaschek (2010). Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol., 85, 1697-1712.
Formanek, J., R. Mackie, H.P. Blaschek (1997). Enhanced butanol production by Clostridium beijerinckii BA101 grown in semidefined P2 medium containing 6 percent maltodextrin or glucose. Appl. Environ. Microbiol., 63, 2306-2310.
Garćia, V., J. Päkkilä, H. Ojamo, E. Muurinen, R.L. Keiski (2011). Challenges in biobutanol production: How to improve the efficiency. Renew. Sustain. Ener. Reviews, 15, 964-980.
Geng, Q.H., C.H. Park and K. Janni (1995). Uptake of organic-acids by Clostridium acetobutylicum B18 under controlled pH and reduced butanol inhibition. Korean J. Chem. Eng., 12, 378-383.
Harris, L.M., R.P. Desai, N.E. Welker, E.T. Papoutsakis (2000). Characterization of recombinant strains of the Clostridium acetobutylicum butyrate kinase inactivation mutant: need for new phenomenological models for solventogensis and butanol inhibition? Biotechnol. Bioeng., 67, 1-11.
Hendriks A.T.W.M. and G. Zeeman (2009). Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresour. Technol., 100, 10-18.
Huang, W.C., D.E. Ramey, and S.T. Yang (2004). Continuous production of butanol by Clostridium acetobutylicum immobilized in a fibrous bed bioreactor. Appl. Biochem. Biotechnol., 113-116, 887-898.
Huang, H., H. Liu, Y.R. Gan (2010). Genetic modification of critical enzymes and involved genes in butanol biosynthesis from biomass. Biotech. Adv., 28, 651-657.
Jiang, Y., C. Xu, F .Dong, Y. Yang, W. Jiang, S. Yang (2009). Disruption of the

246
acetoacetate decarboxylase gene in solvent-producing Clostridium acetobutylicum increases the butanol ratio. Metab. Eng., 11, 284-291.
Kumar, M. and K. Gayen (2011). Developments in biobutanol production: New insights. Appl. Ener., 88, 1999-2012.
Lin, Y.L. and H.P. Blaschek (1983). Butanol production by a butanol-tolerant strain of Clostridium acetobutylicum in extruded corn broth. Appl. Environ. Microbiol., 45, 966-973.
Lee, S.T., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentive butanol production by Clostridia. Biotechnol. Bioeng., 101,209-228.
Mermelstein, L.D., E.T. Papoutsakis, D.J. Petersen, G.N. Bennett (1993). Metabolic engineering of Clostridium acetobutylicum ATCC 824 for increased solvent production by enhancement of acetone formation enzyme-activities using a synthetic acetone operon. Biotechnol. Bioeng., 42, 1053-1060.
Meyer, C.L., J.W. Roos and E.T. Papoutsakis (1986). Carbon monoxide gassing leads to alcohol production and butyrate uptake without acetone formation in continuous cultures of Clostridium acetobutylicum. Appl. Microbiol. Biotechnol., 24, 159-167.
Mosier, N.S., C. Wyman, B.Dale, R.Elander, Y.Y. Lee, M. Holtzapple and M.R. Ladisch (2005). Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96, 673-668.
Mussatto, S.I. and I.C. Roberto (2004). Alternatives for detoxification of diluted-acid lignocellulosic hydrolysates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Naik, S.N., V.V. Goud, P.K. Rout, A.K. Dalai (2010). Production of first and second generation biofuels: a comprehensive review. Renew. Sust. Energy Rev., 14, 578-597.
Nair, R.V. and E.T. Papoutsakis (1994). Expression of plasmid-encoded aad in Clostridium acetobutylicum M5 restores vigorous butanol production. J. Bacteriol., 176, 5843-5846.
Nair, R.V., E.M. Green, D.E. Watson, G.N. Bennett, Papoutsakis (1999). Regulation of the sol locus genes for butanol and acetone formation in Clostridium acetobutylicum ATCC 824 by a putative transcriptional repressor. J. Bacteriol., 181, 319-330.

247
Ni, Y. and Z. Sun (2009). Recent progress on industrial fermentative production of acetone-butanol-ethanol by Clostridium acetobutylicum in China. Appl. Microbiol. Biotechnol., 83, 415-423.
Nigam, P.S. and A. Singh (2011). Production of liquid biofuels from renewable resources. Prog. Ener. Combust. Sci., 37, 52-68.
Papoutsakis, E.T. (2008). Engineering solventogenic clostridia. Curr. Opin. Biotechnol., 19, 420-429.
Paredes, C., I. Rigoutsos, and E.T. Papoutsakis (2004). Transcriptional organization of the Clostridium acetobutylicum genome. Nucleic Acid Res., 32, 1973-1981.
Paredes, C., K.V. Alsaker, and E.T. Papoutsakis (2005). A comparative genomic view of clostridial sporulation and physiology. Nat. Rev., 3, 969-978.
Parekh, M., J. Formanek, and H.P. Blaschek (1998). Development of cost-effective glucose-corn steep medium for production of butanol by Clostridium beijerinckii. J. Ind. Microbiol. Biotechnol., 21, 187-191.
Parekh, M., J. Formanek, and H.P. Blaschek (1999). Pilot-scale production of butanol by Clostridium beijerinckii BA101 using a low-cost fermentation medium based on corn steep water. Appl. Microbiol. Biotechnol., 51, 152-157.
Pfromm, P.H., V. Amanor-Boadu, R. Nelson, P. Vadlani, R. Madl (2010). Bio-butanol vs. bio-ethanol: a technical and economic assessment for corn and switchgrass fermented by yeast or Clostridium acetobutylicum. Biomass Bioenergy, 34, 515-524.
Qureshi, N. and H.P. Blaschek (2001). Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 27, 287-291.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst. Eng., 30, 419-427.
Qureshi, N. and T.C. Ezeji (2008). Butanol, ‘a superior biofuel’ production from agricultural residues (renewable biomass): recent progress in technology. Biofuels, Bioprod. Bioref., 2, 319-330.
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta and H.P. Blaschek (2008).

248
Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.
Qureshi, N., B.C. Saha, B. Dien, R.E. Hector, M.A. Cotta (2010a). Production of butanol (a biofuel) from agricultural residues: Part I-Use of barley straw hydrolysate. Biomass Bioenergy, 34, 559-565.
Qureshi, N., B.C. Saha, R.E. Hector, B. Dien, S. Hughes, S. Liu, L. Iten, M.J. Bowman, G. Sarath, M.A. Cotta (2010b). Production of butanol (a biofuel) from agricultural residues: Part II-Use of corn stover and switchgrass hydrolysates. Biomass Bioenergy, 34, 566-571.
Rao G. and R. Mutharasan (1987). Altered electron flow in continuous cultures of Clostridium acetobutylicum induced by viologen dyes. Appl. Environ. Microbiol., 53, 1232-1235.
Shi, Z. and H.P. Blaschek (2008). Transcriptional analysis of Clostridium beijerinckii NCIMB 8052 and they hyper-butanol-producing mutant BA101 during shift from acidogensis to solventogensis. Appl. Environ. Microbiol., 74, 7709-7714.
Swana, J., Y. Yang, M. Behnam, R. Thompson (2011). An analysis of net energy production and feedstock availability for biobutanol and bioethanol. Bioresour. Technol., 102, 2112-2117.
Tashiro, Y., K. Takeda, G. Kobayashi, K. Sonomoto, A. Ishizaki, S. Yoshino (2004). High butanol production by Clostridium saccharoperbuutylacetonicum N1-4 in fed-batch culture with pH-stat continuous butyric acid and glucose feeding method. J. Biosci. Bioeng., 98, 263-268.
Tashiro, Y., H. Shinto, M. Hayshi, S. Baba, G. Kobayashi, K. Sonomoto (2007). Novel high-efficient butanol production from butyrate by non-growing Clostridium saccharoperbutyl acetonicum N1-4 (ATCC 13564) with methyl viologen. J. Biosci. Bioeng., 104, 238-240.
Terracciano, J.S. and E.R. Kashket (1986). Intracellular conditions required for the initiation of solvent production by Clostridium acetobutylicum. Appl. Environ. Microbiol., 52, 86-91.
Vane, L.M. (2005). Review: A review of Pervaporation for product recovery from biomass fermentation processes. J. Chem. Technol. Biotechnol., 80, 603-629.

249
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuls, Bioprod. Bioref., 2, 553-588.

250
Substrate
Glucose Glucose + cysteine Glucose + butyrate Sugarcane bagasse hydrolysate
Parental Mutant Parental Mutant Parental Mutant Parental Mutant
Acetone (g/L) 1.25 2.31 1.07 2.26 2.42 2.5 0.34 1.69
Ethanol (g/L) 0.21 0.2 0.11 0.16 0.39 0.28 0.18 0.12
Butanol (g/L) 9.84 11.27 6.18 9.38 8.14 9.25 2.05 7.63
ABE (g/L) 11.30 13.78 7.36 11.80 10.95 12.03 2.57 9.44
Butanol yield (g/g) 0.39 0.38 0.34 0.38 0.27 0.33 0.23 0.29
ABE yield (g/g) 0.45 0.46 0.41 0.47 0.36 0.43 0.28 0.36
Butanol productivity (g/L. h) 0.19 0.22 0.10 0.17 0.12 0.13 0.04 0.14
ABE productivity (g/L. h) 0.22 0.27 0.12 0.21 0.16 0.17 0.05 0.17
Residual acids (g/L) 3.39 1.51 4.14 1.69 5.77 1.38 5.30 1.81
Initial sugar (g/L) 36.6 36.0 36 35.8 37.3 35.9 29.0 29.3
Final sugar (g/L) 11.5 6.3 18.1 10.9 7.2 8.0 19.9 3.1
Sugar conversion 68.6% 82.5% 49.8% 69.4% 80.7% 77.8% 31.3% 89.5%
Table 7.1 Comparison and summary of the performance of parental strain and mutant strain under all the scenarios evaluated in this study.
250

0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60 70
Time (h)
Glu
cose
(g
/L)
0
2
4
6
8
10
12
So
lve
nts
an
d a
cid
s (g
/L)
Glucose
Acetone
Ethanol
Butanol
Acetic Acid
Butyric Acid
(a)
0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60 70
Time (h)
Glu
cose
(g
/L)
0
2
4
6
8
10
12
So
lve
nts
an
d a
cid
s (g
/L)
Glucose
Acetone
Ethanol
Butanol
Acetic Acid
Butyric Acid
(b)
Figure 7.1 Fermentation kinetics of C. beijerinckii CC101 and C. beijerinckii CC101-SV6 in synthetic glucose-P2 medium. (a) ABE production of C. beijerinckii CC101, (b) ABE production of C. beijerinckii CC101-SV6.
251
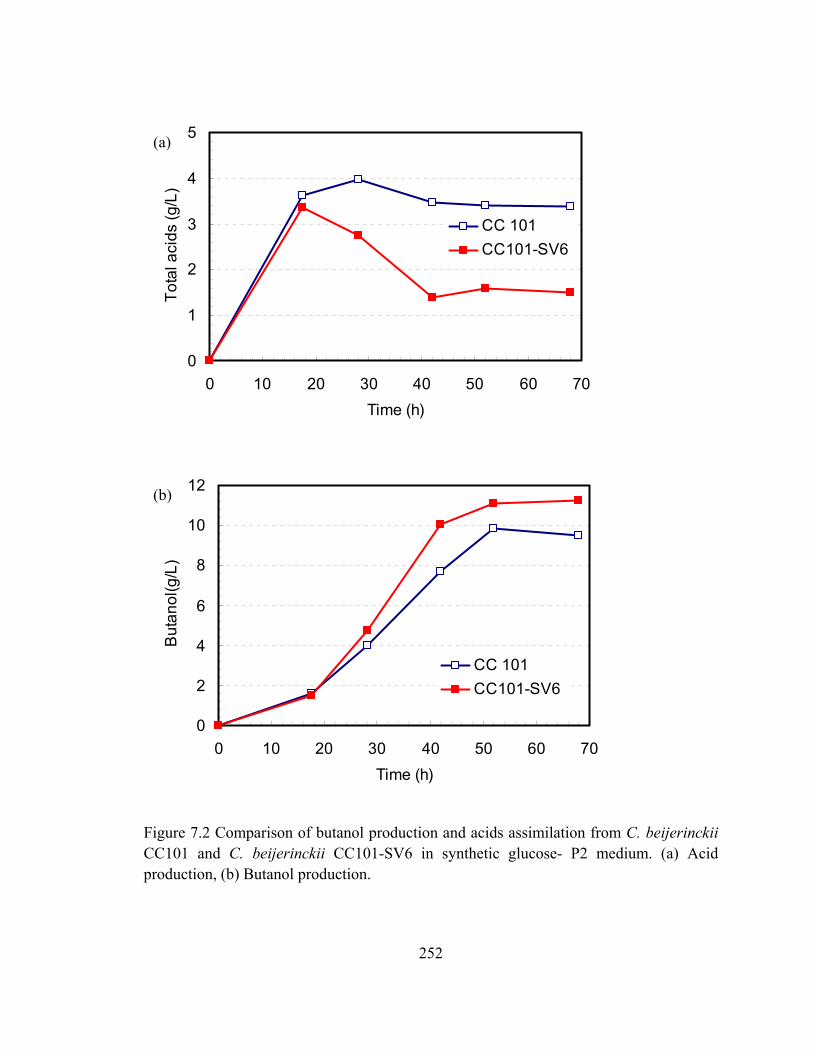
0
1
2
3
4
5
0 10 20 30 40 50 60 70
Time (h)
To
tal a
cid
s (g
/L)
CC 101
CC101-SV6
(a)
0
2
4
6
8
10
12
0 10 20 30 40 50 60 70
Time (h)
Bu
tan
ol(
g/L
)
CC 101
CC101-SV6
(b)
Figure 7.2 Comparison of butanol production and acids assimilation from C. beijerinckii CC101 and C. beijerinckii CC101-SV6 in synthetic glucose- P2 medium. (a) Acid production, (b) Butanol production.
252

0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60 70
Time (h)
Glu
cose
(g
/L)
0
1
2
3
4
5
6
7
So
lve
nts
an
d a
cid
s (g
/L)
Glucose
Acetone
Ethanol
Butanol
Acetic Acid
Butyric Acid
(a)
0
5
10
15
20
25
30
35
40
0 10 20 30 40 50 60 70
Time (h)
Glu
cose
(g
/L)
0
1
2
3
4
5
6
7
8
9
10
So
lve
nts
an
d a
cid
s (g
/L)
Glucose
Acetone
Ethanol
Butanol
Acetic Acid
Butyric Acid
(b)
Figure 7.3 Fermentation kinetics of C. beijerinckii CC101 and C. beijerinckii CC101-SV6 in synthetic glucose-P2 medium with cysteine. (a) ABE production of C. beijerinckii CC101, (b) ABE production of C. beijerinckii CC101-SV6.
253

0
1
2
3
4
5
0 10 20 30 40 50 60 70
Time (h)
To
tal a
cid
s (g
/L)
CC 101
CC101-SV6
(a)
0
1
2
3
4
5
6
7
8
9
10
0 10 20 30 40 50 60 70
Time (h)
Bu
tan
ol(
g/L
)
CC 101
CC101-SV6
(b)
Figure 7.4 Comparison of butanol production and acids assimilation from C. beijerinckii CC101 and C. beijerinckii CC101-SV6 in glucose-P2 medium with cysteine. (a) Acid production, (b) Butanol production.
254
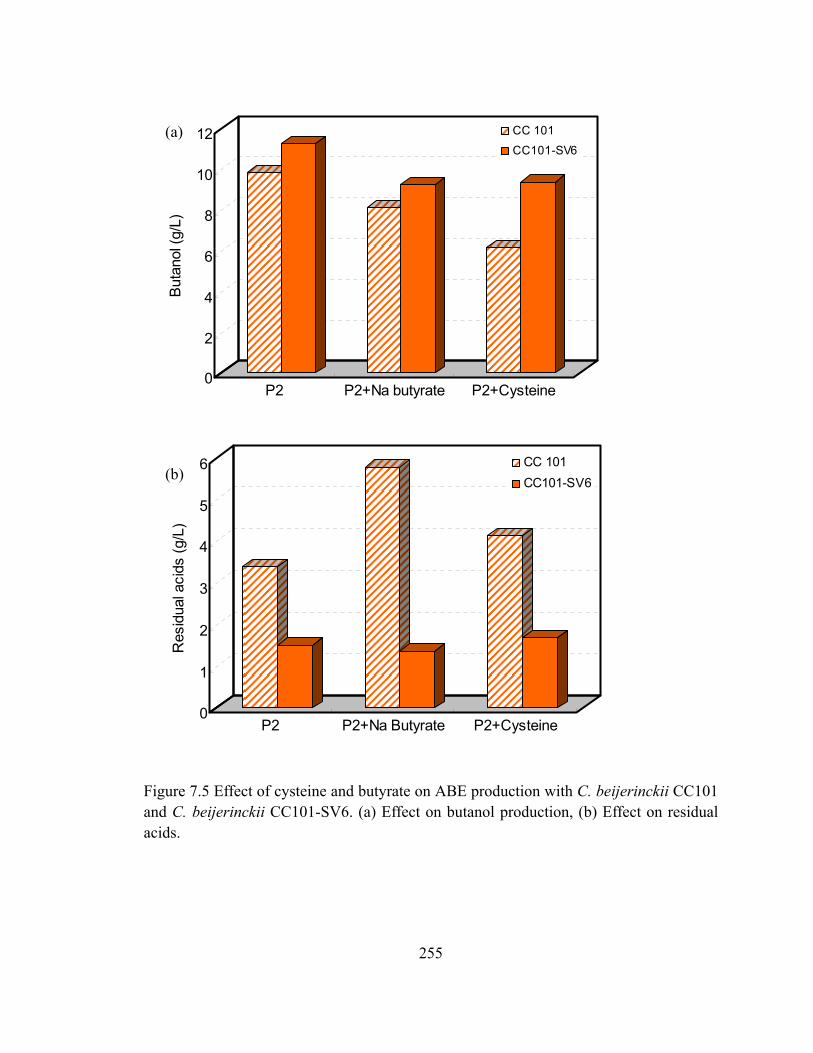
0
2
4
6
8
10
12
But
anol
(g/
L)
P2 P2+Na butyrate P2+Cysteine
CC 101
CC101-SV6
(a)
0
1
2
3
4
5
6
Res
idua
l aci
ds (
g/L)
P2 P2+Na Butyrate P2+Cysteine
CC 101
CC101-SV6
(b)
Figure 7.5 Effect of cysteine and butyrate on ABE production with C. beijerinckii CC101 and C. beijerinckii CC101-SV6. (a) Effect on butanol production, (b) Effect on residual acids.
255

0
2
4
6
8
10
12
14
16
18
20
0 10 20 30 40 50 60
Time (h)
Sug
ars
(g
/L)
0
1
2
3
4
5
6
7
So
lve
nts
an
d a
cid
s (g
/L)
Glucose
Xylose
Acetone
Ethanol
Butanol
Acetic Acid
Butyric Acid
(a)
0
2
4
6
8
10
12
14
16
18
20
0 10 20 30 40 50 60 70 80
Time (h)
Su
ga
rs (
g/L
)
0
1
2
3
4
5
6
7
8
9
So
lve
nts
an
d a
cid
s (g
/L)
Glucose
Xylose
Acetone
Ethanol
Butanol
Acetic Acid
Butyric Acid
(b)
Figure 7.6 ABE production from sugarcane bagasse hydrolysate. (a) Fermentation kinetics of C. beijerinckii CC101, (b) Fermentation kinetics of C. beijerinckii CC101-SV6.
256

257
Chapter 8: Conclusions and Recommendations
8.1 Conclusions
This study investigated butanol production via ABE fermentation from renewable
and sustainable lignocellulosic feedstocks using evolved and engineered mutant strains,
and demonstrated the advantages of alternative separation technique for online butanol
recovery and enhanced overall butanol production in the integrated fermentation process.
As shown in this project, butanol can be produced by ABE fermentation from corn fiber,
cassava bagasse, wood pulp and sugarcane bagasse. Gas stripping is an effective
separation technique for butanol recovery, and enhanced overall butanol production can
be achieved by integrated online butanol recovery with the fermentation process. The
important findings and conclusions of this project are discussed and summarized as
follows.
8.1.1 Butanol production by engineered mutant strains
Two mutant strains, C. beijerinckii JB 200 and C. beijerinckii CC101-SV6, were
investigated and employed in this study as solventogenic bacteria for butanol production.
C. beijerinckii JB 200 was obtained using evolution engineering after adaptation under
increased butanol stress in a fibrous bed bioreactor over an extended period of time,

258
whereas C. beijerinckii CC101-SV6 was obtained by overexpressing the solventogenic
genes. Compared with 22.4 g/L total ABE and 13.5 g/L butanol production by the
parental strain C. beijerinckii ATCC 55025, JB 200 is a hyper-butanol-producing mutant
with 36.4 g/L total ABE and 22.2 g/L butanol production. This result indicates that FBB
is a powerful system for cell mutation and evolution towards enhanced butanol
production. By overexpressing the solventogenic genes of adhE2, ald, ctfA and ctfB,
mutant C. beijerinckii CC101-SV6 exhibits stable butanol production under various
conditions, and demonstrates better acids assimilation and conversion into solvents than
its parental strain C. beijerinckii CC101. By employing these two mutants as the
solventogenic bacteria, stable and enhanced butanol production can be achieved even
under stressful environments, as compared to the parental strains.
8.1.2 Butanol production from lignocellulosic feedstocks
As demonstrated in this project, butanol can be produced by solventogenic Clostridia
beijerinckii in ABE fermentation using corn fiber, cassava bagasse, wood pulp, and
sugarcane bagasse. All the lignocellulosic biomass must be pretreated and hydrolyzed
first in order to release the fermentable sugars that can be utilized by the bacteria in the
fermentation. Acid hydrolysis generates toxic compounds inhibitory to the bacteria in the
subsequent fermentation process due to the degradation of sugars and lignin under severe
conditions, whereas enzymatic hydrolysis is mild and doesn’t pose any inhibition on the

259
fermentation. Detoxification and/or dilution are needed on the acid hydrolysates of
lignocellulosic biomass in order to remove or reduce the fermentation inhibitors present
and to obtain decent butanol production. In the batch ABE fermentation process, 8.8 g/L,
11.35 g/L and 9.44 g/L ABE were obtained from activated carbon detoxified corn fiber
hydrolysate, resin and evaporation detoxified wood pulp hydrolysate, and diluted
sugarcane bagasse hydrolysate, respectively. Inhibition caused by the lignocellulosic
hydrolysate was observed in all the above-mentioned studies. 15.41 g/L ABE were
obtained from enzymatic hydrolysate of cassava bagasse without observing any inhibition.
Butanol production from lignocellulosic hydrolysate is a challenging research topic, and
selecting robust strains and effective detoxification on the lignocellulosic hydrolysate are
two key factors in achieving desired butanol production.
8.1.3 Gas stripping as an alternative butanol recovery technique
Gas stripping is an efficient butanol recovery technique that can be integrated with
fermentation process for online butanol removal. It is effective in concentrating butanol
in the condensate stream, and the concentrated butanol solution eases the downstream
separation process. Lowering the condensate temperature and increasing the gas flow rate
both lead to faster butanol removal rate, but result in lower butanol concentration in the
condensate due to excess removal of water as well. Gas stripping does not remove any
cells or nutrients present in the fermentation broth, and the presence of cells do not have

260
any significant effect on butanol removal rate. Butanol selectivity of 7.4 – 21.4 was
obtained in this study, with an average in between 11.0 – 15.0. Acetone and ethanol
selectivity was found to be in the range of 3.0 – 4.0, indicating that gas stripping is highly
selective towards butanol removal.
8.1.4 Enhanced butanol production in the integrated fermentation process with online
product recovery
Enhanced overall butanol production can be achieved in the integrated fermentation
process with online product recovery by gas stripping, as demonstrated in this study. Gas
stripping can efficiently recover butanol from fermentation broth and relieve the
inhibition caused by butanol. Concentrated substrate feeding is possible in the fed-batch
ABE fermentation when coupled with online butanol removal, reducing the reactor
volume and increasing the volumetric productivity. More efficient sugar conversion can
be achieved in the integrated process than the non-integrated process due to relieved
inhibition and stress on the bacteria, leading to higher overall solvent production. When
operating the fed-batch fermentation process over a long extended period, nutrients
supplementation can rejuvenate the bacteria and maintain a high sugar conversion rate
and solvent productivity. In this study, highly concentrated cassava bagasse hydrolysate
containing 584.4 g/L glucose was utilized and 90.31 g/L ABE were obtained in the
integrated fed-batch fermentation with gas stripping as product recovery, compared to

261
15.4 g/L ABE produced in the batch process. When providing additional nutrients to the
bacteria in the fermentation, 108.5 g/L ABE was produced from the integrated process.
Compared with 44.8 g/L glucose consumed in the control batch fermentation, 244.6 g/L
and 336.9 g/L glucose were utilized in the integrated fed-batch fermentation without and
with nutrient supplementation, respectively. 17.73 g/L ABE were achieved in the gas
stripping integrated batch process using resin detoxified wood pulp hydrolysate,
comparing to 11.35 g/L ABE in the control batch process. The sugar conversion was
improved from 65.6 % to 74.6 % due to the relief of end product butanol inhibition.
8.2 Recommendations
Although an integrated process for butanol production from lignocellulosic biomass
has been developed and demonstrated in this project, many areas still require continuing
research endeavors for improvement and perfection before this process can be
industrialized on a commercial scale and compete with petrochemically-derived butanol.
Some suggestions and recommendations for future research work are listed below.
8.2.1 Improvement on the fermentability of lignocellulosic hydrolysates
The biggest challenge in butanol production by Clostridium beijerinckii from
lignocellulosic biomass encountered is the inhibition on the cells caused by the
hydrolysates. Dilute acid hydrolysis is the most commonly used pretreatment method on

262
lignocellulosic biomass, but inevitably results in degradation products from sugars and
lignin. The degradation products in the hydrolysate are severe fermentation inhibitors.
Enzymatic hydrolysis does not generate fermentation inhibitors, but the process
efficiency is limited by the accessibility and digestibility of hemicellulose and cellulose
due to the rigid structure of lignocellulose. Other pretreatment methods such as steam
explosion or liquid hot water pretreatment can be explored in the future to improve the
digestibility of the lignocellulosic biomass, which can further improve the efficiency of
enzymatic hydrolysis.
In this study, detoxification methods, including overliming and adsorption by
activated carbon and resin, have been examined and adsorption by resin is identified as a
very effective detoxification method. However, detoxification by resin is very expensive
and resin needs to be regenerated. Other detoxification methods such as biological
detoxification and liquid-liquid extraction of the inhibitory compounds can be studied in
the future in the search for an economic and effective procedure to improve the
fermentability of the lignocellulosic hydrolysates.
8.2.2 Optimization of butanol recovery by gas stripping and investigation on alternative
recovery techniques
In this study, gas flow rate and condensate temperature were investigated as two
operating parameters in the gas stripping study. Condensation process is the most

263
energy-intensive step in gas stripping, and condensation temperature in the range of 0 – 5
oC can be evaluated in the future to find an optimal temperature that can efficiently
recover more butanol and less water. In addition, higher condensate temperature is in
favor of saving energy. A relationship between required gas flow rate and reactor volume
should be established in the future to suit the need of different processes and provide
information on the process scale-up. Understanding the relationship between gas flow
rate and butanol recovery rate is also very important in designing the integrated process,
where the butanol production rate and butanol removal rate should reach a desired
equilibrium by carefully controlling the gas stripping process.
Besides gas stripping, there exist many alternative butanol recovery techniques, such
as liquid-liquid extraction and pervaporation. Pervaporation has been widely reported as
an efficient butanol recovery technique that can be integrated with ABE fermentation for
online butanol removal. Pervaporation process can be investigated in the future study as
another feasible alternative butanol recovery technique.
8.2.3 Process development on ABE fermentation
Besides batch and fed-batch fermentations investigated in this study, continuous
fermentation using lignocellulosic biomass can be studied in the future. Continuous
fermentation in the cell-immobilized fibrous bed bioreactor may offer many advantages,
including high reactor productivity and reduced inhibition due to a constant flow of fresh

264
medium. In addition to the feedstocks that have been investigated, other lignocellulosic
biomass can be evaluated in the future for butanol production to broaden the substrate
pool and gain more information on the performance of each type of feedstock, such as
sorghum, corn stover, wheat straw and switchgrass. Evaluating different feedstocks helps
to understand the choices of substrates for butanol production in different regions, and
promotes value-added by products for the processing industry.
More research attention should also be paid on medium formula for ABE
fermentation. Soybean meal, cotton seed protein, corn steep liquor and molasses were
preliminarily investigated in this project as potential nitrogen sources to replace the
expensive yeast extract currently used in the medium formula. The effect of cysteine,
butyric acid and ammonia acetate on butanol production was also investigated. In the
future, an optimized medium formula using these alternative nitrogen sources and
supplementation of additional chemicals can be developed in search for a cost-effective
medium formula for industrial process for economical butanol production.

265
Bibliography
Afschar, A.S., H. Biebl, K. Schaller, and K. Schugerl (1985). Production of acetone and butanol by Clostridium acetobutylicum in continuous culture with cell recycle. Appl. Microbiol. Biotechnol., 22, 394-398.
Ahn, J.H., B.I. Sang, and Y. Um (2011). Butanol production from thin stillage using Clostridium pasteurianum. Bioresour. Technol., 102, 4934-4937
Atsumi, S., A.F. Cann, M.R. Connor, C.R. Shen, K.M. Smith, M.P. Brynildsen, K.J. Chou, T.Hanai, J.C. Liao (2007). Metabolic engineering of Escherichia coli for 1-butnaol production. Metab. Eng., 10, 305-311.
Bahl, H., W. Andersch, K. Braun, and G. Gottschalk (1982). Effect of pH and butyrate concentration on the production of acetone and butanol by Clostridium acetobutylicum grown in continuous culture. European J. Appl. Microbiol. Biotechnol., 14, 17-20.
Badr, H.R., R. Toledo, and M.K. Hamdy (2001). Continuous acetone-ethanol-butanol fermentation by immobilized cells of Clostridium acetobutylicum. Biomass and Bioenergy, 20, 119-132.
Barton, W.E. and A.J. Daugulis (1992). Evaluation of solvents for extractive butanol fermentation with Clostridium acetobutylicum and the use of poly(propylene glycol) 1200. Appl. Microbiol. Biotechnol., 36, 632-639.
Beesch, S.C. (1952). Acetone-butanol fermentation of sugars. Ind. Eng. Chem., 44, 1677-1682.
Biebi, H (2001). Fermentation of glycerol by Clostridium pasteurianum – batch and continuous culture studies. J. Ind. Microbiol. Biotechnol., 27,18-26.
Boddeker, K.W., G. Bengtson and H. Pingel ( 1990). Pervaporation of isomeric butanols. J. Memb. Sci., 54, 1-12.

266
Bock, S.A., S.L. Fox, and W.R. Gibbons (1997). Development of a low-cost, industrially suitable medium for the production of acetic acid from Clostridium thermoaceticum. Biotechnol. Appl. Bioc., 25, 117-125.
Bramorski, A., C.R. Soccol, P. Christen, S. Revah (1998). Fruity aroma production by Ceratocystis fimbriata in solid cultures from agro-industrial wastes. Rev. Microbiol., 29, 208-212.
Bruant, G., M.J. Levesque, C. Peter, S.R. Guiot, L. Masson (2010). Genomic analysis of carbon monoxide utilization and butanol production by Clostridium carboxidivorans strain P7. PLoS ONE, 5 (9), article number e13033.
Cadoche, L. and G.D. Lopez (1989). Assessment of size reduction as a preliminary step in the production of ethanol from lignocellulosic wastes. Biol. Waste., 30, 153-157.
Cara, C., M. Moya, I. Ballesteros, M.J. Negro, A. Gonzalez and E. Ruiz (2007). Influence of solid loading on enzymatic hydrolysis of steam exploded or liquid hot water pretreated olive tree biomass. Process Biochemistry, 42, 1003-1009.
Chang, V.S., B. Burr and M.T. Holtzapple (1997). Lime pretreatment of switchgrass. Appl. Biochem. Biotechnol., 63, 3–19.
Chang, V.S., M. Nagwani and M.T. Holtzapple (1998). Lime pretreatment of crop residues bagasse and wheat straw. Appl. Biochem. Biotechnol., 74, 135–159.
Chang, V.S., M. Nagwani, C.H. Kim and M.T. Holtzapple (2001). Oxidative lime pretreatment of high-lignin biomass. Appl. Biochem. Biotechnol., 94, 1–28.
Chernova, N.I., T.P. Korobkova, and S.V. Kiseleva (2010). Use of biomass for producing liquid fuel: Current state and innovations. Thermal Eng., 57, 937-945.
Chen, C.K. and H.P. Blaschek (1999). Acetate enhances solvent production and prevents degeneration in Clostridium beijerinckii BA 101. Appl. Microbiol. Biotechnol., 52, 1701-73.
Cho, D.H., Y.H. Lee, Y. Um, B.I. Sang, Y.H. Kim (2009). Detoxification of model phenolic compounds in lignocellulosic hydrolysates with peroxidase for butanol production from Clostridium beijerinckii. Appl. Microbiol Biotechnol., 83, 1035-1043.

267
Converti, A., J.M. Dominguez, P. Perego, S.S. Silva and M. Zilli (2000). Wood hydrolysis and hydrolysate detoxification for subsequent xylitol production. Chem. Eng. Technol., 23, 1013-1020.
Cornillot, E., R. Nair, E.T. Papoutsakis, P. Soucaille (1997). The genes for butanol and acetone formation in Clostridium acetobutylicum ATCC 824 reside on a large plasmid whose loss leads to degeneration of the strain. J. Bacteriol., 179, 5442-5447.
Delgenes, J., R. Moletta and J.M. Navarro (1996). Effects of lignocellulose degradation products on ethanol fermentations of glucose and xylose by Saccharomyces cerevisiae, Zymomonas mobilis, Pichia stipitis and Candida shehatae. Enzyme Microb. Technol., 19, 220-225.
Demain, A.L. (2009). Biosolutions to the energy problem. J. Ind. Microbiol. Biotechnol., 36, 319-332.
Dien, B.S., R.B. Hespell, L.O. Ingram and R.J. Bothast (1997). Conversion of corn milling fibrous co-products into ethanol by recombinant Escherichia coli strains K011 and SL40. World J. Microbiol Biotechnol., 13, 619-625.
Dominguez, J.M., C.S. Gong, and G.T. Tsao (1996). Pretreatment of sugar cane bagasse hemicellulose hydrolysate for xylitol production by yeast. Appl. Biochem. Biotech., 57-8, 49-56.
Duff, S.J.B. and W.D. Murray (1996). Bioconversion of forest products industry waste cellulosics to fuel ethanol: a review. Bioresour. Technol., 55, 1-33.
Dürre, P. and H. Bahl (1996). Microbial production of acetone/butanol/isopropanol. In: Roehr M (ed) Biotechnoloy, 2nd edition, vol 6.VCH, Weinheim, pp 229-268.
Dürre, P. (1998). New insights and novel developments in clostridial acetone/ butanol/ isopropanol fermentation. Appl. Microbiol. Biotechnol., 49, 639-648.
Dürre, P. (2007). Biobutanol: An attractive biofuel. Biotechnol. J., 2, 1525-1534.
Earle, M.J. and K.R. Seddon (2000). Ionic liquids. Green solvents for the future. Pure Appl. Chem., 72, 1391-1398.
Eckert, G. and K. Schügerl (1987). Continuous acetone-butanol production with direct

268
product removal. Appl. Microbiol. Biotechnol., 27, 221-228.
Ennis, B.M., N.A. Gutierrez, I.S. Maddox (1986a). The actone-butanol-ethanol fermentation: a current assessment. Process Biochem., 21, 131-147.
Ennis, B., C.T. Marshall, I.S. Maddox, A.H.J. Paterson (1986b). Continuous product recovery by in-situ gas stripping/condensation during solvent production from whey permeate using Clostridium acetobutylicum. Biotechnol. Lett., 8, 725-730.
Ennis, B.M., N. Qureshi and I.S. Maddox (1987). In-line toxic product removal during solvent production by continuous fermentation using immobilized Clostridium acetobutylicum. Enzyme Microbial. Technol., 9, 672-675.
Evans, P.J. and H.Y. Wang (1988). Response of Clostridum acetobutylicum to the presence of mixed extractants. Appl. Environ. Microbiol., 54, 175-192.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2003). Production of butanol by Clostridium beijerinckii BA101 and in-situ recovery by gas stripping. J. Microbiol. Biotechnol., 19, 595-603.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004a). Butanol fermentation research: upstream and downstream manipulations. The Chemical Record, 4, 305-314.
Ezeji, T.C., N. Qureshi, and H.P. Blaschek (2004b). Acetone-butanol-ethanol production from concentrated substrate: reduction in substrate inhibition by fed-batch technique and product inhibition by gas stripping. Appl. Microbiol. Biotechnol., 63, 653-658.
Ezeji, T.C., P.M. Karcher, N. Qureshi, H.P. Blaschek (2005a). Improving performance of a gas stripping-based recovery system to remove butanol from Clostridium beijerinckii fermentation. Bioprocess Biosyst. Eng., 27, 207-214.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2005b). Process for continuous solvent production. United States Patent Application Publication, US patent 20050089979A1.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007a). Bioproduction of butanol from biomass: from genes to bioreactors. Current Opinion in Biotechnol., 18, 220-227.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007b). Butanol production from agricultural residues: impact of degradation products on Clostridium beijerinckii growth and

269
butanol fermentation. Biotechnol. Bioeng., 97, 1460-1469.
Ezeji, T.C., N. Qureshi and H.P. Blaschek (2007c). Production of acetone butanol (AB) from liquefied corn starch, a commercial substrate, using Clostridium beijerinckii coupled with product recovery by gas stripping. J Ind. Microbiol. Biotechnol., 34, 771-777.
Ezeji, T.C. and H.P. Blaschek (2008). Fermentation of dried distillers’ grains and solubles (DDGS) hydrolysates to solvents and value-added products by solventogenic clostridia. Bioresour. Technol., 99, 5232-5242.
Ezeji, T.C., C. Milne, N.D. Price, H.P. Blaschek (2010). Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol., 85, 1697-1712.
Fadeev, A.G., M.M. Meagher, S.S. Kelley, V.V. Volkov (2000). Fouling of poly-[1-(trimethylsilyl)-1-propyne] membranes in pervaporative recovery of butanol from aqueous solutions and ABE fermentation broth. J. Membr. Sci., 173, 133-144.
Fadeev, A.G. and M.M. Meagher (2001). Opportunities for ionic liquids in recovery of biofuels. Chem. Commun., 295-296.
Fadeev, A.G., YaA Selinskaya, S.S. Kelley, M.M. Meagher, E.G. Litvinova, V.S. Khotimsky, V.V. Volkov (2001). Extraction of butanol from aqueous solutions by pervaporation through poly(1-trimethylsilyl-1-propyne). J. Membr. Sci., 186,205-217.
Fadeev, A.G., S.S. Kelley, J.D. McMillan, YaA Selinskaya, V.S. Khotimsky, V.V. Volkov (2003). Effect of yeast fermentation by-products on poly(1-trimethylsilyl-1-propyne) pervaporative performance. J. Membr. Sci., 214,229-238.
Felipe, M.G.A., M.V. Veira, M. Vitolo, I.M. Mancilha, I.C. Roberto and S.S. Silva (1995). Effect of acetic acid on xylose fermentation to xylitol by Candida guilliermondii. J. Basic Microb., 35, 171-177.
Festel, G.W. (2008). Review : Biofuels – Economic aspects. Chem. Eng. Technol., 31, 715-720.
Fond, O., G. Matta Ammouri, H. Petitdemange, and J.M. Engasser (1985). The role of acids on the production of acetone and butanol by Clostridium acetobutylicum. Appl.

270
Microbiol. Biotechnol., 22, 195-200.
Formanek, J., R. Mackie, H.P. Blaschek (1997). Enhanced butanol production by Clostridium beijerinckii BA101 grown in semidefined P2 medium containing 6 percent maltodextrin or glucose. Appl. Environ. Microbiol., 63, 2306-2310.
Frick, C. and K. Schugerl (1986). Continuous acetone-butanol production with free and immobilized Clostridium acetobutylicum. Applied Microbiol. Biotechonol., 25, 186-193.
Gabriel, C.L. (1928). Butanol fermentation process. Ind. Eng. Chem., 20, 1063-1067.
Gabriel, C.L. and F.M. Crawford (1930). Development of the butyl-acetonic fermentation industry. Ind. Eng. Chem., 22, 1163-1165.
Gapes, J.R., D. Nimcevic, and A. Friedl (1996). Long-term continuous cultivation of Clostridium beijerinckii in a two-stage chemostat with on-line solvent removal. Appl. Environ. Microbiol., 62, 3210-3219.
Gapes, J.R. (2000). The economics of acetone-butanol fermentation: Theoretical and market considerations. J. Mol. Microbiol. Biotech., 2, 27-32.
Garćia, V., J. Päkkilä, H. Ojamo, E. Muurinen, R.L. Keiski (2011). Challenges in biobutanol production: How to improve the efficiency. Renew. Sustain. Ener. Reviews, 15, 964-980.
Geng, Q and C-H Park (1994). Pervaporative butanol fermentation by Clostridium acetobutylicum B18. Biotech. Bioeng., 43, 978-986.
Geng, Q.H., C.H. Park and K. Janni (1995). Uptake of organic-acids by Clostridium acetobutylicum B18 under controlled pH and reduced butanol inhibition. Korean J. Chem. Eng., 12, 378-383.
George, H.A., J.L. Johnson, W.E.C. Moore, L.V. Holdeman, J.S. Chen (1983). Acetone, isopropanol, and butanol production by Clostridium beijerinckii (syn. Clostridium butylicum) and Clostridium aurantibutyricum. Appl. Environ Microbiol., 45, 1160-1163.
Girbal, L. and P. Soucaille (1998). Regulation of solvent production in Clostridium acetobutylicum. Trends Biotechnol., 16, 11-16.

271
Gottschalk, J.C. and J.G. Morris (1981). The induction of acetone and butanol production in cultures Clostridium acetobutylicum by elevated concentrations of acetate and butyrate. FEMS Microb. Letters, 12, 385-389.
Groot, W.J. , C.E. Van der Oever and N.W.F. Kossen (1984). Pervaporation for simultaneous product recovery in the butanol/isopropanol batch fermentation. Biotechnol. Lett., 6, 709-714.
Groot, W.J. and K. Ch. A. M. Luyben (1986). In situ product recovery by adsorption in the butanol/isopropanol batch fermentation. Appl. Microbiol. Biotechnol., 25, 29-31.
Groot, W.J., R.G.J.M. van der Lans, K.Ch.A.M. Luyben (1989). Batch and continuous butanol fermentations with free cells: integration with product recovery by gas- stripping. Appl. Microbiol. Biotechnol., 32, 305-308.
Groot, W.J., H.S. Soedjak, P.B. Donck, R.G.J.M. van der Lans, K. Ch. A.M. Luyben, J.M.K. Timmer (1990). Butanol recovery from fermentations by liquid-liquid extraction and membrane solvent extraction. Bioprocess Eng., 5, 203-216.
Groot, W.J., R.G.J.M. van der Lans, and K. Ch. A.M. Luyben (1992). Technologies for butanol recovery integrated with fermentations. Proc. Biochem., 27, 61-65.
Gu, Y., S.Y. Hu, J. Chen, L.J. Shao, H.Q. He, Y.L. Yang, S. Yang, W.H. Jiang (2009). Ammonium acetate enhances solvent production by Clostridium acetobutylicum EA 2018 using cassava as a fermentation medium. J. Ind. Microbiol. Biotechnol., 36, 1225-1232.
Gutierrez, N.A., I.S. Maddox, K.C. Schuster, H. Swoboda, J.R. Gapes (1998). Strain comparison and medium preparation for the acetone-butanol-ethanol (ABE) ferment process using substrate potato. Bioresour. Technol., 66, 263-265.
Hagiwara, R. and Y. Ito (2000). Room temperature ionic liquids of alkylimidazolium cations and fluoroanions. J. Fluor. Chem., 105, 221-227.
Hahn-hagerdal, B., M. Galbe, M.F. Gorwa-Grauslund, G. Liden and G. Zacchi (2006). Bio-ethanol—the fuel of tomorrow from the residues of today. TRENDS in Biotechnol., 12, 549-556.
Harris, L.M., R.P. Desai, N.E. Welker, E.T. Papoutsakis (2000). Characterization of recombinant strains of the Clostridium acetobutylicum butyrate kinase inactivation

272
mutant: need for new phenomenological models for solventogensis and butanol inhibition? Biotechnol. Bioeng., 67, 1-11.
Harris, L.M., N.E. Welker, E.T. Papousakis (2002). Northern, morphological, and fermentation analysis of spo0A inactivation and over-expression in Clostridium acetobutylicum ATCC824. J. Bacteriol., 184, 3586-3597.
Hendriks A.T.W.M. and G. Zeeman (2009). Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresour. Technol., 100, 10-18.
Hickey, P.J., F.P. Juricic and C.S. Slater (1992). The effect of process parameters on the pervaporation of alcohols through organophilic membranes. Separation Sci. Technol., 27, 843-861.
Hipolito, C.N., E. Crabbe, C.M. Badillo, O.C. Zarrabal, M.A. M. Mora, G.P. Flores, M.A.H. Cortazar, A. Ishizaki (2008). Bioconversion of industrial wastewater from palm oil processing to butanol by Clostridium saccharoperbutylacetonicum N1-4 (ATCC 13564). J. Cleaner Prod., 16, 632-638.
Holtzapple, M.T. and R.F. Brown (1994). Conceptual design for a process to recover volatile solutes from aqueous solutions using silicalite. Sep. Technol., 4, 213-229.
Howard, R.L., E. Abotsi, E.L. Jansen van Rensburg and S. Howard (2003). Lignocellulosic biotechnology: issues of bioconversion and enzyme production. African J. Biotechnol., 2(12), 602-619.
Huang J. and M.M. Meagher (2001). Pervaporative recovery of n-butanol from aqueous solutions and ABE fermentation broth using thin-film silicalite-filled silicone composite membranes. J. Membr. Sci., 192, 231-242.
Huang, Y.L., Z. Wu, L. Zhang, C.M. Cheung and S.T. Yang (2002). Production of carboxylic acids from hydrolyzed corn meal by immobilized cell fermentation in a fibrous-bed bioreactor. Bioresource Technol., 82, 51-59.
Huang, W.C., D.E. Ramey, and S.T. Yang (2004). Continuous production of butanol by Clostridium acetobutylicum immobilized in a fibrous bed bioreactor. Appl. Biochem. Biotechnol., 113-116, 887-898.
Huang, H., H. Liu, Y.R. Gan (2010). Genetic modification of critical enzymes and involved genes in butanol biosynthesis from biomass. Biotech. Adv., 28, 651-657.

273
Huddleston, J.G., H.D. Willauer, R.P. Swatloski, A.E. Visser, R.D. Rogers (1998). Room temperature ionic liquids as novel media for “clean” liquid-liquid extraction. Chem Commun., 1765-1766.
Inui, M., M. Suda, S. Kimura, K. Yasuda, H. Suzuki, H. Toda, S. Yamamoto, S. Okino, N. Suzuki, H. Yukawa (2008). Expression of Clostridium acetobutylicum butanol synthetic genes in Escherichia coli. Appl. Microbiol. Biotechnol., 77, 1305-1316.
Izak, P., K. Schwarz, W. Ruth, H. Bahl, U. Kragl (2008). Increased productivity of Clostridium acetobutylicum fermentation of acetone, butanol, and ethanol by pervaporation through supported ionic liquid membrane. Appl. Microbiol. Biotechnol., 78, 597-602.
Jain, M.K., D. Beacom, and R. Datta (1993). Mutant strain of C. acetobutylicum and process for making butanol. United States Patent, US Patent 5192673.
Jesse, T.W., T.C. Ezeji, N. Qureshi, H.P. Blaschek (2002). Production of butanol from starch-based waste packing peanuts and agricultural waste. J. Ind. Microbiol. Biotechnol., 29, 117-123.
Jiang, Y., C. Xu, F .Dong, Y. Yang, W. Jiang, S. Yang (2009). Disruption of the acetoacetate decarboxylase gene in solvent-producing Clostridium acetobutylicum increases the butanol ratio. Metab. Eng., 11, 284-291.
Jones, D.T. and D. Woods (1986). Acetone-butanol fermentation revisted. Microbiol. Reviews, 50, 484-524.
Jonquieres, A. and A. Fane (1997). Filled and unfilled composite GFT PDMS membranes for the recovery of butanol from dilute aqueous solutions: influence of alcohol polarity. J. Membr. Sci., 125, 245-255.
Jonquieres, A., R. Clement, P. Lochon, J. Neel, M. Dresch, B. Chertien (2002). Industrial state-of-art of pervaporation and vapour permeation in the western countries. J. Membr. Sci., 206, 87-117.
Jorgensen, H., J. B. Kristensen and C. Felby (2007). Enzymatic conversion of lignocellulose into fermentable sugars: Challenges and opportunities. Biofuels. Bioprod. Bioref., 1, 119–134.
Kadam, K.L. and M.M. Newman (1997). Development of a low-cost fermentation

274
medium for ethanol production from biomass. Appl. Microbiol. Biotechnol., 47, 625-629.
Kim S.Y., D.K. Oh and J.H. Kim (1999). Evaluation of xylitol production from corn cob hemicellulose hydrolysate by Candida parapsilosis. Biotechonol. Lett., 21, 891-895.
Klinke, H.B., A.B. Thomsen and B.K. Ahring (2004). Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pre-treatment of biomass. Appl. Microbiol. Technol., 66, 10-26.
Knoshaug, E.P. and M. Zhang (2009). Butanol tolerance in a selection of microorganisms. Appl. Biochem. Biotechnol., 153, 13-20.
Kumar, P., D.M. Barrett, M.J. Delwiche and P. Stroeve (2009). Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem., 48, 3713-3729.
Kumar, M. and K. Gayen (2011). Developments in biobutanol production: New insights. Appl. Ener., 88, 1999-2012.
Larsson, S., A. Reimann, N. Nilvebrant, J.J. Jönsson (1999). Comparison of different methods for the detoxification of lignocellulose hydrolysates of spruce. Appl. Biochem. Biotechnol., 77-79, 91-103.
Larrayzo M.A. and L. Puigjaner (1987). Study of butanol extraction through pervaporation in acetobutylic fermentation. Biotechnol. Bioeng., 30, 692-696.
Lee, S.T., J.H. Park, S.H. Jang, L.K. Nielsen, J. Kim, K.S. Jung (2008). Fermentive butanol production by Clostridia. Biotechnol. Bioeng., 101,209-228.
Li, S., V.A. Tuan, J.L. Falconer, R.D. Noble (2003). Properties and separation performance of Ge-ZSM-5 membranes. Micropor. Mesopor. Mater., 58, 137-154.
Li, S-Y, R. Srivastava and R.S. Parnas (2010). Separation of 1-butanol by pervaporation using novel tri-layer PDMS composite membrane. J. Membr. Sci., 363, 287-294.
Lin, Y.L. and H.P. Blaschek (1983). Butanol production by a butanol-tolerant strain of Clostridium acetobutylicum in extruded corn broth. Appl. Environ. Microbiol., 45, 966-973.

275
Liou, J.S.C., D.L. Balkwill, G.R. Drake, R.S. Tanner (2005). Clostridium carboxidivorans sp. nov., a solvent-producing Clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen Clostridium scatologenes strain SL1 as Clostridium drakei sp. nov. Int. J. Syst. Evol. Microbiol., 55, 2085-2091.
Liu, Q., M.H.A. Janssen, F. van Rantwijk, R.A. Sheldon (2005). Room temperature ionic liquids that dissolve carbohydrates in high concentrations. Green Chemistry, 7, 39-42.
Liu, Z., Y.Ying, F. Li, C. Ma, P. Xu (2010). Butanol production by Clostridium beijerinckii ATCC 55025 from wheat bran. J. Ind. Microbiol. Biotechnol., 37, 495-501.
Lu, X.B., Y.M. Zhang, J. Yang and Y. Liang (2007). Enzymatic hydrolysis of corn stover after pretreatment with dilute sulfuric acid. Chem. Eng. Technol., 30, 938-944.
MacDonald, D. G.; N.N. Bakhshi, J.F. Mathews, A. Roychowdhury, P. Bajpai and M. Moo-Young (1983). Alkali treatment of corn stover to improve sugar production by enzymatic hydrolysis. Biotechnol. Bioeng., 25, 2067–2076.
Maddox, I.S. (1982). Use of silicalite for the adsorption of n-butanol from fermentation liquors. Biotechnol. Lett., 4, 759-760.
Maddox, I.S. (1989). The actone-butanol-ethanol fermentation: recent progress in technology. Genet. Eng. Rev., 7, 189-220.
Maddox, I.S., N. Qureshi and K. Roberts-Thomson (1995). Production of acetone-butanol-ethanol from concentrated substrates using Clostridium acetobutylicum in an integrated fermentation-product removal process. Process Biochemistry, 30, 209-215.
Madihah, M.S., A.B. Ariff, K.M. Sahaid, A.A. Suraini, M.I.A. Karim (2001). Direct fermentation of gelatinized sago starch to acetone-butanol-ethanol by Clostridium acetobutylicum. World J. Microbiol. Biotechnol., 17, 567-576.
Malherbe, S. and T.E. Cloete (2003). Lignocellulose biodegradation: fundamentals and applications: A review. Environ. Sci. Biotechnol., 1, 105-114.
Martinez, A., M.E. Rodriguez, M.L. Wells, S.W. York, J.F. Preston and L.O. Ingram (2001). Detoxification of dilute acid hydrolysates of lignocellulose with lime.

276
Biotechnol. Prog., 17, 287-293.
Matsumoto, M., K. Mochiduki, K. Fukunishi, K. Kondo (2004). Extraction of organic acids using imidazolium-based ionic liquids and their toxicity to Lactobacillus rhamnosus. Separ. Purif. Technol., 40, 97-101.
Matsumura, M. and H. Kataoka (1987). Separation of dilute aqueous butanol and acetone solutions by pervaporation through liquid membranes. Biotechnol. Bioeng., 30, 1991-1992.
McCoy, E., E.B. Fred, W.H. Peterson, E.G. Hastings (1926). A cultural study of the acetone butyl alcohol organisms. J. Infect. Dis., 39, 457-483.
Meagher, M.M., N. Qureshi and R.W. Hutkins (1998). Silicalite membrane and method for the selective recovery and concentration of acetone and butanol from model ABE solutions and fermentation broth. U.S. Patent 5,755,967.
Mermelstein, L.D., E.T. Papoutsakis, D.J. Petersen, G.N. Bennett (1993). Metabolic engineering of Clostridium acetobutylicum ATCC 824 for increase solvent production by enhancement of acetone formation enzyme-activities using a synthetic acetone operon. Biotechnol. Bioeng., 42, 1053-1060.
Meyer, C.L., J.W. Roos and E.T. Papoutsakis (1986). Carbon monoxide gassing leads to alcohol production and butyrate uptake without acetone formation in continuous cultures of Clostridium acetobutylicum. Appl. Microbiol. Biotechnol., 24, 159-167.
Mes-Hartree, M., B. E. Dale and W.K. Craig (1988). Comparison of steam and ammonia pretreatment for enzymatic hydrolysis of cellulose. Appl. Microbiol. Biotechnol., 29, 462–468.
Milestone, N.B. and D.M. Bibby (1981). Concentration of alcohols by adsorption on silicalite. J. Chem. Technol. Biotechnol., 31, 732-736.
Mitchell, W.J. (1996). Carbohydrate uptake and utilization by Clostridium beijerinckii NCIMB 8052. Anaerobe, 2, 379-384.
Mosier, N.S., C. Wyman, B.Dale, R.Elander, Y.Y. Lee, M. Holtzapple and M.R. Ladisch (2005). Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96, 673-668.

277
Mussatto, S.I. and I.C. Roberto (2001). Hydrolysate detoxification with activated charcoal for xylitol production by Candida guilliermondii. Biotechnol. Lett., 23, 1681-1684.
Mussatto, S.I. and I.C. Roberto (2004a). Alternatives for detoxification of diluted-acid lignocellulosic hydrolysates for use in fermentative processes: a review. Bioresour. Technol., 93, 1-10.
Mussatto, S.I. and I.C. Roberto (2004b). Optimal experimental condition for hemicellulosic hydrolysate treatment with activated charcoal for xylitol production. Biotechnol. Prog., 20, 134-139.
Mussatto, S.I., J.C. Santos and I.C. Roberto (2004). Effect of pH and activated charcoal adsorption on hemicellulosic hydrolysate detoxification for xylitol production. J. Chem. Technol. Biotechnol., 79, 590-596.
Naik, S.N., V.V. Goud, P.K. Rout, A.K. Dalai (2010). Production of first and second generation biofuels: a comprehensive review. Renew. Sust. Energy Rev., 14, 578-597.
Nair, R.V. and E.T. Papoutsakis (1994). Expression of plasmid-encoded aad in Clostridium acetobutylicum M5 restores vigorous butanol production. J. Bacteriol., 176, 5843-5846.
Nair, R.V., E.M. Green, D.E. Watson, G.N. Bennett, E.T. Papouskis (1999). Regulation of the sol locus genes for butanol and acetone formation in Clostridium acetobutylicum ATCC824 by a putative transcriptional regressor. J. Bacteriol., 181, 319-330.
Napoli, F., G. Olivieri, M.E. Russo, A. Marzocchella, P. Salatino (2010). Butanol production by Clostridium acetobutylicum in a continuous packed bed reactor. J. Ind. Microbiol. Biotechnol., 37, 603-608.
Ni, Y. and Z. Sun (2009). Recent progress on industrial fermentative production of acetone-butanol-ethanol by Clostridium acetobutylicum in China. Appl. Microbiol. Biotechnol., 83, 415-423.
Nielsen, L., M. Larsson, O. Holst, B. Mattiasson (1988). Adsorbents for extractive bioconversion applied to the acetone-butanol fermentation. Appl. Microbiol. Biotechnol., 28, 335-339.
Nielsen, D.R. and K.J. Prather (2009). In situ product recovery of n-butanol using

278
polymeric resins. Biotechnol. Bioeng., 102,811-821.
Nielsen, D.R., E. Leonard, S.H. Yoon, H.C. Tseng, C. Yuan, K.L.J. Parther (2009). Engineering alternative butanol production platforms in heterologous bacteria. Metab. Eng., 11, 262-273.
Nigam, P.S. and A. Singh (2011). Production of liquid biofuels from renewable resources. Prog. Ener. Combust. Sci., 37, 52-68.
Nilvebrant, N.O., A. Reimann, S. Larsson and L.J. Jonsson (2001). Detoxification of lignocellulose hydrolysates with ion exchange resins. Appl. Biochem. Biotechnol., 91, 35-49.
Nuttha, T., S. Navankasattusas and S.T. Yang (2009). Production of lactic acid and ethanol by Rhizopus oryzae integrated with cassava pulp hydrolysis. Bioprocess. Biosyst. Eng., DOI 10.1007/s00449-009-0341-x.
Olsson, L and B. Hahn-Hagerdal (1996). Fermentation of lignocellulosic hydrolysates for ethanol production. Enzyme Microb. Technol., 18, 312-331.
Oudshoorn, A., L.A.M. van der Wielen, and A.J.J. Straathof (2009). Adsorption equilibria of bio-based butanol solutions using zeolite. Biochem. Eng. J., 48, 99-103.
Palmarola-Adrados, B., M. Galbe and G. Zacchi (2004). Combined steam pretreatment and enzymatic hydrolysis of starch-free wheat fibers. Appl. Biochem. Biotechnol., 113, 989-1002.
Palmqvist, E., H. Grage, N.Q. Meinander and B. Hahn-Hagerdal (1999). Main and interaction effects of acetic acid, furfural and p-hydroxybenzoic acid on growth and ethanol productivity of yeasts. Biotechnol. Bioeng., 63, 46-55.
Palmqvist, E. and B. Hahn-Hagerdal (2000a). Fermentation of lignocellulosic hydrolysates. I: inhibition and detoxification. Bioresour. Technol., 74, 17-24.
Palmqvis, E. and B. Hahn-Hagerdal (2000b). Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol., 74, 25-33.
Pandey, A., C.R. Soccol, P. Nigam and V.T. Soccol (2000a). Biotechnological potential of agro-industrial residues. I: sugarcane bagasse. Bioresour. Technol., 74, 69-80.

279
Pandey, A., C.R. Soccol, P. Nigam, V.T. Soccol, L.P.S. Vandenberghe and R. Mohan (2000b). Biotechnological potential of agro-industrial residues. II: cassava bagasse. Bioresour. Technol., 74, 81-87.
Papoutsakis, E.T. (2008). Engineering solventogenic clostridia. Curr. Opin. Biotechnol., 19, 420-429.
Paredes, C., I. Rigoutsos, and E.T. Papoutsakis (2004). Transcriptional organization of the Clostridium acetobutylicum genome. Nucleic Acid Res., 32, 1973-1981.
Paredes, C., K.V. Alsaker, and E.T. Papoutsakis (2005). A comparative genomic view of clostridial sporulation and physiology. Nat. Rev., 3, 969-978.
Park, C., M. Okos and P. Wankat (1989). Acetone-butanol-ethanol (ABE) fermentation in an immobilized cell tickle bed reactor. Biotechnol. Bioeng., 34, 18-29.
Parekh, M., J. Formanek and H.P. Blaschek (1998). Development of a cost-effective glucose-corn steep medium for production of butanol by Clostridium beijerinckii. J. Industrial Microbiol. Biotechnol., 21, 187-191.
Parekh, M., J. Formanek, and H.P. Blaschek (1999). Pilot-scale production of butanol by Clostridium beijerinckii BA 101 using a low-cost fermentation medium used on corn steep water. Appl. Microbiol. Biotechnol., 51, 152-157.
Perez, J., J.M. Dorado, T.D. Rubia and J. Martinez (2002). Biodegradation and biological treatment of cellulose, hemicellulose and lignin: An overview. Int. Microbiol., 5, 53-63.
Pfromm, P.H., V. Amanor-Boadu, R. Nelson, P. Vadlani, R. Madl (2010). Bio-butanol vs. bio-ethanol: a technical and economic assessment for corn and switchgrass fermented by yeast or Clostridium acetobutylicum. Biomass Bioenergy, 34, 515-524.
Pierrot, P., M. Fick, and J.M. Engasser (1986). Continuous acetone-butanol fermentation with high productivity by cell ultrafiltration and recycling. Biotechnol. Lett., 8, 253-256.
Qureshi, N., I.S. Maddox, and A. Friedl (1992). Application of continuous substrate feeding to the ABE fermentation: relief of product inhibition using extraction, perstraction, stripping and pervaporation. Biotechnol. Prog., 8, 382-390.

280
Qureshi, N. and H.P. Blaschek (1999a). Butanol recovery from model solution/ fermentation broth by pervaporation: evaluation of membranes performance. Biomass and Bioenergy, 17, 175-184.
Qureshi, N. and H.P. Blaschek (1999b). Production of acetone butanol ethanol (ABE) by a hyper-producing mutant strain of Clostridium beijerinckii BA101 and recovery by pervaporation. Biotechnol. Prog., 15, 594-602.
Qureshi, N. and H.P. Blaschek (1999c). Fouling studies of a pervaporation membrane with commercial fermentation media and fermentation broth of hydrper-butanol-producing Clostridium beijerinckii BA101. Sep. Sci. Technol., 34, 2803-2815.
Qureshi N. and H.P. Blaschek (2000a). Butanol production using Clostridium beijerinckii BA101 hyper-butanol producing mutant strain and recovery by pervaporation. Appl. Biochem. Biotechnol., 84, 225-235.
Qureshi, N. and H.P. Blaschek (2000b). Economics of butanol fermentation using hyper-butanol producing Clostridium beijerinckii BA 101. Trans. IChem E., 78, 139-144.
Qureshi, N. and H.P. Blaschek (2001a). Recovery of butanol from fermentation broth by gas stripping. Renewable Energy, 22, 557-564.
Qureshi, N. and H.P. Blaschek (2001b). Recent advances in ABE fermentation: hyper-butanol producing Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 27, 287-291.
Qureshi, N. and H.P. Blaschek (2001c). ABE production from corn: a recent economic evaluation. J. Ind. Microbio. Biotechnol., 27, 292-297.
Qureshi, N. and I.S. Maddox (1988). Reactor Design for the ABE fermentation using cells of Clostridium acetobutylicum immobilized by adsorption onto bonechar. Bioprocess Eng., 3, 69-72.
Qureshi, N. and I.S. Maddox (1991). Integration of continuous production and recovery of solvents from whey permeate: use of immobilized cells of Clostridium acetobutylicum in a fluidized bed reactor coupled with gas stripping. Bioproc. Eng., 6, 63-69.

281
Qureshi, N. and I.S. Maddox (1995). Continuous production of acetone-butanol- ethanol using immobilized cells of Clostridium acetobutylicum and integration with product removal by liquid-liquid extraction. J. Ferment. Bioeng., 80, 185-189.
Qureshi, N. and I.S. Maddox (2005). Reduction in butanol inhibition by perstraction: Utilization of concentrated lactose/whey permeate by Clostridium acetobutylicum to enhance butanol fermentation economics. Trans IChemE, Part C, Food and Bioproducts Processing, 83, 43-52.
Qureshi, N., M.M. Meagher and R.W. Hutkins (1999). Recovery of butanol from model solutions and fermentation broth using silicalite/silicone membrane. J. Membr. Sci., 158, 115-125.
Qureshi, N., J. Schripsema, J. Lienhardt and H.P. Blaschek (2000). Continuous solvent production by Clostridium beijerinckii BA 101 immobilized by adsorption onto brick. J. Microbiol. Biotechnol., 16, 377-382.
Qureshi, N., M.M. Meagher, Jicai Huang, R.W. Hutkins (2001a). Acetone butanol ethanol (ABE) recovery by pervaporation using silicate-silicone composite membrane from fed-batch reactor of Clostridium acetobutylicum. J. Membrane Sci., 187, 93-102.
Qureshi, N., A. Lolas and H.P. Blaschek (2001b). Soy molasses as fermentation substrate for production of butanol using Clostridium beijerinckii BA101. J. Ind. Microbiol. Biotechnol., 26, 290-295.
Qureshi, N., L.L. Lai, and H.P. Blaschek (2004). Scale-up of a high productivity continuous biofilm reactor to produce butanol by adsorbed cells of Clostridium beijerinckii. Food Bioprod. Process, 82, 164-173.
Qureshi, N., S. Hughes, I.S. Maddox, M.A. Cotta (2005). Energy-efficient recovery of butanol from model solutions and fermentation broth by adsorption. Bioprocess Biosyst. Eng., 27, 215-222.
Qureshi, N., X. Li, S. Hughes, B.C. Saha and M.A. Cotta (2006). Butanol production from corn fiber xylan using Clostridim acetobutylicum. Biotechnol. Prog., 22, 673-680.
Qureshi, N., B.C. Saha and M.A. Cotta (2007). Butanol production from wheat straw hydrolysate using Clostridium beijerinckii. Bioprocess Biosyst. Eng., 30, 419-427.

282
Qureshi, N. and T.C. Ezeji (2008). Butanol, ‘a superior biofuel’ production from agricultural residues (renewable biomass): recent progress in technology. Biofuels, Bioprod. Bioref., 2, 319-330.
Qureshi, N., T.C. Ezeji, J. Ebener, B.S. Dien, M.A. Cotta and H.P. Blaschek (2008a). Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol., 99, 5915-5922.
Qureshi, N., B.C. Saha, R.E. Hector, M.A. Cotta (2008b). Removal of fermentation inhibitors from alkaline peroxide pretreated and enzymatically hydrolyzed wheat straw: production of butanol from hydrolysate using Clostridium beijerinckii in batch reactors. Biomass Bioenergy, 32, 1353-1358.
Qureshi, N., B.C. Saha, R. E. Hector, S.R. Hughes, M.A. Cotta (2008c). Butanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part I- Batch fermentation. Biomass Bioener., 32, 168-175.
Qureshi, N., B.C. Saha and M.A. Cotta (2008d). Butanol production from wheat straw by simultaneous saccharification and fermentation using Clostridium beijerinckii: Part II- Fed-batch fermentation. Biomass Bioener., 32, 176-183.
Qureshi, N., B.C. Saha, B. Dien, R.E. Hector, M.A. Cotta (2010a). Production of butanol (a biofuel) from agricultural residues: Part I-Use of barley straw hydrolysate. Biomass Bioenergy, 34, 559-565.
Qureshi, N., B.C. Saha, R.E. Hector, B. Dien, S. Hughes, S. Liu, L. Iten, M.J. Bowman, G. Sarath, M.A. Cotta (2010b). Production of butanol (a biofuel) from agricultural residues: Part II-Use of corn stover and switchgrass hydrolysates. Biomass Bioenergy, 34, 566-571.
Rabinovich, M.L., M.S. Melnik and A.V. Bolobova (2002). Microbial cellulases: A review. Appl. Biochem. Microbiol., 38, 305-321.
Rajagopalan, S., R.P. Datar, and R.S. Lewis (2002). Formation of ethanol from carbon monoxide via a new microbial catalyst. Biomass and bioenergy, 23, 487-493.
Rao G. and R. Mutharasan (1987). Altered electron flow in continuous cultures of Clostridium acetobutylicum induced by viologen dyes. Appl. Environ. Microbiol., 53, 1232-1235.

283
Ravi, V.P., R.V. Jasra, and T.S.G. Bhat (1998). Adsorption of phenol, cresol isomers and benzyl alcohol from aqueous solution on activated carbon at 278, 298 and 323 K. J. Chem. Technol. Biot., 71, 173-179.
Reddy, N. and Y. Yang (2005). Biofibers from agricultural byproducts for industrial applications. TRENDS in Biotechnol., 23, 22-27.
Roberto, I.C., M.G.A. Felipe, I.M. Mancilha, M. Vitola, S. Sato and S.S. Silva (1995). Xylitol production by Candida guilliermondii as an approach for the utilization of agroindustrial residues. Bioresour. Technol., 51, 255-257.
Rodrigues, R.C.L.B., M.G.A. Felipe, J.B.A.E. Silva, M. Vitolo, P.V. Gomez (2001). The influence of pH, temperature and hydrolyzate concentration on the removal of volatile and nonvolatile compounds from sugarcane bagasse hemicellulosic hydrolyzate treated with activated charcoal before or after vacuum evaporation. Braz. J. Chem. Eng., 18, 299-311.
Roffler, S.R., H.W. Blanch, and C.R. Wilke (1987a). In-situ recovery of butanol during fermentation, part 1: batch extractive fermentation. Bioprocess Eng., 2, 1-12.
Roffler, S.R., H.W. Blanch, and C.R. Wilke (1987b). In-situ recovery of butanol during fermentation, part 2: fed-batch extractive fermentation. Bioprocess Eng., 2, 181-190.
Roffler, S.R., H.W. Blanch, C.R. Wilke (1988). In situ extractive fermentation of acetone and butanol. Biotechnol. Bioeng., 31, 135-143.
Saha, B.C. (2003). Hemicellulose bioconversion. J Ind Microbiol Biotechnol., 30, 279-291.
Sano, T., H. Yanagishita, Y. Kiyouzumi, F. Mizukami, H. Haraya (1994). Sepation of ethanol/water mixture by silicalite membrane on pervaporation. J. Membr. Sci., 95, 221-228.
Schlote, D. and G. Gottschalk (1986). Effect of cell recycle on continuous butanol-acetone fermentation with Clostridium acetobutylicum under phosphate limitation. Appl. Microbiol. Biotechnol., 24, 1-5.
Schmidt, S.L., M.D. Myers, S.S. Kelley, J.D. McMillan, N. Padukone (1997). Evaluation of PTMSP membranes in achieving enhanced ethanol removal from fermentations by pervaporation. Appl. Biochem. Biotechnol., 63-65, 469-482.

284
Seddon, K.R. (1997). Review: Ionic liquids for clean technology. J. Chem. Tech. Biotechnol., 68, 351-356.
Servinsky, M.D., J.T. Kiel, N.F. Dupuy, C.J. Sund (2010). Transcriptional analysis of different carbohydrate utilization by Clostridium acetobutylicum. Microbiology, 156, 3478-3491.
Shen, C.R. and J.C. Liao (2008). Metabolic engineering of Escherichia coli for 1-butanol and 1-propanol production via keto-acid pathways. Metab. Eng., 10, 312-320.
Shi, Z. and H.P. Blaschek (2008). Transcriptional analysis of Clostridium beijerinckii NCIMB 8052 and they hyper-butanol-producing mutant BA101 during shift from acidogensis to solventogensis. Appl. Environ. Microbiol., 74, 7709-7714.
Silva, E.M. and S.T. Yang (1995). Kinetics and stability of a fibrous-bed bioreactor for continuous production of lactic from unsupplemented acid whey. J. Biotechnol., 41, 59-70.
Somrutai, W., M. Takagi and T. Yoshida (1996). Acetone-butanol fermentation by Clostridium aurantibutyricum ATCC 17777 from a model medium for palm oil mill effluent. J. Ferment. Bioeng., 81, 543-547.
Soni, B.K., P. Soucaille and G.Goma (1987). Continuous acetone-butanol fermentation: a global approach for the improvement in the solvent productivity in synthetic medium. Appl. Microbiol. Biotechnol., 25, 317-321.
Sriroth, K., R. Chollakup, S. Chotineeranat, K. Piyachomkwan and C.G. Oates (2000). Processing of cassava waste for improved biomass utilization. Bioresour. Technol., 71, 63-69.
Sun, Y. and J. Cheng (2002). Hydrolysis of lignocellulosic material from ethanol production: A review. Bioresour. Technol., 83, 1-11.
Swana, J., Y. Yang, M. Behnam, R. Thompson (2011). An analysis of net energy production and feedstock availability for biobutanol and bioethanol. Bioresour. Technol., 102, 2112-2117.
Taylor, F., M.J. Kurantz, N. Goldberg, A.J. McAloon, J.C. Craig, Jr. (2000). Dry-grind process for fuel ethanol by continuous fermentation and stripping. Biotechnol. Prog., 16, 541-547.

285
Teymouri, F., L.L. Perez, H. Alizadeh and B.E. Dale (2004). Ammonia fiber explosion treatment of corn stover. Appl. Biochem. Biotechnol., 113, 951–963.
Taherzadeh, M.J. and K. Karimi (2007). Enzyme-based hydrolysis processes for ethanol from lignocellulosic materials: a review. Bioresour., 2, 707-738.
Tashiro, Y., K. Takeda, G. Kobayashi, K. Sonomoto, A. Ishizaki, S. Yoshino (2004). High butanol production by Clostridium saccharoperbuutylacetonicum N1-4 in fed-batch culture with pH-stat continuous butyric acid and glucose feeding method. J. Biosci. Bioeng., 98, 263-268.
Tashiro, Y., H. Shinto, M. Hayshi, S. Baba, G. Kobayashi, K. Sonomoto (2007). Novel high-efficient butanol production from butyrate by non-growing Clostridium saccharoperbutyl acetonicum N1-4 (ATCC 13564) with methyl viologen. J. Biosci. Bioeng., 104, 238-240.
Terracciano, J.S. and E.R. Kashket (1986). Intracellular conditions required for the initiation of solvent production by Clostridium acetobutylicum. Appl. Environ. Microbiol., 52, 86-91.
Thang, V.H., K. Kanda, and G. Kobayashi (2010). Production of acetone-butanol- ethanol (ABE) in direct fermentation of cassava by Clostridium saccharoperbutylacetonicum N1-4. Appl. Biochem. Biotechnol., 161, 157-170.
Thongchul, N., S. Navankasattusas, and S.T. Yang (2009). Production of lactic acid and ethanol by Rhizopus oryzae integrated with cassava pulp hydrolysis. Bioproc. Biosyst. Eng., 33, 407-416.
Thongsukmak, A. and K.K. Sirkar (2007). Pervaporation membranes highly selective for solvents present in fermentation broth. J. Memb.Sci., 302, 45-48.
Thormann, K. and P. Dürre (2001). Orf5/SolR: a transcriptional repressor of the sol operon of Clostridium acetobutylicum? J. Ind. Microbiol. Biotechnol., 27, 307-313.
Thormann, K., L. Feustel, K. Lorenz, S. Nakotte, P. Dürre (2002). Control of butanol formation in Clostridium acetobutylicum by transcritional activation. J. Bacteriol., 184, 1966-1973.
Toh, S.L.I., J. McFarlane, C. Tsouris, D.W. DePaoli, H. Luo, S. Dai (2006). Room-temperature ionic liquids in liquid-liquid extraction: effects of solubility in

286
aqueous solutions on surface properties. Solvent Extraction and Ion Exchange, 24, 33-56.
Vane, L.M. (2005). Review: A review of Pervaporation for product recovery from biomass fermentation processes. J. Chem. Technol. Biotechnol., 80, 603-629.
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuls, Bioprod. Bioref., 2, 553-588.
Varga, E., K. Reczey and G. Zacchi (2004). Optimization of steam pretreatment of corn stover to enhance enzymatic digestibility. Appl. Biochem. Biotechnol., 113, 509-523.
Volkov, V.V., V.S. Khotimsky, E.G. Litvinova, A.G. Fadeev, YaA Selinskaya, N.A. Plate, J.D. McMillan, S.S. Kelley (1997). Development of novel poly[-1-(trimethylsilyl)-1-propyne] based materials for pervaporation separation in biofuel production. Polym. Mater. Sci. Eng., 77, 339-340.
Volkov, V.V., A.G. Fadeev, V.S. Khotimsky, E.G. Litvinova, YaA Selinskaya, J.D. McMillan, S.S. Kelley (2004). Effects of synthesis conditions on the pervaporation properties of poly[-1-(trimethylsilyl)-1-propyne] useful for membrane bioreactors. J. Appl. Polym. Sci., 91, 2271-2277.
Vrana, D.L., M.M. Meagher, R.W. Hutkins, B. Duffield (1993). Pervaporation of model acetone-butanol-ethanol fermentation product solutions using Polytetrafluoroethylene membranes. Sep. Sci. Technol., 28, 2167-2178.
Wang, B., T. Ezeji, Z. Shi, H. Feng, H.P. Blaschek (2009). Pretreatment and conversion of distiller’s dried grains with solubles for acetone-butanol-ethanol (ABE) production. Transactions of ASABE, 52, 885-892.
Wayman, M. and R. Parekh (1987). Production of acetone-butanol by extractive fermentation using dibutylphthalate as extractant. J. Ferment. Technol., 65, 295-300.
Weber, C., A. Farwick, F. Benisch, D. Brat, H. Dietz, T. Subtil, E. Boles (2010). Trends and challenges in the microbial production of lignocellulosic bioalcohol fuels. Appl. Microbiol. Biotecnol., 87, 1303-1315.
Wei, G.T., Z. Yang and C.J. Chen (2003). Room temperature ionic liquid as a novel medium for liquid/liquid extraction of metal ions. Analytica Chimica Acta, 488, 183-192.

287
Woods, D.R. (1995). The genetic engineering of microbial solvent production. Trends Biotechnol., 13, 259-264.
Wu, Z. and S.T. Yang (2003). Extractive fermentation for butyric acid production from glucose by Clostridium tyrobutyricum. Biotechnol. Bioeng., 82, 93-102.
Yang, X., G.J. Tsai and G.T. Tsao (1994). Enhancement of in situ adsorption on the acetone-butanol fermentation by Clostridium acetobutylicum. Separation Techonol., 4, 81-92.
Yang, X. and G.T. Tsao (1995). Enhanced acetone-butanol fermentation using repeated fed-batch operation coupled with cell recycle by membrane and simultaneous removal of inhibitory products by adsorption. Biotechnol. Bioeng., 47, 444-450.
Yang, S.T. (1996). Extractive fermentation using convoluted fibrous bed bioreactor. United States Patent, US patent 5563069.
Yat, S.C., A. Berger and D.R. Shonnard (2008). Kinetic characterization of dilute surface acid hydrolysis of timber varieties and switchgrass. Bioresour. Technol., 99, 3855-3863.
Yu, M., Y. Zhang, I. Tang, S.T. Yang (2011). Metabolic engineering of Clostridium tyrobutyricum for n-butanol production. Metab. Eng., in press, doi:10.1016/j.ymben.2011.04.002.
Zautsen, R.R.M., F. Maugeri-Filho, C.E. Vaz-Rossell, A.J.J. Straathof, L.A.M. van der Wielen, J.A.M. de Bont (2009). Liquid-liquid extraction of fermentation inhibiting compounds in lignocellulose hydrolysate. Biotechnol. Bioeng., 102, 1354-1360.
Zhang, L., D. Li, L. Wang, T. Wang, L. Zhang, X.D. Chen and Z. Mao (2008). Effect of steam explosion on biodegradation of lignin in wheat straw. Bioresour. Technol., 99, 8512-8515.
Zhang, Y., Y. Ma, F. Yang, C. Zhang (2009). Continous acetone-butanol-ethanol production by corn stalk immobilized cells. J. Ind. Microbiol. Biotechnol., 36, 1117-1121.
Zhao, H., S. Xia and P. Ma (2005). Review: use of ionic liquids as “green” solvents for extractions. J. Chem. Technol. Biotechnol., 80, 1089-1096.

288
Zheng, Y.N., L.Z. Li, M. Xian, Y.J. Ma, J.M. Yang, X. Xu, D.Z. He (2009). Problems with the microbial production of butanol. J. Ind. Microbiol. Biotechnol., 36, 1127-1138.
Zhu, Y. and S.T. Yang (2003). Adaption of Clostridium tyrobutyricum for enhanced tolerance to butyric acid in a fibrous bed bioreactor. Biotechnol. Progress, 19, 365-372.
Zhu, Y., Z. Wu, and S.T. Yang (2002). Butyric acid production from acid hydrolysate of corn fibre by Clostridium tyrobutyricum in a fibrous bed bioreactor. Process Biochem., 38, 657-666.

289
Appendix A: Analytical Procedures
A.1 Gas chromatograph
For analysis of solvents and acids in the fermentation broth, samples were
centrifuged at 13.2 g for at least 5 min to remove any cell bodies or solids. Samples
containing acetone, butanol, ethanol, acetic acid, and butyric acid, were analyzed with a
Shimadzu GC-2014 gas chromatograph (GC) (Shimadzu, Columbia, MD, USA)
equipped with a flame ionization detector (FID) and a 30.0 m fused silica column (0.25m
film thickness and 0.25 mm ID, Stabilwax-DA). External standard and internal standard
methods were both used to analyze the concentration of solvents and acids in the sample.
When using external standard methods, samples were diluted at least 10-fold using 1%
phosphoric acid buffer solution to a final volume of 1 ml in the vial. To reduce the
injection mechanic error margin, internal standard method was later developed.
Isobutanol and isobutyric acid were used as internal standards for analyzing the solvents
and acids present in the sample. An internal standard buffer solution containing 0.5 g/L
isobutanol, 0.1 g/L isobutyric acid, and 1% phosphoric acid was used to dilute each
sample 20 times for acidification and calibration prior to analysis on GC. The final

290
volume in the vial was 1 ml. The gas chromatograph was operated at an injection
temperature of 200 oC with 1 μL of the acidified sample injected by the AOC-20i
Shimadzu auto injector. Column temperature was held at 80 oC for 3 min, raised to 150
oC at a rate of 30 oC/min, and held at 150 oC for 3.7 min. GC chromatogram of the
standard sample using external standard method and internal standard method is shown in
Figure A.1. Typical GC chromatogram of the ABE fermentation samples is shown in
Figure A.2 and A.3. GC chromatogram of the condensate recovered during the gas
stripping process is shown in Figure A.4 and A.5.
A.2. High performance liquid chromatograph
The concentration of carbohydrates (glucose, xylose and arabinose), acids (acetic
acid and butyric acid), and solvent (butanol) were analyzed by a high performance liquid
chromatography (HPLC) with an organic acid column (Bio-Rad HPX-87, ion exclusion
organic acid column, 300 mm × 7.8mm). Samples were centrifuged at 13.2 g for 5 min in
microcentrifuge tubes and diluted 10 times with distilled water prior to analysis on HPLC.
HPLC was run at 45 oC using 0.01N H2SO4 as the eluent at a flow rate of 0.6 ml/min.
15μL sample was injected by an automatic injector (SIL-10Ai) and the running time was
set at 36 min. A refractive index (RI) detector (Shimadzu RID-10A) was set at the range
of 200 to detect the organic compounds in the sample. The HPLC column was installed in
a column oven (CTO-10A) with temperature control at 45 oC. Peak height was used to

291
calculate concentration of sugars in the sample based on the peak height of standard
sample. In this project, HPLC is primarily used to analyze the carbohydrate concentration
in the samples, which can not be analyzed by YSI such as xylose and arabinose. HPLC
chromatogram of the standard sample is shown in Figure A.6. HPLC chromatogram of
several lignocellulosic hydrolysates used in this project is shown in Figure A.7, A.8 and
A.9. HPLC can also analyze the acids and solvent present in the fermentation samples,
and the HPLC chromatogram of several fermentation samples is shown in Figure A.10,
A.11 and A.12.

1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 min
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
5.5
6.0
6.5
7.0
7.5
8.0uV(x100,000) Chromatogram
Ace
ton
e/1
.52
4
Eth
ano
l/1.8
16 But
ano
l/3.5
67
Ace
tic A
cid
/6.0
99
But
yric
Aci
d/7
.679
A
1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 min
-0.25
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
2.25
2.50
2.75
3.00
3.25
3.50
3.75
4.00uV(x100,000) Chromatogram
/1.1
80
Ace
ton
e/1
.40
4/1
.53
8E
tha
nol/1
.65
5
/2.4
04
Isob
uta
nol/2
.60
3 But
ano
l/3.2
37
/5.8
00
Ace
tic A
cid
/5.8
64
Isob
utyr
ic a
cid/
6.7
71
But
yric
Aci
d/7
.366
B
Figure A.1 GC chromatogram of the standard sample containing acetone, butanol, ethanol, acetic acid and butyric acid using external standard and internal standard methods. (A) External standard method (1g/L each), (B) Internal standard method (0.5 g/L each).
292

1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 min
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0uV(x100,000) Chromatogram
Ace
ton
e/1
.52
7
/1.7
50
Eth
ano
l/1.8
49
But
ano
l/3.5
80
/4.1
72
/5.0
81
/5.2
26
Ace
tic A
cid
/6.1
17
/6.3
17
/7.3
43
But
yric
Aci
d/7
.697
Figure A.2 GC chromatogram of 10-fold diluted ABE fermentation sample containing 8.2 g/L acetone, 16.6 g/L butanol, 2.7 g/L ethanol, 5.1 g/L acetic acid and 2.9 g/L butyric acid (External standard method).
293

1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 min
-0.25
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
2.25
2.50
2.75
3.00
3.25
3.50
3.75
4.00uV(x100,000) Chromatogram
/0.8
43
/1.1
77
/1.2
84
Ace
ton
e/1
.40
7
Eth
ano
l/1.6
55
Isob
uta
nol/2
.59
7
But
ano
l/3.1
88
Ace
tic A
cid
/5.8
63
/6.0
61
Isob
utyr
ic a
cid/
6.7
65
/7.0
36
But
yric
Aci
d/7
.368
Figure A.3 GC chromatogram of 20-fold diluted ABE fermentation sample containing 2.54 g/L acetone, 8.15 g/L butanol, 0.21 g/L ethanol, 0.96 g/L acetic acid and 1.66 g/L butyric acid (Internal standard method).
294

1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 min
-0.1
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1.1
1.2
1.3
1.4
1.5
1.6uV(x1,000,000) Chromatogram
/0.8
02
/1.1
84
Ace
ton
e/1
.40
3
Eth
ano
l/1.6
54
/2.0
55
But
ano
l/3.2
42
/3.8
42
Ace
tic A
cid
/5.8
71
But
yric
Aci
d/7
.366
Figure A.4 GC chromatogram of 100-fold diluted condensate sample from gas stripping process containing 26.1 g/L acetone, 166.6 g/L butanol, 4.8 g/L ethanol, 0.46 g/L acetic acid, and 1.0 g/L butyric acid (External standard method).
295
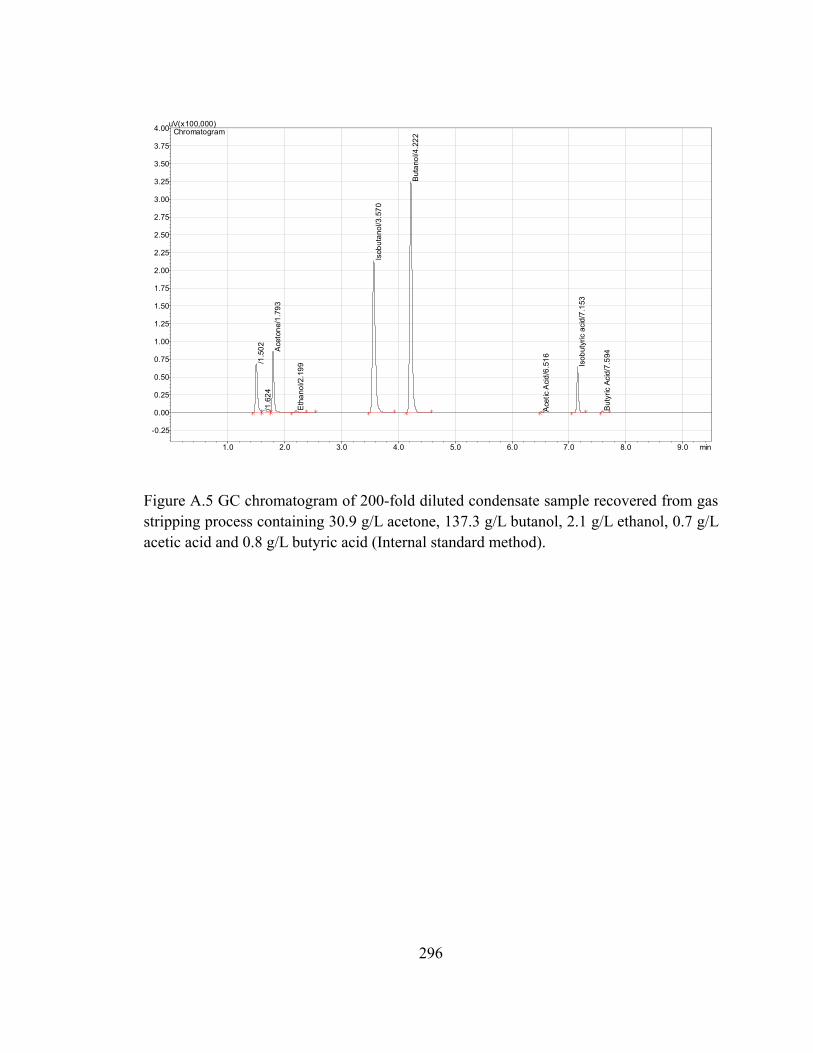
1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 min
-0.25
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
2.25
2.50
2.75
3.00
3.25
3.50
3.75
4.00uV(x100,000) Chromatogram
/1.5
02
/1.6
24
Ace
ton
e/1
.79
3
Eth
ano
l/2.1
99
Isob
uta
nol/3
.57
0
But
ano
l/4.2
22
Ace
tic A
cid
/6.5
16
Isob
utyr
ic a
cid/
7.1
53
But
yric
Aci
d/7
.594
Figure A.5 GC chromatogram of 200-fold diluted condensate sample recovered from gas stripping process containing 30.9 g/L acetone, 137.3 g/L butanol, 2.1 g/L ethanol, 0.7 g/L acetic acid and 0.8 g/L butyric acid (Internal standard method).
296

Minutes0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36
uR
IU
-10.0
-7.5
-5.0
-2.5
0.0
2.5
5.0
7.5
10.0
12.5
15.0
17.5
20.0
22.5
25.0
27.5
30.0
uRIU
-10.0
-7.5
-5.0
-2.5
0.0
2.5
5.0
7.5
10.0
12.5
15.0
17.5
20.0
22.5
25.0
27.5
30.0
Glu
cose
9.4
50
2.0
00
CA
LX
ylo
se 1
0.1
67
2.0
00 C
AL
Ara
bin
ose
11.
317
2.0
00
CA
L 1
2.5
33
0.0
00
Lac
tic A
cid
14
.13
3 2
.000
CA
L
Ace
tic A
cid
14
.91
7 2
.00
0 C
AL
1
9.3
67
0.0
00
Bu
tyric
Aci
d 2
1.5
33
2.0
00 C
AL
Bu
tan
ol
33
.15
0 2
.000
CA
L
RID-10A091510
NameRetention TimeESTD concentration
Figure A.6 HPLC chromatogram of the standard sample containing glucose, xylose, arabinose, acetic acid, lactic acid, butyric acid and butanol (2 g/L each).
297

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
uRIU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
Glu
cose
8.7
67
26
.96
3X
ylo
se 9
.400
22
.66
3
Ara
bin
ose
10
.26
7 1
1.2
41
1
1.4
33 0
.00
0
11.
867
0.0
00
(Lac
tic A
cid
)
0.0
00
BD
L
13
.06
7 0
.000
Ace
tic A
cid
14
.86
7 2
.567
But
yric
Aci
d 2
1.3
67 0
.07
2
(Bu
tan
ol)
0
.00
0 B
DL
RID-10A050811
NameRetention TimeESTD concentration
Figure A.7 HPLC chromatogram of the 10-fold diluted corn fiber hydrolysate containing 27.0 g/L glucose, 22.7 g/L xylose, 11.2 g/L arabinose, 2.6 g/L acetic acid.
298

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
uRIU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
Glu
cose
8.7
67
42
.14
4X
ylo
se 9
.383
1.7
99
Ara
bin
ose
10.
233
0.0
27
1
0.6
00 0
.00
0
11
.43
3 0
.00
0
Lac
tic A
cid
12.
433
0.2
15
1
3.0
50 0
.00
0
13.
667
0.0
00
Ace
tic A
cid
14.
867
0.4
84
(Bu
tyri
c A
cid
)
0.0
00
BD
L
(Bu
tan
ol)
0
.00
0 B
DL
RID-10A050811
NameRetention TimeESTD concentration
Figure A.8 HPLC chromatogram of the 10-fold diluted cassava bagasse hydrolysate containing 42.1 g/L glucose, 1.80 g/L xylose, 0.22 g/L lactic acid and 0.48 g/L acetic acid.
299

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
uRIU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
Glu
cose
8.7
67
9.1
20X
ylo
se 9
.383
39
.67
3
Ara
bin
ose
10
.26
7 2
.121
1
1.9
00 0
.00
0(L
act
ic A
cid
)
0.0
00
BD
L
13
.05
0 0
.000
Ace
tic A
cid
14
.63
3 0
.05
9
(Bu
tyric
Aci
d)
0.0
00
BD
L
(But
ano
l)
0.0
00 B
DL
RID-10A050811
NameRetention TimeESTD concentration
Figure A.9 HPLC chromatogram of the 10-fold diluted wood pulp hydrolysate containing 9.1 g/L glucose, 39.7 g/L xylose, 2.1 g/L arabinose and 0.06 g/L acetic acid.
300

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
uRIU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
Glu
cose
8.7
67
60
.47
1X
ylo
se 9
.40
0 2
9.9
60
Ara
bin
ose
10
.26
7 1
.77
8
Lac
tic A
cid
12
.43
3 0
.232
1
3.65
0 0
.00
0
Ace
tic A
cid
14.
867
6.2
31
(But
yric
Aci
d)
0.
000
BD
L
2
3.3
17
0.0
00
3
2.2
00
0.0
00
(But
ano
l)
0.0
00 B
DL
RID-10A050811
NameRetention TimeESTD concentration
Figure A.10 HPLC chromatogram of the 20-fold diluted sugarcane bagasse hydrolysate containing 60.5 g/L glucose, 30.0 g/L xylose, 1.8 g/L arabinose, 0.2 g/L lactic acid and 6.2 g/L acetic acid.
301

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
uRIU
-15
-10
-5
0
5
10
15
20
25
30
Glu
cose
9.5
17
5.8
30 Xyl
ose
10
.21
7 1
4.1
90
(Ara
bin
ose)
0
.00
0 B
DL
(La
ctic
Aci
d)
0
.000
BD
L
Ace
tic A
cid
14
.96
7 1
.51
3
2
0.4
00
0.0
00
Bu
tyri
c A
cid
21
.48
3 1
.89
4
But
ano
l 33
.10
0 5
.36
5
RID-10A011111
NameRetention TimeESTD concentration
Figure A.11 HPLC chromatogram of 10-fold diluted ABE fermentation sample using glucose and xylose as substrates containing 5.8 g/L glucose, 14.2 g/L xylose, 1.5 g/L acetic acid, 1.9 g/L butyric acid and 5.4 g/L butanol.
302

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
uRIU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
Glu
cose
8.7
67
19.
769
Xyl
ose
9.4
00
11.
530
Ara
bin
ose
10
.26
7 3
.523
Lact
ic A
cid
12
.43
3 0
.968
1
3.0
67
0.0
00
Ace
tic A
cid
14.
867
5.7
34
1
5.8
67
0.0
00
1
7.5
83
0.0
00
Bu
tyric
Aci
d 2
1.4
50
5.7
76
22.1
50
0.0
00
But
ano
l 36
.93
3 3
.81
7
RID-10A050811
NameRetention TimeESTD concentration
Figure A.12 HPLC chromatogram of 10-fold diluted ABE fermentation sample using corn fiber hydrolysate as substrate containing 19.8 g/L glucose, 11.5 g/L xylose, 3.5 g/L arabinose, 1.0 g/L lactic acid, 5.7 g/L acetic acid, 5.8 g/L butyric acid and 3.8 g/L butanol.
303

Minutes
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
uR
IU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
uRIU
-15
-10
-5
0
5
10
15
20
25
30
35
40
45
50
55
60
Glu
cose
8.7
67 2
3.8
81
Xyl
ose
9.3
67
0.6
12
(Ara
bin
ose
)
0.0
00
BD
L
10.
533
0.0
00
1
1.51
7 0
.00
0
Lac
tic A
cid
12.
433
0.5
15
1
3.0
67
0.0
00
Ace
tic A
cid
14
.86
7 5
.529
1
5.8
67
0.0
00
1
7.6
67
0.0
00
1
8.60
0 0
.00
0
But
yric
Aci
d 2
1.4
33
4.7
24
2
2.1
67 0
.00
0
23
.36
7 0
.000
24
.81
7 0
.000
Bu
tan
ol
36
.91
7 7
.267
RID-10A050811
NameRetention TimeESTD concentration
Figure A.13 HPLC chromatogram of 10-fold diluted ABE fermentation sample using cassava bagasse hydrolysate as substrate containing 23.9 g/L glucose, 0.61 g/L xylose, 0.5 g/L lactic acid, 5.5 g/L acetic acid, 4.7 g/L butyric acid and 7.3 g/L butanol.
304

Minutes0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36
uR
IU
-10
-8
-6
-4
-2
0
2
4
6
8
10
12
14
uRIU
-10
-8
-6
-4
-2
0
2
4
6
8
10
12
14
8
.26
7 0
.00
0
Glu
cose
9.4
50
2.3
57
Xyl
ose
10.
183
13
.51
0
11
.05
0 0
.00
0(A
rab
ino
se)
0.
000
BD
L
La
ctic
Aci
d 1
3.9
17 1
.50
8
Ace
tic A
cid
14
.90
0 1
.577
1
6.7
67 0
.00
0
1
9.4
17 0
.00
0
2
0.4
67
0.0
00
Bu
tyri
c A
cid
21.
483
0.7
43
Bu
tan
ol
33.2
17
4.4
92
RID-10A091510
NameRetention TimeESTD concentration
Figure A.14 HPLC chromatogram of 10-fold diluted ABE fermentation sample using wood pulp hydrolysate as substrate containing 2.4 g/L glucose, 13.5 g/L xylose, 2.1 g/L arabinose, 1.5 g/L lactic acid, 1.6 g/L acetic acid, 0.7 g/L butyric acid and 4.5 g/L butanol.
305

306
Appendix B: Evaluation of Liquid-liquid Extraction Using Ionic Liquids for Butanol
Recovery
B.1 Materials and methods
Five solvents were evaluated as potential extractants for butanol recovery using
liquid-liquid extraction, including oleyl alcohol (OA), 2-ethyl-1-hexanol,
1-decyl-3-methyl-imidazolium tetracyanoborate (DMIM-TCB), 1-butyl-3-methyl-1H-
imidazol-3-ium (BMIM-PF6), and commercial diesel. DMIM-TCB and BMIM-PF6 are
two ionic liquids (ILs), which were kindly provided by Merck. Oleyl alcohol and
2-ehtyl-1-hexanol were also provided by Merck to be used as benchmarks. Since butanol
is of research interest to be used as a biofuel in the future, commercial diesel was also
investigated in this study as an extractant since butanol can be blended with diesel as a
fuel additive. The liquid-liquid extraction experiment was done in 15 ml centrifuge tubes
(BD falcon, Fisher Scientific). Fermentation broth and model solution were both studied.
Model solution contained 10.4 g/L acetone, 20.3 g/L butanol, 3.8 g/L ethanol, 4.3 g/L
acetic acid and 5.1 g/L butyric acid. Fermentation broth was obtained from ABE
fermentation using C. beijerinckii ATCC 55025, which contained 5.5 g/L acetone, 12.5

g/L butanol, 0.8 g/L ethanol, 1.6 g/L acetic acid, and 1.5 g/L butyric acid. The cell
concentration in the fermentation broth was 3.3 g/L. The organic/aqueous ratio in the
liquid-liquid extraction experiment was 1:1, with 2 ml of each phase. The mixing of the
two phases was performed using a vortex mixer (Fisher vortex Genie 2, Fisher Scientific)
at full speed for 10 seconds. The mixture was then settled for natural phase separation.
The concentration of acetone, butanol, ethanol, acetic acid and butyric acid was analyzed
by gas chromatograph, and the operating procedures can be referred to Appendix A.
Distribution coefficient and selectivity were used as two parameters determining the
performance of one extractant. They are defined as below:
component
olbu
ILcomponent
D
DS
aqcomponent
componentD
tan
][
][
[component] represents the concentration of one component.
B.2 Results and discussion
Ionic liquid (IL) is a group of salts that exist in the liquid form at low temperature
(<100oC) or room temperature, and is considered as a green and safe solvent due to its
thermally and chemically stable properties (Earle and Seddon, 2000). The miscibility and
hydrophobicity of ILs can be adjusted by manipulating the structure of anions and cations.
It was reported (Zhao et al., 2005) anions determine the water miscibility of ILs, whereas
307

308
cations have more influence on the hydrophobicity of ILs. ILs have been used as
extractants in many areas, such as organic acids (Matsumoto et al., 2004) and biofuels
(Fadeev and Meagher, 2001). [PF6]- based ILs are usually water-immiscible, and
1-butyl-3- methyl-1H-imidazol-3-ium ([BMIM][PF6]) has been identified as a suitable
extractant for butanol recovery (Fadeev and Meagher, 2001).
Among all five solvents evaluated, only BMIM-PF6 was found to be heavier than the
aqueous solutions (model solution and fermentation broth). The rest four solvents,
including DMIM-TCB, were all lighter than the aqueous solutions. BMIM-PF6, diesel,
and hexanol separated very quickly from the aqueous phase, whereas DMIM-TCB and
oleyl alcohol were slighted emulsified with the aqueous solution.
The concentration of acetone, butanol, ethanol, acetic acid, and butyric acid in the
aqueous phase (model solution and fermentation broth) before and after the liquid-liquid
extraction is shown in Figure B.1. Diesel was shown to be the least effective extractant
among all the five solvents evaluated for butanol recovery by liquid-liquid extraction. It
had low affinity towards butanol, acetone and ethanol, resulting in no significant butanol
concentration change in the aqueous phase before and after extraction. From Figure B.1,
it was shown that ethanol and acetic acid concentration remained almost the same before
and after liquid-liquid extraction in all extractants, indicating that all extractants had low
affinity towards these two compounds and did not selectively remove these two
compounds. Ionic liquids DMIM-TCB and BMIM-PF6 and hexanol had higher

309
distribution coefficient for acetone, selectively removing both acetone and butanol; oleyl
alcohol had lower distribution coefficient for acetone, only selectively removing butanol
but not acetone. DMIM-TCB, BMIM-PF6 and oleyl alcohol also had affinity to butyric
acid, extracting a portion of butyric acid from aqueous phase into organic phase.
The distribution coefficient of butanol, acetone, and butyric acid was summarized in
Table B.1 and illustrated in Figure B.2. When using model solution without cell present,
hexanol exhibited best distribution coefficient for butanol, followed by DMIM-TCB,
oleyl alcohol, and then BMIM-PF6 (Table B.1) In the case of fermentation broth which
contained 3.3 g/L cells, the butanol distribution coefficient was very similar between
hexanol and DMIM-TCB, followed by OA, BMIM-PF6, and diesel in a descending order
(Table B.1). It was noticed that traditional solvents oleyl alcohol and hexanol both
exhibited higher Dbutanol and Dacetone in model solution than in fermentation broth with
cells, whereas it was exactly the opposite when using ionic liquids. DMIM-TCB and
BMIM-PF6 both showed higher distribution coefficient for butanol and acetone in
fermentation broth than in model solution. Among all the five extractants evaluated,
DMIM-TCB had the highest Dbutanol and Dacetone in fermentation broth, and hexanol had
highest Dbutanol in model solution. Higher distribution coefficient is desirable since it
reduces the amount of extractant needed to recover a given amount of solvents (acetone
and butanol), reducing the capital cost of the liquid-liquid extraction system (Vane,
2008).

310
Selectivity of butanol over acetone and butanol over butyric acid was summarized in
Table B.3 and illustrated in Figure B.3. Since oleyl alcohol selectively removed mostly
butanol, but not much acetone, the selectivity of butanol over acetone was the highest for
oleyl alcohol among all the evaluated extractants, both in model solution and in
fermentation broth. Since ionic liquid BMIM-PF6 and DMIM-TCB selectively remove
both acetone and butanol, the S butanol/acetone was close to 1. This feature of the ionic liquid
was in favor of recovering both acetone and butanol from the fermentation broth, both of
which are valuable products from the ABE fermentation. Oleyl alcohol and hexanol
showed higher S butanol/acetone in the fermentation broth than in the model solution, whereas
ionic liquids showed similar S butanol/acetone under both conditions. It appeared that most of
extractants evaluated in this study also recovery butyric acid, except for hexanol. Oleyl
alcohol and hexanol both had significantly higher S butanol/butyric acid in the model solution
than in the fermentation broth, whereas ionic liquids showed higher S butanol/butyric acid in the
fermentation broth than in the model solution. The S butanol/butyric acid was improved in the
fermentation broth with ionic liquids, which was favorable since acids were not desired in
the recovered products.
B.3 References
Earle, M.J. and K.R. Seddon (2000). Ionic liquids. Green solvents for the future. Pure Appl. Chem., 72, 1391-1398.
Fadeev, A.G. and M.M. Meagher (2001). Opportunities for ionic liquids in recovery of

311
biofuels. Chem. Commun., 295-296.
Matsumoto, M., K. Mochiduki, K. Fukunishi, K. Kondo (2004). Extraction of organic acids using imidazolium-based ionic liquids and their toxicity to Lactobacillus rhamnosus. Separ. Purif. Technol., 40, 97-101.
Vane, L.M. (2008). Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuels, Bioprod. Bioref., 2, 553-588.
Zhao, H., S. Xia and P. Ma (2005). Review: use of ionic liquids as “green” solvents for extractions. J. Chem. Technol. Biotechnol., 80, 1089-1096.

312
Dbutanol Dacetone Dbutyric acid Extractant Model solution Broth Model solution Broth Model solution Broth
Oleyl alcohol 2.49 1.84 0.13 0.09 2.16 3.05
2-ethyl-1-hexanol 4.57 3.39 1.24 0.51 0.19 0.26
DMIM-TCB 3.30 3.60 1.76 2.26 3.01 1.21
BMIM-PF6 1.09 1.69 0.84 1.16 0.97 0.52
Diesel 0.18 0.23 0.10 0.25 0.13 0.013
Table B.1 Distribution coefficient of butanol, acetone, and butyric acid from different extractants in model solution and fermentation broth.
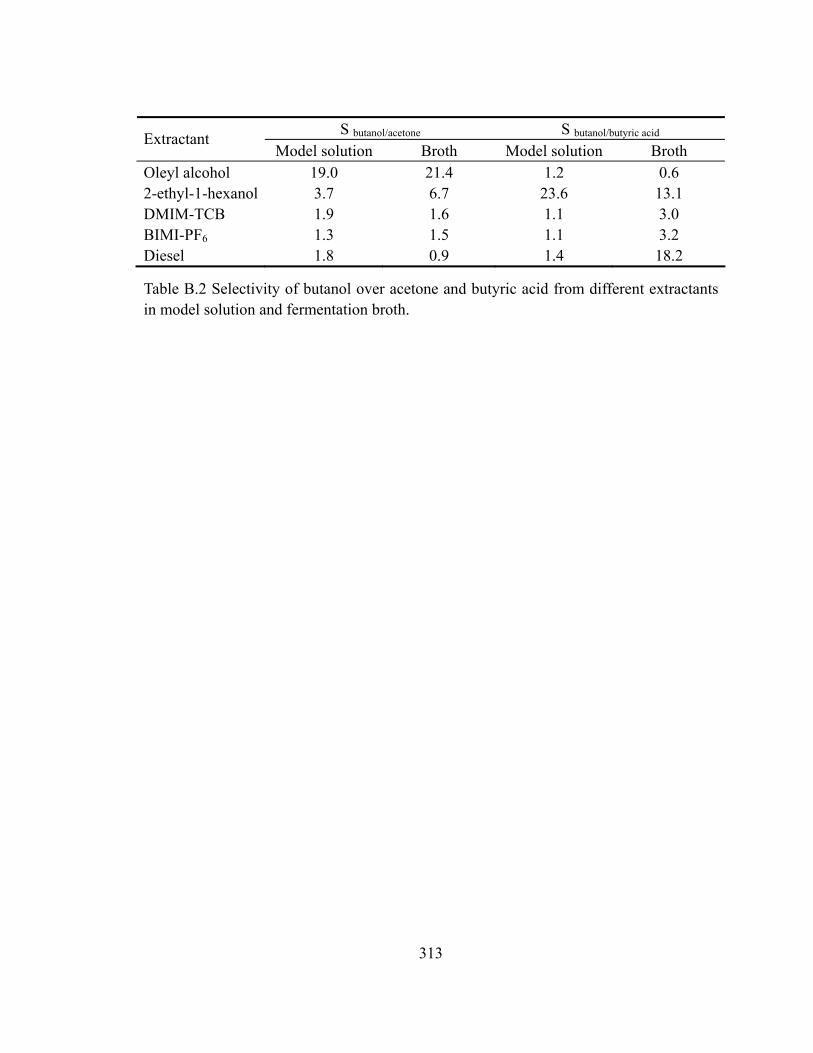
313
S butanol/acetone S butanol/butyric acid Extractant Model solution Broth Model solution Broth
Oleyl alcohol 19.0 21.4 1.2 0.6 2-ethyl-1-hexanol 3.7 6.7 23.6 13.1 DMIM-TCB 1.9 1.6 1.1 3.0 BIMI-PF6 1.3 1.5 1.1 3.2 Diesel 1.8 0.9 1.4 18.2
Table B.2 Selectivity of butanol over acetone and butyric acid from different extractants in model solution and fermentation broth.

0
5
10
15
20
25
Acetone Butanol Ethanol Acetic acid Butyric acid
Co
nce
ntr
atio
n (
g/L
)
Before
After DMIM-TCB
After BMIM-PF6
After diesel
After OA
After hexanol
A
0
2
4
6
8
10
12
14
Acetone Butanol Ethanol Acetic acid Butyric acid
Co
nce
ntr
atio
n (
g/L
)
Before
After DMIM-TCB
After BMIM-PF6
After diesel
After OA
After hexanol
B
Figure B.1 Concentration of acetone, butanol, ethanol, acetic acid, and butyric acid in the aqueous solution before and after the liquid-liquid extraction. (A) Model solution, (B) Fermentation broth.
314

0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5D
-bu
tan
ol
Model solution Fermentation broth
2-ethyl-1-hexanol
DMIM-TCB
Oleyl alcohol
BMIM-PF6
Diesel
A
0
0.5
1
1.5
2
2.5
D-a
ceto
ne
Model solution Fermentation broth
DMIM-TCB
BMIM-PF6
2-ethyl-1-hexanol
Oleyl alcohol
Diesel
B
Figure B.2 Comparison of butanol and acetone distribution coefficient of different extractants in model solution and fermentation broth. (A) Dbutanol, (B) Dacetone.
315
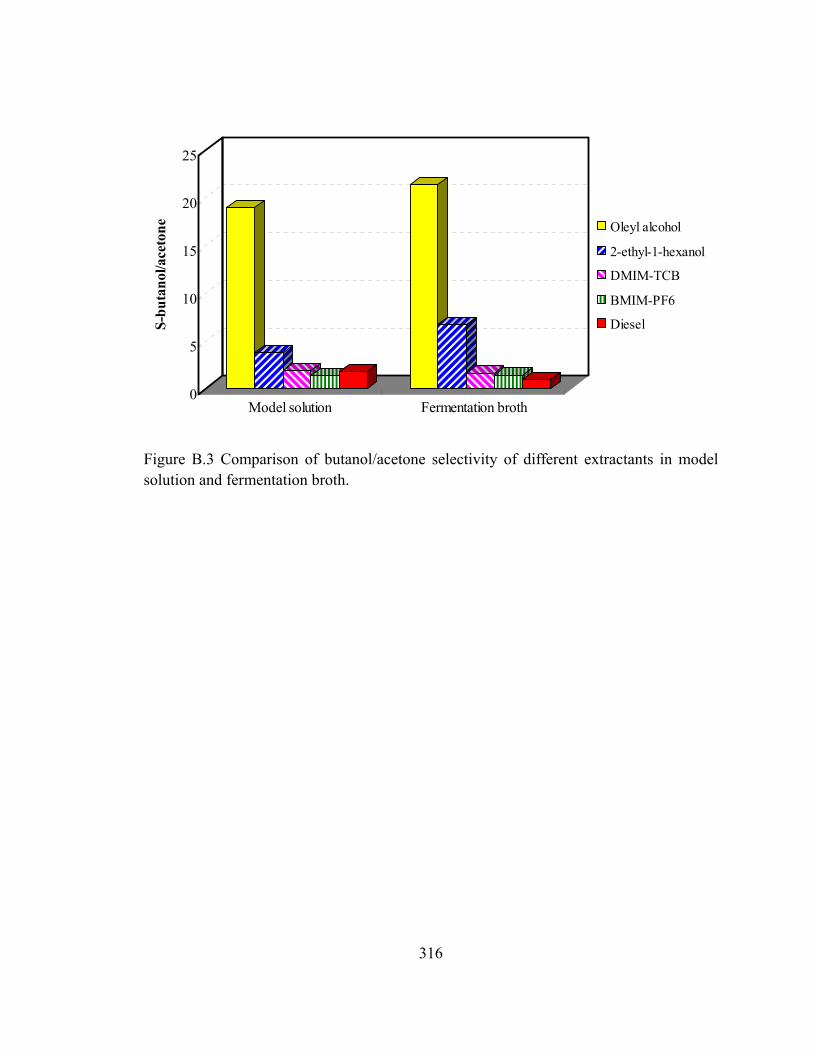
0
5
10
15
20
25S
-bu
tan
ol/a
ceto
ne
Model solution Fermentation broth
Oleyl alcohol
2-ethyl-1-hexanol
DMIM-TCB
BMIM-PF6
Diesel
Figure B.3 Comparison of butanol/acetone selectivity of different extractants in model solution and fermentation broth.
316

317
Appendix C: pSV6 Plasmid Construction and Transformation into
Clostridium beijerinckii CC101
C.1 PCR amplification of the truncated sol operon from C. beijerinckii CC101
The sol operon of C. beijerinckii consists of 4 genes ald, ctfA, ctfB and adc clustered
together in the genome. These genes are primarily responsible for the production of
butanol and shifting from the acidogenic phase to the solventogenic phase. adc gene was
not overexpressed in this study. Primers were designed to amplify the sol operon and clone
it into an overexpression vector containing the aad gene from C. acetobutylicum, or the
adhE2 gene from C. acetobutylicum, which are primarily the genes responsible for the
production of butanol. The truncated sol operon consisting of the ald, ctfA, and ctfB genes
was PCR amplified using designed primers with engineered Bam HI and BglII sites. A 2.9
kb amplicon was amplified and purified using the PCR purification kit.
C.2 Cloning of the t-SOL into pCR2.1 vector
The truncated sol operon was PCR amplified as described earlier and PCR purified.
This was then ligated to the pCR2.1 Vector (Invitrogen). The ligation mixture was

318
incubated overnight at 14 ° C and then transformed into chemically competent E. coli
DH5α cells. Putative transformants were selected on LA+ampicillin using a blue white
selection. Colonies were then transferred to liquid medium LB containing ampicillin at
100µg/ml and allowed to grow overnight. Plasmid DNA was isolated from the putative
clones and digested with HindIII and Sac II to check for the presence of the insert. A 3.0 kb
fragment was obtained upon cutting with these enzymes confirming the presence of the
t-SOL in pCR2.1 vector. The plasmid was designated as pCR2.1 t-SOL and was 6.9 kb in
size. The plasmid was preserved in glycerol stocks at – 80 oC until further use.
C.3 Cloning of the FRT-Hyg-FRT into pMTL-thl-adhE2
The plasmid pMTL-thl-adhE2 was constructed in the lab by another post-doc who
cloned the adhE2 gene from ATCC 824 under the control of the thl promoter. This plasmid
was used to overexpress the adhE2 gene in C. beijerinckii CC101. The pMTL-thl-adhE2
plasmid had the chloramphenicol resistant marker which cannot be used in C. beijerinckii
CC101 as it has a native catP gene which renders the antibiotic resistant to the drug. Thus
the FRT-HygB-FRT cassette was introduced into this plasmid vector so that it can be
successfully transformed into C. beijerinckii CC101 by conjugation. The pMTL-thl-adhE2
was constructed by removing the intron coding region and the ltrA gene from pMTL007
and replacing it with the adhE2 gene from C. acetobutylicum ATCC 824. The thiolase
promoter was cloned upstream to this gene to drive the expression of the ahdE2 gene.

319
The FRT-HYG-FRT cassette was obtained from the Pgem-FRT-HYG plasmid by
NcoI digestion and cloned upstream to the ahdE2 gene expressed under the control of the
thl promoter. The FRT-HYG-FRT cassette has its own promoter and it was cloned
downstream to the fac promoter in pMTL-thl-adhE2 plasmid. The pMTL-thl-adhE2
plasmid was linearized with NcoI and dephosphorylated. The FRT-HYG-FRT cassette
with NcoI engineered sites were then ligated to this vector.
The ligation mixture was incubated overnight at 16 oC and transformed into
chemically competent E. coli DH5α cells. The putative transformants were selected on
LA+ Hygromycin B (100µg/ml). The colonies obtained were again inoculated into liquid
medium and grown overnight at 37 oC and plasmid DNA isolated from them. The plasmid
was checked on the gel and confirmed by digestion with NcoI which gave the removeal of
the FRT-HYG-FRT cassette. The size of the vector was calculated to be 12.7kb. The
plasmid constructed was designated as pSV4 and preserved as glycerol stocks in -80 oC
deep freezer.
C.4 Cloning of the t-SOL(ald + ctfA + ctfB) into pSV4 vector
The truncated sol operon consisting of the ald + ctfA + ctfB gene was earlier cloned
into pCR2.1 vector with engineered HindIII sites. The truncated sol operon was obtained
from pCR2.1 t-SOL as a HindIII fragment. The plasmid pSV4 containing the adhE2 gene
under the control of thl promoter was also digested with HindIII and linearized. The vector

320
was dephosphorylated and the dephosphorylated vector was gel purified and ligated to
HindIII fragment of the truncated sol operon (3.0 kb).
The ligation mixture was then transformed into chemically competent E. Coli DH5α
cells and putative transformants plated on LA containing Hygromycin B (100µg/mL). Two
colonies were obtained on the LA+hygromycin after a few attempts. These were
inoculated into the same liquid medium LB+hygromycin B (100µg/mL) and the plasmid
isolated and confirmed by restriction digestion with HindIII. The plasmid was designated
as pSV6 and preserved in glycerol stocks at -80 oC. Figure C.3 illustrates the constructed
pSV6 plasmid.
C.5 Transformation of pSV6 plasmid into E. coli CAC434
The plasmid pSV6 overexpressing the adhE2 gene under the control of the thiolase
promoter and the ald, ctfA, and ctfB genes under the control of the fac promoter and
containing the FRT-Hyg-FRT marker cassette, was transformed into E. coli CAC434 by
electrotransformation as described earlier. The CAC434 cells were then selected on
LA+hygromycin B (100µg/mL) and grown in liquid medium containing the same
antibiotic. Plasmid pSV6 was isolated from CAC434 and confirmed on the gel and by
restriction digestion. This was then transformed into the host C beijerinckii CC101.

321
C.6 Conjugation of pSV6 harboring E. coli CAC434 cells into C. beijerinckii CC101
by filter mating
The donor strain E. coli CAC434 harboring the pSV6 plasmid was grown till it
reached an O.D.600 of 0.8 and then it was filtered through a low protein binding 0.2µ
Cellulose Acetate Filter membrane (Corning). The membrane was washed once to remove
traces of the medium and the antibiotic with 0.85% NaCl and then the recipient was filtered
through the same membrane. The donor to the recipient ratio was kept to 2:1. The filter was
then removed placed on a 0.5% RCM (Reinforced Clostridia Medium) agar plate and
allowed for filter mating in the anaerobic chamber for 14 h. The cells were then washed
with RCM recovery medium and plated on RCM + D cycloserine (250µg/mL) and
Hygromycin B (250µg/mL). Putative clones were obtained in 60 h and these were then
transferred and restreaked on RCM plates containing D cycloserine (250µg/mL) and
Hygromycin B (250µg/mL). The individual colonies were then picked up and inoculated
into RCM liquid medium containing the same antibiotics. The plasmid DNA was isolated
from C. beijerinckii CC101-SV6 which appeared as a smear on the gel. The plasmid yield
from Clostridium is not as good as that obtained from E. coli. Therefore the purified
plasmid was retransformed into E. coli DH 5 α and then confirmed by isolating the plasmid
from DH5 α and restriction digestion with Hind III which removed the sol operon from the
plasmid backbone. The mutant strain C. beijerinckii CC101-SV6 was preserved as
glycerol stock in -80 oC deep freezer.

Figure C.1 Truncated sol operon (2.9 kb) with ald, ctfA and ctfB genes.
322

Figure C.2 Constructed pMTL-thl-adhE2 plasmid.
323

Figure C.3 Constructed pSV6 plasmid.
324
![Actigomenroute [PDF, 2 blz, 3,41 MB] · 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 60 0 0 0 0 0 5 5 0 60 60 0 0 0 0 0 0 0 0 5 5 0 0 60 60 0 0 0 0 0 0 0 0 0 0](https://img.pdfslide.net/doc/110x75/60c8b2b62084147a4c1c9962/actigomenroute-pdf-2-blz-341-mb-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0.jpg)










![[XLS]mams.rmit.edu.aumams.rmit.edu.au/urs1erc4d2nv1.xlsx · Web view0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0. 0](https://img.pdfslide.net/doc/110x75/5ab434027f8b9a0f058b8cff/xlsmamsrmitedu-view0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0.jpg)












![[XLS] · Web view0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 7 2 0 0 0 0 0 0 0 0 0 0 0 5 4 0 0 0 0 0 0 0 0 0 0 0 5 4 0 0 0 0 0 0 0 0 0 0 0 5 4 0 0 0 0 0 0 0 0 0 0 0 5 4 0 0 0 0](https://img.pdfslide.net/doc/110x75/5aad015d7f8b9a8d678d9907/xls-view0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-7-2-0-0-0-0-0-0-0-0-0.jpg)


