APPLICABILITY OF SEQUENCE DIVERSITY AT MITOCHONDRIAL GENES ON
DIFFERENT TAXONOMIC LEVELS IN GENETICS OF SPECIATION, PHYLOGENETICS
AND BARCODING Yuri Ph. Kartavtsev A.V. Zhirmunsky Institute of
Marine Biology, Vladivostok 690041, Russia; e-mail:
[email protected]
[email protected]@hotmail.com Slide 2
Teacher: Academician, prof. Yuri P. Altukhov, 1992-2006 director,
N.Vavilov Institute of General Genetics, Moscow (Russia) Slide 3
MAIN GOALS 1. CBOL & Fish-BOL 2. SPECIES IDENTIFICATION 3.
SPECIES DEFINITION AND SPECIES ORIGIN: PROBLEMS, RESTRICTIONS.
GENETIC VIEW. Slide 4 1. CBOL & Fish-BOL Slide 5 THE
INTERNATIONAL CBOL PROJECT The CBOL is main global initiative. The
Fish-BOL, its part, has over 5400 species barcoded by Co-1 from
more than 30,000 specimens what makes it unique. P. Hebert and B.
Hanner are preparing a $150M grant application for Genome Canada
only for 2008. Other nations funds in CBOL are also big in some
countries and unions: USA, EU. B. Hanner suggests a recent Fish-BOL
paper on Canadian freshwater fishes for your interest, as well as a
new paper in press that demonstrates barcoding can identify cases
of market substitution in North American seafood. These might be
relevant for our meeting and ensuing discussions! In this year
there will be held third world-wide international conference (Sept.
2008 Chindao, Peoples Republic of China) and many regional meeting
like us were performed. Slide 6 THE INTERNATIONAL FISH-BOL PROJECT
Cochairmen: P. Hebert & B. Ward Slide 7 Fish-BOL CURRENT STATE
(2006 vs 2008) Class Barco- ded Species Prog- ress Class Barco- ded
Species Prog- ress Actinopterygii 3623 5073 27984 13% 18%
Cephalaspidom orphi 9 17 42 21% 40% Elasmobranchii 259 358 968 27%
37% Holocephali 15 37 41% Myxini 7878 70 10% 11% Sarcopterygii 0202
11 0% 18% Slide 8 Registration and Barcoding Utilities (BolD;
www.boldsystems.org) (1) w w.boldsystems.orgw w.boldsystems.org
Slide 9 Registration and Barcoding Utilities (BolD;
www.boldsystems.org) (2) w w.boldsystems.orgw w.boldsystems.org
Slide 10 Registration and Barcoding Utilities (BolD;
www.boldsystems.org) (3) w w.boldsystems.orgw w.boldsystems.org
Slide 11 Chair: Masaki MiyaMasaki Miya Vice Chair: Shunping
HeShunping He Members: Nina Bogutskaya Seinen Chow Shunping He Yuri
Kartavtsev Keiichi Matsuura Masaki Miya Mutsumi Nishida Ekaterina
Vasilieva North East Asian Regional Working Group Slide 12
FISH-BOL. RUSSIA DEVELOPMENT Slide 13 2. SPECIES IDENTIFICATION
Slide 14 Some Objects Fig. 1. Halibut-like flatfish, Hypoglossus
elasodon (A) and obscure flatfish, Pseudopleuronectes obcurus (). A
B Slide 15 INTRODUCTION Mitochondrion DNA (mtDNA) is a ring
molecule of 16-18 kilo-base pairs (kbp) in length. As literature
data show, mtDNA of all fishes has similar organization (Lee et
al., 2001; Kim et al., 2004; Kim et al., 2005; Nagase et al., 2005;
Nohara et al., 2005) and small differences among all vertebrate
animals, including men (Anderson et al., 1981; Bibb et al., 1981;
Wallace, 1992; Kogelnik et al., 2005). The complete content of
whole mitochondrial genome (mitogenome) includes: control region
(CR or D loop), where the site of initiation of replication and
promoters are located, big (16S) and small (12S) rRNA subunits, 22
tRNA and 13 polypeptide genes. Usually in phylogenetic research
single gene sequences are used for both mtDNA and nuclear genome.
However, recently more and more frequent are become complete
mitogenome usage. Japanese scientists are leading here for water
realm organisms. Most popular in phylogenetics are sequences of
cytochrome b (Cyt-b) and cytochrome oxidase 1 (C-1) genes, which
used for taxa comparison at the species - family level (Johns,
Avise, 1998; Hebert et al., 2004; Kartavtsev, Lee, 2006). Many
sequences that bringing the phylogenetic signal obtained for
different taxa at gene 16S rRNA as well. Sequences of separate
genes can dive different phylogenetic signal because of differences
in substitution rates. This is also true for different sections of
genes. Also, under comparison of higher taxa there may be effects
of homoplasy. When numerous taxa available there are problems of
insufficient information capacity of sequences to cover big species
diversity and representative taxa representation is quite important
(Hilish et al., 1996). Nevertheless, for the species
identification, excluding rare cases, fine results are available
even with the usage of short sequences, like -1, with 650 bp. Slide
16 Spacers [ITS-1, 2] mtDNA mtDNA nDNA, nDNA, rDNA rDNA Most
substantiated statistically results Most substantiated
statistically results Statistically significant results
Statistically significant results Species G Genus Family Order C
Class P Phylum Applicability of Different DNA Types in
Phylogenetics and Taxonomy Slide 17 MATERIAL AND METHODS 2. PCR
Amplification Genes DNA 3. Sequence Analysis. In 2006 : 200 seq. 4.
Mol. Phylogenetics & Taxonomy 1. DNA Isolation. Slide 18
Aligned flatfish sequences at -1: our and GenBank data Slide 19
Distance Data Slide 20 p-DISTANCES IN GROUPS OF COMPARISON, Catfish
Fig. 1. Fig. 1. Resulting graph of mean p-distance values at four
levels of differentiation in the catfish species (Siluriformes) for
Cyt-b gene. Groups: 1. Intraspecies, among individuals of the same
species; 2. Intragenus, among species of the same genera; 3.
Intrafamily, among genera of the same family; 4. Intraorder,
families of the order Pleuronectiformes. There are statistically
significant variation. SE: a standard error of mean; F = 124.74,
d.f. = 3; 29, p < 0.0001 (Kartavtsev et al., 2007a, Gene). Slide
21 Fig. 2. Resulting graph of one factor ANOVA and mean p-distance
values at four levels of differentiation in the flatfish species
(Pleuronectiformes) for Cyt-b gene. Groups: 1. Intraspecies, among
individuals of the same species; 2. Intragenus, among species of
the same genera; 3. Intrafamily, among genera of the same family;
4. Intraorder, families of the order Pleuronectiformes.
Statistically significant variation are shown on top of the graph.
SE: a standard error of mean (Kartavtsev et al., 2007b, Marine
Biol.). p-DISTANCES IN GROUPS OF COMPARISON, flatfish Slide 22
p-DISTANCES IN GROUPS OF COMPARISON, turtles Fig. 3. Resulting
graph of ANOVA and mean p-distance values at four levels of
differentiation in turtle species (Testudines) for Cyt-b gene.
Groups: 1. Intraspecies, among individuals of the same species; 2.
Intragenus, among species of the same genera; 3. Intrafamily, among
genera of the same family; 4. Intraorder, families of the same
order. Variation among four groups is statistically significant: F
= 61.87, d.f. = 3; 152, p < 0.000001 (Jung et al., 2006).
-Distances: (1) 2.330.03%, (2) 3.340.48%, (3) 6.410.11% (4)
11.920.37% (Mean SE). Slide 23 p-DISTANCES IN GROUPS OF COMPARISON,
Perciformes Fig. 3. Resulting graph of one factor ANOVA and mean
p-distance values at four levels of differentiation in fish species
(Perciformes) for Co-1 gene sequence data. Comparison groups: 1.
Intraspecies, among individuals of the same species; 2. Intragenus,
among species of the same genera; 3. Intrafamily, among genera of
the same family; 4. Intraorder, families of the order Perciformes.
Variation is statistically significant. Bars are confidence
intervals for mean (95%). Slide 24 Fig. 4. Categorized plot of
distribution of weighted mean p-distances among four groups of
comparison at Cyt-b and Co-1 genes. Groups here: 1. Intra-species,
among individuals of the same species; 2. Intra-sibling species, 3.
Intra-genus, among species of the same genera; 4. Intra-family,
among genera of the same family (Kartavtsev, Lee, 2006).
p-DISTANCES IN GROUPS OF COMPARISON, Review Slide 25 GENETIC
SIMILARITY IN TAXA OF DIFFERENT RANK: MEAN FOR THE GROUPS Fig. 5.
Genetic similarity in taxa of different rank based on protein
markers: mean for the groups. 1. Subspecies, 2. Semispecies and
sibling species, 3. Species, 4. Genera. Intraspecies genetic
distances were measured for many groups of organisms (Lewontin,
1974, Nei, 1987, Altukhov, 1989). Mean genetic similarity on this
level is near I = 0.95 (see details in Kartavtsev, 2005). mtDNA
data were presented above. Thus, data available suggest that in
general a phyletic evolution prevail in animal world, and so far,
the Geographic speciation events (Type 1a) prevail in nature. Do
data presented assume that speciation is always follows the Type 1a
mode? I guess, no. Few examples below let to support this answer.
Slide 26 DISTANCE VS TAXA SPLITTING Has punctuation an impact in
species origin on molecular level? Avise, Ayala, 1976; Kartavtsev
et al., 1980; current No. Pegel et al., 2006 Yes. Number of
Splittings Transformed p-distance r s = 0.22, p < 0.05 Fig. 6.
Plot of p-distance on number of splittings at Cyt-b sequence data
for catfishes and flatfishes. Slide 27 GENETIC DISTANCES AMONG
SPECIES IN SEPARATE ANIMAL GENERA (After Avise, Aquadro, 1982) This
plot illustrate a thought that different animal groups of the same
rank are unequal in structural gene divergence; i.e. the rate of
evolution differ either at genes or at morphology or both. Slide 28
GENETIC DISTATNCES IN TAXA OF SALMON FISHES 1 Populations within
species, 2 Subspecies, 3 - Species D This plot support a thought
that in salmon even a very small structural gene change can create
separate biological species. Slide 29 Comparison of chars Table
2.1. COMPARISON OF ISOZYME ACTIVITY IN THREE WHITEFISH FORMS
(COREGONIDAE) AND GRAYLING (THYMALLIDAE) LOCUS/ FORM LEVELS OF
DIFFERENCES IN ACTIVITY Note. Total number of loci analyzed are:
Whitefish 22, Grayling 23, - Activity do not differ significantly,
+ Iterative activity difference, ++ two-fold difference, +++
three-fold or greater difference Ratio, % EXAMPLES OF REGULATORY
DIVERGENCE IN FISH TAXA Slide 30 WHAT IS MAIN OUTCOME Distance
measure alone is not satisfactory descriptor. Data on intraspecies
diversity (heterozygosity) at structural genes are necessary.
Measures of regulatory genome changes should be necessary to
describe transformative modes of speciation. Other descriptors of
genomic change are required (e.g. chromoseme number, NF, etc.).
Slide 31 3. SPECIES DEFINITION AND SPECIES ORIGIN: PROBLEMS,
RESTRICTIONS. GENETIC VIEW Slide 32 WHAT SPECIES IS? Species is a
biological unity which reproductively isolated from other unities
and consisting from one to several more or less stable populations
of sexually reproducing individuals that occupy certain area in
nature (my definition). In principal points, this is the definition
of BSC (Biological Species Concept). In one of the original BSC
definitions A species is a reproductive community of populations
(reproductively isolated from others) that occupies a specific
niche in nature (Mayr, 1982, p. 273). We will accept BSC for
further discussion, although will keep in mind that it is
restricted mainly to bisexual organisms (Mayr, 1963,
Timofeev-Resovsky et al., 1977, Templeton, 1998). The Linnaean
Species The Biological Species Concept (BSC) (Mayr, 1942, 1963) BSC
Modification II (Mayr, 1982) The Recognition Species Concept
(Paterson, 1978, 1985) The Cohesion Species Concept (Templeton,
1989) Evolutionary Species Concept Simpson (1961) Evolutionary
Species Concept. Wileys (1978) Evolutionary Species Concept. The
Ecological Species Concept (Van Vallen, 1976). The Phylogenetic
Species Concept (Crawcraft, 1983). Slide 33 GENERAL GENETIC
APPROACH: ADVANCES AND LIMITATIONS The problem of biological
species, and speciation are main focus of this report. These
problems took researchers attention since establishing the biology
as a science. Most popular now among biologist is the Synthetic
Theory of Evolution (STE), which part is comprised by the
Biological Species Concept (BSC). Origin and systematic description
of STE concept was presented in fundamental books by Haldane
(1932), Dobzhansky (1937, 1943, 1951), Huxley (1954), Mayr and
co-workers (Mayr, 1942, Mayr, 1963). A popular in Russia summary of
STE became a book by Timofeev-Resovsky with co-authors (1977).
Good, constructive ideas in STE support were developed by Vorontsov
(1980). One of weak point in STE is absence as a rule a possibility
to prove experimentally one of key criteria of BSC i.e.,
reproductive isolation of the species in nature. There are a lot of
other criticisms that were summarized for example by King (1993).
Nowadays, the new controversy between BSC and Phylogenetic Species
Concept arise (Avise, Wollenberg, 1997). The theory of speciation
is also not well developed in STE. Exactly speaking, in a
quantitative meaning there is no theory as real matter at all. In
should be outlined, nevertheless, that many directions of STE and
genetics of speciation are developing. Thus, a diverse analysis
performed to understand a genetic sense and conceptual basements of
speciation (Fox, Morrow, 1981, Grant, 1984, King, 1992). The
genetic basis for creation of a reproductive isolation was
subjected to the analysis too (Leslie, 1982, Templeton, 1981, Nei
et al., 1983, Coyne, 1992). As well there were considered: a
possibility of a sympatric speciation (Bush, 1975, Genermon, 1991),
the role of saltations or revolutions in evolution (Altukhov,
Rychkov, 1970, Carson, 1974, Altukhov, 1985, 1997) and the genetic
differentiation during formation of living forms and taxa (Avise,
1975, Avise, Aquadro, 1982, Nevo, Cleve, 1978, Thorpe, 1982, Nei,
1975, 1987). What in general are the advances and limitations in
contemporary genetic approach? Slide 34 ADVANCES 1. Data reduction
up to genotypic codes (values) give us a possibility to use genetic
theory in the analysis. 2. It is possible to perform a comparative
investigation of a variability among structural and regulatory
elements of the genome and genetic divergence of taxa. 3.
Investigations on protein and nucleotide divergence of species from
nature discovered a Molecular clock. 4. A possibility of
phylogenetic reconstruction occurred: 1) not by similarity, but by
kinship and 2) by in time dating of a divergence. Slide 35
LIMITATIONS 1. Deduction is limited by genotypic descriptions and
genetic theory. 2. Analysis is connected with preliminary laborious
experimental investigation (with its own limitations). 3.
Investigation of a species from nature is frequently limited by
originality or rare repetition of an event (phenomenon). 4.
Genotypic effects of the marker loci on phenotype are weak. 5. The
theory is not sufficiently developed in some directions. Slide 36
WHAT DATA ARE NECESSARY? Data that support (reject) central dogma
of Neodarwinism Evolution can occurred the only on the base of
genetic change. Data on variability at different levels of
biological organization in genetic terms (by loci quantitative
genotypic values AA, ) single-dimensioned data tables (DT). Data on
genotypic values of an individual at the set of loci (genotype AA
Bb) or whole gene sequence set multi-dimensional DT. Complementary
data: Morphology traits, data on abiotic variability etc. (at least
as an expert estimate grouping variables). Slide 37 SCHEMATIC
REPRESENTATION OF SPECIES DIVERGENCE AND ORIGIN (From Dobzhansky,
1955) Fig. 3.1. Dobzhanskys (1955) scheme of in time divergence.
Single species population. B Initial phase of divergence
(subspecies). C Different species. A B C The keystone of STE
(Synthetic Theory of Evolution) may be represented by Dobzhanskys
scheme (Fig. 3.1), in which the gene pool separation is a key to
speciation. If one provides a fact that evolution is possible
without genetic change in lineages, then the evolutionary genetic
paradigm and STE in particular can be rejected. Slide 38 Fig. 3.1.
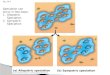
Main Modes of Speciation Bush, 1975) FIG. 3.2. DIAGRAMMATIC
REPRESENTATION OF BASIC MODES OF SPECIATION (From Bush, 1975) The
gene flow breaks are able to create Reproductive Isolating Barriers
(RIB) or Reproductive Isolation Mechanisms (RIM), which in their
turn lead to further origin of species; under different situation
in nature, the different modes of speciation acted (Fig. 3.2).
Neither, the scheme above, nor the paper itself (Bush, 1975),
answer many fundamental questions of speciation. For instance, it
is unclear, what mode is most frequent and is a gene flow the sole
primary factor, that alter gene pools or there are others? In other
words we have to conclude that there is no a theory of speciation
in scientific meaning at all. Slide 39 SPECIATION MODES (SM):
POPULATION GENETIC VIEW ABSENCE OF QUANTITATIVE THEORY OF
SPECIATION (QTS) We have mentioned in preceding section that the
speciation theory in evolutionary genetics is absent in exact
scientific meaning, which expects the ability to predict future by
the theory. In this case this is to predict species origin, or at
least discriminate among several speciation modes on the basis of
some quantitative parameters or their empirical estimates. Attempts
made in this direction (Avise, Wollenberg, 1997, Templeton, 1998)
do not fit the above criteria. That is why we attempted to step in
the discrimination of the speciation modes on the basis of main
population genetic measurements available in literature, and that
may be laid in the frame of a genetic speciation concept. BASEMENT
FOR THE QTS As a basis for the set of evolutionary genetic concepts
we used the descriptions made by Templeton (1981). As a result the
classification scheme for 7 different modes of speciation was
created (Fig. 3.3). This approach leads to quite simple
experimental scheme that permits: (i) to arrange further
investigation of speciation in different groups of organisms, and
(ii) to derive analytical relations for each speciation mode (Fig.
3.4). The approach is based on a set theory but it is a
knowledge-based approach. I believe, this approach is best for such
complicated matter. Slide 40 Fig. 3.3. SPECIATION MODES (SM):
POPULATION GENETIC VIEW (Kartavtsev et al, 2002) DIVERGENCE SM D1.
ADAPTIVED2. CLINALD3. HABITAT Necessary Conditions for Speciation
D1. a) Erection of extrinsic Isolating barriers followed by gene
flow break; b) Pleotropic origin of RIB (Reproductive Isolatiion
Barriers) in long time D2. a) Selection on a cline with isolation
by distance; b) Pleotropic origin of RIB D3. a) Selection over
multiple habitats with no isolation by distance; b) RIB origin by
disruptive selection at genes determined behavior Sufficient
Conditions for Speciation Lack of efficient hybridi- zation in the
zone of contact DESCRIPTORS: D Genetic distance at structural
genes: D T in suggested parent taxa, D S among conspecific demes, D
D among subspecies or sibling species; H D Mean heterozygosity in
suggested daughter population; H p Mean heterozygosity in suggested
parent population; E P Divergence in regulatory genes among
suggested parent taxa; E D Divergence in regulatory genes among
suggested daughter taxa; TM + - Test for modification (positive);
TM - - Test for modification (negative). RIB Reproductive isolation
Barriers. Lack of efficient hybridi- zation outside the zone of
contact Lack of efficient hybridi- zation inside and outside the
zone of contact 1. D T > D S 1 (S) 2. E D = E P 3. H D = H P 4.
TM - 1. D T > D S 2 (S) 2. E D E P 3. H D = H P 4. TM - 1. D T =
D S 3 (S) 2. E D E P 3. H D =< H P 4. TM - Experimentally
measurable features and possible descriptors for the model
(theory), (S) Slide 41 Fig. 3.4. ANALITICAL DESCRIPTION OF SEVEN
TYPES OF SPECIATION MODES 1 (S) {(D T > D S ) (E D = E P ) (H D
= H P ) TM - } (D1) 2 (S) {(D T = D S ) (E D E P ) (H D = H P ) TM
- } (D2) 3 (S) {(D T = D S ) (E D E P ) (H DD D ) (E D E P ) (H D
< H P ) TM - } (T1) 5 (S) {(D T = D D ) (E D = E P ) (H D < H
P ) TM - } (T2) 6 (S) {(D T > D D ) (E D E P ) (H D > H P )
TM - } (T3) 7 (S) {(D T > D S ) (E D E P ) (H D < H P ) TM -
} (T4) Note. Descriptors are explained in previous figure. Slide 42
THE QTSEMPIRICAL PROVING THE QTS EMPIRICAL PROVING Salmon
(Kartavtsev, Mamontov, 1983, Kartavtsev et al., 1983),
Cypriniformes (Kartavtsev et al., 2002), Turtles, flatfishes,
catfishes (Jung et al., 2006, Kartavtsev et al., 2006, 2007). Slide
43 PHYLOGENETICS & BARCODING Slide 44 SPECIES IDENTIFICATION
AND PHYLOGENETICS Identification + Taxonomy Phylogenetics +
Taxonomy Slide 45 Fig. 3.5. Rooted consensus (50%) trees (A-B)
showing phylogenetic interrelationships on the basis of Cyt-b
sequence data for the analyzed flatfish species (Pleuronectiformes)
and four out-group taxa. A tree based on NJ clustering technique
with bootstrap support shown in the nodes (n=1000), B Bayesian
tree; repetition frequencies for n=106 simulated generations are
shown (%) in the nodes. The tree was built based on the TrN+I+G
model and was rooted with the sequences of four out- group species:
three are Perciformes and one is Cypriniformes. The scales in the
left bottom corners indicate relative branch lengths. Slide 46 Fig.
3.6. Consensus (50%) tree showing phylogenetic interrelationships
on the basis of Co-1 sequence data for the analyzed flatfish
species (Pleuronectiformes) and two outgroup taxa. Rooted Bayesian
tree; repetition frequencies (probabilities) for n=10 6 simulated
generations are shown in the nodes (%).The tree was built based on
the TVM+I+G model and rooted with the sequences of two outgroup
species, Perciformes. The scale in the left bottom corners indicate
the relative branch lengths. Slide 47 Fig. 3.7. Rooted consensus
(50%) tree showing phylogenetic interrelationships on the basis of
Cyt-b sequence data for the analyzed flatfish species
(Pleuronectiformes) and three outgroup taxa. Bayesian tree;
repetition frequencies for n=10 6 simulated generations are shown
(%) in the nodes. The trees were built based on the TrN+I+G model,
and rooted with the sequences of outgroup species Perciformes. The
scales in the left bottom corners indicate the relative branch
lengths. (Kartavtsev et al., 2007, Marine Biol.). Slide 48 Fig.
3.8. Consensus (50%) trees showing phylogenetic interrelationships
on the basis of Co-1 sequence data for 7 analyzed perch- like fish
species (Perciformes) and two outgroup sequences. Rooted Bayesian
tree was build for sample purposes; posterior probabilities for
n=10 6 simulated generations are shown in the nodes (%). The tree
was built based on the HKY+G model. Two other numbers in the nodes
show tree bootstrap support based on similar clustering for NJ and
ML techniques; support scores are given in the order NJ/ML/BA.
Outgroup are two sequences of a representative of Cypriniformes.
The scale in the left bottom corner indicate the relative branch
lengths. Slide 49 Conclusions Speciation mode must be specified
with a set of descriptors not exclusively by distances Both Co-1
and Cyt-b are generally good barcoding tools for species
identification For phylogenetic reconstructions we need to cover
both taxa diversity and several genes sequence diversity Slide 50
THANKS FOR ATTENTION! ! Slide 51 FEW RECENT PUBLICATIONS Kartavtsev
YP. 2005. Molecular evolution and population genetics. Far Eastern
State Univ. Press., Vladivostok, 234 p. Kartavtsev YP, Lee J-S.
Analysis of nucleotide diversity at genes Cyt-b and Co-1 on
population, species, and genera levels. Applicability of DNA and
allozyme data in the genetics of speciation. Genetika, 2006. 42:
437-461. Jung S-O, Lee Y-M, Kartavtsev YP, Park I-S, Kim D-S, Lee
J-S. The complete mitochondrial genome of the Korean soft-shelled
turtle Pelodiscus sinensis // DNA Sequence, 2006. 17(6): 471-483.
Sasaki T, Kartavtsev YP, Uematsu T, Sviridov VV, Hanzawa N.
Phylogenetic independence of Far Eastern Leuciscinae (Pisces:
Cyprinidae) inferred from mitochondrial DNA analysis. Gene and
Genetic Systems, 2007. 82: 329-340. Kartavtsev YP, Lee Y-M, Jung
S-O, Byeon H-K, Son Y-, Lee J-S. Complete mitochondrial genome in
the bullhead torrent catfish, Liobagrus obesus (Siluriformes,
Amblycipididae) and phylogenetic considerations. Gene, 2007a. 396:
13-27. Kartavtsev YP, Park T-J, Vinnikov KA, Ivankov VN, Sharina
SN, Lee J-S. Cytochrome b (Cyt-b) gene sequences analysis in six
flatfish species (Pisces, Pleuronectidae) with phylogenetic and
taxonomic insights. Journal Marine Biology, 2007b. 152(4): 757-773.
Slide 52 Terminal taxa: A B C D E F G H Outgroup: TERMSRoot Nodes,
Speciation events Internal nodes Branches Ingroups: group Sister
group Slide 53 A B C D E A C E B D A E C B D Slide 54 . Slide 55 -
. , . Slide 56 A B C A A B B C C a bca b c : Slide 57 - - 4N - -
Slide 58 1. .. (, -. , . , ) 2. .. (, ..., . ) 3. .. (, ...) 4. ..
(, ...) 5. .. (, ...) 6. .. (, -. , . ) 6. .. (, ...) 7. A.A. (,
...) 1. .. (, ...) 2. .. (, ...) 3. .. (, ) 4. .. (, ..., . ) 5. ..
(, ...) 6. .. (, ..., . ) 7. .. (, ) 8. O.. (, ) Slide 59 (4) ..
(...., , . ) .. (...) .. (...) .. (..., . ) .. (...) (4) .. (...)
.. (...) .. () .. ( ) (3) .. (-. , ) .. (...) .. () -, , . (7) ..
(..., . ) .. (..., . ) .. (...) .. (...) .. (..., . ) .. (..., . )
.. (...) Slide 60 GenBank (NCBI) Slide 61 () , 16-18 (.). , (Lee et
al., 2001; Kim et al., 2004; Kim et al., 2005; Nagase et al., 2005;
Nohara et al., 2005) , , (Anderson et al., 1981; Bibb et al., 1981;
Wallace, 1992; Kogelnik et al., 2005). () : (CR D ), , (16S) (12S)
, 22 13 . , , . b (Cyt-b) 1 (C-1), (Johns, Avise, 1998; Hebert et
al., 2004; , , 2006). , , 16S . - / ( ). . , , , (Hilish et al.,
1996, Miya et al., 2001). , , , , 1 (-1, 654 ).


![[MP] 02 - Phylogenetics - biologia.campusnet.unito.it · Molecular Phylogenetics Basis of Molecular Phylogenies Overview ¾Phylogenetics Definitions ¾Genetic Variation and Evolution](https://img.pdfslide.net/doc/110x75/5c6216d809d3f238158b4601/mp-02-phylogenetics-molecular-phylogenetics-basis-of-molecular-phylogenies.jpg)