Embed Size (px)
Citation preview

Supplementary Materials forCRISPR/Cas9-coupled recombineering for metabolic engineering of Corynebacterium
glutamicum
Jae Sung Choa,b, Kyeong Rok Choia,b, Cindy Pricilia Surya Prabowoa, Jae Ho Shina, Dongsoo
Yanga, Jaedong Janga & Sang Yup Leea,c
aMetabolic and Biomolecular Engineering National Research Laboratory, Department of
Chemical and Biomolecular Engineering (BK21 Plus Program), BioProcess Engineering
Research Center, BioInformatics Research Center, and Institute for the BioCentury, Korea
Advanced Institute of Science and Technology, 291, Daehak-ro, Yuseong-gu, Daejeon,
34141, Republic of Korea.
bThese authors contributed equally to this work.
cCorrespondence should be addressed to S.Y.L. ([email protected])
Contents Page
Supplementary Notes 2 – 3
Supplementary Figures 4 – 7
Supplementary Tables 8 – 13
References 14 – 14
1

Note S1. Cloning of endogenous recT gene in E. coli
Cloning of recT gene from E. coli K-12 MG1655 into pTacCC1 vector was first conducted in E. coli DH5ɑ which also harbors the same gene in its chromosome. During the selection of transformants harboring successfully cloned plasmids, PCR analysis indicated that most of the colonies formed carry the desired pTacCC1-recT plasmids. Interestingly, however, sequencing analysis on the plasmids prepared from the colonies read no recT gene cloned, indicating spontaneous loss of the gene while growing the colonies in liquid media. The loss of recT gene was further demonstrated by second round PCR analysis for the streaked colonies grown on master plates. It should be noted that the absence of lacI gene in pTacCC1 leads to uncontrolled expression of the recombinase RecT from its 0.8-kb gene. Ironically, the expressed RecT may have facilitated recombination of chromosomal and plasmid-borne recT genes, damaging the recT genes in the plasmids. As a response to this speculation, we changed the cloning host to E. coli XL1-Blue, which harbors chromosomal lacIQ (Table S1), to minimize the expression of RecT from the successfully cloned pTacCC1-recT. Additional supplementation of ~80 g/L glucose to prevent catabolite activator protein (CAP) from binding to Ptac in the plasmid and enhancing RecT expression enabled successful cloning and maintenance of pTacCC1-recT in XL1-Blue.
The same strategy, however, did not work for cloning the His:RecT vector pTacCC1-HrT. The loss of hexahistidine-tagged recT gene while growing the XL1-Blue transformants was consistently observed even with the supplementation of 80 g/L glucose. It has been repeatedly reported and also observed in our lab that N-terminal fusion of hexahistidine tag improves the expression of proteins (Shin et al., 2016), most likely due to increased translation efficiency. Accordingly, the leaky expression of RecT from XL1-Blue harboring pTacCC1-HrT with glucose supplemented might be high enough to induce recombination of the recT gene in the plasmid, facilitating the loss of the gene from the plasmid. With an observation that successfully cloned pTacCC1-HrT is detectable in most of the colonies formed immediately after transformation, we conceived a strategy of retrieving plasmids directly from the initial colonies formed. Our hypothesis was that successfully cloned pTacCC1-HrT would be among the heterogeneous plasmid population and direct introduction of the prepared plasmid to C. glutamicum will generate colonies with intact pTacCC1-HrT. Proving the hypothesis, C. glutamicum strains with intact pTacCC1-HrT was successfully selected upon introducing the heterogeneous population of plasmid isolated from the harvested colonies.
Note S2. Expression of Cas9 with different codon usage
In our initial efforts to construct plasmid pEKEx1-Cas9, a backbone vector harboring the S. pyogenes cas9 gene, we observed that the plasmid did not result in any transformant upon electroporation of C. glutamicum with pEKEx1-Cas9. It was only when we had cloned the cas9 gene originally codon-optimized for atinomycetal genomes(Tong et al., 2015) that we were able to obtain transformants expressing the Cas9 protein upon IPTG induction in RG media (Fig. S2b). It has been well reported and also observed in our group that the expression of Cas9 at high level retards bacterial cell growth most likely due to non-specific binding of the uncharged Cas9 to the chromosomal DNAs of host organisms. Moreover, a plasmid
2

constitutively expressing Cas9 in the cloning host has rarely been cloned. These reports and observations suggest the cas9 gene with native codon may results in a stronger expression than the codon-optimized cas9 gene in C. glutamicum, resulting in leaky expression of the native cas9 gene at significant levels upon the transformation and culture of the transformants in RG media.
Although direct demonstration of this hypothesis is difficult since viable C. glutamicum colonies harboring pEKEx1-Cas9 do not form, the hypothesis is further supported by another observation: a modified version of pEKEx1-Cas9 (pEKEx1-Cas9spacer) which has a long spacer (49 bp) between its ribosome binding site (RBS) and the first start codon – a feature that generally reduces translation initiation efficiency of a gene – can successfully be introduced to C. glutamicum, although the resulting transformants do not express Cas9 at a level observable on SDS-PAGE (data not shown). If the hypothesis is true, it is a good counterexample to a general notion that a gene with GC content similar to that of the host chromosome has higher translation efficiency – note that the GC content in the codon-optimized cas9 gene is 61.5% as opposed to 35.1% of the native S. pyogenes Cas9 gene while C. glutamicum genome has a GC content of 53.8%.
3

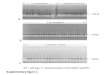
Fig. S1
Adapting electroporation conditions for C. glutamicum transformation for CRISPR/Cas9-coupled recombineering.
Electroporation protocol for C. glutamicum was adapted to improve transformation efficiency by exploiting different media for cell culture (a), distances between electrodes in electroporation cuvettes (b), resistance during electroporation (c), recovery time after electroporation (d) and presence or absence of pTacCC1-HrT (e). Default parameters are NCM media, 1-mm gap electroporation cuvettes and resistance of 200 Ω with two hours of recovery. n = 3, NS, not significant, **P < 0.01, determined by two-tailed Student t-test. Error bars represent standard deviations.
4

Fig. S2
Expression test for RecT and Cas9 in C. glutamicum.
(a) Expression of RecT (~30 kDa) and hexahistidine-tagged RecT (~31 kDa) from C. glutamicum ATCC 13032 harboring either pTacCC1-recT or pTacCC1-HrT. No IPTG was supplemented. Red arrows (►) indicate overexpressed protein bands. φ, control strain with pTacCC1. (b) Expression of hexahistidine-tagged Cas9 (~160 kDa) from C. glutamicum ATCC 13032 pEKEx1-Cas9opt in BHI and RG media. IPTG was supplemented at different concentrations (0 – 2 mM). Note that C. glutamicum strains applied for CRISPR/Cas9-coupled recombineering are recovered in BHI media after electroporation with no IPTG supplemented. Red arrows (◄) indicate overexpressed protein bands.
5
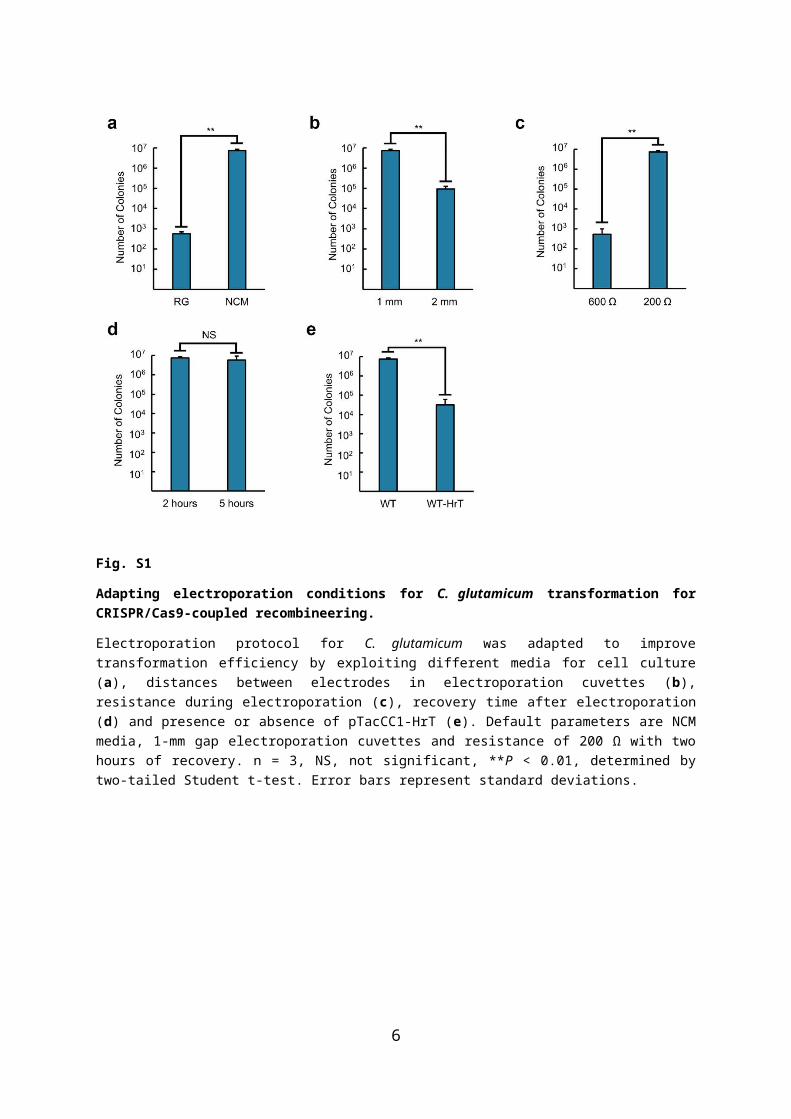
Fig. S3
Serial transformation protocol for CRISPR/Cas9-coupled recombineering.
(a) Schematic overview of the serial transformation protocol. Cas9-sgRNA vector (pCG9-argR1) and ssODN (ssODNargR_400) targeting 400-bp deletion of argR are introduced to the cells. The transformed cells are immediately used for the preparation of competent cells. The competent cells are re-transformed with the same Cas9-sgRNA vector and ssODN and the cycle is repeated indefinitely to enhance the mutant population until positive mutants are found.
6
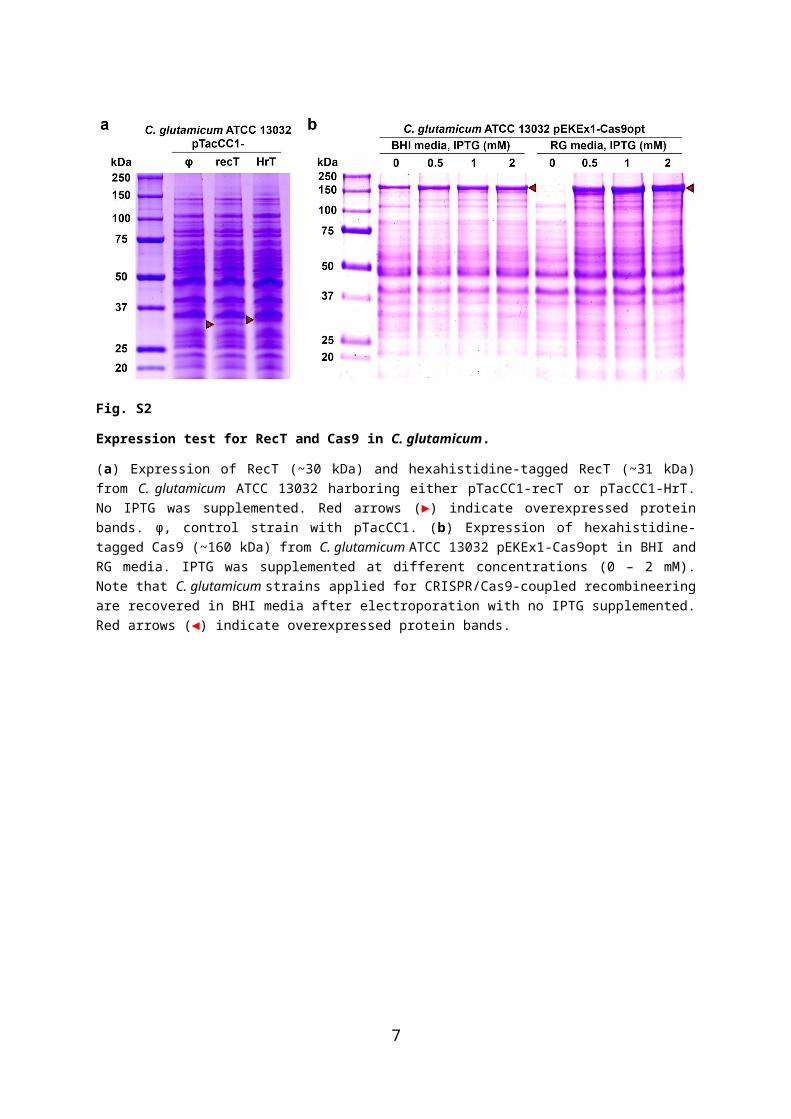
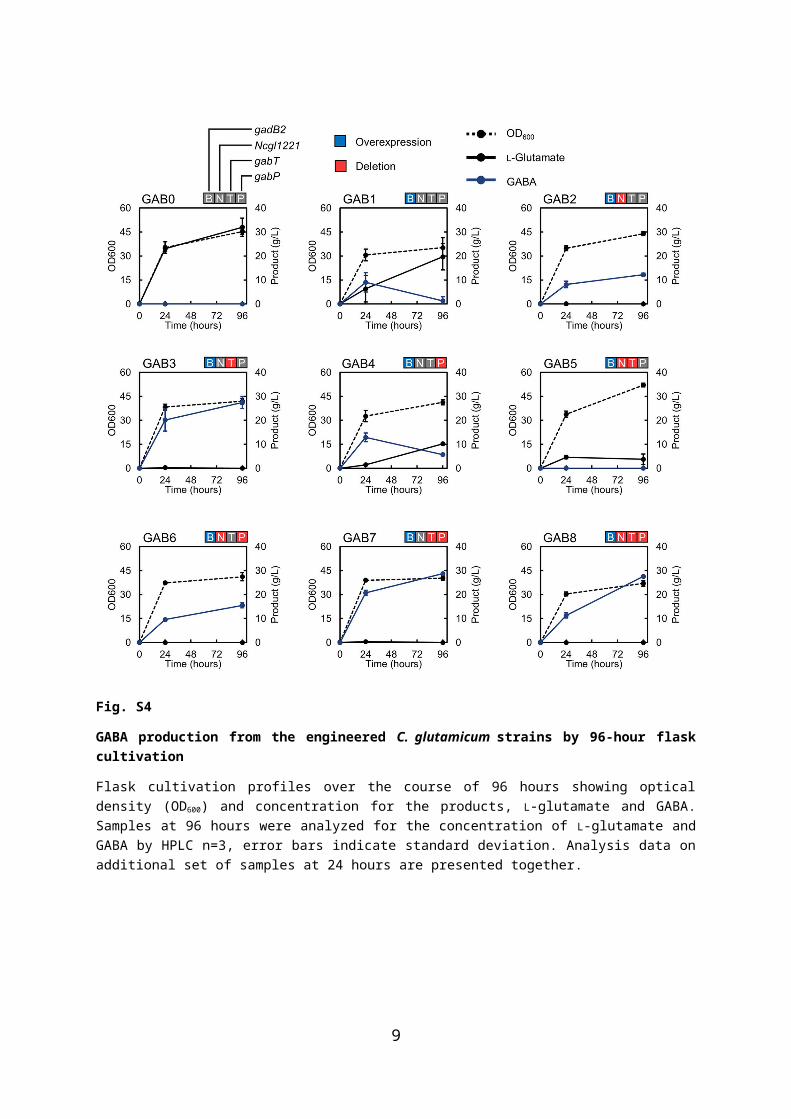
Fig. S4
GABA production from the engineered C. glutamicum strains by 96-hour flask cultivation
Flask cultivation profiles over the course of 96 hours showing optical density (OD 600) and concentration for the products, L-glutamate and GABA. Samples at 96 hours were analyzed for the concentration of L-glutamate and GABA by HPLC n=3, error bars indicate standard deviation. Analysis data on additional set of samples at 24 hours are presented together.
7
Table S1
Strains and plasmids used in this work
Strain/Plasmid Relevant Characteristicsa Source
Escherichia coli strains
DH5α E. coli K-12 F- φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17(rk⁻
mk+) phoA supE44 thi-1gyrA96 relA1 λ- Invitrogen
XL1-Blue E. coli endA1 gyrA96(nalR) thi-1 recA1 relA1 lac glnV44 F'[::Tn10 proAB+ lacIQ lacZΔM15] hsdR17 (rk
⁻ mk+) Stratagene
MG1655 E. coli K-12 F- λ- ilvG– rfb-50 rph-1 Bachmann (1996)
BL21(DE3) E. coli str. B F– ompT gal dcm lon hsdSB(rB–mB
–) λ(DE3 [lacI lacUV5-T7p07 ind1 sam7 nin5]) [malB+]K-12(λS)
Studier and Moffatt (1986)
Corynebacterium glutamicum strains
WT C. glutamicum ATCC 13032 Kinoshita et al. (1958)
WT-HrT WT strain harboring pTacCC1-HrT This study
ARG1 WT-HrT ΔargR harboring pCG9-argR1 This study
ARG2 WT-HrT ΔargR harboring pCG9ts-argR2 This study
WHC2 WT-HrT ΔNcgl1221 harboring pCG9ts-Ncgl1221 This study
WHC3 WT-HrT ΔgabT harboring pCG9ts-gabT This study
WHC4 WT-HrT ΔgabP harboring pCG9ts-gabP This study
WHC5 WT-HrT ΔNcgl1221 ΔgabT harboring pCG9ts-gabT This study
WHC6 WT-HrT ΔNcgl1221 ΔgabP harboring pCG9ts-gabP This study
WHC7 WT-HrT ΔgabT ΔgabP harboring pCG9ts-gabP This study
WHC8 WT-HrT ΔNcgl1221 ΔgabT ΔgabP harboring pCG9ts-gabT This study
WH2 WT-HrT ΔNcgl1221 This study
WH3 WT-HrT ΔgabT This study
WH4 WT-HrT ΔgabP This study
WH5 WT-HrT ΔNcgl1221 ΔgabT This study
WH6 WT-HrT ΔNcgl1221 ΔgabP This study
WH7 WT-HrT ΔgabT ΔgabP This study
WH8 WT-HrT ΔNcgl1221 ΔgabT ΔgabP This study
W2 WT ΔNcgl1221 This study
W3 WT ΔgabT This study
W4 WT ΔgabP This study
W5 WT ΔNcgl1221 ΔgabT This study
W6 WT ΔNcgl1221 ΔgabP This study
W7 WT ΔgabT ΔgabP This study
W8 WT ΔNcgl1221 ΔgabT ΔgabP This study
GAB0 WT harboring pEKEx1 This study
GAB1 WT harboring pGA7 This study
GAB2 W2 harboring pGA7 This study
GAB3 W3 harboring pGA7 This study
GAB4 W4 harboring pGA7 This study
GAB5 W5 harboring pGA7 This study
GAB6 W6 harboring pGA7 This study
GAB7 W7 harboring pGA7 This study
GAB8 W8 harboring pGA7 This study
8
Plasmids
pEKEx1 E. coli – C. glutamicum shuttle vector, pBL1 origin, pUC origin, Ptac, lacIQ, KmR Eikmanns et al. (1991)
pEKTs1 pEKEx1 derivative, temperature-sensitive origin pBL1ts, KmR This study
pCRISPR-Cas9 pGM1190 derivative, codon-optimized S. pyogenes cas9, sgRNA, AmR Tong et al. (2015)
pCRISPR-dCas9 pGM1190 derivative, codon-optimized S. pyogenes dcas9, sgRNA, AmR Tong et al. (2015)
pEKEx1-Cas9spacer pEKEx1 derivative, S. pyogenes cas9, 49-bp spacer between RBS and start codon, KmR This study
pEKEx1-Cas9 pEKEx1 derivative, hexahistidine-tagged S. pyogenes cas9, KmR This study
pEKEx1-Cas9opt pEKEx1 derivative, hexahistidine-tagged, codon-optimized S. pyogenes cas9, KmR This study
pEKEx1-dCas9opt pEKEx1 derivative, hexahistidine-tagged, codon-optimized S. pyogenes dcas9, KmR This study
pEKTs-Cas9opt pEKEx1-Cas9opt derivative, temperature-sensitive origin pBL1ts, KmR This study
pUC19 E. coli cloning vector, pUC origin, ApR Yanisch-Perron et al. (1985)
pWAS ColE1 origin, araC, PBAD-cI(ts2), PR-sRNA scaffold-T1/TE terminator, ApR Na et al. (2013)
pUC19-sgRNA pUC19 derivative, sgRNA scaffold-T1/TE terminator, ApR This study
pEKEx1-argR1 pEKEx1 derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–AGC TCT CAT TTT GCA GAT TT–3ʹ (gs_argR1), KmR This study
pCG9-argR1 pEKEx1-Cas9opt derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–AGC TCT CAT TTT GCA GAT TT–3ʹ (gs_argR1), KmR This study
pCG9-argR2 pEKEx1-Cas9opt derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–ATC GAG ATC TCG GGA CAA GG–3ʹ (gs_argR2), KmR This study
pdCG9-argR1 pEKEx1-dCas9opt derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–AGC TCT CAT TTT GCA GAT TT–3ʹ (gs_argR1), KmR This study
pCG9ts-Ncgl1221 pEKTs1-Cas9opt derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–AAG CCT CGG AGA GAG TGA TC–3ʹ (gs_1221), KmR This study
pCG9ts-gabT pEKTs1-Cas9opt derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–TCG ACA ACG CGT ACC ACG GA–3ʹ (gs_gabT), KmR This study
pCG9ts-gabP pEKTs1-Cas9opt derivative, Ptac-sgRNA-T1/TE terminator with guide sequence 5ʹ–TGG CCA TGT AAT AGG CCA CT–3ʹ (gs_gabP), KmR This study
pCDFDuet-1 E. coli expression vector, CloDF13 origin, two sets of PT7, lacIQ, SmR Novagen
pTac15K E. coli expression vector, p15A origin, Ptac, KmR Lee et al. (2008)
pACYC184 E. coli cloning vector, p15A origin, CmR, TcR Chang and Cohen (1978)
pKCA212-MCS E. coli – C. glutamicum shuttle vector, pCC1 origin, p15A origin, KmR Shin et al. (2016)
pTacCC1 pCC1 origin, Ptac, SmR This study
pTacCC1-recT pTacCC1 derivative, E. coli K-12 MG1655 recT, SmR This study
pTacCC1-HrT pTacCC1 derivative, hexahistidine-tagged E. coli K-12 MG1655 recT, SmR This study
pTacCC1-EGFP pTacCC1 derivative, EGFP, SmR This study
pES0-EGFP pEKEx1 derivative, lacIQ deleted, EGFP, KmR Lab stockpGA7 pEKEx1 derivative, Lactobacillus brevis ATCC 367 gadB2, KmR This study
aKm, kanamycin; Am, apramycin; Sm, spectinomycin; R, resistance; nt, nucleotide.
9

Table S2
M9*_F series single-stranded oligodeoxyribonucleotides (ssODNs) to clone guide sequences
ssODN Sequencea Guide sequence Target description
M9argR1_F
TTGACAATTAATCATCGGCTCGTATAATGTGTG
GAGCTCTCATTTTGCAGATTTGTTTTAGAGCTAGAAATAG
CAAGT
gs_argR1 argR, leading strand
M9argR2_F
TTGACAATTAATCATCGGCTCGTATAATGTGTG
GATCGAGATCTCGGGACAAGGGTTTTAGAGCTAGAAATA
GCAAGT
gs_argR2 argR, lagging strand
M91221_F
TTGACAATTAATCATCGGCTCGTATAATGTGTG
GAAGCCTCGGAGAGAGTGATCGTTTTAGAGCTAGAAATA
GCAAGT
gs_1221 Ncgl1221, leading strand
M9gabT_F
TTGACAATTAATCATCGGCTCGTATAATGTGTG
GTCGACAACGCGTACCACGGAGTTTTAGAGCTAGAAATA
GCAAGT
gs_gabT gabT, leading strand
M9gabP_F
TTGACAATTAATCATCGGCTCGTATAATGTGTG
GTGGCCATGTAATAGGCCACTGTTTTAGAGCTAGAAATA
GCAAGT
gs_gabP gabP, lagging strand
aUnderscored letters indicate 20-nt guide sequence for targeting chromosomal DNA of C. glutamicum ATCC 13032.
10

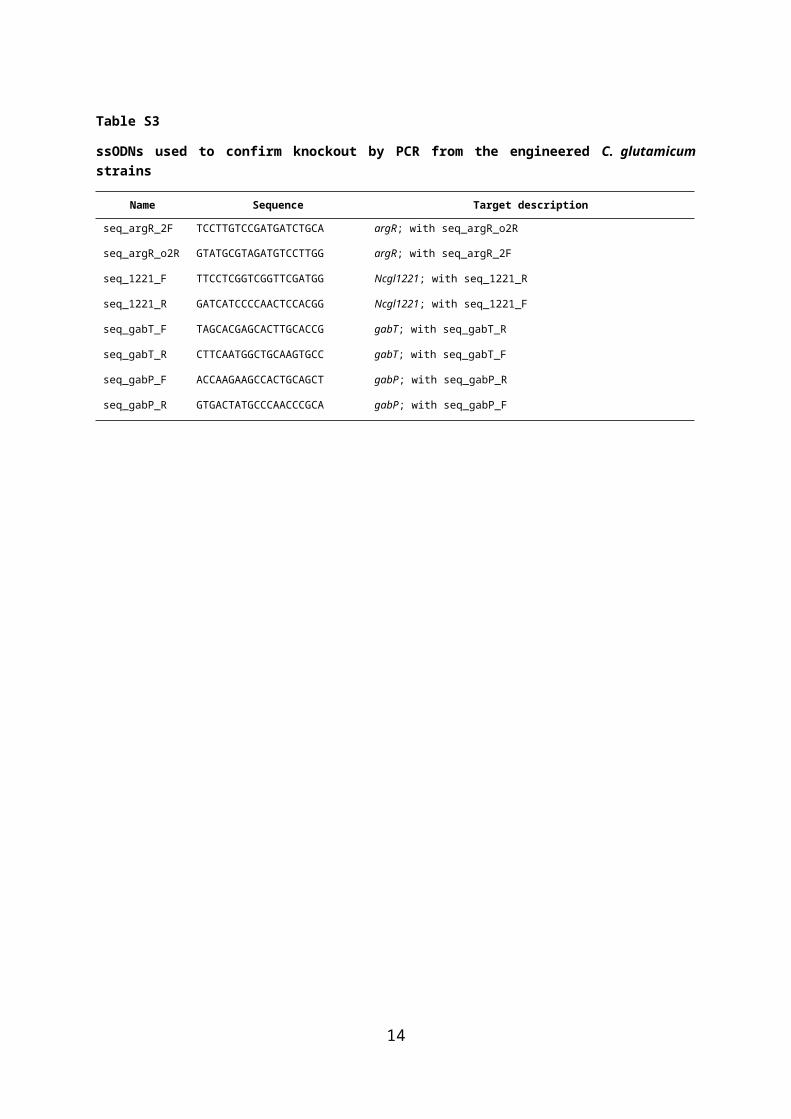
Table S3
ssODNs used to confirm knockout by PCR from the engineered C. glutamicum strains
Name Sequence Target description
seq_argR_2F TCCTTGTCCGATGATCTGCA argR; with seq_argR_o2R
seq_argR_o2R GTATGCGTAGATGTCCTTGG argR; with seq_argR_2F
seq_1221_F TTCCTCGGTCGGTTCGATGG Ncgl1221; with seq_1221_R
seq_1221_R GATCATCCCCAACTCCACGG Ncgl1221; with seq_1221_F
seq_gabT_F TAGCACGAGCACTTGCACCG gabT; with seq_gabT_R
seq_gabT_R CTTCAATGGCTGCAAGTGCC gabT; with seq_gabT_F
seq_gabP_F ACCAAGAAGCCACTGCAGCT gabP; with seq_gabP_R
seq_gabP_R GTGACTATGCCCAACCCGCA gabP; with seq_gabP_F
11

Table S4
ssODNs introduced to C. glutamicum strains for Cas9- and RecT-based knockout
Name Sequencea Description
ssODNargR_400TAATTCACCTAGTTCTTTACCTGTGAGCGGATCACGGGC
GTGCGAGTCACGGGATTTAAGTTTTCCGGTGTTGACGGGGTargR; lagging strand; 400-bp deletion
ssODN1221_250
CCTTACACGTTGAGCATGAAGAGCCAAAGACCTCGCTTA
TCAGAAGATCCTGTCGTTTGGCGGACGTGTCCGCATGAGC
A
Ncgl1221; lagging strand; 250-bp
deletion; for ΔgabT strain generation
ssODN1221_400TCGTGCAAGTCACCGCCGGCAATCAATGGCTGGTCGAAC
GCGCATGAGCACGTCCCTGTTGTTGGGTGCGCTGCTCTTGC
Ncgl1221; lagging strand; 400-bp
deletion; for ΔgabT strain generation
ssODNgabT_100CCGTCGCGCAGTGGGTAAGACATTGGTGCACGGTAGACG
TGACCGCGCCCTTGCCGGTGTAGGCGCGTGCCACCTTGACGgabT; lagging strand; 100-bp deletion
ssODNgabT_300GTGGAAATGGCTGCGAGGAATCCTGGTGCGGGGACGATG
ACGGCTTCGGCGCCAGAGTTAAACAGCGCGCTCTTCTTGTC
gabT; lagging strand; 300-bp deletion;
for ΔgabT strain generation
ssODNgabP_150
GCGCATCGCGGTGAATCCAATGGCGATGAATGAAGTGAA
CATGCCGATGATGATCGCGCGTGGCACTGCCTTCGGGGTG
G
gabP; lagging strand; 150-bp deletion;
for ΔgabP strain generation
ssODNgabP_400AGTGGTGGAAGAACAGTTAGTCCGACAGCAACGGATCGC
TAAGCACTGTCCATCCACACAGCACGATGATGATCGTTGCC
gabP; lagging strand; 400-bp deletion;
for ΔgabP strain generation
aEach 40-nt homology sequence in the ssODN are distinguished by the presence and absence of underscore.
12
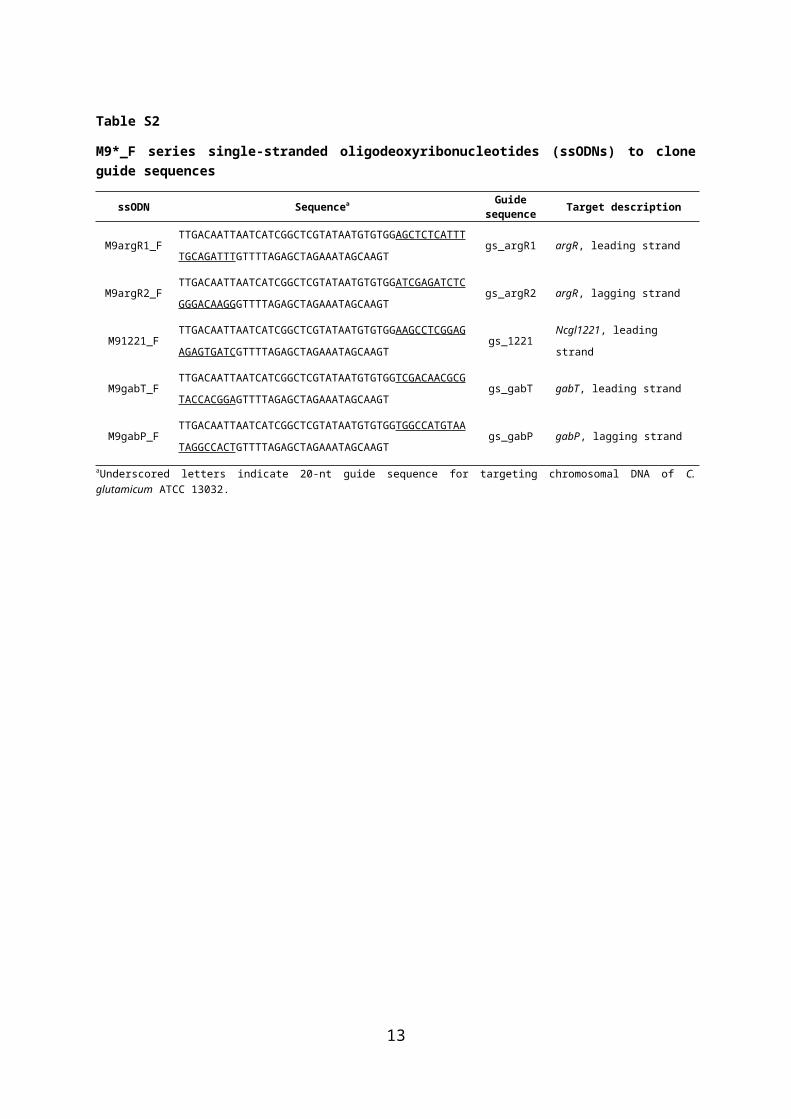
Table S5
Analysis of sgRNA guide sequences through CRISPy-web
Guide sequence
Target gene
Position (bp)a Sequence Strand PAM Number of promiscuous targetsb
0-bp 1-bp 2-bp gs_argR1 argR 20-43 AGCTCTCATTTTGCAGATTT Leading TGG 0 1 10
gs_argR2 argR 115-138 ATCGAGATCTCGGGACAAGG Lagging TGG 0 0 2
gs_1221 Ncgl1221 435-458 GCACTGATTACTACCCAAAT Lagging CGG 0 0 1
gs_gabT gabT 457-480 TCGACAACGCGTACCACGGA Leading CGG 0 0 0
gs_gabP gabP 827-850 TGGCCATGTAATAGGCCACT Lagging GGG 0 0 0aPosition is measured from the beginning of the coding sequence.bNumber of promiscuous targets with 0-, 1-, or 2-bp mismatches of the sgRNA sequence.
13
References
Bachmann, B.J., 1996. Derivations and genotypes of some mutant derivatives of Escherichia coli K-12. In: Neidhardt, F. C., (Ed.), Escherichia coli and Salmonella typhimurium: cellular and molecular biology vol. 2. ASM Press, Washington, DC, pp. 2460-2488.
Chang, A.C., Cohen, S.N., 1978. Construction and characterization of amplifiable multicopy DNA cloning vehicles derived from the P15A cryptic miniplasmid. J. Bacteriol. 134, 1141-1156.
Eikmanns, B.J., Kleinertz, E., Liebl, W., Sahm, H., 1991. A family of Corynebacterium glutamicum/Escherichia coli shuttle vectors for cloning, controlled gene expression, and promoter probing. Gene. 102, 93-98.
Kinoshita, S., Nakayama, K., Akita, S., 1958. Taxonomical study of glutamic acid accumulating bacteria, Micrococcus glutamicus nov. sp. Bull. Agr. Chem. Soc. Japan. 22, 176-185.
Lee, S.Y., Lee, J.W., Song, H., Kim, J.M., Choi, S., Park, J.H., 2008. Recombinant microorganism having an ability of using sucrose as a carbon source. US patent 20110269183.
Na, D., Yoo, S.M., Chung, H., Park, H., Park, J.H., Lee, S.Y., 2013. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 31, 170-174.
Shin, J.H., Park, S.H., Oh, Y.H., Choi, J.W., Lee, M.H., Cho, J.S., Jeong, K.J., Joo, J.C., Yu, J., Park, S.J., Lee, S.Y., 2016. Metabolic engineering of Corynebacterium glutamicum for enhanced production of 5-aminovaleric acid. Microb. Cell Fact. 15, 174.
Studier, F.W., Moffatt, B.A., 1986. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J. Mol. Biol. 189, 113-130.
Tong, Y., Charusanti, P., Zhang, L., Weber, T., Lee, S.Y., 2015. CRISPR-Cas9 based engineering of actinomycetal genomes. ACS Synth. Biol. 4, 1020-1029.
Yanisch-Perron, C., Vieira, J., Messing, J., 1985. Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene. 33, 103-119.
14