Embed Size (px)
Citation preview

Cerebral Cortex November 2011;21:2599--2611
doi:10.1093/cercor/bhr046
Advance Access publication April 5, 2011
Ascl1 Participates in Cajal--Retzius Cell Development in the Neocortex
Rajiv Dixit1, Celine Zimmer2,3, Ronald R. Waclaw4,5, Pierre Mattar1,6, Tarek Shaker1, Christopher Kovach1, Cairine Logan1,
Kenneth Campbell4, Francxois Guillemot2 and Carol Schuurmans1
1Hotchkiss Brain Institute, Alberta Children’s Hospital Research Institute, University of Calgary, Calgary, Alberta, Canada T2N
4N12Division of Molecular Neurobiology, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, UK
3Current address: Institut de biologie du developpement de Marseille Luminy/Unite Mixte de Recherche 6216, 13009 Marseille,
France4Current address: Division of Developmental Biology, Cincinnati Children’s Hospital Medical Center, University of Cincinnati
College of Medicine, Cincinnati, OH 45229, USA5Current address: Division of Experimental Hematology and Cancer Biology, Cincinnati Children’s Hospital Medical Center,
University of Cincinnati College of Medicine, Cincinnati, OH 45229, USA6Current address: Cellular Neurobiology Research Unit, Institut de recherches cliniques de Montreal, Montreal, Quebec, Canada
H2W 1R7
Rajiv Dixit, Celine Zimmer, and Ronald R. Waclaw have contributed equally to this work
Address correspondence to Carol Schuurmans, University of Calgary, 3330 Hospital Drive NW, Calgary, Alberta, Canada T2N 4N1.
Email: [email protected].
Cajal--Retzius cells are essential pioneer neurons that guideneuronal migration in the developing neocortex. During develop-ment, Cajal--Retzius cells arise from distinct progenitor domainsthat line the margins of the dorsal telencephalon, or pallium. Here,we show that the proneural gene Ascl1 is expressed in Cajal--Retzius cell progenitors in the pallial septum, ventral pallium, andcortical hem. Using a short-term lineage trace, we demonstrate thatit is primarily the Ascl1-expressing progenitors in the pallial septumand ventral pallium that differentiate into Cajal--Retzius cells.Accordingly, we found a small, albeit significant reduction in thenumber of Reelin1 and Trp731 Cajal--Retzius cells in the Ascl12/2
neocortex. Conversely, using a gain-of-function approach, we foundthat Ascl1 induces the expression of both Reelin, a Cajal--Retziusmarker, and Tbr1, a marker of pallial-derived neurons, in a subset ofearly-stage pallial progenitors, an activity that declines overdevelopmental time. Taken together, our data indicate that theproneural gene Ascl1 is required and sufficient to promote thedifferentiation of a subset of Cajal--Retzius neurons during earlyneocortical development. Notably, this is the first study that reportsa function for Ascl1 in the pallium, as this gene is best known for itsrole in specifying subpallial neuronal identities.
Keywords: basic-helix-loop-helix, dorsal telencephalon, pallium, proneuralgene, transcription factor
Introduction
The neocortex is comprised of 6 neuronal layers that are
generated in a sequential inside-out manner during develop-
ment. The laminar pattern of the neocortex is established in
part by Cajal--Retzius cells, which are essential pioneer neurons
that guide neuronal migration during development. One of the
critical molecules secreted by Cajal--Retzius cells is the
extracellular protein Reelin, the mutation of which results in
a disruption of neuronal layering (Caviness 1982; Morante-Oria
et al. 2003; Hammond et al. 2006; Pla et al. 2006; Yabut et al.
2007; Wagener et al. 2010). Notably, Reelin has also been
implicated in regulating the tangential and subsequent radial
migration of late-born cortical interneurons (Morante-Oria
et al. 2003; Hammond et al. 2006; Yabut et al. 2007), dendrite
formation (Yabut et al. 2007; Chameau et al. 2009), synapto-
genesis (Borrell et al. 1999), and cortical column formation
(Nishikawa et al. 2002; Janusonis et al. 2004), although some of
these roles remain controversial (Pla et al. 2006; Wagener et al.
2010).
Lineage tracing and birthdating experiments have demon-
strated that Cajal--Retzius cells differentiate between embry-
onic day (E) 10 and E13.5 in mouse, originating at focal sites in
the margins of the dorsal telencephalon (i.e., pallium) (Wood
et al. 1992; Hevner et al. 2003; Takiguchi-Hayashi et al. 2004;
Muzio and Mallamaci 2005). The sites of Cajal--Retzius cell
genesis include the ventral pallium, pallial septum, cortical
hem, presumptive choroid plexus, and rostral telencephalon
(Takiguchi-Hayashi et al. 2004; Bielle et al. 2005; Yoshida et al.
2006; Imayoshi et al. 2008; Zimmer et al. 2010). According to
their pallial sites of origin, Cajal--Retzius cells express pan-
cortical-specific markers (e.g., the T-box transcription factor
Tbr1) and have a glutamatergic neurotransmitter phenotype
(del Rio et al. 1995; Gorski et al. 2002; Hevner et al. 2003).
However, Cajal--Retzius cells are also molecularly heteroge-
neous, as highlighted by their regional-specific marker expres-
sion. For example, Trp73 is expressed in Cajal--Retzius cells
derived from the pallial septum and cortical hem, while Er81/
Etv1 is specific to rostral telencephalic derivatives (Meyer et al.
2002; Takiguchi-Hayashi et al. 2004; Bielle et al. 2005; Zimmer
et al. 2010). The subtype specificity of Cajal--Retzius cells is
thought to be functionally significant, allowing these cells to
participate in patterning the embryonic neocortex through
their regional-specific secretion of distinct morphogens
(Borello and Pierani 2010; Griveau et al. 2010).
The heterogeneity of Cajal--Retzius cells arises in part due to
the involvement of distinct signaling molecules patterning each
progenitor domain. For instance, Fgf8 signaling is implicated in
specifying an Er81/Etv1+Cajal--Retzius cell identity in the
rostral pallium (Zimmer et al. 2010), while Tgfb signaling is
involved in the cortical hem (Siegenthaler and Miller 2008).
Several transcription factors that control Cajal--Retzius cell
development have also been identified. Negative regulators
include the homeodomain transcription factors Pax6
(Stoykova et al. 2003) and Lhx2 (Bulchand et al. 2001; Monuki
et al. 2001), the winged helix factor Foxg1 (Hanashima et al.
2004; Muzio and Mallamaci 2005; Hanashima et al. 2007), the
basic-helix-loop-helix (bHLH) proteins Hes1, Hes3, and Hes5
� The Author 2011. Published by Oxford University Press. All rights reserved.
For permissions, please e-mail: [email protected] from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

(Imayoshi et al. 2008), and the nuclear receptor CoupTFI
(Studer et al. 2005). While some of these factors appear to
directly suppress Cajal--Retzius cell generation (e.g., CoupTFI),
others may act indirectly as they are required to pattern the
pallial signaling domains from which Cajal--Retzius cells arise.
Examples of indirect factors are Foxg1 (Hanashima et al. 2004;
Muzio and Mallamaci 2005; Hanashima et al. 2007) and Lhx2
(Bulchand et al. 2001; Monuki et al. 2001), both of which are
required to prevent expansion of the Wnt-rich cortical hem
into neocortical domains. In contrast, Hes1/3/5 are required
to promote a non-neural choroid plexus fate at the expense
of a neural (i.e., Cajal--Retzius cell generating) cortical hem
identity (Imayoshi et al. 2008).
Transcription factors that act in a positive fashion to
promote Cajal--Retzius cell development include the T-box
factor Tbr1 (Hevner et al. 2001) and the LIM homeodomain
transcription factor Lhx5 (Miquelajauregui et al. 2010), as well
as Trp73 (transformation related protein 73), all of which are
expressed postmitotically and act as differentiation factors. In
addition, the homeodomain genes Emx1 and Emx2 are
required to establish the Wnt-rich cortical hem domain and
thus likely act indirectly to promote cortical hem--derived
Cajal--Retzius cell differentiation (Mallamaci et al. 2000;
Shinozaki et al. 2002; Muzio and Mallamaci 2005; von Frowein
et al. 2006). The proneural bHLH gene Neurogenin (Neurog) 2
is also required and partially sufficient to specify a Cajal--Retzius
cell fate (Imayoshi et al. 2008). Nevertheless, some Cajal--
Retzius cells are generated in the absence of Neurog2 function
(Imayoshi et al. 2008), suggesting that additional proneural
genes may be involved in specifying the identity of these
pioneer neurons.
The proneural bHLH gene achaete-scute-complex-homolog-
like-1 (Ascl1) has a well-established role in ventral telence-
phalic (i.e., subpallial) progenitors, where it is robustly
expressed and is required and sufficient to specify a ventral,
GABAergic neuronal identity (Casarosa et al. 1999; Horton et al.
1999; Britz et al. 2006; Wang et al. 2009). However, Ascl1 is also
expressed at lower levels in pallial progenitors (Britz et al.
2006), where its function(s) remain poorly defined. Here, we
show that Ascl1 is expressed in a subset of Cajal--Retzius cell
progenitors in the cortical hem, pallial septum, and ventral
pallium. Furthermore, using loss-of-function analyses, we show
that Ascl1 is indeed required for the differentiation of a subset
of Cajal--Retzius cells. Finally, using a gain-of-function approach,
we demonstrate that at early stages of Cajal--Retzius cell
development (E10.5), Ascl1 can promote the expression of
Tbr1, a pallial-specific marker (Hevner et al. 2001), and Reelin,
a Cajal--Retzius cell marker, in a subset of pallial progenitors, an
activity that declines over developmental time.
Materials and Methods
Animals and GenotypingTo generate staged embryos, timed pregnant matings were set up, with
the morning of the vaginal plug considered E0.5. The Ascl1 and
Neurog2GFPKI mutant alleles were maintained on a CD1 outbred
background (Charles River). Genotyping of Ascl1 mutant and wild-type
alleles (Casarosa et al. 1999), the Ascl1:GFP transgene (Gong et al.
2003), and Neurog2 wild-type and Neurog2GFPKI mutant alleles (Britz
et al. 2006) were performed using polymerase chain reaction as
previously described. Timed pregnant CD1 females were also used for
all electroporation experiments.
Tissue Processing and RNA In Situ HybridizationEmbryos were dissected at the specified embryonic stages in ice-cold
phosphate-buffered saline (PBS, pH 7.5). Embryos were transferred to
4% paraformaldehyde (PFA)/13 PBS and fixed overnight at 4 �C.Embryos were then washed 3 3 10 min in 13 PBS and transferred to
20% sucrose/13 PBS overnight at 4 �C. Fixed embryos were then
embedded and frozen in Tissue Tek OCT (Sakura Finetek USA, Inc.)
prior to cryosectioning at 10 lm and mounting on Superfrost plus
slides (Fisher Scientific). RNA in situ hybridization was performed as
previously described (Alam et al. 2005), using digoxygenin (dig)-
labeled riboprobes that were generated with a 10X dig-labeling mix
according to the manufacturer’s instructions (Roche Diagnostics
GmbH). Linearized templates and corresponding riboprobes were
generated with the following restriction enzyme/polymerase combi-
nations: Ascl1 (XbaI/Sp6), Reln (EcoR I/T3), Trp73 (Sal I/T3), and
Wnt3a (SacI/Sp6).
ImmunostainingEmbryos were processed as described above. Immunostaining was
performed on 10 lm cryostat sections. Cryosections were blocked
either in 10% normal goat or donkey serum/13 PBS/0.1% Triton X-100
(PBT) or in 3% bovine serum albumin/13 TBST (Tris-buffered saline:
25 mM Tris--HCl, pH 7.4/0.14 M NaCl/0.1% Triton X-100). Primary
antibodies were incubated on sections for 2 h at room temperature or
overnight at 4 �C in blocking solution. Sections were washed 3 3 10
min in PBT or TBST and then incubated with secondary antibodies
for 1 h at room temperature. Sections were then washed 3 3 10 min
in PBT or TBST and counterstained 5 min with 4,6-diamidino-2-
phenylindole (DAPI, 1/10 000, Sigma). Finally, sections were washed
3 3 10 min in PBS or TBST and mounted in AquaPolymount
(Polysciences, Inc.). Some immunolabeling was processed using
secondary antibodies coupled to horseradish peroxidase and a Dia-
minobenzidine substrate, according to the manufacturer’s instruc-
tions (VectaStain ABC kit, Vector Laboratories). Primary antibodies
included: rabbit anti-calretinin (1/2000; Swant), mouse anti-Ascl1 (1/
200; BD Biosciences), rabbit anti-Neurog2 (1/20) (gift of David
Anderson), rabbit anti-Neurog2 (1/500; gift of Masato Nakafuku),
rabbit anti-GFP (1/500; Chemicon), sheep anti-GFP (1/750; Bio-
genesis), rabbit anti-Tbr1 (1/3000; Chemicon), mouse anti-Reelin (1/
500; Chemicon), rabbit anti-pan-Dlx antibody (1/500; gift from Grace
Panganiban), rabbit anti-pan-Otx antibody (1/500; Baas et al. 2000),
rabbit anti-calbindin D-28k (1:1000; Swant), and mouse-anti-Trp73 (1/
200; Lab Vision). Secondary antibodies included goat or donkey anti-
mouse or anti-rabbit antibodies conjugated to Alexa488 (1/500;
Invitrogen Canada Inc.), Cy3 or Cy5 (1/500; Jackson Immunore-
search).
In Utero Electroporation and Embryo CultureIn utero electroporation was performed as previously described (Saito
and Nakatsuji 2001; Langevin et al. 2007). Expression vectors were
generated by subcloning Ascl1 into pCIG2, which contains a chicken
b-actin/Cytomegalovirus promoter/enhancer and an IRES2-EGFP
cassette for visualization of transfected cells (Britz et al. 2006; Mattar
et al. 2008). DNA was purified using a Qiagen Endotoxin free kit
(Qiagen). An electric current (7 pulses of 40--50 mV) was applied
across the uterine wall surrounding injected embryos with 5 mm
platinum tweezer-style electrodes (Protech) and a BTX square wave
electroporator (VWR, CanLab). The uterus was then returned to the
abdominal cavity and embryos were allowed to develop to appropriate
developmental stages. Electroporation and culture of E10.5 embryos
were performed as previously described (Hand et al. 2005; Friocourt
et al. 2008; Gohlke et al. 2008).
Imaging, Cell Counts, and StatisticsImaging was carried out on a Leica TCS 4D confocal microscope or
a Leica DMRXA2 microscope using OpenLab software (Improvision).
For lineage analyses, 2 confocal photographs (212 3 212 lm; stacks of
3 lm) per area per embryo (n = 2) were analyzed. For the
electroporation data: 2--3 confocal photographs (212 3 212 lm;
stacks of 3 lm) per area per embryo (n = 4) were analyzed. For
counting of RNA in situ hybridization signal, 3 digital images were
2600 Ascl1 Function in Cajal--Retzius Cells d Dixit et al.Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

captured for each embryo (n = 3 per genotype). Error bars reflect
standard error of the mean. Student’s 2-tailed t-tests were performed
with P values denoted as follows: <0.05*, <0.01**, <0.005***.
Results
Ascl1 Is Expressed in Cajal--Retzius Cell ProgenitorDomains in the Pallium
Ascl1 is a proneural gene that is expressed at high levels in
subpallial progenitors, where it is required and sufficient to
specify a GABAergic interneuron identity (Casarosa et al. 1999;
Horton et al. 1999; Britz et al. 2006; Wang et al. 2009).
However, Ascl1 is also expressed at lower levels in pallial
progenitors (Britz et al. 2006), where its role has yet to be
elucidated. To determine if Ascl1 is expressed in pallial
territories that give rise to Cajal--Retzius cells, we first
performed RNA in situ hybridization between E10.5 and
E12.5, when the bulk of Cajal--Retzius cells differentiate (Wood
et al. 1992; Hevner et al. 2003; Takiguchi-Hayashi et al. 2004).
Cajal--Retzius cells are derived from progenitor domains that
line the pallial margins, including the pallial septum, ventral
pallium, and cortical hem (schematic; Fig. 1A,B). In E10.5
coronal sections through the rostral telencephalon, Ascl1
transcripts were detected at high levels in the lateral ganglionic
eminence (lge) of the subpallium and at lower levels in
scattered pallial progenitors, including in the pallial septum and
ventral pallium (Fig. 1C,C#). In more caudal sections, Ascl1
transcripts were detected in the lge, as well as the ventral
pallium and cortical hem, which serve as lateral and caudome-
dial signaling centers, respectively, and are major sources of
Cajal--Retzius cells (Fig. 1D--D$). Similar expression profiles
were apparent at E11.5 (Fig. 1E,E#,F--F$) and E12.5 (Fig.
1G,G#,H--H$), with high Ascl1 transcript levels detected in
subpallial progenitors and lower Ascl1 expression levels
throughout the pallial ventricular zone (vz), including in the
pallial septum, ventral pallium, and cortical hem. To confirm
that Ascl1 transcripts were translated into protein in pallial
progenitors, we also performed immunolabeling at E10.5 (Fig.
2A--L and Supplementary Fig. S1B--B#), E11.5 (Supplementary
Fig. S1C--C#), and E12.5 (Fig. 1I,I#,J--J$ J$ and Supplementary
Fig. S1D--D#). At all stages, Ascl1 protein was detected at low
levels in pallial progenitors compared with its robust expres-
sion in the vz of the subpallium. Within the pallium, Ascl1
protein was detected at higher levels in the cortical hem and at
lower levels in the ventral pallium and pallial septum (Fig. 1I,J,J$).To confirm that Ascl1 was indeed expressed in progenitors
that had a pallial identity, rather than in subpallial progenitors
that had migrated tangentially into the dorsal telencephalon,
we performed coimmunolabeling with Ascl1 and a series of
pallial markers. Focusing on the E10.5 ventral pallium, we
observed that the vast majority of Ascl1+progenitors coex-
pressed Pax6, a paired homeodomain transcription factor that
shows a bias toward higher expression levels in progenitors on
the pallial side of the pallial--subpallial border (Fig. 2A--D).
Similarly, we found that in the E10.5 pallium, 38.2 ± 4.1% Ascl1+
progenitors coexpressed Neurog2 (n = 2; Fig. 2E--H), a proneu-
ral bHLH transcription factor that is exclusively expressed
on the pallial side of the boundary and which is required for
the generation of a subset of Cajal--Retzius neurons (Imayoshi
et al. 2008). Notably, proneural proteins are expressed in a ‘‘salt-
and-pepper’’ fashion, and only 10.2 ± 1.2% of E10.5 pallial
progenitors (labeled by DAPI) expressed Ascl1 (n = 4) while
6.4 ± 0.8% expressed Neurog2 (n = 2; Fig. 2E--H). We also noted
temporal and spatial differences in Ascl1 and Neurog2 ex-
pression in pallial progenitors. Ascl1 was detected at higher
levels than Neurog2 within the E10.5 pallial septum, while at
later stages (E11.5 and E12.5), Ascl1 was expressed at lower
levels than Neurog2 in the rostromedial pallium (Supplemen-
tary Fig. S1).
Ascl1 and Neurog2 are therefore expressed in distinct tem-
poral and spatial patterns in Cajal--Retzius cell progenitor do-
mains, suggesting they may have nonoverlapping roles in
regulating the differentiation of these pioneer neurons.
Ascl1 Is Expressed in Cajal--Retzius Cell Lineages ofPallial Origin
To better assess the extent to which Ascl1 is expressed in
Neurog2+pallial lineages, we performed short-term lineage
tracing, taking advantage of the perdurance of the GFP reporter
in Neurog2GFPKI/+ embryos (Fig. 2I--L). An analysis of GFP and
Ascl1 coexpression revealed that 41.2% (n = 2 embryos, 438
progenitors) of GFP+pallial progenitors coexpressed Ascl1 in
E10.5 Neurog2GFPKI/+ cortices. Conversely, 66.2% of Mash1
+
progenitors and 58.8% of GFP+progenitors were single positive
in E10.5 Neurog2GFPKI/+ cortices (Fig. 2I--L). These results
suggest either that: 1) Ascl1 is expressed in both Neurog2-
expressing and non-expressing pallial lineages (Nieto et al.
2001) and/or 2) Ascl1 precedes the onset of Neurog2
expression, or persists after GFP is degraded, in a common,
Neurog2-expressing pallial lineage. Regardless of the interpre-
tation, we can conclude that pallial lineages are spatially and/or
temporally heterogeneous with respect to Ascl1 and Neurog2
expression.
We next asked if pallial progenitors expressing Ascl1 indeed
give rise to Cajal--Retzius cells by performing short-term lineage
tracing with Ascl1:GFP BAC transgenic embryos (Parras et al.
2007). We quantified the number of GFP+cells that coex-
pressed Reelin, a pan-Cajal--Retzius cell marker (Schiffmann
et al. 1997; Alcantara et al. 1998), and Trp73, which labels
Cajal--Retzius cells derived from the pallial septum and cortical
hem (Meyer et al. 2002, 2004). GFP-labeled Cajal--Retzius cells
were separately quantified in the ventral pallium/piriform
cortex in lateral domains, the pallial septum in the rostral
midline, and the cortical hem in the caudal midline (as shown
in schematic; Fig. 3A). In E11.5 Ascl1:GFP cortices, a significant
proportion of Reelin+and Trp73
+cells coexpressed GFP in the
ventral pallium/piriform cortex (34 ± 1.5% GFP+Reelin
+/Reel-
in+; 10 ± 0.2% GFP
+Trp73
+/Trp73
+; n = 2; Fig. 3B--D) and pal-
lial septum (29 ± 2.6% GFP+Reelin
+/Reelin
+; 38 ± 7.1%
GFP+Trp73
+/Trp73
+; n = 2; Fig. 3B#,C#,D), while many fewer
Cajal--Retzius cells in the cortical hem were GFP+(2 ± 0.6%
GFP+Reelin
+/Reelin
+; 1 ± 1.7% GFP
+Trp73
+/Trp73
+; n = 2; Fig.
3B$,C$,D).Reelin is also expressed in some cortical interneurons,
although it is not thought to be expressed in these cells until
late embryonic stages (Alcantara et al. 1998). However, to
confirm that the GFP+Reelin
+cells in the E11.5 Ascl1
+lineage
trace were indeed Cajal--Retzius neurons and not Reelin+
interneurons, we performed triple immunolabeling with
the cortical-specific neuronal marker Tbr1 (Hevner et al.
2001). The presence of GFP+Reelin
+Tbr1
+cells in the cortical
marginal zone of E11.5 Ascl1:GFP embryos confirmed that
Cerebral Cortex November 2011, V 21 N 11 2601Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

Figure 1. Ascl1 is expressed in Cajal--Retzius cell progenitor domains. (A,B) Schematic representation of a coronal section through the E11.5 telencephalon showing the majorCajal--Retzius cell progenitor domains (green; boxed areas), including the ventral pallium (A,B), pallial septum (at rostral levels; A), and cortical hem (at caudal levels; B). (C--H)Distribution of Ascl1 transcripts in coronal sections through the E10.5 (C,C#,D--D#D$), E11.5 (E,E#,F--F#F$), and E12.5 (G,G#,H--H#H$) telencephalon, focusing on rostral(C,C#,E,E#,G,G#) and medial (D--D#D$,F--F#F$,H--H#H$) levels. Black arrowheads mark the pallial--subpallial border, which is demarcated by the transition from low to high Ascl1transcript levels. Boxed areas in C,E,G are in the pallial septum, which are magnified in C#,E#,G#, respectively. Boxed areas in D,F,H are in the ventral pallium and cortical hem,which are magnified in D#,F#,H# and D#D$,F#F$,H#H$, respectively. (I,J) Ascl1 protein expression in coronal sections through the E12.5 telencephalon at rostral (I,I#) and caudal(J-J$) levels. Arrowheads mark the pallial--subpallial border. The boxed area in I is in the pallial septum, which is magnified in I#, and the boxed areas in J are in the ventralpallium and cortical hem, which are magnified in J#,J$, respectively. vp, ventral pallium; ps, pallial septum; ch, cortical hem; mge, medial ganglionic eminence.
2602 Ascl1 Function in Cajal--Retzius Cells d Dixit et al.Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

Ascl1-expressing progenitors give rise to Cajal--Retzius cells
with a pallial-specific regional identity (Fig. 3E--H).
In E11.5 Ascl1:GFP transgenics, GFP was detected on both
sides of the pallial/subpallial border (Fig. 3B--C), consistent
with the expression of Ascl1 in both pallial and subpallial
progenitors (Fig. 1C--J). This raised the remote possibility that
GFP+Reelin
+and GFP
+Trp73
+cells in E11.5 Ascl1:GFP trans-
genics may have a subpallial origin, an interpretation that
would go against the prevailing view that Cajal--Retzius cells are
derived from pallial progenitors (Wood et al. 1992; Hevner et al.
2003; Takiguchi-Hayashi et al. 2004; Bielle et al. 2005; Muzio
and Mallamaci 2005). Nevertheless, we set out to provide
additional support for the pallial origin of Cajal--Retzius cells,
particularly those in the ventral pallium/piriform cortex, which
overlie the subpallium. Short-term lineage tracing in E11.5
Neurog2GFPKI/+ embryos revealed that the vast majority (if not
all) Reelin+(Fig. 3I--I$,K,L--L$) and Trp73
+(Fig. 3J,J$) Cajal--
Retzius cells were GFP+in the ventral pallium/piriform cortex,
pallial septum, and cortical hem. Furthermore, a significant
proportion of Reelin+cells underlying the basal telencephalon
(Fig. 3K,M--M$), which are thought to be derived in part from
the pallium and thalamic eminence (Abellan et al. 2009; Tissir
et al. 2009), were also GFP+in E11.5 Neurog2
GFPKI/+ embryos.
We can thus conclude that the vast majority of Cajal--Retzius
cells present in the E11.5 neocortex and piriform cortex
(where our Ascl1 lineage tracing was focused) are pallial in
origin, while marginal zone cells underlying the basal telen-
cephalon may have a mixed origin.
Taken together, we conclude that Ascl1-expressing progen-
itors give rise to Cajal--Retzius cells, particularly in the pallial
septum and ventral pallium. However, a long-term lineage trace
(e.g., using an Ascl1-CreKI line crossed with a reporter) would
be required to validate this regional bias.
Ascl1 Is Required for the Generation of Cajal--RetziusNeurons
To test if Ascl1 is required for the generation of Cajal--Retzius
cells, we performed cell counts along the rostral-to-caudal
extent of the neocortical marginal zone in wild-type and
Ascl1–/– embryos sectioned along the sagittal plane (Fig. 4A,B).
Cell counts were not performed in a regional-specific manner
as the loss of one Cajal--Retzius cell population is known to be
compensated for by alterations in the migratory routes of Cajal--
Retzius cells derived from other progenitor domains (Bielle
et al. 2005; Pla et al. 2006; Griveau et al. 2010). At E12.5, 1.35-
fold fewer Reelin+Cajal--Retzius cells were detected in Ascl1
–/–
cortices compared with wild-type controls (wild-type: 126.4 ±4.0; n = 3 vs. Ascl1
–/–: 93.1 ± 3.4 Reelin+cells/field; n = 2; P <
0.0001; Fig. 4C,C#,D,D#,G). Similarly, there was a 2.2-fold
decrease in Trp73+Cajal--Retzius cells in E12.5 Ascl1
–/– cortices
(wild-type: 31.2 ± 1.9; n = 3 vs. Ascl1–/–: 14.2 ± 2.1 Trp73
+cells/
Figure 2. Ascl1 is coexpressed with pallial-specific markers. (A--D) Confocal images of Pax6 (green), Ascl1 (red), and DAPI (nuclear marker, blue) expression in theE10.5 pallium, focusing on the pallial--subpallial border (demarcated by dotted line and white arrowhead). Boxed cell in panels A--D is magnified in D to demonstrate thatsome DAPIþ pallial progenitors in the ventral pallium coexpress Ascl1 and Pax6. (E--H) Confocal images of Neurog2 (green), Ascl1 (red), and DAPI (nuclear marker, blue)expression in the E10.5 pallium, focusing on the pallial--subpallial border (demarcated by dotted line and white arrowhead). Boxed cell in E--H is magnified in H todemonstrate that some DAPIþ pallial progenitors in the ventral pallium coexpress Ascl1 and Neurog2. (I--L) Confocal images of GFP (green), Ascl1 (red), and DAPI(nuclear marker, blue) expression in E10.5 Neurog2GFPKI/þ pallium, focusing on the pallial--subpallial border (demarcated by dotted line and white arrowhead). Boxedcell in I--L is magnified in L to demonstrate that some DAPIþ pallial progenitors in the ventral pallium coexpress Ascl1 and GFP in E10.5 Neurog2GFPKI/þ cortices. vp,ventral pallium.
Cerebral Cortex November 2011, V 21 N 11 2603Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

field; n = 2; P < 0.0001; Fig. 4E,E#,F,F#,G). Notably, the reduction
in Cajal--Retzius cell number, including Trp73+cells, which are
derived from the pallial septum and cortical hem, was not due
to a mispatterning of medial signaling centers. Indeed, Wnt3a
was expressed in the hem of both wild-type and Ascl1–/– E12.5
cortices (Fig. 4H,I).
To determine if the reduction in Cajal--Retzius cell number
observed at E12.5 was due to a delay in the differentiation of
these cells in Ascl1 mutants, we also performed cell counts at
E15.5 and E18.5, analyzing both sagittal and coronal sections
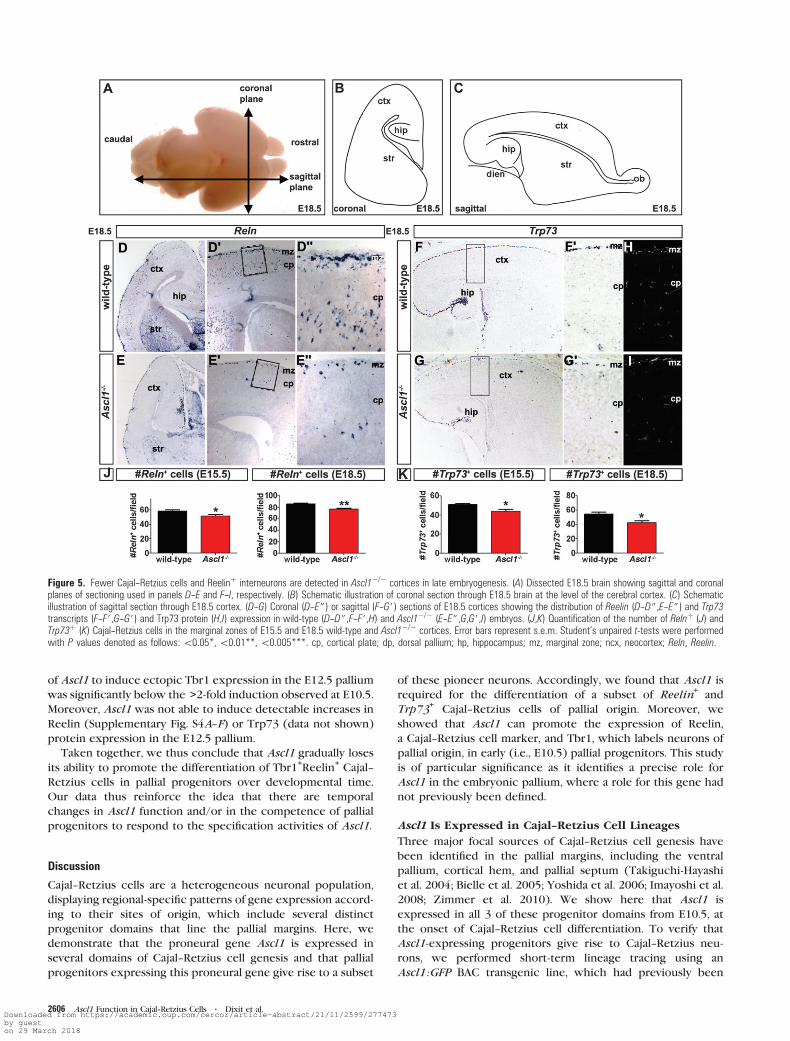
through the neocortex (Fig. 5A--C). At E15.5, the number of
Reelin+(wild-type: 58.6 ± 2.1; n = 3 vs. Ascl1
–/–: 51.8 ± 2.0
Reelin+cells/field; n = 3; P = 0.03; Fig. 5J) and Trp73
+(wild-
type: 50.9 ± 1.4; n = 3 vs. Ascl1–/–:44.0 ± 2.1 Trp73
+cells/field;
n = 3; P = 0.01; Fig. 5K) Cajal--Retzius cells was reduced inAscl1–/–
cortices compared with wild-type controls. The presence of
fewer Reelin+(wild-type: 85.9 ± 1.7; n = 3 vs. Ascl1
–/–: 77.0 ± 1.7
Reelin+cells/field;n = 6; P = 0.006; Fig. 5D--D#,E--E#,J) and Trp73+
(wild-type: 54.4 ± 2.8; n = 5 vs. Ascl1–/–: 42.6 ± 2.5 Trp73
+cells/
field; n = 3; P = 0.01; Fig. 5F,F#,G,G#,H,I,K) Cajal--Retzius cells inthe Ascl1
–/– marginal zone was maintained even at E18.5, when
neocortical neurogenesis is complete (Takahashi et al. 1999).
Ascl1 is therefore required for the generation of a subset of
Cajal--Retzius cells, including Trp73+cells, which are specifi-
cally derived from the cortical hem and pallial septum.
Ascl1 Is Required for the Differentiation of Reelin+
Cortical Interneurons
By E18.5, Reelin is not only expressed in Cajal--Retzius cells in
the upper marginal zone but also in interneurons of subpallial
origin that are interspersed throughout the lower marginal
zone and in neocortical layers II--VI (Fig. 5D--D$) (Alcantara
et al. 1998; Morante-Oria et al. 2003). In E18.5 Ascl1–/– cortices,
the number of Reelin+cells was reduced not only in the
marginal zone but also throughout the cortical plate (Fig. 5E--
E$). The reduction in Reelin+cells in E18.5 Ascl1
–/– cortices is
consistent with the loss of cortical interneurons previously
reported in these mutants (Casarosa et al. 1999; Horton et al.
1999). We verified that fewer cortical interneurons were
generated in E18.5 Ascl1–/– cortices by examining the
expression of several additional interneuron markers. Calreti-
nin, which labels both Cajal--Retzius cells of pallial origin and
cortical plate interneurons, was expressed in fewer cells in
E18.5 Ascl1–/– cortices (Supplementary Fig. S2A,B), as was
Figure 3. Cajal--Retzius cells are derived from Ascl1þ lineages. (A) Schematic representation of coronal section through the E11.5 telencephalon showing sites of analysis in thefollowing figures (red, boxed areas), including the ventral pallium/piriform cortex, pallial septum (at rostral levels), and cortical hem (at medial levels). (B--B$,C--C$) Coexpressionof GFP (green) and Reelin (red; B--B$) or GFP (green) and Trp73 (red; C--C$) in E11.5 Ascl1:GFP cortex at the level of the ventral pallium/piriform cortex (B--C), pallial septum(B#,C#), and cortical hem (B$,C$). Boxed cells in each panel are shown at higher magnification in panels to the right. Yellow dotted line in B,C marks the pallial--subpallial border.Scale bar: 53 lm. (D) Graph showing the percentage of Cajal--Retzius cells labeled with Reelin (red bar) or Trp73 (yellow bar) that derive from the Ascl1 lineage (i.e., GFPþ) in theventral pallium/piriform cortex, pallial septum, and cortical hem. (E--H) Confocal images of GFP (green, E,H), Reelin (red, F,H), and Tbr1 (blue, G,H) expression in E11.5 Ascl1: EGFPpallium, focusing on the pallial--subpallial border (demarcated by dotted line). Boxed cell in E--H is magnified in H to demonstrate that some GFPþ cells in the ventral palliumcoexpress Reelin and Tbr1. (I--M) Coexpression of GFP (green) and Reelin (red; I--I$,K,L--L$,M--M$) or GFP (green) and Trp73 (red; J--J$) in E11.5 Neurog2GFPKIþ/- cortex. Highmagnification images are at the level of the ventral pallium/piriform cortex (I,J,L--L$), pallial septum (I$,J$), cortical hem (I$,J$), or basal telencephalon (M--M$). Boxed cells inI--J$ are shown at higher magnification in panels to the right. Yellow dotted line in I--J marks the pallial--subpallial border. Scale bar in I is 53 lm. ch, cortical hem; mge, medialganglionic eminence; pc, piriform cortex; ps, pallial septum; vp, ventral pallium.
2604 Ascl1 Function in Cajal--Retzius Cells d Dixit et al.Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

calbindin, which labels interneurons in the lower marginal
zone and infragranular layers (Supplementary Fig. S2C,D). A
reduction in cortical interneurons in E18.5 Ascl1–/– cortices
was also evident using pan-Dlx (Supplementary Fig. S2E,F) and
pan-Otx (Supplementary Fig. S2G,H) antibodies, the former
labeling interneurons of subpallial origin, and the latter labeling
both infragranular neurons in the cortical plate and interneur-
ons in supragranular layers and in the marginal zone.
We thus conclude that Ascl1 is required for the generation
of most cortical interneurons, including those that initiate
Reelin expression in the late embryonic period.
Induction of Cajal--Retzius Cell Markers by Ascl1
To test if Ascl1 is sufficient to promote a Cajal--Retzius cell
identity, we misexpressed this gene in the E10.5 pallium using
a bicistronic vector coexpressing GFP and Ascl1 and compared
the results with a control vector expressing only b-galactosi-dase (b-gal; Fig. 6I). Electroporated brains were cultured for 2
days in vitro (DIV) and then immunolabeled with antibodies to
Reelin, a pan-Cajal--Retzius cell marker, and Tbr1, a marker of
postmitotic neurons of pallial derivation (Hevner et al. 2001).
Strikingly, Ascl1 promoted a 2.1-fold increase in the number
of cells that expressed Tbr1 compared with control electro-
porations (control: 23.2 ± 1.3% b-gal+ cells coexpressed Tbr1+;
n = 4; 1092 electroporated cells; Fig. 6A--D,J vs. 47.6 ± 4.4%
GFP+cells coexpressed Tbr1
+after Ascl1 overexpression; n = 4
733 electroporated cells; Fig. 6E--H,J; P < 0.0001). Moreover,
2.1-fold more Ascl1-electroporated cells expressed both Tbr1
and Reelin (i.e., 14.9 ± 1.8% of GFP+cells wereGFP
+Tbr1
+Reelin
+;
n = 4; 733 electroporated cells; P = 0.0009; Fig. 6E--H,J) compared
with control electroporations (i.e., 7.2 ± 0.9% b-gal+ cells were
b-gal+Tbr1+Reelin+;n = 4; 1092 electroporated cells; Fig. 6A--D,J).Notably, Ascl1 did not induce the expression of other Cajal--
Retzius markers in this assay, including calretinin (Supplemen-
tary Fig. S3A--H) and Trp73 (data not shown).
The cell fate specification properties of the proneural genes,
including Ascl1, are both spatially and temporally restricted
(Britz et al. 2006; Mattar et al. 2008). We therefore tested if
Ascl1 could promote the expression of Cajal--Retzius cell
markers in the E12.5 pallium, at the tail end of their
differentiation period. An advantage of the E12.5 stage was
our ability to efficiently use in utero electroporation to
introduce expression constructs into the pallium, allowing for
normal in vivo development posttransfection (Fig. 7G) (Saito
and Nakatsuji 2001; Langevin et al. 2007; Mattar et al. 2008). In
E12.5 / E15.5 pallial electroporations, Ascl1 promoted a
modest, albeit significant, 1.06-fold increase in the number of
cells that expressed Tbr1 compared with control electro-
porations (Ascl1: 93.09 ± 1.03% GFP+cells coexpressed Tbr1
+;
n = 3; 3990 GFP+electroporated cells vs. control pCIG2: 87.24 ±
1.60% GFP+
cells coexpressed Tbr1+; n = 3; 2826 GFP
+
electroporated cells; Fig. 7A--F,H). However, the minimal ability
Figure 4. Fewer Cajal--Retzius cells are generated in E12.5 Ascl1�/� cortices. (A) Dissected E12.5 brain showing plane of sectioning (marked by line; sagittal plane) used inpanels C--F,H,I. (B) Schematic illustration of sagittal section through E12.5 telencephalon/diencephalon. (C--F) RNA in situ hybridization on sagittal sections of E12.5 wild-type(C,C#,E,E#) and Ascl1�/� (D,D#,F,F#) cortices showing expression of Reelin (C--D#) and Trp73 (E--F#). (E) Reelinþ and Trp73þ cell quantitation in the E12.5 preplate. Error barsrepresent standard error of the mean (s.e.m.). Student’s unpaired t-tests were performed with P values denoted as follows:\0.05*,\0.01**,\0.005***. (H,I) Expression ofWnt3a in E12.5 cortical hem in wild-type (H) and Ascl1�/� (I) cortices. ch, cortical hem; ctx, cortex; dienc, diencephalon; ge, ganglionic eminences; pp, preplate.
Cerebral Cortex November 2011, V 21 N 11 2605Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018
of Ascl1 to induce ectopic Tbr1 expression in the E12.5 pallium
was significantly below the >2-fold induction observed at E10.5.
Moreover, Ascl1 was not able to induce detectable increases in
Reelin (Supplementary Fig. S4A--F) or Trp73 (data not shown)
protein expression in the E12.5 pallium.
Taken together, we thus conclude that Ascl1 gradually loses
its ability to promote the differentiation of Tbr1+Reelin
+Cajal--
Retzius cells in pallial progenitors over developmental time.
Our data thus reinforce the idea that there are temporal
changes in Ascl1 function and/or in the competence of pallial
progenitors to respond to the specification activities of Ascl1.
Discussion
Cajal--Retzius cells are a heterogeneous neuronal population,
displaying regional-specific patterns of gene expression accord-
ing to their sites of origin, which include several distinct
progenitor domains that line the pallial margins. Here, we
demonstrate that the proneural gene Ascl1 is expressed in
several domains of Cajal--Retzius cell genesis and that pallial
progenitors expressing this proneural gene give rise to a subset
of these pioneer neurons. Accordingly, we found that Ascl1 is
required for the differentiation of a subset of Reelin+and
Trp73+
Cajal--Retzius cells of pallial origin. Moreover, we
showed that Ascl1 can promote the expression of Reelin,
a Cajal--Retzius cell marker, and Tbr1, which labels neurons of
pallial origin, in early (i.e., E10.5) pallial progenitors. This study
is of particular significance as it identifies a precise role for
Ascl1 in the embryonic pallium, where a role for this gene had
not previously been defined.
Ascl1 Is Expressed in Cajal--Retzius Cell Lineages
Three major focal sources of Cajal--Retzius cell genesis have
been identified in the pallial margins, including the ventral
pallium, cortical hem, and pallial septum (Takiguchi-Hayashi
et al. 2004; Bielle et al. 2005; Yoshida et al. 2006; Imayoshi et al.
2008; Zimmer et al. 2010). We show here that Ascl1 is
expressed in all 3 of these progenitor domains from E10.5, at
the onset of Cajal--Retzius cell differentiation. To verify that
Ascl1-expressing progenitors give rise to Cajal--Retzius neu-
rons, we performed short-term lineage tracing using an
Ascl1:GFP BAC transgenic line, which had previously been
Figure 5. Fewer Cajal--Retzius cells and Reelinþ interneurons are detected in Ascl1�/� cortices in late embryogenesis. (A) Dissected E18.5 brain showing sagittal and coronalplanes of sectioning used in panels D--E and F--I, respectively. (B) Schematic illustration of coronal section through E18.5 brain at the level of the cerebral cortex. (C) Schematicillustration of sagittal section through E18.5 cortex. (D--G) Coronal (D--E$) or sagittal (F--G#) sections of E18.5 cortices showing the distribution of Reelin (D--D$,E--E$) and Trp73transcripts (F--F#,G--G#) and Trp73 protein (H,I) expression in wild-type (D--D$,F--F#,H) and Ascl1�/� (E--E$,G,G#,I) embryos. (J,K) Quantification of the number of Relnþ (J) andTrp73þ (K) Cajal--Retzius cells in the marginal zones of E15.5 and E18.5 wild-type and Ascl1�/� cortices. Error bars represent s.e.m. Student’s unpaired t-tests were performedwith P values denoted as follows:\0.05*,\0.01**,\0.005***. cp, cortical plate; dp, dorsal pallium; hp, hippocampus; mz, marginal zone; ncx, neocortex; Reln, Reelin.
2606 Ascl1 Function in Cajal--Retzius Cells d Dixit et al.Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

reported to faithfully recapitulate the endogenous expression
of Ascl1 (Parras et al. 2007). In Ascl1:GFP BAC transgenics, we
found that GFP was primarily expressed in Cajal--Retzius cells
that were derived from the ventral pallium and pallial septum.
Surprisingly, despite relatively high Ascl1 transcript and protein
expression in the cortical hem, very little GFP expression was
detected in Cajal--Retzius cells lining the caudomedial marginal
zone (i.e., hem derived) in Ascl1:GFP BAC transgenics. We
speculate that the ability of Ascl1 to specify a Cajal--Retzius cell
fate in the cortical hem may be prevented by the formation of
complexes between Ascl1 and inhibitory factors specifically
present in cortical hem progenitor cells. However, we cannot
rule out the possibility that the Ascl1:GFP BAC transgenic line
does not faithfully label all cells derived from the Ascl1 lineage.
Future long-term lineage tracing experiments using an Ascl1-
creKI line would be necessary to image the full complement of
Ascl1-derived pallial neurons.
What are the signals that regulate Ascl1 expression in the
pallium? Previous reports demonstrated that Ascl1 can be
induced by Fgf signaling in the telencephalon (Kuschel et al.
2003; Gutin et al. 2006), the activity of which is high in the
pallial septum (Zimmer et al. 2010), where Ascl1 is expressed
at high levels from E10.5. Conversely, Wnts repress Ascl1
expression (Backman et al. 2005), yet we found high Ascl1
transcript and protein levels in the cortical hem, which is a rich
source of Wnt signaling (Grove et al. 1998; Lee et al. 2000;
Shimogori et al. 2004). However, the hem is also under the
influence of Bmp and Tgfb signaling (Yoshida et al. 2006;
Hanashima et al. 2007; Siegenthaler and Miller 2008), and it may
be that these signals combined are permissive (or instructive)
for Ascl1 expression.
Ascl1 Is Required in Cajal--Retzius Cell Lineages
Previous clonal analyses of Ascl1–/– pallial progenitors in
a Neurog2-negative lineage revealed that Ascl1 is required to
regulate neuronal differentiation in these cells in an in vitro
clonal assay (Nieto et al. 2001). To date, this has been the only
indication that Ascl1 may have a pallial-specific function. Here,
we report the first in vivo evidence supporting a role for Ascl1
in pallial progenitors. Specifically, we found that Ascl1 is
required for the generation of a subset of Cajal--Retzius cells,
with decreased numbers of both Reelin+and Trp73
+Cajal--
Retzius neurons observed in Ascl1–/– cortices. We suggest that
the reduction in Trp73+Cajal--Retzius cells in Ascl1 mutants,
which are derived from both the cortical hem and the pallial
septum (Nieto et al. 2001; Meyer et al. 2002; Takiguchi-Hayashi
et al. 2004; Bielle et al. 2005), mainly corresponds to Cajal--
Retzius cells derived from the pallial septum for several rea-
sons. Firstly, our short-term lineage tracing (albeit with
the caveats outlined above) suggests that not many Ascl1-
expressing progenitors in the cortical hem differentiate into
Cajal--Retzius cells, whereas the pallial septum is a rich source
of Ascl1-derived Cajal--Retzius cells. Secondly, we show here
that the cortical hem expression of Wnt3a is not perturbed in
Ascl1–/– cortices, suggesting that there are no major patterning
defects in this signaling center in the absence of Ascl1 function.
While additional studies would be required to confirm that the
cortical hem is a functional signaling center in Ascl1 mutants,
we suggest that this is the case. Indeed, the cortical hem is
known to pattern adjacent hippocampal domains (Galceran
et al. 2000; Bulchand et al. 2001), which are normal in Ascl1
mutants (data not shown). Finally, we found that in the E10.5
pallial septum, more progenitors express high levels of Ascl1
protein as compared with Neurog2, a proneural transcription
factor that is also required for the generation of a subset of
Cajal--Retzius cells (Imayoshi et al. 2008), suggesting that Ascl1
may have a nonredundant function in this territory at the
earliest stages of Cajal--Retzius cell differentiation. Should Ascl1
indeed function in the early pallial septum, which is Fgf
signaling dependent (Zimmer et al. 2010), it would be
Figure 6. Ascl1 promotes the differentiation of Tbr1þReelinþ Cajal--Retzius cells in the early E10.5 pallium. (A--H) Triple immunostaining on coronal sections of E10.5 þ 2DIVtelencephalon electroporated with nls-b-gal (b-galactosidase) (A--D) or Ascl1-IRES-GFP (E--H) using antibodies directed against b-gal (A--D) or GFP (E--H) and Reelin (red) andTbr1 (blue). (I) Schematic illustration of the pallial area analyzed in A--H. (J) Quantification of the percentage of markerþ (i.e., b-galþ in A--D or GFPþ in E--H) electroporated cellscoexpressing Tbr1 over the total number of electroporated cells. Quantification of the percentage of markerþ (i.e., b-galþ in A--D or GFPþ in E--H) electroporated cellscoexpressing Tbr1 and Reelin over the total number of electroporated cells. Error bars represent s.e.m. Student’s unpaired t-tests were performed with P values denoted asfollows:\0.05*,\0.01**,\0.005***. mz, marginal zone.
Cerebral Cortex November 2011, V 21 N 11 2607Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

interesting in the future to determine if Ascl1 pallial function(s)
in Cajal--Retzius cell generation depends on Fgf signaling.
Given that Cajal--Retzius cells are reduced in number in
Neurog2–/– (Imayoshi et al. 2008) and Ascl1
–/– cortices, both
genes must have nonredundant roles in regulating the
differentiation of these cells. At first glance, the reduction in
Cajal--Retzius cell number in Neurog2 mutants is somewhat
surprising given the pallial-specific upregulation of Ascl1
expression previously reported in these embryos (Fode et al.
2000; Schuurmans et al. 2004). However, we have found that
Ascl1 is not upregulated in the Neurog2–/– pallium at E10.5,
with increased Ascl1 transcription not evident until ~E12.5(data not shown), when Cajal--Retzius cell genesis is almost
complete. We thus favor the idea that Neurog2 and Ascl1
function in nonoverlapping populations of pallial progenitors
destined to differentiate into distinct types of Cajal--Retzius
cells. Alternatively, we suggest that Neurog2 and Ascl1 may
function in the same lineages but at different times in lineage
progression.
Ascl1 Fate Specification Properties Are TemporallyRegulated in the Pallium
Proneural genes promote both generic aspects of neuronal
differentiation and specify neuronal subtype identities (Talikka
et al. 2002). In the embryonic telencephalon, Ascl1 has
previously been shown to specify a subpallial, GABAergic in-
terneuron identity (Casarosa et al. 1999; Horton et al. 1999;
Britz et al. 2006; Wang et al. 2009). Here, we misexpressed
Ascl1 in the E10.5 pallium, and to our surprise, found that Ascl1
could induce the expression of Tbr1, a pallial-specific neuronal
marker, as well as Reelin, a pan-Cajal--Retzius cell marker. This
suggests that Ascl1 can play an instructive role in specifying
a cortical-specific Cajal--Retzius cell identity in early pallial
progenitors. However, Ascl1 was not sufficient to induce all
Cajal--Retzius cell markers, including calretinin and Trp73. We
suggest that the inability ofAscl1 to induce calretinin expression,
which is normally excluded from pallial septum-derived Cajal--
Retzius cells (Bielle et al. 2005), may reflect a specific role for
Ascl1 in promoting a pallial septum-specific identity. In contrast,
it is not so easy to explain the inability of Ascl1 to induce
Trp73 expression, a marker of Cajal--Retzius cells derived from
the pallial septum and cortical hem (Takiguchi-Hayashi et al.
2004; Bielle et al. 2005). We thus suggest that Ascl1 does not
have the capacity to fully initiate a Cajal--Retzius cell differen-
tiation program, possibly because it is only one component of a
complex of transcription factors that might together be re-
quired. Additional studies examining the combinatorial effects
of different transcription factor complexes on Cajal--Retzius
cell differentiation would be required to further dissect the
transcriptional cascades that are involved in Cajal Retzius cell
fate specification and differentiation.
We have previously shown that the proneural activities of Ascl1
and Neurog2 are temporally and spatially regulated in the
embryonic telencephalon (Schuurmans et al. 2004; Britz et al.
2006; Mattar et al. 2008). Here, we found that the ability of Ascl1
to promote the expression of Cajal--Retzius cell markers is
strikingly reduced by E12.5. Instead, at this later developmental
stage, we previously showed that Asc1l promotes the ectopic
expression of subpallial markers (Britz et al. 2006). Accordingly,
we found here that Ascl1 is required for the differentiation of
a subset of Reelin+Cajal--Retzius cells and almost all Reelin
+
cortical interneurons, reflecting a requirement for Ascl1 in both
Figure 7. Ascl1 has a reduced ability to promote Tbr1 expression in the E12.5 pallium. (A--F) Expression of GFP (green, A,C,D,F) and Tbr1 (red, B,C,E,F) in brains electroporated atE12.5 with pCIG2 (A--C) or pCIG2-Ascl1 (D--F) expression vectors and harvested and analyzed at E15.5. (G) Schematic illustration of coronal section through E15.5 pallium,showing site of analysis (boxed area). (H) Quantification of the percentage of GFPþ cells coexpressing Tbr1. Error bars represent s.e.m. Student’s unpaired t-tests were performedwith P values denoted as follows:\0.05*,\0.01**,\0.005***. ctx, cortex; ig, infragranular layers; iz, intermediate zone; mz, marginal zone; str, striatum; sg, supragranularlayers; svz, subventricular zone.
2608 Ascl1 Function in Cajal--Retzius Cells d Dixit et al.Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

pallial and subpallial domains, respectively. However, we suggest
that the Reelin+Cajal--Retzius cells and Reelin
+cortical inter-
neurons are derived from distinct progenitor domains, an idea
supported not only by lineage studies but also by genetic analyses.
For example, the homeodomain genes Cux1/2 are required for
the generation of Reelin+interneurons in the cortical plate but
not for the generation of Reelin+Cajal--Retzius neurons (Cubelos
et al. 2008). It would thus be of interest in the future to examine if
there are genetic interactions between Ascl1 and Cux1/2 in
subpallial progenitors.
We showed that Ascl1 is expressed at higher levels in subpallial
versus pallial progenitors. This raises the question of whether the
dose of Ascl1 plays an instructive role in cell fate specification,
with the elevated levels of endogenous Ascl1 expression observed
in subpallial progenitors preferentially promoting an interneuron
fate, while the lower Ascl1 expression levels observed in pallial
progenitors promoting a Cajal--Retzius cell fate. However, if Ascl1
gene dosage were simply instructive, we would not expect that
the high levels of Ascl1 expression obtained with our electro-
poration technique would promote the differentiation of Cajal--
Retzius cells at any embryonic stage. Instead, our data support the
idea that there are either changes in Ascl1 itself (e.g., through
posttranslational modifications) and/or temporal changes in
the competence of pallial progenitors to respond to the fate
specification properties of Ascl1. Notably, precedents exist for
temporal changes in the abilities of different transcription factors
to specify cell fates. For instance, the homeobox gene Gsx2
preferentially specifying a striatal projection neuron identity early
in telencephalic development, while later it promotes an olfactory
bulb interneuron identity (Waclaw et al. 2009).
Supplementary Material
Supplementary material can be found at: http://www.cercor
.oxfordjournals.org/
Funding
Canadian Institutes of Health Research (CIHR) Operating Grant to
C.S. (grant number MOP-44094) and Medical Research Council
Institutional Funds to F.G. C.S. is an Alberta Heritage Foundation
for Medical Research (AHFMR) Senior Scholar. C.K. and P.M. were
supported by a CIHR Training Grant in Genetics, Child De-
velopment and Health. P.M. was also supported by a CIHR Canada
Graduate Scholarship,; Alberta Heritage Foundation for Medical
Research Studentship; and a Heart and Stroke Foundation
Studentship and C.K. received a QEII Scholarship. R.D. was
supported by a CIHR Canada HOPE Fellowship. C.Z. was
supported by a Medical Research Development Fellowship.
Notes
We thank Siew-lan Ang, Masato Nakafuku, David Anderson, Flora
Vaccarino, and Grace Panganiban for generously providing reagents.
We thank Nicole Gruenig and Laidlaw Green for technical assistance
with animal maintenance and genotyping, Dawn Zinyk for help
preparing figures, and Deborah Kurrasch for critical reading of the
manuscript. Conflict of Interest : None declared.
References
Abellan A, Menuet A, Dehay C, Medina L, Retaux S. 2009. Differential
expression of LIM-homeodomain factors in Cajal-Retzius cells of
primates, rodents, and birds. Cereb Cortex. 20:1788--1798.
Alam S, Zinyk D, Ma L, Schuurmans C. 2005. Members of the Plag gene
family are expressed in complementary and overlapping regions in
the developing murine nervous system. Dev Dyn. 234:772--782.
Alcantara S, Ruiz M, D’Arcangelo G, Ezan F, de Lecea L, Curran T,
Sotelo C, Soriano E. 1998. Regional and cellular patterns of reelin
mRNA expression in the forebrain of the developing and adult
mouse. J Neurosci. 18:7779--7799.
Baas D, Bumsted KM, Martinez JA, Vaccarino FM, Wikler KC,
Barnstable CJ. 2000. The subcellular localization of Otx2 is cell-
type specific and developmentally regulated in the mouse retina.
Brain Res Mol Brain Res. 78:26--37.
Backman M, Machon O, Mygland L, van den Bout CJ, Zhong W,
Taketo MM, Krauss S. 2005. Effects of canonical Wnt signaling on
dorso-ventral specification of the mouse telencephalon. Dev Biol.
279:155--168.
Bielle F, Griveau A, Narboux-Neme N, Vigneau S, Sigrist M, Arber S,
Wassef M, Pierani A. 2005. Multiple origins of Cajal-Retzius cells
at the borders of the developing pallium. Nat Neurosci. 8:
1002--1012.
Borello U, Pierani A. 2010. Patterning the cerebral cortex: traveling with
morphogens. Curr Opin Genet Dev. 20:408--415.
Borrell V, Del Rio JA, Alcantara S, Derer M, Martinez A, D’Arcangelo G,
Nakajima K, Mikoshiba K, Derer P, Curran T, et al. 1999. Reelin
regulates the development and synaptogenesis of the layer-
specific entorhino-hippocampal connections. J Neurosci. 19:
1345--1358.
Britz O, Mattar P, Nguyen L, Langevin LM, Zimmer C, Alam S,
Guillemot F, Schuurmans C. 2006. A role for proneural genes in
the maturation of cortical progenitor cells. Cereb Cortex.
16:i138--i151.
Bulchand S, Grove EA, Porter FD, Tole S. 2001. LIM-homeodomain gene
Lhx2 regulates the formation of the cortical hem. Mech Dev.
100:165--175.
Casarosa S, Fode C, Guillemot F. 1999. Mash1 regulates neurogenesis in
the ventral telencephalon. Development. 126:525--534.
Caviness VS, Jr. 1982. Neocortical histogenesis in normal and reeler
mice: a developmental study based upon [3H]thymidine autoradi-
ography. Brain Res. 256:293--302.
Chameau P, Inta D, Vitalis T, Monyer H, Wadman WJ, van Hooft JA.
2009. The N-terminal region of reelin regulates postnatal dendritic
maturation of cortical pyramidal neurons. Proc Natl Acad Sci U S A.
106:7227--7232.
Cubelos B, Sebastian-Serrano A, Kim S, Redondo JM, Walsh C, Nieto M.
2008. Cux-1 and Cux-2 control the development of Reelin expressing
cortical interneurons. Dev Neurobiol. 68:917--925.
del Rio JA, Martinez A, Fonseca M, Auladell C, Soriano E. 1995.
Glutamate-like immunoreactivity and fate of Cajal-Retzius cells in
the murine cortex as identified with calretinin antibody. Cereb
Cortex. 5:13--21.
Fode C, Ma Q, Casarosa S, Ang SL, Anderson DJ, Guillemot F. 2000. A
role for neural determination genes in specifying the dorsoventral
identity of telencephalic neurons. Genes Dev. 14:67--80.
Friocourt G, Kanatani S, Tabata H, Yozu M, Takahashi T, Antypa M,
Raguenes O, Chelly J, Ferec C, Nakajima K, et al. 2008. Cell-
autonomous roles of ARX in cell proliferation and neuronal
migration during corticogenesis. J Neurosci. 28:5794--5805.
Galceran J, Miyashita-Lin EM, Devaney E, Rubenstein JL, Grosschedl R.
2000. Hippocampus development and generation of dentate gyrus
granule cells is regulated by LEF1. Development. 127:469--482.
Gohlke JM, Armant O, Parham FM, Smith MV, Zimmer C, Castro DS,
Nguyen L, Parker JS, Gradwohl G, Portier CJ, et al. 2008.
Characterization of the proneural gene regulatory network during
mouse telencephalon development. BMC Biol. 6:15.
Gong S, Zheng C, Doughty ML, Losos K, Didkovsky N, Schambra UB,
Nowak NJ, Joyner A, Leblanc G, Hatten ME, et al. 2003. A gene
expression atlas of the central nervous system based on bacterial
artificial chromosomes. Nature. 425:917--925.
Gorski JA, Talley T, Qiu M, Puelles L, Rubenstein JL, Jones KR. 2002.
Cortical excitatory neurons and glia, but not GABAergic neurons,
are produced in the Emx1-expressing lineage. J Neurosci. 22:
6309--6314.
Cerebral Cortex November 2011, V 21 N 11 2609Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

Griveau A, Borello U, Causeret F, Tissir F, Boggetto N, Karaz S, Pierani A.
2010. A novel role for Dbx1-derived Cajal-Retzius cells in early
regionalization of the cerebral cortical neuroepithelium.. PLoS Biol.
8:e1000440.
Grove EA, Tole S, Limon J, Yip L, Ragsdale CW. 1998. The hem of the
embryonic cerebral cortex is defined by the expression of multiple
Wnt genes and is compromised in Gli3-deficient mice. Develop-
ment. 125:2315--2325.
Gutin G, Fernandes M, Palazzolo L, Paek H, Yu K, Ornitz DM,
McConnell SK, Hebert JM. 2006. FGF signalling generates ventral
telencephalic cells independently of SHH. Development. 133:
2937--2946.
Hammond V, So E, Gunnersen J, Valcanis H, Kalloniatis M, Tan SS. 2006.
Layer positioning of late-born cortical interneurons is dependent on
Reelin but not p35 signaling. J Neurosci. 26:1646--1655.
Hanashima C, Fernandes M, Hebert JM, Fishell G. 2007. The role of
Foxg1 and dorsal midline signaling in the generation of Cajal-Retzius
subtypes. J Neurosci. 27:11103--11111.
Hanashima C, Li SC, Shen L, Lai E, Fishell G. 2004. Foxg1 suppresses
early cortical cell fate. Science. 303:56--59.
Hand R, Bortone D, Mattar P, Nguyen L, Heng JI, Guerrier S, Boutt E,
Peters E, Barnes AP, Parras C, et al. 2005. Phosphorylation of
Neurogenin2 specifies the migration properties and the dendritic
morphology of pyramidal neurons in the neocortex. Neuron.
48:45--62.
Hevner RF, Neogi T, Englund C, Daza RA, Fink A. 2003. Cajal-Retzius
cells in the mouse: transcription factors, neurotransmitters, and
birthdays suggest a pallial origin. Brain Res Dev Brain Res.
141:39--53.
Hevner RF, Shi L, Justice N, Hsueh Y, Sheng M, Smiga S, Bulfone A,
Goffinet AM, Campagnoni AT, Rubenstein JL. 2001. Tbr1
regulates differentiation of the preplate and layer 6. Neuron.
29:353--366.
Horton S, Meredith A, Richardson JA, Johnson JE. 1999. Correct
coordination of neuronal differentiation events in ventral
forebrain requires the bHLH factor MASH1. Mol Cell Neurosci.
14:355--369.
Imayoshi I, Shimogori T, Ohtsuka T, Kageyama R. 2008. Hes genes and
neurogenin regulate non-neural versus neural fate specification in
the dorsal telencephalic midline. Development. 135:2531--2541.
Janusonis S, Gluncic V, Rakic P. 2004. Early serotonergic projections to
Cajal-Retzius cells: relevance for cortical development. J Neurosci.
24:1652--1659.
Kuschel S, Ruther U, Theil T. 2003. A disrupted balance between Bmp/
Wnt and Fgf signaling underlies the ventralization of the Gli3 mutant
telencephalon. Dev Biol. 260:484--495.
Langevin LM, Mattar P, Scardigli R, Roussigne M, Logan C, Blader P,
Schuurmans C. 2007. Validating in utero electroporation for the
rapid analysis of gene regulatory elements in the murine telen-
cephalon. Dev Dyn. 236:1273--1286.
Lee SM, Tole S, Grove E, McMahon AP. 2000. A local Wnt-3a signal is
required for development of the mammalian hippocampus. De-
velopment. 127:457--467.
Mallamaci A, Mercurio S, Muzio L, Cecchi C, Pardini CL, Gruss P,
Boncinelli E. 2000. The lack of Emx2 causes impairment of Reelin
signaling and defects of neuronal migration in the developing
cerebral cortex. J Neurosci. 20:1109--1118.
Mattar P, Langevin LM, Markham K, Klenin N, Shivji S, Zinyk D,
Schuurmans C. 2008. Basic helix-loop-helix transcription factors
cooperate to specify a cortical projection neuron identity. Mol Cell
Biol. 28:1456--1469.
Meyer G, Cabrera Socorro A, Perez Garcia CG, Martinez Millan L,
Walker N, Caput D. 2004. Developmental roles of p73 in Cajal-
Retzius cells and cortical patterning. J Neurosci. 24:9878--9887.
Meyer G, Perez-Garcia CG, Abraham H, Caput D. 2002. Expression of
p73 and Reelin in the developing human cortex. J Neurosci.
22:4973--4986.
Miquelajauregui A, Varela-Echavarria A, Ceci ML, Garcia-Moreno F,
Ricano I, Hoang K, Frade-Perez D, Portera-Cailliau C, Tamariz E, De
Carlos JA, et al. 2010. LIM-homeobox gene Lhx5 is required for
normal development of Cajal-Retzius cells. J Neurosci. 30:
10551--10562.
Monuki ES, Porter FD, Walsh CA. 2001. Patterning of the dorsal
telencephalon and cerebral cortex by a roof plate-Lhx2 pathway.
Neuron. 32:591--604.
Morante-Oria J, Carleton A, Ortino B, Kremer EJ, Fairen A, Lledo PM.
2003. Subpallial origin of a population of projecting pioneer
neurons during corticogenesis. Proc Natl Acad Sci U S A.
100:12468--12473.
Muzio L, Mallamaci A. 2005. Foxg1 confines Cajal-Retzius neurono-
genesis and hippocampal morphogenesis to the dorsomedial
pallium. J Neurosci. 25:4435--4441.
Nieto M, Schuurmans C, Britz O, Guillemot F. 2001. Neural bHLH genes
control the neuronal versus glial fate decision in cortical progen-
itors. Neuron. 29:401--413.
Nishikawa S, Goto S, Hamasaki T, Yamada K, Ushio Y. 2002. Involvement
of reelin and Cajal-Retzius cells in the developmental formation of
vertical columnar structures in the cerebral cortex: evidence from
the study of mouse presubicular cortex. Cereb Cortex. 12:
1024--1030.
Parras CM, Hunt C, Sugimori M, Nakafuku M, Rowitch D, Guillemot F.
2007. The proneural gene Mash1 specifies an early population of
telencephalic oligodendrocytes. J Neurosci. 27:4233--4242.
Pla R, Borrell V, Flames N, Marin O. 2006. Layer acquisition by cortical
GABAergic interneurons is independent of Reelin signaling. J
Neurosci. 26:6924--6934.
Saito T, Nakatsuji N. 2001. Efficient gene transfer into the embryonic
mouse brain using in vivo electroporation. Dev Biol. 240:237--246.
Schiffmann SN, Bernier B, Goffinet AM. 1997. Reelin mRNA expression
during mouse brain development. Eur J Neurosci. 9:1055--1071.
Schuurmans C, Armant O, Nieto M, Stenman JM, Britz O, Klenin N,
Brown C, Langevin LM, Seibt J, Tang H, et al. 2004. Sequential
phases of cortical specification involve neurogenin-dependent and
-independent pathways. EMBO J. 23:2892--2902.
Shimogori T, Banuchi V, Ng HY, Strauss JB, Grove EA. 2004. Embryonic
signaling centers expressing BMP, WNT and FGF proteins interact
to pattern the cerebral cortex. Development. 131:5639--5647.
Shinozaki K, Miyagi T, Yoshida M, Miyata T, Ogawa M, Aizawa S, Suda Y.
2002. Absence of Cajal-Retzius cells and subplate neurons associ-
ated with defects of tangential cell migration from ganglionic
eminence in Emx1/2 double mutant cerebral cortex. Development.
129:3479--3492.
Siegenthaler JA, Miller MW. 2008. Generation of Cajal-Retzius neurons
in mouse forebrain is regulated by transforming growth factor beta-
Fox signaling pathways. Dev Biol. 313:35--46.
Stoykova A, Hatano O, Gruss P, Gotz M. 2003. Increase in reelin-positive
cells in the marginal zone of Pax6 mutant mouse cortex. Cereb
Cortex. 13:560--571.
Studer M, Filosa A, Rubenstein JL. 2005. The nuclear receptor COUP-TFI
represses differentiation of Cajal-Retzius cells. Brain Res Bull.
66:394--401.
Takahashi T, Goto T, Miyama S, Nowakowski RS, Caviness VS, Jr. 1999.
Sequence of neuron origin and neocortical laminar fate: relation to
cell cycle of origin in the developing murine cerebral wall. J
Neurosci. 19:10357--10371.
Takiguchi-Hayashi K, Sekiguchi M, Ashigaki S, Takamatsu M, Hasegawa H,
Suzuki-Migishima R, Yokoyama M, Nakanishi S, Tanabe Y. 2004.
Generation of reelin-positive marginal zone cells from the caudome-
dial wall of telencephalic vesicles. J Neurosci. 24:2286--2295.
Talikka M, Perez SE, Zimmerman K. 2002. Distinct patterns of
downstream target activation are specified by the helix-loop-helix
domain of proneural basic helix-loop-helix transcription factors.
Dev Biol. 247:137--148.
Tissir F, Ravni A, Achouri Y, Riethmacher D, Meyer G, Goffinet AM.
2009. DeltaNp73 regulates neuronal survival in vivo. Proc Natl Acad
Sci U S A. 106:16871--16876.
von Frowein J, Wizenmann A, Gotz M. 2006. The transcription
factors Emx1 and Emx2 suppress choroid plexus development and
promote neuroepithelial cell fate. Dev Biol. 296:239--252.
Waclaw RR, Wang B, Pei Z, Ehrman LA, Campbell K. 2009. Distinct
temporal requirements for the homeobox gene Gsx2 in
2610 Ascl1 Function in Cajal--Retzius Cells d Dixit et al.Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018

specifying striatal and olfactory bulb neuronal fates. Neuron.
63:451--465.
Wagener RJ, David C, Zhao S, Haas CA, Staiger JF. 2010. The
somatosensory cortex of reeler mutant mice shows absent layering
but intact formation and behavioral activation of columnar somato-
topic maps. J Neurosci. 30:15700--15709.
Wang B, Waclaw RR, Allen ZJ 2nd, Guillemot F, Campbell K. 2009. Ascl1
is a required downstream effector of Gsx gene function in the
embryonic mouse telencephalon. Neural Dev. 4:5.
Wood JG, Martin S, Price DJ. 1992. Evidence that the earliest generated
cells of the murine cerebral cortex form a transient population in
the subplate and marginal zone. Brain Res Dev Brain Res. 66:
137--140.
Yabut O, Renfro A, Niu S, Swann JW, Marin O, D’Arcangelo G. 2007.
Abnormal laminar position and dendrite development of interneur-
ons in the reeler forebrain. Brain Res. 1140:75--83.
Yoshida M, Assimacopoulos S, Jones KR, Grove EA. 2006. Massive loss of
Cajal-Retzius cells does not disrupt neocortical layer order. De-
velopment. 133:537--545.
Zimmer C, Lee J, Griveau A, Arber S, Pierani A, Garel S, Guillemot F.
2010. Role of Fgf8 signalling in the specification of rostral
Cajal-Retzius cells. Development. 137:293--302.
Cerebral Cortex November 2011, V 21 N 11 2611Downloaded from https://academic.oup.com/cercor/article-abstract/21/11/2599/277473by gueston 29 March 2018





























