Embed Size (px)
Citation preview

J. Cell Sd. 56, 263-279 (1982) 263Printed in Great Britain © Company of Biologists Limited 1982
ASCOSPORE DEVELOPMENT IN THE
FISSION YEASTS SCHIZOSACCHAROMYCES
POMBE AND S. JAPONICUS
KENJI TANAKA* AND AIKO HIRATAInstitute of Applied Microbiology, The University of Tokyo,Bunkyo-ku, Tokyo 113, Japan
SUMMARY
The fine structure of ascospore formation in the fission yeasts Schizosaccharomyces pombeand Schizosaccharomyces japonicus var. japonicus was studied by serial thin-sectioning andelectron microscopy. The morphogenetic events were almost the same in both species.Ascospore development was initiated by the formation of the forespore membrane on thecytoplasmic side of the differentiated nucleus-associated organelle (NAO) in the intervalbetween meiosis I and II in S. pombe, or during the post-meiotic nuclear division in S. japonicus,and the process proceeded almost synchronously through the two or four nuclei in the ascus.The forespore membrane developed by fusion of the cytoplasmic vesicles and this was clearlydemonstrated in S. japonicus where the behaviour of vesicles involved in the foresporemembrane development could be traced as they were marked by the presence of electron-densegranules. The staining technique, by phosphotungustic acid-chromic acid (PTA-CA) aftertreatment with periodic acid, was used to attempt to elucidate the origin and the nature ofthe forespore membrane. The method specific to plasmalemma-type membranes stained bothascus and ascospore plasmalemmas; the forespore membrane was not stained at first butdeveloped the same affinity for stain as the plasma membrane in the course of ascosporedevelopment. The results suggest that the forespore membrane did not come directly fromthe ascus plasma membrane, but from another membrane system such as the endoplasmicreticulum. Spore wall material was deposited in the space between the inner and outer leafletsof the forespore membrane.
INTRODUCTION
Ascospore development in yeasts seems to be one of the most advantageous systemsfor the study of cell differentiation. Ascospore formation in bakers' yeast involveszygote formation by conjugation between sexually compatible cells, meiotic divisionsand ascospore development in the ascus. Ascospore delimitation in Saccharomycescerevisiae has been observed to be initiated at the site of spindle-pole bodies ornucleus-associated organelles (NAOs) of the second meiotic spindles, by the develop-ment of a pair of unit membranes (Moens & Rapport, 1971; Peterson, Gray & Ris,1972), between which the spore wall is subsequently laid down (Lynn & Magee,1970; Beckett, Illingworth & Rose, 1973). However, the origin of the ascospore-delimiting membrane has not been clarified.
• Present address: Institute of Medical Mycology, Nagoya University School of Medicine,Showa-ku, Nagoya 466, Japan.

264 K. Tanaka and A. Hirata
The fission yeast Schizosaccharomyces pombe has been used widely for cell-cycleand genetic studies, and the ultrastructure of ascospore formation also has beenstudied by several investigators (Conti & Naylor, i960; Yoo, Calleja & Johnson,1973). They have established the basic outline of events in the process leading tothe formation of ascospores, but have not revealed any details of the nuclear structuresin the process, because they used fixation with permanganate exclusively for studiesof the membrane structures. Olson, Eden, Egel-Mitani & Egel (1978) have providedelectron micrographs of sections of S. pombe nuclei undergoing meiosis. Our ownexperience of early phases of this process has been similar (Hirata & Tanaka, 1982)and the conclusions have been incorporated into Fig. 1.
This paper describes the ultrastructural details of the origin and formation of theascospore-delimiting membranes of glutaraldehyde-osmium tetroxide fixed cellsduring ascosporogenesis in S. pombe and in Schizosaccharomyces japonicus var.japonicus as demonstrated by serial thin-sectioning and electron microscopy.
MATERIALS AND METHODS
Organisms and culture for sporulation
S. pombe strain I190 a homothallic strain and S. japonicus Yukawa et Maki var. japonicusIFO 1713 were used in this study.
For the sporulation of S. pombe, cells were cultured in presporulation medium, whichcontained 3 % glucose and 0-5 % yeast extract, and harvested at the early stationary phaseof growth. After washing once in distilled water, cells were inoculated into 100 ml of sporulationmedium (SSL) in conical flasks at a density of about 5 x 10' cells per ml (Egel, 1971). Theywere shaken at 30 °C. In these conditions sporulation was completed within 24 h.
Sporulation of S. japonicus was brought about by following the method described byTsuboi, Ohashi, Takahara & Hayashibe (1978). Modified Burkholder medium (B medium)supplemented with 0̂ 5 % yeast extract and 0-5 % malt extract (BYM medium) was preparedfor the presporulation culture, which was kept in the dark to avoid light-induced sexualflocculation. Sporulation was carried out in B medium, from which (NH4),SO4 was omitted(B — N medium), in the light with shaking, at 26 °C.
Electron microscopy
For electron microscopy aliquots were taken at intervals from the sporulation culture, andcvi vol. of glutaraldehyde fixative was added before centrifugation. Centrifuged cells wereresuspended in fresh fixative at room temperature for 2 to 3 h, and then stored in a refrigerator.The fixative was a 3 % glutaraldehyde solution in potassium phosphate buffer (pH 7-0). Fixed
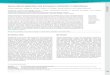
Fig. 1. A diagrammatic representation of meiosis and the development of ascosporesin S. pombe and S. japonicus var. japonicus. A to D are common to the two species.A. A zygote with two nuclei (dotted), each associated with a single NAO. B. Fusionof the two nuclei is preceded by fusion of both NAOs. c. A fused and elongatednucleus, with a fused NAO at one end containing a number of linear elements, whichis thought to be a stage of meiotic prophase. D. Meiosis I nucleus with a spindle(parallel lines) between a pair of NAOs. E and F, from S. pombe. E. Foresporemembranes of double membrane sacs originating on the cytoplasmic sides of theNAOs cover the half-spindles of meiosis II in the two nuclei. F. Four ascosporeswith thick walls. Spore nuclei carry their respective NAOs. c, H and 1, fromS. japonicus. c. Two nuclei with spindles at meiosis II in the ascus. H. Foresporemembranes developing on the differentiated NAOs of the post-meiotic spindles offour nuclei. Development seems to proceed synchronously through the four nuclei.1. An eight-ascospore ascus.
Ascospore development in fission yeast
A
265
Fig. 1
266 K. Tanaka and A. Hirata
fm
or
V.30L1
' • %
c

Ascospore development in fission yeast 267
cells were washed several times in buffer, and treated with Zymolyase 60000 (Kirin BreweryCo. Ltd, Takasaki, Japan) (o-i mg/ml) in the same buffer at 30 °C. The disintegration of thecell wall was examined with a phase-contrast microscope. The Zymolyase-treated cells werewashed again in buffer and postfixed in 2 % OsO4 for 2 h at room temperature. After washingin distilled water, they were soaked in a 0-5 % (w/v) aqueous solution of uranyl acetate for2 h and embedded in agar blocks. The cells were dehydrated by passing them through a seriesof increasing concentrations of ethanol and absolute acetone, and then they were embeddedin Spurr's resin. Serial sections were obtained with a Dupont diamond knife on a Porter-Blum MT-2 ultramicrotome and treated by one of the following methods. (1) Sections weremounted on Formvar-coated single-slot grids and stained with uranyl acetate and lead citrate.(2) In order to apply specific staining to the plasmalemma, sections were collected usinga small plastic sheet with a hole (3-0 mm diameter) punched through it at one end. Sectionsfloating on a film of distilled water within the hole were treated with 1 % periodic acid andstained with phosphotungustic acid plus chromic acid (PTA-CA) (Roland, Lembi & Morr6,1972). Sections were viewed with a JEOL 200 CX electron microscope at 100 kV.
RESULTS
Ascospore formation in the fission yeast involves the developmental stages of cellflocculation, cell fusion, karyogany, meiotic nuclear divisions, ascospore delimitation bythe development of double unit membranes or forespore membrane, and spore wallmaturation. The morphological changes in the meiotic nuclei and the process leading toascospore formation were almost the. same in 5. pombe (Hirata & Tanaka, 1982) and S.japonicus (Hirata & Tanaka, unpublished observations), except that the latter underwentone additional nuclear division before ascospore delimitation began. This is illustratedin Fig. 1. Conjugation of the two haploid cells resulted in a binucleate zygote (A)where karyogamy had occurred by fusion of the two nucleus-associated organelles(NAOs) (B). At the presumed prophase in meiosis I, the zygote nucleus with a fusedNAO at one end was elongated and contained a number of filamentous linearelements (c) (Olson et al. 1978). Nuclear division was effected by a spindle consistingof microtubules connecting NAOs at opposite poles (D). In 5. pombe ascosporedelimitation first became apparent at the start of meiosis II (E) and a four-sporedascus was produced (F). In 5. japonicus this event is delayed because in this speciesmeiosis is followed by another round of nuclear division (G, H). The primary envelopesfor the eight spores about to be created begin to be formed simultaneously near theNAOs at the spindle poles of the dividing haploid nuclei even before these mitoseshave been completed (H, I).
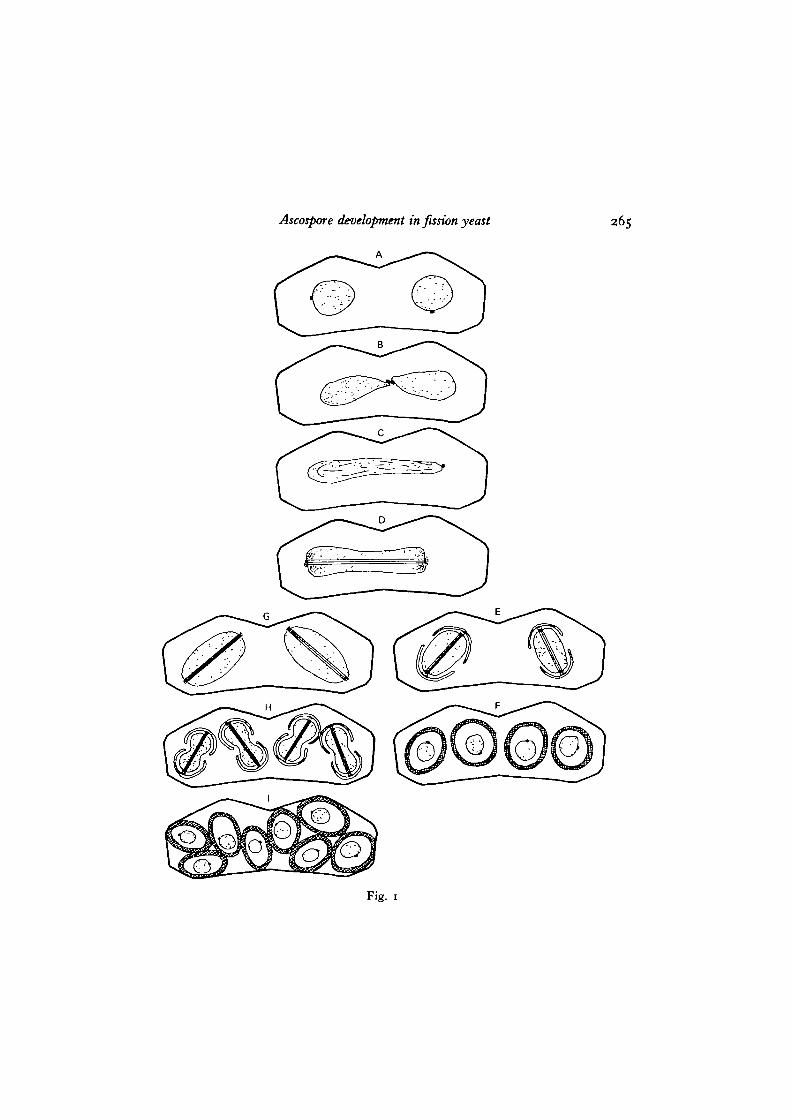
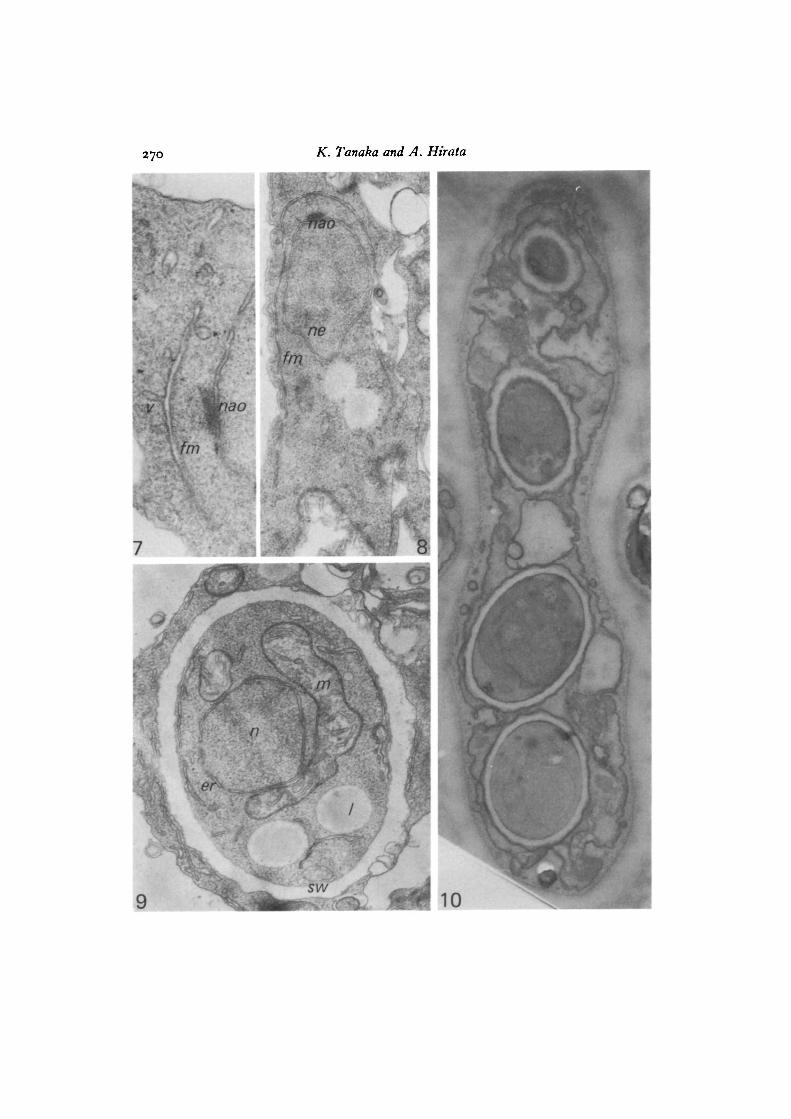
Figs. 2-10. S. pombe.Fig. 2. A zygote at meiosis II. Two nuclei that are the products of meiosis I seem
to be at almost the same stage of meiotic division as shown by the presence ofspindle microtubules (arrow). This indicates that meiosis II is proceeding syn-chronously between the two nuclei, x 16coo.
Fig. 3. Two consecutive sections (A, B) show the multi-plaque structure of themodified NAO (nao) at meiosis II and the forespore membrane (fin) developingon it. Traces of spindle microtubules can be seen (mt). A is an enlarged view of oneof the two nuclei in the sections shown in Fig. 2. Associations of endoplasmicreticulum (er) and vacuolar membranes with forespore membrane can be seen,x 60000.
268 K. Tanaka and A. Hirata
ne
\
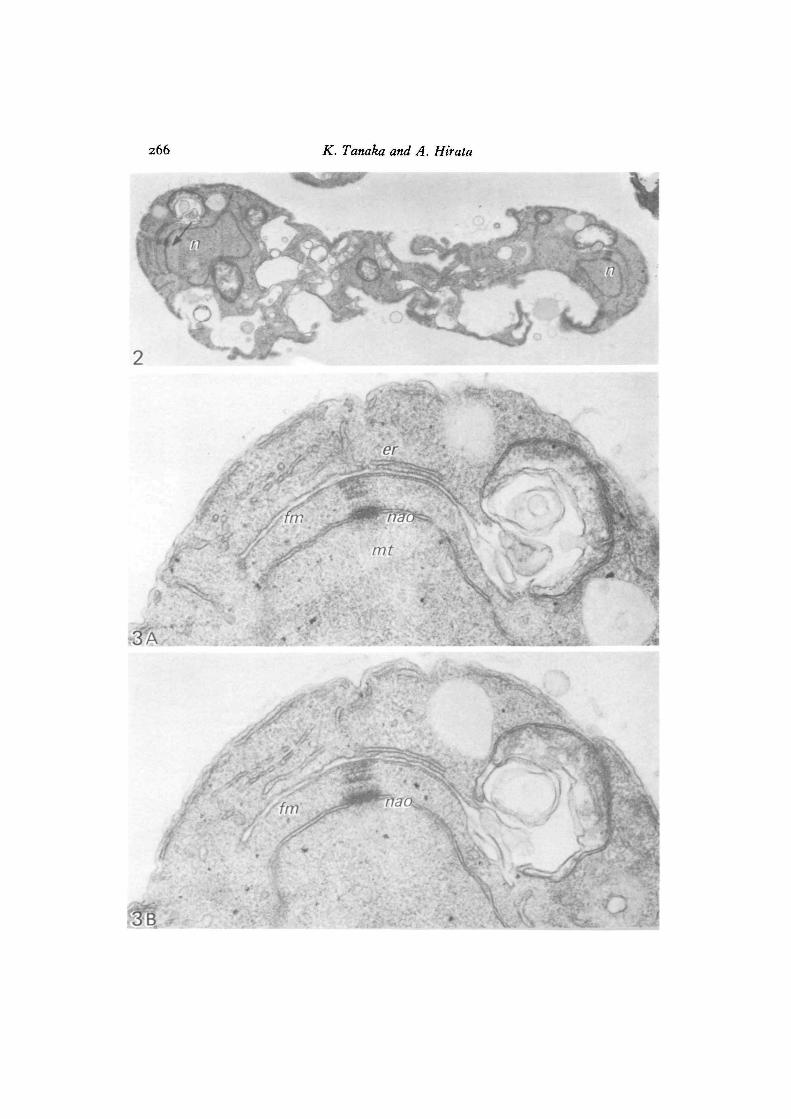
Fig. 4. An ascus stained with PTA-CA in which the two nuclei at meioais IIare associated with the developing forespore membrane. The ascus plasmalemma(pm) with large imaginations, and one segment of forespore membrane, are stainedpositively, x 12000.
Fig. 5. The left-hand nucleus in Fig. 4 at higher magnification. Forespore mem-branes (Jm) in the upper part are not stained, while those accompanying a fewvesicles in the lower part are stained to medium density. The nuclear envelope(ne) is not stained, pm, plasmalemma. x 42720.
Fig. 6. The right-hand nucleus in Fig. 4, enlarged, where one segment of foresporemembrane (Jm) is positively stained to the same extent as that of the ascus plasma-lemma (pm), while the other is definitely unstained (lower part, arrow), ne, nuclearenvelope, x 42720.

Ascospore development in fission yeast 269
S. pombe
The two products of meiosis I underwent meiosis II almost synchronously (Fig. 2).The NAO of the dense plaque embedded in the nuclear envelope of the meiosis Ispindle has now elaborated several layers of dense plaques, and the foresporemembrane is being developed immediately adjacent to the outermost plaque on thecytoplasmic side (Fig. 3 A, B). The forespore membranes at both NAOs might beinitiated at the same time and developed at the same rate, because serial-sectionanalysis has showed that the forespore membranes developed to a similar extenton both sides, concomitant with spindle elongation. The forespore membrane wassometimes connected to flattened vesicles or tubular reticulum, which was definitelyconnected to the smooth endoplasmic reticulum running along the plasmalemma(Fig. 3 A, B). Fusion of cytoplasmic vesicles with the forespore membrane can alsobe seen in Fig. 7. Fig. 8 shows a nucleus (meiosis being finished but the foresporemembrane not being closed) where the layered plaques of the meiosis II NAO areno longer clear and have dedifferentiated into a single dense NAO on the nuclearenvelope.
When sections of the ascospore-forming ascus were subjected to PTA-CA staining,not all of the forespore membranes showed positive reactions. The ascus plasmalemmaand membranes or vesicles derived from it were stained positively, but the nuclearenvelope, endoplasmic reticulum and mitochondrial membrane were not stained(Fig. 4). The forespore membranes surrounding both nuclei at meiosis II seemedto be developed to a similar extent, but they were not uniformly stained by PTA-CA.The forespore membranes surrounding one of the meiotic nuclei were only faintlystained (Fig. 5), while those of the other nucleus were asymmetrically stained; oneleaf of the membranes being stained positively and the other negatively (Fig. 6).
Wall material was deposited between the double unit membranes; thus the innermembrane facing the nucleus constituted the plasmalemma of the ascospore proto-plasm. The nucleus, mitochondria, endoplasmic reticula, lipid granules and othergranules were enclosed in the ascospore (Fig. 9). In mature asci with four ascospores,PTA-CA clearly stained the plasmalemmas of both ascus and ascospores as well asthe outer membranes or spore coats of the ascospores, while membranes of thecytoplasmic organelles were left unstained (Fig. 10).
S. japonicus var. japonicus
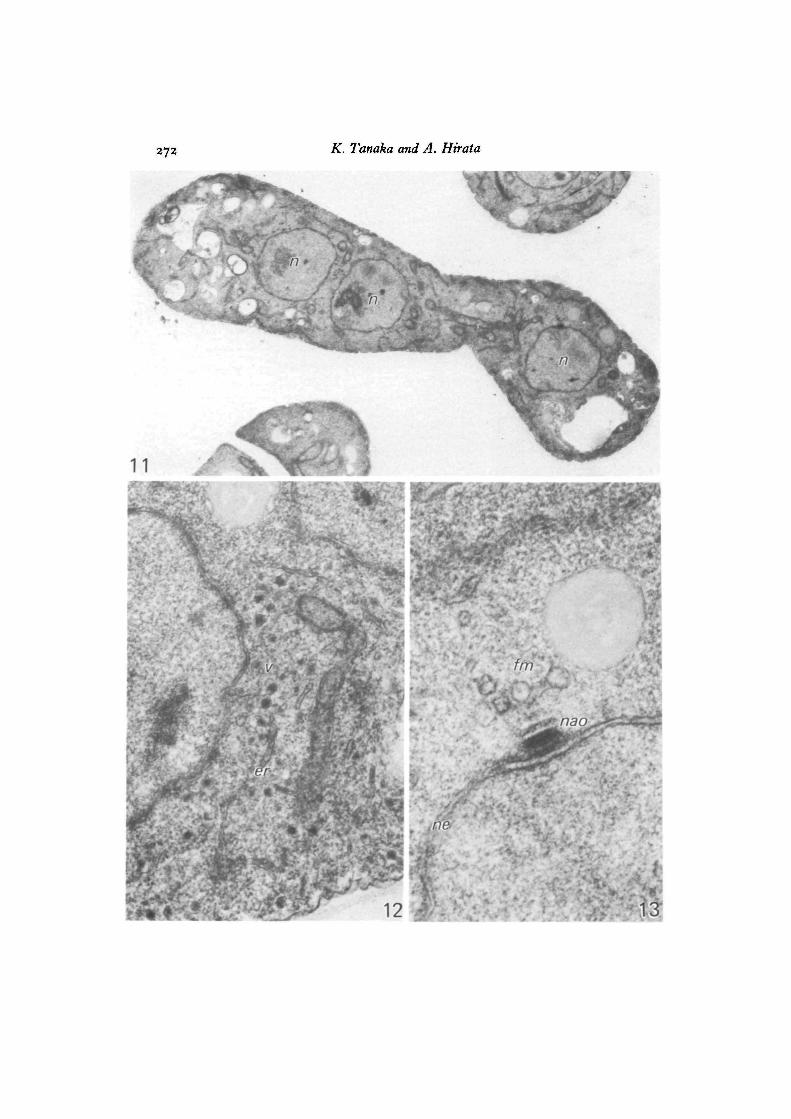
Ascospore formation in S. japonicus began in the ascus that contained four nucleiafter two successive meiotic divisions (Fig. 1). The four-nuclei ascus contained alarge number of dense granules in the cytoplasm (Fig. 11), which were enclosedin vesicles 50-80 nm in diameter and seen abundantly in the region of the smoothendoplasmic reticulum (Fig. 12). The nuclei bore differentiated NAOs of multi-layered plaques, in the proximity of which a few cytoplasmic vesicles were seen toaggregate to form incipient forespore membranes (Fig. 13). Fig. 14 shows the lastor post-meiotic nuclear division by a spindle consisting of microtubules that connectopposite NAOs. No part of the nuclear envelope was disrupted. Forespore membranes
270 K. Tanaka and A. Hirata

Ascospore development in fission yeast 271
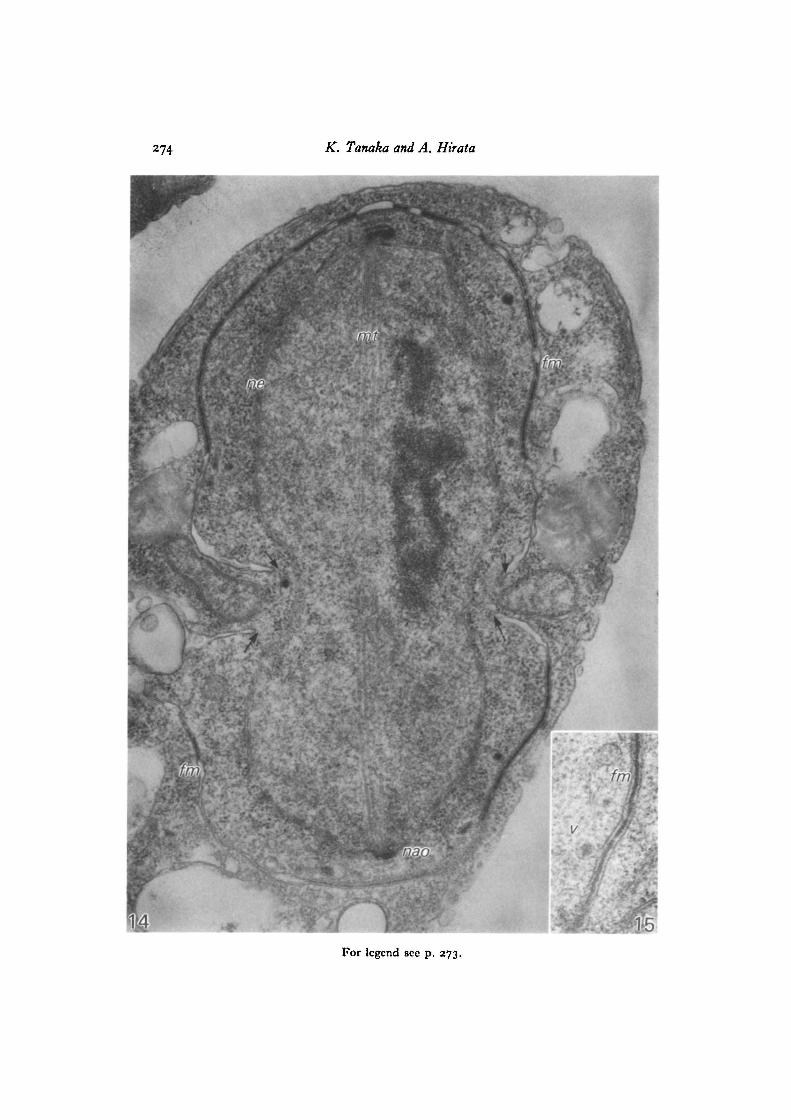
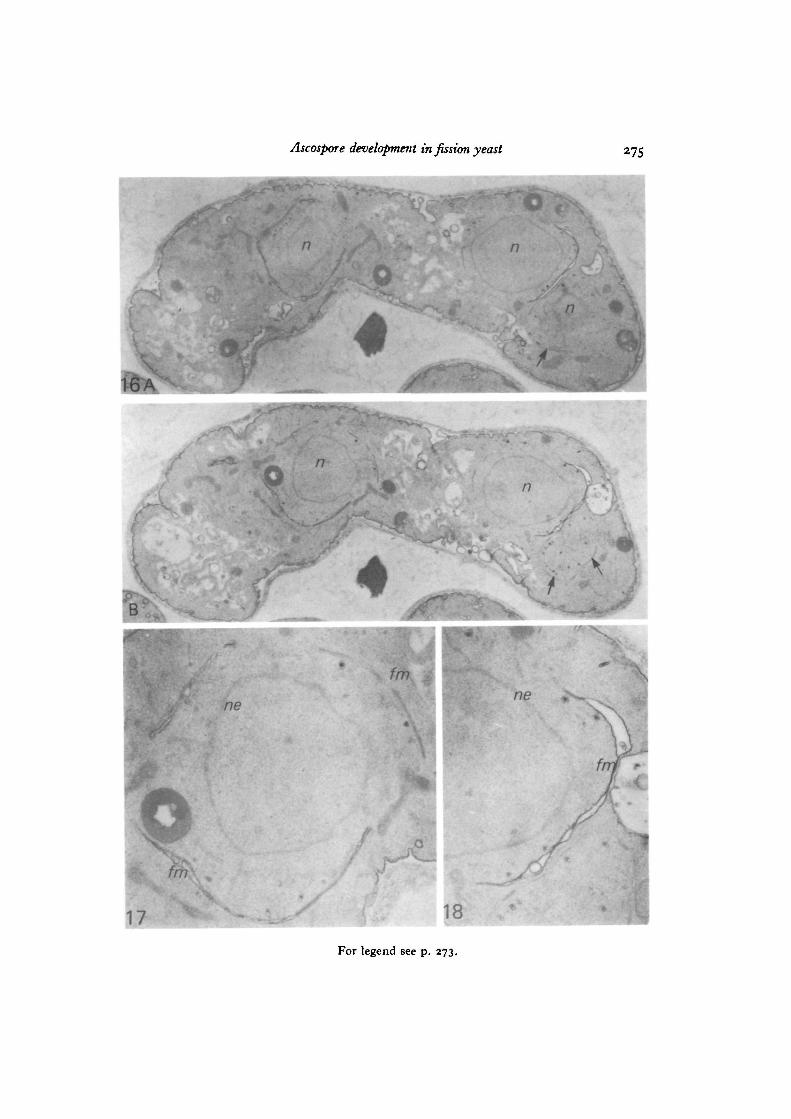
formed on the outermost plaque of the modified NAOs at opposite poles seemed toenvelop each half-spindle to a similar extent. Pairs of unit membranes of the fore-spore membrane were usually closely apposed to one another and each pair wasseparated by a space about 27 nm in diameter, in the middle of which electron-densematerial formed a distinct layer. In places, particularly at the advancing or growingfront of the forespore membrane, double membranes were separated giving theappearance of a flattened sac. Cytoplasmic vesicles carrying dense material weremostly found in the cytoplasm between the nuclear envelope and the foresporemembrane (Figs. 14, 16). One such vesicles can be seen apparently in the process offusion with the inner leaf of the forespore membrane, thus adding its share to thedense matter that at this time begins to accumulate in the space between the twoleaves of the forespore membrane (Fig. 15). We conjecture that the numerous vesicleswith dark centres seen in the vicinity of developing forespore membranes in Figs. 16and 17 have the same function, since Johnson, Yoo & Calleja (1973) in their studyof cell plate growth in S. pombe have shown examples of ' dense round bodies' thatsuggest participation in that process.
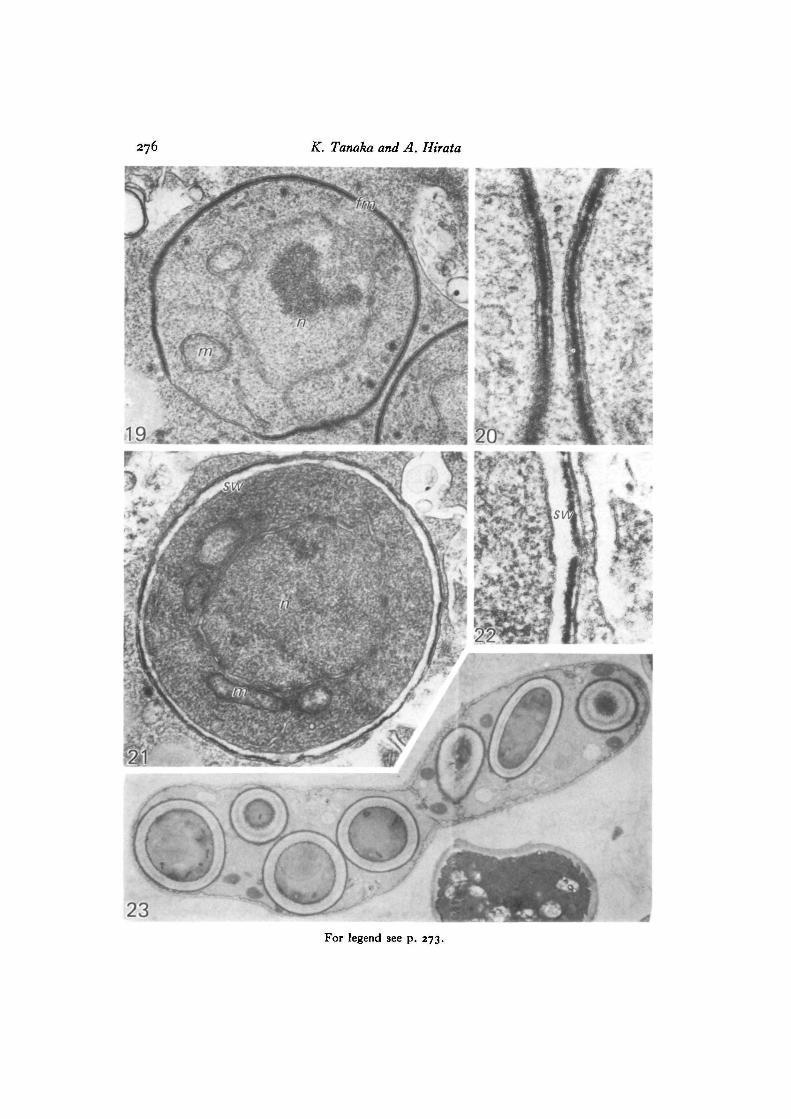
In order to determine the origin and nature of the cytoplasmic vesicle membraneand the forespore membrane, sections were treated by the PTA-CA method. Theresults were the same as in S. pombe, and two serial sections were presented todemonstrate the reliability of the staining technique (Fig. 16A, B). Membranes ofdense vesicles were in general not stained by PTA-CA, suggesting that their originwas in the cytoplasmic membrane system; the forespore membranes were notuniformly stained. Parts of the double membranes joined by a layer of dense materialwere not stained, while other parts with swollen intercisternal spaces were stained(Figs. 17, 18). The developing forespore membrane finally encircled the dividedascospore nucleus and a pair of unit membranes joined by a dense layer could beseen (Figs. 19, 20). Further maturation of the ascospore wall seemed to proceed bydeposition of wall material between the inner leaf of the forespore membrane andthe dense intermediate layer (Figs. 21, 22). A section of seven ascospores with athick spore wall within a matured eight-spored ascus is shown in Fig. 23, in whichthe ascus and ascospore plasmalemmas and the spore coat were definitely stainedby the PTA-CA method.
Fig. 7. Cytoplasmic vesicles (v) fusing with the forespore membrane (Jm). x 75coo.Fig. 8. The forespore membrane (Jm) fairly well developed, but not completely
encircling the nucleus, which has completed meiosis II. x 35000.Fig. 9. The spore wall (no) has developed between the two unit membranes of
the forespore membrane. Nucleus (n), mitochondria (mi), ER (er), lipid globule (/)and other vesicles are enclosed in the spore, x 35000.
Fig. 10. A four-spore ascus. The ascospores are not fully mature. The section isstained with PTA-CA, by which method ascus and spore plasma membranes aswell as the peripheries of the spore coat are stained, while nuclei and mitochondriawithin the spores are not stained, x 16200.
272 K. Tanaka and A. Hirata
11
ne
* • %
12 13

Ascospore development in fission yeast 273
Figs. 11-23. S.japonicus var. japonicus.Fig. 11. A zygote after meiosis II, three of the four nuclei (n) can be seen. Note
the occurrence of dense granules in the cytoplasm, x 35000.Fig. 12. Part of the cytoplasm of a cell at a stage similar to the one shown in
Fig. 11. Dense granules are enclosed in vesicles (v) that seem to be derived fromsmooth endoplasmic reticulum («•). x 41000.
Fig. 13. Part of the right-hand nucleus shown in Fig. n , enlarged. A few cyto-plasmic vesicles forming the initial forespore membrane (Jm) are seen close to theNAO (nao) on the nuclear envelope (ne), from which multi-plaques are differentiating,x 80000.
Fig. 14. A nucleus with a spindle at the post-meiotic division. The nuclearenvelope (ne) is preserved and spindle microtubules (mi) connect the modifiedNAOs at both poles. A pair of forespore membranes (Jm) are enveloping each half-spindle and seem to have developed to a similar extent on both sides (indicated byarrows). The two adjacent membranes of the forespore membrane are in mostregions joined by a layer of dense material and in places, particularly at the advancingfronts of the membranes, are swollen into flattened sacs. Vesicles carrying densegranules are seen exclusively in the cytoplasm enclosed by the developing foresporemembrane, x 41000.
Fig. 15. A cytoplasmic vesicle (v) earring dense material is fused with the innerleaf of the forespore membrane (fin), x 80000.
Fig. I6A, B. TWO serial sections (but with four intervening sections skipped) ofan ascus at the post-meiotic division stained by the PTA-CA method. Two setsof forespore membranes surrounding the two nuclei (n) and part of the foresporemembrane of the third nucleus (arrows on the right-hand side) can be seen, x 9770.
Fig. 17. The foreapore membranes (Jm) around the left-hand nucleus of Fig. I6B,enlarged. The regions of the forespore membrane joined to a dense layer are notstained, but those with a swollen cisternal space are stained, ne, nuclear envelope,x 27140.
Fig. 18. The forespore membrane (Jm) around the right-hand nucleus of Fig. I6B,enlarged. The swollen parts of the forespore membrane are stained, while most of themembranes of vesicles with dense granules and the nuclear envelope are not stained.ne, nuclear envelope, x 27 140.
Fig. 19. The foreapore membrane (Jm) is closed and includes a nucleus (n), mito-chondria (m) and other organelles. Most of the forespore membrane is filled withmaterial, x 40000.
Fig. 20. Part of the forespore membrane in the section adjacent to that shownin Fig. 19, enlarged. Pairs of unit membranes of the forespore membrane closelyapposed to one another; the space in the middle of each pair contains electron-densematerial forming a distinct layer, x 120000.
Fig. 21. A maturing ascospore. Wall material (no) seems to be deposited betweena layer of dense material and the inner leaf of the forespore membrane, x 40000.
Fig. 22. Higher magnification of part of Fig. 21. x 120000.Fig. 23. Seven ascospores in the eight-spore ascus are shown by PTA-CA staining.
Ascus, ascospore plasmalemmas and spore coat are distinctly stained, x 7360.
274 K. Tanaka and A. Hirata
fmj
X-
For legend see p. 273.
Ascospore development in fission yeast 275
n
n
t
n
n
\
17 18
For legend see p. 273.
276 K. Tanaka and A. Hirata
23For legend see p. 273.

Ascospore development in fission yeast 277
DISCUSSION
Ultrastructural changes in the fission yeast S. pombe during ascospore developmenthave been reported by seveial authors (Conti & Naylor, i960; Yoo et al. 1973), butthey were all results obtained on cells fixed with permanganate, which impaired thenucleoplasm and therefore gave information only on the membrane structure. Thispaper reports the fine structure of ascospore development in the two species ofSchizosaccharomyces fixed with glutaraldehyde and osmium tetroxide and attemptsto clarify cytochemically the origin of the ascospore wall by using PTA-CA staining.
Ascospore development began in the interval between meiosis I and II in S. pombe,producing four-spored asci, and during a post-meiotic division in S.japonicus, whereeight-spored asci were produced. The earliest morphological event in ascosporedevelopment was the appearance of the forespore membrane initially on the dif-ferentiating nucleus-associated organelle (NAO) of a multi-plaque structure, andthis was clearly demonstrated at the four-nuclei stage of S. japonicus (Fig. 13). Themulti-plaque structure of the NAO during the last nuclear division in ascosporedevelopment in the ascus was commonly observed among the ascomycetes. Ashton& Moens (1979), in discussing the taxonomic significance of the ultrastructure ofthe NAO, concluded that NAO structure was relatively consistent in higher asco-mycetes, while variety could be found in the hemiascomycetes. According to theircategorization of NAO morphology at meiosis II, the NAO of S. pombe may belongto type II, which consist of three plaques. This was also in accord with their un-published observations. It also seems to be a common feature among ascomycetesthat the forespore membrane or prospore wall is initiated as a disk-shaped vesicleon the cytoplasmic side of the NAO.
In ascospore development, it was established that ascospore wall material isdeposited between the double membrane envelope or forespore membrane, whichis derived from fusion of cytoplasmic vesicles. However, the origins of the vesiclesand the wall material remain unknown. Some authors claim that vesicles are producedby the imagination of the ascus plasmalemma (Bandoni, Bisalputra & Bisalputra,1967; Syrop & Beckett, 1972), while others have proposed that they are derivedby blebbing from nuclear membrane (Carroll, 1967; Oso, 1969; Yoo et al. 1973) orendoplasmic reticulum (ER) (Lynn & Magee, 1970; Black & Gorman, 1971). Theformer presumed that the plasmalemma-type characteristics of the membrane wouldbe conserved, while the latter assumed that there would be a change from ER- toplasmalemma-type during ascospore development, considering that the inner mem-brane of the double forespore membrane would constitute the ascospore plasma-lemma. In this study we made observations from which we conclude that the foresporemembrane was derived from the fusion of vesicles that originated from endoplasmicreticulum: in S. pombe the forespore membrane was observed in direct continuitywith the ER and in S. japonicus the behaviour of vesicles carrying dense materialpresented more direct evidence for the origin of the forespore membrane.
Syrop & Beckett (1972) demonstrated the membrane continuity, by showing thatthe spore-delimiting membranes were formed by the invagination of the ascus

278 K. Tanaka and A. Hirata
plasmalemma, using the PTA-CA method, which preferentially stains plasmalemma-type membrane. When the same technique was applied in this study to ascosporedevelopment in the fission yeast, results were not consistent enough to concludethat the double membrane had a single origin. We found some vesicles stainedwith PTA-CA near the unstained forespore membrane, which was stained withmedium density as development proceeded and eventually very densely as it formedthe ascospore plasmalemma. The conclusion reached was that the forespore mem-brane with ER characteristics at the early stage of its development might changeprogressively to acquire plasmalemma characteristics, this process of membranemodification possibly being facilitated chemically or by adding plasmalemma-derived vesicles to the forespore membrane. This process of chemical modificationseemed to progress asynchronously between the separate nuclei in an ascus, whilemorphological development went on synchronously.
Ascospore wall material was thought to be deposited between the double leafletsof the forespore membrane, but where it was synthesized and how it was depositedare not known. It was clearly demonstrated, at least in S. japonicus, that part of thewall material was carried by the vesicles fused with the forespore membrane, as wasshown by the behaviour of the dense material carried by the vesicles. Thickening ofthe ascospore wall seemed to occur after the delimitation of the ascospore wascomplete, and in S. japonicus this seemed to proceed between the layer of densematerial and the inner leaf of forespore membrane. The synthesis and mode ofdeposition of the wall material require further investigation.
This work was supported in a part by grants nos. 344001 and 56470127 from the Ministryof Education, Science and Culture, Japan.
REFERENCES
ASHTON, M. L. & MOENS, P. B. (1979). Ultrastructure of sporulation in the hemiascomycetesAtcoidea corymbosa, A. rubescent, Cephaloascus fragrans, and Saccharomycopsis capsularis.Can.J. Bot. 57, 1259-1284.
BANDONI, R. J.( BISALPUTRA, A. A. & BISALPUTRA, T. (1967). Ascospore development inHansenula anomala. Can. J. Bot. 45, 361-366.
BECKETT, A., ILLINGWORTH, R. F. & ROSE, A. H. (1973). Ascospore wall development inSaccharomyces cerevisiae. J. Bad. 113, 1054-1057.
BLACK, S. H. & GORMAN, C. (1971). The cytology of Hansenula. III. Nuclear segregation anddevelopment during ascosporogenesis in Hansenula taingei. Arch. Mikrobiol. 79, 231-248.
CARROLL, G. C. (1967). The ultrastructure of ascospore delimitation in Saccobolus kerverniJ. Cell Biol. 33, 218-224.
CoNTi, S. F. & NAYLOR, H. B. (i960). Electron microscopy of ultrathin sections of Schizo-saccharomyces octosporus. III. Ascosporogenesis, ascospore structure, and germination. J.Bad. 79, 417-425.
EGEL, R. (1971). Physiological aspects of conjugation in fission yeast. Planta 98, 89-96.HIRATA, A. & TANAKA, K. (1982). Nuclear behaviour during conjugation and meiosis in the
fission yeast Schizosaccharomyces pombe. J. gen. appl. Microbiol. 28 (In Press.)JOHNSON, B. F., YOO, B. Y. & CALLEJA, G. B. (1973). Cell division in yeasts: Movement of
organelles associated with cell plate growth of Schizosaccharomyces pombe. J. Bad. 115,358-366.
LYNN, R. R. & MAGEE, P. T. (1970). Development of the spore wall during ascospore formationin Saccharomyces cerevisiae. J. Cell Biol. 44, 688-692.

Ascospore development in fission yeast 279
MOENS, P. B. & RAPPORT, E. (1971). Spindles, spindle plaques, and meiosis in the yeastSaccharomycet cerevisiae (Hansen). J. Cell Biol. 50, 344-361.
OLSON, L. W., EDEN, U., EGEL-MITANI, M. & EGEL, R. (1978). Asynaptic meiosis in fissionyeast? Hereditas 89, 189-199.
Oso, B. A. (1969). Electron microscopy of ascus development in Ascobolus. Arm. Bot. 33,205-209.
PETERSON, J. B., GRAY, R. H. & Ris, H. (1972). Meiotic spindle plaques in Saccharomycescerevisiae. J. Cell Biol. 53, 837-841.
ROLAND, J.-C, LEMBI, C. A. & MORRE, D. J. (1972). Phosphotungustic acid-chromic acid as aselective electron dense stain for plasma membranes of plant cells. Stain Technol. 47, 159-200.
SYROP, M. J. & BECKETT, A. (1972). The origin of ascospore-delimiting membranes inTaphrina deformam. Arch. Mikrobiol. 86, 185-191.
TSUBOI, M., OHASHI, K., TAKAHARA, M. & HAYASHIBE, M. (1978). Sexual process in eight-spored ascus-forming yeast, SMzosaccharomyees japonicus. I. Culture medium for thesynchronous sexual process. Plant & Cell Pkytiol. 19, 1327-1332.
Yoo, B. Y., CALLEJA, G. B. & JOHNSON, B. F. (1973). Ultrastructural changes of the fissionyeast (Schizosaccharomycet pombt) during ascospore formation. Arch. Mikrobiol. 91, 1-10.
(Received 21 August 1981)