Embed Size (px)
Citation preview

J. theor. Biol. (1997) 184, 353–358
0022–5193/97/030353+06 $25.00/0/jt960297 7 1997 Academic Press Limited
Asymmetric Light Competition and Founder Control in Plant Communities
M R* J B†
*Department of Biology, Imperial College, Silwood Park, Ascot Berkshire, SL5 7PY, U.K.†Department of Ecology and Evolution, University of Chicago, 1101 E. 57th Street, Chicago,
IL 60637, U.S.A.
(Received on 20 June 1996, Accepted in revised form on 11 October 1996)
We explore the effects of asymmetric competition for light on the outcome of interspecific competitionin simple models of resource competition. The models assume that species only differ in their allocationof resource to roots and photosynthetic tissues. Previous studies, which assume symmetric competitionfor light, have found that all two-species equilibria are unstable leading to founder control, and thatas species become more similar in their allocation of resources to roots and photosynthetic tissues sothe likelihood of founder control is reduced. In contrast when competition for light is asymmetric asspecies become more similar the likelihood of founder control increases. Vegetative successions in themodel can proceed in one of two directions—from nutrient specialist, with a high allocation to roots,to light specialists, with low allocation to roots, or vice versa. Low nutrient, low disturbance habitatstend to have successions running from light specialists to nutrient specialists, whereas high nutrient,high disturbance habitats have successions in the opposite direction. When light competition issymmetric, we demonstrate that only successions from leaf specialists to root specialists can becomearrested. In contrast, when light competition is asymmetric either type of succession may be arrested.Hence, asymmetric competition can have important effects on the outcome of inter-specific competitionand on ecosystem properties.
7 1997 Academic Press Limited
Introduction
The importance of plant morphology in determininghow plants forage for resources, and how this, inturn, affects competitive ability has long fascinatedecologists (Harper, 1977; Tilman, 1988). Intuitively,one would expect that differences in competitiveability will be smallest between species with similarforaging strategies. As a result, initial differences inbiomass or density might be sufficient to overcomeany innate competitive superiority, making theoutcome of competition contingent on initial con-ditions (Keddy & Shipley, 1989). The extent to whichinitial densities or biomasses determine the outcomeof competition is an important, unresolved questionin community ecology. When initial conditionsdetermine the outcome of competition we havefounder control, and this can be a potent forcepromoting diversity in spatially structured systems,
providing dispersal rates are low (Levin, 1974;Reynolds & Pacala, 1993).
Reynolds and Pacala (henceforth R&P) haveanalysed a model for resource competition betweenspecies that differ only in their allocation of resourcesto photosynthetic tissues and roots (Reynolds &Pacala, 1993). In their analysis, R&P assume that, ona per gram basis, leaves and roots of the competingspecies have an equivalent impact on light and soilresources respectively [i.e. resource use is perfectlysymmetrical (Weiner, 1990)]. They demonstrate thatfor any two allocation strategies there exist habitatswhere the outcome of competition is foundercontrolled. Since regions of founder control arerevealed in their analysis, the outcome of competitionis not determined solely by the allocation strategies ofthe competing species, and initial conditions candetermine the identity of the competitive dominant.However, in contrast to the intuitive reasoning

. . 354
described above, in R&P’s model as species becomemore similar in their allocation strategies thelikelihood of founder control is reduced. This issurprising since one might expect initial conditions tobecome more, not less, important in determining theoutcome of competition as species become moresimilar.
Both the analysis of R&P and the analysispresented here, are motivated by Tilman’s complexcomputer model ALLOCATE which simulates plantcompetition for light and soil nutrients (Tilman,1988). In this model, the light level that a speciesexperiences is determined by the total biomass at allheights above it. Hence, Tilman’s model includesasymmetric competition for light, tall individualsshade short ones but not vice versa (see alsoKohyama, 1993). Here we extend the analyticalresults of R&P to determine the extent to which theoutcome of competition is determined by initialconditions, both with symmetric and asymmetriccompetition for light.
The Model
Following R&P, where a more complete descrip-tion of the model is given, we begin by assumingthat species differ only in their allocation ofresources to roots and leaves, and that the percapita rate of change of biomass of species j is givenby
dBj
dt=Bj MING
G
G
K
k
rNAj
N+ kN−R
rL(1−Aj)L+ kL
−RGG
G
L
l−Bjd, j=1, 2(1)
where Bj is the biomass of species j, N is the amountof available soil nutrient, L is the light availability, Aj
is the fraction of biomass allocated to root by speciesj, r is the per captia maximal rate of plant growth, Ris the per capita respiration rate, d is the per capitaloss rate and kn and kL are the half saturationcoefficients for nutrient and light limited growthrespectively. Following R&P, we model soil nutrientdynamics as a linear function of resources suppliedand resources consumed,
dNdt
= a0T−N− s2
j=1
Bjp.1− s
2
j=1
MIN GG
G
K
k
rNAj
N+ kN−R
rL(1−Aj)L+ kL
−RGG
G
L
lBjp, (2)
where T is the total amount of all forms of soilnutrient in a habitat, a is the mineralisation rate andp is the plant tissue nutrient concentration. Light isassumed to equilibrate instantaneously,
L=L0
1+ s2
j=1
aBj(1−Aj)
, (3)
where L0 is the solar constant and a is the per biomasslight decay rate. The models contain no stagestructure, and populations are characterised solely bytheir biomass. As a result population growth isentirely contingent on vegetative growth, and themodels are only strictly appropriate for vegetativesuccessions.
For this system of equations, R&P demonstratethat monocultures are always dynamically stable:they derive the identity of the initial and finaldominant in succession, and also demonstrate that forall species pairs there exist habitats where competitionis founder controlled. Using R&P’s terminology Ai isthe allocation strategy of the initial dominant and Af
that of the final dominant. The final dominant isby definition uninvadable by all other allocationstrategies. We use the terms final dominant andoptimum allocation strategy interchangeably. R&Palso prove that any allocation strategy other than Af
can be invaded by a strategy closer to Af by somesufficiently small amount (o). Hence, we have thecounter-intuitive result that in any particular habitat,we can always choose species pairs that are sufficientlysimilar such that founder control does not occur. Asa direct consequence, if there are a large number ofintermediate strategies between Ai and Af thensuccession will never be arrested.
We extend R&P’s analysis to determine the impactof asymmetric light competition on founder control.Unlike R&P, we assume that during an invasion theresident species, B1, will have a greater proportion ofits leaves in full sunlight and therefore will experiencea higher light level,
L1 =L0
1+ a1B1(1−A1), (4)
where a1 is the resident’s light decay rate. An invadingspecies, B2 will experience lower light levels becausemost of its leaves are shaded by the resident, hence
L2 =L0
1+ a2B1(1−A1), (5)
where, a2 is the invader’s light decay rate. Forsymmetric light competition we have a1 = a2, the casestudied by R&P, whereas for asymmetric light

0.53
0.4
–10.30
Allocation to roots
Gro
wth
rat
e of
inva
der
0.40
0
0.2
–0.2
–0.4
–0.6
–0.8
0.33 0.35 0.38 0.43 0.45 0.48 0.50
(a) Af A1
0.53
0.2
–1.20.30
Allocation to roots
Gro
wth
rat
e of
inva
der
0.40
0
–0.2
–0.4
–0.6
–0.8
0.33 0.35 0.38 0.43 0.45 0.48 0.50
(b) AfA1
–1
355
competition a1 Q a2. This is of course a grosssimplification of competition for light, because in realcanopies some leaves are in full light whereas othersare in deep shade. However, we interpret the lightlevels each species experiences as a mean level whichdepends on whether the species is at equilibrium orinvading an established stand.
First we establish some notation. Let
dB1
dt= f1(B1, N, L), and
dB2
dt= f2(B2, N, L) (6)
where the functional forms of f1(B1, N, L) andf2(B2, N, L) are given by Eqn (1). Invasion into amonoculture of species 1 by species 2 will succeed if,at the equilibrium nutrient and light levels for species1, 1f2/1B2 q 0. The analysis is divided into two parts,first we explore the likelihood of founder controlwhen species have similar allocation strategies. Thenwe look at founder control of the optimum allocationstrategy.
Founder control of species with similar allocationstrategies
R&P demonstrate that, in any particular habitat,we can always find pairs of species with allocationstrategies that are sufficiently similar that foundercontrol does not occur. Here we explore the effect ofasymmetric light competition on this result. We dothis by asking if pairs of allocation strategies existsuch that each member cannot invade a monocultureof the other. Consider the case where species 1allocates a greater proportion of resources to roots
than the final dominant (i.e. A1 qAf). In this casespecies 1 is light limited at equilibrium. Withsymmetric light use, R&P show that this strategy isalways invadable by species 2 which has an allocationstrategy closer to Af by some small amount (o) andwhich is still light limited. Hence, species 2 iscompetitively dominant to species 1 and there is nofounder control. However, with asymmetric lightcompetition this is no longer true because,
limo4 0
1f2
1B2=
rL2(1−A1)L2 + kL
−R− dQ dB1
dt=0. (7)
This demonstrates that when competition for light isasymmetric and competing species are light limitedspecies 1 cannot invade species 2 and vice versa. Inthis case we have founder control for all sufficientlysimilar allocation strategies. This result is illustratedin Fig. 1(a) where it is shown that when there isasymmetric light competition then there is a zonearound species 1 where all nearby strategies fail toinvade (the growth rates for all similar strategies arenegative). Note, that the zone of founder controlaround a strategy increases as light competitionbecomes more asymmetric. For example, when lightcompetition is only weakly asymmetric (a1 =0.001,a2 =0.0011) then the dominant, Af, can invadespecies 1. However, when a2 =0.005 then invasion ofspecies 1 by even the competitive dominant will fail.Therefore, we find that when we have asymmetriccompetition for light as strategies become moresimilar so the likelihood of founder control isincreased. This contrasts with symmetric competitionwhere the likelihood of founder control is reduced asstrategies become more similar.
F. 1. Per capita growth rate of an invading strategy as a function of allocation to roots. The lines mark the invader’s growth ratewhen there is (1) symmetric light competition a1 = a2 =0.001—solid lines, (2) asymmetric light competition a1 =0.001 and a2 =0.0011—thedashed lines and (3) asymmetric light a1 =0.001 and a2 =0.005—the dotted lines. A1 marks the allocation strategy of the resident, Af theallocation strategy of the final dominant. (a) The resident has a higher allocation to roots than the dominant and so sufficiently similarstrategies are founder controlled providing a2 q a1 (i.e. there is asymmetric light competition). (b) The resident has a lower allocation toroots than the dominant and founder control of similar strategies only occurs if a2 is sufficiently large. Note, however, that Af—the finaldominant—cannot invade allocation strategy, A1, and so founder control occurs for sufficiently dissimilar strategies even when there issymmetric light competition.

. . 356
When species 1 allocates less to roots than thecompetitive dominant (i.e. A1 QAf) then it will benutrient limited at equilibrium. With symmetriccompetition R&P show that species 1 is alwaysinvadable by a strategy A2 =A1 + e which is alsonutrient limited. This is illustrated by the solid line inFig. 1(b), where the growth rates of similar strategiesthat are closer to Af are positive. Again, species 2 isthe competitive dominant and there is no foundercontrol. This result also holds for moderate degrees ofasymmetric light competition [a2 is not too large,Fig. 1(b) dashed line]. However,
lima24a
1f2
1B2= −R− dQ 0, (8)
indicating that as light use becomes sufficientlyasymmetric then a zone of founder control isintroduced [Figure 1(b), dotted line].
In summary when species are light limited then wehave founder control in all habitats for sufficientlysimilar strategies, provided competition for light isasymmetric. When similar species are nutrient limitedat equilibrium then founder control will only occur iflight use is sufficiently asymmetric. These resultssuggest an interesting pattern: in habitats with lowsoil nutrients and high light, Af will be large (i.e. highallocation to roots), most alternative strategies will benutrient limited, and similar strategies will only befounder controlled if light consumption is sufficientlyasymmetric. In contrast, in high soil nutrient habitatsAf will be small, most alternative strategies will belight limited, and hence similar strategies will often befounder controlled.
Founder control of the optimum allocation strategy
The results presented above assess the likelihoodof founder control between similar species. Whensimilar species are not founder controlled thenR&P demonstrate that succession will proceed inan ordered sequence to the dominant, providingsufficient intermediate species are present. When someintermediate species are absent then, even withsymmetric light competition, succession may becomearrested. However, if the optimum strategy caninvade all other strategies present in a particularhabitat then the endpoint of succession is always thecompetitive dominant, Af. Here we explore howasymmetric light competition alters the range ofstrategies into which the competitive dominant cansuccessfully invade. Biologically, this corresponds tothe introduction of the competitive dominant into amonoculture of a subordinate, as in the experimentsof Wedin and Tilman (1993).
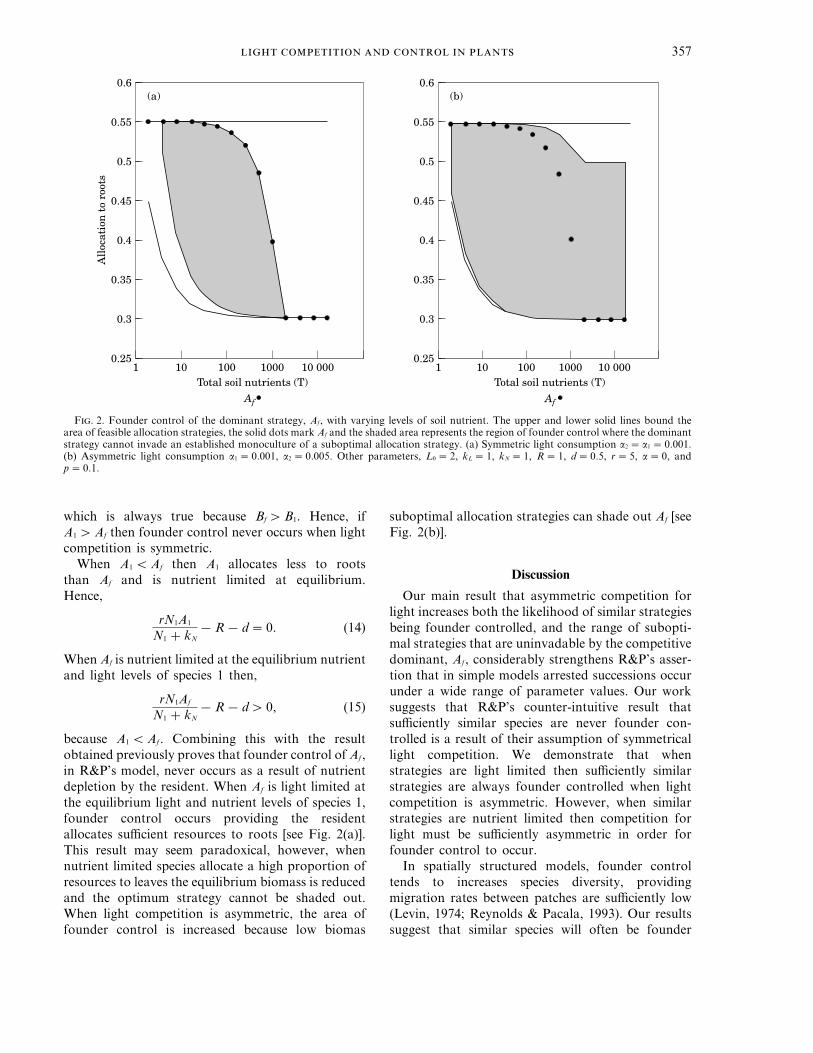
The procedure used was to determine theequilibrium biomass, light and nutrient levels for amonoculture of a suboptimal allocation strategy andthen numerically evaluate the condition for invasionby the dominant. By systematically searching therange of feasible strategies, we determined the set ofstrategies which were uninvadable by the competitivedominant. We repeated this procedure for a widerange of parameters. A representative plot showingthe area of founder control in relation to soil nutrientsis shown in Fig. 2. As expected, as soil nutrient levelsincrease the optimal allocation to roots is reduced.This occurs because biomass increases as soilnutrients become less limiting. This in turn makescompetition for light more intense, which favoursincreased allocation to leaves.
We found that when light competition is symmetricthen the zone of founder control is largest atintermediate levels of soil nutrients [Fig. 2(a)]. Fromthe figure it appears that with symmetric lightcompetition, Af is never founder controlled whenA1 qAf. In order to prove this, note that A1 allocatesmore to roots than Af and is therefore light limited atequilibrium. Since,
rL1(1−A1)L1 + kL
−R− d=0, (9)
when A1 is at equilibrium, Af can invade if it is lightlimited at the equilibrium light and nutrient levels ofspecies 1 (i.e. N1, L1) because
rL1(1−Af)L1 + kL
−R− dq 0, (10)
which is always true given A1 qAf. If, however, Af isnutrient limited at N1, L1 then we need to demonstratethat,
rN1Af
N1 + kN−R− dq 0, (11)
in order to prove that founder control does not occur.R&P show that Af is given by the followingexpression,
Af =0(d+R)r 1 01+
kNaaT−Bf p(a+ d)1 (12)
where Bf is the maximum biomass obtainable, whichoccurs when individuals allocate Af of resources toroots. Solving for N1 from eqns (1–3) and substitutingfor Af and N1 in eqn (11), and rearranging, we obtain
1+kNa
aT−Bf p(a+ d)q 1+
kNaaT−B1p(a+ d)
(13)
Af ·
0.6
0.251
Total soil nutrients (T)
All
ocat
ion
to
root
s
0.5
0.55
0.45
0.4
0.35
0.3
10 100 1000 10 000
(a)
Af ·
0.6
0.251
Total soil nutrients (T)
0.5
0.55
0.45
0.4
0.35
0.3
10 100 1000 10 000
(b)
357
F. 2. Founder control of the dominant strategy, Af, with varying levels of soil nutrient. The upper and lower solid lines bound thearea of feasible allocation strategies, the solid dots mark Af and the shaded area represents the region of founder control where the dominantstrategy cannot invade an established monoculture of a suboptimal allocation strategy. (a) Symmetric light consumption a2 = a1 =0.001.(b) Asymmetric light consumption a1 =0.001, a2 =0.005. Other parameters, L0 =2, kL =1, kN =1, R=1, d=0.5, r=5, a=0, andp=0.1.
which is always true because Bf qB1. Hence, ifA1 qAf then founder control never occurs when lightcompetition is symmetric.
When A1 QAf then A1 allocates less to rootsthan Af and is nutrient limited at equilibrium.Hence,
rN1A1
N1 + kN−R− d=0. (14)
When Af is nutrient limited at the equilibrium nutrientand light levels of species 1 then,
rN1Af
N1 + kN−R− dq 0, (15)
because A1 QAf. Combining this with the resultobtained previously proves that founder control of Af,in R&P’s model, never occurs as a result of nutrientdepletion by the resident. When Af is light limited atthe equilibrium light and nutrient levels of species 1,founder control occurs providing the residentallocates sufficient resources to roots [see Fig. 2(a)].This result may seem paradoxical, however, whennutrient limited species allocate a high proportion ofresources to leaves the equilibrium biomass is reducedand the optimum strategy cannot be shaded out.When light competition is asymmetric, the area offounder control is increased because low biomas
suboptimal allocation strategies can shade out Af [seeFig. 2(b)].
Discussion
Our main result that asymmetric competition forlight increases both the likelihood of similar strategiesbeing founder controlled, and the range of subopti-mal strategies that are uninvadable by the competitivedominant, Af, considerably strengthens R&P’s asser-tion that in simple models arrested successions occurunder a wide range of parameter values. Our worksuggests that R&P’s counter-intuitive result thatsufficiently similar species are never founder con-trolled is a result of their assumption of symmetricallight competition. We demonstrate that whenstrategies are light limited then sufficiently similarstrategies are always founder controlled when lightcompetition is asymmetric. However, when similarstrategies are nutrient limited then competition forlight must be sufficiently asymmetric in order forfounder control to occur.
In spatially structured models, founder controltends to increases species diversity, providingmigration rates between patches are sufficiently low(Levin, 1974; Reynolds & Pacala, 1993). Our resultssuggest that similar species will often be founder

. . 358
controlled and hence, when migration rates are low,this provides a mechanism that allows similar speciesto coexist. Coexistence as a result of founder controland low dispersal rates differs from the competition-colonisation hypothesis which depends on a trade-offbetween competitive ability and colonisation success.According to this hypothesis weaker competitorsare only able to persist if they colonise sites morerapidly than they are excluded from them, because asuperior competitor will always be able to exclude theresident when it arrives at a site (Levins, 1969;Tilman, 1994). Thus, we may distinguish two types ofpriority effect, one is transitory and occurs in modelswhere an inferior competitor will be excluded bycompetitively superior species when it arrives at a site.The other is long-term and the identity of thecompetitive dominant depends on the initial con-ditions—once a species is established at a site itcannot be excluded.
Founder control has important implicationsfor succession. In R&P’s model, the direction ofsuccession can proceed in one of two directions—from nutrient specialist, with a high allocation toroots, to light specialists, with low allocation to roots,or vice versa. Low nutrient, low disturbance habitatstend to have successions running from light specialiststo nutrient specialists, whereas high nutrient, highdisturbance habitats have successions in the oppositedirection (Reynolds & Pacala, 1993). When lightcompetition is symmetric, our results demonstratethat only successions from leaf specialists to rootspecialists can become arrested. In contrast, whenlight competition is asymmetric either type ofsuccession may be arrested.
We believe that the inability of suboptimalstrategies to reduce soil nutrients to a level that Af
cannot invade is a result of a fundamental differencebetween the ways that light and soil nutrients areconsumed. Since light is a directional resource, it isalways consumed in proportion to leaf biomass. Incontrast, soil nutrients are both consumed andreleased as a result of the death of plant tissues. This
will tend to limit the level to which nutrients canbe reduced. The model also assumed that speciesonly differ in their allocation of biomass to rootsand leaves. However, in perennial grasses nitrogenmineralisation rates have been shown to be correlatedwith tissue nitrogen concentration and the allocationof biomass above and below ground (Wedin &Tilman, 1990). Therefore, further studies of a widerange of resource competition models that incorpor-ate physiological changes which are correlated withdifferences in allocation strategy are required todetermine the generality of this result. Neverthelessour study clearly demonstrates the importance ofunderstanding the degree of asymmetry in resourcecapture when trying to interpret competitive inter-actions.
Work funded by the Royal Society (MR) and NSF grantDEB-9496331 (JB).
REFERENCES
H, J. L. (1977). Population Biology of Plants. London:Academic Press.
K, P. A. & S, B. (1989). Competitive hierarchies inherbaceous plant communities. Oikos 54, 234–241.
K, T. (1993). Size-structured tree populations in gap-dynamic forest—the forest architecture hypothesis for stablecoexistence of species. J. Ecol. 81, 131–143.
L, S. A. (1974). Dispersion and population interactions.Am. Nat. 108, 207–228.
L, R. (1969). Some demographic and genetic consequences ofenvironmental heterogeneity for biological control. Bull. Ent.Soc. Am. 15, 237–240.
R, H. L. & P, S. W. (1993). An analytical treatmentof root-to-shoot ratio and plant competition for soil nutrient andlight. Am. Nat. 141, 51–70.
T, D. (1988). Plant Strategies and the Dynamics and Structureof Plant Communities. Princeton, NJ: Princeton University Press.
T, D. (1994). Competition and biodiversity in spatiallystructured habitats. Ecology 75, 2–16.
W, D. & T, D. (1990). Species effects on nitrogen cycling:a test with perennial grasses. Oecologia 84, 433–441.
W, D. & T, D. (1993). Competition among grasses alonga nitrogen gradient: initial conditions and mechanisms ofcompetition. Ecol. Mono. 63, 199–229.
W, J. (1990). Asymmetric competition in plant populations.Trends Ecol. Evol. 5, 360–364.





























