Embed Size (px)
Citation preview

3/19/10
1
Aspartate carbamoyl Transferase (ATCase)
Manickam Sugumaran University of Massachusetts
Boston, MA 02125
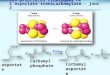
ATCase is the key enzyme responsible for the biosynthesis of pyrimidines
N-Carbamoyl aspartateAspartic acidCarbamoyl phosphate
+H2N COOH
COOHOP
O NH2
OO
O NH2
O NH
COOH
COOH
N
C
O
N
NH2
OHOH
OO
PO
PO
PO
O O O
O OO
Cytidine triphosphate (CTP)
ATCase
Pi

3/19/10
2
ATCase follows ordered Bi Bi reaction Carbamoyl phosphate binds to the enzyme first. Aspartate then comes
and attacks the carbonyl carbon of carbamoyl phosphate. This results in a tetrahedral intermediate formation. The transition state then breaks down to produce carbamoyl aspartate (which then comes out of the
enzyme first) and inorganic phosphate.
E - Carbamoyl Phosphate
Carbamoyl Phosphate
Carbamoyl AspartateAspartate
InorganicPhosphate
EE - Carbamoyl Phosphate + Aspartate
E - Carbamoyl Aspartate + Phosphate
E - phosphateE
Succinic acid is a competitive inhibitor (against aspartate) for the enzyme.
N-Carbamoylaspartate
Succinic acid (competitive inhibitor)
OP
O NH2
OO
O
Aspartic acidCarbamoyl phosphate
+H2N COOH
COOH
H2N
O
N COOH
COOH
H
COOH
COOH
Pi

3/19/10
3
Consistent with the reaction mechanism, N‐(phosphanacetyl)‐L‐aspartic acid, the bisubstrate, transition state analog of the enzyme, serves as a
potent inhibitor for ATCase (GKi = 10‐8 M).
Pi
N
H
HOHO
OHO
OO
O
OP H2N
OC
Transition State
N-Carbamoylaspartate
H2N NH COOH
OCOOH
OP
O NH2
OO
O
Aspartic acidCarbamoyl phosphate
+H2N COOH
COOH
Transition State analog
N-Phosphonacetyl L-aspartic acid
N
HOHO
OHO
OC
O
OP
OC
H
H
Biosynthesis of CTP
Glutamine + 2 ATP + O=C=O ---> Glutamate + 2 ADP + Pi + Carbamoyl phosphate 1
7
8
Enzymes:1. Carbamoyl phosphate synthetase2. Aspartate transcarbamoylase3. Dihydroorotase4. Dihydroorotate dehydrogenase5. Orotate Phosphoribosyl transferase6. Orotidylate decarboxylase7. Nucleoside monophosphate kinase8. Nucleotide diphosphate kinase9. CTP synthetase
NADNADH
+ Aspartate
N-Carbamoyl aspartate2
3
4
5
6
Dihydroorotate
Orotate
Orotidylate (OMP)
Uridylate (UMP)
H2O
UDP
UTP
CTP9
PRPPPPi
CO2
ATP ADP
ADPATP
ATP Gln +Glu + ADP

3/19/10
4
Biosynthesis of CTP and regulation (in E.Coli)
H2N
COOHNH2
OCO2
UMP
UDP
+ 2 ATP
COOHH2N
COOH
COOHNH
CO
OHH2N
O
Carbamoyl Phosphate
Aspartate
Glutamine+
+ 2 ADP + Pi
COOHNH
CO
O
N HH
ATCase
Dihydroorotase
Dihydroorotatedehydrogenase
COOHNH
CO
O
NH
OP
NH2
O
O
O O
N-carbamoyl aspartate
Dihydrooratate
Oratate
OHOH
OOP
O OO
COOHN
CO
O
NH
PRPPPPi
Oratate Phosphoribosyl transferase
Orotidylate
Orotidylate decarboxylase
UTP
CTP
Effect of ATP (activator) and CTP (inhibitor) on ATCase
[S] mM
v
+ CTP
+ ATP Control
Activator (ATP) decreases sigmoidity; Inhibitor (CTP) increases sigmoidity
Aspartate

3/19/10
5
The significance of CTP/ATP regulation
CTP (end product inhibition) is essential to turn off the synthetic pathway when excess CTP accumulates.
When high concentration of purines accumulate and energy rich environment exist, ATP activation helps to produce enough pyrimidines for DNA replication.
ATCase can be modified by pcmb. Modified ATCase exhibits Michaelis Menten type kinetics and is not
responsive to either ATP or CTP.
+ ClHg COOH
Enzyme thiol p-chloro mercuribenzoate
HS
OH
SH
+
Hg COOHS
Regenerated Enzyme
SH
SO
HHg COOH
[S] mM
v
[S] mM
v
[S] mM
v

3/19/10
6
Catalytic and regulatory sites are located on different subunits of ATCase.
p‐hydroxymercuribenzoate dissociates ATCase into catalytic subunits (C3) and regulatory subunits (R2).
C3 and R2 can be separated by centrifugation and/or ion exchange chromatography.
Isolated C3 shows ATCase activity, but exhibits only typical Michaelis Menten kinetics. It is also non responsible to allosteric effectors ‐ ATP and/or CTP.
Regulatory subunit binds to ATP and CTP but does not have any detectable ATCase activity.
Reconstituion of regulatory and catalytic subunits (after the removal of mercurial) generates the fully fiunctional ATCase.
Catalytic and regulatory sites are located on different subunits of ATCase.
ATCase treated with p‐hydroxymercuribenzoate (pCMB) does not exhibit allosteric kinetics. This desensitization is caused by the separation of catalytic subunits (C3) and regulatory subunits (R2) by the reaction with mercurials.
ATCase Subunits
pCMB 2
3+
C
RR
C R C C
R
CR
R
C
C
R
CR C

3/19/10
7
Catlytic subunit C3 and regulatory subunit R2 can be separated by centrifugation and/
or ion exchange chromatography.
Distance migrated
C6R6
R2C3
ATCase Subunits
pCMB 2
3+
C
RR
C R C C
R
CR
R
C
C
R
CR C
Native ATCase: (C6R6) Allosteric kinetics Catalytic subunit alone : (C3) Michaelis Menten kinetics
Regulatory subunit: (R2) No enzyme activity; but binds to ATP or CTP.
Regulatory Subunits bind to ATP/CTP; but no catalytic activity.
3 R R
The i
The i
The i2 C C C
ATCase
The i
The i
The i
The i
The i
The i
C
R
C R C R
R C
R
C R C
[S] mM
v
[S] mM
v

3/19/10
8
Reconstitution of catalytic and regulatory subunits restores the fully functional ATCase
The image cann
The image cann
The image cann
2
3 +
R
C
R
C C
The image cann
The image cann
The image cann
The image cann
The image cann
The image cann
C
R
C R
C R
R C
R
C
R C
Reconstitution
The reconstituted enzyme now shows allosteric properties.
Catalytic subunits show normal M.M. kinetics & insensitive to ATP/CTP
Subunit structural arrangement in ATCase
Top
Bottom
Bottom
Bottom
Top
Top
Bottom
Bottom
Bottom
Top
Top
Top
ATCase has a three fold axis of symmetry
C
C
C
C
CC
RR
RR
RR

3/19/10
9
Crystal structure of ATCase
Ligand interaction with ATCase
Aspartate exhibits positive homotropic interaction.
ATP shows positive heterotropic interaction.
CTP shows negative heterotropic interaction.

3/19/10
10
Effects of ATP and CTP
The end product metabolism, CTP inhibits ATCase reaction allosterically.
The activator ATP competes to the same site at which CTP binds; but the results are different. While ATP enhances the affinity of ATCase for its substrates; CTP decreases the affinity.
Allosteric effects of ATCase can be explained by concerted model
The T state is more closed (tight). Therefore it is poorly accessible to the substrate.
The R (relaxed) state is more open and allows the substrate to bind to it freely.

3/19/10
11
Substrate binding to one subunit causes conformational changes in
other subunits also.
Tight state Relaxed state
Substrate
Nitration experiment
C(NO2)4
Native enzyme Nitrotyrosine containing modified enzyme
Nitration of native enzyme produces a modified enzyme which has yellow color. Alteration in its conformation can now be monitored in the visible specrum
OH OH
NO2
C(NO2)4

3/19/10
12
Active site is not in direct geometric relation with subunit interaction site.
Reconstitution of normal catalytic trimer with nitrotyrosine containing trimer generates the dodecamer. Succinate binding to the normal catalytic trimer, causes conformation changes in the nitrotyrosine containing trimer also. This confirm that conformational changes in one subunit are communicated to the others even though the second one does not bind to the substrate or its analog.
Tight state Relaxed state
Succinate