Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 374:70-83 (1996)
Atypical and Typical Neuroleptic Treatments Induce Distinct Programs
of Transcription Factor Expression in the Striatum
NOBORU HIROI AND ANN M. GRAYBIEL Department of Brain and Cognitive Sciences, Massachusetts Institute of Technology,
Cambridge, Massachusetts 02 139
ABSTRACT Atypical and typical neuroleptics, when administered chronically, can bring about pro-
found but contrasting changes in schizophrenic symptoms and motor activation and dramati- cally modulate brain neurochemistry. To explore the transcriptional events that might be involved in this neurochemical regulation, we used immunohistochemistry and immunoblot- ting to examine the expression patterns of two bZip transcription factors, c-Fos and FosB, in the striatum of rats treated acutely and chronically with neuroleptic drugs of different classes. Typical and atypical neuroleptic drugs produced contrasting regulatory effects on a FosB-like protein of ca. 36-39 kDa, the molecular weight of truncated FosB (AFosB). Chronic treatments with two typical neuroleptics, haloperidol and metoclopramide, but not with the atypical neuroleptic clozapine, led to markedly enhanced FosB-like immunoreactivity in the caudoputa- men. Further, c-Fos-like protein in the striatum, considered a marker for the induction of antipsychotic actions by neuroleptic treatments, was downregulated by chronic treatment with the two potent antipsychotic drugs tested, but not by chronic treatment with metoclopramide, which has low antipsychotic efficacy but induces extrapyramidal side effects. These results suggest that chronic treatments with neuroleptics having different effects on cognitive and motor behavior induce different long-term changes in transcription factor expression in the striatum. Nevertheless, we found that neuroleptics of both classes regulated transcription factor expression in overlapping populations of striatal neurons expressing enkephalin or DARPP-32. Contrasting patterns of transcriptional regulation in these neurons may thus contribute to the distinct neurochemical and behavioral effects that characterize neuroleptics of different classes. o 1996 Wiley-Liss, Inc.
Indexing terms: neuroleptics, striosome, Fos, FosB, extrapyramidal side effects
Neuroleptic treatment remains the single most reliable way to treat schizophrenic symptoms. These drugs alter second-messenger systems in neurons by blocking neuro- transmitter receptors. They are thought thereby to counter- act the actions of endogenous neurotransmitters. The receptors affected include subclasses of dopamine, seroto- nin, and acetylcholine receptors, all of which are strongly represented in the circuitry of the basal ganglia and frontal lobes (Graybiel, 1990; Seeman, 1990; Cleghorn et al., 1991; Gerfen, 1992; Ellenbroek, 1993; Sunahara et al., 1993).
Based on their contrasting chronic effects on schizo- phrenic symptoms, neuroleptics are divided into two classes. Typical neuroleptics attenuate the positive symptoms of schizophrenia. Atypical neuroleptics, including clozapine, attenuate both positive and negative schizophrenic symp- toms (Gross and Langner, 1966; Angst et al., 1971; Acken-
heil et al., 1976; Chouinard and Annable, 1976; Casey, 1989; Hippius, 1989; Cleghorn et al., 1991). This general- ized action of clozapine on schizophrenic symptoms and its lack of motor side effects suggest that clozapine may act selectively on the specific neuronal systems dysfunctional in the brains of schizophrenics. By contrast, as typical neuroleptics have little effect on negative symptoms of schizophrenia and can induce disabling motor side effects, they may have only partial and less selective actions within
Accepted July 2, 1996. N. Hiroi is now at Yale University School of Medicine, Department of
Psychiatry, Division of Molecular Psychiatry, 34 Park Street, New Haven, CT 06508.
Address reprint requests to Ann M. Graybiel, MIT, E25-618, Cambridge, MA 02139. E-mail: [email protected]
O 1996 WILEY-LISS, INC.

NEUROLEPTICS AND STRIATUM
this brain circuitry (Baldessarini and Tarsy, 1980; Braude et al., 1983; Ellenbroek, 1993).
A key feature of neuroleptics of both classes is that they become increasingly effective as antipsychotics with pro- longed use. Similarly, most of the extrapyramidal side effects accompanying typical neuroleptic treatment in- crease with time or appear only after repeated treatment. This time course suggests that neuroleptics may induce gradual changes in the neural systems they affect. Conse- quently, long-term changes in gene expression are likely to occur in these neurons (see Berridge, 1986; Goelet et al., 1986; Comb et al., 1987; Sheng and Greenberg, 1990; Morgan and Curran, 1991; Hyman and Nestler, 1993).
In initial tests of this idea, we and others have shown that acute treatments with neuroleptics of either type enhance expression of c-Fos-like transcription factor in the rat striatum and that typical and atypical neuroleptics do so in different anatomical patterns (Hiroi et al., 1991; Hiroi and Graybiel, 1992, 1993a,b; Nguyen et al., 1992; Robertson and Fibiger, 1992). Interestingly, TRE/AP-1-like binding sites to which c-Fos can bind are in the 5’ region of genes encoding a number of neuropeptides that are regulated by neuroleptics in the striatum (see Graybiel, 1990; Angulo and McEwen, 1994), including prodynorphin (Naranjo et al., 1991), neurotensin (Kislauskis and Dobner, 19901, and cholecystokinin (Monstein, 1993). Such changes in neuro- peptide levels have been suggested to contribute to the therapeutic effects of neuroleptics (Bissette and Nemeroff, 1985). c-Fos and related gene products are thus among the candidate transcription factors that could bridge between the acute cell surface receptor occupancy by neuroleptic drugs and the gradual, long-lasting neural changes induced when they are given chronically.
We demonstrate here that neuroleptics of the typical and atypical classes bring about contrasting changes in the inducibility of FosB-like and c-Fos-like proteins in subpopu- lations of striatal neurons.
71
MATERIALS AND METHODS Animals
Male Sprague-Dawley rats (180-300 g, Charles River) were used in the experiments. Animals were kept under a 12- 12-hour light-dark cycle with food and water ad libitum.
Drug treatments For acute treatment, each rat was given a single intraperi-
toneal injection of clozapine (10, 20, or 40 mg/kg, Sandoz Research Institute, East Hanover, NJ), haloperidol (0.25, 0.5, or 1.0 mg/kg, Sigma Chemical Co., St. Louis, MO), metoclopramide (0.25,0.5, 1.25,2.5,5, or 10 mg/kg, Sigma Chemical Co., St. Louis, MO), or the corresponding vehicle as control, and was perfused 2 hours later. Another set of animals was acutely treated with clozapine (20 mglkg, i.p.) or with vehicle and perfused after 8 hours. As a positive control for the double-staining procedures, some rats were injected with &hetamine (5 mg/kg, Sigma Chemical Co., St. Louis, MO) or 0.9% saline (1 ml/kg) and were perfused 2 hours later.
Clozapine was dissolved in 1 M tartaric acid and diluted with distilled water (dH20). Vehicle was made by mixing tartaric acid and water. The drug and vehicle solutions were adjusted to pH 5-6 with 1 N NaOH. Haloperidol was dissolved in 1 M acetic acid and diluted with d H 2 0 , and its vehicle was the same concentration of acetic acid. Both
solutions were adjusted to ca. pH 7 with 1 N NaOH. Metoclopramide HC1 was dissolved in 0.9% saline, pH 7.4. Its vehicle was 0.9% saline.
For chronic drug treatments, clozapine (n = 121, haloperi- do1 (n = 81, and metoclopramide (n = 12) were injected i.p. at doses of 20 mg/kg, 0.5 mglkg, and 5 mglkg, respectively. The 5-mg/kg dose of metoclopramide in the rat was esti- mated to be equivalent to the dose in humans that results in extrapyramidal side effects without significant antipsy- chotic effects (Lavy et al., 1978; Wood, 1978). Animals were randomly assigned to one of four groups for each drug. One group received repeated injections of vehicle for 8 days and an acute injection of vehicle on the 9th day (W). The second group was given repeated injections of vehicle for 8 days and an acute injection of clozapine (VC), haloperidol (VH), or metoclopramide (VM) on the 9th day. The third group received repeated injections of clozapine (CV), halo- peridol ( H V ) , or metoclopramide ( M V ) for 8 days and then an acute injection of vehicle on the 9th day. The fourth group was given repeated injections of clozapine (CC), haloperidol (HH), or metoclopramide (MM) for 8 days and an acute injection of the same drug on the 9th day.
All animals were deeply anesthetized 2 hours after the last vehicle or drug injection on the 9th day and were perfused with 0.9% saline followed by 4% paraformalde- hyde in 0.1 M Na-K phosphate buffer.
Immunohistochemical procedures Brains postfixed in 4% paraformaldehyde in 0.1 M Na-K
phosphate buffer for 1-2 hours and cryoproteded in 20% glycerol overnight were cut at 15 or 20 km on a freezing microtome.
Immunostaining was carried out with the following antisera: 1) a polyclonal antiserum against c-Fos, 1:200 (Oncogene Science, Inc., Uniondale, NY); 2) a polyclonal antiserum against the conserved M-peptide sequence in c-Fos and Fos-related antigens (FRAs), 1:5,000 (kindly donated by Dr. M.J. Iadarola); 3) a polyclonal antiserum against FosB/AFosB, 1:200 or 1:400 (Santa Cruz Biotech- nology, Santa Cruz, CA); 4) a monoclonal antibody against calbindin-DzsK, 1:1,000 (Sigma Chemical Co., St. Louis, MO); 5) a polyclonal antiserum against calretinin, 1:2,500 (Chemicon, Temecula, CA); 6) a polyclonal antiserum against methionine-enkephalin, 1:2,000 (INCstar Corp., Stillwater, MN); 7) a polyclonal antiserum against dynor- phin B, 1:1,000 or 1:2,000 (Drs. E. Weber and M. Sonders); 8) a monoclonal antibody against choline acetyltransferase (CUT), 1:800 (INCstar Corp., Stillwater, MN); 9) a mono- clonal antibody against dopamine- and CAMP-regulated phosphoprotein (DAFtPP-32), 1:20,000 (kind gift of Drs. P. Greengard, H. Hemmings, and E.L. Gustafson); 10) a monoclonal antibody against parvalbumin, 1: 1,000 (Sigma Chemical Co., St. Louis, MO).
No immunoreactivity was detected when these antisera were preadsorbed with their respective peptides or when the primary antisera were omitted. We refer to immuno- staining observed with the polyclonal antisera against c-Fos and FosB as “c-Fos-like” and “FosB-like” immunoreactivi- ties. We follow similar conventions to refer to immunostain- ing with the other antisera listed. We attempted to analyze FRA induction immunohistochemically with the M peptide antiserum used for immunoblotting (Iadarola et al., 1989), but there were high constitutive levels of Fos/FRA staining in the caudoputamen and nucleus accumbens, as also reported by Dilts et al. (1993). This constitutive expression,

72
particularly in the nucleus accumbens, made it difficult to analyze neuroleptic-induced c-FosiFRA induction (cf. Deutch et al., 1992; Dilts et al., 1993). The data therefore were excluded from analysis.
Immunohistochemistry was used to mark different neu- ronal populations of the striatum (Graybiel, 1990; Gerfen, 1992) as follows: dynorphin-like immunoreactivity as a marker of striatal projection neurons that predominantly project to the substantia nigraiventral tegmental area complex and entopeduncular nucleus (Gerfen and Young, 1989; Reiner and Anderson, 1990); met-enkephalin immu- noreactivity as a marker of striatal neurons that predomi- nantly project to the globus pallidusiventral pallidum (Gerfen and Young, 1989; Reiner and Anderson, 1990; Kalivas et al., 1993); NADPH-diaphorase as a marker of striatal interneurons that co-express somatostatin (Vincent et al., 1983; Vincent and Johansson, 19831, neuropeptide Y (Sandell et al., 1986), nitric oxide synthase (Hope et al., 1991), and GABA (Kubota et al., 1993); parvalbumin as a marker of another set of GABA-containing striatal interneu- rons (Kubota et al., 1993); calretinin as a marker of a third set of GABA-containing striatal interneurons (Kubota et al., 1993); choline acetyl transferase (ChAT) as a marker of striatal cholinergic interneurons; and DARPP-32 as a marker of neurons that predominantly (Walaas and Green- gard, 1984), but not exclusively (Anderson and Reiner, 1991; LeMoine et al., 19911, express D1-class dopamine receptors in the striatum.
Single antiserum immunohistochemistry. Free- floating sections were stained by the indirect avidin-biotin (ABC) technique. Sections were treated for 10 minutes with 3% HzOz in 0.01 M Na phosphate buffer containing 0.2% Triton X-100 and 0.9% NaCl (PBS, pH 7.4), were then treated for 30 minutes with 5% or 10% normal serum of the species in which the secondary antiserum was raised, and were incubated in primary antiserum overnight at 4°C. Sections were then incubated in the secondary antiserum for 1 hour and subsequently in avidin-biotin-peroxidase complex (1: 170, Vector Laboratories, Burlingame, CA) for 1 hour. Horseradish peroxidase conjugated with biotin was devel- oped with diaminobenzidine (DAB; Sigma Chemical Co., St. Louis, MO) in the presence of hydrogen peroxide (0.001%). Each step was followed by washes in PBS (3 x 10 minutes).
Dual antiserum immunohistochemistry. Staining for two antigens in single neurons was carried out either by nickel-intensified DAB vs. DAB double staining or by silver-intensified immunogold vs. DAB double staining. The nickel-intensified DAB method produced different col- ors: gray-black for nickel-intensified DAB and brown for DAB. Sections were stained with c-Fos or FosB antiserum with nickel-intensified DAB first, and then with antiserum against ca1bindin-Dz8K, calretinin, or parvalbumin followed by DAB development. For nickel-DAB staining with antise- rum against dynorphin or ChAT, the order was reversed. After the first staining was completed, the remaining horseradish peroxidase was blocked by treatment with 0.6% H202 for 45 minutes. The remaining biotin and avidin were blocked, respectively, by treatment with blocking avidin (30 minutes) and blocking biotin (30 minutes).
The second method, followed for DARPP-32 and met- enkephalin dual staining, marked one antigen with silver- intensified colloidal gold particles and the second with brown-colored DAB (Berretta et al., 1992). Sections were incubated in the primary antiserum for c-Fos (1:200) or FosB (1:400) overnight at 4"C, and then in goat anti-rabbit
N. HIROI AND A.M. GRAYBIEL
IgG linked to colloidal gold (1:200 or 1:400, AuroProbe LM GAR, Amersham, Arlington Heights, IL) overnight at 4°C. The colloidal gold was detected by silver enhancement (IntenSE LM, Amersham, Arlington Heights, IL) for 20 minutes. This staining was followed by DAB staining for met-enkephalin or DARPP-32. Briefly, we used 0.25 M Tris saline for the enkephalin staining and 0.1 M Tris saline for the DARPP-32 staining. Sections were incubated consecu- tively in 10% methanol containing 3% HzOz for 5 minutes, 2% bovine serum albumin for 30 minutes, and in the primary antiserum overnight at 4°C. Sections were then incubated in secondary IgG raised against the species in which the primary antiserum was raised, and in avidin- biotin complex. Horseradish peroxidase conjugated to bio- tin was detected by DAB in the presence of HzOz. The fact that c-Fos and FosB antisera stained nuclei, whereas other markers stained the cytoplasm, makes it unlikely that double labeling was the result of crossreactivity.
NADPH diaphorase histochemistry was carried out ac- cording to the protocol of Vincent et al. (1983). For dual staining, sections were washed overnight in PBS containing 0.2% Triton X-100 and were then stained for c-Fos or FosB with DAB chromogen.
Statistical analysis In selected sets of sections from the chronic neuroleptic
treatment groups, the numbers of immunoreactive nuclei were analyzed with the aid of a Biocom Image Analyzer (Les Ulis, France). The numbers of immunoreactive c-Fos- or FosB-positive nuclei were counted in specific representative regions of both hemispheres within 1) the anterior nucleus accumbens (A1.70 mm, Paxinos and Watson, 1986), 2) the ventral caudoputamen, 3) the medial caudoputamen, 4) the dorsal caudoputamen, and 5) the lateral caudoputamen (all at A1.O-1.2 mm, Paxinos and Watson, 1986; see Saper, 1996). These levels showed the maximal induction exhib- ited in sections through the nucleus accumbens and caudo- putamen. For each section, rectangular (0.5 x 1.0-mm) areas were chosen from charts by reference to reproducible landmarks. These counting rectangles were superposed 1) on the anterior nucleus accumbens (NA), horizontally oriented, 2) on the ventral caudoputamen (VCP), horizon- tally oriented and 0.5 mm dorsal to the anterior commis- sure, 3) on the middle of the medial caudoputamen (MCP), parallel and adjacent to the lateral ventricle, 4) on the dorsal tip of the caudoputamen (DCP), parallel and adja- cent to the corpus callosum, and 5 ) on the lateral tip of the caudoputamen (LCP), parallel and adjacent to the corpus callosum. Counts from both hemispheres of each animal were combined, and these summed values were analyzed by a two-way, one-repeated measure design analysis of vari- ance (ANOVA) with planned comparisons, with the chronic treatments as an independent factor and the striatal loci as a repeated factor.
Immunoblotting Immunoblotting was carried out on homogenates of
striatal tissue from both acutely and chronically treated animals. For the acute study, rats were decapitated 2 hours after injection of clozapine (20 and 40 mgikg), haloperidol (0.5 and 1 mgikg), metoclopramide (5 and 10 mgikg), or vehicle. For the chronic study, rats were given eight daily injections of clozapine (20 mgikg, i.p.1, haloperidol (0.5 mgikg, i.p.1, or metoclopramide (5 mgikg, i.p.1. These

NEUROLEPTICS AND STRIATUM 73
animals were then given injections of vehicle (CV, HV, and M V ) or the respective neuroleptics (CC, HH, and MM) and euthanized 2 hours later on the 9th day. Controls were given vehicle injections for 8 days, and tissue was taken 2 hours after the last vehicle injections, on the 9th day (W).
Brains were rapidly removed, and samples from the nucleus accumbens and from the caudoputamen were dissected at 0°C. They were homogenized in 20 mM HEPES buffer containing 0.4 M NaCI, 20% glycerol, 5 mM MgC12, 0.5 mM EDTA, 0.1 mM EGTA, 1% Nonidet-P40, 10 pg/p1 leupeptin, 0.1 mM p-aminobenzamidine, 1 pg/ p1 pepstatin, 0.5 mM PMSF, and 5 mM DTT and were left at 4°C for 20 minutes. Samples were centrifuged at 15,OOOg for 20 minutes. Supernatants were used for analysis.
Samples were separated by sodium dodecyl sulfate- polyacrylamide gel electrophoresis (SDS-PAGE, 10% acryl- amide/0.4% bis-acrylamide gel) and were transferred to polyvinylidene fluoride (PVDF) membranes (Immobilon-P, Millipore Corp., Bedford, MA). Transfer membranes were incubated with 4% dry milk for 60 minutes at room temperature and then with polyclonal FRA antiserum (1:4,000, anti-FRA antiserum, a gift of Dr. M. Iadarola, NIH) or with FosB antiserum raised against a sequence shared by FosB and AFosB (FosBiAFosB antiserum, 1:1,000, Santa Cruz Biotech., Santa Cruz, CAI overnight at room temperature. The membranes were washed and incu- bated with the rabbit anti-goat IgG peroxidase conjugate (1:4,000, Sigma Chemical Co., St. Louis, MO) for 2 hours. After washing, the membranes were developed with a chemiluminescence kit (Renaissance, DuPont NEN, Bos- ton, MA) and exposed to high-performance luminescence detection film (Hyperfilm-MP, Amersham, Arlington Heights, IL).
For control experiments, primary antisera were incu- bated with an excess amount of the corresponding peptide overnight at 4"C, and membranes were stained with the preadsorbed antibody. Bands that were eliminated by this procedure were considered to be specific proteins. All experiments were carried out at least in triplicate.
RESULTS Acute treatments with typical and atypical neuroleptics induce contrasting patterns of expression of c-Fos-like and FosB-like
proteins in the striatum Basal levels of c-Fos-like immunoreactivity in the stria-
tum were minimal, and vehicle injections did not induce an appreciable increase in immunoreactivity. Immunostained nuclei appeared occasionally in the dorsal corner of the shell division of the nucleus accumbens and in the medial caudoputamen. By contrast, considerable FosB-like immu- noreactivity was expressed constitutively in the nucleus accumbens, and there was a low level of FosB-like immuno- reactivity in the dorsal caudoputamen as well. Each drug induced the transcription factors in a dose-dependent man- ner.
Clozapine. Acute low-dose clozapine ( 5 and 10 mg/kg) induced c-Fos-like immunoreactivity almost exclusively in the nucleus accumbens. The induction was specific to the anterior and calbindin-poor dorsomedial parts. In the cau- doputamen, only a few scattered c-Fos-positive nuclei appeared; they were in the medial caudoputamen. At higher doses (20 and 40 mgikg), c-Fos-positive nuclei appeared throughout the anterior and dorsomedial parts of the
nucleus accumbens and in the medial caudoputamen. In addition, at the highest (40 mg/kg) dose, immunoreactive nuclei appeared along the dorsal margin of the caudoputa- men and in striosomes, identified by immunostaining for ~ a 1 b i n d i n - D ~ ~ ~ (Fig. 1A).
The induction of FosB-like protein was much weaker. Following acute treatment with clozapine, there was little or no induction of FosB immunostaining at low doses, and with higher doses, induction was found mainly in the nucleus accumbens and in the dorsolateral caudoputamen. Quantitative analysis of the 20 mgikg material did show weak induction in the medial and ventral caudoputamen (136% and 132% of control, respectively), but this was not significant (medial, F(1,8) = 1.52, P > .05; ventral, F(1,8) = 1.49, P > .05, see Fig. 4B, W and VC). In the nucleus accumbens, there were high levels of constitutive expression of FosB-like immunoreactivity, but statistically significant increases were detected (W vs. VC, F(1,8) = 20.60, P < ,011 (see Fig. 4B). The increase in FosB-like immunoreactivity was also detectable by immunoblotting for nucleus accumbens samples (see below). Because of the negative results in the caudoputamen at low dose levels, we examined the possibility that acute clozapine (20 mg/kg) would induce more FosB-like immunoreactivity at longer (&hour) survival times. No such augmented induction was detectable.
Single injections of haloperidol or metoclopramide induced c-Fos-like immuno- reactivity in the caudoputamen as well as in the nucleus accumbens at all dose levels. At the lowest doses (0.25 mg/kg for haloperidol, 1.25 and 2.5 mgikg for metoclopra- mide), c-Fos-positive nuclei appeared through the entire anterior nucleus accumbens and anterior caudoputamen. Farther caudally, c-Fos-positive nuclei were progressively less widely distributed in the caudoputamen, assuming a dorsolateral position. This general pattern was maintained at higher doses (0.5 and 1 mg/kg for haloperidol, 5 and 10 mg/kg for metoclopramide), but the expression in the caudoputamen became more intense and, for metoclopra- mide, even more widespread. Both haloperidol and metoclo- pramide induced c-Fos-like immunoreactivity in the entire nucleus accumbens and in both striosome and matrix compartments of the caudoputamen (Fig. lB, see also Fig. 4A: VH and VM).
The two typical neuroleptics also reliably induced FosB- like immunoreactivity in the caudoputamen and nucleus accumbens (see Fig. 4B). The distributions of FosB-positive nuclei were not identical for the two drugs, in that the induction of FosB-like protein was mainly dorsal and lateral with low doses of haloperidol and medial with low doses of metoclopramide. The induction levels in the other regions of the caudoputamen increased with increasing doses (see also Fig. 4B, VH and VM). Despite high levels of constitu- tive expression of FosB-like immunoreactivity in the nucleus accumbens, increased FosB-like immunoreactivity could be detected there as well with high doses of haloperidol and metoclopramide. Statistical analysis confirmed these obser- vations (W vs. VH, F(1,20) = 10.26, P < .01; W vs. VM, F(1,25) = 66.86, P < .01) (see Fig. 4B). No striosomei matrix patterning of FosB immunostaining was seen either with haloperidol or metoclopramide treatment (data not shown).
Haloperidol and metoclopramide.
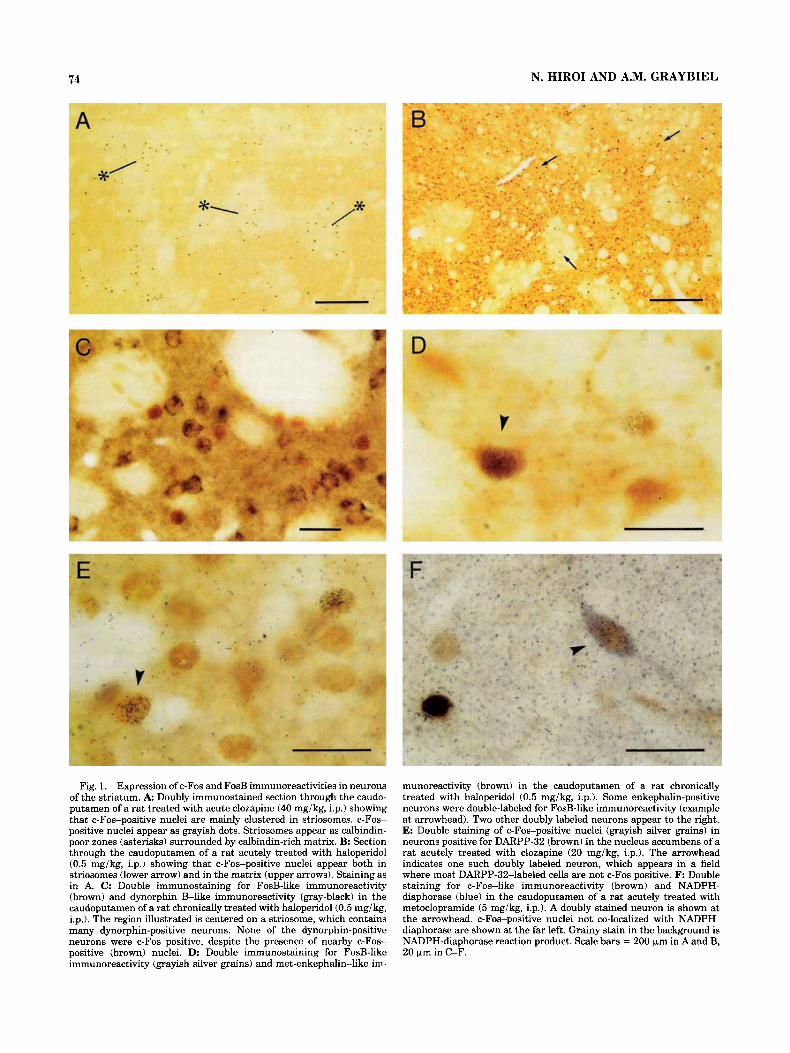
74 N. HIROI AND A.M. GRAYBIEL
Fig. 1. Expression of c-Fos and FosB immunoreactivities in neurons of the striatum. A Doubly immunostained section through the caudo- putamen of a rat treated with acute clozapine (40 mgikg, i.p.1 showing that c-Fos-positive nuclei are mainly clustered in striosomes. c-Fos- positive nuclei appear as grayish dots. Striosomes appear as calbindin- poor zones (asterisks) surrounded by calbindin-rich matrix. B: Section through the caudoputamen of a rat acutely treated with haloperidol (0.5 mg/kg, i.p.1 showing that c-Fos-positive nuclei appear both in striosomes (lower arrow) and in the matrix (upper arrows). Staining as in A. C: Double immunostaining for FosB-like immunoreactivity (brown) and dynorphin E l i k e immunoreactivity (gray-black) in the caudoputamen of a rat chronically treated with haloperidol (0.5 mgikg, i.p.1. The region illustrated is centered on a striosome, which contains many dynorphin-positive neurons. None of the dynorphin-positive neurons were c-Fos positive, despite the presence of nearby c-Fos- positive (brown) nuclei. D: Double immunostaining for FosB-like immunoreactivity (grayish silver grains) and met-enkephalin-like im-
munoreactivity (brown) in the caudoputamen of a rat chronically treated with haloperidol (0.5 mg/kg, i.p.). Some enkephalin-positive neurons were double-labeled for FosB-like immunoreactivity (example at arrowhead). Two other doubly labeled neurons appear to the right. E: Double staining of c-Fos-positive nuclei (grayish silver grains) in neurons positive for DARPP-32 (brown) in the nucleus accumbens of a rat acutely treated with clozapine (20 mg/kg, i.p.). The arrowhead indicates one such doubly labeled neuron, which appears in a field where most DARPP-32-labeled cells are not c-Fos positive. F: Double staining for c-Fos-like immunoreactivity (brown) and NADPH- diaphorase (blue) in the caudoputamen of a rat acutely treated with metoclopramide (5 mg/kg, i.p.). A doubly stained neuron is shown at the arrowhead. c-Fos-positive nuclei not co-localized with NADPH- diaphorase are shown at the far left. Grainy stain in the background is NADPH-diaphorase reaction product. Scale bars = 200 km in A and B, 20 pm in C-F.
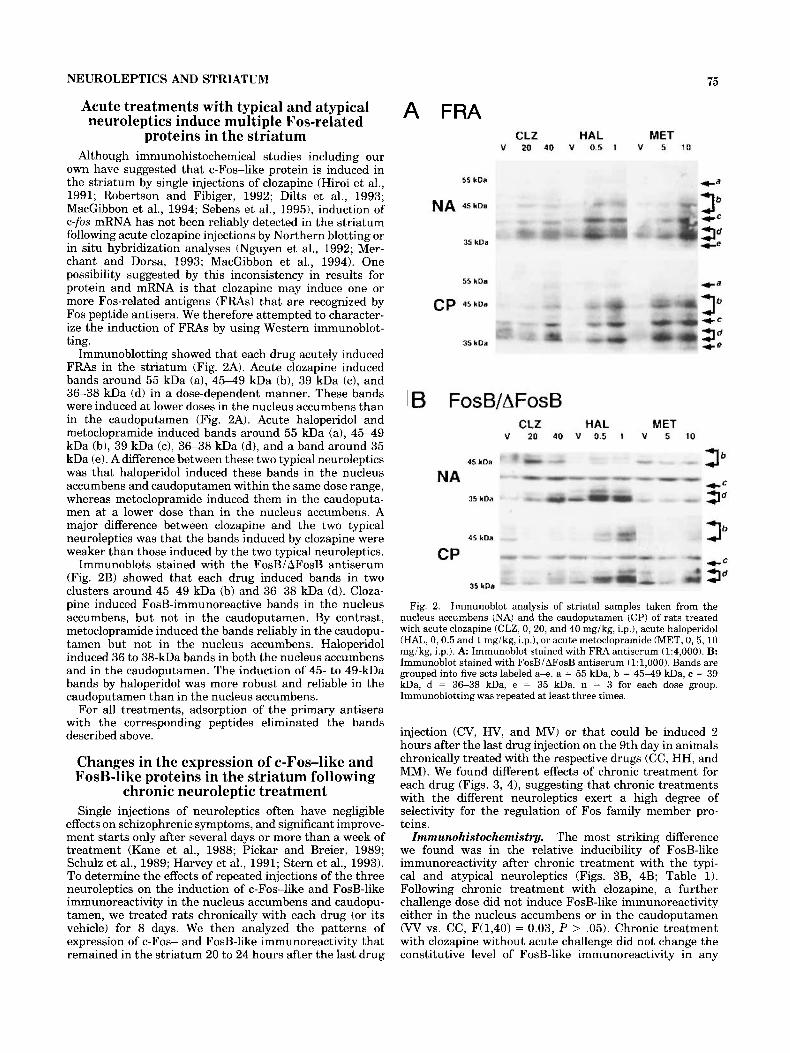
NEUROLEPTICS AND STRIATUM 75
Acute treatments with typical and atypical neuroleptics induce multiple Fos-related
proteins in the striatum Although immunohistochemical studies including our
own have suggested that c-Fos-like protein is induced in the striatum by single injections of clozapine (Hiroi et al., 1991; Robertson and Fibiger, 1992; Dilts et al., 1993; MacGibbon et al., 1994; Sebens et al., 19951, induction of c-fos mRNA has not been reliably detected in the striatum following acute clozapine injections by Northern blotting or in situ hybridization analyses (Nguyen et al., 1992; Mer- chant and Dorsa, 1993; MacGibbon et al., 1994). One possibility suggested by this inconsistency in results for protein and mRNA is that clozapine may induce one or more Fos-related antigens (FRAs) that are recognized by Fos peptide antisera. We therefore attempted to character- ize the induction of FRAs by using Western immunoblot- ting.
Immunoblotting showed that each drug acutely induced FRAs in the striatum (Fig. 2A). Acute clozapine induced bands around 55 kDa (a), 45-49 kDa (b), 39 kDa (c), and 36-38 kDa (d) in a dose-dependent manner. These bands were induced at lower doses in the nucleus accumbens than in the caudoputamen (Fig. 2A). Acute haloperidol and metoclopramide induced bands around 55 kDa (a), 45-49 kDa (b), 39 kDa (c), 36-38 kDa (d), and a band around 35 kDa (e). A difference between these two typical neuroleptics was that haloperidol induced these bands in the nucleus accumbens and caudoputamen within the same dose range, whereas metoclopramide induced them in the caudoputa- men at a lower dose than in the nucleus accumbens. A major difference between clozapine and the two typical neuroleptics was that the bands induced by clozapine were weaker than those induced by the two typical neuroleptics.
Immunoblots stained with the FosB/ AFosB antiserum (Fig. 2B) showed that each drug induced bands in two clusters around 45-49 kDa (b) and 36-38 kDa (d). Cloza- pine induced FosB-immunoreactive bands in the nucleus accumbens, but not in the caudoputamen. By contrast, metoclopramide induced the bands reliably in the caudopu- tamen but not in the nucleus accumbens. Haloperidol induced 36 to 38-kDa bands in both the nucleus accumbens and in the caudoputamen. The induction of 45- to 49-kDa bands by haloperidol was more robust and reliable in the caudoputamen than in the nucleus accumbens.
For all treatments, adsorption of the primary antisera with the corresponding peptides eliminated the bands described above.
Changes in the expression of c-Fos-like and FosB-like proteins in the striatum following
chronic neuroleptic treatment Single injections of neuroleptics often have negligible
effects on schizophrenic symptoms, and significant improve- ment starts only after several days or more than a week of treatment (Kane et al., 1988; Pickar and Breier, 1989; Schulz et al., 1989; Harvey et al., 1991; Stern et al., 1993). To determine the effects of repeated injections of the three neuroleptics on the induction of c-Fos-like and FosB-like immunoreactivity in the nucleus accumbens and caudopu- tamen, we treated rats chronically with each drug (or its vehicle) for 8 days. We then analyzed the patterns of expression of c-Fos- and FosB-like immunoreactivity that remained in the striatum 20 to 24 hours after the last drug
A FRA CLZ HAL MET
V 20 40 V 0.5 1 V 5 10
55 kDa
NA 45 kDa
35 kDa
55 kDa
cp 45 kDa
35 kDa
c=
3 c= ce 3"
3" 4 - e
B FosB/AFosB CLZ HAL MET
V 20 40 V 0.5 1 V 5 10
45 kDa
4 - c NA
35 kDa 3"
45 kDa
CP
35 kDa
Fig. 2. Immunoblot analysis of striatal samples taken from the nucleus accumbens (NA) and the caudoputamen (CP) of rats treated with acute clozapine (CLZ, 0,20, and 40 mgikg, i.p.1, acute haloperidol (HAL, 0,0.5 and 1 mgikg, i.p.), or acute metoclopramide (MET, 0,5,10 mgikg, i.p.1. A Immunoblot stained with FRA antiserum (1:4,000). B: Immunoblot stained with FosBiAFosB antiserum (1:1,000). Bands are grouped into five sets labeled a+. a = 55 kDa, b = 4 5 4 9 kDa, c = 39 kDa, d = 36-38 kDa, e = 35 kDa. n = 3 for each dose group. Immunoblotting was repeated at least three times.
injection (CV, HV, and M V ) or that could be induced 2 hours after the last drug injection on the 9th day in animals chronically treated with the respective drugs (CC, HH, and MM). We found different effects of chronic treatment for each drug (Figs. 3, 41, suggesting that chronic treatments with the different neuroleptics exert a high degree of selectivity for the regulation of Fos family member pro- teins.
Zmmunohistochemistry. The most striking difference we found was in the relative inducibility of FosB-like immunoreactivity after chronic treatment with the typi- cal and atypical neuroleptics (Figs. 3B, 4B; Table 1). Following chronic treatment with clozapine, a further challenge dose did not induce FosB-like immunoreactivity either in the nucleus accumbens or in the caudoputamen (W vs. CC, F(1,40) = 0.03, P > .05). Chronic treatment with clozapine without acute challenge did not change the constitutive level of FosB-like immunoreactivity in any
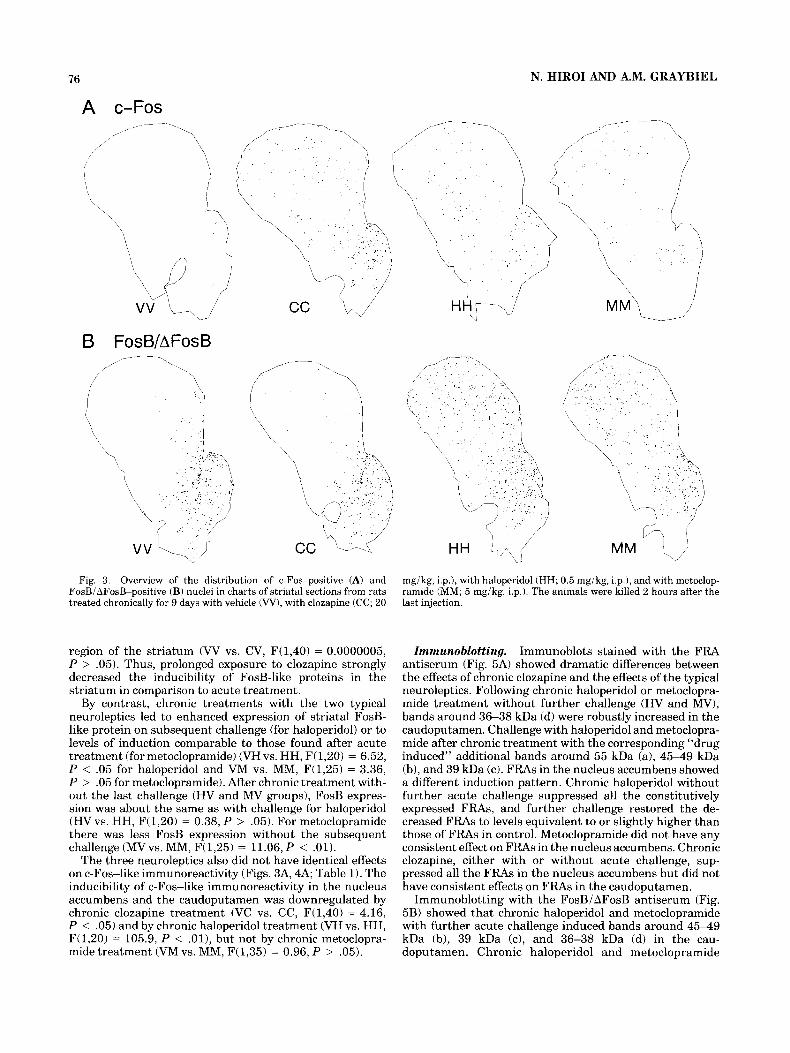
76 N. HIROI AND A.M. GRAYBIEL
A C-FOS
B FosB/AFosB
Fig. 3. Overview of the distribution of c-Fos-positive (A) and FosBiAFosB-positive (B) nuclei in charts of striatal sections from rats treated chronically for 9 days with vehicle (W), with clozapine (CC; 20
mgikg, i.p.), with haloperidol (HH; 0.5 mgikg, i.p.1, and with metoclop- ramide (MM; 5 mgikg, i.p.). The animals were killed 2 hours after the last injection.
region of the striatum (W vs. CV, F(1,40) = 0.0000005, P > ,051. Thus, prolonged exposure to clozapine strongly decreased the inducibility of FosB-like proteins in the striatum in comparison to acute treatment.
By contrast, chronic treatments with the two typical neuroleptics led to enhanced expression of striatal FosB- like protein on subsequent challenge (for haloperidol) or to levels of induction comparable to those found after acute treatment (for metoclopramide) (VH vs. HH, F(1,20) = 6.52, P < .05 for haloperidol and VM vs. MM, F(1,25) = 3.36, P > .05 for metoclopramide). After chronic treatment with- out the last challenge (HV and MV groups), FosB expres- sion was about the same as with challenge for haloperidol (HVvs. HH, F(1,20) = 0.38, P > ,051. For metoclopramide there was less FosB expression without the subsequent challenge (MVvs. MM, F(1,25) = 11.06, P < .01).
The three neuroleptics also did not have identical effects on c-Fos-like immunoreactivity (Figs. 3A, 4A; Table 1). The inducibility of c-Fos-like immunoreactivity in the nucleus accumbens and the caudoputamen was downregulated by chronic clozapine treatment (VC vs. CC, F(1,40) = 4.16, P < .05) and by chronic haloperidol treatment (VH vs. HH, F(1,20) = 105.9, P < . O l ) , but not by chronic metoclopra- mide treatment (VM vs. MM, F(1,35) = 0.96, P > .05).
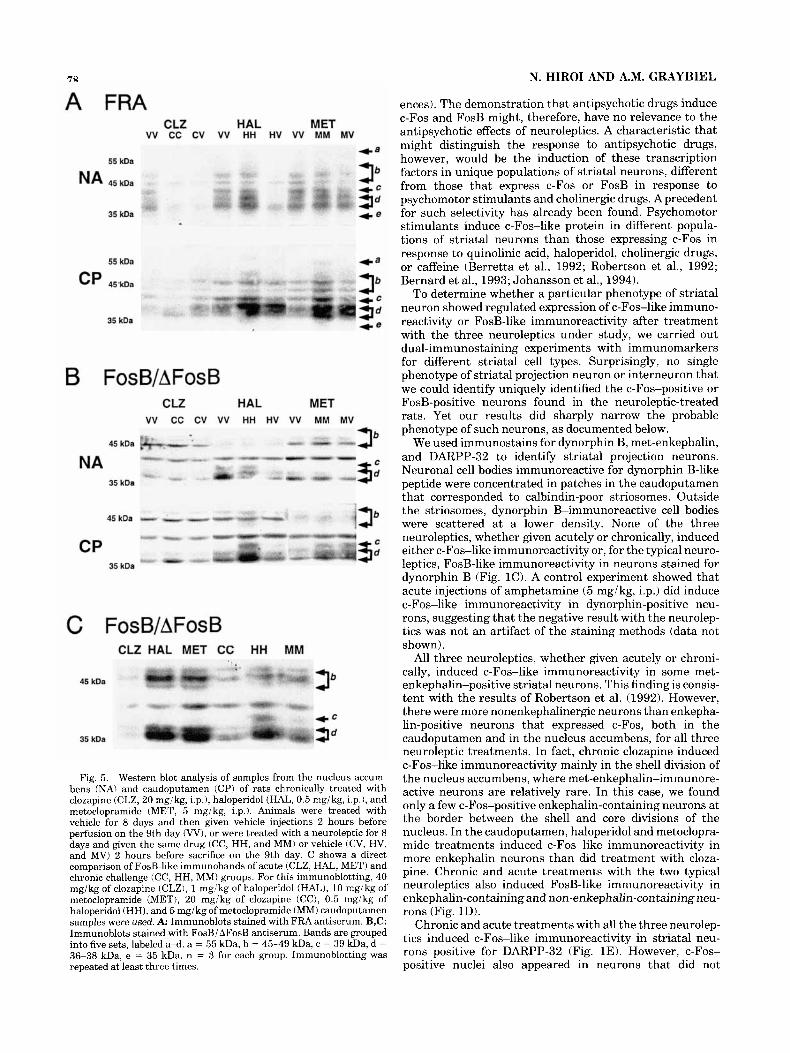
Immunoblotting. Immunoblots stained with the FRA antiserum (Fig. 5A) showed dramatic differences between the effects of chronic clozapine and the effects of the typical neuroleptics. Following chronic haloperidol or metoclopra- mide treatment without further challenge (HV and M V ) , bands around 36-38 kDa (d) were robustly increased in the caudoputamen. Challenge with haloperidol and metoclopra- mide after chronic treatment with the corresponding “drug induced” additional bands around 55 kDa (a), 45-49 kDa (b), and 39 kDa (c). FRAs in the nucleus accumbens showed a different induction pattern. Chronic haloperidol without further acute challenge suppressed all the constitutively expressed FRAs, and further challenge restored the de- creased FRAs to levels equivalent to or slightly higher than those of FRAs in control. Metoclopramide did not have any consistent effect on FRAs in the nucleus accumbens. Chronic clozapine, either with or without acute challenge, sup- pressed all the FRAs in the nucleus accumbens but did not have consistent effects on FRAs in the caudoputamen.
Immunoblotting with the FosBi AFosB antiserum (Fig. 5B) showed that chronic haloperidol and metoclopramide with further acute challenge induced bands around 45-49 kDa (b), 39 kDa (c), and 36-38 kDa (d) in the cau- doputamen. Chronic haloperidol and metoclopramide
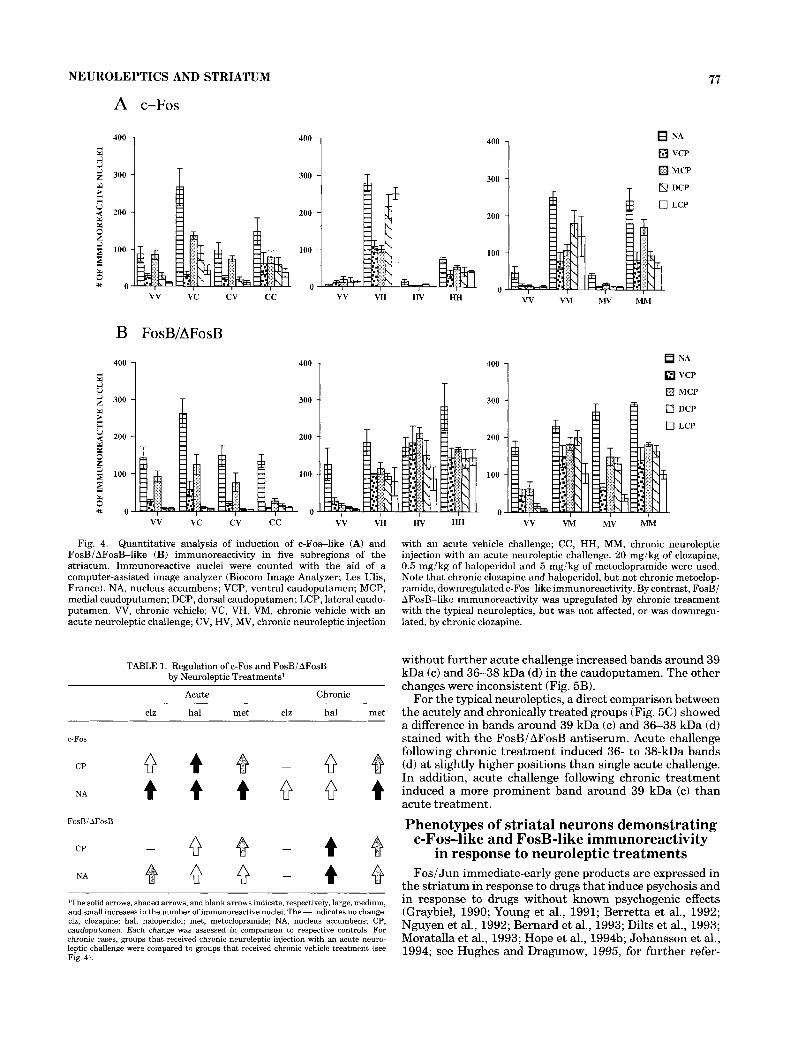
NEUROLEPTICS AND STRIATUM 77
A C-FOS
NA 400 400 400
3 VCP
2 300 300
c) U
w MCP
3011
5 8 g 100 ion 3 8
U
W
z
200 d 200 200
100
0 0 " 0
VV VRI hw MM
B FosB/AFosB
400 1 400 1 NA
VCP 3 u MCP 2 300 300 w 5 i; 200 200 2 g 100 100
2 8
0 z
0 " 0 vv vc cv cc V V V H H V H H VV VM M v MM
Fig. 4. Quantitative analysis of induction of c-Fos-like (A) and FosBi AFosB-like (B) immunoreactivity in five subregions of the striatum. Immunoreactive nuclei were counted with the aid of a computer-assisted image analyzer (Biocom Image Analyzer; Les Ulis, France). NA, nucleus accumbens; VCP, ventral caudoputamen; MCP, medial caudoputamen; DCP, dorsal caudoputamen; LCP, lateral caudo- putamen. W, chronic vehicle; VC, VH, VM, chronic vehicle with an acute neuroleptic challenge; CV, HV, MV, chronic neuroleptic injection
TABLE 1. Regulation of c-Fos and FosBiAFosB by Neuroleptic Treatments'
Acute Chronic
clz ha1 met clz hal met
c-Fos
CP 0 4 - Q NA 4 4 4 0 0 4
CP - 0 - 4 NA 0 0 - 4
FosBi AFosB
'The solid arrows, shaded arrows, and blank arrows indicate, respectively, large, medium, and small increases in the number of immunoreactive nuclei. The - indicates no change. clz, clozapine; hal, haloperidol; met, metoclopramide; NA, nucleus accumbens; CP, caudoputamen. Each change was assessed in comparison to respective controls For chronic cases, groups that received chronic neuroleptic injection with an acute neuro- leptic challenge were compared to groups that received chronic vehicle treatment (see Fig. 4).
with an acute vehicle challenge; CC, HH, MM, chronic neuroleptic injection with an acute neuroleptic challenge. 20 mgikg of clozapine, 0.5 mgikg of haloperidol and 5 mgikg of metoclopramide were used. Note that chronic clozapine and haloperidol, but not chronic metoclop- ramide, downregulated c-Fos-like immunoreactivity. By contrast, FosBi AFosB-like immunoreactivity was upregulated by chronic treatment with the typical neuroleptics, but was not affected, or was downregu- lated, by chronic clozapine.
without further acute challenge increased bands around 39 kDa (c) and 36-38 kDa (d) in the caudoputamen. The other changes were inconsistent (Fig. 5B).
For the typical neuroleptics, a direct comparison between the acutely and chronically treated groups (Fig. 5C) showed a difference in bands around 39 kDa (c) and 36-38 kDa (d) stained with the FosBi AFosB antiserum. Acute challenge following chronic treatment induced 36- to 38-kDa bands (d) at slightly higher positions than single acute challenge. In addition, acute challenge following chronic treatment induced a more prominent band around 39 kDa (c) than acute treatment.
Phenotypes of striatal neurons demonstrating c-Fos-like and FosB-like immunoreactivity
in response to neuroleptic treatments Fos/Jun immediate-early gene products are expressed in
the striatum in response to drugs that induce psychosis and in response to drugs without known psychogenic effects (Graybiel, 1990; Young et al., 1991; Berretta et al., 1992; Nguyen et al., 1992; Bernard et al., 1993; Dilts et al., 1993; Moratalla et al., 1993; Hope et al., 1994b; Johansson et al., 1994; see Hughes and Dragunow, 1995, for further refer-
78
A FRA CLZ HAL MET
VV CC CV VV HH HV VV MM MV
55 kDa
NA 45kDa
35 kDa
55 kDa
cp 4SkOa
35 kDa
B FosB/AFosB CLZ HAL MET
VV CC CV VV HH HV VV MM MV
45 kDa 3” NA
35 kDa
45 kDa
CP 35 kDa
C FosBlAFosB CLZ HAL MET CC HH MM
45 kDa 3”
5.“ 3” 52
+= 35 kDa
Fig. 5. Western blot analysis of samples from the nucleus accum- bens (NA) and caudoputamen (CP) of rats chronically treated with clozapine (CLZ, 20 mgikg, i.p.1, haloperidol (HAL, 0.5 mgikg, i.p.), and metoclopramide (MET, 5 mgikg, i.p.1. Animals were treated with vehicle for 8 days and then given vehicle injections 2 hours before perfusion on the 9th day (W), or were treated with a neuroleptic for 8 days and given the same drug (CC, HH, and MM) or vehicle (CV, HV, and M V ) 2 hours before sacrifice on the 9th day. C shows a direct comparison of FosB-like immunobands of acute (CLZ, HAL, MET) and chronic challenge (CC, HH, MM) groups. For this immunoblotting, 40 mgikg of clozapine (CLZ), 1 mgikg of haloperidol (HAL), 10 mgikg of metoclopramide (MET), 20 mgikg of clozapine (CC), 0.5 mgikg of haloperidol (HH), and 5 mgikg of metoclopramide (MM) caudoputamen samples were used. A: Immunoblots stained with FRA antiserum. B,C: Immunoblots stained with FosBi AFosB antiserum. Bands are grouped into five sets, labeled a-d. a = 55 kDa, b = 45-49 kDa, c = 39 kDa, d = 36-38 kDa, e = 35 kDa. n = 3 for each group. Immunoblotting was repeated at least three times.
N. HIROI AND A.M. GRAYBIEL
ences). The demonstration that antipsychotic drugs induce c-Fos and FosB might, therefore, have no relevance to the antipsychotic effects of neuroleptics. A characteristic that might distinguish the response to antipsychotic drugs, however, would be the induction of these transcription factors in unique populations of striatal neurons, different from those that express c-Fos or FosB in response to psychomotor stimulants and cholinergic drugs. A precedent for such selectivity has already been found. Psychomotor stimulants induce c-Fos-like protein in different popula- tions of striatal neurons than those expressing c-Fos in response to quinolinic acid, haloperidol, cholinergic drugs, or caffeine (Berretta et al., 1992; Robertson et al., 1992; Bernard et al., 1993; Johansson et al., 1994).
To determine whether a particular phenotype of striatal neuron showed regulated expression of c-Fos-like immuno- reactivity or FosB-like immunoreactivity after treatment with the three neuroleptics under study, we carried out dual-immunostaining experiments with immunomarkers for different striatal cell types. Surprisingly, no single phenotype of striatal projection neuron or interneuron that we could identify uniquely identified the c-Fos-positive or FosB-positive neurons found in the neuroleptic-treated rats. Yet our results did sharply narrow the probable phenotype of such neurons, as documented below.
We used immunostains for dynorphin B, met-enkephalin, and DARPP-32 to identify striatal projection neurons. Neuronal cell bodies immunoreactive for dynorphin B-like peptide were concentrated in patches in the caudoputamen that corresponded to calbindin-poor striosomes. Outside the striosomes, dynorphin B-immunoreactive cell bodies were scattered at a lower density. None of the three neuroleptics, whether given acutely or chronically, induced either c-Fos-like immunoreactivity or, for the typical neuro- leptics, FosB-like immunoreactivity in neurons stained for dynorphin B (Fig. 10. A control experiment showed that acute injections of amphetamine (5 mgikg, i.p.) did induce c-Fos-like immunoreactivity in dynorphin-positive neu- rons, suggesting that the negative result with the neurolep- tics was not an artifact of the staining methods (data not shown).
All three neuroleptics, whether given acutely or chroni- cally, induced c-Fos-like immunoreactivity in some met- enkephalin-positive striatal neurons. This finding is consis- tent with the results of Robertson et al. (1992). However, there were more nonenkephalinergic neurons than enkepha- lin-positive neurons that expressed c-Fos, both in the caudoputamen and in the nucleus accumbens, for all three neuroleptic treatments. In fact, chronic clozapine induced c-Fos-like immunoreactivity mainly in the shell division of the nucleus accumbens, where met-enkephalin-immunore- active neurons are relatively rare. In this case, we found only a few c-Fos-positive enkephalin-containing neurons at the border between the shell and core divisions of the nucleus. In the caudoputamen, haloperidol and metoclopra- mide treatments induced c-Fos-like immunoreactivity in more enkephalin neurons than did treatment with cloza- pine. Chronic and acute treatments with the two typical neuroleptics also induced FosB-like immunoreactivity in enkephalin-containing and non-enkephalin-containing neu- rons (Fig. 1D).
Chronic and acute treatments with all the three neurolep- tics induced c-Fos-like immunoreactivity in striatal neu- rons positive for DARPP-32 (Fig. 1E). However, c-Fos- positive nuclei also appeared in neurons that did not

NEUROLEPTICS AND STRIATUM 79
express DARPP-32. Similarly, the two typical neuroleptics, haloperidol and metoclopramide, induced FosB-like immu- noreactivity both in neurons immunoreactive for DARPP-32 and in neurons lacking DARPP-32 immunoreactivity.
Taken together, these results suggest that c-Fos- and FosB-like proteins are not induced at detectable levels in dynorphin B-positive projection neurons in response to any of the neuroleptics we tested, but that subsets of met- enkephalin-positive and DARPP-32-positive neurons do have such a response. These neuroleptics are known to differentially up- or downregulate some striatal neuropep- tides (e.g., met-enkephalin, see Graybiel, 1990; Angulo and McEwen, 1994), which makes quantitative comparisons among different treatment groups difficult. For this reason, no attempt was made to assess the percentage of these neuronal types coexpressing c-Fos or FosB.
We tested for the induction of c-Fos- and FosB-like immunoreactivity in four classes of striatal interneurons, expressing, respectively, choline acetyltransferase (CUT) , calretinin, parvalbumin, and NADPH-diaphorase (nitric oxide synthase). None of these interneuronal types showed a response to chronic or acute treatment with any of the three neuroleptics except for a small number of NADPH- diaphorase-positive neurons that did express c-Fos-like immunoreactivity in each of the drug groups (Fig. 1F). The NADPH-diaphorase-containing interneurons were not FosB positive in the haloperidol or metoclopramide-treated cases.
DISCUSSION Our findings demonstrate that typical and atypical neuro-
leptics, when given chronically, differ sharply in the tran- scription factor responses they induce in striatal neurons. Chronic treatments with typical neuroleptics strongly in- duced FosBi AFosB-like proteins, whereas chronic treat- ment with clozapine did not. Within the typical neuroleptic class, neuroleptics with high and low antipsychotic efficacy also differed in the profile of c-Fos-like and FosB-like proteins they induced in striatal neurons. We conclude that the major classifications of neuroleptic drugs based on their behavioral effects have neurobiological markers at the level of transcription factor regulation.
Acute treatments with atypical and typical neuroleptics induce region-specific expression
of C-FOS, FosBlAFosB, and other FRAs in the striatum
All three neuroleptics, given acutely, induced a FRA with an electrophoretic mobility of ca. 55 kDa, the apparent molecular weight of c-Fos. This indication that the neurolep- tics induced c-Fos contrasts with the reported difficulty in detecting increases in c-fos mRNA following clozapine treatment at the same range of doses (Nguyen et al., 1992; Merchant and Dorsa, 1993; MacGibbon et al., 1994). This contrast raises the possibility that clozapine acts at the translational level to activate the expression of c-Fos.
Acute treatments with all three neuroleptics induced other FRA bands around 45-49 kDa, 39 kDa, and 36-38 kDa, and acute treatment with the two typical neuroleptics additionally induced a 35-kDa FRA. The electrophoretic mobilities of some of these proteins correspond to those of known FRAs. The apparent molecular weights of the 45- to 49-kDa and 36- to 39-kDa bands correspond, respectively, to those of full-length FosB and an alternatively spliced form of FosB (truncated FosB or AFosB) (Mumberg et al.,
1991; Nakabeppu and Nathans, 1991). We detected bands at the same positions with FosBiAFosB antiserum, support- ing the possibility that at least some of these bands in fact correspond to FosB and AFosB. Both in the immunoblots and in our immunohistochemical study, we found stronger responses in the caudoputamen to acute treatment with the typical neuroleptics than to acute treatment with clozapine. In the nucleus accumbens, acute clozapine and haloperidol treatments induced stronger FosBi AFosB expression than did acute metoclopramide treatments (see Fig. 2 ) . These findings suggest that neuroleptics of both types acutely induce full-length FosB and AFosB in the striatum, but that the induction of these proteins is stronger with the typical neuroleptics in the caudoputamen and weaker in the nucleus accumbens. The identity of the 35-kDa FRA, not as clearly recognized with anti-FosBi AFosB antiserum as with the FRA antiserum, remains unclear.
Although the Western blot analysis indicated complex patterns of expression of different FosiFRA proteins in- duced by acute treatments with the different neuroleptics, our immunohistochemical analysis suggests that atypical and typical neuroleptics can be distinguished by the predomi- nant induction of c-Fos and FosBl AFosB in region-specific patterns within the striatum. Acute treatment with cloza- pine induced greater c-Fos and FosB expression in the nucleus accumbens than in the caudoputamen. Haloperidol and metoclopramide induced the opposite pattern of induc- tion. Previous studies have shown that atypical and typical neuroleptics differentially induce electrophysiological changes in these two striatal regions (White and Wang, 1983; Chiodo and Bunney, 1983). The present study shows that the differential effects of atypical and typical neurolep- tics can be observed even at the level of transcriptional regulation.
Chronic treatments with atypical and typical neuroleptics differ in their effects on c-Fos and
FosBlAFosB expression in the striatum Chronic treatment with typical neuroleptics strongly
regulated expression of a set of FRAs. The most prominent were bands at 36-38 kDa and at 45-49 kDa in the caudopu- tamen, expressed at augmented levels following chronic haloperidol and metoclopramide treatment. Some of these bands were detected at the same positions by FosBiAFosB antiserum, and may include truncated FosB, as discussed above. For the same reason, some of the 45- to 49-kDa FRAs induced following chronic haloperidol and metoclop- ramide with further acute challenge may correspond to full-length FosB. We also detected strong induction of the 39-kDa band with FosBiAFosB antiserum in the caudopu- tamen following chronic haloperidol and metoclopramide treatments, with or without further acute challenge. Thus, chronic treatments with haloperidol and metoclopramide appear to augment expression of AFosB and its variants in the caudoputamen. Acute challenges with haloperidol or metoclopramide after the chronic treatments additionally increase full-length FosB in the caudoputamen. The FosB- like immunoreactivity we detected by immunohistochemis- try thus seems to contain at least two different FosB species with different kinetics of inducibility and expression.
In contrast to these results for the typical neuroleptics, chronic clozapine treatment led to no increase and even downregulated inducibility of both c-Fos and FosBl AFosB in the striatum. Thus, chronic treatments with atypical and typical neuroleptics induce different sets of transcription

80
factors. These findings strongly suggest that, in addition to regional specificity of their effects in the striatum, neurolep- tics when given chronically bring about expression of unique combinations of transcription factors in the stria- tum.
FRAs recognized around 35 kDa are strongly induced by chronic treatment with a number of different agents, and a general involvement of these proteins in long-term neuro- chemical regulation has been suggested (Bronstein et al., 1994; Hope et al., 1994a,b; Chen et al., 1995). The 36- to 39-kDa FRAs that we detected with the FRA and FosB/ AFosB antisera appeared to follow the temporal kinetics of the 35-kDa FRAs that previous studies have documented.
Atypical and typical neuroleptics induce c-Fos and FosB in overlapping populations
of striatal neurons The coexpression patterns we found by double immuno-
staining demonstrate two major points. First, all three neuroleptics tend to induce c-Fos-like immunoreactivity in striatal projection neurons expressing immunoreactivity for met-enkephalin or DARPP-32. We found no such induc- tion in the class of projection neurons expressing dynorphin B-like immunoreactivity. Second, there was no obvious distinction between the classes of neurons that expressed c-Fos- or FosB-like immunoreactivity in response to treat- ments with typical and atypical neuroleptics. We also observed many c-Fos-positive nuclei in striatal neurons that did not express DARPP-32, enkephalin, or NADPH diaphorase. We were unable to identify the phenotype of these neurons.
I t should be emphasized that our results for neuropep- tides expressed in striatal projection neurons are not inclu- sive for all species of enkephalin-related and dynorphin- related peptides. These neuropeptides are derived from their precursor forms, which contain sequences correspond- ing to met-enkephalin, leu-enkephalin, and their related peptides for enkephalin (Yoshikawa et al., 1984) and dynor- phin A, dynorphin B, leumorphin and a-neoendorphin for dynorphin (Civelli et al., 1985). Our results in no way exclude the possibility that the neuroleptic treatments we administered affected neurons expressing neuropeptides other than those detected with the met-enkephalin and dynorphin B antisera we used.
We have no independent measure of what proportion of neurons expressing met-enkephalin or dynorphin B were detectable in our immunohistochemical experiments. As a consequence, some of the peptide-negative striatal neurons that we found to express c-Fos- or FosB-like immunoreac- tivity could represent false-negatives. We note, however, that chronic treatment with typical neuroleptics has been shown to upregulate met-enkephalin-like immunoreactiv- ity (see Angulo and McEwen, 1994). Nevertheless, we still observed c-Fos- and FosB-immunoreactive nuclei in met- enkephalin-negative as well as in met-enkephalin-positive neurons. Our observation that c-Fos and FosB immunoreac- tivities were expressed in DARPP-32-positive as well as DARPP-32-negative neurons also seems unlikely to be due exclusively to technical problems, as acute psychomotor stimulants induce c-Fos exclusively in DARPP-32-positive neurons with the same antiserum and staining protocols (Berretta et al., 1992). Rather, this finding suggests that neurons expressing this protein phosphatase inhibitor can express c-Fos and FosB in response to neuroleptic treat- ment, but that DARPP-32 cannot be solely involved in the
N. HIROI AND A.M. GRAYBIEL
induction cascade leading to the expression of c-Fos and FosB/ AFosB. We are least confident of complete staining of all dynorphin B-containing striatal neurons, as the immu- nohistochemistry for dynorphin B proved difficult. We nevertheless observed many dynorphin B-immunoreactive striatal neurons, yet found almost no double-labeling of these cells. We also succeeded in obtaining c-Fosidynorphin B double labeling in positive control sections from amphet- amine-treated cases.
The expression patterns we found are strikingly different from those induced by acute or chronic psychomotor stimu- lant treatment (Berretta et al., 1992; Moratalla et al., 1996). Following treatment with amphetamine or cocaine, there is robust expression of c-Fos and JunB-like immuno- reactivity in dynorphin-immunopositive projection neu- rons, but very little expression in enkephalin-positive projec- tion neurons. The one point of overlap with the expression patterns we found for neuroleptics was for DARPP-32 neurons, many of which are immunoreactive for FosiJun proteins following psychomotor stimulant treatment and some of which expressed immunoreactivity for c-Fos or FosB/AFosB in response to neuroleptic treatment. Thus, our findings strongly support the notion that different patterns of neuronal stimulation can activate different sets of immediate-early genes in different populations of neu- rons within a given neural system (Wisden et al., 1990; Berretta et al., 1992; Moratalla et al., 1993).
The dissociation we observed between the effects of chronic treatments with atypical and typical neuroleptics involved the type of transcription factor chronically induced rather than the type of striatal neurons expressing each transcription factor. c-Fos was induced by the two potent antipsychotics (clozapine and haloperidol). c-Fos showed marked downregulation after repeated treatment with halo- peridol, but metoclopramide, which has a weak antipsy- chotic effect and induces strong extrapyramidal side effects, potently induced c-Fos both acutely and chronically. Cloza- pine, given chronically, downregulated the inducibility of c-Fos. FosB immunoreactivity, on the other hand, was induced chronically by both of the two typical neuroleptics, but not by clozapine. Thus, the expression patterns of c-Fos and FosB immunoreactivities were different for the differ- ent neuroleptics. Nevertheless, both c-Fos and FosB immu- noreactivities were expressed in the same types of neurons (met-enkephalin-positive and DARPP-32-positive neu- rons). We did see some differences in responsive neuronal phenotypes on comparing the effects of acute treatments with typical and atypical neuroleptics. Clozapine induced c-Fos in the shell division of the nucleus accumbens, where met-enkephalin-positive neurons are sparse, and haloperi- do1 and metoclopramide induced c-Fos in more met- enkephalin-positive neurons than did clozapine. These drugs exert their therapeutic effects only after repeated treatment, however, at which point clozapine and haloperi- do1 induce negligible c-Fos. Thus, the therapeutic relevance of the differential induction of c-Fos in different neuronal populations is questionable.
Functional correlates of the drug-selective and treatment-selective patterns
of c-Fos and FosB expression in the striatum A long-standing view is that the antipsychotic effects of
neuroleptics are exerted through the mesolimbic dopamine system, which projects to the nucleus accumbens, the striatal region selectively innervated by the hippocampal

NEUROLEPTICS AND STRIATUM 81
formation (Kelley and Domesick, 1982). Hippocampal abnor- mality has been implicated in the etiology of positive symptoms of schizophrenia (Bogerts et al., 19931, and functional abnormality of the hippocampal-nucleus accum- bens circuit might underlie some aspects of schizophrenic symptoms (Stevens, 1973). By contrast, the motor side effects of neuroleptics are thought to depend for their action on the nigrostriatal dopamine system, which inner- vates the caudoputamen (see Anden, 1972; White and Wang, 1983; Carlsson, 1988).
Even probing only Fos-family gene products in one brain region, we found that typical and atypical neuroleptic treatments regulate different gene products in different temporal and spatial patterns for each drug. Thus, different anatomical distribution patterns of induced gene products alone, so far emphasized as distinguishing the neural responses to different classes of neuroleptics, do not ad- equately reflect the selectivity of the drugs. In particular, our findings cast doubt on the induction of c-Fos as an adequate positive marker of neuronal systems activated to bring about therapeutic efficacy of neuroleptics (Robertson and Fibiger, 1992) or the genesis of extrapyramidal side effects (Sebens et al., 1995). Our results suggest that it is more likely to be the absence than the presence of c-Fos expression that is a candidate neuronal marker for the status of neurons relevant to antipsychotic actions.
An alternative possibility for a clinically relevant positive marker is a form of FosB-like immunoreactivity which, according to our immunoblotting analysis, might be a truncated form of FosB or related protein species. Both our results and those of Dragunow (1990) show that acute haloperidol treatment induces FosB-like immunoreactivity in the nucleus accumbens and in the caudoputamen, and our work suggests that acute treatment with metoclopra- mide, another typical neuroleptic, also induces the putative truncated form of FosB in the striatum. Chronic treat- ments with haloperidol and metoclopramide, but not with clozapine, also raised the level of FosB-like immunostaining in the caudoputamen.
This pattern suggests that although the expression of FosB-like proteins (putatively including AFosB) does not parallel the antipsychotic actions of neuroleptics, it does correlate strongly with the generation of extrapyramidal side effects. Chronic treatments with the typical neurolep- tics, but not clozapine, raised the level of the putative AFosB in the caudoputamen, mirroring the delayed onset of some motor side effects the drugs induce in humans (Baldessarini and Tarsy, 1980; Ellenbroek, 1993). The induction of putative AFosB was selectively found in the anterior caudoputamen, a region through which typical neuroleptics are thought to exert their motor side effects (Ellenbroek et al., 1985).
What could be the functional effects of such differential activation? The fact that at least some enkephalin- containing striatal neurons express these changes is particu- larly interesting. Enkephalin is a peptide marker for the striatal projection neurons giving rise to the movement- diminishing indirect pathway of the basal ganglia. The downregulation of c-Fos-like protein but upregulation of FosB-like protein in these enkephalin-containing projection neurons could reflect changes in transcriptional regulation related to the genesis of motor side effects by typical neuroleptics. This is likely to be only one effect of the neuroleptics, however, for nonenkephalinergic neurons also exhibited the regulatory changes. Of more general interest
is the fact that both of the comparisons we made in this study, between typical and atypical neuroleptics and be- tween neuroleptics with potent and weak antipsychotic actions, suggest that neuroleptics distinguished by their behavioral actions induce drug-specific patterns of transcrip- tional regulation as well.
ACKNOWLEDGMENTS It is a pleasure to acknowledge the support of NIDA grant
DA08037, the Stanley Foundation, the Dystonia Medical Research Foundation, and the Hascoe Family Foundation. N. Hiroi was supported by a fellowship from the Human Frontier Science Program. We thank Drs. S. Berretta, R. Moratalla, F.-C. Liu, and Ms. D. Major and Mr. G. Holm for their technical advice, Ms. A. Rosales, Mr. A. Patel, Mr. J. Ledwith, Mr. E. Yellin, Ms. A. Siddiqui, Ms. M. Ferrell, and Ms. Z. Sachs for their excellent assistance, and Mr. H. Hall, who is responsible for the photography. We are grateful to Drs. P. Greengard, H. Hemmings, E.L. Gustafson, P.C. Emson, E. Weber, M. Sonders, and M.J. Iadarola for their generous gifts of antisera. We acknowledge the Sandoz Pharmaceutical Co. for its gift of clozapine. We dedicate this article to S.H.-S.
LITERATURE CITED Ackenheil, M., H. Beau, A. Burkhart, A. Franke, and W. Packa (1976)
Anti-psychotishe Wirksamkeit im Verhaltnis zum plasmaspiegel von Clozapine. Arzneimittel forschung 26:1156-1158.
Anden, N.E. (1972) Dopamine turnover in the corpus striatum and the limbic system after treatment with neuroleptic and anti-acetylcholine drugs. J. Pharm. Pharmacol. 24.905-906.
Anderson, K.D., and A. Reiner 11991) Immunohistochemical localization of DARPP-32 in striatal projection neurons and striatal interneurons: implication for the localization of D1-like dopamine receptors on differ- ent types of striatal neurons. Brain Res. 568:235-243.
Angst, J., D. Bente, P. Berner, H. Helmann, H. Helmchen, and H. Hippius (1971) Das klinische irkungsbild von Clozapin. Pharmakopsychiatry 4201-211.
Angulo, J.A., and B.S. McEwen (1994) Molecular aspects of neuropeptide regulation and function in the corpus striatum and nucleus accumhens. Brain Res. Rev. 19:l-28.
Baldessarini, R., and D. Tarsy (1980) Dopamine and the pathophysiology of dyskinesias induced by anti-psychotic drugs. Annu. Rev. Neurosci. 3r23-41.
Bernard, V., B. Dumartin, E. Lamy, and B. Bloch 11993) Fos immunoreactiv- ity after stimulation or inhibition of muscarinic receptors indicates anatomical specificity for cholinergic control of striatal efferent neurons and cortical neurons in the rat. Eur. J. Neurosci. 5:1218-1225.
Berretta, S., H.A. Robertson, and A.M. Graybiel (1992) Dopamine and glutamate agonists stimulate neuron-specific transcriptional activation of c-fos in the striatum. J. Neuropbysiol. 68:767-777.
Berridge, M. (1986) Second messenger dualism in neuromodulation and memory. Nature 323294-295.
Bissette, G., and C.B. Nemeroff (1985) Do neuropeptide systems mediate some of the effects of antipsychotic drugs? Prog. Clin. Biol. Res. 192.349-353.
Bogerts, B., J.A. Lieberman, M. Ashtari, R.M. Bilder, G. Degreef, G. Lerner, C. Johns, and S. Masiar (1993) Hippocampus-amygdala volumes and psychopathology in chronic schizophrenia. Biol. Psychiatry 33:236-246.
Braude, W., T . Barnes, and S. Gore (1983) Clinical characteristics of akathisia: a systematic investigation of acute psychiatric inpatient admissions. Br. J. Psychiatry 143:139-150.
Hong 11994) Role of a 35 kDa fos-related antigen (FRA) in the long-term induction of striatal dynorphin expression in the 6-hydroxydopamine- lesioned rats. Mol. Brain Res. 23:191-203.
Carlsson, A. (1988) The current status of the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 1: 179-203.
Bronstein, D.M., H. Ye, K.R. Pennypacker, P.M. Hudson, and

82 N. HIROI AND A.M. GRAYBIEL
Casey, D.E. (1989) Clozapine: neuroleptic-induced EPS and tardive dyskine- sia. Psychopharmacology (Berl.) 99:47-53.
Chen, J . 3 , H.E. Nye, M.B. Kelz, N. Hiroi, Y. Nakabeppu, B.T. Hope, and E.J. Nestler (1995) Regulation of A FosB and FosB-like proteins by electroconvulsive seizure and cocaine treatments. Mol. Pharmacol. 48: 880-889.
Chouinard, G., and L. Annable (1976) Clozapine in the treatment of newly admitted schizophrenic patients. A pilot study. J. Clin. Pharm. Ther. 16:289-297.
Chiodo, L.A., and B.S. Bunney (1983) Typical and atypical neuroleptics: differential effects of chronic administration on the activity of A9 and A10 midbrain dopaminergic neurons. J. Neurosci. 3:1607-1619.
Civelli, O., J. Douglass, A. Goldstein, and E. Herbert (1985) Sequence and expression of the rat prodynorphin gene. Proc. Natl. Acad. Sci. U.S.A. 82:42914295.
Cleghorn, J.L., R.B. Zipursky, and S.J. List (1991) Structural and functional brain imaging in schizophrenia. J. Psychiatry Neurosci. 16:53-74.
Comb, M., S.E. Hyman, and H.M. Goodman (1987) Mechanisms of trans- synaptic regulation of gene expression. Trends Neurosci. 10:473-478.
Deutch, A.Y., M.C. Lee, and M.J. Iadarola (1992) Regionally specific effects of atypical antipsychotic drugs on striatal Fos expression: the nucleus accumbens shell as a locus of antipsychotic action. Mol. Cell. Neurosci. 3:332-341.
Dilts, R.P., Jr., T.E. Helton, and J.F. McGinty (1993) Selective induction of Fos and FRA immunoreactivity within the mesolimbic and mesostriatal dopamine terminal fields. Synapse 13:251-263.
Dragunow, M. (1990) Presence and induction of FosB-like immunoreactivity in neural, but not non-neural, cells in adult rat brain. Brain Res. 533:324-328.
Ellenbroek, B.A. (1993) Treatment of schizophrenia: a clinical and preclini- cal evaluation of neuroleptic drugs. Pharmacol. Ther. 57:l-78.
Ellenbroek, B.A., M. Schwarz, K.-H. Sontag, R. Jaspers, and A. Cools (1985) Muscular rigidity and delineation of a dopamine-sensitive neostriatal subregion: tonic EMG activity in rats. Brain Res. 345:132-140.
Gerfen, C.R. (1992) The neostriatal mosaic: multiple levels of compartmen- tal organization. Trends Neurosci. 15:133-138.
Gerfen, C.R., and W.S. Young 111 (1989) Distribution of striatonigral and striatopallidal peptidergic neurons in both patch and matrix compart- ments: an in situ hybridization histochemistry and fluorescent retro- grade tracing study. Brain Res. 460:161-167.
Goelet, P., V.F. Castelluci, S. Schacher, and E.R. Kandel(1986) The long and the short of long-term memory-a molecular framework. Nature 322: 419422.
Graybiel, A.M. (1990) Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci. 13,244-254.
Gross, H., and E. Langner (1966) The active profile of a novel broad- spectrum neuroleptic of the benzodiazepine group. Wien Med. Wochen- schr. 40:814-816.
Harvey, P.D., M. Davidson, P. Powchik, J. Schmeidler, R. McQueeney, R. Kaminsky, and K.L. Davis (1991) The course and clinical predictors of treatment response in schizophrenia. Schizophr. Res. 5:161-166.
Hippius, H. (1989) The history of clozapine. Psychopharmacology (Berl.)
Hiroi, N., and A.M. Graybiel(1992) Effects of chronic clozapine treatment on induction of Fos-like proteins in the striatum. Soc. Neurosci. Abstr. 18: 1043.
Hiroi, N., and A.M. Graybiel (1993a) Typical and atypical neuroleptics stimulate contrasting patterns of neuropeptides and FOSiFRA expres- sion in the striatum. SOC. Neurosci. Abstr. 19:129.
Hiroi, N., and A.M. Graybiel (1993b) Atypical and typical neuroleptics differentially activate Fos family members in striatal neurons. Eur. J. Neurosci. [Suppl.] 6:263.
Hiroi, N., H.A. Robertson, and A.M. Graybiel (1991) Fos is induced by the antipsychotic drug clozapine in the rat striatum. SOC. Neurosci. Abstr. 17:685.
Hope, B.T., G.J. Michael, K.M. Knigge, and S.R. Vincent (1991) Neuronal NADPH diaphorase is a nitric oxide synthase. Proc. Natl. Acad. Sci. U.S.A. 88:2811-2814.
Hope, B., M.B. Kelz, R.S. Duman, and E.J. Nestler (1994a) Chronic electroconvulsive seizure (ECS) treatment results in expression of a long-lasting AP-1 complex in brain with altered composition and charac- teristics. J. Neurosci. 14:4318-4328.
Hope, B., H.E. Nye, M.B. Kelz, D.W. Self, M.J. Iadarola, Y. Nakabeppu, R.S. Duman, and E.J. Nestler (1994b) Induction of a long-lasting AP-1
[Suppl.] 99:s3-s5.
complex composed of altered Fos-like proteins in brain by chronic cocaine and other chronic treatments. Neuron 13:1235-1244.
Hughes, P., and M. Dragunow (1995) Induction of immediate-early genes and the control of neurotransmitter-regulated gene expression within the nervous system. Pharmacol. Rev. 47:133-178.
Hyman, S.E., and E.J. Nestler (19931 The Molecular Foundations of Psychiatry. Washington, DC: American Psychiatric Press.
Iadarola, M.J., C.L. Yeung, G. Draisci, and M.A. Ruda (1989) Prolonged elevation of multiple Fos immunoreactive proteins in rat spinal cord: western blot and immunocytochemical analysis. Soc. Neurosci. Abstr. 15.468.
Johansson, B., K. Lindstrom, and B.B. Fredholm (1994) Differences in the regional and cellular localization of c-fos messenger RNA induced by amphetamine, cocaine, and caffeine in the rat. Neuroscience 59:837-849.
Kalivas, P.W., L. Churchill, and M.A. Klitenick (1993) GABA and enkephalin projects from the nucleus accumbens and ventral pallidum to the ventral tegmental area. Neuroscience 57:1047-1060.
Kane, J.M., G. Honigfeld, J. Singer, H. Meltzer, and Clozaril Collaborative Study Group (1988) Clozapine for the treatment-resistant schizo- phrenic. Arch. Gen. Psychiatry 45r789-796.
Kelley, A.E., and V.B. Domesick (1982) The distribution of the projection from the hippocampal formation to the nucleus accumbens in the rat: an anterograde- and retrograde-horseradish peroxidase study. Neurosci- ence 7:2321-2335.
Kislauskis, E., and P.R. Dobner (1990) Mutually dependent response elements in the cis-regulatory region of the neurotensinineuromedin N gene integrate environmental stimuli in PC12 cells. Neuron 4:783-795.
Kubota, Y., S. Mikawa, and Y. Kawaguchi (1993) Neostriatal GABAergic interneurones contain NOS, calretinin or parvalbumin. Neuroreport 5905-208.
Lavy, S., E. Melamed, and S. Penchas (1978) Tardive dyskinesia associated with metoclopramide. BMJ 2(6105):77-78.
LeMoine, C., E. Normand, and B. Bloch (1991) Phenotypical characteriza- tion of the rat striatal neurons expressing the D1 dopamine receptor gene. Proc. Natl. Acad. Sci. U.S.A. 88:4205-4209.
MacGibbon, G.A., P.A. Lawlor, R. Bravo, and M. Dragunow (1994) Clozapine and haloperidol produce a differential pattern of immediate-early gene expression in rat caudate-putamen, nucleus accumbens, lateral septum and islands of Calleja. Mol. Brain Res. 232-32 .
Merchant, K.M., and D.M. Dorsa (1993) Differential induction of neuroten- sin and c-fos gene expression by typical versus atypical antipsychotics. Proc. Natl. Acad. Sci. U.S.A. 90:3447-3451.
Monstein, H.J. (1993) Identification of an AP-1 transcription factor binding site within the human cholecystokinin (CCK) promoter. Neuroreport 4:195-197.
Moratalla, R., E.A. Vickers, H.A. Robertson, B.H. Cochran, and A.M. Graybiel ( 1993) Coordinate expression of c-fos and junB is induced in the rat striatum by cocaine. J. Neurosci. 13:423-433.
Moratalla, R., B. Elibol, M. Vallejo, and A.M. Graybiel (1996) Network-level changes in inducible Fos-Jun proteins in the striatum during chronic cocaine treatment and withdrawal. Neuron 17:147-156.
Morgan, J.I., and T. Curran (1991) Stimulus-transcription coupling in the nervous system: involvement of the inducible proto-oncogenes fos and jun. Annu. Rev. Neurosci. 14:421451.
Mumberg, D., F.C. Lucibello, M. Schuermann, and R. Muller (1991) Alternative splicing of fosB transcripts results in differentially expressed mRNAs encoding functionally antagonistic proteins. Genes Dev. 5:1212- 1223.
Nakabeppu, Y., and D. Nathans (1991) A naturally occurring truncated form of FosB that inhibits FosiJun transcriptional activity. Cell 64:751-759.
Naranjo, J.R., B. Mellstrom, M. Achaval, and P. Sassone-Corsi (1991) Molecular pathway of pain: fosijun-mediated activation of a noncanoni- cal AP-1 site in the prodynorphin gene. Neuron 6:607-617.
Nguyen, T.V., B.E. Kosofsky, R. Birnbaum, B.M. Cohen, and S.E. Hyman (1992) Differential expression of c-Fos and zif268 in rat striatum after haloperidol, clozapine, and amphetamine. Proc. Natl. Acad. Sci. U.S.A. 89:42704274.
Paxinos, G., and C. Watson (1986) The Rat Brain in Stereotaxic Coordinates, 2nd ed. New York: Academic Press.
Pickar, D., and A. Breier (1989) Clinical and biochemical effects of neurolep- tic treatment: implications for the pathophysiology of schizophrenia. In S.C. Schulz and C.A. Tamminga (eds): Schizophrenia: Scientific Progress. New York: Oxford University Press, pp. 285-301.

NEUROLEPTICS AND STRIATUM 83
Reiner, A., and K.D. Anderson (1990) The patterns of neurotransmitter and neuropeptide co-occurrence among striatal projection neurons: conclu- sions based on recent findings. Brain Res. Rev. 15:251-265.
Robertson, G.S., and H.C. Fibiger (1992) Neuroleptics increase c-fos expres- sion in the forebrain: contrasting effects of haloperidol and clozapine. Neuroscience 46:3 15-328.
Robertson, G.S., S.R. Vincent, and H.C. Fibiger (1992) D1 and D2 dopamine receptors differentially regulate c-fos expression in striatonigral and striatopallidal neurons. Neuroscience 49285-296.
Sandell, J.H., A.M. Graybiel, and M.F. Chesselet (1986) A new enzyme marker for striatal compartmentalization: NADPH diaphorase activity in the caudate nucleus and putamen of the cat. J. Comp. Neurol. 243:326-334.
Saper, C.B. (1996) Any way you slice it: a new Journal policy for the use of unbiased counting methods. J. Comp. Neurol. 364:5.
Schulz, S.C., R.R. Conley, E.M. Kahn, and J. Alexander (1989) Nonre- sponders to neuroleptics: a distinct subtype. In S.C. Schulz and C.A. Tamminga (eds): Schizophrenia: Scientific Progress. New York: Oxford University Press, pp. 341-357.
Sebens, J.B., T. Koch, G.J. Ter Horst, and J. Korf (1995) Differential Fos-protein induction in rat forebrain regions after acute and long-term haloperidol and clozapine treatment. Eur. J. Pharmacol. 273:175-182.
Seeman, P. (1990) Atypical neuroleptics: role of multiple receptors, endog- enous dopamine, and receptor linkage. Acta Psychiatr. Scand. 82(Suppl. 358): 14-20.
Sheng, M., and M.E. Greenberg (1990) The regulation and function of c-fos and other immediate early genes in the nervous system. Neuron 41477-485.
Stern, R.G., R. Kahn, P.D. Harvey, F. Amin, S.H. Apter, and J. Hirschowitz (1993) Early response to haloperidol treatment in chronic schizophrenia. Schizophr. Res. 10:165-171.
Stevens, J.R. (1973) An anatomy of schizophrenia? Arch. Gen. Psychiatry 29: 177-189.
Sunahara, R.K., P. Seeman, H.H.M. Van Tol, and H.B. Niznik (1993) Dopamine receptors and antipsychotic drug response. Br. J. Psychiatry 163(Suppl. 22):31-38.
Vincent, S.R., and T. Johansson (1983) Striatal neurons containing both somatostatin- and avian pancreatic polypeptide (APP)-like immunoreac- tivities and NADPH-diaphorase activity: a light and electron microscopic study. J. Comp. Neurol. 21 7364-270.
Vincent, S.R., 0. Johansson, T. Hokfelt, L. Skirboll, R.P. Elde, L. Terenius, J. Kimmel, and M. Goldstein (1983) NADPH-diaphorase: a selective histochemical marker for striatal neurons containing both somatostatin and avian pancreatic polypeptide (APP)-like immunoreactivities. J. Comp. Neurol. 217:252-263.
Walaas, S.I., and P. Greengard (1984) DARPP-32, a dopamine- and adeno- sine 3':5'-monophosphate-regulated phosphoprotein enriched in dopa- mine-innervated brain regions. I. Regional and cellular distribution in the rat brain. J. Neurosci. 4:84-98.
White, F.J., and R.Y. Wang (1983) Differential effects of classical and atypical antipsychotic drugsonA9 and A10 dopamine neurons. Science221:1054- 1057.
Wisden, W., M.L. Errington, S. Williams, S.B. Dunnett, C. Waters, D. Hitchcock, G. Evan, T.V. Bliss, and S.P. Hunt (1990) Differential expression of immediate early genes in the hippocampus and spinal cord. Neuron 43303-614.
Wood, G. (1978) An adverse reaction to metoclopramide therapy. Br. J. Oral Surg. 15:278-280.
Young, S.T., L.J. Porrino, and M.J. Iadarola (1991) Cocaine induces striatal c-Fos-immunoreactive proteins via dopaminergic D1 receptors. Proc. Natl. Acad. Sci. U S A . 88:1291-1295.
Yoshikawa, K., C. Williams, and S.L. Sabol(1984) Rat brain preproenkepha- lin mRNA. cDNA cloning, primary structure, and distribution in the central nervous system. J. Biol. Chem. 259:14301-14308.





























