Embed Size (px)
Citation preview

AUXINAS
Hormônios de crescimento

Introdução Hormônios vegetais ou fitormônios: substâncias produzidas
pelas plantas que geralmente em baixas concentrações causam respostas fisiológicas.
Reguladores de crescimento são substâncias sintéticas que atuam como hormônios.
Auxinas foram o 1º grupo a ser descoberto. Final do séc. XIX, Charles Darwin observou curvaturas de
plântulas de gramíneas em resposta à iluminação lateral. Substância produzida nos ápices difundia-se de coleóptilos para
blocos de Agar. Auxina foi a denominação proposta por Frits Went (1926-
Holanda). Palavra de origem grega, auxein, que significa crescer ou
aumentar. 1946 deu-se isolamento e caracterização química do ácido indolil
-3- acético (AIA), auxina natural mais ativa de plantas.

Coleóptilos

Peter Boysen-Jensen- Fisiologista dinamarques

Arpad Paál - Hungria
Frits Warmolt Went-Fisiologista holandês

Biossíntese de AIA- ácido indolil-3-acéticoPrecursores
1) Aminoácido triptofano.
2) Indol-3-glicerol fosfato ou indol.
Locais de síntese:
Meristema apical caulinar.
Folhas jovens.
Frutos jovens e sementes.
Pouco em ápices radiculares.

GUS- gene que codifica a enzima Beta-Glucuronidase, que não ocorre em plantas.
Esse gene foi clonado ao promotor de um gene SAUR cuja expressão ocorre em locais com alta concentração de auxinas e sua expressão é dose-dependente.
Dessa forma, ele indica onde há auxinas.
A Beta-Glucuronidase, pode ser histoquimicamente marcada com corantes.
5-Bromo-4-chloro-1H-indol-3-yl β-D-glucopyranosiduronic acid

Biossíntese de auxinas a partir de triptofano

Outras auxinas naturais menos ativas que AIA. Ácido 4- cloro-indol-3-acético Ácido fenilacético Ácido indolil- 3- butírico (AIB)

Auxinas sintéticasAuxinas sintéticas

Transporte de auxinas
Transporte pode ser polar basípeto, ou seja, do ápice para a base da planta.
Ou acrópeto, do ápice radicular para o córtex radicular
Pode ocorrer de célula a célula e/ou via floema.
Nos caules, folhas e raízes pode ocorrer no parênquima vascular e através do floema.

Transporte polar em hipocótilos

Transporte polar de auxinas -Modelo quimiosmótico do transporte
As H+ ATPases de membranas celulares geram gradiente eletroquímico através da membrana.
Hidrolisam ATP e H2O.
Fazem o bombeamento de prótons H+ para paredes celulares (pH 5,0) e OH- para citosol (pH 7,0).
Parede celular
pH 5,0
Citosol
pH 7,0

AIA é um ácido fraco e lipofílico.
Nas paredes celulares predomina AIAH (protonado) devido ao pH
ao redor de 5,0.
No citosol, grande parte é AIA¯ (aniônico) devido ao pH ao redor de
7,0
AIAH entra na célula por duas vias:
1) Pela difusão simples.
2) Pelo co-transporte com H+ mediado pelas proteínas AUX
transportadoras de AIA, também chamadas de permeases.
Neste caso, usa o transporte ativo secundário.
AIA sai da base celular via proteína transportadora de efluxo PIN
(pin shaped inflorescences, isolada de Arabidopsis).

PIN

Na parede celular da parte apical da célula, ocorre o co-transporte de AIA junto ao HH++, pelos carregadores AUX ou pela difusão.
O AIA¯ sai da parte basal do citoplasma através das proteínas carregadoras PIN.

Proteínas PIN não são fixas na
membrana.
São transportadas para
compartimentos celulares.
Retornam à membrana quando
necessárias.
Seu movimento é dirigido pelo
citoesqueleto de actina.
Redistribuição lateral de PIN
ocorre após estímulos luminosos
ou de gravidade.


Mutantes de Arabidopsis pin1 apresentam falhas na produção de
proteína PIN.
Ausência de distribuição lateral de PIN.
Transporte de AIA é apenas basal.
Meristema apical da inflorescência tem haste desprovida de
órgãos florais devido ao bloqueio no transporte lateral de
auxinas.
Auxinas atuam na sinalização de eventos para organogênese
de meristema apical de inflorescência.
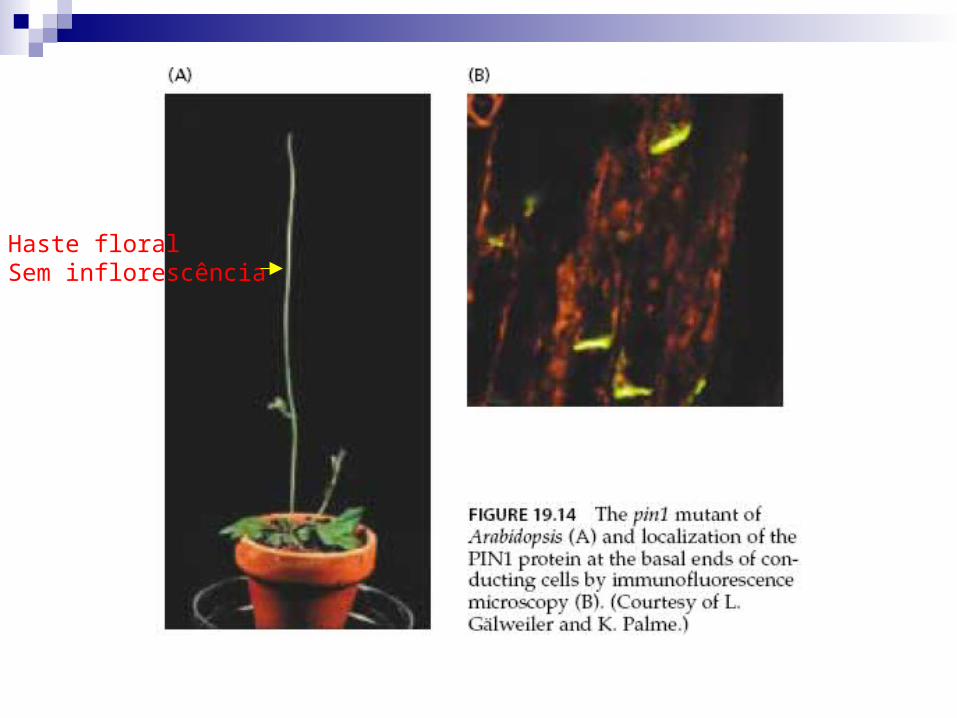
Haste floralSem inflorescência

Anatomia de raízes
Localização das proteínas AUX 1 em raízes por imunolocalização
As proteínas AUX localizam-se na columela, na parte lateral da coifa e nos tecidos do estelo .

Evidência do transporte polar de auxinas
Base caulinar

MODO DE AÇÃO DAS AUXINAS
Auxinas são substâncias lipofílicas. Têm a capacidade de atravessar membranas celulares por
difusão ou carregadores. Moléculas de auxinas precisam apresentar anel aromático para ter
atividade. Devem se ligar a um receptor protéico nas membranas. Proteínas receptoras de auxinas são chamadas de ABP
(proteínas ligadoras de auxina) e proteínas Rx. Há três sítios celulares de receptores: RE, membrana
plasmática e tonoplasto. Maior parte ocorre no RE. ABP1 tem alta afinidade e causa expansão celular. Rx tem baixa afinidade e causa divisão celular.
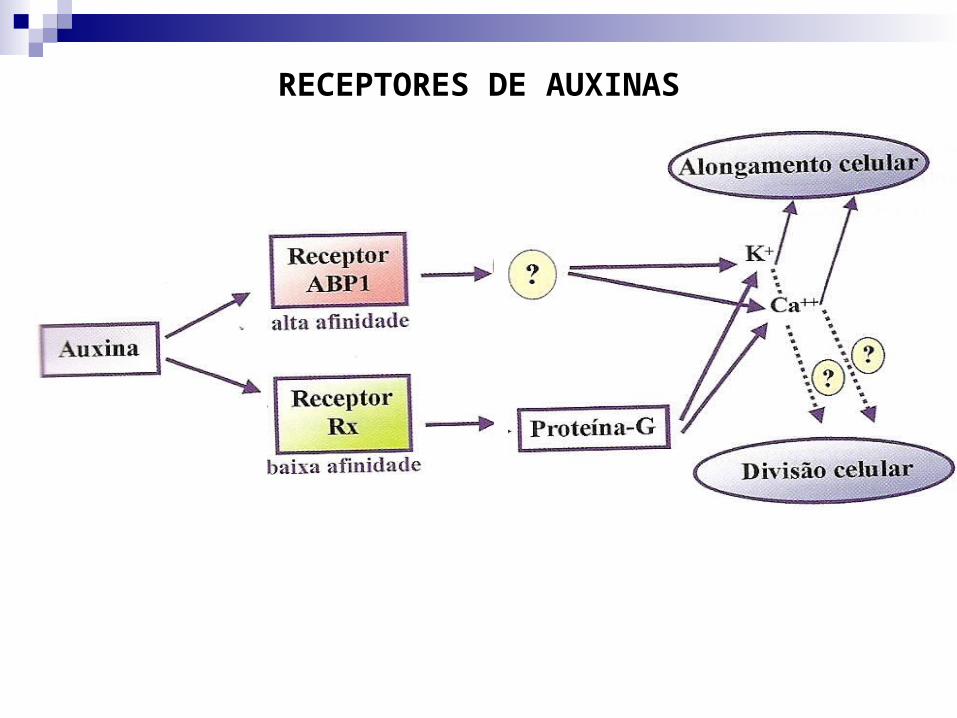
RECEPTORES DE AUXINAS

PRINCIPAIS EFEITOS FISIOLÓGICOS DE AUXINAS
Ativação de divisão celular.
Indução de crescimento celular por alongamento e expansão.
Indução de diferenciação celular.
Diferenciação de tecidos vasculares.
Desenvolvimento radicular e indução de enraizamento.
Fototropismo e gravitropismo (ou geotropismo).
Desenvolvimento de flores e frutos.
Controle de abscisão foliar.
Manutenção de dominância apical .

DIVISÃO CELULAR Auxinas + citocininas causam proliferação celular de tecidos de folhas,
raízes , caules e gemas no cultivo “in vitro”. Auxinas isoladas causam produção de calos “ in vitro” . Auxina aumenta atividade de CDK/a (quinase a dependente de ciclina)
ativada por ciclina D3 na transição de G1 para S.
Ciclina D3 (CYC/D3) é ativada por citocininas.
Ativação
CDK/a , CYC/D3

CRESCIMENTO CELULARMECANISMOS DE ALONGAMENTO CELULAR
Para a célula expandir deve ocorrer afrouxamento das paredes
celulares, por ativação enzimática.
E diminuição de Ψ celular pelo aumento de micromoléculas
solúveis.
Ocorre relaxamento de estresse ou seja, redução da pressão
hidrostática celular (ou do potencial de pressão Ψ p) e de potencial
hídrico (Ψ celular).
Ψ celular fica mais negativo.
Isto causa aumento da absorção de água que auxilia a
vacuolização celular.

Estímulo de alongamento celular em segmentos de coleóptilos de aveia

Hipótese do crescimento ácido: acidificação de paredes celulares
Ação de auxinas
AIA causa ativação de H+ ATPases da membranas
celulares.
Ou
Síntese de novas H+ ATPases nas membranas
celulares.

Modelo de ativação de H+ ATPase de membrana plasmática

Ativação ou síntese de ATPases
ATIVAÇÃO

SÍNTESE

Consequências do abaixamento de pH de paredes celulares
Ativação de hidrolases de parede celular: celulases, hemicelulases,
glucanases e pectinases.
Hidrólise de polímeros de parede: celuloses e hemiceluloses.
Deslizamento de polímeros da parede celular.
Produção e ação de proteínas expansinas que quebram pontes H
entre microfibrilas de celulose e hemicelulose.
Aumento de absorção de água e de solutos, principalmente K+.
Em coleóptilos de milho há aumento de 5 a 7 X na expressão de
genes de canais de K+ triplicando os canais e aumentando K+
celular.
Crescimento de caules e hipocótilos estimulado em
concentrações de 10-6 a 10-5 M de AIA.



PAREDE CELULAR –CÉLULAS DE GAMETÓFITOS DE Acrostichum danaeifolium Langsd. , Fisch. (Polypodiopsida, Pteridaceae)

Aumento de K+
Aumento de absorção de água
Aumento de potencial de pressão

Diferenciação radicular e de tecidos vasculares
Auxinas podem ser transportadas dos meristemas apicais caulinares para raízes.
Ocorre também síntese de auxinas nos ápices de raízes. Raízes laterais são formadas a partir de células do periciclo
sensíveis às auxinas. Primórdio radicular atravessa córtex e emerge pela epiderme. Crescimento de raízes é estimulado em concentrações de 10-
10 a 10-9 M de AIA e inibido em concentrações maiores. Raízes adventícias formam-se em caules ou pecíolos devido ao
estresse (via etileno), como por exemplo, alagamentos. Células sofrem desdiferenciação. Entram no ciclo celular, sofrem divisão e diferenciação com
formação de um novo meristema radicular.

Indução de raiz lateral
Diferenciação de tecidos vasculares: auxinas produzidas nos meristemas caulinares apicais e citocininas produzidas nos meristemas apicais de raízes induzem diferenciação de xilema e floema.

Estímulo do enraizamento de estacas caulinares/foliares.
Auxinas estimulam enraizamento caulinar. Raízes adventícias são formadas no periciclo caulinar.
500 mg L-1 1000 mg L-1 2000 mg L-1 4000 mg L-1


Indução de tecido vascular


Diferenciação de raízes em cultura de tecido.
Crescimento de raízes estimulado com 10-10M e 10-9 M de AIA e inibido a partir de 10-8 M, possivelmente devido à síntese de etileno.
Folha
caloraiz

Fototropismo
Fototropismo é a curvatura de caules e coleóptilos em direção a um
estímulo luminoso.
Ocorre transporte assimétrico de AIA em resposta ao estímulo
luminoso lateral.
A concentração de AIA torna-se maior no lado mais sombreado,
que cresce mais.

Evidência de distribuição lateral de auxinas pelo estímulo luminoso

• Luz azul é absorvida pelas fototropinas (flavoprotéinas autofosforiláveis) que causam transporte assimétrico de auxinas.• Isso gera maior crescimento no lado sombreado, onde há mais
AIA.

Gravitropismo ou geotropismo
Acúmulo de AIA na zona de alongamento da raíz.
Em raízes na posição horizontal auxinas da coifa migram para
parte inferior.
Proteínas PIN3 acumuladas no lado inferior aumentam transporte
de AIA.
Aumentos de concentração de AIA causam inibição de
crescimento celular em raízes.

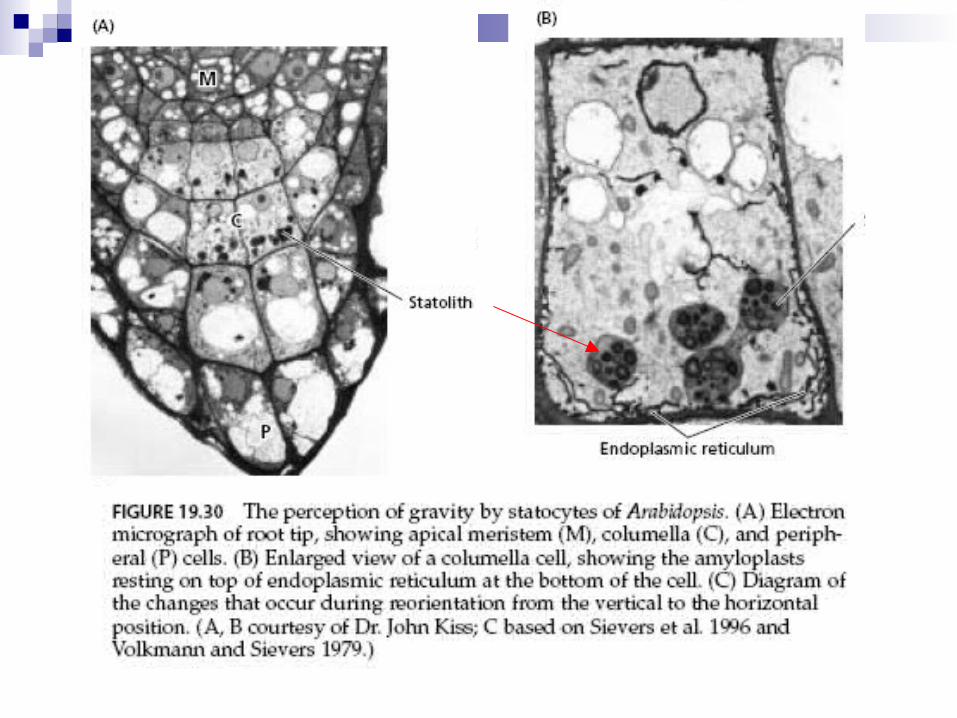
Gravitropismo ou geotropismo

Teoria da tensogridade - integridade tensiva Raízes possuem na coifa, células especiais, os estatocitos com
estatolitos (grãos de amido móveis) que percebem a orientação
gravitacional.
Sedimentação de estatolitos causa rompimento de citoesqueleto,
mudança da tensão sobre RE, acúmulo de proteínas PIN 3 e
canais de Ca2+ na membrana.
Ocorrem alterações de atividades celulares.
Raízes crescendo verticalmente - PIN3 tem distribuição uniforme nas
células da columela e AIA também.
Partes aéreas (caules e ramos) – têm bainha de amido circundando
tecidos vasculares, que se orienta conforme a gravidade.




Dominância apical
Dominância apical é a inibição do crescimento das gemas axilares
pelo meristema apical.
Bloqueio de divisão celular e alongamento celular nas gemas axilares.
Auxinas produzidas pelos meristemas apicais caulinares bloqueiam a
divisão celular das gemas axilares.
Parece haver a ativação de genes envolvidos com inibição de divisão
celular.
A remoção da gema apical reduz os níveis de auxinas e as gemas
axilares se desenvolvem.

Em plantas intactas, as gemas axilares não se desenvolvem.Quando o ápice é removido, as gemas axilares crescem.Quando auxina em lanolina é colocadaNo ápice, as gemas axilares não desenvolvem

Crescimento de frutos e indução de floração Indução de crescimento de frutos: ex: morango.
a) Fruto normalb) Fruto com remoção de aquêniosc) Fruto com remoção de uma fileira de aquênios

Promoção de floração em bromeliáceas. Promoção de flores femininas em plantas dióicas (via etileno), em cucurbitáceas, por
exemplo. Auxinas estimulam a síntese de etileno

Referências bibliográficas consultadas
Kerbauy, G.B. 2004. Fisiologia Vegetal. Guanabara
Koogan, 452p.
Raven, P.H., Evert,R.F. & Eichhorn, S.E. 2001. Biologia
Vegetal, 6ª Edição, Guanabara Koogan.,906 p.
Taiz ,L. & Zeiger, E. 2004. FISIOLOGIA VEGETAL. 3ª
EDIÇÃO. ARTMED, 719P.
Taiz, L. & Zeiger, E.2006. Plant Physiology. Sinauer
Associates, Inc, Publishers, 705p.





























