Embed Size (px)
Citation preview

Axis formation in zebrafish
Wolfgang Driever
Massachusetts General Hospital and Harvard Medical School, Boston, USA
Recent advances in our understanding of axis formation and patterning in zebrafish relate the developmental mode of this aspiring genetic model
organism to higher vertebrates. The effect of UV irradiation and lithium
treatment, as well as detailed early lineage analyses, have shed some light
on dorsoventral axis formation. However, the molecular mechanism of axis
formation, as well as the identity of a fish Nieuwkoop center, are still open
issues. A Vgl homolog is expressed in zebrafish, and activin as well as the
mouse nodal gene product have been demonstrated to induce mesoderm and
ectopic axes, respectively, in zebrafish. The zebrafish organizer is defined by
the expression domains of goosecoicf, axial, and liml. The cyclops gene is
involved in maintaining goosecoid expression in axial mesoderm of the
head. Large mutagenesis screens provide the basis for a genetic analysis of
axis formation.
Current Opinion in Genetics & Development 1995, 5:61 O-61 8
Introduction
All vertebrate zygotes, after fertilization, undergo a series of cell divisions to generate a mass of cells from which
the embryo will form. The first patterning step is the establishment of the embryonic axis. At a morphological
level, vertebrate embryos display diversity at this stage.
Some zygotes, such as Xenopus, initially possess clear animal/vegetal polarity and develop dorsoventral polarity
during the first cleavage. For other vertebrates, however, the axes are not predictable before the onset of
gastrulation. At the molecular level, though, there are
clear indications for common themes in axis formation. Many different vertebrate embryos respond in a similar
fashion to the application of inducing agents, such as
lithium, fibroblast growth factor (FGF) or transforming growth factor (TGF)p family members. Gene expression
patterns are homologous, and mutations in homologous
genes result in similar phenotypes in zebrafish and mice
[l-3]. Comparative studies among different vertebrates
have been very important in identifying common basic principles of axis formation.
Traditional vertebrate experimental systems provided
either access to genetic manipulation (mice) or em-
bryological manipulation (chicken, frogs, etc.). Over
the past decade, the zebrafish (Danio rerio) has become
popular for developmental studies because its short
generation time and high fecundity make mutational analysis possible, and the small translucent embryos
are excellent subjects for embryological studies and manipulations. In this review, I will focus on the progress
made during the past two years in our understanding of
axis formation and patterning in zebrafish and will relate this work to classic studies on pattern formation in fish.
Early post-fertilization development
Zebrafish develop in a manner typical of teleosts, and morphological aspects of their development and a normal table of developmental stages have been described in detail [4*]. When eggs are laid, yolk and cytoplasm are intermixed and the egg is surrounded by
a transparent chorion (Fig. 1). The animal/vegetal axis is preset during oogenesis and sperm can enter the egg
only at the future animal pole through the micropyle, a specialization in the otherwise sperm-impermeable chorion [5]. After fertilization, cytoplasm streams to
the animal pole as it segregates from the yolk. About
30 minutes after fertilization, the cytoplasm forms the
blastodisc at the animal pole and surrounds the vegetal
yolk mass as a thin yolk cytoplasmic layer.
At 40 minutes post-fertilization, the first meroblastic
cleavage occurs. Four more cleavages occur in stereo-
typic orientation at 15-minute intervals, followed by five
synchronous, but not oriented, cleavages. During these cleavages, the marginal vegetal blastomeres maintain
large cytoplasmic bridges with the yolk cell. These 10
cleavages generate a mound of blastomeres on top of
the vegetal yolk cell. Subsequently, during mid-blastula
transition, activation of zygotic transcription coincides
with the generation of the first three separate lineages of the embryo [6,7]. Two of these lineages, the enveloping
layer forming the outer surface of the blastoderm and the yolk syncytial layer deriving from the collapse of
vegetal marginal blastomeres into the yolk cell, constitute extraembryonic lineages. The third lineage, termed the
deep cell layer, will form the embryo proper. At the
end of the mid-blastula transition, the first coordinated cell movements occur in the embryo. The cells of the
610
Abbreviations
CNscentral nervous system; FGF-fibroblast growth factor; TCF-transforming growth factor.
0 Current Biology Ltd ISSN 0959-437X

Axis formation in zebrafish Driever 611
blastoderm spread vegetal-ward over the yolk cell during epiboly [8,9*,10]. No dorsoventral asymmetry has been detected during these stages.
Castrulation
Shortly before the onset of gastrulation movements, the future dorsoventral axis becomes obvious through an asymmetry in the thickness of the blastoderm [ll]. The formation of the hypoblast (the mesendodermal germ layer) is initiated on the dorsal side of the embryo and soon continues all around the margin. Formation of the hypoblast leads to appearance of the germ-ring, a thickened marginal region all around the blastoderm rim. On the dorsal side forms a pronounced thickening, the embryonic shield [12]. Gastrulation in zebrafish is characterized by movement of single cells rather than coherent cell layers. In clear distinction to the involution of layers of cells in amphibia, ingression might be a more precise term to describe the movements of cells during hypoblast formation [10,13*,14,15*].
Soon, molecular differences between epiblast and hy- poblast can be detected. For instance, ~0~2 transcripts, present throughout the blastula, become restricted to the epiblast during gastrulation [ 16,171. As gastrulation continues, the hypoblast extends toward the animal pole, while epiboly expands both layers towards the vegetal pole. At the same time, both hypoblast and epiblast cells converge from ventral and lateral positions toward the dorsal side. Mediolateral intercalation of cells converging to the dorsal side leads to elongation of the embryonic shield along the anteroposterior axis of the embryo [12,18]. At the end of epiboly, the embryo extends along the dorsal side of the yolk sphere, with the head positioned at the former animal pole and the tailbud developing at the former vegetal pole of the egg.
Axis formation, fate maps and determination
At the onset of gastrulation, analysis of cellular fates reveals that the endoderm will derive from the vegetal- most marginal blastomeres (Fig. 1). Mesoderm forms from the vegetal one-third of the blastoderm, whereas ectoderm originates from the animal half of the blastoderm. Neuroectoderm in particular derives from the dorsal section of the animal half [19]. Notochord derives from the dorsal side, where the shield forms, whereas somitic mesoderm, blood and heart develop fi-om lateral and ventral positions, respectively (see also [20]). The organization of the zebrafish fate map is similar to that of Xenoplrs [19].
Attempts have been made to establish a fate map based on the first cleavages, which are of stereotype orientation and, at the eight-cell stage, produce an asymmetric array of two x four cells on top of the yolk cell [21]. Individual blastomeres at the eight-cell stage were labeled with different fluorescent lineage tracers. A correlation between the second cleavage plane and the dorsoventral axis was suggested, but three independent
studies were later able to demonstrate that no correlation exists between any of the early cleavage planes and the dorsoventral axis [22”,23”,24], confirming earlier studies [25]. These earlier studies scored the position of dorsal tissue at the shield stage (rather than at the end of somitogenesis) [21], which afforded a less ambiguous assignment of dorsal position. A mechanism by which localized (dorsal?) determinants are distributed in a stereotype fashion among the early blastomeres in correlation to the cleavage planes can be excluded from being involved in axis formation in zebrafish.
Portions of the zebra&h embryo retain their relative position in the early embryo from blastula to gastrula and, consequently, rudiments of a fate map can be observed prior to gastrula stages [23”,26,27]. For instance, the descendants of central blastomeres (located at the center of the blastoderm when viewed from the animal pole) at the 64-cell stage frequently end up in the animal-most portion of the early gastrula and give rise to neural structures, as well as ectoderm and mesoderm. The fate of central blastomeres is obscured, as their descendants are subject to extensive cell mixing during epiboly. In contrast, marginal blastomeres are subject to less extensive mixing [28-l. Thus, it has been possible to demonstrate that cardiac progenitors are located at the margin of the blastula-stage embryo [20,29].
Very detailed fate maps have been generated for the shield region [13*] and for the neuroectoderm at the onset of gastrulation [30*]. For the epiblast of the shield region, some degree of radial organization has been observed: the outermost cell layer frequently gives rise to endoderm, the second to notochord, and the third to neuroectoderm. In contrast to amphibians, cell lineages present in the epiblast of the shield region (neuroectodemal, notochordal, endodermal and, to a lesser extent, somitic) are spatially less well segregated and represent intermingled populations of cells. Whether these findings relate to different patterning mechanisms is unknown. Within the neuroectoderm, a predictable order in the precursors of presumptive major brain sub- divisions indicates that cell mixing in the neuroectoderm is less pronounced [30*].
When do cells become irreversibly committed to a spe- cific fate? Heterotopic and heterochronic transplantation experiments [31] reveal that cells are not committed to a certain fate at late-blastula stages (5 hours post-fertil- ization), when tissue-restricted lineages arise. However, when hypoblast cells are transplanted heterotopically to the epiblast at mid-gastrula (8 hours post-fertilization), they will predominantly give rise to hypoblast fates. Thus, cells become committed to a specific germ layer at mid-gastrula stages. Earlier tissue-specific lineage restrictions do not necessarily reflect the state of commitment of the cell.
Dorsoventral patterning
Establishment of the dorsoventral axis is best understood in Xenopus (reviewed in [32,33]). Cortical rotation
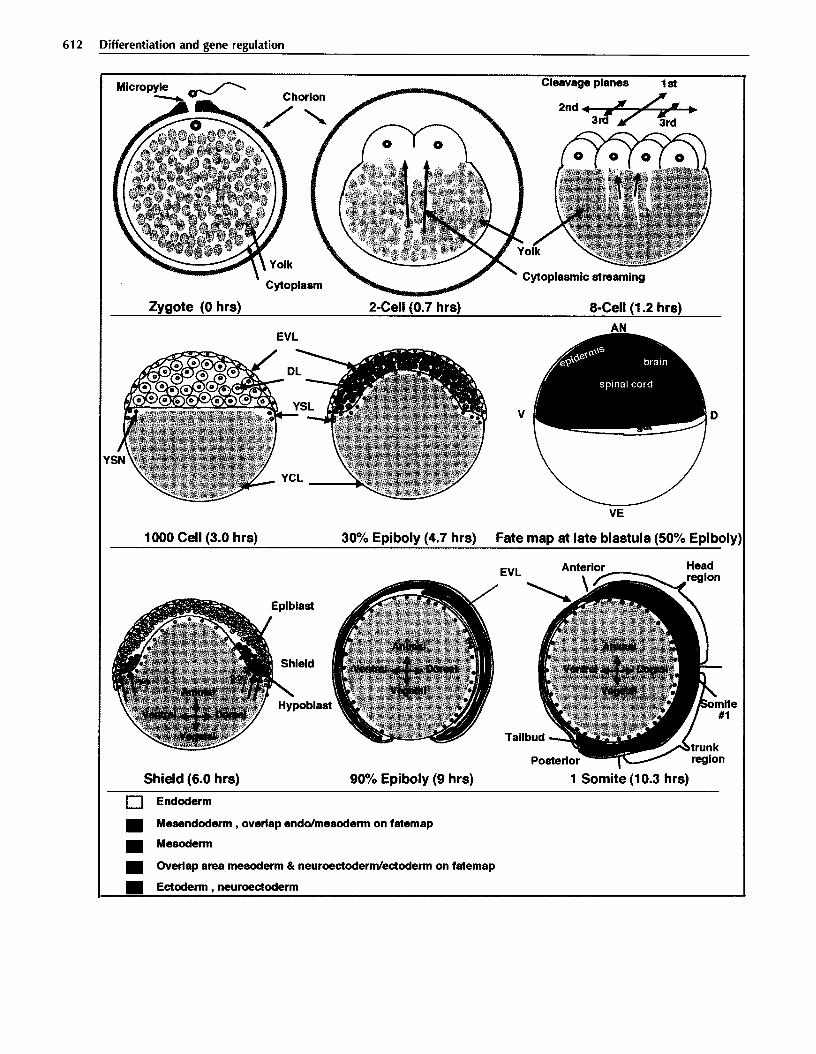
612 Differentiation and gene regulation
Micropy’- _ Chorlon
Cleavage planes
’ Cytoplasm
Zygote (0 hrs) 2-Cell(0.7 hrs) a-Cell (1.2 hrs)
AN
VE
1000 Cell (3.0 hrs) 30% Epiboly (4.7 hrs) Fate map at late blastula (50% Epiboly
Shield (6.0 hrs) 90% Epiboly (9 hrs)
Anterior Head region
He 1
1 Somite (10.3 hrs)
0 Endoderm
q Mesendoderm , overlap enddmesoderm on tatemap
n Mesoderm
n Overlap area mesoderm C neuroectodermkctoderm on fatemap
n Ectoderm , neuroectoderm

Axis formation in zebrafish Driever 613
during the first cell cycle is microtubule dependent, W sensitive, and results in the formation of the Nieuwkoop center in dorsal vegetal blastomeres. The Nieuwkoop center appears to induce the Spemann organizer in the dorsal blastopore lip, which again is the source of signals patterning the dorsoventral and anteroposterior axes. Are there similarities to zebrafish development?
W irradiation of zebrafish eggs between 10 and 25 minutes after fertilization depletes dorsal structures and results in radially symmetric embryos [8]. Within the zebrafish yolk cytoplasmic layer at that stage, there is a prominent parallel array of microtubules oriented in animal/vegetal direction [8]. However, the target of W action is unknown-so far, no evidence exists that cortical rotation takes place in zebrafish, and the target of W action could be maternal RNAs as well as the microtubules.
Similarities to amphibia are evident in the sensitivity to lithium chloride exposure during cleavage stages, which results in dorsalized zebrafish embryos [34]. Upon lithium treatment, dorsal genes such as postmid are expressed all around the margin of the gastrula, whereas the expression of ventral and posterior genes such as eve1 are suppressed [35]. In lithium-treated radialized embryos, which appear to have no dorsoventral axis, the anteroposterior order of gene expression is maintained between the animal pole and the vegetal margin at late-gastrulation stages [36-l. The lithium-sensitive period is between the 16-cell and 1024-cell stage [34], before mid-blastula transition. Therefore, the activities of maternal factors appear to be responsible for the initiation of dorsoventral axis formation.
Experiments performed in other teleosts indicate a possible involvement of the yolk syncytial cell in the specification of dorsal fates. Transplantation of younger rainbow trout blastoderms onto gastrula-stage yolk syncytial cells, from which the blastoderm had been removed, led to development of dorsal structures by those cells located atop the dorsal side of the host yolk cell [37]. Further, the capacity of the blastoderm to differentiate autonomously after separation from the yolk has been tested for several teleosts. Fundulur blastoderms from embryos older than 32-cell stage [38] and salmon or loach blastoderms Gem embryos after mid-blastula
stage [39,40] differentiate into embryonic structures, often with well defined axes. In cgntrast, younger blastoderms were reported to form morphologically less differentiated cell aggregates, sometimes with columnar epithelia, which might indicate ectoderm. In a few cases, incompletely differentiated nervous tissue was reported. Oppenheimer [38] suggested that some substances are required to be passed on from the yolk cell to the blastomeres to induce differentiation. Whether these postulated substances are Nieuwkoop center derived dorsal signals and/or general mesoderm inducers remains to be demonstrated.
Other observations are consistent with an involvement of the yolk syncytial cell in axis formation. Blastomeres appear to have equal developmental potentials prior to onset of gastrulation [41], and transplantation of blastomeres prior to gastrulation never results in the induction of a second axis in hosts. The developmental modes of some other teleosts make it difficult to consider dorsalizing determinants in blastomeres. Teleosts with annual lifestyles often go through a phase of dispersion of blastomeres at the end of epiboly, and blastomere move- ment is amoeboid [42,43]. Reaggregation at one site on the yolk syncytial cell precedes embryo formation. Reaggregation could be guided by specialization of a region of the yolk cell, or by sorting out of blastomeres that would have differentiated into separate lineages before the migratory phase, similar to chick mesoderm formation [44].
So far, no zebrafish mutations have been described in maternal-effect genes involved in the establishment of the dorsoventral axis. The maternal-effect mutation jantrs can give rise to axis duplications in the embryo [22**]. Mutant blastoderms tend to split at the 4-16-cell stage into two half-blastoderms, which remain next to each other and continue cleavage and epiboly independently. Only in the fraction of embryos with dorsal structures induced at the place where both blastoderms touch each other are two embryonic shields formed adjacent to each other, resulting in axis duplications. It is important to note that only one dorsal center is induced per embryo, not one per blastoderm. It is not yet known from where the dorsal-inducing signal (the zebrafish homolog of the Nieuwkoop center?) derives.
Fig. 1. Early development of zebrafish. The development of zebrafish embryos from fertilization to the end of gastrulation is represented by schematic drawings of mid-saggital sections of the embryos. Developmental times are in hours post-fertilization at 28’C. Only for the zygote and the two-cell embryo is the chorion shown. Fertilization occurs at a structurally specific site, the micropyle. In the zygote, yolk (grey) and cytoplasm (light blue) are mixed, but separate during the first two hours of development by cytoplasmic streaming to the animal pole (blue arrows). The stereotypical cleavage planes are indicated above the eight-cell embryo. The lOOO-cell embryo represents the mid-blastula. The different embryonic and extraembryonic lineages can be clearly distinguished: DL, deep layer (embryo proper); EVL, enveloping layer; YSL, yolk syncytial layer; YSN, yolk syncytial nuclei; YCL, yolk cytoplasmic layer. Fate map at 50% epiboly, just before the onset of gastrulation: V, ventral; D, dorsal; AN, animal pole; VE, vegetal pole. During fate-mapping experiments, significant regions of overlap have been found. Orange represents areas with cells that give rise to both endodermal and mesodermal fates. Purple represents areas where cells with both mesodermal and ectodermal or neuroectodermal fate were found. At the shield stage, gastrulation movements have created the epiblast. For the first time, the dorsoventral axis can be identified by a morphological criterion: the thickening of the embryonic shield on the dorsal side. The red arrows represent hypothetical mesoderm-inducing signals, the green arrow a dorsal-inducing signal. At 90% epiboly during late gastrula, the hypoblast has almost reached the animal pole. Convergence of cells to the dorsal side results in a thickening of the embryonic axis. On the ventral side of the yolk, only very thin layers remain. At the one-somite stage, gastrulation is complete and the major regions along the anteroposterior axis (head, trunk, and tail) can be distinguished. The organization of germlayers in the tailbud (green) is not known for zebrafish.

614 Differentiation and acne regulation
If the zebrafish Nieuwkoop center were a structure
autonomously generated during blastula stages in the
blastoderm itself, one would expect to find two spatially
separated dorsal centers in some of thejanus blastoderms,
which is never the case. However, a Nieuwkoop center
could form during the first cleavages and segregate
among prospective dorsal marginal blastomeres or with
the yolk syncytial cell. Whether maternal gene products
are localized in the zebrafish yolk syncytial cell, and
whether there is any contribution to patterning by
transcription from the yolk syncytial nuclei, needs to be
determined.
Mesoderm induction
In Xenopus, activin, Vg-1, wnt-1 1, bone morphogenetic
protein 4, and FGF have been suggested to be major
components of the mesoderm-induction pathway (for review, see [45]). In contrast to the vegetal localization
of Vgl (DVR-1) mRNA in Xenopus, the maternally expressed zebrafish zDVR-1 mRNA is equally dis-
tributed among all blastomeres, but appears to be absent from the vegetal yolk cell [46]. Processing of the inactive Vgl precursor is highly controlled, and
the active form can induce dorsal mesoderm [47].
As zDVR-1 is processed in Xenupus and can induce mesoderm, it is likely to be a true Vgl homolog
(C Dohrmann, DS Kessler, DA Melton, personal communication). The reason for the difference in
localization between zebrafish and Xenopus is unknown. Helde and Grunwald [46] suggest that the indeterminate
state of the zebrafish blastomeres and the more extensive cell mixing, when compared to Xenopus, require that all blastomeres harbor equivalent maternal information for
developmental potential. Therefore, it could be possible
that zDVR-1 is processed to the active form in only a subset of the zebrafish blastomeres.
The mouse nodal gene product, another member of the
TGFB family, when injected into zebrafish embryos,
causes the formation of ectopic shields and, subsequently, axes including notochord and somites, but no head
duplications [48*]. The mouse nodal gene is disrupted
in a mouse mutant deficient in mesoderm formation
[49]. The axis-determining function of nodal might
be mediated by activating early-response genes such as
~~yoose& and lirlr 1 [48’].
The role of zebrafish activin has been studied in embryos
of a similar teleost, medaka [50-l. Activin PB is expressed during oogenesis and from the late-blastula stage on.
To distinguish between maternal and zygotic activin,
distinct mutant forms were generated. Embryos injected with RNAs encoding dominant-negative processing-
defective activins, which act upon cotranslation on the
cellular form by dimer formation, do not generate a mesoderm-deficient phenotype in fish. In contrast, dominant-negative activins, which appear to interfere with the activin receptors, do result in mesoderm-defi-
cient phenotypes. The authors conclude that maternally derived activin receptor activating activity might be
sufficient for mesoderm formation. However, as the
dominant-negative activin might interfere with TGFP
family receptors other than activin receptors, a specific
role for the maternal activins has yet to be demonstrated.
Activin has the ability to induce in zebrafish animal
cap explants mesodermal markers, such as no tail [l]
(the zebrafish Bruchyury homolog), snail1 [51], and the
hepatocyte nuclear factor B (HNF3p) homolog axial [52]. Zygotic expression of both snail1 [51,53] and no tail is initially established within the whole germ-ring,
prospective mesendodermal cells. Animal cap cells transplanted to the germ-ring start to express no tail [l]. Thus activins, and/or other related mesoderm-inducing
factors, are active in the germ-ring. The role of FGFs
has not yet been investigated, but FGF receptors are
expressed in the early embryo [54].
The ‘fish organizer’
Classic transplantation experiments demonstrate that the zebrafish shield has organizer activities similar to
the dorsal blastopore lip of amphibia. In the teleosts F&tr/us [55] and zebrafish [56], shield mesoderm, when ectopically transplanted to the ventral side of a host embryo, induces secondary axis formation. The
molecular nature of the signals from the organizer appears to be conserved, as zebrafish shield can induce
axis duplications in amphibia [57].
The earliest gene known to be expressed specifically in the organizer region is the zebrafish goosecoid
homolog [34,58,59]. Transcripts are detected as early
as late-blastula stages in one-half of the blastula, in a graded distribution with a high point at the future dorsal side. Cells expressing goosecoid are among the
first to enter the hypoblast, and expression continues in the anterior hypoblast until onset of somitogenesis.
The Roosecoid-positive cells at this stage are located in the prechordal plate (axial head mesoderm), which continues to express the hlxl homeodomain protein during
further development [60]. Cells expressing goosecoid in
the late-blastula most likely define the region of the
early organizer and the head organizer, whereas late
goosemid- and h/xl-expressing cells appear to constitute
axial head mesoderm, the prechordal plate, to which
activities in dorsoventral patterning of the head have
been attributed [61-63]. It appears that at late-gastrula
stages, goosecoid expression spreads also to the inner cell
layer of the epiblast. This might reflect some of the interactions between hypoblast and epiblast, known as
vertical induction [64*]. The fate of these inner epiblast
cells is unknown, but is suggested to be neuroectoderm
[64’].
Midline signaling and dorsoventral patterning
Patterning along the dorsoventral midline of the embryo proper includes dorsoventral specification within the CNS (Rohon Beard cells, motoneurons, and floorplate,

Axis formation in zebrafish Driever 615
among others), formation and differentiation of the notochord, and an endodermal layer of cells between notochord and yolk. The prominent role of axial mesoderm in patterning of the trunk of zebrafish has become obvious from the analysis of the no tail mutation [2], the zebrafish homolog of mouse Bruchyury/T [3]. Embryos mutant for no tail do not develop differentiated notochord, although chordamesodermal precursor cells are present in the trunk. It has been suggested that signaling from these precursors is responsible for the fact that no tail mutants have a normal floorplate in the trunk [2]. However, requirements for patterning of paraxial mesoderm appear to also depend on signaling from differentiated notochord, as rzo tail mutants have abnormal somite patterning. The tail phenotype is much more severe, similar to Brachyury in mice. The tail is truncated, axial mesoderm is absent, and floorplate does not form in the remnants of the tail spinal cord. Expression of the ventro-posterior marker eve1 in the tailbud is absent [35].
The phenotype as well as the expression pattern define three aspects of no tail activity [l]. First, it is widely expressed in the tailbud and is involved in morphogenesis of the tail. Second, its expression in chordamesoderm during gastrulation appears to be required for differentiation into notochord. Third, during gastrulation, no tail is also expressed in the entire germ-ring. Cells from paraxial, lateral, and ventral positions of the germ-ring develop largely normally in no tail mutant embryos. It has been reported, however, that expression of snail occurs at reduced levels during gastrulation of no tail mutant embryos [51,53]. Thus, no tail might contribute toward establishment of ventral and lateral mesoderm, even though it is not the limiting regulator.
Zebrafish cyclops mutants are characterized by a deletion of some ventral parts of the brain and the floorplate [65,66*]. The deletion of ventral midline cells is cell autonomous, whereas other missing ventral midbrain and forebrain structures result from non-autonomous interactions. In mutant embryos, the expression of goosecoid in the epiblast is absent and, during late gastrulation, is reduced in the hypoblast. Thus, cyclops is also involved in specification of mesoderm in the zebrafish head [64*].
Wild-type floorplate cells, when transplanted into the cyclops ventral CNS midline, can rescue neighboring mutant cells to form midline structures [65,66-l. Thus, floorplate cells can homogenetically induce cyclops neighbors to form floorplate. The pathway for homogenetic induction might be distinct from the initial vertical floorplate-inducing signals originating in axial mesoderm [65,67].
A simple notion that cyclops might be defective in receiving the floorplate-inducing signals is challenged by the fact that goosecoid expression is reduced in the hypoblast. Furthermore, analysis of a differentiation marker, a(I) collagen type II, detects abnormalities in
cyclops mutant notochord [68], indicating that defects in axial mesoderm might be responsible in part for the phenotype.
The zebrafish HNF3p homolog axial [52] is first expressed just prior to gastrulation on the dorsal side of the embryo and continues to be expressed in dorsal axial mesendoderm (first the organizer region, later prechordal plate, notochord, and endoderm) along the whole axis until the end of gastrulation. Subsequently, axial expression is turned on in ventral neural plate cells (future forebrain and floorplate) adjacent to axial- expressing mesodermal cells. In cyclops mutant embryos, axial is not expressed in the neuroectoderm. Thus, cyclops is a component of the genetic pathway leading to axial activation. The cyclops gene is also involved in activation of sonic hedgehog expression in ventral CNS [67]. The sonic hedgehog gene is initially expressed in the shield hypoblast, next in axial mesoderm, and then in the ventral midline of the CNS. Ectopic expression of sonic hedgehog leads to ectopic activation of axial expression in the midbrain, where it is usually restricted to the floorplate, indicating that sonic hedgehog is one of the signals inducing ventral structures in the CNS. In cyclops mutants, sonic hedgehog is expressed in axial mesoderm, but not in the CNS.
The most prominent phenotype of spadetail mutants is the absence of trunk muscles [69]. Detailed fate mapping and the analysis of snail 1 and snail2 expression in spadetail embryos have demonstrated that the mutation is deficient in convergence and, possibly, specification of paraxial and lateral mesoderm in the trunk as welI as the head [53,70].
Further prospects for genetic analysis of pattern
formation
The unique features of zebrafish as a genetic system [71-73,74*] make it possible to identify a wide variety of mutations affecting pattern formation. So far, only a small number of mutations have been published (and discussed in this review). However, large-scale screens for embryonic phenotypes have been completed during the past few years [75*,76’], and results from the screens, including several hundred novel mutant loci, are expected to be published at the end of this year. Although these mutations will not suffice for solving the problem of pattern formation, they will provide a vast set of novel entry points from which to study axis formation, patterning, and organogenesis. In addition to the existing chemically induced mutations, novel techniques are emerging to induce insertional mutations [77*].
Current rapid improvements in genome resources, such as the genetic linkage map of zebrafish [78*] (for which the number of markers has already doubled within only one year after publication), and large insert libraries will make possible a molecular analysis of novel mutations in the near future. This will be important in order not only

616 Differentiation and gene regulation
to understand interactions between zebrafish mutations by gene expression analysis, but also, on a molecular level, to understand how patterning pathways identified in other vertebrate systems, such as Xenopus and mice, relate to zebrafish development.
Conclusions and perspectives
Despite significant advances in our understanding of zebrafish development, many important questions re- main. How is the dorsoventral axis initially specified? Is there a Nieuwkoop center in the embryo, and where is it? What is the source of the mesoderm-inducing signal? Where is Vg-1 processed? Whether the yolk syncytial cell plays a role as a source of localized signals and whether the yolk syncytial nuclei are transcriptionally active and contribute to patterning are interesting hypotheses to challenge.
Can zebrafish be a model for axis formation in mammals? Early blastomeres, similar to the inner cell mass in mice, appear developmentally non-restricted and equivalent. What role do the ‘extraembryonic’ tissues play in zebrafish is an issue as important as the one of activities of extraembryonic tissues at the onset of mammalian gastrulation.
The recently identified mutations should provide tools to answer some of these questions. The two mutations cydop.c and no tail have provided important information on the mechanism of midline signaling, and recent genetic screens have identified >20 additional mutations involved in dorsoventral patterning and midline signaling (C Niisslein-Volhard et al., personal communication; W Driever et al., unpublished data). Detailed genetic pathways should be constructed from these mutants over the next few years.
Zebrafish will be useful in the unraveling of develop- mental mechanisms by comparative studies. This applies not only to phenotypic and experimental analyses, but also to the study of gene regulation. With ~400 million years’ divergence, only sequences of functionally significant regulatory elements are conserved between mammals and zebrafish (or the puffer fish, another novel fish model system for genome analysis) [79-811. The combination of genetic, embryological, molecular, and comparative studies in zebrafish should enable us to dissect the components of maternal and zygotic genetic contributions to axis formation and to elucidate developmental pathways in the early embryo.
Acknowledgements
Thanks for discussion and comments on earlier versions of the
review to I>errk StempIe, Alexander Schier, Eliza Shah, and
Lilianna Solnlca-Krczel. Thanks to Uill Trevarrow for reference
to thr work of Wourrns on annual fishes. The author is sup-
ported by National Institutes of Health grant HI129761 and by
Umtol-Myers Squibb.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as: . . .
1.
2.
3.
4. .
of special inter&t _ of outstanding interest
Schulte-Merker S, Ho RK, Herrmann BC, NOsslein-Volhard C: The protein product of the zebrafish homologue of the mouse T gene is expressed in nuclei of the germ ring and the notochord of the early embryo. Development 1992, 116:1021-1032.
Halpern ME, Hp RK, Walker C, Kimmel CB: Induction of muscle pioneers and floor plate is distinguished by the zebrafish no tail mutation. Cell 1993, 75:99-l 11,
Schulte-Merker S, VanEeden FJM, Halpern ME, Kimmel CB, Niisslein-Volhard C: No tail (Ntl) is the zebrafish homologue of the mouse T (Brachyury) gene. Development 1994, 120:1009-1015.
Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF: Stages of embryonic development of the zebrafish. Dev Dynam 1995, 202:253-310.
Presents the most detailed description to date ofzebrafish embryogenesis, including a complete staging series as reference for future work.
5. Wolenski JS, Hart NH: Scanning electron microscope studies of sperm incorporation into the zebrafish (Brachydanio) egg. / Exp zoo/ 1987, 243:259-273.
6. Kane DA, Warga RM, Kimmel CB: Mitotic domains in the early embryo of the zebrafish. Nature 1992, 360:735-737.
7. Kane DA, Kimmel CB: The zebrafish midblastula transition. Development 1993, 119:447-456.
8. Strahle U, Jesuthasan S: Ultraviolet irradiation impairs epiboly in zebrafish embryos: evidence for a microtubule-dependent mechanism of epiboly. Development 1993, 119:909-919.
9. Solnica-Krezel L, Driever W: Microtubule arrays of the . zebrafish yolk cell: organization and function during epiboly.
Development 1994, 120:2443-2455. This paper refines the analysis of forces that drive epiboly and suggests that epibolic movements of the yolk syncytial nuclei and the blastoderm are not coupled. Epiboly of the yolk syncytial nuclei is absolutely, and of the blastoderm only partially, dependent on microtubules. Epibolic movements of the enveloping layer also involve endocytosis of membrane vesicles.
10. Solnica-Krezel L, Stemple DL, Driever W: Transparent things: cell fates and cell movements during early embryogenesis of zebrafish. Bioessays 1995, in press.
11. Schmitz B, Campos-Ortega JA: Dorso-ventral polarity of the zebrafish embryo is distinguishable prior to the onset of gastrulation. Roux Arch Dev Bio/ 1994, 203:374-380.
12. Warga RM, Kimmel CB: Cell movements during epiboly and gastrulation in zebrafish. Development 1990, 108:569%580.
13. Shih J, Fraser 5: The distribution of tissue progenitors within . the shield region of the zebrafish gastrula. Development 1995,
in press. A paper that presents provocative new ideas on the organization of the fate map at the dorsal margin, the embryonic shield. A salt-and-pepper distribution of neuroectoderm, axial mesoderm, and endoderm is found for the center region of the shield. Fufiher, a radial organization of fates within the different cell layers of the hypoblast is suggested.
14. Ballard WW: A new fate map for Salmo gairdncri. ) Exp Zoo/ 1973, 184:49-74.
15. Collaro A, Balker JA, Keller R: A phylogenetic perspective on . teleost gastrulation. Am Nat 1994, 144:133-l 52. An excellent comparison of cell behavior, cell movements, and morphology of gastrulating teleosts and other vertebrates.
16. Takeda H, Matsuzaki T, Oki T, Miyagawa T, Amanuma H: A novel POU domain gene, zebrafish pou2: expression and roles of two alternatively spliced twin products in early development. Genes Dev 1994, 8:45-59.

Axis formation in zebrafish Driever 617
17. Hauptmann G, Gerster T: Pou-2-a zebrafish gene active during cleavage stages and in the early hindbrain. Mech Dev 1995, 51:127-l 38.
18. Kane DA, Warga RM: Domains of movement in the zebrafish gastrula. Semin Dev Viol 1994, 5:101-l 09.
19. Kimmel CB, Warga RM, Schilling TF: Origin and organization of the zebrafish fate map. Development 1990, 108:581-594.
20. Lee RK, Stainier DY, Weinstein BM, Fishman MC: Cardiovascu- lar development in the zebrafish. II. Endocardial progenitors are sequestered within the heart field. Development 1994, 120:3361-3366.
21. Strehlow D, Gilbert W: A fate map for the first cleavages of the zebrafish. Nature 1993, 361:451-453.
22. Abdelilah S, Solnica-Krezel L, Stainier DY, Driever W: . . Implications for dorsoventral axis determination from the
zebrafish mutation janus. Nature 1994, 370:468-471. The first publication of a maternal-effect mutation in zebrafish. The janus mutation results in embryos with two adjacent blastoderms. The authors discuss the implications of the fact that in all cases, only one position along the combined margins will give rise to dorsal tissue.
23. . .
Helde KA, Wilson ET, Cretekos CJ, Crunwald DJ: Contribution of early cells to the fate map of the zebrafish gastrula. Science 1994, 265:517-520.
The analysis of embryos labeled with lineage-tracers at the eight-cell stage for expression of goosecoid RNA at shield stage settles a controversy, in finding that no correlation exists between the second cleavage plane and the future dorsoventral axis.
24. Wacker S, Herrmann K, Berking S: The orientation of the dorsal/ventral axis of zebrafish is influenced by gravitation. Roux Arch Dev Viol 1994, 203:281-283.
25. Kimmel CB, Warga RM: Tissue-specific cell lineages originate in the gastrula of the zebrafish. Science 1986, 231:365-368.
26. Wilson ET, Helde KA, Grunwald DJ: Something’s fishy here-rethinking cell movements and cell fate in the zebrafish embryo [review]. Trends Cenef 1993, 9:348-352.
27. Strehlow D, Heinrich G, Gilbert W: The fates of the blastomeres of the 16-cell zebrafish embryo. Development 1994, 120:1791-1798.
28. Wilson ET, Cretekos CJ, Helde KA: Cell mixing during early . epiboly in the zebrafish embryo. Dev Cenet 1995, in press. Analysis of the extent of cell mixing in different regions of the blastula provides an explanation for the observation that marginal blastomeres give rise to a more restricted range of fates than animal or central blastomeres, which are subject to more extensive mixing during epiboly. These findings explain controversies in previous fate-mapping attempts.
29. Stainier DY, Lee RK, Fishman MC: Cardiovascular development in the zebrafish. I. Myocardial fate map and heart tube formation. Developmenf 1993, 119:3 l-40.
30. Woo K, Fraser S: Order and coherence in the fate map of the zebrafish nervous system. Development 1995, in press.
iresents a very detailed fate map of the regions of the CNS at the shield stage as well as during late gastrulation. An excellent reference for the interpretation of gene expression patterns, and a first good framework to evaluate the movements of neuroectodermal precursors during gastrulation.
31.
32.
33.
34.
35.
Ho RK, Kimmel CB: Commitment of cell fate in the early zebrafish embryo. Science 1993, 261: 109-l 11.
Gerhart J, Danilchik M, Doniach T, Roberts S, Rowning B, Stewart R: Cortical rotation of the Xenopus egg: consequences for the anteroposterior pattern of embryonic dorsal development. Development 1989, Suppl:37-51.
50. Wittbrodt J, Rosa FM: Disruption of mesoderm and axis . formation in fish by ectopic expression of activin variants:
the role of maternal activin. Genes Dev 1994, 8:1448-l 462. Two dominant-negative activin variants are analyzed for their effects on fish development upon RNA injection into one-cell embryos. One variant inhibits native translated cellular activin precursor proteins by forming processing-defective heterodimers. The other competes for binding of mature processed cellular activin to receptor. The cotranslatlonal form does not inhibit normal axis formation, indicating that zygotic activin transcription is not necessary for axis formatton.
Kessler DS, Melton DA: Vertebrate embryonic induction: meso- dermal and neural patterning. Science 1994, 266:596-604.
51. Hammerschmidt M, Ntisslein-Volhard C: The expression of a zebrafish gene homologous to Drosophila snail suggests a conserved function in invertebrate and vertebrate gastrulation. Development 1993, 119:1107-1118.
Stachel SE, Grunwald DJ, Myers PZ: lithium perturbation and goosecoid expression identify a dorsal specification pathway in the pregastrula zebrafish. Development 1993, 117:1261-l 274.
52. Strahle U, Blader P, Henrique D, lngham PW: Axial, a zebrafish gene expressed along the developing body axis, shows altered expression in cyclops mutant embryos. Genes Dev 1993, 7:1436-l 446.
Joly JS, Joly C, Schulte-Merker S, Boulekbache H, Condamine 53. Thisse C, Thisse B, Schilling TF, Postlethwait JH: Structure of H: The ventral and posterior expression of the zebrafish the zebrafish snail1 gene and its expression in wild-type,
homeobox gene eve1 is perturbed in dorsalized and mutant embryos. Development 1993, 119:1261-1275.
36. Xu Q, Holder N, Patient R, Wilson SW: Spatially regulated . expression of three receptor tyrosine kinase genes during
gastrulation in the zebrafish. Development 1994, 120:287-299. The expression of three receptor tyrosine kinases sheds an interesting light on regionalization in neuroectoderm and axial mesoderm. Ex- pression of receptor tyrosine kinases in dorsalized embryos indicates that the anteroposterior order of gene expression is maintained in dorsalized-radialized embryos.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48. .
Long WL: The role of the yolk syncytial layer in determination of the plane of bilateral symmetry in rainbow trout, Sahno gairdneri Richardson. / Exp Zoo/ 1983, 228:91-97.
Oppenheimer JM: The development of isolated blastoderms of Fundulus heteroclitus. J Exp Zoo/ 1936, 72:247-269.
DeVillers C: In vitro explantations of fish blastoderms (Salmo, Esox.) [in French]. Experientia 1947, 3:71-74.
Kostomarova AA: The differentiation capacity of isnlated loach (Misgurnus fossilis 1.) blastoderm. / Embryo/ Exp Morph 1969, 22:407-430.
Lin S, Long W, Chen J, Hopkins N: Production of germ-line chimeras in zebrafish by cell transplants from genetically pigmented to albino embryos. Proc Nat/ Acad Sci USA 1992, 89:4519-4523.
Carter CA, Wourms JP: Cell behaviour during early development in the South American annual fishes of the genus Cynolebias. / Morphol 1991, 210:247-266.
Wourms JP: The developmental biology of annual fishes. II. Naturally occurring dispersion and reaggregation of blastomeres during the development of annual fish eggs. / Exp Zoo/ 1972, 182:169-200.
Stern CD: Mesoderm induction and development of the embryonic axis in amniotes. Trends Cenet 1992, 8:158-163.
Slack JM: Inducing factors in Xenopus early embryos. Curr Biol 1994, 4:116-l 26.
Helde KA, Grunwald DJ: The DVR-1 (Vgl) transcript of zebrafish is maternally supplied and distributed throughout the embryo. Dev Biol 1993, 159:418-426.
Thomsen CH, Melton DA: Processed Vgl protein is an axial mesoderm inducer in Xenopus. Cell 1993, 74:433-441.
Toyama R, O’Connell ML, Wright CVE, Kuehn MR, Dawid IB: Nodal induces ectopic goosecoid and liml expression and axis duplication in zebrafish. Development 1995, 121:383-391,
An application of ‘Xenopus genetics’ (i.e. injection of RNAs encoding growth factors into early embryos) in zebrafish, and the first demonstra- tion of the inductive activities of TCFB family molecules in zebrafish.
49. Zhou X, Sasaki H, Lowe L, Hogan BLM, Kuehn MR: Nodal is a novel TCF-p-like gene expressed in the mouse node during gastrulation. Nature 1993, 361:543-547.

618 Differentiation and gene regulation
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64. .
Thisse C, Thisse B, Halpern ME, Postlethwait JH: Coosecoid expression in neuroectoderm and mesendoderm is disrupted in zebrafish cyclops gastrulas. Dev Biol 1994, 164:420-429.
A detailed study of the expression of goosecoid in wild-type and cyclops mutant embryos reveals the spreading of goosecoid expression to the hypoblast and demonstrates that cyclops acts in the shield and head mesoderm. This is an important paper for understanding the role of head axial mesoderm in patterning the ventral forebrain.
65. Hatta K, Kimmel CB, Ho RK, Walker C: The cyclops mutation blocks specification of the floor plate of the zebrafish central nervous system. Nature 1991, 350:339-341.
66. Hatta K, Puschel AW, Kimmel CB: Midline signaling in the . primordium of the zebrafish anterior central nervous system.
Proc Nat/ Acad Sci USA 1994, 91:2061-2065. Transplantation experiments between wild-type and cyclops mutant embryos suggest a cell autonomous requirement for cyclops activity in the ventral midline cells of the forebrain, whereas patterning of the wider ventral part of the forebrain missing in cyclops mutants appears to depend on non-autonomous signals deriving from the ventral midline cells. These experiments define the signaling pathways in the ventral zebrafish forebrain.
67. Krauss S, Concordet JP, lngham PW: A functionally conserved homolog of the Drosophila segment polarity gene hh is expressed in tissues with polarizing activity in zebrafish embryos. Cell 1993, 75:1431-1444.
68. Yan Y-L, Hatta K, Riggleman B, Postlethwait JH: Expression of a type II collagen gene in the zebrafish embryonic axis. Dev Dynam 1995, 203:363-376.
69. Ho RK, Kane DA: Cell-autonomous action of zebrafish spt- 1 mutation in specific mesodermal precursors. Nature 1990, 348:728-730.
70. Thisse C, Thisse B, Postlethwait JH: Expression of snai12, a second member of the zebrafish snail family, in cephalic
spadetail and no tail mutant embryos. Development 1993, 119:1203-1215.
Thisse B, Thisse T, Weston J: A novel FGF receptor (Z-FCFR4) is dynamically expressed in mesoderm and neuroectoderm during early embryogenesis. Dev Dynam 1995, 203:377-391.
Oppenheimer J: Transplantation experiments on developing teleosts (Fundulus and Perca). J Exp Zoo/ 1936, 72:409-437.
Ho R: Axis formation in the embryo of the zebrafish Brachydanio rerio. Sem Dev t?io/ 1992, 3:53-64.
Oppenheimer JM: Structures developed in amphibians by implantation of living fish organizers. Proc Sot Exp Biol Med 1936, 34:461-463.
Schulte-Merker S, Hammerschmidt M, Beuchle D, Cho KW, DeRobertis EM, Nijsslein-Volhard C: Expression of zebrafish goosecoid and No tail gene products in wild-type and mutant No tail embryos. Development 1994, 120:843-852.
DeRobertis EM, Fainsod A, Cant LK, Steinbeisser H: The evolution of vertebrate gastrulation. Developmenf 1994, 116(suppl):ll7-124.
Fjose A, Izpisua-Belmonte JC, Fromental-Ramain C, Duboule D: Expression of the zebrafish gene hlx-1 in the prechordal plate and during CNS development. Development 1994, 120:71-81.
Adelmann HB: The development of the prechordal plate and mesoderm of Amblystoma punctatum. / Morphol 1932, 54:1-67.
Adelmann HB: The problem of cyclopia. Part II. Quart Rev Bio/ 1936, 11:284-304.
Mangold 0: The problem of determination. Ill. The vertebrate eye in development and regeneration [in German]. Ergeb d Biol 1931, 7:193-403.
mesoderm and presumptive neural crest of wild type and spadetail mutant embryos. Dev Biol 1995, in press.
71. Streisinger C, Walker C, Dower N, Knauber D, Singer F: Production of clones of homozygous diploid zebra fish (Erachydanio rerio). Nature 1981, 291:293-296.
72. Mullins MC, Nijsslein-Volhard C: Mutational approaches to studying embryonic pattern formation in the zebrafish. Curr Opin Genet Dev 1993, 3:648*54.
73. Driever W, Stemple D, Schier A, Solnica-Krezel L: Zebrafish: genetic tools for studying vertebrate development. Trends Genet 1994, 10: 152-l 59.
74. Riley BR, Grunwald DL: Efficient induction of point mutations . allowing recovery of specific locus mutations in zebrafish. Proc
Nafl Acad Sci USA 1995, 92:5997-6001. This paper presents genetic experimental strategies that make allele screens and screens for novel mutations feasible with a compararively small investment in animal facilities.
75. Mullins MC, Hammerschmidt M, Haffter P, Ntisslein-Volhard . C: large-scale mutagenesis in the zebrafish: in search of
genes controlling development in a vertebrate. Curr 6iol 1994, 4:189-202.
This paper outlines the strategies for saturation mutagenesis in zebrafish. The paper deals with logistic aspects of a screen, mutagenesis efficiency, and range of phenotypes obtained in pilot screens.
76. Solnica-Krezel L, Schier AF, Driever W: Efficient recovery of . EN&induced mutations from the zebrafish germline. Genetics
1994, 136:1401-1420. The most detailed analysis of mutagenesis conditions and mutagen-dose efficiency in zebrafish. The essential ‘how to’ paper if a genetic screen in zebrafish is to be planned.
77. Lin S, Caiano N, Culp P, Burns JC, Friedmann T, Yee . JK, Hopkins N: Integration and germ-line transmission of
a pseudotyped retroviral vector in zebrafish. Science 1994, 265:666-669.
Demonstration of high-efficiency germline transmission of Moloney murine leukemia wrus based retroviruses, which have been pseudotyped for the vesicular stomatitis virus surface glycoprotein G, in zebrafish, potentially representing a strategy of choice for future Insertional mutagenesis screens.
78. Postlethwait JH, Johnson SL, Midson CN, Talbot WS, Gates . M, Ballinger EW, Africa D, Andrew R, Carl T, Elsen JS ef
al.: A genetic linkage map for the zebrafish. Science 1994, 264:699-703.
Presents a zebrafish map based on randomly amplified polymorphic DNA (RAPD) markers. The paper demonstrates how the special features of zebrafish genetics, such as making haploid embryos, can be exploited for rapid mapping of mutations and cloned genes in zebrafish.
79. Westeriield M, Wegner J, Jegalian BC, DeRobertis EM, Piischel AW: Specific activation of mammalian Hox promoters in mosaic transgenic zebrafish. Genes Dev 1992, 6:591-598.
80. Aparicio S, Morrison A, Gould A, Gilthorpe J, Chaudhuri C, Rigby P, Krumlauf R, Brenner S: Detecting conserved regulatory elements with the model genome of the Japanese puffer fish, Fugu rubripes. Proc Nat/ Acad Sci USA 1995, 92:1684-l 688.
81. Brenner S, Elgar C, Sandford R, Macrae A, Venkatesh B, Aparicio S: Characterization of the pufferfish Wugu) genome as a compact model vertebrate genome. Nature 1993, 366~265-267.
W Ikievrr, Cardiovascular Research Center, Massachusetts
General Hospital Thirteenth Street, Building 149 (Mail Code
1494X1), Charlestown, Massachusetts 02129-2060, USA.





























