Embed Size (px)
Citation preview

falciparum. Genome Biol. 6, R26
1471-4922/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.
doi:10.1016/j.pt.2009.07.003 Available online 4 September 2009
Update Trends in Parasitology Vol.25 No.10
AcknowledgementsThis work was supported by the European Commission (BioMalPar:Network of Excellence) and the DFG (SFB 544). ML is a member of theCellnetworks excellence cluster.
References1 Snow, R.W. et al. (2005) The global distribution of clinical episodes of
Plasmodium falciparum malaria. Nature 434, 214–2172 Mita, T. et al. (2003) Recovery of chloroquine sensitivity and low
prevalence of the Plasmodium falciparum chloroquine resistancetransporter gene mutation K76T following the discontinuance ofchloroquine use in Malawi. Am. J. Trop. Med. Hyg. 68, 413–415
3 Mita, T. et al. (2004) Expansion of wild type allele rather than backmutation in pfcrt explains the recent recovery of chloroquine sensitivityof Plasmodium falciparum in Malawi. Mol. Biochem. Parasitol. 135,159–163
4 Wang, X. et al. (2005) Decreased prevalence of the Plasmodiumfalciparum chloroquine resistance transporter 76T markerassociated with cessation of chloroquine use against P. falciparummalaria in Hainan, People’s Republic of China. Am. J. Trop. Med.Hyg 72, 410–414
5 Kublin, J.G. et al. (2003) Reemergence of chloroquine-sensitivePlasmodium falciparum malaria after cessation of chloroquine usein Malawi. J. Infect. Dis. 187, 1870–1875
6 Hayward, R. et al. (2005) pfmdr1mutations associated with chloroquineresistance incurafitness cost inPlasmodiumfalciparum.Mol.Microbiol.55, 1285–1295
7 Shinondo, C.J. et al. (1994) Effect of pyrimethamine resistance onsporogony in a Plasmodium berghei / Anopheles stephensi model.Exp. Parasitol. 78, 194–202
8 Rosario, V.E. et al. (1978) Persistence of drug-resistant malariaparasites. Lancet 1, 185–187
9 Schneider, P. et al. (2008) Does the drug sensitivity of malaria parasitesdepend on their virulence? Malar J. 7, 257
10 Mackinnon, M.J. et al. (2002) Virulence in rodent malaria: hostgenotype by parasite genotype interactions. Infect. Genet. Evol. 1,287–296
Research Focus
Nottingham, NG7 2UH, UK
Corresponding author: Pleass, R.J. ([email protected]).
11 Ekland, E.H. and Fidock, D.A. (2007) Advances in understanding thegenetic basis of antimalarial drug resistance.Curr. Opin.Microbiol. 10,363–370
12 Mita, T. et al. (2009) Indigenous evolution of Plasmodium falciparumpyrimethamine resistance multiple times in Africa. J. Antimicrob.Chemother. 63, 252–255
13 Abali, E.E. et al. (2008) Regulation of human dihydrofolate reductaseactivity and expression. Vitam. Horm. 79, 267–292
14 Thaithong, S. et al. (2001) Plasmodium falciparum: gene mutationsand amplification of dihydrofolate reductase genes in parasites grownin vitro in presence of pyrimethamine. Exp. Parasitol. 98, 59–70
15 Dharia, N.V. et al. (2009) Use of high-density tiling microarrays toidentify mutations globally and elucidate mechanisms of drugresistance in Plasmodium falciparum. Genome Biol. 10, R21
16 Sanchez, C.P. et al. (2008) Dissecting the components of quinineaccumulation inPlasmodium falciparum.Mol.Microbiol. 67, 1081–1093
17 Sanchez, C.P. et al. (2007) Is PfCRT a channel or a carrier? Twocompeting models explaining chloroquine resistance in Plasmodiumfalciparum. Trends Parasitol. 23, 332–339
18 Valderramos, S.G. and Fidock, D.A. (2006) Transporters involved inresistance to antimalarial drugs. Trends Pharmacol. Sci. 27, 594–601
19 Borges-Walmsley, M.I. et al. (2003) Structure and function of effluxpumps that confer resistance to drugs. Biochem. J. 376, 313–338
20 Litman, T. et al. (2001) From MDR to MXR: new understandingof multidrug resistance systems, their properties and clinicalsignificance. Cell. Mol. Life. Sci 58, 931–959
21 Jansen, G. et al. (1998) A structurally altered human reduced folatecarrier with increased folic acid transport mediates a novel mechanismof antifolate resistance. J. Biol. Chem. 273, 30189–30198
22 Assaraf, Y.G. and Goldman, I.D. (1997) Loss of folic acid exporterfunction with markedly augmented folate accumulation in lipophilicantifolate-resistant mammalian cells. J. Biol. Chem. 272, 17460–17466
23 Martin, R.E. et al. (2005) The ‘permeome’ of the malaria parasite: anoverview of the membrane transport proteins of Plasmodium
B-cells get the T-cells but antibodies get the worms
Richard J. Pleass and Jerzy M. Behnke
Institute of Genetics, and Parasite Biology and Immunogenetics Research Group, Queen’s Medical Centre, University of
Two recent papers published in Immunity and Cell Host& Microbe underline the great importance of B cells andof antibodies (Abs) in orchestrating crucial T helper celltype 2 (Th2) protective immune responses to gastroin-testinal nematodes. The findings in animal models nowraise major questions as to how B cells and Abs carry outthese functions in humans. Here we discuss recent tech-nological advances in humanizing animal models at thelevel of both Abs and their Fc-receptors, that mightprovide some answers.
From polyclonal to monoclonalWe have read with great interest the important work byWojciechowski et al. [1] and the highly complimentaryarticle by McCoy et al. [2], in which the authors demon-strate that B cells and antibodies (Abs) play an essential
role in protection to the gastrointestinal nematode, Helig-mosomoides polygyrus, now renamed Heligmosomoidesbakeri [3]. The two papers highlight that B cells are crucialto generation of T helper cell type 2 (Th2) responses thatlead to control of parasites. Although this expansion andmaturation of protective Th2 cells by B cells was indepen-dent of Ab synthesis, the ability to limit parasites wasclearly attributable to the presence of specific Ab. Theimportant contribution made by Abs was demonstratedby passive transfer of immune serum (or the IgG fraction)into naı̈ve recipient C57BL/6 mice [2]. Surprisingly, pass-ive transfer of immune serum intom�MTmicewaswithoutprotective effect [1], presumably as a consequence of agenetic defect in the mice (or some other uncharacterizeddeficiency) on which the functionality of Abs relies, andthat indirectly prevents Abs from clearing parasites?
In contrast to the work of Wojciechowski and McCoyusing passive transfer of immune serum containing poly-clonal Abs (principally IgG1), many groups have shown
443
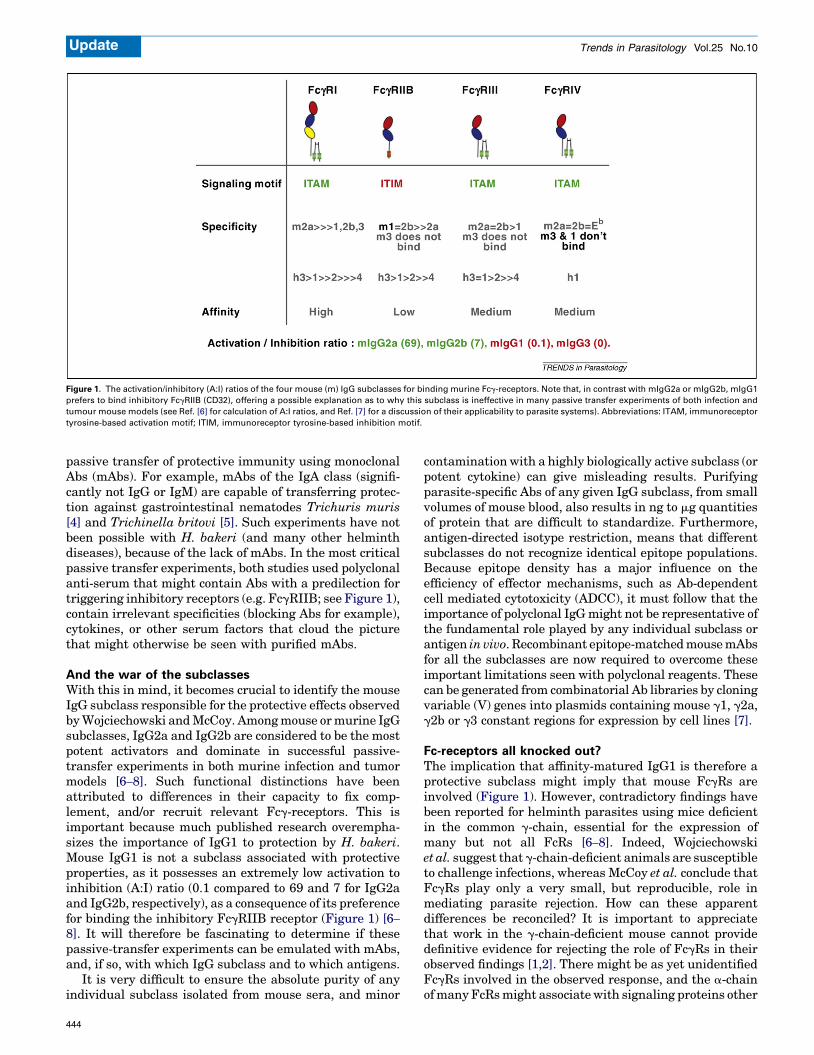
Figure 1. The activation/inhibitory (A:I) ratios of the four mouse (m) IgG subclasses for binding murine Fcg-receptors. Note that, in contrast with mIgG2a or mIgG2b, mIgG1
prefers to bind inhibitory FcgRIIB (CD32), offering a possible explanation as to why this subclass is ineffective in many passive transfer experiments of both infection and
tumour mouse models (see Ref. [6] for calculation of A:I ratios, and Ref. [7] for a discussion of their applicability to parasite systems). Abbreviations: ITAM, immunoreceptor
tyrosine-based activation motif; ITIM, immunoreceptor tyrosine-based inhibition motif.
Update Trends in Parasitology Vol.25 No.10
passive transfer of protective immunity using monoclonalAbs (mAbs). For example, mAbs of the IgA class (signifi-cantly not IgG or IgM) are capable of transferring protec-tion against gastrointestinal nematodes Trichuris muris[4] and Trichinella britovi [5]. Such experiments have notbeen possible with H. bakeri (and many other helminthdiseases), because of the lack of mAbs. In the most criticalpassive transfer experiments, both studies used polyclonalanti-serum that might contain Abs with a predilection fortriggering inhibitory receptors (e.g. FcgRIIB; see Figure 1),contain irrelevant specificities (blocking Abs for example),cytokines, or other serum factors that cloud the picturethat might otherwise be seen with purified mAbs.
And the war of the subclassesWith this in mind, it becomes crucial to identify the mouseIgG subclass responsible for the protective effects observedbyWojciechowski andMcCoy. Amongmouse ormurine IgGsubclasses, IgG2a and IgG2b are considered to be the mostpotent activators and dominate in successful passive-transfer experiments in both murine infection and tumormodels [6–8]. Such functional distinctions have beenattributed to differences in their capacity to fix comp-lement, and/or recruit relevant Fcg-receptors. This isimportant because much published research overempha-sizes the importance of IgG1 to protection by H. bakeri.Mouse IgG1 is not a subclass associated with protectiveproperties, as it possesses an extremely low activation toinhibition (A:I) ratio (0.1 compared to 69 and 7 for IgG2aand IgG2b, respectively), as a consequence of its preferencefor binding the inhibitory FcgRIIB receptor (Figure 1) [6–
8]. It will therefore be fascinating to determine if thesepassive-transfer experiments can be emulated with mAbs,and, if so, with which IgG subclass and to which antigens.
It is very difficult to ensure the absolute purity of anyindividual subclass isolated from mouse sera, and minor
444
contamination with a highly biologically active subclass (orpotent cytokine) can give misleading results. Purifyingparasite-specific Abs of any given IgG subclass, from smallvolumes of mouse blood, also results in ng to mg quantitiesof protein that are difficult to standardize. Furthermore,antigen-directed isotype restriction, means that differentsubclasses do not recognize identical epitope populations.Because epitope density has a major influence on theefficiency of effector mechanisms, such as Ab-dependentcell mediated cytotoxicity (ADCC), it must follow that theimportance of polyclonal IgGmight not be representative ofthe fundamental role played by any individual subclass orantigen in vivo. Recombinant epitope-matchedmousemAbsfor all the subclasses are now required to overcome theseimportant limitations seen with polyclonal reagents. Thesecan be generated from combinatorial Ab libraries by cloningvariable (V) genes into plasmids containing mouse g1, g2a,g2b or g3 constant regions for expression by cell lines [7].
Fc-receptors all knocked out?The implication that affinity-matured IgG1 is therefore aprotective subclass might imply that mouse FcgRs areinvolved (Figure 1). However, contradictory findings havebeen reported for helminth parasites using mice deficientin the common g-chain, essential for the expression ofmany but not all FcRs [6–8]. Indeed, Wojciechowskiet al. suggest that g-chain-deficient animals are susceptibleto challenge infections, whereas McCoy et al. conclude thatFcgRs play only a very small, but reproducible, role inmediating parasite rejection. How can these apparentdifferences be reconciled? It is important to appreciatethat work in the g-chain-deficient mouse cannot providedefinitive evidence for rejecting the role of FcgRs in theirobserved findings [1,2]. There might be as yet unidentifiedFcgRs involved in the observed response, and the a-chainofmany FcRsmight associatewith signaling proteins other

Update Trends in Parasitology Vol.25 No.10
than the common g-chain. For example, mouse IgG3-opso-nized Cryptococcus neoformans can still be phagocytosedby macrophages from these same FcRg-/- mice [9]. Thiseffect is probably mediated via an undefined FcR that doesnot require g-chain for function because, of the knownFcRs, only murine FcgRI binds mouse IgG3 [10] (seeFigure 1). More problematic is that FcRg-/- mice have beenfound to express partially functional FcgRI in more recentmouse knockouts [11,12]. It is now known that the a-chainof FcgRI can mediate major histocompatibility complex(MHC) class II Ag presentation without active g-chainsignaling [13,14], and that the a-chain can interact withPeriplakin to control receptor endocytosis and IgG bindingcapacity [13,14]. These issues might therefore be moreclearly resolved in mice deficient in a-chains for theseFcRs, for example, in quintuplet Fc-deficient mice usedrecently to highlight the importance of FcgRIV to thefunction of mouse IgE [15]. If the assertion made by theauthors as to the importance of IgG1 is correct, might oneexpect greater levels of protection in mice deficient inFcgRIIB, that are no longer able to bind IgG1 and aretherefore incapable of mounting inhibitory responses(Figure 1)?
Neutralizing Fabs or regulatory FcsAnother possibility is that the protection offered by pas-sively transferred IgG is mediated not only by FcgRs, butalso by other mechanisms. For example, in malaria, Abscan inhibit processing of key parasite antigens and thesubsequent invasion of red blood cells by mechanisms thatare independent of Fc-receptors [16]. The passive transferof Fabs or F(ab’)2, derived from their specific IgG (free ofFc), would have provided evidence for the role of the Fc overthe Fab in their findings. However, that passively trans-ferred Abs do not protect in m-MT animals would argueagainst Fc-independent mechanisms of Ab action.
One unifying hypothesis that fits the data obtained isthat Abs simply neutralize the immunomodulatory factorssecreted by the highly immune suppressive adult worms,thereby allowing Th2-driven responses to gain dominance,together with ensuing inflammatory responses, leading toprotection [17,18]. Indeed, earlier work from our group [19]demonstrated that IgG1 (almost undetectable IgG2a) wasthe predominant subclass found at mucosal surfaces, andmuch of it was directed against excretory/secretory (ES)molecules from adult worms. That IgG1 is the major sub-class found at mucosal surfaces in mice makes sense inlight of the low A:I ratio ascribed to this subclass [6,7]. Ithas also recently been shown that the Fc portion of IgGcontains T-cell epitopes that are capable of specificallyactivating CD4+CD25HiFoxP3
+ natural T regulatory cells(Tregs) [20]. In vivo administration of Fc ‘tregitopes’resulted in suppression of immune responses to knownimmunogens, and suggests a potential immunosuppres-sive activity of polyclonal IgG1, as seen during H. bakeriprimary infections with adult worms [17,18,21]. Theimmunosuppressive effects of adult worms and the poly-clonal IgG1 they induce might therefore be related to theactivity of Tregs. Such an abundance of polyclonal IgG1might tip the resulting immune response towards toler-ance of worms rather than their elimination [20]. The
identification of the molecules seen by these IgG1 s andhow they interfere with adult worms must be priorities forfuture research. Understanding the role (if any), of theother subclasses, e.g. by chimerization of V genes fromthese IgG1s, might yield novel findings concerning therole of the Fc in this process. However, a mechanism ofsurvival based purely on the induction of Tregs is unlikelyto explain why adult worms can be transplanted directlyinto the intestines of immune mice where they quicklyestablish and survive for many weeks [22].
Human, mouse or human–mouse chimeras?Immunity in rodents might also bear little resemblance toimmunity in humans. Now that the importance of Abs incontrolling nematode infections in rodents has been firmlyestablished, making the system more relevant to humansbecomes critical. This could be addressed by engineeringhuman mAbs for testing in mouse models transgenic forhuman Fc-receptors, as has recently been done withmalaria to highlight the importance of human FcgRI[7,23]. That this approach might hold promise for investi-gating Ab-mediated immunity to nematodes has beendemonstrated by the observation that human IgG canpassively protect against larval Strongyloides stercoralisin recipient mice [24]. Similar approaches have been usedto examine human immune responses to whipworms byreconstitution of SCIDmice with human lymphocytes [25].The relevance of mouse models to infection in humans isparticularly important to address, because knockout miceare increasingly being used in models of parasite infection.It is therefore vital to recognize the differences betweenFcR systems in mice and in humans. For example, thereare no mouse equivalents of human FcgRIIA, FcgRIIC,FcgRIIIB and FcaRI [7,8]. Looking at FcgRIIA in particu-lar, this receptor has its own unique immunoreceptortyrosine-based activation (ITAM) motif and is thereforecapable of signal transduction and phagocytosis in theabsence of the common g-chain or other associated sub-units. Such careful considerations will prove important inthe design of Ab-based therapies and vaccination protocolsfor helminth diseases of humans and animals.
It is clear from these two important publications that Abis an important component of immune responses leading tothe control of parasites. The priority now is to identify theantigens involved and to elucidate the Fc-receptor/IgGsubclasses that contribute optimally to this protection.Only detailed and precise answers to these questions willlead to accurate in vitro assays that better reflect ourexpectations for future vaccines.
AcknowledgementsWe thank the Wellcome Trust, the Medical Research Council UK and theSir Halley Stewart trust for funding work in our laboratories.The authors declare no conflicts of interest
References1 Wojciechowski, W. et al. (2009) Cytokine-producing effector B cells
regulate type 2 immunity to H. polygyrus. Immunity 30, 421–4332 McCoy, K.D. et al. (2008) Polyclonal and specific antibodies mediate
protective immunity against enteric helminth infection. Cell HostMicrobe 16, 362–373
3 Behnke, J. and Harris, P. (2009) Heligmosomoides bakeri orHeligmosomoides polygyrus? Am. J. Trop. Med. Hyg. 80, 684–685
445

1
1
1
1
1
1
1471-4922/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.
doi:10.1016/j.pt.2009.07.001 Available online 4 September 2009
Update Trends in Parasitology Vol.25 No.10
4
4 Roach, T.I. et al. (1991) Trichuris muris: antigen recognition andtransfer of immunity in mice by IgA monoclonal antibodies. Parasit.Immunol. 13, 1–12
5 Inaba, T. et al. (2003) Monoclonal IgA antibody-mediated expulsion ofTrichinella from the intestine of mice. Parasitology 126, 591–598
6 Nimmerjahn, F. and Ravetch, J.V. (2005) Divergent immunoglobulin g
subclass activity through selective Fc receptor binding. Science 310,1510–1512
7 Pleass, R.J. (2009) Fc-receptors and immunity to malaria: frommodelsto vaccines. Parasit. Immunol. 31, 529–538
8 Pleass, R.J. and Woof, J.M. (2001) Fc receptors and immunity toparasites. Trends Parasitol. 17, 545–551
9 Yuan, R. et al. (1998) Ab-mediated modulation of Cryptococcusneoformans infection is dependent on distinct Fc receptor functionsand IgG subclasses. J. Exp. Med. 187, 641–648
0 Gavin, A.L. et al. (1998) Identification of the mouse IgG3 receptor:implications for antibody effector function at the interface betweeninnate and adaptive immunity. J. Immunol. 160, 20–23
1 Ioan-Facsinay, A. et al. (2002) FcgammaRI (CD64) contributessubstantially to severity of arthritis, hypersensitivity responses, andprotection from bacterial infection. Immunity 16, 391–402
2 Barnes, N. et al. (2002) FcgammaRI-deficient mice show multiplealterations to inflammatory and immune responses. Immunity 16,379–389
3 van Vugt, M.J. et al. (1999) The FcgammaRIa (CD64) ligand bindingchain triggers major histocompatibility complex class II antigenpresentation independently of its associated FcR gamma-chain.Blood 94, 808–817
4 Beekman, J.M. et al. (2004) Direct interaction between FcgammaRI(CD64) and periplakin controls receptor endocytosis and ligandbinding capacity. Proc. Natl. Acad. Sci. USA 101, 10392–10397
5 Mancardi, D.A. et al. (2008) FcgammaRIV is a mouse IgE receptor thatresembles macrophage FcepsilonRI in humans and promotes IgE-induced lung inflammation. J. Clin. Invest. 118, 3738–3750
46
16 Collins, C.R. et al. (2009) An inhibitory antibody blocks interactionsbetween components of the malarial invasion machinery. PLoS Pathog5 (1), e1000273. Epub 2009 Jan 23
17 Pleass, R.J. and Bianco, A.E. (1994) The role of adult worms insuppressing functional protective immunity to Heligmosomoidespolygyrus bakeri challenge infections. Parasit. Immunol. 16,619–628
18 Behnke, J.M. (1987) Evasion of immunity by nematode parasitescausing chronic infections. Adv. Parsitol. 26, 1–71
19 Pleass, R.J. and Bianco, A.E. (1996) Irradiated larval vaccination andantibody responses evaluated in relation to the expression of immunityto Heligmosomoides polygyrus. Parasitol. Res. 82, 445–453
20 De Groot, A.S. et al. (2008) Activation of natural regulatory T cells byIgG Fc-derived peptide ‘Tregitopes’. Blood 112, 3303–3311
21 Chapman, C.B. et al. (1979) IgG1 hypergammaglobulinaemia inchronic parasitic infections in mice: evidence that the responsereflects chronicity of antigen exposure. Aus. J. Exp. Biol. Med. Sci57, 389–400
22 Robinson, M. et al. (1988) Immunity to adult Heligmosomoidespolygyrus (Nematospiroides dubius): survival or rejection of adultworms following transplantation to mice refractory to larvalchallenge. J. Helminthol. 62, 221–231
23 McIntosh, R.S. et al. (2007 May 18) The importance of humanFcgammaRI inmediating protection tomalaria. PLoS Pathog 3 (5), e72
24 Kerepesi, L.A. et al. (2004) Human immunoglobulin G mediatesprotective immunity and identifies protective antigens againstlarval Strongyloides stercoralis in mice. J. Infect. Dis. 189, 1282–
129025 Taylor, M.D. and Else, K.T. (2002) Human Trichuris-specific antibody
responses in vaccinated hu-PBL-SCID mice. Parasit. Immunol. 24,1–13





























