Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY,0021-9193/99/$04.0010
Dec. 1999, p. 7154–7160 Vol. 181, No. 23
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Bacillus subtilis yckG and yckF Encode Two Key Enzymes of theRibulose Monophosphate Pathway Used by Methylotrophs,
and yckH Is Required for Their ExpressionHISASHI YASUEDA,* YOSHIO KAWAHARA, AND SHIN-ICHI SUGIMOTO
Fermentation and Biotechnology Laboratories, Ajinomoto Co., Inc.,Kawasaki-ku, Kawasaki-shi, 210-0801, Japan
Received 14 June 1999/Accepted 14 September 1999
The ribulose monophosphate (RuMP) pathway is one of the metabolic pathways for the synthesis of com-pounds containing carbon-carbon bonds from one-carbon units and is found in many methane- and methanol-utilizing bacteria, which are known as methylotrophs. The characteristic enzymes of this pathway are 3-hexu-lose-6-phosphate synthase (HPS) and 6-phospho-3-hexuloisomerase (PHI), neither of which was thought toexist outside methylotrophs. However, the presumed yckG gene product (YckG) of Bacillus subtilis shows aprimary structure similar to that of methylotroph HPS (F. Kunst et al., Nature 390:249–256, 1997). We havealso investigated the sequence similarity between the yckF gene product (YckF) and methylotroph PHI(Y. Sakai, R. Mitsui, Y. Katayama, H. Yanase, and N. Kato, FEMS Microbiol. Lett. 176:125–130, 1999) andfound that the yckG and yckF genes of B. subtilis express enzymatic activities of HPS and PHI, respectively. Bothof these activities were concomitantly induced in B. subtilis by formaldehyde, with induction showing depen-dence on the yckH gene, but were not induced by methanol, formate, or methylamine. Disruption of either genecaused moderate sensitivity to formaldehyde, suggesting that these enzymes may act as a detoxification systemfor formaldehyde in B. subtilis. In conclusion, we found an active yckG (for HPS)-yckF (for PHI) gene structure(now named hxlA-hxlB) in a nonmethylotroph, B. subtilis, which inherently preserves the RuMP pathway.
The ribulose monophosphate (RuMP) pathway is involvedin formaldehyde fixation in many methylotrophs (11, 15, 17,22). Typically, the RuMP pathway has two characteristic en-zymes, 3-hexulose-6-phosphate synthase (HPS) and 6-phos-pho-3-hexuloisomerase (PHI), and it also shares some en-zymes with the pentose phosphate pathway and the glycolyticor Entner-Doudoroff pathway. PHI catalyzes isomerization be-tween fructose 6-phosphate and hexulose 6-phosphate, whichis synthesized from ribulose 5-phosphate (Ru5P) and formal-dehyde by HPS. Although HPS and PHI have been detected inmany methylotrophs (3, 12, 18–20, 23, 32, 34), the genes en-coding these enzymes remain largely unknown. The hps gene,encoding HPS, was first cloned from an obligate methylotroph,Methylomonas aminofaciens 77a (44), and cloning of the gene(rmpB) encoding PHI from M. aminofaciens 77a (35) and afacultative methylotroph, Mycobacterium gastri MB19 (28), wasrecently reported. Neither enzyme was thought to exist outsidemethylotrophs, but sequence analysis of the Bacillus subtilisgenome (24, 41) has suggested that yckG may be a homolog ofthe gene encoding HPS (44) in M. aminofaciens 77a. The yckGgene is located between yckF and yckH, at about 32° on theB. subtilis genome map (24) (Fig. 1a). Here, we present datashowing that a nonmethylotroph, B. subtilis, contains genes(yckG and yckF) encoding two key enzymes of the RuMPpathway (HPS and PHI) and that the activities of both en-zymes can be induced by formaldehyde with dependence onthe yckH gene. These findings indicate the presence of char-acteristic enzymes from the RuMP pathway in a nonmethyl-otroph. The significance of detection of these enzymes in B.
subtilis is discussed from the aspect of molecular evolution ofthe RuMP pathway.
MATERIALS AND METHODS
Bacterial strains, oligonucleotide primers, and plasmids. The wild-type strainB. subtilis 168 (1) was purchased from the American Type Culture Collection(Rockville, Md.), and Escherichia coli JM109 (45) was obtained from TakaraShuzo Co. (Kyoto, Japan). B. subtilis YD101 (yckG-deficient mutant) and YD102(yckF-deficient mutant) were constructed, as described below, by inserting thespectinomycin resistance (spc) DNA fragment from pDG1726 (13) (BacillusGenetic Stock Center, Columbus, Ohio) into the individual genes of the chro-mosome of strain 168. Strains YD111 (yckH::spc) and YD112 (yckH::spc) werealso constructed from strain 168 in the same way (Fig. 1a). Strain YD121 (yckHyckH1) contained both the intact yckH gene and the defective one (Fig. 1b).
The potential open reading frame (ORF) of the yckG or yckF gene was clonedby PCR amplification (29) using the oligonucleotide primer pair BsYck-G1(59-GAGTATCGATAAAATGGAATTACAGCTTGCATTAGACCTCGT-39;the translation start codon is underlined, and the restriction enzyme site [ClaI]for ligation to the vector is italicized) plus BsYck-G6 (59-AATTGTGGATCCCATTGAGAATTTCCGCTACGTATTCAGTCG-39; the restriction enzyme site[BamHI] for ligation to the vector is italicized) or BsYck-F1 (59-AAGCATCGATAAAATGAAAACGACTGAATACGTAGCGGAA-39) plus BsYck-F2 (59-ATCTTGGATCCGGTTGTGTGATGTTATTCAAGGTTTGCG-39). The re-gion containing the yckG and yckF genes was amplified by PCR using the primerpair BsYck-G1 plus BsYck-F2. PCR was performed with Takara LA Taq DNApolymerase (Takara Shuzo) for 28 cycles with purified B. subtilis genomic DNA.The PCR products were purified with a QIAquick gel extraction kit (Qiagen).The fragments thus obtained were ligated behind the E. coli trp promoter tomediate overexpression in E. coli (46). Each amplified DNA fragment wasdigested with restriction enzymes ClaI and BamHI, and the fragments wereintroduced into the vector to construct the expression vectors pT-Bsb-yckG6,pT-Bsb-yckF1, and pT-Bsb-yckGF1 for the overexpression of yckG, yckF, andyckGF, respectively. The fragment containing yckH was amplified by PCR usingprimers BsYck-H1 (59-CAATGTTAACGTCAGGCTTTTGCTGGATCACTTCTGGCA-39; the restriction enzyme site for cloning of the amplified DNA frag-ment is italicized) and BsYck-H2 (59-GTGTACCGAATTCGTTTTTGTGCATCCGTTAAAGGGTA-39). After digestion with HpaI and EcoRI, the yckHfragment was cloned into pACYC184 (Nippon Gene, Tokyo, Japan) treated withBst1107I and EcoRI to construct pAY-Bsb-yckH.
For the purpose of gene disruption, the spc fragment was introduced into thecloned yckG and yckF genes to construct pT-Bsb-yckG-Sp5 and pT-Bsb-yckF-Sp6, respectively. The spc gene was also cloned into the PvuII site in the yckHgene to construct two plasmids, pAY-Bsb-yckH-SpA and pAY-Bsb-yckH-SpB, in
* Corresponding author. Mailing address: Fermentation and Biotech-nology Laboratories, Ajinomoto Co., Inc., 1-1, Suzuki-cho, Kawasaki-ku,Kawasaki-shi, 210-0801, Japan. Phone: 81-44-245-5972. Fax: 81-44-222-0129. E-mail: [email protected] or Hisashi [email protected]..
7154
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

which the direction of the spc gene was the same as and opposite, respectively,that of the yckG gene (Fig. 1a).
Construction of B. subtilis mutants. B. subtilis mutants YD101, YD102,YD111, and YD112 were constructed by the introduction of pT-Bsb-yckG-Sp5,pT-Bsb-yckF-Sp6, pAY-Bsb-yckH-SpA, and pAY-Bsb-yckH-SpB, respectively,into the wild-type strain. YD121 was generated from the wild-type strain throughhomologous recombination with the yckH region by a Campbell-type event.Transformation of B. subtilis was performed by the competent-cell method (1),and spectinomycin (100 mg/ml) was used for selection of the transformants.
Preparation of cell extract from E. coli overexpressing the yckG and yckFgenes. E. coli JM109 was transformed with the expression plasmids (36). Trans-formants were grown at 37°C for 12 h in 5 ml of M9-Casamino Acids medium(36) supplemented with thiamine-HCl (2 mg/ml), indoleacrylic acid (25 ml/ml),and ampicillin (100 ml/ml). The cells were harvested by centrifugation and sus-pended in buffer A (50 mM potassium phosphate [pH 7.6], 1 mM MgCl2). Aftercentrifugation, the washed cell pellet was resuspended in 1 ml of buffer B (50 mMpotassium phosphate [pH 7.6], 5 mM MgCl2, 1 mM dithiothreitol). A cell extractwas prepared by sonication with a disrupter (Bioruptor; Cosmobio, Tokyo, Ja-pan) at maximum power with a standard oscillator. The sonicate was clarified bycentrifugation at 15,000 3 g for 20 min at 2°C, and the supernatant thus obtainedwas used as the cell extract.
SDS-PAGE analysis. Sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE) (14% polyacrylamide) was performed by the methodof Laemmli (25), using a ready-made gel system (TEFCO, Tokyo, Japan). Sam-ples of cultured cells were collected by centrifugation, and the cell pellets weresuspended in gel loading buffer and heated at 100°C for 5 min. Aliquots of 5 mlwere loaded onto an SDS-polyacrylamide gel, which was stained with PhastGelBlue R (Coomassie blue R-350; Pharmacia AB). Molecular weight standardsconsisted of a marker mix consisting of seven recombinant proteins of 15, 25, 35,50, 75, 100, and 150 kD (Novagen, Madison, Wis.).
Induction of expression of the yckG and yckF genes in B. subtilis. Afterovernight growth in Luria-Bertani (LB) medium (36), cultures of wild-type andmutant B. subtilis were diluted to 1:20 in fresh medium. Cells were cultured at37°C with shaking until the mid-log phase, and then formaldehyde or anotherreagent (methanol, formate, or methylamine) was added as specified in the text.Other cultures were exposed to the following stresses: heat shock, provided byculture at 48°C; salt stress, created by adding NaCl at a final concentration of 5%(wt/vol); and acid shock, provided by adding 0.5 N HCl to reduce the pH of theculture from 6.8 to 5.2. Incubation was continued for 90 min under these con-ditions. Next, the cells were harvested, washed, and resuspended in 50 mMpotassium phosphate buffer (pH 7.5) containing 3 mM MgCl2 and 1 mM dithio-threitol. The cell extract was prepared for assay by sonication and centrifugationto remove debris.
Assay of HPS and PHI activities. HPS activity was determined as describedpreviously (3), with some modifications. The reaction mixture (0.95 ml) consisted
of 50 mM potassium phosphate (pH 7.6), 5 mM MgCl2, 5 mM ribose-5-phos-phate (Ri5P) (Nacalai Tesque, Inc., Kyoto, Japan), 2.5 mM NADP (Sigma, St.Louis, Mo.), 10 U of phosphoribose isomerase (PRI) (Sigma), 10 U of glucose-6-phosphate dehydrogenase (Boehringer Mannheim), 10 U of phosphoglucoseisomerase (PGI) (Boehringer Mannheim), 50 U of recombinant PHI purifiedfrom E. coli overproducing M. aminofaciens 77a PHI (35), and the test sample ofcell extract. The mixture was preincubated for 3 min at 37°C to achieve equilib-rium of Ru5P and Ri5P (catalyzed by PRI), and then the reaction was started byadding formaldehyde at a final concentration of 5 mM. In the PHI assay, re-combinant M. aminofaciens 77a HPS (35) was used instead of recombinant PHI,and both were eliminated from the reaction mixture when the serial reaction ofHPS and PHI enzymes was assayed. Activity was assessed by monitoring thechange in optical density (OD) at 340 nm with NADP as a cofactor, and cor-rection was made for formaldehyde-independent activity. The protein concen-tration was determined by the method of Bradford (8) with a Bio-Rad proteinassay kit (Bio-Rad Laboratories, Richmond, Calif.) and bovine serum albumin asthe standard.
Assay of other enzymes. NAD-dependent methanol dehydrogenase (MDH)and glutathione-dependent formaldehyde dehydrogenase (GS-FDH) were as-sayed by using the B. subtilis cell extract treated with formaldehyde. MDH ac-tivity was assayed as described by Arfman et al. (2); GS-FDH activity was assayedas described by Attwood. (6).
Effect of yckG and yckF mutations on cell growth in the presence of formal-dehyde or NaCl. B. subtilis 168 (wild type) and two isogenic mutant strains(YD101 and YD102) were cultured at 30°C with shaking in LB medium con-taining spectinomycin when necessary. Overnight cultures were inoculated intowarmed fresh LB medium, or into medium containing formaldehyde or NaCl,and cell growth and lysis profiles were automatically recorded at an OD of 660nm with a Bio-photorecorder (Advantec, Tokyo, Japan).
13C NMR analysis of the products obtained with [13C]formaldehyde as thesubstrate. A reaction mixture containing 50 mM potassium phosphate (pH 7.6),5 mM MgCl2, 5 mM Ri5P, 10 U of PRI, and 10 U of PGI was prepared. Afterpreincubation at 37°C for 10 min to produce Ru5P from Ri5P, cell extractcontaining the yckG gene product (YckG) and yckF gene product (YckF) wasadded to the mixture. The serial YckG and YckF reaction was started by additionof [13C]formaldehyde as the substrate. After incubation at 37°C for 20 min,EDTA was added to the mixture to chelate Mg21, which is required for HPSactivity. The total activity in the mixture was assayed with 13C nuclear magneticresonance (NMR) spectroscopy by monitoring the decrease of the resonancesignal for the 13C atom of formaldehyde and the appearance of signals for the Catoms of fructose 6-phosphate and glucose 6-phosphate, which was yielded byaddition of PGI. In the negative control sample, EDTA was added to the mixtureimmediately after addition of the cell extract.
NMR spectra were obtained at 100 MHz with a Fourier transform (FT) NMRspectrometer (JNM-A400; JEOL, Tokyo, Japan), using 0.6-ml samples contain-ing 15% D2O in 5-mm-diameter tubes to provide a lock signal. A total of 256transients were acquired by using 32k data points. Gated proton decoupling wasused, and 45° pulses were applied, with a 2.5-s relaxation delay between pulses togive a repetition time of 3.9 s. During acquisition, the samples were kept at 25°C.[13C]formaldehyde (99% 13C atom) was purchased from Cambridge IsotopeLaboratories (Andover, Mass.).
RNA analysis. B. subtilis cells were grown to mid-log phase in LB medium at37°C. After the addition of formaldehyde at final concentration of 0.5 mM,culture was continued for 45 min. Then RNA was extracted by using an ISOGENkit (Nippongene, Toyama, Japan) and treated with RNase-free DNase (TakaraShuzo). About 1 mg of RNA was mixed with the YGF-R2 oligonucleotide DNA(59-TTCAGGCCGGTTGTGTGATGC-39), and cDNA was synthesized by usinga reverse transcription (RT) kit (RNA LA PCR kit; Takara Shuzo). YGF-R2corresponded to the minus strand of DNA just behind the yckF ORF. Then a20-ml aliquot of the cDNA mixture was combined with DNA primers YGF-F1(59-ATGGAATTACAGCTTGCATTAGACC-39) and YGF-R1 (59-TGTGATGCTATTCAAGGTTTGC-39), LA Taq DNA polymerase, and PCR buffer(Takara Shuzo) in a volume of 100 ml. RT-PCR was carried out according to themanufacturer’s instructions. YGF-F1 and YGF-R1 corresponded to the plusstrand of DNA encoding the N-terminal region of YckG and to the minus strandof DNA encoding the C-terminal region of YckF, respectively. As a negativecontrol, mixture without RT was used for PCR amplification.
Computer analysis. Nucleotide and amino acid sequences were analyzed withthe Genetyx-Mac computer program (Software Development Co., Tokyo, Ja-pan). Alignment of deduced amino acid sequences was performed by comparisonwith entries in the SWISS-PROT database (release 35.0).
RESULTS
Overproduction and enzyme activity of the yckG and yckFgene products. To identify the orthologs of the gene encodingPHI, FASTA searches (30, 31) of the microorganism proteinsequence database were performed, using the amino acid se-quence of the rmpB gene product (PHI) (35) of M. amino-faciens 77a as the query sequence. As a result, the primary
FIG. 1. Diagram of the chromosomes of wild-type B. subtilis and its mutants.(a) Map of the yckF-yckG (phi-hps) and yckH loci showing the protein codingregions at 32° on the B. subtilis genome (24). Locations and orientations of thecoding regions are indicated by thin black arrows, and potential transcriptionterminators, as proposed in the SubtiList data bank (41), are shown. The hps, phi,and yckH genes are expanded and depicted as thick black arrows below the map.Insertion of a spc cassette (thick white arrow) leading to disruption of hps instrain YD101, phi in YD102, or yckH in YD111 or YD112 is shown. (b) Chro-mosomal structure of strain YD121, which contains the disrupted and intactyckH genes. Heavy and thin lines depict vector and chromosomal DNA, respec-tively. Open boxes indicate the yckG, yckF, and yckH genes. The black box showsthe spc gene, which disrupts the yckH gene. Thin arrows show the direction oftranscription.
VOL. 181, 1999 ONE-CARBON COMPOUND METABOLISM IN B. SUBTILIS 7155
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

sequence of the yckF gene product (YckF) in B. subtilis showedthe highest structural similarity with that of PHI (Fig. 2). TheyckF gene is situated five nucleotides downstream of the yckGgene in the B. subtilis genome (Fig. 1a), and the yckG geneproduct (YckG) is known to show structural similarity withHPS (24). Thus, both putative genes involved in one-carboncompound metabolism were situated side-by-side in the B. sub-tilis genome. Because B. subtilis cannot utilize methanol as itssole carbon source (i.e., it is a nonmethylotroph), the detectionof such genes in this microorganism was interesting with re-spect to function and evolution.
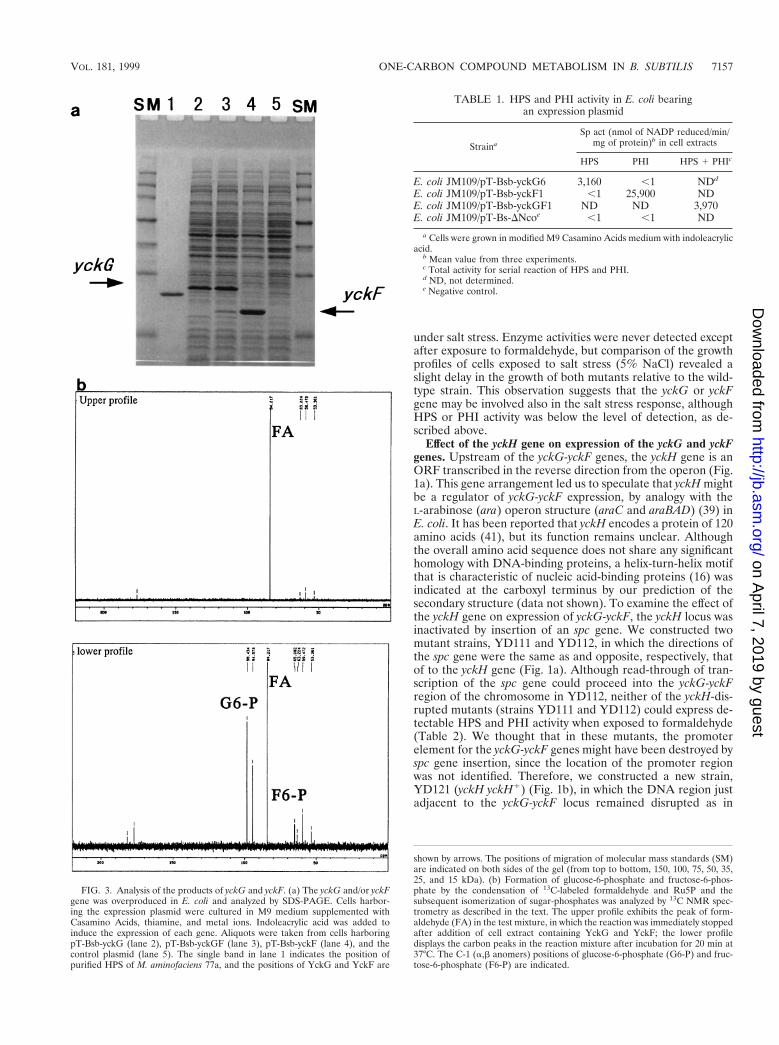
We first expressed the yckG- and/or yckF ORF in E. coli toevaluate whether the translated product of each ORF dis-played HPS or PHI activity. After each corresponding DNAfragment with the postulated ORF was amplified by PCR, thefragments were inserted into the expression plasmid and over-production was stimulated (Fig. 3a). As a result, the yckG geneyielded a dense protein band with the expected molecular massof 23 kDa on SDS-PAGE, and the yckF gene yielded a proteinwith a molecular mass of 19 kDa. Furthermore, overexpressionof a DNA fragment containing the yckG and yckF genes in theintact gene structure led to concomitant overproduction ofboth ORF products (Fig. 3a, lane 3), showing that terminationof transcription did not occur between the yckG and yckFgenes in E. coli.
The HPS and PHI activities of cell extracts were examinedby the enzyme assay method. E. coli overexpressing yckG oryckF showed HPS or PHI activity, respectively (Table 1). Toconfirm that formaldehyde was incorporated into pentosephosphate, we used the 13C NMR technique with [13C]form-aldehyde. Preincubation of the reaction mixture containingRi5P, PRI and other chemicals yielded Ru5P, which is anacceptor of formaldehyde in the reaction with HPS. Additionof a cell extract containing YckG and YckF to the mixturecaused the appearance of the corresponding resonance signalsfor [13C]fructose-6-phosphate and [13C]glucose-6-phosphate,along with a decrease of the [13C]formaldehyde signal (Fig.3b). This showed that the serial reaction catalyzed by both geneproducts could progress, because YckG produced hexulose-6-phosphate from formaldehyde and Ru5P, while YckF and PGIthen created fructose-6-phosphate and glucose-6-phosphate,respectively. Since the two enzymes were previously thoughtto be unique to methylotrophs (11, 15), it was surprising that
B. subtilis contained nucleotide sequences that could expressthese enzyme activities.
Expression of yckG and yckF genes by B. subtilis. To inves-tigate whether the yckG and yckF genes intrinsically expressedany enzyme activity in B. subtilis, we assessed extracts of cellsgrown under various culture conditions. The extract of wild-type cells cultured in LB medium showed negligible HPS orPHI activity, but both activities were detected after culture inthe presence of formaldehyde (Table 2). When various con-centrations of formaldehyde were tested, significant HPS andPHI activity was found in the cell extract at formaldehydeconcentrations of around 0.5 mM. At a concentration of over1 mM, however, both activities almost disappeared.
Formaldehyde is produced from methanol by methanol de-hydrogenase and is also oxidized to formate by formaldehydedehydrogenase in some methylotrophs (11). Therefore, we ex-amined whether the synthesis of HPS and PHI could be in-duced by methanol or formate, but neither enzyme activity wasdetected after cells were grown in medium containing metha-nol or formate (Table 2). On the other hand, since formalde-hyde is also synthesized by oxidation of methylamine in severalmethylotroph species (26), we looked for HPS or PHI activityin extracts of B. subtilis grown in medium containing methyl-amine hydrochloride (0.4 or 1.2% [wt/vol], final concentration)but did not observe the induction of these enzymes. Therefore,the nonmethylotroph B. subtilis possessed two key enzymes ofthe RuMP pathway, and the expression of both enzymes wasinduced by formaldehyde.
Analysis of yckG and/or yckF disruption mutants. We nextexamined whether the yckG or yckF gene encoded the ob-served formaldehyde-inducible HPS or PHI activity. We con-structed a yckG mutant (strain YD101) and a yckF mutant(strain YD102) by insertional mutagenesis (Fig. 1a) and ana-lyzed the formaldehyde-inducible HPS or PHI activity of eachstrain. After induction with formaldehyde, we detected no sig-nificant enzyme activity in the YD101 strain and only HPS ac-tivity in the YD102 strain (Table 2). Thus, we identified YckGand YckF as the only formaldehyde-inducible HPS and PHI en-zymes, respectively, and our results also suggested that a polareffect occurred between the yckG and yckF genes. To confirmthe transcription unit of the two genes, total RNA was isolatedfrom wild-type B. subtilis cells exposed to formaldehyde andanalyzed by RT-PCR. The presence of mRNA species (of atleast 1.2 kb) spanning the yckG ORF to the yckF ORF (datanot shown) was revealed, suggesting that the two genes (yckGand yckF) were expressed as a polycistronic mRNA from anoperon.
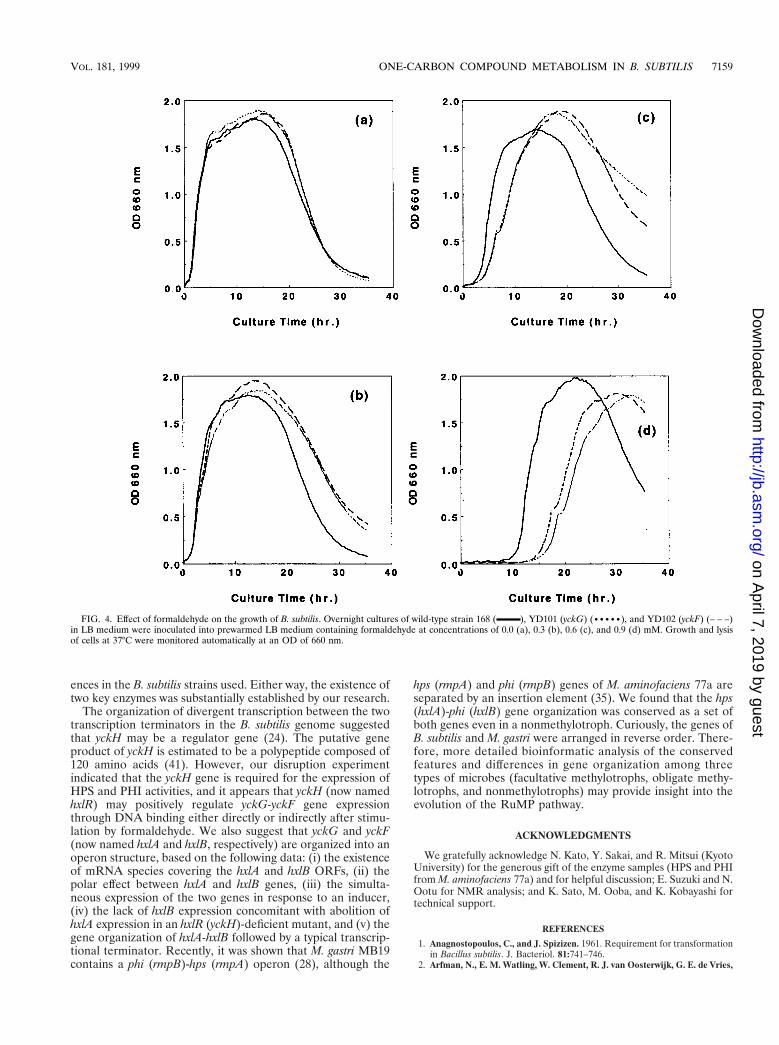
We also investigated the effect of formaldehyde on the phe-notype of these mutants. In LB medium, the wild-type strainand the two mutant strains all showed typical growth and lysiscurves (Fig. 4a). However, after addition of formaldehyde tothe medium, the mutants showed a prolonged lag phase rela-tive to the growth profile of the wild-type strain, which wasdependent on the concentration of formaldehyde added (Fig.4). These results indicated that the yckG-yckF system mightendow B. subtilis with the ability to detoxify formaldehyde pro-duced via endogenous metabolism or environmental changes.
It appeared that the cells were under stress when exposed toformaldehyde, since growth was arrested for a few hours in itspresence (Fig. 4c and d). To examine whether the increase ofHPS and PHI activities after exposure to formaldehyde wasdue to a general stress response rather than to formaldehydeitself, we assessed the levels of these enzymes in cells grownunder heat (increase from 30 to 48°C), acid (decrease from pH6.8 to 5.2), and salt (addition of 5% NaCl) stress. We alsoexamined the growth profiles of wild-type and mutant cells
FIG. 2. Alignment of amino acid sequences of YckF of B. subtilis (24) andPHI of M. aminofaciens 77a (35). Identical amino acids and conservative replace-ments are indicated by asterisks and dots, respectively. Amino acids are num-bered from the N terminus of each protein.
7156 YASUEDA ET AL. J. BACTERIOL.
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
under salt stress. Enzyme activities were never detected exceptafter exposure to formaldehyde, but comparison of the growthprofiles of cells exposed to salt stress (5% NaCl) revealed aslight delay in the growth of both mutants relative to the wild-type strain. This observation suggests that the yckG or yckFgene may be involved also in the salt stress response, althoughHPS or PHI activity was below the level of detection, as de-scribed above.
Effect of the yckH gene on expression of the yckG and yckFgenes. Upstream of the yckG-yckF genes, the yckH gene is anORF transcribed in the reverse direction from the operon (Fig.1a). This gene arrangement led us to speculate that yckH mightbe a regulator of yckG-yckF expression, by analogy with theL-arabinose (ara) operon structure (araC and araBAD) (39) inE. coli. It has been reported that yckH encodes a protein of 120amino acids (41), but its function remains unclear. Althoughthe overall amino acid sequence does not share any significanthomology with DNA-binding proteins, a helix-turn-helix motifthat is characteristic of nucleic acid-binding proteins (16) wasindicated at the carboxyl terminus by our prediction of thesecondary structure (data not shown). To examine the effect ofthe yckH gene on expression of yckG-yckF, the yckH locus wasinactivated by insertion of an spc gene. We constructed twomutant strains, YD111 and YD112, in which the directions ofthe spc gene were the same as and opposite, respectively, thatof to the yckH gene (Fig. 1a). Although read-through of tran-scription of the spc gene could proceed into the yckG-yckFregion of the chromosome in YD112, neither of the yckH-dis-rupted mutants (strains YD111 and YD112) could express de-tectable HPS and PHI activity when exposed to formaldehyde(Table 2). We thought that in these mutants, the promoterelement for the yckG-yckF genes might have been destroyed byspc gene insertion, since the location of the promoter regionwas not identified. Therefore, we constructed a new strain,YD121 (yckH yckH1) (Fig. 1b), in which the DNA region justadjacent to the yckG-yckF locus remained disrupted as in
FIG. 3. Analysis of the products of yckG and yckF. (a) The yckG and/or yckFgene was overproduced in E. coli and analyzed by SDS-PAGE. Cells harbor-ing the expression plasmid were cultured in M9 medium supplemented withCasamino Acids, thiamine, and metal ions. Indoleacrylic acid was added toinduce the expression of each gene. Aliquots were taken from cells harboringpT-Bsb-yckG (lane 2), pT-Bsb-yckGF (lane 3), pT-Bsb-yckF (lane 4), and thecontrol plasmid (lane 5). The single band in lane 1 indicates the position ofpurified HPS of M. aminofaciens 77a, and the positions of YckG and YckF are
TABLE 1. HPS and PHI activity in E. coli bearingan expression plasmid
Straina
Sp act (nmol of NADP reduced/min/mg of protein)b in cell extracts
HPS PHI HPS 1 PHIc
E. coli JM109/pT-Bsb-yckG6 3,160 ,1 NDd
E. coli JM109/pT-Bsb-yckF1 ,1 25,900 NDE. coli JM109/pT-Bsb-yckGF1 ND ND 3,970E. coli JM109/pT-Bs-DNcoe ,1 ,1 ND
a Cells were grown in modified M9 Casamino Acids medium with indoleacrylicacid.
b Mean value from three experiments.c Total activity for serial reaction of HPS and PHI.d ND, not determined.e Negative control.
shown by arrows. The positions of migration of molecular mass standards (SM)are indicated on both sides of the gel (from top to bottom, 150, 100, 75, 50, 35,25, and 15 kDa). (b) Formation of glucose-6-phosphate and fructose-6-phos-phate by the condensation of 13C-labeled formaldehyde and Ru5P and thesubsequent isomerization of sugar-phosphates was analyzed by 13C NMR spec-trometry as described in the text. The upper profile exhibits the peak of form-aldehyde (FA) in the test mixture, in which the reaction was immediately stoppedafter addition of cell extract containing YckG and YckF; the lower profiledisplays the carbon peaks in the reaction mixture after incubation for 20 min at37°C. The C-1 (a,b anomers) positions of glucose-6-phosphate (G6-P) and fruc-tose-6-phosphate (F6-P) are indicated.
VOL. 181, 1999 ONE-CARBON COMPOUND METABOLISM IN B. SUBTILIS 7157
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

YD111 but the intact yckH gene was located away from theyckG-yckF region past the intervening sequence of the inte-grated plasmid. We examined YD121 for formaldehyde-in-duced HPS or PHI activity and again detected the induction ofboth activities. This indicated that the yckH gene is requiredfor the expression of HPS and PHI activities.
DISCUSSION
Recently, bioinformatics analysis has suggested that hps genehomologs may be widespread in bacteria (33). We also founda structural similarity between PHI from a methylotroph (35)and YckF from B. subtilis and noticed that the hps homolog(yckG) and phi homolog (yckF) were adjacent in the B. subtilisgenome. In E. coli, it was reported that the SgaH (ORF o216)and SgbH (ORF o220) proteins exhibit sequence similarity tothe HPS of M. aminofaciens (33). Therefore, we assessed theenzymatic activity of the sgaH and sgbH gene products fromstrain W3110 by overexpression of the corresponding genes butcould detect no HPS activity. We also examined HPS activity inextracts of E. coli cells cultured under several stresses, includ-ing formaldehyde exposure, but the activity was below the levelof detection (data not shown). Although HPS activity in E. colimight be induced by a specific effector, it appears from ourresults that E. coli does not possess the enzyme. Our homologysearch indicated that the E. coli genome does not contain ahomolog of the rmpB (phi) gene (data not shown). Thesefindings suggested that the E. coli hps homologs may possess afunction other than hexulose phosphate synthesis or that dur-ing its evolution the bacterium has accumulated mutationswhich impede the expression of HPS activity. According to theevolutionary history of prokaryotes based on phylogeneticanalysis (43), B. subtilis is more proximal than E. coli to thebranch point of the three kingdoms, Archaea, Bacteria, andEucarya (7). Therefore, it appears that the presence of a set ofhps and phi genes in the genome is critical, and the existence ofboth genes in the B. subtilis genome suggests that the geneproducts were of significant value for this microorganism in itsoriginal environment.
Expression of the yckG and yckF genes was induced inB. subtilis by formaldehyde, similar to what has been found fortwo gram-positive methylotrophs, Arthrobacter P1 (27) and Ba-cillus methanolicus (4). Formaldehyde is produced from meth-
anol by MDH in the first stage of the utilization of carbon andenergy sources by methylotrophs. In B. methanolicus, the syn-thesis of HPS is also enhanced by culture with methanol (2),but methanol could not induce HPS or PHI activity in B. sub-tilis. Having found that HPS and PHI enzymes exist in B. sub-tilis, we examined the activity of MDH in this microbe. Thestructure of the cytoplasmic NAD-dependent MDH of B. meth-anolicus C1 was described by De Vries et al. (9). Using theirsequence information, we performed a homology search forthe mdh gene in the B. subtilis genome database and found nosignificant homologs. In addition, we could not detect NAD-dependent MDH activity in cell extracts of B. subtilis culturedunder various conditions (data not shown). Thus, it appearsthat B. subtilis does not possess an NAD-dependent MDHhomolog derived from B. methanolicus. However, it is inter-esting that the possible yeaC gene product of B. subtilis (24, 41)displays similarity to the product of mxaR (moxR) required foractivity of the pyrrolo-quinoline quinone (PQQ)-linked MDHin a gram-negative methylotroph (42). Besides the methanoloxidation pathway, formaldehyde is also synthesized frommethylamine in several species of methylotrophs which areable to utilize methylamine as the sole source of carbon andenergy (21, 26, 27). The yckG-yckF system might be involved inmethylamine metabolism, since some heterotrophs are able touse methylamine as a nitrogen source, and methylamine, whichis more stable than formaldehyde, is widely distributed in ma-rine environment (21). However, no detectable HPS or PHIactivity was found in the extract of B. subtilis cells cultured withmethylamine, and methylamine could not support the growthof B. subtilis as a nitrogen source (data not shown).
This study also indicated a possible role of YckG and YckFin the detoxification of formaldehyde. As noted by Attwoodand Quayle (5), it seems possible that the HPS and PHI systemis very efficient for trapping free formaldehyde. However, B.subtilis may also equip other detoxification systems for form-aldehyde, since our yckG or yckF mutants did not show markedformaldehyde sensitivity. In E. coli, GS-FDH is the primaryenzyme that detoxifies formaldehyde (14). Therefore, it is pos-sible that B. subtilis also possesses a reliable detoxificationsystem employing the corresponding enzyme. Indeed, our ho-mology search showed that the adhB gene product in B. subtilisis similar to GS-FDH in Methylobacter marinus (40), althoughwe detected no of GS-FDH activity in cell extracts from B.subtilis exposed to formaldehyde, as was the case for the meth-anotroph. In addition, we found that both mutants (yckG andyckF) were slightly more sensitive to salt (NaCl) stress than thewild-type strain, although the HPS or PHI activity in cellsstressed by salt was too low to be measured. This implies thatthe promoter responsible for the salt stress response mightexist in front of both genes or that salt stress could induce theaccumulation of formaldehyde during metabolism. The rela-tionship between the salt stress response and the function ofyckG and yckF remains to be determined.
The results presented here demonstrate that a nonmethyl-otroph, B. subtilis, can synthesize two key enzymes (HPS andPHI) that were previously believed to be specific to methyl-otrophs employing the RuMP pathway. In B. subtilis, the pen-tose phosphate pathway has been identified by metabolic fluxanalysis (37, 38). Therefore, we suggest that B. subtilis pre-serves the RuMP pathway. Although De Wulf assumed thepresence of this pathway in B. subtilis on the basis of a prelim-inary physiological experiment (10), his conclusion that formicacid in the culture medium was effectively utilized by B. subtilisvia the RuMP pathway is not compatible with our results,because formic acid could not induce HPS and PHI activities inthe present study. This discrepancy might be related to differ-
TABLE 2. HPS and PHI activities in B. subtilis 168 and deletionmutants under different growth conditions
Strain Growthconditionsa
Sp act (nmol ofNADPH produced/min/mg of protein)b
in cell extracts
HPS PHI HPS 1 PHI
168 wild type LB ,1 ,1 NDc
LB 1 0.5 mM FA 690 ND 310LB 1 1.0 mM FA 95 ND ,1LB 1 2.0 mM FA ,1 ND ,1LB 1 2% (vol/vol)
MeOH,1 ND ,1
LB 1 0.5 mM FM ,1 ND ,1YD101 (yckG) LB 1 0.5 mM FA ,1 ,1 NDYD102 (yckF) LB 1 0.5 mM FA 290 ,1 NDYD111 (yckH) LB 1 0.5 mM FA ,1 ,1 ,1YD112 (yckH) LB 1 0.5 mM FA ,1 ,1 ,1YD121 (yckH yckH1) LB 1 0.5 mM FA 1,050 ND 200
a FA, formaldehyde; MeOH, methanol; FM, formate.b Mean value from three experiments.c ND, not determined.
7158 YASUEDA ET AL. J. BACTERIOL.
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
ences in the B. subtilis strains used. Either way, the existence oftwo key enzymes was substantially established by our research.
The organization of divergent transcription between the twotranscription terminators in the B. subtilis genome suggestedthat yckH may be a regulator gene (24). The putative geneproduct of yckH is estimated to be a polypeptide composed of120 amino acids (41). However, our disruption experimentindicated that the yckH gene is required for the expression ofHPS and PHI activities, and it appears that yckH (now namedhxlR) may positively regulate yckG-yckF gene expressionthrough DNA binding either directly or indirectly after stimu-lation by formaldehyde. We also suggest that yckG and yckF(now named hxlA and hxlB, respectively) are organized into anoperon structure, based on the following data: (i) the existenceof mRNA species covering the hxlA and hxlB ORFs, (ii) thepolar effect between hxlA and hxlB genes, (iii) the simulta-neous expression of the two genes in response to an inducer,(iv) the lack of hxlB expression concomitant with abolition ofhxlA expression in an hxlR (yckH)-deficient mutant, and (v) thegene organization of hxlA-hxlB followed by a typical transcrip-tional terminator. Recently, it was shown that M. gastri MB19contains a phi (rmpB)-hps (rmpA) operon (28), although the
hps (rmpA) and phi (rmpB) genes of M. aminofaciens 77a areseparated by an insertion element (35). We found that the hps(hxlA)-phi (hxlB) gene organization was conserved as a set ofboth genes even in a nonmethylotroph. Curiously, the genes ofB. subtilis and M. gastri were arranged in reverse order. There-fore, more detailed bioinformatic analysis of the conservedfeatures and differences in gene organization among threetypes of microbes (facultative methylotrophs, obligate methy-lotrophs, and nonmethylotrophs) may provide insight into theevolution of the RuMP pathway.
ACKNOWLEDGMENTS
We gratefully acknowledge N. Kato, Y. Sakai, and R. Mitsui (KyotoUniversity) for the generous gift of the enzyme samples (HPS and PHIfrom M. aminofaciens 77a) and for helpful discussion; E. Suzuki and N.Ootu for NMR analysis; and K. Sato, M. Ooba, and K. Kobayashi fortechnical support.
REFERENCES
1. Anagnostopoulos, C., and J. Spizizen. 1961. Requirement for transformationin Bacillus subtilis. J. Bacteriol. 81:741–746.
2. Arfman, N., E. M. Watling, W. Clement, R. J. van Oosterwijk, G. E. de Vries,
FIG. 4. Effect of formaldehyde on the growth of B. subtilis. Overnight cultures of wild-type strain 168 ( ), YD101 (yckG) (•••••), and YD102 (yckF) (– – –)in LB medium were inoculated into prewarmed LB medium containing formaldehyde at concentrations of 0.0 (a), 0.3 (b), 0.6 (c), and 0.9 (d) mM. Growth and lysisof cells at 37°C were monitored automatically at an OD of 660 nm.
VOL. 181, 1999 ONE-CARBON COMPOUND METABOLISM IN B. SUBTILIS 7159
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

W. Harder, M. M. Attwood, and L. Dijkhuizen. 1989. Methanol metabolismin thermotolerant methylotrophic Bacillus strains involving a novel catabolicNAD-dependent methanol dehydrogenase as a key enzyme. Arch. Micro-biol. 152:280–288.
3. Arfman, N., L. Bystrykh, N. I. Govorukhina, and L. Dijkhuizen. 1990. 3-Hex-ulose-6-phosphate synthase from thermotolerant methylotrophic BacillusC1. Methods Enzymol. 188:391–397.
4. Arfman, N., K. J. de Vries, H. R. Moezelaar, M. M. Attwood, G. K. Robinson,M. van Geel, and L. Dijkhuizen. 1992. Environmental regulation of alcoholmetabolism in thermotolerant methylotrophic Bacillus strains. Arch. Micro-biol. 157:272–278.
5. Attwood, M. M., and J. R. Quayle. 1984. Formaldehyde as a central inter-mediary metabolite of methylotrophic metabolism, p. 315–323. In R. L.Crawford and R. S. Hanson (ed.), Proceedings of the 4th InternationalSymposium on Microbial Growth on C1 Compounds. American Society forMicrobiology, Washington, D.C.
6. Attwood, M. M. 1990. Formaldehyde dehydrogenases from methylotrophs.Methods Enzymol. 188:314–327.
7. Barns, S. M., C. F. Delwiche, J. D. Palmer, and N. R. Pace. 1996. Perspec-tives on archeal diversity, thermophily and monophyly from environmentalrRNA sequences. Proc. Natl. Acad. Sci. USA 93:9188–9193.
8. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye bind-ing. Anal. Biochem. 72:248–251.
9. De Vries, G. E., N. Arfman, P. Terpstra, and L. Dijkhuizen. 1992. Cloning,expression, and sequence analysis of the Bacillus methanolicus C1 methanoldehydrogenase gene. J. Bacteriol. 174:5346–5353.
10. De Wulf, P. 1998. Presence of the ribulose monophosphate pathway inBacillus subtilis. Microbiology 144:596–597. (Comment.)
11. Dijkhuizen, L., P. R. Levering, and G. E. de Vries. 1992. The physiology andbiochemistry of aerobic methanol-utilizing Gram-negative and Gram-posi-tive bacteria, p. 149–181. In J. C. Murrell and H. Dalton (ed.), Methane andmethanol utilizers. Plenum Press, New York, N.Y.
12. Ferenci, T., T. Strøm, and J. R. Quayle. 1974. Purification and properties of3-hexulose phosphate synthase and phospho-3-hexuloisomerase from Methy-lococcus capsulatus. Biochem. J. 144:477–486.
13. Guerout-Fleury, A.-M., K. Shazand, N. Frandsen, and P. Stragier. 1995.Antibiotic-resistance cassettes for Bacillus subtilis. Gene 167:335–336.
14. Gutheil, W. G., B. Holmquist, and B. L. Vallee. 1992. Purification, charac-terization, and partial sequence of the glutathione-dependent formaldehydedehydrogenase from Escherichia coli: a class III alcohol dehydrogenase.Biochemistry 31:475–481.
15. Hanson, R. S., and T. E. Hanson. 1996. Methanotrophic bacteria. Microbiol.Rev. 60:439–471.
16. Honikoff, S., G. W. Haughn, J. M. Calvo, and J. C. Wallance. 1988. A largefamily of bacterial activator proteins. Proc. Natl. Acad. Sci. USA 85:6602–6606.
17. Johnson, P. A., and J. R. Quayle. 1965. Microbial growth on C1 compounds.Synthesis of cell constituents by methane- and methanol-grown Pseudomo-nas methanica. Biochem. J. 95:859–867.
18. Kato, N., H. Ohashi, T. Hori, Y. Tani, and K. Ogata. 1977. Properties of3-hexulose phosphate synthase and phospho-3-hexuloisomerase of a meth-anol-utilizing bacterium, 77a. Agric. Biol. Chem. 41:1133–1140.
19. Kato, N., H. Ohashi, Y. Tani, and K. Ogata. 1978. 3-Hexulosephosphatesynthase from Methylomonas aminofaciens 77a: purification, properties andkinetics. Biochim. Biophys. Acta 523:236–244.
20. Kato, N., N. Miyamoto, M. Shimao, and C. Sakazawa. 1988. 3-Hexulosephosphate synthase from a new facultative methylotroph, Mycobacteriumgastri MB19. Agric. Biol. Chem. 52:2659–2661.
21. Kelly, D. P., G. Malin, and A. P. Wood. 1993. Microbial transformations andbiogeochemical cycling of one-carbon substrates containing sulphur, nitro-gen or halogens, p. 47–64. In J. C. Murrell and D. P. Kelly (ed.), Microbialgrowth on C1 compounds. Intercept Ltd., Andover, Mass.
22. Kemp, M. B., and J. R. Quayle. 1967. Microbial growth on C1 compounds.Uptake of (14C)formaldehyde and (14C)formate by methane-grown Pseudo-monas methanica and determination of the hexose labelling pattern afterbrief incubation with (14C)methanol. Biochem. J. 102:94–102.
23. Kemp, M. B. 1974. Hexulose phosphate synthase from Methylococcus cap-sulatus makes D-arabino-3-hexulose phosphate. Biochem. J. 139:129–134.
24. Kunst, F., N. Ogasawara, I. Moszer, et al. 1997. The complete genomesequence of the Gram-positive bacterium Bacillus subtilis. Nature (London)390:249–256.
25. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature (London) 227:680–685.
26. Large, P., and J. Green. 1984. Oxidation of mono-, di-, and trimethylamineby methanotrophic yeasts: properties of the microsomal and peroxisomalenzymes involved and comparison with bacterial enzyme systems, p. 155–164.In R. L. Crawford and R. S. Hanson (ed.), Microbial growth on C1 com-pounds. American Society for Microbiology, Washington, D.C.
27. Levering, P. R., L. M. Croes, and L. Dijkhuizen. 1985. Regulation of meth-ylamine and formaldehyde metabolism in Arthrobacter P1. Formaldehyde isthe inducing signal for the synthesis of the RuMP cycle enzyme hexulosephosphate synthase. Arch. Microbiol. 144:272–278.
28. Mitsui, R. 1998. Organization and regulation of the genes involved in theribulose monophosphate pathway in methylotrophic bacteria. Ph.D. thesis.Kyoto University, Kyoto, Japan.
29. Mullis, K. B., and F. A. Faloona. 1987. Specific synthesis of DNA in vitro viaa polymerase catalyzed chain reaction. Methods Enzymol. 155:335–350.
30. Pearson, W. R., and D. J. Lipman. 1988. Improved tools for biologicalsequence comparison. Proc. Natl. Acad. Sci. USA 85:2444–2448.
31. Person, W. R. 1990. Rapid and sensitive sequence comparison with FASTPand FASTA. Methods Enzymol. 183:63–98.
32. Quayle, J. R. 1982. 3-Hexulose-6-phosphate synthase from Methylomonas(Methylococcus) capsulatus. Methods Enzymol. 90:314–319.
33. Reizer, J., A. Reizer, and M. H. Saier, Jr. 1997. Is the ribulose monophos-phate pathway widely distributed in bacteria? Microbiology 143:2519–2520.(Comment.)
34. Sahm, H., H. Schutte, and M.-R. Kula. 1982. 3-Hexulose-phosphate synthasefrom Methylomonas M15. Methods Enzymol. 90:319–323.
35. Sakai, Y., R. Mitsui, Y. Katayama, H. Yanase, and N. Kato. 1999. Organi-zation of the genes involved in the ribulose monophosphate pathway in anobligate methylotrophic bacterium, Methylomonas aminofaciens 77a. FEMSMicrobiol. Lett. 176:125–130.
36. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
37. Sauer, U., V. Hatzimanikatis, H.-P. Hohmann, M. Manneberg, A. P. G. M.van Loon, and J. E. Bailey. 1996. Physiology and metabolic fluxes of wild-type and riboflavin-producing Bacillus subtilis. Appl. Environ. Microbiol. 62:3687–3696.
38. Sauer, U., V. Hatzimanikatis, J. E. Bailey, M. Hochuli, T. Szyperski, and K.Wuthrich. 1997. Metabolic fluxes in riboflabin-producing Bacillus subtilis.Nature Biotechnol. 15:448–452.
39. Schleif, R. 1992. Regulation of the L-arabinose catabolic operon araBAD, p.643–665. In S. L. McKnight and K. R. Yamamoto (ed.), Transcriptionalregulation, vol. 2. Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, N.Y.
40. Speer, B. S., L. Chistoserdova, and M. E. Lidstrom. 1994. Sequence of thegene for a NAD(P)-dependent formaldehyde dehydrogenase (class III alco-hol dehydrogenase) from a marine methanotroph Methylobacter marinusA45. FEMS Microbiol. Lett. 121:349–356.
41. SubtiList. 1997. Data release R14.2. In I. Mozer and A. Danchin (ed.),http://www.pasteur.fr./Bio/SubtiList.html.[Online.] Institut Pasteur, Paris,France.
42. Van Spanning, R. J. M., C. W. Wansell, T. De Boer, M. J. Hazelaar, H.Anazawa, N. Harms, L. F. Oltmann, and A. H. Stouthamer. 1991. Isolationand characterization of the moxJ, moxG, moxI, and moxR genes of Paracoc-cus denitrificans: inactivation of moxJ, moxG, and moxR and the resultanteffect on methylotrophic growth. J. Bacteriol. 173:6948–6961.
43. Woese, C. R. 1987. Bacterial evolution. Microbiol. Rev. 51:221–271.44. Yanase, H., K. Ikeyama, R. Mitsui, S. Ra, K. Kita, Y. Sakai, and N. Kato.
1996. Cloning and sequence analysis of the gene encoding 3-hexulose-6-phosphate synthase from the methylotrophic bacterium, Methylomonasaminofaciens 77a, and its expression in Escherichia coli. FEMS Microbiol.Lett. 135:201–205.
45. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. Improved M13 phagecloning vectors and host strains: nucleotide sequences of the M13mp18 andpUC19 vectors. Gene 33:103–119.
46. Yasueda, H., K. Nakanishi, Y. Kumazawa, K. Nagase, M. Motoki, and H.Matsui. 1995. Tissue-type transglutaminase from red sea bream (Pagrusmajor): sequence analysis of the cDNA and functional expression in Esche-richia coli. Eur. J. Biochem. 232:411–419.
7160 YASUEDA ET AL. J. BACTERIOL.
on April 7, 2019 by guest
http://jb.asm.org/
Dow
nloaded from





























