Embed Size (px)
Citation preview

Serodiugnosis and Immunotherapy in Infectious Disease (1989) 3, 149-l 59
Review article
Bacterial heat-shock proteins and serodiagnosis
Introduction
The application of hybridoma and recombi- nant DNA techniques to the study of bacterial antigens and virulence factors has yielded some remarkable and unexpected findings. One star- tling discovery is that heat-shock proteins feature as immunodominant antigens in the mammalian immune response to many mi- crobial pathogens. These proteins are synthe- sized in large amounts by cells in response to a rapid increase in temperature (‘heat shock’)‘J. They are among the most highly conserved of all proteins, and are produced by all living cells, both eucaryotic and procaryotic’.*. That these proteins are important antigens in many bac- terial pathogens has implications for serodiag- nosis of bacterial infection, vaccine develop- ment, and the study of autoimmunity. This review will give an account of the discovery of heat-shock protein antigens, consider their dis- tribution among pathogenic bacteria, discuss why they should be so immunogenic, and whether they are of use in serodiagnosis, and finally speculate on future avenues for research on these proteins. I shall concentrate on pro- caryotic pathogens-readers interested in para- site heat-shock proteins should consult the review by Newport, Culpepper & Agabian’.
Mycobacterial beat-shock proteins
During the early 1980’s many different labora- tories raised monoclonal antibodies against Mycohacterium tuberculosis and Mycobacter- ium leprae to characterize the antigens of these organisms. The antigenic specificity and cross- reactivity of over 50 anti-mycobacterial monoclonals were compared in two WHO- sponsored workshops in 1984 and 198V. De- spite both the large number of proteins present in mycobacteria, and the variations in experi- mental protocols used, one particular protein,
of molecular weight 65 kD, was recognized by antibodies from every laboratory involved. Richard Young and colleagues used monoclo- nal antibodies against this and other mycobac- terial proteins to probe genomic DNA libraries made from M. leprae and M. tuberculosis DNA6J. They cloned the genes for the 65 kD antigens of both mycobacterial species, as well as the genes for several other monoclonally- defined antigens”,‘. The 65 kD antigen gene of M. bovis BCG was cloned by a Dutch group, who probed their genomic library with rabbit hyperimmune serum*. Simultaneously Belgian researchers purified the 65 kD protein from filtrates of zinc-deficient BCG cultures, and determined the N-terminal sequence of this proteinlo.
The genes encoding the 65 kD antigen of M. tuberculosis, M. leprae and BCG and several other of the genes first cloned by Young’s group have now been sequenced9,“-“. Compari- son of the derived amino-acid sequences of these antigens with other known sequences has shown that several of the most prominent mycobacterial antigens are very similar to known heat-shock proteins. The derived amino-acid sequence of the 65 kD antigen of M. tuberculosis is identical in 54% of residues to that of GroEL, a major heat-shock protein of E. cofi’*. The derived sequence of the BCG protein is identical to that of the M. tuberculo- sis antigen, while the M. Ifprae 65 kD antigen sequence is identical in 81% of its amino-acid sites to the other two mycobacterial homolo- gues9J2.1x,
Mycobacterial antigens of molecular weights 70 kD (M. leprae) and 71 kD (M. tuberculosis) are closely related to similar sized heat-shock proteins found in E. coli and many eucaryotes (the hsp70 family). The equivalent protein in E. coli, DnaK is identical in 5 1% of the sequence of its carboxy-terminal half to the same region in the M. leprae protein’5m’7. Even the C-termi- nus of Drosophila melanogaster hsp70 is identi-
149 0888-0786/89/030149 + 11 $03.00/O cr 1989 Academic Press Limited

150 Review article
cal in 42% of residues to the same region of the M. leprae antigen”. An I8 kD monoclonally- defined antigen of M. leprae is identical in 3 I % of 127 amino acids to a soybean heat-shock protein, 17.5E, one of a member of related eucaryotic low molecular weight heat-shock proteins14. Recently, Baird and colleagues have shown that a l&l2 kD antigen of BCG, termed BCG-a, is identical in 45% of its derived amino-acid sequence to GroES, a heat-shock protein of E. coli (where it is part of the same operon as GroEL)19.
In addition to the sequence data, there is some experimental evidence to suggest that the most prominent mycobacterial protein anti- gens, the 65 kD and 70/71 kD proteins, actually function as heat-shock proteins in mycobac- teria. Shinnick and colleagues report that a 62 kD protein of M. smegmatis, which cross- reacts with monoclonal antibodies against the 65 kD antigen, is synthesized in large amounts after heat-shock treatment of M. smegmatis cells’*. A protein of molecular weight 72 kD is also synthesized in response to heat shock by M. smegmatis-it probably corresponds to the 70171 kD antigen of M. tuberculosis and M. leprue’*. Shinnick and colleagues have also shown that the promoter for the 65 kD antigen gene is regulated by an increase in temperature. Escherichiu coli cells in which the 1acZ gene has been placed under the control of this promoter express more beta-galactosidase at 42°C than at lower temperaturesIn.
Heat-shock protein antigens in other bacteria
Polyclonal antisera and some monoclonal anti- bodies reactive with the mycobacterial 65 kD antigens bind to similar-sized proteins in extracts from a large variety of bacteria, includ- ing pathogens as diverse as Streptococcus pyo- genes and Campylobacter jejuni, or Treponema pallidurn and Clostridium dtJiciF’. It has recently become obvious that these mycobac- terial antigens are examples of a protein anti- gen termed ‘common antigen’ first described in Pseudomonas aeruginosa over ten years ago, and known to be present in over 50 different bacterial specieszo. Cross-reactive proteins are found even in archaebacteria, suggesting that the gene for this protein dates back over three billion years to the common ancestor of all living organismszo. Indeed, plants, animals and fungi harbour proteins related to GroEL and the mycobacterial 65 kD proteins2’-23.
Although proteins related to GroEL and the
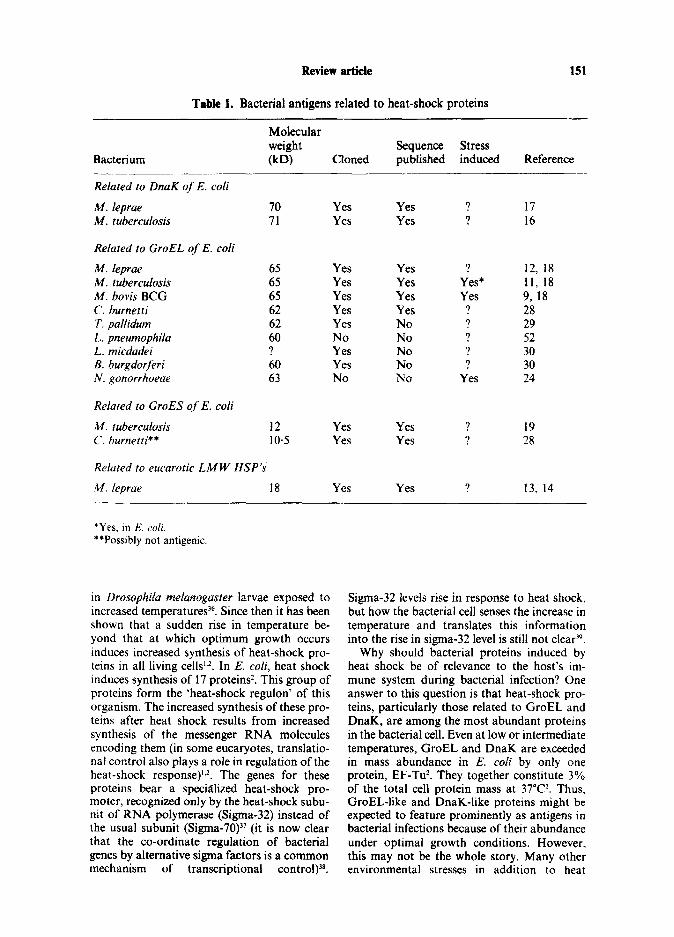
mycobacterial 65 kD antigens (henceforth all termed ‘GroEL-like proteins’) probably occur in all bacterial species, in only a few pathogenic bacteria have they been shown to be induced by heat shock, or other stresses (see Table 1). Neisseria gonorrhoeae produces a 63 kD pro- tein in response to a variety of environmental stresses, and a monoclonal to this protein binds to the 65 kD antigen of M. tuberculosis24. C56, thought to be the GroEL-like protein of Safmo- nella typhimurium, is induced by heat-shock, and antibodies to the molecule are dominant in the murine antibody response to this organ- ism25.26. Pseudomonas aeruginosa produces a 61 kD heat-shock protein, which cross-reacts immunologically with GroEL2’.
Further evidence that GroEL-like proteins are important bacterial antigens comes from several studies in which polyclonal sera were used to probe genomic libraries from various bacterial pathogens. Recombinants containing the gene for the GroEL-like protein of Coxiellu burnettii have been identified with both animal serum and serum from a Q fever patient2*. The derived amino acid sequence of the 62 kD Cox- iella antigen is identical in 55% of the first 520 amino acids to that of the 65 kD antigen of M. leprae2*. A seemingly non-antigenic 10.5 kD protein, related to GroES, is encoded by a gene within the same operon as the Coxiellu GroEL- like genez8. The genes for the GroEL-like pro- teins of Treponema pallidurn and Borrelia burg- dorferi have been cloned using similar approaches, but sequence data has yet to be published 29.30.
Although proteins related to DnaK are induced by heat shock in Salmonella typhi- murium and Pseudomonas aeruginosa25.27, DnaK-like proteins have yet to be shown to be important antigens in bacteria other than mycobacteria. Members of the hsp 70 family are definitely important antigens in eucaryotic parasites such as Plasmodium sp., and Schisto- soma SP.“-‘~ Whether their apparent absence as antigens in bacterial pathogens is because they are generally not antigenic in procaryotic pathogens, or because they have not yet been studied adequately is not known. A similar point applies to other bacterial heat-shock proteins, such as those related to the M. leprae 18 kD antigen and to GroES.
Why are heat-shock proteins antigens?
The heat-shock response was first described in 1962 as a pattern of chromosome puffing seen
Review article
Table 1. Bacterial antigens related to heat-shock proteins
151
Molecular weight Sequence Stress
Bacterium 00 Cloned published induced Reference -__ Related to DnaK of E. coli
M. leprae IQ Yes Yes ? 17 M. tuberculosis 71 Yes Yes ? 16
Related to GroEL of E. coli
h4. ieprae 65 Yes Yes ? 12, 18 M. tuberculosis 65 Yes Yes Yes* 11, 18 M. bovis BCG 65 Yes Yes Yes 9, 18 C. burnetti 62 Yes Yes ? 28 T. pallidurn 62 Yes No ? 29 L. pneumophila 60 No No ? 52 L. micdadei ? Yes No ? 30 B. burgdorferi 60 Yes No ? 30 N. gonorrhoeae 63 NO No Yes 24
Related to GroES of E. coli
M. tubertulosis 12 Yes Yes . 7 19 C. burnetti** 10.5 Yes Yes ? 28
Related to eucarotic LMW HSP’s
M. leprae 18 Yes Yes 7 13, 14
*Yes, in E. coli. **Possibly not antigenic.
in Drosophila meianogaster larvae exposed to increased temperatures 36. Since then it has been shown that a sudden rise in temperature be- yond that at which optimum growth occurs induces increased synthesis of heat-shock pro- teins in all living cells ‘J. In E. coli, heat shock induces synthesis of 17 proteins2. This group of proteins form the ‘heat-shock regulon’ of this organism. The increased synthesis of these pro- teins after heat shock results from increased synthesis of the messenger RNA molecules encoding them (in some eucaryotes, translatio- nal control also plays a role in regulation of the heat-shock response)‘J. The genes for these proteins bear a specialized heat-shock pro- moter, recognized only by the heat-shock subu- nit of RNA polymerase (Sigma-32) instead of the usual subunit (Sigma-70)” (it is now clear that the co-ordinate regulation of bacterial genes by alternative sigma factors is a common mechanism of transcriptional control)38.
Sigma-32 levels rise in response to heat shock, but how the bacterial cell senses the increase in temperature and translates this information into the rise in sigma-32 level is still not cleaP.
Why should bacterial proteins induced by heat shock be of relevance to the host’s im- mune system during bacterial infection? One answer to this question is that heat-shock pro- teins, particularly those related to GroEL and DnaK, are among the most abundant proteins in the bacterial cell. Even at low or intermediate temperatures, GroEL and DnaK are exceeded in mass abundance in E. co/i by only one protein, EF-TtP. They together constitute 3% of the total cell protein mass at 37”C*. Thus, GroEL-like and DnaK-like proteins might be expected to feature prominently as antigens in bacterial infections because of their abundance under optimal growth conditions. However. this may not be the whole story. Many other environmental stresses in addition to heat

152 Review article
shock cause increased synthesis of heat-shock proteins such as GroEL and DnaK’,*. Exposing bacteria to noxious stimuli such as hydrogen peroxide, ethanol, quinolones, or U.V. irradia- tion induces synthesis of novel proteins’JJS. Similarly, new proteins are also produced after amino-acid or glucose deprivation”‘. The groups of proteins induced by these stresses contain some proteins common to all stress regulons, as well as some proteins produced only in response to a very particular stress (e.g. catalase is induced by hydrogen peroxide)25. GroEL-like and DnaK-like proteins are pro- duced in response to most stresses, and hence are better termed ‘stress proteins’, rather than being narrowly defined as heat-shock proteins (see Figure 1).
It seems likely that stress proteins are recog- nized so readily by the host’s immune system because they are produced in massive amounts by bacteria during infection’5,4’. The host en- vironment is very stressful to the invading microbe15.4’. The temperature is higher than that of the external environment, and if fever is present, may be at the upper limit of the bacterium’s viable temperature range. Host
body fluids, especially during the acute phase response, are very low in metal ions, such as iron and zinc. Bacteria, once phagocytosed, are exposed to intensive oxidative stress in the phagolysosome. Extracellular organisms may be lysed by complement. Faced with such in- tense stresses, some of which probably act in synergy, the bacteria are likely to switch on both defensive and offensive regulons. (Note that virulence factors are often co-ordinately regulated and induced by stressful environmen- tal conditions42,43.) One can perhaps view the bacterial stress response as a mirror image of the host’s acute-phase and inflammatory res- ponses (see Figure 2). Interestingly, synthesis of host stress proteins is increased in many of the host cells involved in inflammation”‘.
No-one has yet shown directly that bacterial stress proteins are produced in large amounts during infection. However, in vitro findings support the above scenario. GroEL and GroES constitute 15% of cell protein mass in E. cofi grown at high temperature*. The 65 kD antigen accounts for about 9% of all cellular protein in M. bovis BCG grown in zinc-deficient cul- tures’O. Oxidative stress is known to increase
Figure I. Schematic diagram showing overlap brtwrrn ihrrr diffrrrnt stress rrgulonr in Salmon#//o fyphimurium (barrd on dota in ref. 25)
Common core stress proteins, e.g. GroEL, DnaK homologues
Figure 1.

Review article 153
Figure 2.Thr hort/pathogsn interaction
Acute-Dhase r( Serum Zinc falls 1 GroEL EL DnaK svnthesis 1
Stress resoonse
Figure 2.
synthesis of GroEL-like and DnaK-like pro- teins in Enterobacteriaceae25. Nonetheless, it would be interesting if large amounts of bacter- ial stress proteins could be demonstrated dir- ectly in infected tissues, such as in abscesses, or granulomas.
Functions of stress proteins
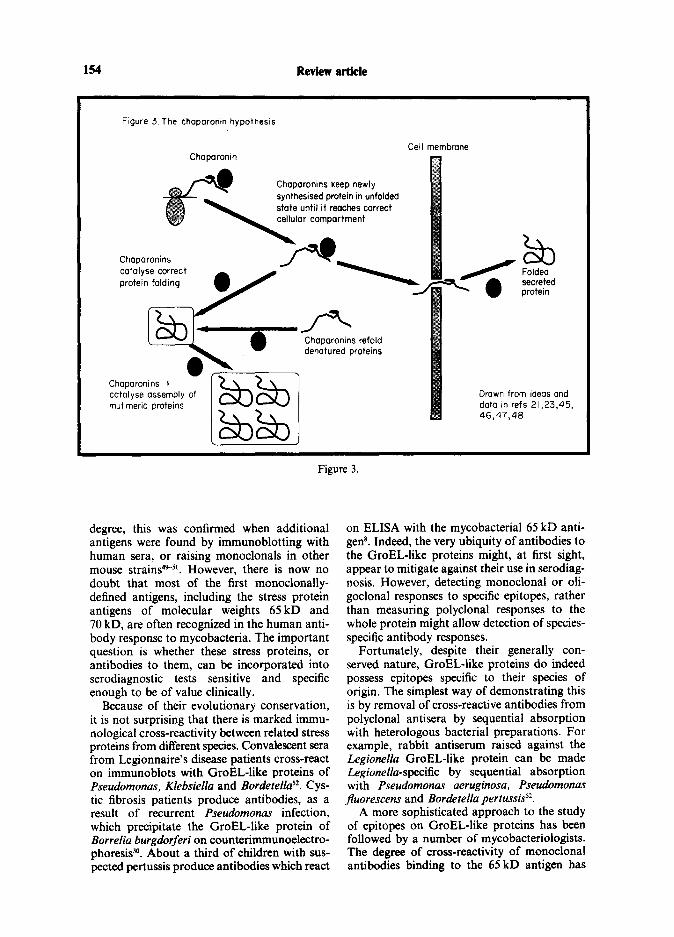
The value of some of the bacterial proteins induced by stress is obvious-for example, the production of catalase by Salmonella typhimur- ium in response to hydrogen peroxide protects the organism against oxidative stress*‘. How- ever, the functions of many other stress pro- teins are less clear, particularly those related to DnaK and GroEL. Drawing on many years of experimental work on these proteins, Ellis and colleagues have suggested that these two groups of molecules belong to a class of pro- teins termed ‘chaparonins’21.45. The proposed role of these chaparonins is to ensure that newly synthesized polypeptides fold into the correct tertiary and quaternary structures, and to ‘chaparone’ them as they move across mem- branes, either during secretion across the cell
membrane, or, in eucaryotes, during transloca- tion from one cellular compartment to another. Translocation across membranes requires pro- teins to be in an unfolded state, and chaparo- nins are thought to catalyse the folding and unfolding events before and after transloca- tion. In addition, those molecules probably refold proteins which have been denatured by heat-shock and other stresses (see Figure 3). Ellis and colleagues most recently invoked the chaparonin hypothesis after finding two types of GroEL-like polypeptides in a multimeric protein which catalyses assembly of the abun- dant plant enzyme Rubisco. Other evidence has recently been produced in support of their idea from studies of both eucaryotic and procaryo- tic DnaK-like and GroEL-like proteins23,w8.
Stress proteins-their importance in serodiagnosis
After the WHO-sponsored workshops on anti- mycobacterial monoclonal antibodies, it was often suggested that the limited set of antigens defined by such monoclonals was an artefact of the Ba1b.C mouse’s immune system@. To some
154 Review article
Figure 3 The chaparontn hypothesis
Chaparonin
Chaparonins catalyse correct protein folding
CC ?II membrane
/ -’ Chaparomns refold denatured proteins
I
Folded secreted protein
Chaparonins 3 catalyse assembly of mutlmeric proteins
Drawn from ideas and dato In refs 21,23,45, 46,47,40
Figure 3.
degree, this was confirmed when additional antigens were found by immunoblotting with human sera, or raising monoclonals in other mouse strainPsl. However, there is now no doubt that most of the first monoclonally- defined antigens, including the stress protein antigens of molecular weights 65 kD and 70 kD, are often recognized in the human anti- body response to mycobacteria. The important question is whether these stress proteins, or antibodies to them, can be incorporated into serodiagnostic tests sensitive and specific enough to be of value clinically.
Because of their evolutionary conservation, it is not surprising that there is marked immu- nological cross-reactivity between related stress proteins from different species. Convalescent sera from Legionnaire’s disease patients cross-react on immunoblots with GroEL-like proteins of Pseudomonas, Klebsiella and Bordetella”. CYS- tic fibrosis patients produce antibodies, as a result of recurrent Pseudomonas infection, which precipitate the GroEL-like protein of Borrelia burgdorferi on counterimmunoelectro- phoresi9. About a third of children with sus- pected pertussis produce antibodies which react
on ELISA with the mycobacterial 65 kD anti- gen*. Indeed, the very ubiquity of antibodies to the GroEL-like proteins might, at first sight, appear to mitigate against their use in serodiag- nosis. However, detecting monoclonal or oli- goclonal responses to specific epitopes, rather than measuring polyclonal responses to the whole protein might allow detection of species- specific antibody responses.
Fortunately, despite their generally con- served nature, GroEL-like proteins do indeed possess epitopes specific to their species of origin. The simplest way of demonstrating this is by removal of cross-reactive antibodies from polyclonal antisera by sequential absorption with heterologous bacterial preparations. For example, rabbit antiserum raised against the Legionella GroEL-like protein can be made Legionella-specific by sequential absorption with Pseudomonas aeruginosa, Pseudomonas Jluorescens and Bordetella pertussis52.
A more sophisticated approach to the study of epitopes on GroEL-like proteins has been followed by a number of mycobacteriologists. The degree of cross-reactivity of monoclonal antibodies binding to the 65 kD antigen has

Review article 155
been examined in detail, studying the binding of monoclonals to epitopes on GroEL-like pro- teins in several dozen bacterial species by immunoblotting*“. Epitopes have been mapped to continuous amino-acid sequences on the 65 kD mycobacterial antigen using both recombinant DNA and synthetic peptide approaches5’-55. Indeed, the mycobacterial 65 kD antigen is now probably the most thor- oughly defined of all bacterial antigens in terms of B and T cell epitopes.
Given this intense study, has the 65 kD anti- gen proven useful in the serodiagnosis of tuber- culosis and leprosy? Unfortunately not, at least, not so far. High antibody titres to this protein are found on ELISA in tuberculosis patients, household contacts, and in healthy contro1P. Thus, measuring polyclonal res- ponses to this antigen allows no discrimination between healthy and infected individuals. Anti- body titres to a monoclonally-defined epitope on the protein (TB78 epitope) were higher in smear-positive tuberculosis patients’ sera than in controls in a recent study, but monoclonals to other mycobacterial antigens give far better resultP. However, when considering these dis- appointing results with tuberculosis patients,’ sera one must realise that only one of the many possible epitopes on the GroEL-like molecule has been investigated-perhaps better discrimi- nation can be obtained with other monoclonals or with synthetic peptide epitopes. Coates and colleagues, in studying the antibody response in Chinese smear-negative tuberculosis patients by immunoblotting, found that 14/34 patients recognised the 65 kD molecule, as compared with 7/36 controls, again a difference not marked enough to be useful diagnostically”. Interestingly four patients recognized a lo- 12 kD molecule, probably the mycobacterial GroES-like protein, whilst no control sera con- tained antibodies to this moleculeso.
Disappointing results have also been obtained when studying sera of leprosy patients. Rather surprisingly, no antibody to the 65 kD antigen could be detected in a study of leprosy patient’s sera by immunoblotting”. Technical, rather than biological reasons may account for this lack of success in detecting anti-65 kD antigen antibodies in leprosy sera- GroEL-like proteins are notoriously unstable under denaturing conditionP.
The usefulness of the 70 kD antigen of M. ieprue in the serodiagnosis of leprosy is uncer- tain. Eight of fifteen individual BL/LL sera precipitated the antigen in an early study by Britton and colleagues, but no BT/TT sera
reacted with it”. There is no published data on the frequency of anti-70 kD antigen antibodies in healthy controls or household contacts of leprosy patients.
Thus, to date, the cloned mycobacterial heat-shock proteins have been unhelpful in serodiagnosis.
Future prospects
Despite the disappointing results of the studies on serodiagnosis of mycobacterial disease, the discovery of the dominance of highly conserved stress proteins in the human antibody response to bacterial pathogens opens many exciting new avenues for research. Given their import- ance in bacterial diseases as diverse as Lyme disease, Legionellosis, Q fever, tuberculosis, leprosy and gonorrhoea, it seems likely that stress proteins figure prominently as antigens in almost all invasive bacterial diseases. The way is now open to define the heat-shock responses in all important bacterial pathogens. A recent paper on the heat-shock response in Pseudomo- nas aeruginosa shows how heat-shock proteins can be defined in a given microorganism by simple polyacrylamide gel electrophoresisl’. A more sophisticated approach is 2-D gel electro- phoresis. One potential adjunct to these approaches would be to blot proteins induced by heat shock or other stress (e.g. oxidative stress) in a given bacterium onto nitrocellulose, and then probe directly with the human sera from patients. This approach is likely to pick up antigens not evident when cell sonicates from bacteria grown under optimal conditions. without stress, are bIotted and probed with human sera. An alternative avenue for research is to attempt to clone the genes for heat-shock proteins of one pathogen and screen by probes derived from the equivalent gene of another pathogen. This approach depends on the high degree of conservation in both the genes and proteins involved in the heat-shock response. Bardwell & Craig have shown that this strategy is feasible. They identified the homologue in E. coil of the eucaryotic hsp38 heat-shock pro- teirP. They probed E. coli genomic DNA with Drosophila hsp83 probes on Southern blots washed at low stringency, and then cloned the hybridizing sequences in a partial DNA libraryS8. A related approach, is to use pub- lished sequence data to devise oligonucleotide probes which correspond to those short patches of a heat-shock protein’s sequence which are most highly conserved (for an example, see Figure 4). This approach has already been tried

Review Article
Esch. coli (amino acids 163-8) Ile Ala Olu Ala Met Asp
Cuxielle bornetti (amino acids 163-8) Ile Ala Qlu Ala Met Asp
Mycubecterlum sp. (amino acids 163-8) Ile Ala Glu Ala Met (31~
Mixed probe sequence ATTGCTGAAGCTATGGA c c cc
Probeefficlencycanbeimproved ifcodon preferences ofspecies under study are known
Figure 4. A short conserved sequence,common to the GroEL-like proteins of three distontly related bacterial genera, which can be used to design on oligonucleotide probe for use with other bacterial pathogens
Figure 4.
with other proteins, which contain small patches of amino acid sequences conserved between distantly related organisms59.
An further area of research is the study of the link between autoimmunity and heat-shock proteins. T cells responding to an epitope in the mycobacterial65 kD antigen (amino acids 1% 188) have been shown to induce immunopatho- logy in a rat model of autoimmune disease (adjuvant arthritis)w. Surprisingly a T cell clone reacting to this same epitope was also able to protect against this disease@. Douglas Young and colleagues have shown that human T cells can recognize epitopes on this molecule which are conserved between the human and myco- bacterial proteins4’. Antibodies to a member of the human hsp70 family have been found in over a third of sera from patients with systemic lupus erythematosi#. Antibodies to another high molecular weight heat-shock protein are also found in 39% of patients with ankylosing spondylitisg,u. Given the high degree of conser- vation between human and bacterial heat- shock proteins, it is quite plausible that bacter- ial heat-shock proteins are involved in trigger- ing autoimmune reactions. This possibility makes them less attractive as subunit vaccines,
despite their high antigenicity. One final question of interest is the relation-
ship between stress and virulence. Many viru- lence factors are co-ordinately regulated by environmental conditions. In an intriguing short paper Sokolovic & Goebel have recently suggested that the synthesis of listerolysin, a known virulence factor of Listeria monocyfo- genes, may be increased after heat shocka. How many other virulence factors are induced in response to the same stresses that upregulate the synthesis of the common core stress pro- teins?
In conclusion, the unexpected finding that the first mycobacterial protein antigens to be well characterized are heat-shock proteins has already given the heat-shock response a new prominence in medical research. These heat- shock proteins, already present over three bil- lion years ago in the common ancestor of all living organisms, are certain to keep scientists, especially those interested in serodiagnosis, busy for a long time to come.
MARK PALLEN Department of Medical Microbiology,
St Bartholomew’s Hospital,London ECIA 7BF

Review Article 157
1.
2.
3.
4.
5.
6.
7.
8.
9.
IO.
11.
12.
13.
14.
References
Lindquist S. The heat shock response. Ann Rev Biochem 1986; 55: 1151-91. Neidhardt FC, VanBogelen RA, Vaughn V. The genetics and regulation of heat-shock proteins. Ann Rev Genet 1984; 18: 2955329. Newport G, Culpepper J, Agabian N. Parasite heat-shock proteins. Parasitology Today 1988: 4: 30&12. Engers HD. Letter to the Editor. Results of a World Health Organisation-sponsored work- shop on monoclonal antibodies to Mycobacrer- ium leprae. Infec Immun 1985; 48: 60%5. Engers HD. Letter to the Editor. Results of a World Health Organisation-sponsored work- shop to characterise antigens recognised by Mycobacterium-specific monoclonal antibodies. Infect Immun 1986; 51: 718-20. Young RA, Bloom BR, Grosskinsky CM, Ivanyi J, Thomas D, Davis RW. Dissection of Myco- bacterium tuberculosis antigens using recombi- nant DNA. Proc Nat1 Acad Sci USA 1985; 82: 2583-7. Young RA, Mehra V. Sweetser D er al. Genes for the major protein antigens of Mycobacrerium leprue. Nature 1985; 316: 450-2. Thole JER, Dauwerse HG, Das PK, Groothius DG, Schouls LM, van Embden JDA. Cloning of Mycobacterium bovis BCG DNA and expression of antigens in Escherichiu coli. Infect lmmun 1985; SO: 80&6. Thole JER, Kenlen WJ, Kolk AHJ, Groothius DG, Schouls LM, va Embderi JDA. Charactcri- sation, sequence determination and immunoge- nicity of a 65-kilodalton of Mycobacterium bovis BCG expressed in Escherichia coli K12. Infect Immun 1987; 55: 146675. De Bruyn J, Bosmans R, Turneer M et al. Purification, partial characterisation and identifi- cation of a skin-reactive protein antigen of Mycobacterium bovis BCG. Infect Immun 1987; 55: 245-2. Shinnick TM. The 65-kilodalton antigen of Mycobacterium tuberculosis. J Bacterial 1987; 169: lOE(r8. Mehra V, Sweetser D, Young RA Efficient mapping of protein antigenic determinants. Proc Nat1 Acad Sci USA 1986; 83: 7013-7. Booth RJ, Harris DP, Love JM, Watson JD. Antigenic proteins of Mycobacterium feprue. Complete sequence of the gene of the 18 kDA protein, 1988; 140: 597-601. Nerlund AH, Mustafa AS, Sweetser D, Godal T, .~ Young RA. A protein antigen of Mycobacterium feprae is related to a family of small heat shock proteins. J Bacterial 1988; 170: 5919-21.
15. Young DB. Stress-induced proteins and the human immune response to leprosy. Microbial Sciences 1988; 5: 143-52.
16. Mehlert, A, Young DB. Biochemical and antige- nit characterisation of the Mycobacterium ruber- culosis 71 kDA antigen: a member of the 70 kDA
17.
18.
19.
20.
21.
22.
23.
24.
25.
heat-shock protein family. (submitted for publi- cation). Garsia RJ, Hellquist L, Booth RJ er al. Homo- logy of the 70-kilodalton antigens from Myco- bacterium leprae and Mycobacterium bovi.7 with the Mycobacterium mberculosis 7 I-kD antigen and with the conserved Heat Shock Protein 70 of eucaryotes. Infect Immun 1989; 57: 204-12. Shinnick TM, Vodkin MH. Williams JC. The Mycobacrerium tuberculosis 65-kilodalton anti- gen is a heat-shock protein which corresponds to common antigen and to the Escherichia coli GroEL protein. Infect Immun 1988; 56: 446451. Baird PN, Hall LMC, Coates ARM. A major antigen from Mycobucterium tu6ercufosi.s which is homologous to the heat-shock protein GroES from E. coli and the htpA gene product of Coxiella burnetti. Nucl Acids Res 1988; 16: 9047. Thole JER, Hindersson P, de Bruyn J er (11. Antigenic relatedness of a strongly immunogenic 65 kDa mycobacterial protein antigen with a similarly sized ubiquitous bacterial common antigen. Microbial Pathogenesis 1988; 4: 71-83. Hemmingsen SM. Woodford C, van der Vies SM, Tilly K, Dennis D, Georgopoulos CP. Hen- drix RW, Ellis RJ. Homologous plant and bac- terial proteins chaparone oligomeric protein as- sembly. Nature 1988; 33: 33&l. McMullin TW, Hallberg RL. A highly evolutio- narily conserved mitochondrial protein is struc- turaly related to the protein encoded by the E. coli groEL gene. Mol Cell Biol 1988; 8: 371.-5. Reading DS. Hallberg RL, Myers AM. Charac- terisation of the yeast HSP60 gene coding for a mitochondrial assembly factor. Nature 1989; 337: 655-9. Pannekoek Y, Wee1 JFL, Hopman CTP, van Putten JPM. Gonococcal stress proteins, carriers of “common bacterial antigens”? Abstract of 20th Lunteren Lectures on Molecular Genetics, Lunteren, Netherlands September 1988; 27-30. Morgan RW. Christman MF, Jacobson FS, Storz G, Ames BN. Hydrogen-peroxide-indu- cible proteins in Salmonella typhimurium overlap with heat-shock and other stress proteins. Proc Nat1 Acad Sci USA 1986; 83: 805963.
26. Young DB. Stress-induced proteins and the im- mune response to leprosy. Microbial Sciences 1988;s: 143-51.
27. Allan B, Linseman M, MacDonald LA, Lam JS, Kropinski AM. Heat shock response of P.reu&- mows aeruginosa 1988; 170: 3668-74.
28. Vodkin MH, Williams JC. A heat shock operon in Coxiellu burnefii produces a major antigen . 1 . . nomologous to a protem m both mycobacterta and Escherichiu coli. J Bacterial 1988: 170: 1227. 34.
29. Hinderssen P, Knudsen JD, Axelsen NH. Clon- ing and expression of Treponema pallidurn com- mon antigen (Tp-4) in kcherichia co/i K12. J Gen Microbial 1987: 133: 587-96.
30. Hansen K, Bangsborg JM, Fjordvang H, Peder- sen NS, Hinderssen P. Immunochemical charac-

158 Review Article
terisation of an isolation of the gene for a Borre- lia burgdorferi immunodominant 60-kilodalton antigen common to a wide-range of bacteria. Infect Immun 1988; 56: 3057-3.
31. Ardeshir F, Flint JE, Richman SJ, Reese RT. A 75 kD merozoite surface protein of Plasmodium falciparum which is related to 70 kD heat-shock proteins. Embo Journal 1987; 6: 493-9.
32. Bianco AE, Favaloro JM, Burkot TR et al. A repetitive antigen of Plasmodium falciparum that is homologous to heat shock protein 70 of Droso- phila melanogasrer. Proc Nat1 Acad Sci USA
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44
45
1986; 83: 87137. Scallon BJ, Bogitsh BJ, Carter CE. Cloning of a Schistosoma japonicum gene encoding a major immunogen recognised by hyperinfected rabbits. Anal Biochem Parsitol 1987; 24: 23745. Hedstrom R, Culpepper, J. Harrison RA, Aga- bian N, Newport G. A major immunogen in Schistosoma mansoni infections is homologous to heat-shock protein Hsp-70. J Exp Med 1987; 165: 143&5. Peterson MG, Crewther PE, Thompson JK et al. A second antigenic heat shock protein of Plasmo- diumfalciparum. DNA 1988; 7: 71-8. Ritossa F. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia 1962; 18: 571-3. Cowing DW, Bardwell JC, Craig EA, Woolford C, Hendrix RW, Gross CA. Consensus sequence for Escherichia coli heat shock gene promoters. Proc Nat1 Acad Sci USA 1985; 82: 2679-83. Hellman JD, Chamberlin MJ. Structure and function of bacterial sigma factors. Ann Rev Biochem 1988; 57: 839-72. Straus DB, Walter WA, Gross CA. The heat shock response of E. coli is regulated by changes in the concentration of sigma-32. Nature 1987; 329: 248-52. Jenkins DE, Schultz JE, Martin A. Starvation- induced cross-protection between heat and H202 challenge in Escherichia coli. J Bacterial 1988; 170: 39104. Young DB, Mehlert A, Bal V, Mendez-Samperio P, Ivanyi J, Lamb JR. Stress proteins and the immune response to mycobacteria-antigens as virulence factors? Antonie van Leenwenhoek 1988; 54: 43 1-9. Miller V, Mekalanos JJ. Genetic analysis of the cholera toxin positive regulatory gene ToxR. J Bacterial 1985; 163: 580-5. Stibitz S, Weiss AA, Falkow S. Genetic analysis of a region of the Bordptella pertussis chromo- some encoding HLA and the pleiotropic regula- tory locus vir. J Bacterial 1988; 170: 290413. Polla BS. A role for heat-shock proteins in inflammation. Immunol Today 1988; 9: 134-7. Ellis J. Proteins as molecular chaparones. Nature 1987; 328: 378-9.
46. Pelham H. Heat-shock proteins coming in from the cold. Nature 1988; 332: 7767.
47. Cheng MY, Hart1 F-U, Martin J, Pollock RA, Kalonsek F, Newpert W et al. Mitochondrial
48.
49.
50.
51.
52.
53.
54.
55.
56
57
58
59.
heat-shock protein hsp-60 is essential for as: sembly of proteins imported into yeast mitochon- dria. Nature 1989; 337: 62&l. Bochkareva ES Lissin NM, Girshovich AS. Transient association of newly synthesised infolded proteins with the heatshock GroEL protein. Nature 1988; 336: 254-7. Ljungqvist L, Worsaae A, Heron I. Antibody responses against Mycobacterium tuberculosis in 11 inbred strains of mice: novel monoclonal antibody specificities generated by fusions, using spleens from Balb.BlO and CBA/J mice. Infect Immun 1988; 56: 19948. Coates ARM, Nicolai H, Pallen MJ, Guy A. Chaparas SD, Mitchison DA. The 45 kilodalton molecule of Myrobacrerium tuberculosis identi- fied by immunoblotting and monoclonal anti- bodies as antigens in patients with tuberculosis. Br J Exp Path01 (in press). Vega-Lopez F, Stroker NG, Locniskas MF, Dockrell HM, Grant KA, McAdam KPWJ. Recognition of mycobacterial antigens by sera from patients with leprosy. J Clin Microbial 1988; 26: 24749. Plikaytis BB, Carlone GM, C-P Pau, Wilkinson HW. Purified 60-kilodalton Legionella protein antigen with Legionella-specific and non-specific epitopes. J Clin Microbial 1987; 25: 2080-4. Thole JER, van Schooten WCA, Keulen WJ et al. Use of recombinant antigens expressed in Escherichia coli K-12 to map B-cell and T-cell epitopes on the immunodominant 65-kilodalton protein of Mycobacferium bovis BCG. Infect Immun 1988; 56: 1633-1640. Buchanan TM, Nomaguchi H, Anderson DC ef al. Characterisation of antibody-reactive epi- topes on the 65 kilodalton protein of Mycobac- terium leprae. Infect Immun 1987; 55: 100%3. Anderson DC, Barry MG, Buchanan TM. Exact definition of species-specific and cross-reactive epitopes of the 65-kilodalton protein of Myco- bacrerium leprae using synthetic peptides. J lmmunol 1988; 141: 607-13. Jackett PS, Bothamley GH, Batra HV, Mistry A, Young DB, Ivanyi J. Specificity of antibodies to immunodominant mycobacterial antigens in pul- monary tuberculosis. J Clin Microbial 1988; 26: 2313-8. Britton WJ, Gar.la RJ, Hellquist L, Watson JD, Basten A. The characterisation and immunoreac- tivity of a 70 kD protein common to Mycobacter- ium leprae and Mycobacterium bovis (BCG). Lepr Rev 1986; 57: Suppl 2,67-75. Bardwell JCA, Craig EA. Eukaryotic Mr-83,000 heat shock protein has a homologue in Escheri- chia coli. Proc Nat1 Acad Sci USA 1987; 84: 5177781. Reichardt JKV, Berg P. Conservation of short patches of amino acid sequence amongst proteins with a common function but evolutionarily dis- tinct origins: implications for cloning genes and structure-function analysis. Nucl Acids Res 16: 9017-26.

Review Article 159
60. Van Eden W, Thole JER, van der Zee J et al. ber of the hsp70 family of heat shock proteins in Cloning of the mycobacterial epitope recognized systemic lupus erythematosis. J Exp Med 1988; by T lymphocytes in adjuvant arthritis. Nature 168: 1475-80. 1988; 331: 171-3. 62. Sokolovic Z, Goebe.1 W. Synthesis of listeriolysin
61. Minota S, Cameron B, Welch WJ, Winfield JB. in Listeria monocytogenes under heat shock con- Autoantibodies to the constitutive 73-kD mem- ditions. Infect Immun 1989: 57: 295-98.
(Manuscript accepted 10th January 1989)





























