Embed Size (px)
Citation preview

Vol. 144, No. 1JOURNAL OF BACTERIOLOGY, Oct. 1980, p. 375-3810021-9193/80/10-0375/07$02.00/0
Bacteriophages of Methanotrophic BacteriaF. M. TYUTIKOV,* I. A. BESPALOVA, B. A. REBENTISH, N. N. ALEKSANDRUSHKINA, AND A. S.
KRIVISKY
All- Union Research Institute for Genetics and Selection ofIndustrial Microorganisms; Institute ofMolecular Biology, USSR Academy of Sciences; and Moscow State University, Moscow, USSR
Bacteriophages of methanotrophic bacteria have been found in 16 out of 88studied samples (underground waters, pond water, soil, gas and oil installationwaters, fermentor cultural fluids, bacterial paste, and rumen of cattle) taken indifferent geographic zones of the Soviet Union. Altogether, 23 phage strains wereisolated: 10 strains that specifically lysed only Methylosinus sporium strains, 2strains that each lysed 1 of 5 Methylosinus trichosporium strains studied, and 11strains that lysed Flavobacterium gasotypicum and, at the same time, 1 M.sporium strain. By fine structure, the phages were divided into two types (withvery short or long noncontractile tails); by host range and serological properties,they fell into three types. One-step growth characteristics of the phages differedonly slightly; the latent period varied from 6 to 8 h, the rise period varied from 4to 6 h, and the average burst size was 100. All phages had guanine- and cytosine-rich double-stranded deoxyribonucleic acid consisting of common nitrogen bases.The molecular mass of the deoxyribonucleic acid as determined by restrictionendonuclease analysis was 29.4 x 106 for M. sporium phages and 44 x 106 for F.gasotypicum phages. By all of the above-mentioned properties, all phages withineach of the groups were completely identical to one another, but differed fromphages of other groups. Bacteriophages lysing M. sporium and M. trichosporiumGB2 were identical to phages Ml and M4, respectively, which were isolatedearlier in the German Democratic Republic on the same methanotrophic species.
Considerable importance has been attachedrecently to research into the biology and taxon-omy of methanotrophic bacteria. This includesmorphology and physiology of these microorga-nisms (3, 20), their metabolism and ecology (6,22), possible utilization in prospecting for oil andnatural gas (8), and control of methane accu-mulation in mines (6). Studies aimed at usingthese microorganisms in industrial biosynthesisof protein are also under way (6).The last decade was marked by a wide utili-
zation of bacteria and actinomycetes for indus-trial production ofbiologically active substances,proteins, and nutritional products, and phagesare often a cause of disturbances in the techno-logical process. Production of protein mass byusing methane-oxidizing bacteria has also beenaccompanied by various disturbances.
Since experimental data on specific bacterio-phages of methanotrophic bacteria were notavailable until recently, these disturbances wereseldom associated with bacteriophages.
Several years ago, microbiologists of the In-stitute of Technical Chemistry, German Demo-cratic Republic, with one of us (A.S.K.) detectedand isolated bacteriophages of methanotrophicbacteria (21). More detailed information onthese phages was reported by Tyutikov et al.
(17). These data allowed us to reconsider thedeviations from the normal technological proc-ess observed in the biological production of pro-tein from natural gas and to relate them, in somecases, at least, with the lysis of bacteria byspecific phages.The present study was aiined at the isolation
of new phages specific to various methane-oxi-dizing bacteria, the investigation of their distri-bution in sources of natural and industrial originin the Soviet Union and the systematization ofthe isolated phage strains according to theirultrastructure, plaque morphology, host range,serological properties, one-step growth charac-teristics, and nucleic acid properties.
MATERIALS AND METHODSAll manipulation with phages was performed as
described by Adams (1) and Stent (13).Bacteria. Eighteen strains of the following nine
species of methane-oxidizing bacteria were used in thestudy: Methylosinus sporium, two strains; Methylosi-nus trichosporium, five strains; Flavobacterium gas-otypicum, one strain; Methylocystis impressionis, onestrain; Methylocystisparvus, one strain; Methylocystisfuscus, one strain; Methylococcus capsulatus, threestrains; Methylomonas methanica, three strains; andMethylobacter bovis, one strain. Five strains werereceived from the Institute of Technical Chemistry,
375
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

376 TYUTIKOV ET AL.
GDR Academy of Sciences, Leipzig; five strains werefrom the All-Union Research Institute for Biosyn-thesis of Protein Matter, Moscow; and seven strainswere from the Institute of Biochemistry and Physiol-ogy of Microorganisms, USSR Academy of Sciences,Puschino-na-Oke.Media. Puschino liquid medium (4) contained the
following (in grams per liter of water): KNO3, 1;KH2PO4, 0.75; Na2HPO4.12H20, 4.0; MgS04.7H20,0.4; CaCl2, 0.02. Solid medium contained Puschinoliquid medium supplemented with 1.8 and 0.7% Difcoagar for bottom and top agar layers, respectively. Allexperiments were done in gaseous mixture (1:1 ratioof methane to air), and cultures were incubated at30°C for 4 to 8 days.
Isolation of phages. Phages were isolated fromvarious natural sources without enrichment (see be-low) since most of the samples contained organicmatter which is known to prevent the growth of meth-ane-oxidizing bacteria (8).The phage titer was determined by the agar layer
technique. High-titer phage was obtained by themethod of Swanstrom and Adams (14).Bacteriophage fine structure. The fine structure
was investigated by using negative contrasting underan EM-1OOB electron microscope at 30 kV and mag-nification on the screen of 50,000.Each newly isolated phage was designated with
lower case letters according to the parent cultureseparated by a hyphen from the origin number; e.g.,phage gb4-9 was isolated from sample 9 of cultureGB4.
Antigenic properties of phage. The antigenicproperties were studied by neutralization with rabbitphage antiserum. The following phages were used forobtaining antisera: cm4-9 (2.9 x 109 plaque-formingunits [PFU]), cmfl-11 (1.2 x 1010 PFU), M4 (1 x 1010PFU), and Ml (2 x 109 PFU). Phages Ml and M4were isolated earlier in the German Democratic Re-public (21).The rabbits received nine phage injections. The
first two injections were performed subcutaneously;portions of 0.2 ml of phage with complete Freundadjuvant (Difco) (1:1) were administered into fivedifferent sites, and the injections were separated by a7-day interval. The subsequent seven injections werewithout adjuvant and consisted of 2-ml portions givenat 3-day intervals. The rabbits were bled 10 days afterthe last injection.
This procedure yielded highly active sera. Inacti-vation constants (K) for sera against phages gb4, cm4,cml-11, and gb2 equaled, respectively, 170, 127, 544,and 742. The neutralization reaction with those serawas carried out with phage M4 and the 28 phagestrains isolated in the course of this study.
One-step growth. The classical technique ofAdams (1) was slightly modified since frequentchanges of the gaseous medium while taking the sam-ples resulted in complete suppression of the growth.Instead of adsorption tubes, we used shaker flasks (500ml) with spouts near the bottom which were fittedwith rubber hoses. The flasks with hoses were steri-lized at 1 atm for 30 min. A 1-day culture (in thelogarithmic phase of growth) was infected with phageat a multiplicity of infection of 0.1 (the bacterial titer
was determined in a Goryaev counting chamber undera light microscope), incubated for 30 min (for adsorp-tion) in a water bath at 300C, and diluted in Puschinomedium 104- and 106-fold. Both dilutions (50 ml) weretransferred to sterile shaker flasks; the flasks werefilled with the gaseous medium and incubated for 24h on the shaker. Every 2 h, 0.5-ml samples were takenfrom the rubber hoses by using a sterile syringe witha thin needle, and 0.1-ml portions were seeded onplates containing indicator lawns. The number ofplaques was counted after a 4-day incubation in adesiccator containing the gas mixture. These methodsallowed us to ensure steady growth conditions and toobtain more or less reproducible results.
Isolation of phage DNA. DNase and RNase(Serva; 2 to 5 ,ug/ml) were added to cleared lysateswhich were then left overnight at 40C. After centrifu-gation at 5,500 rpm for 20 min in the cold, the phagewas precipitated by adding 0.5 M NaCl and 10% poly-ethylene glycol (Serva; 6,000 molecular weight) or byhigh-speed centrifugation (Measuring & ScientificEquipment model 25 centrifuge; 24,000 rpm for 2 h).The phage pellet was suspended with a magneticstirrer in 0.05 M Tris-hydrochloride-0.01 M MgCl (pH7.6) and centrifuged for 20 min at 10,000 rpm (Mea-suring & Scientific Equipment centrifuge; 18). Thesupernatant was transferred into centrifuge tubes (38.5ml) of a Beckman SW27 rotor onto a 10-ml layer ofCsCl (1.4 g/ml) and spun for 2.5 to 4 h on a Spinco L5-50 at 24,000 rpm at 200C. The pellet was resuspendedin 2 to 3 ml of Tris-hydrochloride-0.1 M NaCl buffer(pH 7.4) and treated 2 or 3 times with 1 to 2 volumesof water-phenol mixture (88% phenol, 12% water, 0.02volume of 1 M Tris-hydrochloride buffer [pH 7.0], and0.001 volume of 5 M KOH). Phenol was removed bycentrifugation at 5,000 rpm for 20 min followed byextraction with ether or dialysis against 0.lx SSC (lxSSC is 0.15 M NaCl plus 0.015 M sodium citrate).DNA was sedimented by adding 1 volume of coldisopropyl alcohol and stored in 0.lx SSC at 40C. Insome cases, DNA was extracted from phage by alter-nately heating to 65°C and cooling to -70°C. Com-plete hydrolysis ofDNA was carried out in sealed glassampoules at 1000C in 57% perchloric acid for 1 h.Chromatographic separation, elution, and spectropho-tometry of nitrogen bases were performed as describedby Vanyushin (18). Measurements of melting temper-ature and guanine-plus-cytosine-content calculationswere carried out by the method of Marmur and Doty(7).Treatment with restriction endonucleases. The
enzymes employed were EcoRI (isolated by themethod of Greene et al. [5]), BamHI (isolated by themethod of Wilson and Young [19]), SalGI (2), HindIIand HindIII (10), and PstI (12). Cleavage ofDNA withBamHI, HindII, HindIII, and PstI enzymes was per-formed at 370C for 1 h in a mixture (100 to 200 pl)containing 50 mM NaCl, 6 mM Tris-hydrochloride(pH 7.4), 6 mM MgCl, and 6 mM 2-mercaptoethanol.The incubation mixture for EcoRI consisted of 50mMNaCl, 100 mM Tris-hydrochloride (pH 7.5), 5 mMMgCl, and 10 mM 2-mercaptoethanol. Restrictionwith SalGI was carried out in a solution of 100 mMNaCl, 6 mM Tris-hydrochloride (pH 7.9), 10 mMMgCl, and 6mM 2-mercaptoethanol. After incubation,
J. BACTERIOL.
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
BACTERIOIPHAGES OF METHANOTROPHIC BACTERIA 377
the samples were heated for 5 to 7 min at 65°C, rapidlycooled to 0°C, and analyzed in 1% agarose gels (11).
Analysis of endonuclease restriction pattern.Analysis of the endonuclease restriction pattern wasdone by using electrophoretograms of the gels. Thegels were photographed in UV light with VCh film,and the electrophoretic mobility of each fragment wasmeasured on an ACM-2M compactor (USSR) or aServa laser scanner. The molecular weights of frag-ments were determined by using as markers restric-tions of phage DNA with known molecular weight(15).
RESULTSDistribution of phages. The samples con-
sisted of soil and water taken in different geo-graphic and climatic zones of the USSR, tripe ofcattle, protein paste of methane-oxidizing bac-teria, and cultural fluid of pilot fermentor in-stallations.From 88 samples, we isolated 30 phage strains
that lysed M. sporium, M. trichosporium, and F.gasotypicum species and no phages that wereactive on Methylococcus, Methylomonas, orMethylocystis genera. It should be noted that allisolated phages were apparently present at highconcentration in the samples; the enrichmenttechnique is unfortunately not applicable to suchstrict autotrophs as methane-oxidizing bacteria(6).Phages of methanotrophic bacteria are rather
widely spread in the environment. They werefound in two cattle rumen samples out of sixstudied. The samples were taken from differentclimatic and geographic zones of the Soviet Un-ion. This fact indicates a rather high probabilityof encountering them in the production of pro-tein from natural gas. The phages were found in34% of the samples investigated, whereas in thepaste of methane-oxidizing bacteria and in fer-mentor cultural fluid, they were present in 60%of the cases. In water samples taken from gasand oil installations, the phages were found in100% of the cases, which is due to the fact thatthis water is an ecological niche for methano-trophic bacteria (6). Our failure to detect phagesactive on Methylococcus, Methylomonas, andMethylocystis bacteria does not indicate theirabsence in the environment since we had only afew indicator strains from these genera. Besides,many strains form poor lawns on solid media,thus decreasing the probability of detectingphages specific to them.Ultrastructures of the phages. Phages of
28 isolated strains were studied under the elec-tron microscope. Two phages, gb2-80 and cmfl-80, isolated from sample 80, were not examinedby electron microscope analysis. All of thephages fell into the two morphological groups
described earlier (17). The first group consistedof phages with short tails. These phages lysedM. sporium and M. trichosporium. Phages ofthe second group had heads with long tails andlysed F. gasotypicum.The first phage group had short tails ending
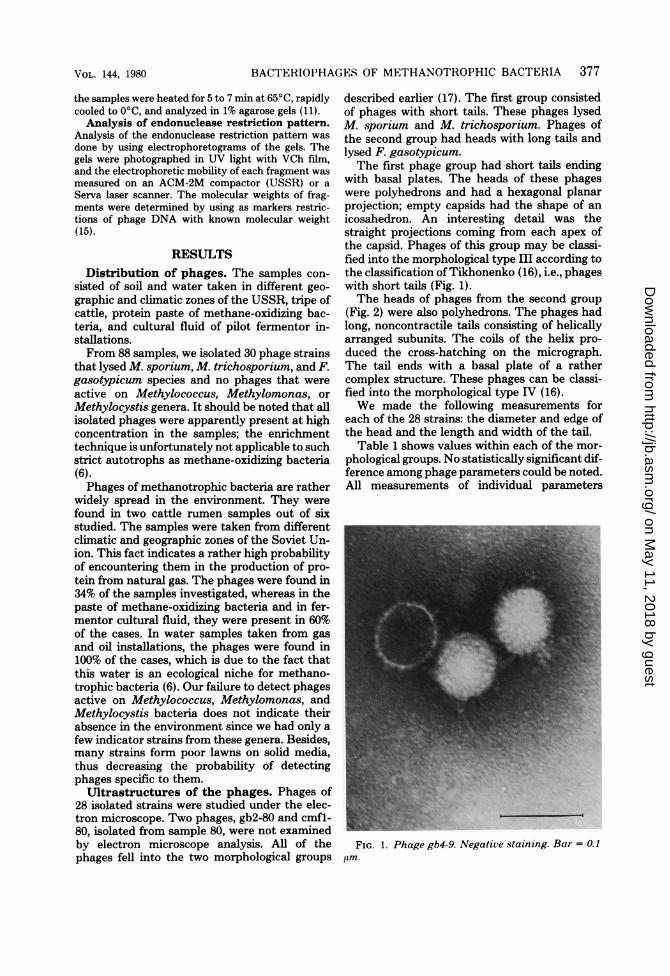
with basal plates. The heads of these phageswere polyhedrons and had a hexagonal planarprojection; empty capsids had the shape of anicosahedron. An interesting detail was thestraight projections coming from each apex ofthe capsid. Phages of this group may be classi-fied into the morphological type III according tothe classification of Tikhonenko (16), i.e., phageswith short tails (Fig. 1).The heads of phages from the second group
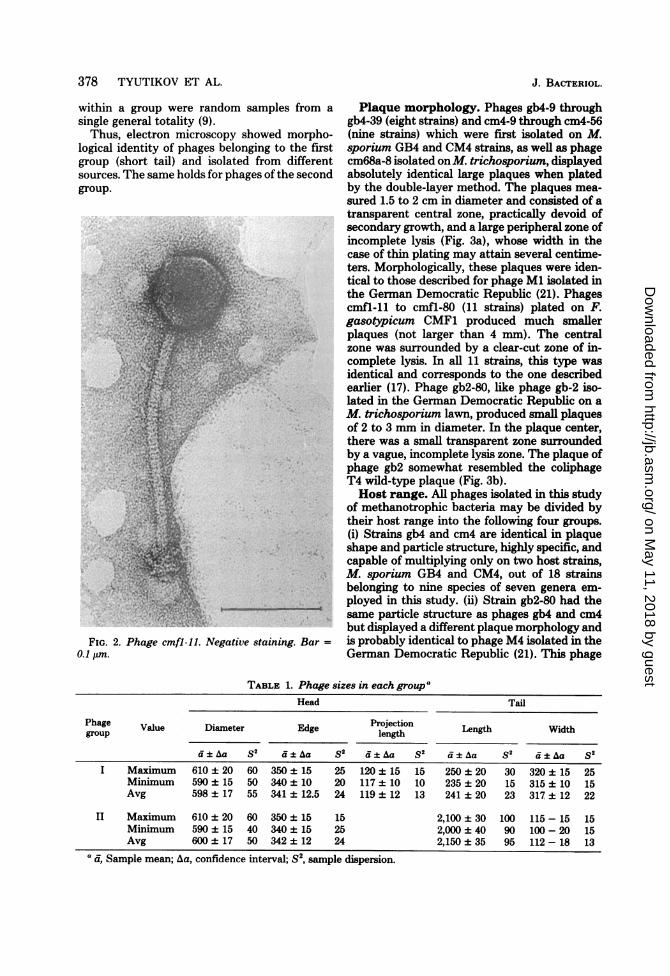
(Fig. 2) were also polyhedrons. The phages hadlong, noncontractile tails consisting of helicallyarranged subunits. The coils of the helix pro-duced the cross-hatching on the micrograph.The tail ends with a basal plate of a rathercomplex structure. These phages can be classi-fied into the morphological type IV (16).We made the following measurements for
each of the 28 strains: the diameter and edge ofthe head and the length and width of the tail.
Table 1 shows values within each of the mor-phological groups. No statistically significant dif-ference among phage parameters could be noted.All measurements of individual parameters
_ A~~~~~~~~~~It I. _
FIG. 1. Phage gb4-9. Negative staining. Bar = 0.1,Am.
VOL. 144, 1980
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
378 TYUTIKOV ET AL.
within a group were random samplsingle general totality (9).Thus, electron microscopy showed
logical identity of phages belonging tigroup (short tail) and isolated fromsources. The same holds for phages of tgroup.
FIG. 2. Phage cmfl-l1. Negative staini0.1 ,um.
es from a Plaque morphology. Phages gb4-9 throughgb4-39 (eight strains) and cm4-9 through cm4-56
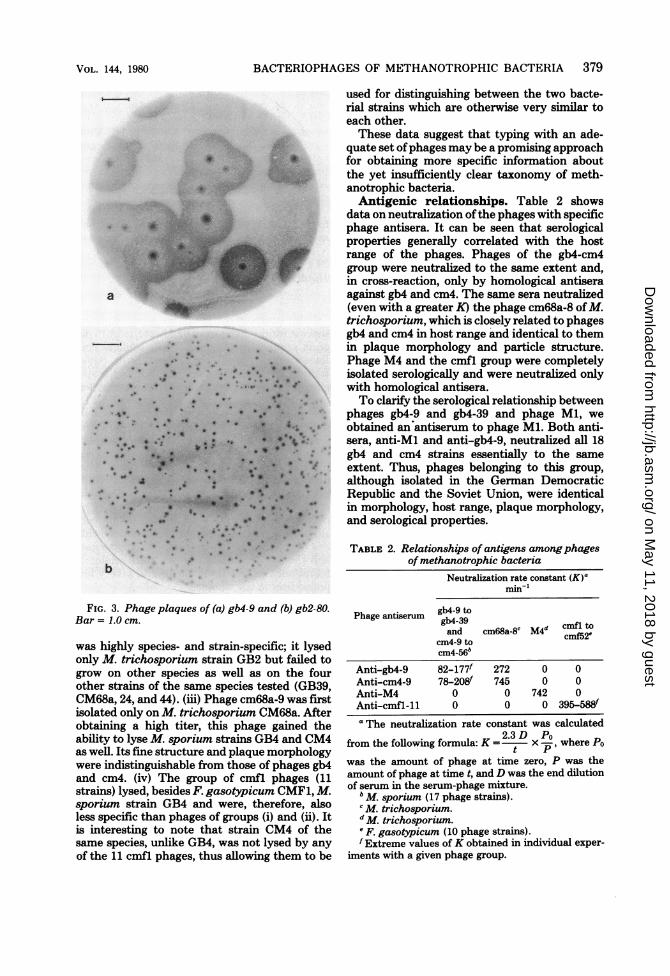
morpho- (nine strains) which were first isolated on M.O the first sporium GB4 and CM4 strains, as well as phagedifferent cm68a-8 isolated onM. trichosporium, displayed
the second absolutely identical large plaques when platedby the double-layer method. The plaques mea-sured 1.5 to 2 cm in diameter and consisted of atransparent central zone, practically devoid ofsecondary growth, and a large peripheral zone ofincomplete lysis (Fig. 3a), whose width in thecase of thin plating may attain several centime-ters. Morphologically, these plaques were iden-tical to those described for phage Ml isolated inthe German Democratic Republic (21). Phagescmfl-11 to cmfl-80 (11 strains) plated on F.gasotypicum CMF1 produced much smallerplaques (not larger than 4 mm). The centralzone was surrounded by a clear-cut zone of in-complete lysis. In all 11 strains, this type wasidentical and corresponds to the one describedearlier (17). Phage gb2-80, like phage gb-2 iso-lated in the Gernan Democratic Republic on aM. trichosporium lawn, produced small plaquesof 2 to 3 mm in diameter. In the plaque center,there was a small transparent zone surroundedby a vague, incomplete lysis zone. The plaque ofphage gb2 somewhat resembled the coliphageT4 wild-type plaque (Fig. 3b).Host range. All phages isolated in this study
of methanotrophic bacteria may be divided bytheir host range into the following four groups.(i) Strains gb4 and cm4 are identical in plaqueshape and particle structure, highly specific, andcapable of multiplying only on two host strains,M. sporium GB4 and CM4, out of 18 strainsbelonging to nine species of seven genera em-ployed in this study. (ii) Strain gb2-80 had thesame particle structure as phages gb4 and cm4but displayed a different plaque morphology and
cng. Bar = is probably identical to phage M4 isolated in theGerman Democratic Republic (21). This phage
TABLE 1. Phage sizes in each groupaHead Tail
Phage Value Diameter Edge Projection Length Width
dI±Aa s2 d+±Aa S2 d±Aa s2 ci±a S2 d±Aa S2
I Maximum 610 ± 20 60 350 ± 15 25 120 ± 15 15 250 ± 20 30 320 ± 15 25Miniimum 590 ± 15 50 340 ± 10 20 117 ± 10 10 235 ± 20 15 315 ± 10 15Avg 598 ± 17 55 341 ± 12.5 24 119 ± 12 13 241 ± 20 23 317 ± 12 22
II Maximum 610 ± 20 60 350 ± 15 15 2,100 ± 30 100 115-15 15Minimum 590 ± 15 40 340 ± 15 25 2,000 ± 40 90 100-20 15Avg 600 ± 17 50 342 ± 12 24 2,150 ± 35 95 112-18 13
a a, Sample mean; Aa, confidence interval; S2, sample dispersion.
J. BACTERIOL.
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
BACTERIOPHAGES OF METHANOTROPHIC BACTERIA 379
.
a
0 ~~** .e;
g* X
*
* gjf i* S r .. '> 0 ' -
40*
* #
~ ~
. X9
9.S
b
FIG. 3. Phage plaques of (a) gb4-9 and (b) gb2-80.Bar = 1.0 cm.
was highly species- and strain-specific; it lysedonly M. trichosporium strain GB2 but failed togrow on other species as well as on the fourother strains of the same species tested (GB39,CM68a, 24, and 44). (iii) Phage cm68a-9 was firstisolated only on M. trichosporium CM68a. Afterobtaining a high titer, this phage gained theability to lyse M. sporium strains GB4 and CM4as well. Its fine structure and plaque morphologywere indistinguishable from those of phages gb4and cm4. (iv) The group of cmfl phages (11strains) lysed, besides F. gasotypicum CMF1, M.sporium strain GB4 and were, therefore, alsoless specific than phages of groups (i) and (ii). Itis interesting to note that strain CM4 of thesame species, unlike GB4, was not lysed by anyof the 11 cmfl phages, thus allowing them to be
used for distinguishing between the two bacte-rial strains which are otherwise very similar toeach other.These data suggest that typing with an ade-
quate set ofphages may be a promising approachfor obtaining more specific information aboutthe yet insufficiently clear taxonomy of meth-anotrophic bacteria.Antigenic relationships. Table 2 shows
data on neutralization ofthe phages with specificphage antisera. It can be seen that serologicalproperties generally correlated with the hostrange of the phages. Phages of the gb4-cm4group were neutralized to the same extent and,in cross-reaction, only by homological antiseraagainst gb4 and cm4. The same sera neutralized(even with a greater K) the phage cm68a-8 ofM.trichosporium, which is closely related to phagesgb4 and cm4 in host range and identical to themin plaque morphology and particle structure.Phage M4 and the cmfl group were completelyisolated serologically and were neutralized onlywith homological antisera.To clarify the serological relationship between
phages gb4-9 and gb4-39 and phage Ml, we
obtained an antiserum to phage Ml. Both anti-sera, anti-Mi and anti-gb4-9, neutralized all 18gb4 and cm4 strains essentially to the same
extent. Thus, phages belonging to this group,although isolated in the German DemocraticRepublic and the Soviet Union, were identicalin morphology, host range, plaque morphology,and serological properties.
TABLE 2. Relationships of antigens among phagesof methanotrophic bacteria
Neutralization rate constant (K)amin-'
Phage antiserum gb4-39and cm68a-8" M4d cmfl2tcm4-9 to cf2
cm4-56b
Anti-gb4-9 82-177f 272 0 0Anti-cm4-9 78-208f 745 0 0Anti-M4 0 0 742 0Anti-cmfl-11 0 0 0 395-588fa The neutralization rate constant was calculated
2.3D Pofrom the following formula: K = x -, where Powas the amount of phage at time zero, P was theamount of phage at time t, and D was the end dilutionof serum in the serum-phage mixture.
b M. sporium (17 phage strains).'M. trichosporium.d M. trichosporium.'F. gasotypicum (10 phage strains).f Extreme values ofK obtained in individual exper-
iments with a given phage group.
VOL. 144, 1980
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

380 TYUTIKOV ET AL.
One-step growth. An essential requirementfor reproducible results on one-step growthcurves ofphages ofmethanotrophic bacteria wasthe constancy of the gaseous medium. Changesin the medium caused by frequent opening ofadsorption flasks resulted in complete suppres-sion of the growth. Shaking of the flasks wasalso found to be necessary for phage develop-ment.The differences among phages of different
groups were very slight. All the phages werecharacterized by a latent period of many hoursand a rather small burst size. Similar results onthe latent period and burst size were obtainedby Werner and Wunsche (Abstr. Int. Symp.Acad. Sci. USSR Sci. Cent. Biol. Res., p. 174,1977) for phage Ml of M. sporium, which isidentical to phages gb4 and cm4.Phage nucleic acid. Phages Ml and cm4 of
M. sporium, M4 of M. trichosporium, and cmflof F. gasotypicum were investigated. Alkalinehydrolysis (0.75 M NaOH) and treatment withpancreatic RNase under standard conditions(37°C for 1 h) did not cause an increase in UVadsorption, indicating that the nucleic acid wasDNA. This was confirmed by a positive Dischereaction on deoxyribose. Separation of acid hy-drolysis (57% HC104 for 1 h at 1000C) productsby thin-layer chromatography on cellulose (Fil-trak) showed the presence of the four commonbases (Table 3). The typical melting curve pro-file for double-stranded DNA and the UV ad-sorption spectra were also typical of DNA ofusual composition. The guanine-plus-cytosinecontent varied from 58.6 to 64.1 mol%.Restriction endonuclease analysis of
phage DNA. Restriction endonuclease treat-ment was applied to the DNA of nine phages ofthe gb4-cm4 group (gb4-9, gb4-12, gb4-14, gb4-39, cm4-11, cm4-12, cm4-21, cm4-39, and cm4-56) lysing M. sporium and eight phages growingon F. gasotypicum (cmfl-11, cmfl-12, cmfl-14,cmfl-21, cmfl-27, cmfl-39, cmfl-50, and cmfl-56). All phages within each of the groups werecleaved absolutely identically by all restrictionenzymes used. EcoRI and PstI were inactive in
TABLE 3. Properties ofphage nucleic acidsMelt- DNA composition (mol%)
Phage ingstrain temp Ade- Gua- Cyto- Thy-
(0C) nine nine sine mine
Mla 94.6 19.6 30.35 30.35 19.6cm4-9a 94.5 19.0 31.0 31.0 19.0M4b 93.2 21.0 29.3 20.3 21.0cmfl-llc 95.6 17.5 32.0 32.0 17.6
a M. sporium.b M. trichosporium.F. gasotypicum.
the gb4-cm4 group and produced 4 and over 15fragments, respectively, after digestion of cmflDNA. SalGI and HindII gave a large number(over 30) of small fragments of cmfl DNA and3 and over 15 fragments, respectively, ofthe gb4-cm4 group DNA. BamHI yielded five differentfragments in both cases, whereas treatment withHindIII produced three or seven fragments ofthe gb4-cm4 or cmfl DNA, respectively.
Estimate of the molecular mass of cmfl DNAfrom the sum of fragment lengths after treat-ment with EcoRI, BamHI, and Hindm was 44.1x 106. Phages of the gb4-cm4 group had DNA of29.4 x 106 daltons (measured by BamHI andHindIII fragments).
DISCUSSIONIn the course of this study, we detected phages
specific for methane-oxidizing bacteria in 16 outof 88 samples investigated. Some samples con-tained phages specific for different hosts. Sevensamples which were taken mostly from the spe-cific habitat of methanotrophic bacteria (fer-mentor cultural fluid, water of gas and oil in-stallations, and rumen of cattle) contained twodifferent phages. In seven samples, plaques werefound after primary plating on lawns of bothstrains GB4 and CM4, and in three samples,plaques were found only on one of them. Afterpropagation to high titers, all 10 strains couldgrow equally well on both host strains. Since gb4phages were identical to the cm4 group by allcharacteristics so far studied, it is to be con-cluded that each ofthese samples contained onlyone strain capable of growing on M. sporium,and the fact that some of them were ofiginallyfound only on one lawn was due to a very lowphage titer in the samples.The total number of phage strains isolated
from 88 samples thus equals 23: 10 strains of thegb4-cm4 type which lysed only M. sporium, 11cmfl strains growing on F. gasotypicum and on1 of the M. sporium strains, and 2 strains whichlysed-only 1 of 5 M. trichosporium strains. Nophages were found to be able to grow on the sixother species of methanotrophic bacteria inves-tigated. This was probably due to the low con-centration of phages in the samples and thefailure to apply the enrichment technique (theculture stops growth because of high organicmatter content).The above evidence suggests that phages of
the gb4-cm4 and cmfl groups are rather widelyspread in the Soviet Union and may cause con-siderable danger for industrial production ofpro-tein from natural gas under continuous and non-sterile conditions. Essentially, these two groupsrepresent individual strains inhabiting differentecological niches and geographic regions. A good
J. BACTERIOL.
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

BACTERIOPHAGES OF METHANOTROPHIC BACTERIA 381
example is provided by phages gb4 and cm4,isolated in the Soviet Union, and phage Ml,isolated in the German Democratic Republic.The number of types of phages lysing metha-notrophic bacteria in the environment appearsto be limited. Thus, it would be useful to obtainmutants ofindustrial cultures ofmethanotrophicbacteria that are resistant to most phages.
LITERATURE CITED1. Adams, M. N. 1959. Bacteriophages. Wiley, Interscience,
New York.2. Arrand, J. K., P. A. Myers, and R. J. Roberts. 1978. A
new restriction endonuclease from Streptomyces albus.J. Mol. Biol. 118:127-135.
3. Foster, J. W., and R. H. Davis. 1966. A methane-de-pendent coccus with notes on classification and nomen-clature of obligate methane-utilizing bacteria. J. Bac-teriol. 91:1924-1931.
4. Galchenko, B. N. Shishkina, V. S. Tyurin, and Y. A.Protsenko. 1975. Isolation of pure cultures of meth-anotrophic bacteria and their properties. Mikrobiologia44:844-850. (In Russian.)
5. Greene, P. J., M. G. Betlach, and H. W. Boyer. 1974.The EcoRI restriction endonuclease. Methods Mol.Biol. 7:87-111.
6. Krasnikov, E. J., Y. R. Malashenko, and V. L. Ro-manovskaya. 1974. The biology of microorganismsthat assimilate gaseous hydrocarbons. Prog. Microbiol.2:125-152. (In Russian.)
7. Marmur, J., and P. Doty. 1962. Determination of thebase composition of deoxyribonucleic acid from its ther-mal denaturation temperature. J. Mol. Biol. 5:109.
8. Mogilevsky, G. A. 1970. A bacterial technique for pros-
pecting oil and natural gases. Razved. Nedr 12:315-319.(In Russian.)
9. Nalimov, V. V. 1971. The theory of experiment, p. 62.Nauka Publishers, Moscow.
10. Old, R., K. Murray, and C. Kaizes. 1975. Recognitionsequence of restriction endonuclease III from Hemoph-
ilus influenzae. J. Mol. Biol. 92:331-339.11. Sharp, P. A., B. Sugden, and K. J. Sambrook. 1973.
Detection of two restriction endonuclease activities inHaemophilus parainfluenzae using analytical agarose-ethidium bromide electrophoresis. Biochemistry 12:3055-3063.
12. Smith, D. L., F. K. Blattner, and J. Davies. 1976. Theisolation and partial characterisation of a new restric-tion endonuclease from Providencia stuartii. NucleicAcids Res. 3:345-353.
13. Stent, G. S. 1963. Molecular biology of bacterial viruses.W.H. Freeman & Co., San Francisco, Calif.
14. Swanstrom, M., and M. H. Adams. 1951. Agar layermethod for production of high-titer phage stocks. Proc.Soc. Exp. Biol. Med. 78:372.
15. Thomas, M., and R. W. Davis. 1975. Studies on thecleavage of bacteriophage lambda DNA with EcoRIrestriction endonuclease. J. Mol. Biol. 91:315-328.
16. Tikhonenko, A. S. 1968. The ultrastructure of bacterialviruses (In Russian). Nauka Publishers, Moscow.
17. Tyutikov, F. M., N. N. Belyaevf, L. C. Smirnova, A.S. Tikhonenko, and A. S. Krivisky. 1976. Isolationof bacteriophages of methane-oxidizing bacteria. Mik-robiologia 6:1056-1062. (In Russian.)
18. Vanyushin, B. F. 1964. The determination of nucleic acidbase composition, p. 236. In V. N. Orekhovitch (ed.),Modern methods in biochemistry (In Russian). Medit-sina Publishers, Moscow.
19. Wilson, G. A., and F. E. Young. 1975. Isolation ofsequence specific endonuclease (BamI) from Bacillusamyloliquefaciens H. J. Mol. Biol. 97:123-125.
20. Wittenbury, R., K. C. Phillips, and J. F. Wilkinson.1970. Enrichment, isolation and some properties ofmethane-utilizing bacteria. J. Gen. Microbiol. 61:205-218.
21. Wunsche, L., B. Werner, H. Hauche, W. J. Kadel-mann, U. Hilger, A. S. Krivisky, W. W. Jessipowa,A. S. Tikhonenko, and I. Zimmermann. 1977. Na-chweis und erste Characterisierung von Bakteriophagenobligat methanassimilierender Bakterien. Z. AUg. Mik-robiol. 17:321-323.
22. Zinger, A. C. 1966. The microflora ofunderground waters.Mikrobiologia 35:357-360. (In Russian.)
VOL. 144, 1980
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from





























