Embed Size (px)
Citation preview

ORIGINAL PAPER
Behavioral responses to echolocation calls from sympatricheterospecific bats: implications for interspecific competition
Yuanyuan Li & Jing Wang & Walter Metzner & Bo Luo &
Tinglei Jiang & Siliu Yang & Limin Shi & Xiaobin Huang &
Xinke Yue & Jiang Feng
Received: 9 April 2013 /Revised: 23 December 2013 /Accepted: 23 December 2013# Springer-Verlag Berlin Heidelberg 2014
Abstract Mutual recognition is the product of species coex-istence, and has direct effects on survival and reproduction ofanimals. Bats are able to discriminate between sympatricdifferent heterospecifics based on their echolocation calls,which has been shown both in free-flying and captive bats.To date, however, the factors that may determine the behav-ioral responses of bats to echolocation calls from sympatricheterospecifics have rarely been tested, especially under well-controlled conditions in captive bats. Hence, we aimed attackling this question by performing playback experiments(habituation–dishabituation) with three horseshoe bat specieswithin the constant-frequency bat guild, which included big-eared horseshoe bats (Rhinolophus macrotis), Blyth’s horse-shoe bats (Rhinolophus lepidus), and Chinese horseshoe bats(Rhinolophus sinicus). We studied the behavioral responses ofthese three species to echolocation calls of conspecifics, to
other two species, and to another heterospecifics bat,Stoliczka’s trident bat (Asellisus stoliczkanus), which alsobelongs to this guild. We found that the three rhinolophidspecies displayed a series of distinct behaviors to heterospecificecholocation but few to conspecific calls after habituation,suggesting that they may have been able to discriminate sym-patric heterospecific echolocation calls from those of conspe-cifics. Interestingly, the behavioral responses to heterospecificcalls were positively correlated with the interspecific overlapindex in trophic niche, whereas call design had only a minoreffect. This implies that the behavioral responses of these bats toheterospecific echolocation calls may be related to the degree ofinterspecific food competition.
Keywords Sympatric bats . Echolocation calls . Speciesrecognition . Behavioral responses . Interspecific competition
Introduction
In complex animal communities, mutual recognition is aprerequisite for interspecific interactions (Bee 2006;Qvarnström et al. 2006). Mutual recognition between sympat-ric competitors is also beneficial (Cody 1969, 1973). A centralproblem in understanding heterospecific interactions in suchcomplex animal communities is to what degree interspecificresource competition and the design of communication signalsdetermines behavioral responses of animals to sympatricheterospecifics. Competitors at the same trophic level oftenrequire similar resources and need to avoid similar predators,so information transfer based on mutual recognition betweenthese species may be particularly valuable (reviewed inGoodale et al. 2010). For example, nine-spined sticklebacks(Pungitius pungitius) find appropriate foraging patches bymonitoring the success of three-spined sticklebacks(Gasterosteus aculeatus) (Coolen et al. 2003), or redwings
Communicated by C. Voigt
Y. Li : J. Wang : B. Luo : T. Jiang : S. Yang :X. Huang :X. Yue :J. FengJilin Key Laboratory of Animal Resource Conservation andUtilization, Northeast Normal University, Changchun, Jilin,People’s Republic of China
Y. Li : J. Wang : B. Luo : T. Jiang : S. Yang :X. Huang :X. Yue :J. Feng (*)Key Laboratory for Wetland Ecology and Vegetation Restoration ofNational Environmental Protection, Northeast Normal University,Changchun, Jilin, People’s Republic of Chinae-mail: [email protected]
W. Metzner (*)Department of Integrative Biology and Physiology, UCLA,Los Angeles, CA 90095-1606, USAe-mail: [email protected]
L. ShiCollege of Life Science, Yunnan Normal University, Kunming,Yunnan, People’s Republic of China
Behav Ecol SociobiolDOI 10.1007/s00265-013-1680-9

(Turdus iliacus) can own a profitable breeding patch depend-ing on the abundance of resident tit species (Forsman et al.1998). Another beneficial effect of mutual recognition be-tween sympatric competitors is that it enables these animalsto coexist by lowering the strength of interspecific competi-tion (reviewed in Grether et al. 2009) or interspecific aggres-sion (Korner et al. 2000).
Fight between competitors is costly both in time and energyinvested, and it increases the risk of predation. Consequently,natural selection should favor mutual recognition betweencompetitors, and aggressive interactions should depend uponthe degree of competitive overlap (Moore 1978). Listening toacoustic signals produced by other animals represents animportant way of species recognition in amphibians (e.g.,Ryan 1983), birds (e.g., Lynch and Baker 1991), and mam-mals (e.g., Schuchmann and Siemers 2010). In birds, similarcall design can facilitate interspecific recognition and influ-ence the behavioral responses of birds (Marler 1957; Hurd1996). For example, swamp and song sparrows (Melospizaspp.) respond to each other’s distress calls, which are alike inlength, carrier frequency, and frequency range, but not to thoseof white-throated sparrows (Zonotrichia), which are very dif-ferent (Stefanski and Falls 1972).
Interspecific competition between sympatric bat speciescan be weakened by the partitioning of diets (Arlettaz et al.1997), feeding grounds (Campbell et al. 2007), or roosts(Jacobs and Barclay 2009). However, within most bat com-munities, interspecific competition due to overlapping re-quirements and interactions is still high (e.g., Schoeman andJacobs 2011). Interspecific recognition may therefore be nec-essary to adjust the strength of interspecific interactions. Inbats, both social and echolocation calls can be used for socialcommunication (reviewed by Fenton 2003; Jones and Siemers2011), in spite of echolocation calls havingmainly evolved forthe purpose of orientation and foraging (e.g., Griffin 1958;Griffin et al. 1960). Bats can recognize sympatricheterospecifics based on their echolocation calls, which hasbeen confirmed both in free-flying (Barclay 1982; Übernickelet al. 2013; Dorado-Correa et al. 2013) and captive bats(Schuchmann and Siemers 2010; Voigt-Heucke et al. 2010).However, to date, the factors which may affect behavioralresponses of bats to echolocation calls from sympatricheterospecifics have rarely been tested, especially in captivebats under well-controlled conditions. To the best of ourknowledge, only one study tested this question usingcaptive bats (Noctilio albiventris) and showed that thedegree of social interactions may affect a bat’s response(Voigt-Heucke et al. 2010).
Studies on free-flying bats have demonstrated that thebehavioral responses of bats eavesdropping on the echoloca-tion calls of another bat species mainly depends on the simi-larity in echolocation call design (call structure, duration, andbandwidth) (e.g., Myotis yumanensis and Lasiurus borealis;
Balcombe and Fenton 1988). Additionally, both overlappingdiets (e.g., Myotis lucifugus; Barclay 1982) and similar hunt-ing ecology (Übernickel et al. 2013) can affect a bat’s responseto sympatric heterospecifics. Within a bat guild, similar for-aging ecology and overlapping diets are likely to intensifyinterspecific food competition. The trophic niche overlapindex has been used to determine the degree of overlapin food utilization (Schoener 1968) between two sym-patric species during the same period. This index canalso determine the relative amounts of interspecific foodcompetition (Abrams 1980).
In the present study, we therefore aimed at exploring towhat degree the similarity of call design and the interspecificfood competition affected the behavioral responses of bats toecholocation calls from sympatric heterospecifics. We carriedout playback experiments (habituation–dishabituation) withcaptive bats of the constant-frequency (CF) bat guild includ-ing three rhinolophid species [big-eared horseshoe bats(Rhinolophus macrotis), Blyth’s horseshoe bats (Rhinolophuslepidus), and Chinese horseshoe bats (Rhinolophus sinicus)]and one hipposiderid species, the Stoliczka’s trident bat(Asellia stoliczkanus). Only individuals of the threerhinolophid species were used in playback experiments. Al-though the echolocation calls of rhinolophid bats are generallyvery similar in their composition (a long CF componentinitiated and terminated by brief FM components), they differin their peak frequency, duration, and bandwidth among spe-cies. Thus, the interspecific differences in echolocation-callcharacteristics (peak frequency, duration, and bandwidth) canreflect the difference of call design between species. The callsof A. stoliczkanus are usually much shorter than rhinolophidcalls and lack an initial upward FM component. All the fourspecies we studied usually forage close to or within vegetation(Schnitzler and Kalko 2001), and moths make up a largecomponent of their diets. Consequently, interspecific foodcompetition (this paper mainly refers to interspecific trophicniche overlap index) may be intense depending on the degreeof overlap in their diets.
We hypothesized that the three horseshoe bat species areable to discriminate echolocation calls of heterospecifics fromthose of conspecifics. We also predicted that the intensity oftheir responses differed to echolocation calls from differentsympatric heterospecifics, and that bats displayed the weakestresponse to echolocation calls of A. stoliczkanus. Specifically,we tested two hypotheses: (1) the behavioral responses of bats(i.e., the relative number of each behavior analyzed—see“Materials and methods”—to repeated playbacks ofheterospecific echolocation calls) are negatively correlat-ed with heterospecific differences in echolocation-callcharacteristics (peak frequency, duration, and band-width); and (2) the behavioral responses of bats arepositively correlated with the heterospecific overlap in-dex in trophic niche.
Behav Ecol Sociobiol

Materials and methods
Study area and bats
All experiments were approved by the Department ofWildlifeAdministration in Shuanghe Township, Yunnan province,China (24°30′N, 102°20′E), where the colonies of the batsused in this experiment were located. We conducted our workfrom July to September 2012. The species used in our studywere R. macrotis (colony size, N=80–100), R. lepidus(N=50–60), R. sinicus (N>200), and A. stoliczkanus (N<50).The former three species belong to the Rhinolophidae andwere used for both recording their echolocation calls and forthe playback experiments. The fourth species belongs to theHipposideridae and was only used for recording and subse-quent playback to rhinolophid bats. R. macrotis roosted mostlyin Xianren cave, while the other three species roosted inLongxu cave. Xianren and Longxu caves are located south-west of Shuanghe Township roughly 500 m apart with avertical distance of less than 150 m.
Collection and husbandry of bats
Bats were captured at the cave entrances with mist or hand netsimmediately after sunset, and sex, age (Brunet-Rossinni andWilkinson 2009), and reproductive status were determined(Racey 2009). A total of 17 adult bats (12 ♂, 5 non-lactating♀) were caught for each species except forA. stoliczkanus (only5 ♂ and 5 non-lactating ♀). All captured bats were maintainedin cages (200 cm×140 cm×50 cm) for a maximum duration of10 days. Each cage only contained individuals of the samespecies. Cages were set up in different rooms of the laboratorybuilding to avoid interspecific communication. Room temper-ature and relative humidity were maintained similar to thosefound in the caves (temperature 21–23 °C, relative humidity68–72 %). We adjusted the illumination according to the localphotoperiod. Bats were fed mealworms and given water beforethe sound recordings and experiments to keep them motivated.At all other times, bats had free access to mealworms and water.After the conclusion of the experiments, all bats were releasedat their respective site of capture.
Calls recording and generation of playback files
Following the approach used by Schuchmann and Siemers(2010), we tested the behavioral responses of threerhinolophid species to heterospecific calls. Echolocation callsof all four species (ten individuals were recorded per species, 5♂, 5 ♀) were recorded. Bats were placed in the experimentalbox individually (Fig. 1). When the bat was hanging from theperch and was motionless, we recorded its echolocation callsusing a condenser ul t rasound microphone (typeCM16/CMPA; Avisoft Bioacoustics, Berlin, Germany). The
microphone was facing directly towards the perch and wasconnected to an ul t rasound recording inter face(UltraSoundGate 116; Avisoft Bioacoustics) with a samplingrate of 375 kHz at 16 bits/sample. For each bat, two soundfiles, each 60 s long, were recorded constantly.
For each bat, we selected two series of calls, each 10 s long,that had a signal-to-noise ratio (SNR) better than 30 dB toobtain the sound files that we then used as playback signals.The call series were further divided into shorter segments withdurations ranging from 4 to 1,000 ms. Five playback files,each 20 s long, were then created for each species throughrandom combination of these short call segments usingAvisoft-SASLab Pro 4.3. Each of the playback files includedecholocation pulses from random four (2♂, 2♀) individuals often recorded bats, and at least one of the four individuals isdifferent between any two playback files since we wanted batsto classify the playback files by species, not by individuals orsex. For each file, the number of echolocation pulses from everybat was random and not less than 3. The length of the intervalsbetween echolocation pulses was not standardized. The call ratein the playback files was approximately 15 calls/s (Schuchmannand Siemers 2010). We normalized the playback sounds so thatthe peak amplitude of the weakest call was approximately−30 dB. Each playback file was high-passed at 20 kHz.
Playback experiments
Playbacks of echolocation calls from the three horseshoe batsspecies and from A. stoliczkanus were only presented tohorseshoe bats (mostly because we were only able to catch
computer
UltraSoundGate Player 116
UltraSoundGate 116
bat Sony HDR PJ760E
soundabsorbing foam box
40cm
60cm
80cm
speaker
microphone
Fig. 1 Experimental box (lined with sound-absorbing foam). Awoodenbasket served as a perch for the bats and was installed on one short side ofthe box. An infrared digital video camera (Sony Handycam HDR-PJ760E, Japan) monitored the bat’s behavior and a speaker (UltrasonicDynamic Speaker Vifa; Avisoft Bioacoustics, Germany) directed straighttowards the perch was used to deliver the playback signals, and acondenser ultrasonic microphone picked up the calls emitted by the testbat. In order to avoid recording of the playbacks, the microphone wasplaced on the right side of the loudspeaker. Adapted from Schuchmannand Siemers (2010)
Behav Ecol Sociobiol

five male individuals of A. stoliczkanus). For each horseshoebat species, we tested 12 adult male bats to eliminate theinfluence of gender. Experiments were conducted in the ex-perimental box which was lined with sound-absorbing foam(Fig. 1). A lid covered the box not to disturb the animalsduring experiments.
The moment a bat was placed on the listening perch, webegan to present a playback of conspecific calls (PCC—play-back of conspecific calls) through a speaker (Ultrasonic Dy-namic Speaker Vifa; Avisoft Bioacoustics) in an infinite loopuntil the bat eventually habituated. The speaker was connectedto an ultrasound playback interface (UltraSoundGate Player116; Avisoft Bioacoustics). Habituation meant that the batsbecame completely motionless (no echolocation; no head, ear,or other body movements; no stretching of wings or legs).After the bat remained habituated for 20 s, we began the testphase by switching from PCC to PCC′ (PCC′—another play-back also of conspecific calls but differs from PCC) or to aplayback of heterospecific calls (PHC—playback ofheterospecific calls). If the tested bat resumed any behavioralactivity (for details, see below), we considered this as indica-tion that the bat had discriminated between the two playbacks(before and after switch) (according to Schuchmann andSiemers 2010), and this bat was scored as responding. Foreach species, four types of playback combinations were pre-sented separately: One PCC–PCC′ combination was used forcontrol (including 20 unique playback file pairs), and threedifferent PCC–PHC combinations were used for testing dis-crimination of heterospecific calls, each containing 25 uniqueplayback file pairs (e.g., for R. macrotis—PCC–PCC’ = Rm–Rm; PCC-PHC = Rm–Rl, Rm–Rs, and Rm–As).
Each bat was tested randomly with one combination perday. We then calculated the proportion of responding bats foreach combination [the number of responding bats/total num-ber of bats tested (12)×100 %].
Behavioral responses
The behaviors of bats to PCC′ and PHC after habituation toPCC were determined over a period of 20 s by analyzing thevideo recordings (the responses decreased dramatically forperiods longer than 20 s after switching playback). All behav-iors occurred were analyzed:
1. Leg movement: we defined this behavior as a positionalchange of the bat’s legs. Each move was counted.
2. Nodding: each movement of the head towards the chestwas counted. If a bat only looked up, it was counted ashalf of a behavioral response.
3. Body movement: this behavior was characterized by anexpansion or rotation of the bat’s body. Each expansion orrotation was counted. If we only observed the contraction
of a bat’s body but no subsequent extension, it wascounted as half of a behavioral response.
4. Wing stretching: each stretching of one wing wascounted.
5. Echolocation: we counted the number of echolocationpulses emitted by test bats.
For each playback combination, less than three individualsdisplayed leg movement or wing stretching. These two be-haviors were thus excluded from the analysis to minimizeeffects of individual variation.
Measurement of call parameters
For each individual recording, five echolocation pulses thathad a SNR better than 30 dB were chosen randomly todetermine their peak frequency, duration, and bandwidth(measured at −12 dB). Mean values of these parameters werecalculated for each species. We then calculated separately thedifferences of these three parameters between any two species.Only absolute values were used for analysis.
For each playback combination, all presented playbackpairs (two playbacks before and after the switch) were classi-fied into two groups: one group included pairs in which thebats remained habituated after switching playbacks, while theother group included those pairs in which the bats showedresponses. For each playback pair, wemeasured peak frequen-cy, duration, and bandwidth (measured at −12 dB) of the lastcall of first playback (before switch) and the first call of thesecond playback (after switch), and then calculated the differ-ence of each of the three call parameters between these twocalls. Only the absolute values were used. All call parameterswere measured in Avisoft-SASLab Pro 4.3 using 512 FFTsizeand an overlap of 87.5 %.
Dietary analysis
Bats were captured with mist nets at the cave entrance whenthey returned from foraging in June of 2013. Captured batswere sexed and put into cloth bags to collect feces. Bats werereleased at the site of their capture after approximately 6 h.Air-dried fecal pellets from each bat were preserved for sub-sequent laboratory analysis. We examined the remains of preyitems in fecal pellets from each of the four species followingKunz and Whitaker (1983). Individual pellets were analyzedfor insect remains by softening the samples in 75 % ethanoland glycerol and teasing them apart under a 6×–50× binoculardissecting microscope. One dropping of an individual wasclassified as one sample. Twelve pellets were analyzed foreach bat; if less than 12, all the feces would be analyzed. Insectremains in the feces were identified up to the order level bycomparing them with insects trapped at our studied site. After
Behav Ecol Sociobiol
the insect parts had been identified, we estimated the percentvolume for each order following Whitaker (1988).
Calculation of trophic niche
The degree of trophic niche overlap between pairs of the fourbat species was calculated using Schoener’s niche overlapindex (Schoener 1970):
NOI ¼ 1− 1=2ð ÞX
i¼1
n
p i; jð Þ��� −p i;kð Þ
�� !
Where p(i,j) and p(i,k) are the proportion of the prey species iin the diets of bat species j and k, respectively.
Statistical analyses
Pearson chi-square tests were used to test whether the propor-tion of bats exhibiting any response after switching playbacksdiffered significantly among the nine heterospecific test com-binations (PCC–PHC) or among the four types of playbackcombinations (one PCC–PCC′, three PCC–PHC) for eachhorseshoe bat species. If it differed significantly among thefour types of playback combinations, Fisher exact test wasused for post hoc pairwise comparisons.
For each horseshoe bat species, comparison of counts ofhead nodding, body movement, and echolocation calls per20 s after switching playbacks was performed by either re-peated measures ANOVA or Friedman tests, followed byBonferroni or Dunn multiple comparison post hoc tests, de-pending on the distribution of the data. In the post hoc tests,we tested whether the counts of each behavior differed signif-icantly between any two test combinations.
In order to test whether the call parameters peak frequency,duration, and bandwidth differed significantly among the fourbat species, one-way ANOVA tests or Kruskal–Wallis H testswere performed, depending on the data distribution.
For each playback combination, the difference be-tween two calls (before and after switch) in each callparameter (peak frequency, duration, and bandwidth) wascompared between responding and non-responding bats usinga Mann–Whitney U test.
For the three horseshoe bat species tested, canonical corre-lation analysis (Arnold and Bennett 1988) was performed totest for the effects of interspecific differences of call charac-teristics on bats’ responses. Interspecific differences includedthree variables: the interspecific difference in peak echoloca-tion frequency (x1), in call duration (x2), and in bandwidth (x3).The response contained three variables: the counts of nodding(y1), body movement (y2), and echolocation pulses (y3).
We performed a simple correlation analysis to test whetherthere was a significant correlation between the interspecifictrophic niche overlap index and the number of each behavior.
We tested the normality of the data distribution usingKolmogorov–Smirnov tests. All statistical tests were per-formed with SPSS 16.0 or GraphPad Instat version 3.0.
Results
Number of responding bats to different playbackcombinations
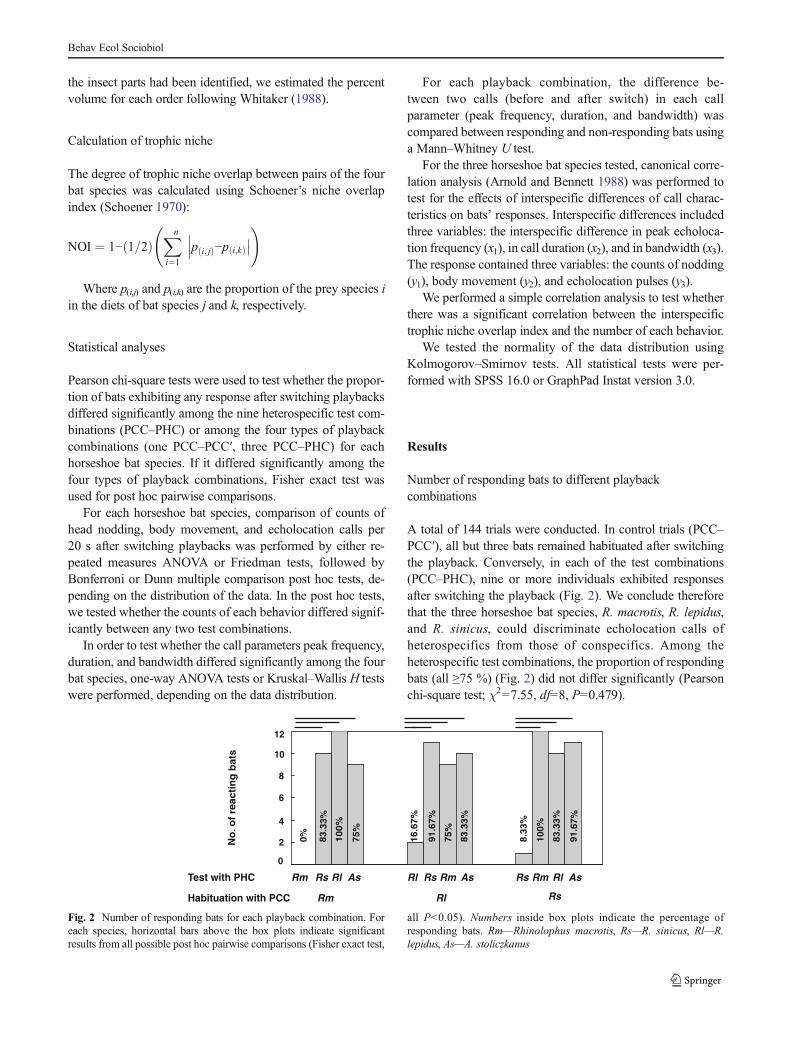
A total of 144 trials were conducted. In control trials (PCC–PCC′), all but three bats remained habituated after switchingthe playback. Conversely, in each of the test combinations(PCC–PHC), nine or more individuals exhibited responsesafter switching the playback (Fig. 2). We conclude thereforethat the three horseshoe bat species, R. macrotis, R. lepidus,and R. sinicus, could discriminate echolocation calls ofheterospecifics from those of conspecifics. Among theheterospecific test combinations, the proportion of respondingbats (all ≥75 %) (Fig. 2) did not differ significantly (Pearsonchi-square test; χ2=7.55, df=8, P=0.479).
No
. of
reac
tin
g b
ats
Test with PHC
Habituation with PCC
Rm Rs Rl As
Rm
Rl Rs Rm As
Rl
Rs Rm Rl As
Rs
0
2
4
6
8
10
12
0% 83.3
3%
100%
75%
16.6
7%
91.6
7%
75%
83.3
3%
8.33
%
100%
83.3
3%
91.6
7%
Fig. 2 Number of responding bats for each playback combination. Foreach species, horizontal bars above the box plots indicate significantresults from all possible post hoc pairwise comparisons (Fisher exact test,
all P<0.05). Numbers inside box plots indicate the percentage ofresponding bats. Rm—Rhinolophus macrotis, Rs—R. sinicus, Rl—R.lepidus, As—A. stoliczkanus
Behav Ecol Sociobiol

For each horseshoe bat species, the proportion ofresponding bats differed significantly among the four typesof playback combinations (Fig. 2; Pearson chi-square test; Rm,χ2=30.88, df=3, P<0.001; Rl, χ2=18.75, df=3, P<0.001; Rs,χ2=31.06, df=3, P<0.001). The horizontal bars above the boxplots indicate significant results from all possible post hocpairwise comparisons (Fisher exact test; all P<0.013).
The behavioral responses to different playback combinations
A total of nine playback combinations (PCC–PHC) weretested to determine the behavioral responses of the threehorseshoe bat species, R. macrotis, R. lepidus, and R. sinicus,to heterospecific echolocation calls. All the three speciesdisplayed a series of behaviors to heterospecific echolocationcalls, including nodding, body movement, wing stretching,leg movement, and echolocation. Nodding, body movement,and echolocation occurred in all playback combinations.Wing stretching was displayed in all but one combination(Rs–Rm), and leg movements occurred in all but three combi-nations (Rl–Rs, Rs–Rl, and Rs–Rm) (Table 1).
For each species, the behavioral response to heterospecificplaybacks depended significantly on the species that theheterospecific call represented (nodding—Friedman test; Rm,Fr=6.13, P=0.046; Rs, Fr=8.06, P=0.018; RM ANOVA; Rl,F=5.50, P=0.012; body movements—Friedman test; Rm,Fr=15.96, P<0.001; RM ANOVA; Rs, F=4.70, P=0.025;Rl, F=5.41, P=0.010). The number of echolocation pulseselicited by different PHC was not significantly different inR. macrotis (Friedman test; Fr=5.43, P=0.085) and R. sinicus(RM ANOVA; F=3.51, P=0.081), but significantly differentin R. lepidus (Friedman test; Fr=8.24, P=0.016) (Fig. 3a).Both R. macrotis and R. lepidus showed significantly fewerbehaviors (nodding and body movement) when exposed toplayback from R. sinicus compared with playbacks from theother species (Bonferroni or Dunn’s corrected; all P<0.049).Similarly, R. sinicus exhibited significantly fewer behaviorswhen exposed to playback from R. macrotis compared withplayback from A. stoliczkanus (Bonferroni or Dunn’scorrected; all P<0.010) (Fig. 3b, c).
Effects of echolocation call on species discrimination
The average resting frequencies of R. macrotis, R. lepidus,R. sinicus, and A. stoliczkanuswere 57.34±0.51 kHz, 91.81±0.91 kHz, 81.91±1.38 kHz, and 125.90±3.03 kHz, re-spectively. The durations (one-way ANOVA test, N=200,F=319.53, P<0.001) and peak frequencies (one-wayANOVA test, N=200, F=1.40, P<0.001) of the echoloca-tion calls differed significantly between the four species,but the bandwidth did not (Kruskal–Wallis H test, χ2=2.96,df=3, P=0.398). Ta
ble1
Behavioralresponses
toecholocatio
ncalls
from
conspecifics
(PCC′)or
heterospecifics(PHC)afterhabituationto
PCC
Com
binatio
nsRm
Rl
Rs
Behaviors
Rm
Rl
As
Rs
Rl
Rm
As
Rs
Rs
Rl
As
Rm
Nodding
02.40
(0–6.5)
1.67
(0–2.5)
0.95
(0.5–1.5)
0.17
(0–1)
1.68
(0.5–4)
2.25
(0.5–4)
1.14
(0.5–2.5)
0.10
(0–1)
1.20
(0.5–1.5)
1.50
(0.5–4.5)
0.56
(0.5–1)
Wingstretching
00.42
(0–2)
0.50
(0–2)
0.09
(0–1)
00.5(0–3)
0.5(0–2)
0.13
(0–0.5)
00.33
(0–2)
1.06
(0–3)
0
Leg
movem
ent
00.25
(0–1)
0.33
(0–1)
0.09
(0–1)
00.17
(0–2)
0.92
(0–4)
00
00.78
(0–2)
0
Bodymovem
ent
01.15
(0–3.5)
1.28
(0–3)
0.35
(0–1.5)
0.50
(0–0.5)
1.14
(0.5–3.5)
0.94
(0.5–1.5)
0.23
(0–1.5)
0.25
(0–1)
1.10
(0–2)
1.10
(0.5–2.5)
0.61
(0–1)
Echolocation
044
(0–252)
57(0–157)
24(0–64)
1(0–2)
43(0–108)
60(0–193)
4(0–21)
1(0– 1)
19(0–86)
23(0–105)
7(0–26)
The
numberof
behavioralresponsesisgivenas
meanandrangein
parentheses.Allbehaviorsaregivenas
totaln
umberoccurringwith
inthe20-strialp
hase
Rm—R.m
acrotis,R
l—R.lepidus,R
s—R.sinicus,A
s—A.stoliczkanus
Behav Ecol Sociobiol
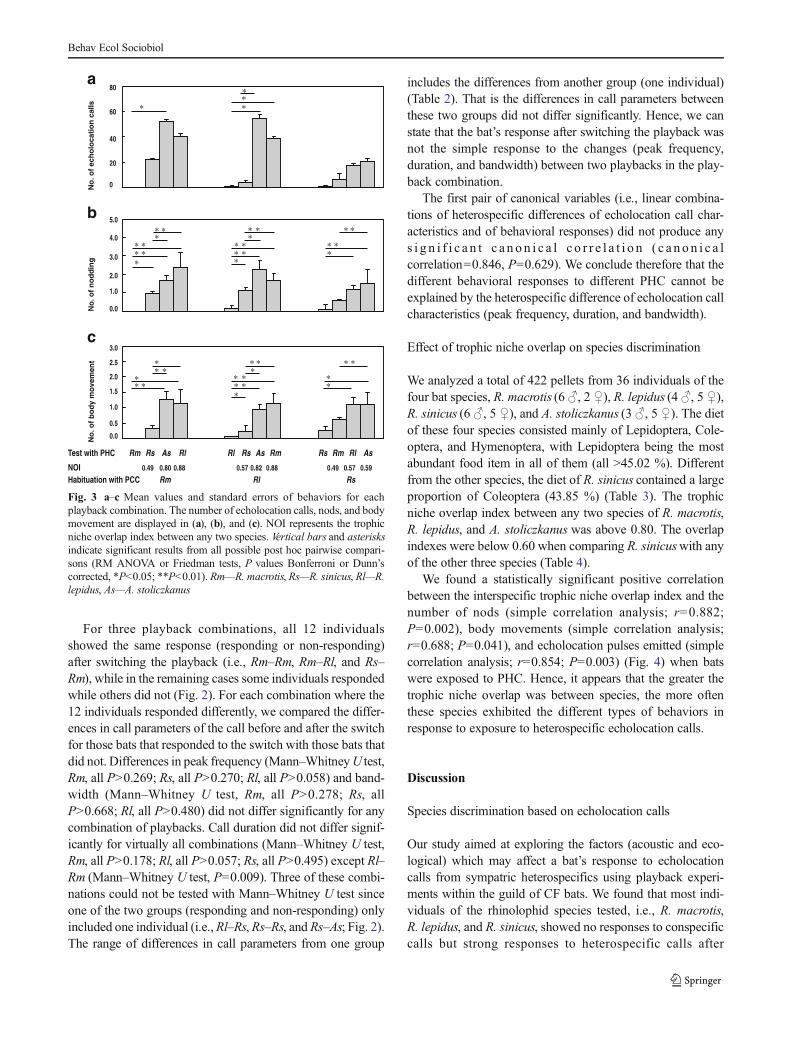
For three playback combinations, all 12 individualsshowed the same response (responding or non-responding)after switching the playback (i.e., Rm–Rm, Rm–Rl, and Rs–Rm), while in the remaining cases some individuals respondedwhile others did not (Fig. 2). For each combination where the12 individuals responded differently, we compared the differ-ences in call parameters of the call before and after the switchfor those bats that responded to the switch with those bats thatdid not. Differences in peak frequency (Mann–WhitneyU test,Rm, all P>0.269; Rs, all P>0.270; Rl, all P>0.058) and band-width (Mann–Whitney U test, Rm, all P>0.278; Rs, allP>0.668; Rl, all P>0.480) did not differ significantly for anycombination of playbacks. Call duration did not differ signif-icantly for virtually all combinations (Mann–Whitney U test,Rm, all P>0.178; Rl, all P>0.057; Rs, all P>0.495) except Rl–Rm (Mann–Whitney U test, P=0.009). Three of these combi-nations could not be tested with Mann–Whitney U test sinceone of the two groups (responding and non-responding) onlyincluded one individual (i.e.,Rl–Rs, Rs–Rs, and Rs–As; Fig. 2).The range of differences in call parameters from one group
includes the differences from another group (one individual)(Table 2). That is the differences in call parameters betweenthese two groups did not differ significantly. Hence, we canstate that the bat’s response after switching the playback wasnot the simple response to the changes (peak frequency,duration, and bandwidth) between two playbacks in the play-back combination.
The first pair of canonical variables (i.e., linear combina-tions of heterospecific differences of echolocation call char-acteristics and of behavioral responses) did not produce anys i g n i f i c a n t c a n on i c a l c o r r e l a t i o n ( c a n on i c a lcorrelation=0.846, P=0.629). We conclude therefore that thedifferent behavioral responses to different PHC cannot beexplained by the heterospecific difference of echolocation callcharacteristics (peak frequency, duration, and bandwidth).
Effect of trophic niche overlap on species discrimination
We analyzed a total of 422 pellets from 36 individuals of thefour bat species, R. macrotis (6♂, 2 ♀), R. lepidus (4♂, 5♀),R. sinicus (6 ♂, 5 ♀), and A. stoliczkanus (3 ♂, 5 ♀). The dietof these four species consisted mainly of Lepidoptera, Cole-optera, and Hymenoptera, with Lepidoptera being the mostabundant food item in all of them (all >45.02 %). Differentfrom the other species, the diet of R. sinicus contained a largeproportion of Coleoptera (43.85 %) (Table 3). The trophicniche overlap index between any two species of R. macrotis,R. lepidus, and A. stoliczkanus was above 0.80. The overlapindexes were below 0.60 when comparing R. sinicuswith anyof the other three species (Table 4).
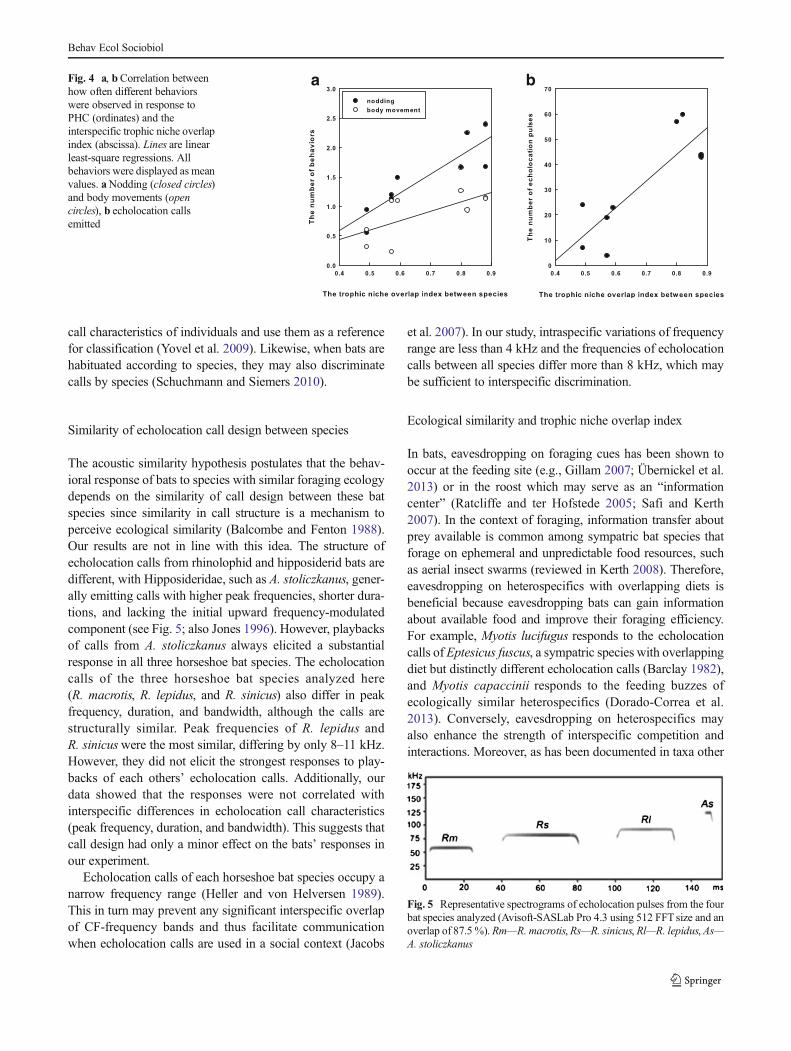
We found a statistically significant positive correlationbetween the interspecific trophic niche overlap index and thenumber of nods (simple correlation analysis; r=0.882;P=0.002), body movements (simple correlation analysis;r=0.688; P=0.041), and echolocation pulses emitted (simplecorrelation analysis; r=0.854; P=0.003) (Fig. 4) when batswere exposed to PHC. Hence, it appears that the greater thetrophic niche overlap was between species, the more oftenthese species exhibited the different types of behaviors inresponse to exposure to heterospecific echolocation calls.
Discussion
Species discrimination based on echolocation calls
Our study aimed at exploring the factors (acoustic and eco-logical) which may affect a bat’s response to echolocationcalls from sympatric heterospecifics using playback experi-ments within the guild of CF bats. We found that most indi-viduals of the rhinolophid species tested, i.e., R. macrotis,R. lepidus, and R. sinicus, showed no responses to conspecificcalls but strong responses to heterospecific calls after
Test with PHC
0.0
0.0
0
NOIHabituation with PCC
No
. of
bo
dy
mo
vem
ent
No
. of
no
dd
ing
No
. of
ech
olo
cati
on
cal
ls
Rm RsRs
Rs
Rs RmRm
Rm
As AsAs Rl
Rl
RlRl
0.490.49 0.80 0.880.88 0.570.57 0.82 0.59
0.5
1.0
1.0
1.5
2.0
2.0
2.5
3.0
3.0
4.0
5.0
80
60
40
20
a
b
c
Fig. 3 a–c Mean values and standard errors of behaviors for eachplayback combination. The number of echolocation calls, nods, and bodymovement are displayed in (a), (b), and (c). NOI represents the trophicniche overlap index between any two species. Vertical bars and asterisksindicate significant results from all possible post hoc pairwise compari-sons (RM ANOVA or Friedman tests, P values Bonferroni or Dunn’scorrected, *P<0.05; **P<0.01). Rm—R. macrotis, Rs—R. sinicus, Rl—R.lepidus, As—A. stoliczkanus
Behav Ecol Sociobiol
habituation. This suggested that these species were indeedable to discriminate calls of heterospecifics from those ofconspecifics. Interestingly, there was no significant negativecanonical correlation between the interspecific differences incall parameters (peak frequency, duration, and bandwidth) andthe behavioral responses of the bats. Finally, in contrast to ourprediction, playbacks of calls from A. stoliczkanus alwayselicited a substantial response in all three horseshoe bat spe-cies.We therefore conclude that echolocation call design had aminor impact on the bats’ responses. Instead, our data showthat the responses depended on the interspecific trophic nicheoverlap index. This implies that the underlying reasons for thedifferent responses to different heterospecific calls may havebeen related to interspecific food competition.
Echolocation pulses are often relatively reliable badges ofspecies identity, in spite of the task specificity of echolocationpulse structure (searching for food or capturing it; e.g., Par-sons and Jones 2000; Russo and Jones 2002). Thus, they havebeen shown to be used by bats to recognize sympatricheterospecifics (Schuchmann and Siemers 2010; Voigt-Heucke et al. 2010). Our results support this finding.
It is well known that echolocation calls, especially of CFspecies, commonly differ in frequency depending on thespecies but also on the sex of the bat for a given species(reviewed in Jones and Siemers 2011). However, in our study,bats were habituated according to species since each playbackfile contained echolocation pulses from both male and femaleindividuals of each of the species. When bats are trained todiscriminate conspecific individuals, bats learn the average
Table2
The
differencesin
callparametersforresponding
andnon-responding
bats
Com
binatio
nsRm–R
s(10,2)
Rm–A
s(9,3)
Rl–Rl
(2,10)
Rl–Rs
(11,1)
Rl–Rm
(9,3)
Rl–As
(10,2)
Rs–Rs
(1,11)
Rs–Rl
(10,2)
Rs–As
(11,1)
Differences
ofcalls
Peak frequency
Responding
(22.69–23.39)
(69.84–71.68)
(0.15,0.60)
(9.83–10.43)
(32.97–33.42)
(36.72–38.20)
(2.11)
(11.80–12.55)
(44.43–46.15)
Non-
responding
(22.84,23.29)
(70.11–71.05)
(0.00–0.76)
(10.13)
(33.12–33.42)
(38.26,38.29)
(1.81–2.52)
(11.95,12.40)
(45.91)
Bandw
idth
Responding
(0.00–0.16)
(0.72–1.38)
(0.14,0.76)
(0.00–0.60)
(0.00–0.30)
(0.60–1.41)
(0.16)
(0.00–0.45)
(0.00–0.46)
Non-
responding
(0.01,0.15)
(0.75–0.97)
(0.14–0.61)
(0.15)
(0.15–0.31)
(0.72,1.03)
(0.01–0.46)
(0.00,0.46)
(0.15)
Duration
Responding
(9.64–25.21)
(18.28–35.11)
(6.36,10.94)
(0.21–13.72)
(0.17–15.84)
(30.03–39.81)
(3.91)
(4.54–22.52)
(37.58–48.57)
Non-
responding
(11.19,23.28)
(20.90–35.11)
(0.30–12.39)
(6.68)
(5.39–13.62)
(35.76,40.23)
(0.09–12.72)
(19.31,25.56)
(47.41)
Undertheplayback
combinatio
ns,the
numbersof
responding
andnon-responding
batsaregivenorderly.The
differencesincallparametersaregivenas
range(alldifferencesaredisplayediftheindividuals
less
than
three)
inparentheses.Allcombinatio
nsaredisplayedexcept
threethatall1
2individualsshow
edthesamereactionaftersw
itching
theplayback
Rm—R.m
acrotis,R
l—R.lepidus,R
s—R.sinicus,A
s—A.stoliczkanus
Table 3 Dietary composition (percentage of volume) of four bat species
Dietcomposition
R. macrotis(n=96)
R. lepidus(n=108)
R. sinicus(n=132)
A. stoliczkanus(n=86)
Lepidoptera 96 84 45 78
Coleoptera 1 4 44 15
Hymenoptera 3 9 5
Diptera 1 2
Orthoptera 2 2
Hemiptera 6
Others 2 1
n number of feces samples analyzed
Table 4 Trophic niche overlap index (0–1) between any two species
R. macrotis R. sinicus R. lepidus A.stoliczkanus
R. macrotis – 0.49 0.88 0.79
R. sinicus 0.49 – 0.57 0.61
R. lepidus 0.88 0.57 – 0.82
A. stoliczkanus 0.79 0.61 0.82 –
Behav Ecol Sociobiol
call characteristics of individuals and use them as a referencefor classification (Yovel et al. 2009). Likewise, when bats arehabituated according to species, they may also discriminatecalls by species (Schuchmann and Siemers 2010).
Similarity of echolocation call design between species
The acoustic similarity hypothesis postulates that the behav-ioral response of bats to species with similar foraging ecologydepends on the similarity of call design between these batspecies since similarity in call structure is a mechanism toperceive ecological similarity (Balcombe and Fenton 1988).Our results are not in line with this idea. The structure ofecholocation calls from rhinolophid and hipposiderid bats aredifferent, with Hipposideridae, such as A. stoliczkanus, gener-ally emitting calls with higher peak frequencies, shorter dura-tions, and lacking the initial upward frequency-modulatedcomponent (see Fig. 5; also Jones 1996). However, playbacksof calls from A. stoliczkanus always elicited a substantialresponse in all three horseshoe bat species. The echolocationcalls of the three horseshoe bat species analyzed here(R. macrotis, R. lepidus, and R. sinicus) also differ in peakfrequency, duration, and bandwidth, although the calls arestructurally similar. Peak frequencies of R. lepidus andR. sinicuswere the most similar, differing by only 8–11 kHz.However, they did not elicit the strongest responses to play-backs of each others’ echolocation calls. Additionally, ourdata showed that the responses were not correlated withinterspecific differences in echolocation call characteristics(peak frequency, duration, and bandwidth). This suggests thatcall design had only a minor effect on the bats’ responses inour experiment.
Echolocation calls of each horseshoe bat species occupy anarrow frequency range (Heller and von Helversen 1989).This in turn may prevent any significant interspecific overlapof CF-frequency bands and thus facilitate communicationwhen echolocation calls are used in a social context (Jacobs
et al. 2007). In our study, intraspecific variations of frequencyrange are less than 4 kHz and the frequencies of echolocationcalls between all species differ more than 8 kHz, which maybe sufficient to interspecific discrimination.
Ecological similarity and trophic niche overlap index
In bats, eavesdropping on foraging cues has been shown tooccur at the feeding site (e.g., Gillam 2007; Übernickel et al.2013) or in the roost which may serve as an “informationcenter” (Ratcliffe and ter Hofstede 2005; Safi and Kerth2007). In the context of foraging, information transfer aboutprey available is common among sympatric bat species thatforage on ephemeral and unpredictable food resources, suchas aerial insect swarms (reviewed in Kerth 2008). Therefore,eavesdropping on heterospecifics with overlapping diets isbeneficial because eavesdropping bats can gain informationabout available food and improve their foraging efficiency.For example, Myotis lucifugus responds to the echolocationcalls of Eptesicus fuscus, a sympatric species with overlappingdiet but distinctly different echolocation calls (Barclay 1982),and Myotis capaccinii responds to the feeding buzzes ofecologically similar heterospecifics (Dorado-Correa et al.2013). Conversely, eavesdropping on heterospecifics mayalso enhance the strength of interspecific competition andinteractions. Moreover, as has been documented in taxa other
a bFig. 4 a, bCorrelation betweenhow often different behaviorswere observed in response toPHC (ordinates) and theinterspecific trophic niche overlapindex (abscissa). Lines are linearleast-square regressions. Allbehaviors were displayed as meanvalues. aNodding (closed circles)and body movements (opencircles), b echolocation callsemitted
Fig. 5 Representative spectrograms of echolocation pulses from the fourbat species analyzed (Avisoft-SASLab Pro 4.3 using 512 FFT size and anoverlap of 87.5 %). Rm—R. macrotis, Rs—R. sinicus, Rl—R. lepidus, As—A. stoliczkanus
Behav Ecol Sociobiol

than bats, eavesdropping may promote increased aggression(Johnstone 2001).
All four species we studied appear to prefer very similarforaging habitats (close to or within vegetation; e.g., Schnitz-ler and Kalko 2001) and similar prey items (moths representedthe majority in their diets). Sympatric species may visit thesame or adjacent feeding grounds. Hence, eavesdropping onheterospecifics and potentially competing for the same preymay be common. (It is important to note, however, that ourstudy was not aimed at testing eavesdropping during foraging.Hence, any statements relating to eavesdropping are, ofcourse, purely speculative.)
Our data show that heterospecific trophic niche overlapwas higher betweenR.macrotis,R. lepidus, andA. stoliczkanusthan between these three species and R. sinicus. This is con-sistent with our observation that R. macrotis and R. lepidusdisplayed the weakest response to the echolocation calls ofR. sinicus. In general, we found that in our playback experi-ments the behavioral responses to heterospecific calls waspositively correlated with the trophic niche overlap indexbetween species. Thus, our results imply that different re-sponse to echolocation calls from different sympatricheterospecifics may be related to interspecific foodcompetition.
The function of behaviors
Consistent with the result of Voigt-Heucke et al. (2010), somebehaviors (e.g., nodding) of bats observed in our experimentcan be considered indicators of emotional arousal. Within thecage, bats displayed the behaviors occurred in our experimentto conspecific or heterospecific other individuals, and finallyall bats kept silence. Although we did not systematicallyinvestigate if these behaviors also occurred naturally, captivebats also often exhibited these behaviors when they weredisturbed (e.g., when opening the cage door and placing foodcontainers into the cage) (personal observations). In other batspecies, nodding may be related to olfactory cues and used forterritorial scent marking (e.g., Saccopteryx bilineata; Casperset al. 2009), and wing stretching may express aggressiontowards intruders in foraging areas (e.g., Cynopterus sphinx;Singaravelan and Marimuthu 2008). Interestingly, the specieswe studied here displayed the same behaviors as Noctilioalbiventris, a different, non-related bat species, under similarbehavioral conditions related to arousal (Voigt-Heucke et al.2010). Therefore, the behaviors that we observed in responseto heterospecific calls may indicate emotional arousal andplay an important role in mediating social interactions.
Species coexistence
Coexisting animals must differ in at least one niche dimensionto avoid excessive competition, e.g., in feeding grounds
(Campbell et al. 2007), diets (Siemers and Swift 2006), activ-ity duration (Jiang et al. 2013), or day roosts (Jacobs andBarclay 2009). Within the bat guild we studied, the differentspecies used more than one strategy to further coexistence.First, the diet overlap between R. sinicus and the other threespecies was low. This may have been the major reason forR. sinicus being the most abundant species in the study area.Second, the different species occupied different roost siteswithin a cave, or even roosted in different caves (personalobservations). Finally, we believe our evidence suggests thatmutual recognition may contribute in enabling them to coex-ist: The strength of heterospecific interactions can be regulatedby heterospecific recognition, meaning that bats may beattracted by feeding buzzes of ecologically similarheterospecifics, but not to those with no overlapping require-ments (Dorado-Correa et al. 2013; Übernickel et al. 2013).Additionally, the response strength of bats may depend on thedegree of social interactions between species (Voigt-Heuckeet al. 2010). This may reduce energy costs associated withunnecessary heterospecific interactions, thus facilitating spe-cies coexistence. Mutualistic and competitive interactions arefrequent and complicated within bat communities (Findley1993). Thus, investigating the interactions among sympatricbat species is likely to help us understand the mechanisms ofspecies coexistence.
Acknowledgments The work was funded by the National NaturalScience Foundation of China (grant no. 31030011, 31270414,31100280, 31100281). We would like to thank Mr. Fushun Yang andhis family, Lifen Xie, and Xiong Guo for their help in the field, andJinhong Luo for his invaluable suggestions and comments on the manu-script. We also thank Songtao Yang for his help in making the experi-mental box and Lei Wang for his help in diet analysis of bats.
Ethical standards All experiments were approved by the Departmentof Wildlife Administration in Shuanghe Township, Yunnan province,China.
References
Abrams P (1980) Some comments on measuring niche overlap. Ecology61:44–49
Arlettaz R, Perrin N, Hausser J (1997) Trophic resource partitioning andcompetition between the two sibling bat species Myotis myotis andMyotis blythii. J Anim Ecol 66:897–911
Arnold SJ, Bennett AF (1988) Behavioural variation in natural popula-tions. V. Morphological correlates of locomotion in the garter snake(Thamnophis radix). Biol J Linn Soc 34:175–190
Balcombe JP, FentonMB (1988) Eavesdropping by bats: the influence ofecholocation call design and foraging strategy. Ethology 79:158–166
Barclay RMR (1982) Interindividual use of echolocation calls:eavesdropping by bats. Behav Ecol Sociobiol 10:271–275
Bee MA (2006) Individual recognition in animal species. In: Naguib M(ed) The encyclopedia of language and linguistics, , 2nd edn.Elsevier, London, pp 617–626
Behav Ecol Sociobiol

Brunet-Rossinni AK, Wilkinson GS (2009) Methods for age estimationand the study of senescence in bats. In: Kunz TH, Parsons S (eds)Ecological and behavioral methods for the study of bats. The JohnHopkins University Press, Baltimore, pp 315–325
Campbell P, Schneider CJ, Zubaid A, Adnan AM, Kunz TH (2007)Morphological and ecological correlates of coexistence inMalaysian fruit bats (Chiroptera: Pteropodidae). J Mammal 88:105–118
Caspers B,Wibbelt G, Voigt CC (2009) Histological examinations of facialglands in Saccopteryx bilineata (Chiroptera, Emballonuridae), andtheir potential use in territorial marking. Zoomorphology 128:37–43
Cody ML (1969) Convergent characteristics in sympatric bird popula-tions: a possible relation to interspecific territoriality. Condor 71:223–234
CodyML (1973) Character convergence. Annu Rev Ecol Syst 4:189–211Coolen I, Bergen YV, Day RL, Laland KN (2003) Species difference in
adaptive use of public information in sticklebacks. Proc R Soc LondB 270:2413–2419
Dorado-Correa AM, Goerlitz HR, Siemers BM (2013) Interspecificacoustic recognition in two European bat communities. FrontPhysiol 4:192
Fenton MB (2003) Eavesdropping on the echolocation and social calls ofbats. Mammal Rev 33:193–204
Findley JS (1993) Bats: a community perspective. Cambridge UniversityPress, Cambridge
Forsman JT, Mönkkönen M, Helle P, Inkeröinen J (1998) Heterospecificattraction and food resources in migrants’ breeding patch selectionin northern boreal forests. Oecologia 115:278–286
Gillam EH (2007) Eavesdropping by bats on the feeding buzzes ofconspecifics. Can J Zool 85:795–801
Goodale E, Beauchamp G, Magrath RD, Nieh JC, Ruxton GD (2010)Interspecific information transfer influences animal communitystructure. Trends Ecol Evol 25:354–361
Grether GF, Losin N, Anderson CN, Okamoto K (2009) The role ofinterspecific interference competition in character displacement andthe evolution of competitor recognition. Biol Rev 84:617–635
Griffin DR (1958) Listening in the dark. Yale University Press, NewHaven
Griffin DR, Webster FA, Michael CR (1960) The echolocation of flyinginsects by bats. Anim Behav 8:141–154
Heller KG, von Helversen O (1989) Resource partitioning of sonarfrequency bands in rhinolophoid bats. Oecologia 80:178–186
Hurd CR (1996) Interspecific attraction to the mobbing calls of black-capped chickadees (Parus atricapillus). Behav Ecol Sociobiol 38:287–292
Jacobs DS, Barclay RMR (2009) Niche differentiation in two sympatricsibling bat species Scotophilus dinganiiand Scotophilus mhlanganii.J Mammal 90:879–887
Jacobs DS, Barclay RMR, Walker MH (2007) The allometry of echolo-cation call frequencies of insectivorous bats: why do some speciesdeviate from the pattern? Oecologia 152:583–594
Jiang TL, Lu GJ, Sun KP, Luo JH, Feng J (2013) Coexistence ofRhinolophus affinis and Rhinolophus pearsoni revisited. ActaTheriol 58:47–53
Johnstone RA (2001) Eavesdropping and animal conflict. Proc Natl AcadSci U S A 98:9177–9180
Jones G (1996) Does echolocation constrain the evolution of body size inbats? Symp Zool Soc Lond 69:111–128
Jones G, Siemers BM (2011) The communicative potential of bat echo-location pulses. J Comp Physiol A 197:447–457
Kerth G (2008) Causes and consequences of sociality in bats. Bioscience58:737–746
Korner P, Whiting MJ, Willem J, Ferguson H (2000) Interspecific ag-gression in flat lizards suggests poor species recognition. Afr JHerpetol 49:139–146
Kunz TH, Whitaker JO (1983) An evaluation of fecal analysis fordetermining food habits of insectivorous bats. Can J Zool 61:1317–1321
Lynch A, Baker AJ (1991) Increased vocal discrimination by learning insympatry in 2 species of chaffinches. Behaviour 116:109–126
Marler P (1957) Specific distinctiveness in the communication signals ofbirds. Behaviour 11:13–39
Moore RF (1978) Interspecific aggression: toward whom should a mock-ingbird be aggressive? Behav Ecol Sociobiol 3:173–176
Parsons S, Jones G (2000) Acoustic identification of twelve species ofecholocating bat by discriminant function analysis and artificialneural networks. J Exp Biol 203:2641–2656
Qvarnström A, Haavie J, Sæther SA, Eriksson D, Pärt T (2006) Songsimilarity predicts hybridization in flycatchers. J Evol Biol 19:1202–1209
Racey PA (2009) Reproductive assessment of bats. In: Kunz TH, ParsonsS (eds) Ecological and behavioral methods for the study of bats. TheJohn Hopkins University Press, Baltimore, pp 249–264
Ratcliffe JM, ter Hofstede HM (2005) Roosts as information centres:social learning of food preferences in bats. Biol Lett 1:72–74
Russo D, Jones G (2002) Identification of twenty-two bat species(Mammalia: Chiroptera) from Italy by analysis of time-expandedrecordings of echolocation calls. J Zool 258:91–103
Ryan MJ (1983) Frequency modulated calls and species recognition in aNeotropical frog. J Comp Physiol 150:217–221
Safi K, Kerth G (2007) Comparative analyses suggest that informationtransfer promoted sociality in male bats in the temperate zone. AmNat 170:465–472
Schnitzler H-U, Kalko EKV (2001) Echolocation by insect-eating bats.Bioscience 51:557–569
Schoeman MC, Jacobs DS (2011) The relative influence of competitionand prey defences on the trophic structure of animalivorous batensembles. Oecologia 166:493–506
Schoener TW (1968) The Anolis lizards of Bimini: resource partitioningin a complex fauna. Ecology 49:704–726
Schoener TW (1970) Nonsynchronous overlap of lizards in patchy hab-itats. Ecology 51:408–418
Schuchmann M, Siemers BM (2010) Behavioral evidence forcommunity-wide species discrimination from echolocation calls inbats. Am Nat 176:72–82
Siemers BM, Swift SM (2006) Differences in sensory ecology contributeto resource partitioning in the bat Myotis bechsteinii and Myotisnattereri (Chiroptera Vespertilionidae). Behav Ecol Sociobiol 59:373–380
Singaravelan N, Marimuthu G (2008) In situ feeding tactics ofshort-nosed fruit bat (Cynopterus sphinx) on mango fruits:evidence of extractive foraging in a flying mammal. J Ethol26:1–7
Stefanski RA, Falls JB (1972) A study of distress calls of song, swampand white-throated sparrows (Aves: Fringillidae). II. Interspecificresponses and properties used in recognition. Can J Zool 50:1513–1525
Übernickel K, Tschapka M, Kalko EKV (2013) Selective eavesdroppingbehaviour in three Neotropical bat species. Ethology 119:66–76
Voigt-Heucke SL, Taborsky M, Dechmann DKN (2010) A dualfunction of echolocation: bats use echolocation calls to iden-tify familiar and unfamiliar individuals. Anim Behav 80:59–67
Whitaker JO (1988) Food habits of insectivorous bats. In: Kunz TH (ed)Ecological and behavioral methods for the study of bats.Smithsonian Institution Press, Washington, pp 171–189
Yovel Y, Melcon ML, Franz MO, Denzinger A, Schnitzler H-U (2009)The voice of bats: how greater mouse-eared bats recognize individ-uals based on their echolocation calls. PLoS Comput Biol 5:e1000400
Behav Ecol Sociobiol





























