Embed Size (px)
Citation preview

Aus dem Institut für Tierhygiene, Tierschutz und Nutztierethologieder Tierärztlichen Hochschule Hannover
Belastung von Schlachtschweinen in zwei Zuführungssystemen zur Elektrobetäubung und die Auswirkungen
auf das Wohlbefinden
INAUGURAL – DISSERTATION
Zur Erlangung des Grades einer Doktorinder Veterinärmedizin
(Dr. med. vet.)durch die Tierärztliche Hochschule Hannover
Vorgelegt vonMelanie Außel
aus Rheda-Wiedenbrück
Hannover 2001

Wissenschaftliche Betreuung: Univ.-Prof. Dr. J. Hartung
1.Gutachter: Univ.-Prof. Dr. J. Hartung
2. Gutachter: Univ.-Prof. Dr. K.-H. Waldmann
Tag der mündlichen Prüfung: 23. November 2001
Gefördert durch das Bundesministerium für Verbraucherschutz, Ernährung undLandwirtschaft, Bonn (Projektträger: Bundesanstalt für Landwirtschaft und Ernährung,Frankfurt)

Für Ben


Aus der vorliegenden Dissertation wurde bereits folgender Teil veröffentlicht:
AUßEL, M., M. MARAHRENS, H. HIEGEMANN u. J. HARTUNG (1999):Das Verhalten von Schlachtschweinen in zwei unterschiedlichen Zuführungssystemen zurElektrobetäubung.23. Kongreß der DVG, Bad Nauheim, 13. - 16. April 1999, Poster Nr. 73


Inhaltsverzeichnis
1 Einleitung 11
2 Literaturübersicht 132.1 Grundlagen der Belastungsbewertung 132.1.1 Stresstheorien und Modelle 132.1.1.1 Zusammenhang zwischen Belastung und Wohlbefinden 172.1.2 Die Belastungsreaktion 192.1.2.1 Zentrale Ebene 192.1.2.2 Periphere Ebene 212.1.2.2.1 Hypothalamo-Hypophysär-Adrenale Achse 212.1.2.2.2 Sympatho-Adrenomedulläres System 232.2 Belastungsindikatoren beim Schwein 272.2.1 Herzfrequenz 272.2.2 Körpertemperatur 292.2.3 Catecholamine 302.2.4 Cortisol 312.2.5 Lactat 332.2.6 Verhalten 342.3 Die Zuführung zur Betäubung 36
3 Eigene Untersuchungen 393.1 Material, Methoden und Tiere 403.1.1 Tiere 403.1.2 Transport 413.1.3 Schlachtbetriebsmanagement und bauliche Einrichtungen 413.1.4 Untersuchungsparameter 453.1.4.1 Erfassung der Klimaverhältnisse 453.1.4.2 Herzfrequenz 453.1.4.3 Körpertemperatur 473.1.4.4 Katheterisierung der Schweine 493.1.4.5 Gewinnung und Behandlung der Blutproben 493.1.4.6 Biochemische Untersuchungen (Laboruntersuchungen) 503.1.4.7 Ethologische Untersuchungen 533.1.5 Statistische Auswertung der Befunde 563.2 Befunde 583.2.1 Physiologische Belastungsindikatoren 593.2.1.1 Herzfrequenz 593.2.1.2 Körpertemperatur 613.2.2 Biochemische Belastungsindikatoren 633.2.2.1 Noradrenalin 633.2.2.2 Adrenalin 653.2.2.3 Adrenalin und Noradrenalin im Stichblut 673.2.2.4 Cortisol 693.2.2.5 Lactat 71

3.2.3 Ethologische Befunde 733.2.3.1 Verhalten im Wartestall 733.2.3.2 Dauer der Vereinzelung und des Zutriebs 753.2.3.3 Verhalten während der Vereinzelung 763.2.3.4 Verhalten während des Zutriebs 783.2.3.5 Einsatz von Treibhilfen 803.2.3.6 Anzahl der Verhaltenswechsel 82
4 Diskussion 844.1 Belastungsreaktion der Schweine vor und nach dem Einbau
des neuen Zuführungssystems anhand physiologischer undhämatologischer Belastungsindikatoren
85
4.2 Belastungsreaktion der Schweine vor und nach dem Einbaudes neuen Zuführungssystems anhand ethologischer Parameter
93
4.3 Bedeutung der baulichen Gestaltung des Zuführungssystemsfür das Wohlbefinden der Schweine
99
5 Schlussfolgerungen 102
6 Zusammenfassung 104
7 Summary 107
8 Literaturverzeichnis 110
9 Tabellenanhang 141

Abkürzungsverzeichnis
Abb. AbbildungACTH Adrenocorticotropes HormonATP AdenosintriphosphatAVP Arginin-VasopressinB SchlachtbandBE BlutentnahmeBHZP Bundeshybrid-Zuchtprogrammbpm. beats per minutebzw. beziehungsweiseC CelsiusCBG Corticosteroid-bindendes-GlobulinCoA Coenzym Acm ZentimeterCo2 KohlendioxidCRH Corticotropin-Releasing HormoneDBH Dopamin-ß-hydroxylaseDOPA 3,4 DihydroxyphenylalaninDHBA DihydroxybenzylaminEDTA EthylendiamintetraacetatEGTA Ethylenbis (oxyethylennitrilo)-Tetraessigsäureg GrammG GaugeGABA Gamma-Amino-ButtersäureHF HerzfrequenzHHA Hypothalamo-Hypophysär-AdrenalHPLC HochdruckflüssigkeitschromatografieHVL HypophysenvorderlappenI.E. Internationale Einheitenkg KilogrammKT KörpertemperaturLDH Lactat-DehydrogenaseLKW LastkraftwagenLSM Least Squares Meansm Meterm2 QuadratmeterM Molaritätml Millilitermm Millimetermmol MillimolMtlw. Mittelwertn AnzahlNA NoradrenalinNAD Nicotinamid-adenin-dinucleotidNADH reduziertes Nicotinamid-adenin-dinucleotidNADP Nicotinamid-dinucleotid-phosphat

NADPH reduziertes Nicotinamid-adenin-dinucleotid-phosphatng Nanogrammnm Nanometernmol NanomolNNM NebennierenmarkNNR Nebennierenrindep Signifikanz der DifferenzenPNMT Phenylethanolamin-N-methyltransferasePOMC ProopiomelanocortinS StandardabweichungS/min Schläge pro MinuteSAM Sympatho-Adrenomedulläres-Systemsec. SekundeT TransportTab. TabelleTemp. UmgebungstemperaturTH TyrosinhydroxylaseTh 1-5 Treibhilfen 1-5TierSchlV Tierschutz-SchlachtverordnungU Umtriebu. undUK United KingdomUSA United States of AmericaVh 1-6 Verhaltensweisen 1-6VIP Vasoaktives intestinales Peptidvs. versusvV vor VorladenW WartestallW min Wartestall minimumZ Zuführungz.B. zum BeispielZNS Zentrales Nervensystem

11
1 Einleitung
Angesichts eines allgemein zunehmenden Interesses für das Wohlbefinden
landwirtschaftlicher Nutztiere ist in den letzten Jahren auch der aus Tierschutzsicht äußerst
sensible Bereich der Schlachtung in den Vordergrund gerückt. Bei der Schlachtung sind die
Tiere nicht nur im unmittelbaren Schlachtvorgang sondern auch während des Aufenthaltes im
Wartestall und besonders bei der Zuführung zur Betäubung erheblichen Belastungen
ausgesetzt, obwohl die Tierschutz-Schlachtverordnung (1997) in § 3 fordert, auch die mit der
Schlachtung in Zusammenhang stehenden Vorgänge so schonend wie möglich zu gestalten
und den Tieren dabei nicht mehr als unvermeidbare Aufregung, Schmerzen, Leiden und
Schäden zuzufügen. Um Schlachtschweine der gesetzlich vorgeschriebenen Betäubung
zuzuführen, kommen allerdings häufig Systeme zum Einsatz, die in erster Linie darauf
ausgerichtet sind, die vom Schlachtbetrieb geforderte möglichst lückenlose Auslastung der
Betäubungsanlage und hohe Schlachtzahlen zu gewährleisten. So ist in europäischen
Schlachtbetrieben eine als „lining up“ bezeichnete Aufreihung der Tiere im Zutrieb üblich.
Dabei werden Treibgänge verwendet, durch die die Schweine einzeln hintereinander zur
Betäubungsfalle gelangen. Kritisch ist dabei vor allem die dem Eintrieb in diese Gänge
vorausgehende Vereinzelung der Tiere zu beurteilen. Die von den Tieren verlangte aktive
Fortbewegung in Richtung Betäubung kann häufig nur durch den mitunter massiven Einsatz
von Treibhilfen erreicht werden. Beides führt zu einer erheblichen Belastung der Tiere, die
das Wohlbefinden beeinträchtigt und die Fleischqualität nachteilig beeinflussen kann. Seit
längerer Zeit wird daher gefordert, Zuführungssysteme für Schweine verhaltensgerechter zu
entwickeln und sie so zu gestalten, dass das Vorwärtsgehen der Schweine ohne
Zwangsmaßnahmen erreicht werden kann. Dies würde einen schonenden Umgang der Treiber
mit den Tieren ermöglichen und einen Teil der Belastung von den Schlachtschweinen nehmen
helfen. Anlagen dieser Art sind bisher jedoch selten und wurden in der Praxis noch nicht im
Detail untersucht.
In der vorliegenden Arbeit wurde geprüft, inwieweit sich die Zuführung zur Betäubung durch
bauliche Maßnahmen tiergerechter und belastungsärmer gestalten lässt. Dazu wurde in einem
kommerziellen Schlachtbetrieb ein herkömmliches Zuführungssystem untersucht und nach
Umbau des Schlachtbetriebes mit einem neuartigen, nach ethologischen Gesichtspunkten
entwickelten System verglichen. Zur Einschätzung der Belastung und Beurteilung des
Wohlbefindens der Schweine wurden physiologische, biochemische und ethologische
Untersuchungen durchgeführt. Die der Zuführung zur Betäubung

12
vorgeschaltetenVerfahrensabschnitte wie Aufenthalt im Wartestall und Transport wurden zur
Kennzeichnung der Vorbelastung in die Untersuchungen einbezogen.

13
2 Literaturübersicht
2.1 Grundlagen der Belastungsbewertung
2.1.1 Stresstheorien und Modelle
Zur Kennzeichnung von Belastungssituationen und/oder -reaktionen bei Mensch und Tier hat
sich sowohl im populären, als auch im wissenschaftlichen Sprachgebrauch weitgehend der
Begriff “Stress” etabliert. In der Literatur gibt es viele Versuche, eine umfassende Definition
von “Stress” zu geben, ohne dass dies bisher gelungen wäre (BROOM und JOHNSON 1993,
LADEWIG 1994). Ein wesentlicher Grund hierfür ist die Nutzung des Terminus als
Sammelbegriff für verschiedene Aspekte des Stressgeschehens. So werden beispielsweise
häufig sowohl die auf den Organismus einwirkenden Umweltereignisse (Stressstimuli), als
auch die darauffolgenden Körperreaktionen (Stressreaktionen) vom Begriff “Stress” umfasst
(LADEWIG 1987, SMIDT et al. 1988, BROOM und JOHNSON 1993).
Eine für die Erforschung der Körperreaktionen auf belastende Umweltereignisse wesentliche
Beobachtung machte W.B.CANNON (1915, 1928): Er wies nach, dass eine Bedrohung des
Organismus zu einem erhöhten Adrenalinspiegel im venösen Blut der Nebenniere führt.
Diese Erkenntnis war Grundlage für die Formulierung seines Stresskonzepts über die
Notwehrfunktion des Nebennierenmarks. Hierbei wird durch Adrenalin eine erhöhte
Sauerstoff- und Energiebereitstellung in Skelettmuskulatur und Gehirn erreicht, die den
Organismus auf körperliche Aktivität im Sinne von Flucht und/oder Kampf vorbereitet,
wodurch die Überlebenschancen in Notfallsituationen steigen (“Fight and Flight Syndrome”).
Stress definierte SELYE (1936, 1950) als eine unspezifische, auf eine Vielfalt von
schädlichen Reizen (Stressoren) gleichartig erfolgende Reaktion des Organismus, die zu
Nebennierenhypertrophie, Atrophie lymphatischer Organe und gastrointestinaler Ulceration
(sog. “Stresstrias”) führen kann. Er unterschied drei Stadien der Stressantwort, die er als
“generalisiertes Adaptationssyndrom” zusammenfasste: 1. die “Alarmreaktion”, die neben
einer Aktivierung des peripheren sympathischen Nervensystems hauptsächlich durch eine
gesteigerte Corticosteroidfreisetzung aus den Nebennieren gekennzeichnet ist, 2. das
“Adaptationsstadium”, in dem die Widerstandsfähigkeit des Organismus erhöht ist, und 3. das
“Stadium der Erschöpfung” mit Zusammenbruch der Anpassung. Weiterhin unterschied er
zwischen positivem Eustress und negativem Distress. Auch WIEPKEMA und KOOLHAAS

14
(1993) unterscheiden zwischen physiologischem und pathologischem Stress, wohingegen
andere Autoren den Begriff ausschließlich dann verwenden, wenn von einem Tier extreme
Anpassungen an ungünstige oder schädliche Umweltbedingungen verlangt werden (RÜGER
et al. 1990, DANTZER 1994).
Die älteren linearen “Stimulus-Belastungsantwort-Modelle” (DANTZER 1994, TOATES
1995) wurden durch transaktionale Konzepte, die die Interaktionen zwischen Individuum und
Belastungssituation unter Beteiligung des kognitiven ZNS einbeziehen, ergänzt. Nach
MASON (1971), BURCHFIELD (1979) und von HOLST und SCHERER (1988) führen
allein die Stimuli, die eine emotionelle Erregung, zum Beispiel Angst auslösen, zu
Belastungsreaktionen, wobei Reize und Ereignisse schon aufgrund ihrer Unbekanntheit oder
“Neuheit” (novelty) zu Stressoren werden können (WIEPKEMA und KOOLHAAS 1993).
Dieses Konzept wurde als “Kognitive Mediator Theorie” bezeichnet (LADEWIG 1994).
Transaktionale Konzepte gehen insgesamt davon aus, dass die Stressreaktion eines
Individuums von seinen Möglichkeiten zur Konfrontation einer Belastungssituation mit
Entwicklung von Bewältigungsstrategien (“coping”) abhängt, und somit
Belastungsantworten nicht nur unspezifisch, sondern auch differenziert, stressorabhängig und
individuell geprägt sein können (LAZARUS 1966, WEISS 1972, HENRY und STEPHENS
1977, LAZARUS und FOLKMANN 1984, LEVINE et al. 1989, deBOER et al. 1990,
DANTZER 1994). “Coping” wird von LAZARUS und LAUNIER (1978) als “das Gesamt der
sowohl aktionsorientierten wie intrapsychischen Anstrengungen, die ein Individuum
unternimmt, um externale und internale Anforderungen, die seine Ressourcen beanspruchen
oder übersteigen, zu bewältigen” definiert. Zur Ausbildung von Bewältigungsstrategien ist
sowohl die Möglichkeit eines Individuums, Einfluss auf eine Situation nehmen zu können,
das heisst seine Möglichkeit zur Kontrolle, als auch das Vermögen, das Vorkommen oder
Nicht-Vorkommen eines bestimmten Ausgangs vorauszusehen, das heisst die
Vorhersehbarkeit (predictability) von entscheidender Bedeutung (WEISS 1971, HENRY und
STEPHENS 1977, ARTHUR 1987, WIEPKEMA und KOOLHAAS 1992). So entwickelten
zum Beispiel Ratten, die die Verabfolgung eines Elektroschocks einerseits aktiv unterbrechen
und andererseits durch ein Lichtsignal verlässlich vorhersehen konnten (Konditionierung)
weitaus geringere Stresssymptome als Ratten, bei denen das Lichtsignal in keinem
Zusammenhang mit der Verabfolgung des Elektroschocks stand. Die aus diesem Experiment
ersichtliche positive Auswirkung der Vorhersehbarkeit erklären einige Autoren damit, dass
das Nicht-Erscheinen von Warnsignalen offensichtlich selbst wie ein „Signal“ wirkt, dass
eine Phase der Sicherheit anzeigt („safety signal“) (WEINBERG und LEVINE 1980,
TORTORA 1983, SCHWITZGEBEL 1992). Die Konfrontation mit unkontrollierbaren

15
und/oder nicht vorhersehbaren intensiven Stressoren (“major life events”) kann zu einem
“erlernte Hilflosigkeit” genannten Überforderungseffekt führen, der durch eine Verringerung
der Regulations- und Adaptationsfähigkeit gekennzeichnet ist und sogar zum Tod führen kann
(SELIGMAN 1975, MAIER und SELIGMAN 1976, WEINBERG und LEVINE 1980,
LEVINE et al. 1989, WIEPKEMA und KOOLHAAS 1993). Negative Auswirkungen ergeben
sich ebenso, wenn die Vorhersehbarkeit von positiven Ereignissen (zum Beispiel Erhalt von
Futter) gering ist (DANTZER et al. 1980, DANTZER und MORMEDE 1983, CARLSTEAD
1986). WIEPKEMA und KOOLHAAS (1993) weisen darauf hin, dass eine durch reizarme
Haltungsbedingungen hervorgerufene sehr hohe Vorhersehbarkeit und/oder Kontrollierbarkeit
ebenfalls schädlich sein kann. Grundlage für eine kognitive Kontrolle über und erfolgreiche
Anpassung an wechselnde Umweltbedingungen ist die individuelle Möglichkeit zur
“...adaptiven Verhaltensänderung als Folge individueller Informationsaufnahme,
Informationsverarbeitung und Informationsspeicherung (Erfahrung) auf der Grundlage
phylogenetisch vorgebildeter und artspezifisch modifizierbarer nervöser Strukturen” (SINZ
1974), das heisst die Lernfähigkeit eines Individdums (WEINBERG und LEVINE 1980,
NICOL 1996). Gleichzeitig mit assoziativen Lernprozessen, die bei höher entwickelten
Wirbeltieren hauptsächlich in synaptisch eng verknüpften Bereichen der Großhirnrinde und
des Limbischen Systems stattfinden (HUETHER 1996), geht der Aufbau eines persönlichen
“Erfahrungsschatzes” einher, der im Falle einer wiederholten Konfrontation mit weniger
intensiven Stressoren, wie z.B. einer nicht schmerzhaften Immobilisation für kurze Zeit, die
Stressreaktion des Individuums verringern oder ganz ausbleiben läßt (Habituation) (deBOER
et al. 1990, WIEPKEMA und KOOLHAAS 1992, LADEWIG 1994). Die Gewöhnung an
einen Stressor ist dabei spezifisch und wird nicht auf andere Stressoren übertragen (KANT et
al. 1985). Intensive (zum Beispiel schmerzhafte) Stressoren führen dagegen auch nach
zahlreichen Wiederholungen nicht zu einer Gewöhnung. Vielmehr bleibt die
Belastungsantwort unverändert oder es setzt eine die Belastungsantwort verstärkende
Sensibilisierung ein (ARTHUR 1987, KONARSKA et al. 1990). “Coping” beinhaltet somit
beim höheren Wirbeltier das Vermögen, Situationen anhand der gemachten Erfahrungen zu
bewerten und Konsequenzen von Handlungen zu verstehen, um so die adaequate Strategie
auswählen zu können (WIEPKEMA und KOOLHAAS 1992, DANZTER 1994, TSCHANZ
1995). Wenn vorhandene Bewältigungsstrategien fehlschlagen oder nicht ausführbar sind, wie
im Falle einer nicht gegebenen Fluchtmöglichkeit, kommt es nach Meinung zahlreicher
Autoren zu Stressreaktionen (von HOLST 1977, BROOM und JOHNSON 1993,
WECHSLER 1993, WIEPKEMA und KOOLHAAS 1993, HUETHER 1996). Abbildung 1
veranschaulicht ein Konzept, das die außerordentliche Komplexität und Dynamik im
Stressgeschehen erfasst und als “Stundenglas-Modell” bezeichnet wird (VEITH-FLANIGAN

16
und SANDMAN 1985). Stress umfasst dabei alle möglichen extraindividuellen Ereignisse,
die in der Lage sind, ein breites Spektrum von intraindividuellen Reaktionen hervorzurufen,
nachdem sie durch einen komplexen Filter von individuellen Unterschieden gegangen sind.
Die “extraindividuellen Ereignisse” (Stressoren) können nach qualitativen
(physisch/emotional) und quantitativen Eigenschaften (Dauer/Intensität) unterschieden
werden (LADEWIG 1994). Für den Prozess der individuellen Filterung spielen sowohl die
früheren Erfahrungen des Individuums, als auch die genetische Disposition eine Rolle
(DANTZER und MORMEDE 1985, SMIDT et al. 1988, von BORELL und LADEWIG
1989).
2.1.1.1 Zusammenhang zwischen Belastung und Wohlbefinden
Abb. 1: Das „Stundenglas“ – Modell des Stress – Konzeptes (nach Veith – Flanigan undSandman 1985)

17
Übermäßige Belastung (Stress) als Folge unzureichender oder nicht anwendbarer
Bewältigungsstrategien wird in der Literatur vielfach als ein Zustand angesehen, der das
Wohlbefinden beeinträchtigen kann (van PUTTEN 1982, BROOM 1991, BROOM und
JOHNSON 1993, DANZTER 1994, LAMBOOIJ et al. 1995, WIEPKEMA und KOOLHAAS
1993, HUETHER 1996). Der Begriff Wohlbefinden, der auch im deutschen Tierschutzgesetz
Verwendung findet, ist im veterinärmedizinischen Sprachgebrauch relativ neu und drückt die
aus ethischem Interesse geäußerte Sorge des Menschen um die Befindlichkeit von in seiner
Obhut lebenden Tieren aus (DANTZER 1994). Das Wohlbefinden eines Tieres wird von
einigen Autoren als “ein Zustand von geistiger und körperlicher Gesundheit, der ein Leben in
Harmonie mit sich selbst und der Umwelt anzeigt” charakterisiert (LORZ 1973, van PUTTEN
1982, DUNCAN und DAWKINS 1983, WIEPKEMA und KOOLHAAS 1992). Diese
Definition stimmt mit dem häufig geäußerten Schluss überein, dass physische und/oder
psychische Leiden Wohlbefinden ausschließen (DANTZER und MORMEDE 1981,
LADEWIG und von BORELL 1988, GREGORY 1998). Durch körperliche
Beeinträchtigungen wie Hunger, Durst, Schmerz oder Krankheit hervorgerufenes Leiden
stand lange Zeit im Mittelpunkt des Interesses (DANZTER 1994), während die psychische
Leidensfähigkeit von Tieren bis heute im wissenschaftlichen Schrifttum nicht unumstritten ist
(WIEPKEMA und KOOLHAAS 1992, SAMBRAUS 1999). Zahlreiche Autoren gehen
jedoch inzwischen davon aus, dass das Wohlbefinden höherer Wirbeltiere sowohl durch die
Nichtbefriedigung von Bedürfnissen (WIEPKEMA und KOOLHAAS 1993, DANTZER
1994, GREGORY 1998), als auch durch weithin als unangenehm bewertete emotionale
Phänomene wie Angst (SAMBRAUS 1982, GRAY 1982, ROWAN 1988, STAUFFACHER
1993) beeinträchtigt werden kann. Die Annahme einer grundsätzlichen, dem Menschen
vergleichbaren Empfindungsfähigkeit (SAMBRAUS 1995, 1998, BEKOFF 1997) wird durch
die evolutionsbiologische Tatsache, dass Merkmale von Vertretern verschiedener Tierarten
homolog sind, wenn sie sich von denselben Merkmalen ihrer gemeinsamen Vorfahren
ableiten lassen (REMANE 1952), und somit neuronale (“hard-wired brain circuits”) und
hormonelle Systeme bei Menschen und höheren Wirbeltieren homolog sind, begründet
(PANKSEPP 1982, TOATES 1986, BUCHHOLTZ 1993). Transaktionale Stresskonzepte
beinhalten das Erscheinen von kognitiven und emotionalen Fähigkeiten und leisten einen
Beitrag zur Integration der psychischen Komponente in die wissenschaftliche Diskussion um
Wohlbefinden (WIEPKEMA und KOOLHAAS 1993, DANTZER 1994). Abbildung 2
veranschaulicht ein Konzept, wonach Belastungssituationen einen bestimmten “emotionalen
Zustand” hervorrufen, der von Furcht bis Freude variieren kann (WIEPKEMA und
KOOLHAAS 1992) und von dem die Reaktion des Individuums abhängt. Bestimmte Mimik,
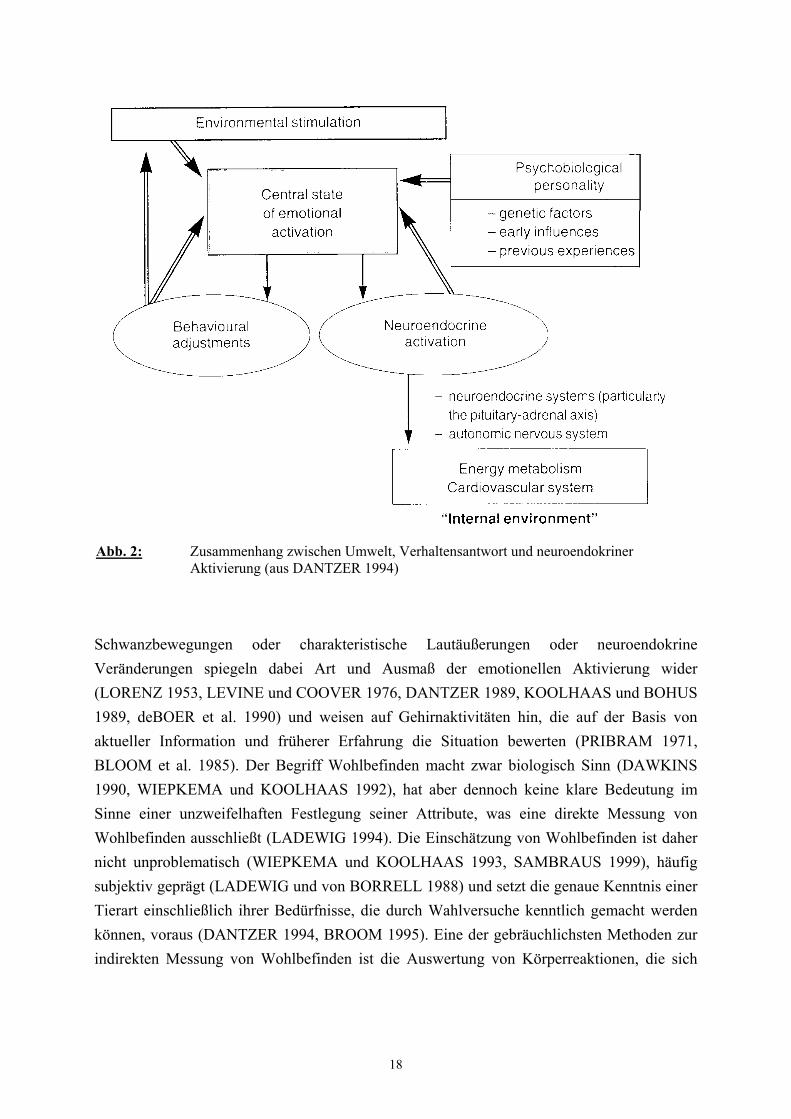
18
Schwanzbewegungen oder charakteristische Lautäußerungen oder neuroendokrine
Veränderungen spiegeln dabei Art und Ausmaß der emotionellen Aktivierung wider
(LORENZ 1953, LEVINE und COOVER 1976, DANTZER 1989, KOOLHAAS und BOHUS
1989, deBOER et al. 1990) und weisen auf Gehirnaktivitäten hin, die auf der Basis von
aktueller Information und früherer Erfahrung die Situation bewerten (PRIBRAM 1971,
BLOOM et al. 1985). Der Begriff Wohlbefinden macht zwar biologisch Sinn (DAWKINS
1990, WIEPKEMA und KOOLHAAS 1992), hat aber dennoch keine klare Bedeutung im
Sinne einer unzweifelhaften Festlegung seiner Attribute, was eine direkte Messung von
Wohlbefinden ausschließt (LADEWIG 1994). Die Einschätzung von Wohlbefinden ist daher
nicht unproblematisch (WIEPKEMA und KOOLHAAS 1993, SAMBRAUS 1999), häufig
subjektiv geprägt (LADEWIG und von BORRELL 1988) und setzt die genaue Kenntnis einer
Tierart einschließlich ihrer Bedürfnisse, die durch Wahlversuche kenntlich gemacht werden
können, voraus (DANTZER 1994, BROOM 1995). Eine der gebräuchlichsten Methoden zur
indirekten Messung von Wohlbefinden ist die Auswertung von Körperreaktionen, die sich
Abb. 2: Zusammenhang zwischen Umwelt, Verhaltensantwort und neuroendokrinerAktivierung (aus DANTZER 1994)

19
aufgrund akuter oder chronischer Belastungen ergeben (FRASER et al. 1975, DANTZER und
MORMEDE 1981, SMIDT et al. 1988, WIEPKEMA und KOOLHAAS 1993, LADEWIG
1994, GREGORY 1998). Neben der Erfassung von objektiv messbaren körperlichen Schäden
(TSCHANZ 1982, 1987) und Leistungsparametern (DANTZER 1994) werden in der Literatur
die Verhaltensebene (van PUTTEN 1982, WECHSLER 1993, TSCHANZ 1995,
BUCHHOLTZ 1996, MENCH und MASON 1997) und die Erhebung physiologischer und
blutgetragener Daten (WIEPKEMA und KOOLHAAS 1993, BROOM 1995, LAMBOOIJ
1995) als geeignet angesehen, empfindliche Indikatoren für übermäßige Belastung und damit
einhergehendes eingeschränktes Wohlbefinden zu liefern. Aufgrund der dynamischen
Beziehung zwischen Stress und Wohlbefinden besteht Einigkeit über die Notwendigkeit, eine
möglichst große Bandbreite von Parametern zu erheben und zur gegenseitigen Interpretation
zu nutzen, wobei insbesondere die Kombination von ethologischen und physiologisch-
biochemischen Daten als sinnvoll erachtet wird (DANTZER und MORMEDE 1981,
LADEWIG und von BORELL 1988, SMIDT et al. 1988, DANTZER 1994, COCKRAM und
MITCHELL 1999). BROOM und JOHNSON (1993) geben eine detaillierte Übersicht über
Indikatoren, die sowohl ungestörtes, als auch eingeschränktes Wohlbefinden anzeigen.
WIEPKEMA und KOOLHAAS (1993) weisen darauf hin, dass die Erfassung der
ethologischen und physiologischen Antwort auf eine Belastung die Prozesse reflektiert, die
der Organismus zur notwendigen Anpassung an eine Situation nutzt, und daher die erhobenen
Parameter immer in ihrem spezifischen Zusammenhang gesehen werden müssen, um
Aussagen bezüglich einer Beeinträchtigung des Wohlbefindens machen zu können.
2.1.2 Die Belastungsreaktion
2.1.2.1 Zentrale Ebene
Während einer Belastungssituation erscheinen im Säugetierorganismus viele verschiedene
neurochemische Reaktionen (OLIVERIO 1987). Durch seine Fähigkeit zur Sekretion von
hypophyseotropen Freisetzungshormonen einerseits und seine Lage als Knotenpunkt eines
ausgedehnten nervalen Systems andererseits gilt der Hypothalamus als der wichtigste zentrale
Integrator und Regulator der Belastungsreaktion (LADEWIG 1994). Durch ein kompliziertes
Netz von Nervenbahnen steht der Hypothalamus sowohl mit höheren Gehirnstrukturen, wie
Neocortex und dem subkortikalen Anteil des Limbischen Systems (Amygdalakomplex und
Hippocampus), in denen Funktionen wie Emotion, Aggression und Motivation angesiedelt
sind (LADEWIG 1994, HUETHER 1996), als auch mit niederen Hirnstrukturen, wie

20
Hirnstamm und Rückenmark, in enger Verbindung. Über afferente Nervenbahnen erhält er
Informationen über exogene und endogene Reize, die nach “Abstimmung” mit den
verschiedenen Strukturen in Reaktionen umgewandelt und über efferente Nervenbahnen bzw.
über Hormone weitergeleitet werden (HENRY und STEPHENS 1977). So führen beim
Auftreten von Stressoren entstehende und zum Hypothalamus geleitete Nervenimpulse zu
einer über die Neurotransmitter Noradrenalin, Gamma-Amino-Buttersäure (GABA),
Acetylcholin, Histamin und Serotonin gesteuerten Sekretion des im paraventrikulären
Nukleus des Hypothalamus gebildeten Peptids Corticotropin-Releasing Hormone (CRH)
(HUETHER 1996). Das CRH wird aus den terminalen Axonen der neurosekretorischen
Zellen des Hypothalamus in ein Pfortadersystem ausgeschüttet, und gelangt so auf kurzem
Blutweg zum Hypophysenvorderlappen (HVL; Adenohypophyse), wo es in den
Proopiomelanocortin (POMC)-Zellen die Freisetzung von ACTH (Adrenocorticotropes
Hormon, Corticotropin), ß-Endorphin und anderen POMC-Derivaten bewirkt (THUN und
SCHWARTZ-PORSCHE 1994). Neben CRH besitzen auch Arginin-Vasopressin (AVP),
Oxytocin und Catecholamine die Fähigkeit, ACTH aus der Hypophyse freizusetzen (RIVIER
und VALE 1983, AXELROD und REISINE 1984). THUN und SCHWARTZ-PORSCHE
(1994) beschreiben unterschiedlich schnell wirksame Feed-back-Systeme, die regulierend in
das Hypothalamus-Hypophysen-System eingreifen: So hemmen peripher freigesetzte
Glucocorticoide über ein “langes” negatives Feed-back sowohl die Ausschüttung des CRH
aus dem Hypothalamus, als auch die des ACTH aus der Hypophyse. Weiterhin wird die
Ausschüttung von CRH über ein “kurzes” negatives Feed-back durch ACTH gehemmt. Durch
ein positives, “ultrakurzes” oder “Autofeed-back-System” kann das CRH seine eigene
Ausschüttung im Streß fördern.
Außer im Hypothalamus wird CRH noch in weiten extrahypothalamischen Bereichen des
ZNS gebildet und wird als ein Schlüsselhormon in der integrierenden Belastungsantwort
bezeichnet (OLIVERIO 1987, CHROUSOS und GOLD 1992, THUN und SCHWARTZ-
PORSCHE 1994). Das Neuropeptid CRH kann an Synapsen des ZNS im Sinne eines
Neurotransmitters wirken, wodurch die Funktion ganzer Neuronenpopulationen modifiziert
und koordiniert und so zum Beispiel Verhaltensreaktionen kontrolliert werden (DÖCKE
1994). Durch seine Wirkung als Comodulator und Feinregulator an Synapsen ist das CRH
auch an der kognitiven Reizverarbeitung in höheren ZNS-Strukturen beteiligt. Weiterhin
spielt es eine Rolle bei der Einbeziehung des zentralen catecholaminergen und des peripheren
sympathischen Nervensystems in die Belastungsantwort (Übersicht bei TOATES 1995).
2.1.2.2 Periphere Ebene

21
2.1.2.2.1 Hypothalamo-Hypophysär-Adrenale Achse
Die Aktivierung der Hypothalamo-Hypophysär-Adrenalen Achse (HHA-Achse) in
physischen und psychischen Belastungssituationen spielte schon früh in der Stressforschung
eine zentrale Rolle (LADEWIG 1994). Die Ausschüttung von Glucocorticoiden (Syn.:
Corticosteroiden) wie Cortisol, Cortison und Corticosteron aus der Zona fasciculata der
Nebennierenrinde (NNR) bildet dabei den letzten Schritt einer neuroendokrinen Kaskade, in
der Hypothalamus, Hypophyse und Nebennierenrinde zu einem geschlossenem Regelkreis
zusammengeschaltet sind (ALLEN et al. 1973). Das durch CRH freigesetzte ACTH ist der
wichtigste physiologische Regulator der Nebennierenrindenfunktion, indem es die
Freisetzung der NNR-Hormone, insbesondere der Glucocorticoide, fördert und außerdem für
die Aufrechterhaltung der NNR-Struktur und die Bereitstellung der
Hormonausgangssubstanzen (Cholesterin) sorgt (AXELROD und REISINE 1984, THUN und
SCHWARTZ-PORSCHE 1994). Da die Nebennierenrinde nicht in der Lage ist, größere
Mengen von Glucocorticoiden zu speichern, muss sie bei vermehrtem Bedarf mit einer
erhöhten Biosynthese reagieren, die außer durch ACTH auch durch den Nervus splanchnicus
ermöglicht wird, wobei als Neurotransmitter das Vasoaktive intestinale Peptid (VIP),
Catecholamine, das Neuropeptid Y sowie CRH fungieren (THUN und SCHWARTZ-
PORSCHE 1994). Die Beteiligung der Catecholamine an der Stimulation der
Glucocorticoidsynthese einerseits, und die Förderung der Freisetzung von Catecholaminen
aus dem Nebennierenmark durch Glucocorticoide andererseits verdeutlicht, dass es auf der
Ebene der Nebennieren zu einer gegnseitigen positiven Beeinflussung der sympathischen und
corticoiden Belastungsantwort kommt (VOIGT 1995).
Neben der basalen Sekretion erscheinen bei Glucocorticoiden, ACTH und CRH
vorübergehend Schübe (Episoden) erhöhter Sekretion (HELLMANN et al. 1970, THUN et al.
1981). Diese episodische Sekretion wird bei Mensch (KRIEGER 1979) und Tier, mit
Ausnahme des Hundes, von einem lichtgebundenen circadianen Rhythmus überlagert, für den
große episodische Schübe mit hoher Frequenz und Amplitude in den frühen Morgenstunden
und kontinuierliche Abnahme der Gipfelkonzentrationen im Tagesverlauf charakteristisch
sind (THUN 1987, LADEWIG und SMIDT 1989, LADEWIG 1994, THUN und
SCHWARTZ-PORSCHE 1994). Die Tagesperiodizität von ACTH und den Glucocorticoiden
ist eng korreliert, während das CRH eine dem Cortisol gegenläufige diurnale Rhythmik
aufweist (GARRICK et al. 1987). Obwohl eine Aktivierung der HHA-Achse in
Belastungssituationen dazu führen kann, dass die ultradianen Schwankungen der

22
Cortisolsekretion überspielt werden, wird Cortisol auch in solchen Situationen in raschen,
kurzen Schüben sezerniert (THUN und SCHWARTZ-PORSCHE 1994). LADEWIG (1994)
weist darauf hin, dass eine zuverlässige Aussage über die Aktivität der Nebennierenrinde in
Ruhe- und Belastungssituationen nur mittels belastungsarmer und frequenter oder
kontinuierlicher Blutentnahme möglich ist.
Durch ihre Wirkung auf den Intermediärstoffwechsel sind Glucocorticoide in der Lage, in
Belastungssituationen für eine vermehrte Energiebereitstellung und längerfristige Erhöhung
des Blutglucosespiegels zu sorgen. Diese der Catecholaminwirkung synergistischen Effekte
resultieren einerseits aus einer gesteigerten hepatischen Gluconeogenese aus glucoplastischen
Aminosäuren, die wiederum aus einem vermehrten Katabolismus von Strukturproteinen
stammen, andererseits aus einer verminderten, der Insulinwirkung entgegengesetzten,
peripheren Glucoseutilisation (DANTZER 1994, THUN und SCHWARTZ-PORSCHE
1994). Eine Hemmung der Glucoseaufnahme in Skeletmuskelzellen, die in die
Belastungsantwort einbezogen sind, findet nicht statt (SAPOLSKY 1994). Durch Förderung
der Wirkung von Catecholaminen wirken Glucocorticoide lipolytisch, wodurch vermehrt
Glycerol und freie Fettsäuren entstehen, die in der Leber wiederum zur Gluconeogenese
genutzt werden können (THUN und SCHWARTZ-PORSCHE 1994). Neben den
metabolischen Effekten haben Glucocorticoide auch eine ausgeprägte antiinflammatorische
Wirkung (LÖSCHER 1994). So wird die Ausschüttung von Histamin und die Freisetzung von
lysosomalen Enzymen durch einen membranstabilisierenden Effekt herabgesetzt, durch
Hemmung der Cyclooxygenase und indirekte Hemmung der Phospholipase A2 die
Prostaglandin- und Leukotriensynthese gehemmt und durch Beeinträchtigung der Bildung
von Zytokinen und der Aktivierung von T-Lymphozyten das lymphatische System
unterdrückt. Die Funktion dieser Wirkungen scheint darin zu liegen, den Organismus in
Belastungssituationen vor einer übermäßigen Abwehrreaktion zu schützen, bzw. diese in
bestimmten Grenzen zu halten (MUNCK et al. 1984, LUMPKIN 1987, LADEWIG 1994).

23
2.1.2.2.2 Sympatho-Adrenomedulläres -System
Ebenso wie für die HHA-Achse wurde auch für das Sympatho-Adrenomedulläre-System
(SAM) schon früh eine stressbedingte Aktivierung nachgewiesen (LADEWIG 1994). Das
SAM stellt eher eine funktionelle Einheit als eine anatomisch definierte Struktur dar
(DANTZER 1994). Es wird durch den sympathischen Zweig des vegetativen Nervensystems
und das Nebennierenmark (NNM), das einer Mischung aus peripherem Ganglion und
Hormondrüse entspricht, repräsentiert. Übergeordnete vegetative Zentren in der Medulla
oblongata und im Zwischenhirn, die mit verschiedenen Strukturen des ZNS in Verbindung
stehen (Hypothalamus, Cortex, Limbisches System), regulieren die Funktion des SAM
(DÖCKE und KEMPER 1994). Die in den Seitenhörnern des Brust- und Lendenmarks
entspringenden präganglionären sympathischen Fasern werden in den Grenzstrang-, Zervikal-
und Mesenterialganglien, in denen die Signalübertragung cholinerg durch Acetylcholin
erfolgt, auf postganglionäre Fasern umgeschaltet, die das Endorgan adrenerg durch
Noradrenalin erregen (PENZLIN 1996). Das Nebennierenmark als spezialisiertes
sympathisches Ganglion wird über präganglionäre sympathische Fasern cholinerg innerviert
(DÖCKE und KEMPER 1994), wobei die elektischen Nervenimpulse in hormonale Signale
umgesetzt werden. Eine Aktivierung des SAM führt zur Ausschüttung von Adrenalin und zu
einem kleineren Teil von Noradrenalin aus den chromaffinen Zellen in den Blutkreislauf. Aus
den varikösen Auftreibungen der postganglionären sympathischen Fasern als Transmitter
sezerniertes Noradrenalin gelangt nur zu einem kleinen Teil in den Blutstrom und entfaltet
Hormonwirkung in der Peripherie (DANTZER 1994). Nach LADEWIG (1994) soll als
Neurotransmitter an Synapsen von sympathisch innervierten Blutgefäßwänden wirksames
Noradrenalin allerdings zu einem erheblichen Anteil ins Blut fließen (spillover) und so den
größten Teil des im Plasma vorhandenen Noradrenalins ausmachen. YAMAGUCHI und
KOPIN (1979) sehen 65 % des sich im Plasma befindlichen Noradrenalins als aus dem NNM
stammend an.
Eine Übersicht über die in den chromaffinen Zellen des NNM und den varikösen
Auftreibungen der sympathischen postganglionären Fasern stattfindende Synthese des
Noradrenalins findet sich bei DÖCKE und KEMPER (1994). Grundsätzlich läuft sie
folgendermaßen ab: Die Synthese des Noradrenalins beginnt mit der Hydroxylierung der
Aminosäure Tyrosin zu 3,4-Dihydroxyphenylalanin (DOPA), dem
geschwindigkeitsbegrenzenden Schritt, der durch das Enzym Tyrosinhydroxylase (TH)
katalysiert wird. Aus DOPA entsteht durch Einwirkung der DOPA-Decarboxylase Dopamin.
Die nächste, durch Dopamin-ß-hydroxylase (DBH) katalysierte Reaktion führt vom Dopamin

24
zum Noradrenalin. Im NNM wird außerdem ein Teil des Noradrenalins durch die
Phenylethanolamin-N-methyltransferase (PNMT) zu Adrenalin umgewandelt. In einer
physischen oder psychischen Belastungssituation werden Catecholamine nicht nur verstärkt
freigesetzt, sondern es kommt durch Aktivitätssteigerung und vermehrte Produktion der
beteiligten Enzyme zu einer deutlichen Erhöhung ihrer Syntheserate (McCARTY et al. 1988).
So führt ein erhöhter Impulsfluss in sympathischen Nervenfasern zu einer Verdreifachung der
Tyrosinhydroxylase- und einer Verdoppelung der Dopamin-ß-hydroxylase Aktivität
(DANTZER 1994). Ebenso erhöht sich die Aktivität und die Syntheserate der PNMT
deutlich, was jedoch nicht nerval, sondern humoral durch Glucocorticoide stimuliert wird.
Die Adrenalinsynthese im NNM steht demnach in enger Beziehung zum HHA-System
(POHORECKY und WURTMAN 1971).
Die Freisetzung der beiden Catecholamine läuft weder unter Ruhe-, noch unter
Belastungsbedingungen parallel (Mc CARTY et al. 1981). Zum Einen ist dies durch die zwei
möglichen Herkünfte des Noradrenalins bedingt: Da Noradrenalin auch von den
sympathischen Nerven an das Blut abgegebeben wird, ist dessen Konzentration im Plasma –
im Gegensatz zur gewöhnlich geringeren Produktion im NNM – stets höher als die des
Adrenalin (DÖCKE und KEMPER 1994). Zum Anderen wird die Ausschüttung auch von der
Art des Stressors beeinflusst. So erhöht sich bei vorwiegend psychischen, mit Angst
verbundenen Belastungen (z.B. die Konfrontation einer Katze mit bellenden Hunden)
besonders die Adrenalinsekretion (CANNON 1915, MASON 1968, LADEWIG 1994).
Aktive Abwehrreaktionen, die mit einer erhöhten Muskelaktivität verbunden sind, führen
dagegen vor allem zu einer Noradrenalinfreisetzung. Das SAM ist insgesamt ein System zur
Überwindung akuter Notfallsituationen (CANNON 1928). Die sympathische Innervation
sorgt in Belastungssituationen für eine sekundenschnelle Aktivierung der Erfolgsorgane. Die
Freisetzung der Catecholamine in den Blutstrom erfolgt ebenfalls schnell. So findet nach
GOLDSTEIN (1987) die Catecholaminausschüttung aus dem NNM schon 1 bis 2 Sekunden
nach Wahrnehmung eines Reizes statt. Während einer Elektrostimulation des Sympathikus
kommt es bei der Ratte zu einem schnellen Anstieg der Plasma-Adrenalinkonzentration, die
innerhalb von 20 Sekunden ihren Höhepunkt erreicht, während der Anstieg der
Noradrenalinkonzentration deutlich langsamer verläuft und nach etwa 3 Minuten zu einem
relativ konstanten Level (steady state) führt (YAMAGUCHI und KOPIN 1979). Neben der
Freisetzung ins Blut erfolgt auch die Elimination der Catecholamine aus dem Blut schnell.
DÖCKE und KEMPER (1994) geben als biologische Halbwertszeit 20 Sekunden bis 10
Minuten an. Nach FERRERIA und VANE (1967) liegt die Halbwertszeit der Catecholamine

25
im Blutplasma des Menschen bei etwa 30 Sekunden. LÖSCHER (1994) gibt sie für alle
Tierarten mit 1 bis 2 Minuten an.
Die Catecholamine üben ihre Wirkungen über spezifische Rezeptoren (Adrenozeptoren) aus.
Prinzipiell wird zwischen α- und β- Rezeptoren unterschieden (AHLQUIST 1948), wobei
jedoch verschiedene Untergruppen bestehen. Die Adrenozeptoren weisen eine
unterschiedliche Empfindlichkeit auf die drei Substanzen Adrenalin, Noradrenalin und
Isoproterenol auf: Noradrenalin wirkt am stärksten auf α-Rezeptoren und Isoproterenol auf β-
Rezeptoren, während Adrenalin eine Mittelstellung einnimmt (DÖCKE und KEMPER 1994).
Abbildung 3 veranschaulicht die wichtigsten Wirkungen der Catecholamine in
Belastungssituationen, die durch eine sekundenschnell erfolgende Kreislaufanpassung und
Energiemobolisation die Reaktionsfähigkeit des Organismus erhöhen (PENZLIN 1996).
Sowohl die kardiovaskulären, als auch die metabolischen Wirkungen der Catecholamine
werden durch Glucocorticoide unterstützt (permissive Wirkung) (DÖCKE und KEMPER
1994).
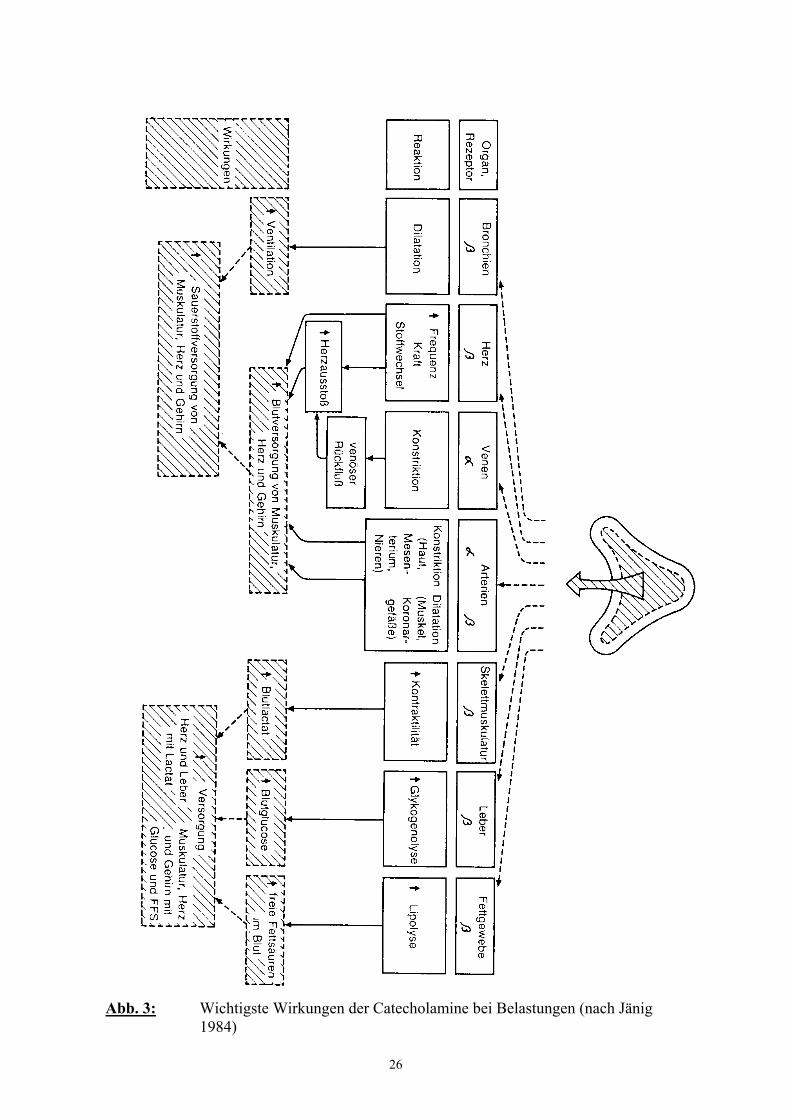
26
Abb. 3: Wichtigste Wirkungen der Catecholamine bei Belastungen (nach Jänig1984)

27
2.2 Belastungsindikatoren beim Schwein
2.2.1 Herzfrequenz
Mit dem Begriff Herzfrequenz wird gemeinhin die Schlagfrequenz des Herzens bezeichnet.
Sie wird in Schlägen pro Minute angegeben und ist mit Herzfrequenzmessern relativ einfach
und genau zu erfassen. Erregt sich ein Tier oder muss es Muskelarbeit verrichten, steigt die
Schlagfrequenz des Herzens an. Die Herzfrequenz ist somit ein geeigneter Parameter, um
Belastungen bei Tieren zu beurteilen (GEERS et al. 1994, BROOM 1995, SCHÜTTE et al.
1995 b, MARAHRENS et al. 1995b). Besondere Vorteile der Herzfrequenzmessung sind die
Möglichkeit der stressarmen Gewinnung durch nichtinvasive Methoden (SCHÜTTE et al.
1995 a) und die sehr kurze Latenzzeit zwischen Belastung und nachfolgender Reaktion
(NIEZGODA et al. 1993).
Die Erregung des Herzens erfolgt normalerweise durch den Sinusknoten als physiologischen
Schrittmacher. Bedingt durch seine Autonomie kann das Herz zwar auch ohne äußere
Nervenversorgung schlagen, doch ist eine Anpassung an einen wechselnden Bedarf des
Organismus an intakte Herznerven gebunden. Die so genannten efferenten Herznerven, Äste
des Nervus vagus und des Sympathikus beeinflussen die Herzfunktion, wobei die
Herzfrequenz durch die zum Sinusknoten verlaufenden Fasern des Nervus vagus vermindert
und durch die des Sympathikus erhöht wird (SELLER 1996). Neben einer auf nervalem Wege
erfolgenden Erhöhung der Schlagfrequenz kommt es in Belastungssituationen durch die
Aktivierung des SAM zusätzlich zu einer humoralen Beeinflussung. Die aus dem
Nebennierenmark freigesetzten Catecholamine Noradrenalin und Adrenalin wirken am
Herzen positiv chrono- und inotrop, das heisst, sie erhöhen die Schlagfrequenz und die
Kontraktionskraft (SLOAN et al. 1996). Eine übergeordnete Regulation dieser Parameter
findet in neocorticalen Strukturen des Gehirns durch unterschiedlich starke Aktivierung des
parasympathischen und sympahischen Nervensystems statt (GALOSY et al. 1979, SHAPIRO
et al. 1993). Während in Belastungssituationen die sympathische Aktivität gesteigert ist
(HENRY et al. 1986), überwiegt bei Beendigung der Situation oder Anpassung an diese
wieder der Einfluß des Nervus vagus, so dass die Herzfrequenz wieder abfällt. Dies kann
auch bei weiterhin erhöhten Catecholaminspiegeln der Fall sein (OBRIST et al. 1974, 1982,
BOHUS et al. 1987). Belastungen, die die Herzfrequenz ansteigen lassen, können physischer,
zum Beispiel körperliche Arbeit, oder psychischer Natur sein. Schon von MICKWITZ und
GRUND (1970) wiesen darauf hin, dass psychische Erregung zu einer Erhöhung der
Herzfrequenz führen kann, die der nach erzwungener körperlicher Belastung gleich kommt.
Unter Umständen können psychische Faktoren dazu führen, dass die Herzfrequenz dauerhaft

28
auf einem erhöhten Niveau bleibt, wie es zum Beispiel bei Tieren in Anbindehaltung im
Vergleich zu solchen, die sich frei bewegen können, der Fall ist (MÜLLER et al. 1988,
SCHOUTEN et al. 1991).
Die Ruheherzfrequenz eines ausgemästeten Schweines mit einem Gewicht zwischen 90 und
110 kg liegt bei etwa 90- 100 Schlägen pro Minute (AUGUSTINI 1976, BICKHARDT
1992). Das Herz-Kreislaufsystem des Hausschweins weist eine besondere Empfindlichkeit
gegenüber Belastungen auf (DOMEL und LEISTNER 1977, SACKMANN 1988, SCHÜTTE
et al. 1994). Die sich unter der modernen Leistungszucht entwickelnde Zunahme des
Körpergewichts bei gleichzeitig sehr hohem Magerfleischanteil hat zu einigen nachteiligen
anatomischen und physiologischen Besonderheiten geführt, zu denen das relativ geringe
Herzgewicht und die funktionelle Insuffizienz zu rechnen sind (von ENGELHARDT 1963,
UNSHELM 1971, STEGEMANN 1984). Belastungen führen bei Schweinen schnell zu
erheblichen Herzfrequenzsteigerungen, die unter Umständen dazu führen, dass das Herz nicht
mehr in der Lage ist, eine ausreichende Sauerstoffversorgung zu gewährleisten. So haben
Untersuchungen gezeigt, dass die Herzfrequenz von Schweinen beim Begehen steiler Rampen
sprunghaft ansteigt und oft einen Bereich von mehr als 200 Schlägen pro Minute erreicht, in
dem die Herzfunktion stark beeinträchtigt ist, weil die Diastolenzeit so verkürzt ist, dass die
Kontraktion der Vorhöfe schon beginnt, bevor die Kammersystole beendet ist (van PUTTEN
und ELSHOF 1978, MAYES und JESSE 1980, LAMBOOIJ et al. 1995). Der Grad der
Herzfrequenzerhöhung ist abhängig von der Steilheit der Rampe (van PUTTEN und ELSHOF
1978, GRANDIN 1982). MAYES und JESSE (1980) stellten fest, dass beim Hinaufgehen,
zum Beispiel einer Laderampe, ein stärkerer Anstieg als beim Hinabgehen erreicht wird.
Entsprechend der offensichtlich großen Belastung, die das Begehen von Rampen für
Schweine darstellt, wurde in einer Reihe von Untersuchungen über den Transport von
Schweinen festgestellt, dass es regelmäßig beim Be- und Entladen zur stärksten Erhöhung der
Herzfrequenz kommt (AUGUSTINI 1976, AUGUSTINI und FISCHER 1982, SCHÜTTE et
al. 1995b, STEFFENS et al. 1995, BARTON-GADE 1996, 1997). MARAHRENS et al.
(1997) untersuchten die Auswirkungen der emotionalen und körperlichen Belastungen, denen
Schweine während des Aufenthalts im Schlachtbetrieb ausgesetzt sind, auf die Herzfrequenz.
Sie registrierten den Verlauf von der Ankunft im Schlachtbetrieb bis zum Einsetzen der
Elektrobetäubung und fanden bei der Zuführung zur Elektrobetäubung mit einer mittleren
Herzfrequenz von 225 Schlägen pro Minute im Vergleich zum Abladen (180 S/min),
Aufenthalt in der Wartebucht (130 S/min) und Umtrieb (155 S/min) die höchsten Werte.
Auch FLOß et al. (1997) stellten in einem Systemvergleich zweier Schlachtbetriebe die
stärkste Erhöhung der Herzfrequenz bei der Zuführung zur Betäubung fest. Jedoch
unterschied sich die Herzfrequenzerhöhung in den verschiedenen Schlachtbetrieben

29
erheblich, was offenbar durch die Art der Zuführung und die baulichen und organisatorischen
Bedingungen beeinflusst wurde.
2.2.2 Körpertemperatur
Neben der Herzfrequenz wird auch die Körpertemperatur als physiologischer Parameter zur
Einschätzung von Belastungen herangezogen (AUGUSTINI 1976, GEERS et al. 1992,
SCHÜTTE et al. 1995b). Mit modernen Methoden ist auch hierbei eine kontinuierliche
Erfassung von Daten möglich, ohne an das Tier herantreten zu müssen (SCHÜTTE et al.
1995a, PARROT et al. 1998).
Die Konstanthaltung der Körpertemperatur in bestimmten physiologischen Grenzen
(Homöothermie) ist ein wesentliches Kennzeichen von Säugetieren und hat im Organismus in
Kompensationssituationen höchste Priorität, wobei wiederum das Hauptbestreben darin liegt,
die Temperatur im ZNS aufrecht zu erhalten (BLIGH 1985). Ermöglicht wird die
Homöothermie durch Thermoregulation. Neben peripheren Thermozeptoren in der Haut
befinden sich im Hypothalamus zentrale Thermozeptoren, die die Kerntemperatur registrieren
und mit einem bestimmten Sollwert vergleichen. Bei Abweichungen kann durch
physiologische und auf der Verhaltensebene erfolgende Reaktionen eine Gegenregulation
erreicht werden. Die erste Strategie ist im allgemeinen die Ausführung eines geeigneten
Verhaltens, wie zum Beispiel das Aufsuchen von Schatten oder windgeschützten Plätzen
(INGRAM und DAUNCEY 1985). In der Regel stehen jedoch landwirtschaftlichen
Nutztieren nur sehr beschränkte Möglichkeiten zur Verfügung, ihr Verhaltensrepertoire zu
nutzen (COCKRAM und MITCHELL 1999), so dass häufig nur die Thermoregulation auf
physiologischer Ebene bleibt. So wird bei Kälte eine verstärkte Wärmebildung unter anderem
durch periphere Vasokonstriktion und durch Zittern, das einem Anheben der Stoffwechselrate
dient (THOMPSON 1977), erreicht. Um einer Überwärmung des Organismus entgegen zu
wirken, wird die Wärmeabgabe über die Haut durch Weitstellen der Gefäße stimuliert.
Weiterhin wird Verdunstung genutzt, wobei Rinder und Pferde vornehmlich schwitzen und
zusätzlich Wärme durch hochfrequente Nasenatmung abgeben, während Schweine sich
bevorzugt suhlen oder, wenn dies nicht möglich ist, bei geöffnetem Maul hecheln
(GREGORY 1996). Insgesamt weist die Thermoregulation des Schweines Schwächen auf
(DOMEL und LEISTNER 1977, HOLZER et al. 1992). So ist die evaporative Wärmeabgabe
beim Schwein im Vergleich mit anderen Tierarten nur unzureichend ausgebildet
(VERHAGEN et al. 1987) und die isolierende Speckschicht erschwert eine erhöhte
Wärmeabgabe zusätzlich. Besonders die Kombination aus hohen Umgebungstemperaturen
und motorischer Aktivität führt beim Schwein zu einer erheblichen Störung des

30
Wärmehaushalts (STEINHARDT 1966, MEYER 1970). Die durchschnittliche
Körpertemperatur von Schweinen mit einem Gewicht von 90-110 kg liegt bei 39°C
(AUGUSTINI 1976). Physische und/oder psychische Belastungen lassen die
Körpertemperatur schnell ansteigen (HONKAVAARA 1989, TRUNKFIELD et al. 1991,
MONIN et al. 1995). So kommt es auf dem Transport von Schweinen im allgemeinen zu
einer Erhöhung der Körpertemperatur um 1°C (BROOM 1995). Bei hohen Ladedichten ist
eine Wärmeabgabe nur sehr eingeschränkt möglich (KLAWITTER 1971, AUGUSTINI und
FISCHER 1982, von MICKWITZ 1982), so dass es zu Temperaturerhöhungen um mehr als
1°C und schlimmstenfalls sogar zum Transporttod kommen kann (TARRANT 1989,
GREGORY 1996). KLONT und LAMBOOIJ (1995a) führen aus, dass nicht nur der
Transport zum, sondern auch die Belastungen im Schlachtbetrieb einen Anstieg der
Körpertemperatur von Schlachtschweinen zur Folge haben können.
2.2.3 Catecholamine
Neben dem Cortisol gelten die Catecholamine als „Stresshormone“ deren Bestimmung im
Plasma als Nachweis der endokrinen Belastungsreaktion eine bedeutende Rolle spielt (SHAW
und TUME 1992, LADEWIG 1994).
Der Catecholamingehalt der Nebennieren weist ebenso wie das Verhälntnis von Adrenalin zu
Noradrenalin starke tierartliche Unterschiede auf. Beim Schwein enthalten die Nebennieren
insgesamt 2150 µg/g an Catecholaminen, wobei der Adrenalinanteil 51 % beträgt
(HOLZBAUER und SHARMAN 1972). Der Catecholamingehalt im Plasma ist stark von der
Blutentnahmemethode abhängig. Ruhewerte können nur unter Verwendung von Kathetern
und unter größtmöglicher Vermeidung von Erregung gewonnen werden (DÖCKE und
KEMPER 1994). Dennoch ist auch dann mit weiteren Beeinflussungen, etwa durch die
Haltungsbedingungen, zu rechnen. So weisen beim Schwein Tiere aus Einzelhaltung höhere
Plasma-Catecholaminwerte auf als Tiere aus Gruppenhaltung (KEMPER 1977, BÜHLER et
al. 1978). Die in der Literatur als Ruhewerte für das Schwein angegebenen
Catecholaminkonzentrationen variieren entsprechend der außerordentlich starken
Abhängigkeit von den Versuchsbedingungen zwischen den einzelnen Untersuchern. DALIN
et al. (1993) fanden bei ovariohysterektomierten Jungsauen, die an den Kontakt mit
Menschen gewöhnt waren, mit 0,18 nmol/l für Adrenalin und 0,9 nmol/l für Noradrenalin
vergleichbar niedrige Werte. TRÖGER und WOLTERSDORF (1988) geben als Ruhewerte
für Schlachtschweine 1,2 nmol/l Adrenalin und 6,6 nmol/l Noradrenalin an. In
Untersuchungen von FORSLID und AUGUSTINSSON (1988) lagen die unter
Ruhebedingungen ermittelten Konzentrationen für Adrenalin und Noradrenalin mit 0,8

31
nmol/l zu 2,2 nmol/l etwa doppelt so hoch wie die von HANNON (1990) und MARAHRENS
et al. (1997) gefundenen Werte. Eine Vielzahl von unterschiedlichen spezifischen Reizen
führt zu einer vermehrten Ausschüttung von Catecholaminen (USDIN et al. 1976, van LOON
et al. 1988). Beim Schwein bedingen zum Beispiel Zwangsmaßnahmen, wie das Fixieren,
einen Anstieg des Catecholamingehalts, besonders des Noradrenalins, im Plasma
(JOHANSSON et al. 1982, DeROTH et al. 1989). DALIN et al. (1993) stellten nach dem
Verladen und während eines einstündigen Transports von Schweinen eine Verdreifachung der
Adrenalinkonzentration im Vergleich zum Ruhewert fest. Der Noradrenalingehalt stieg nach
dem Verladen ebenfalls um etwa das Dreifache des Basalwertes, fiel jedoch unmittelbar
danach wieder bis auf das Ausgangsniveau ab. Auch die Erregung und physische Belastung
von Schweinen unter Schlachtbedingungen führt zu einem erheblichen Anstieg der
Catecholaminkonzentration im Plasma, wobei insbesondere nach elektrischer Betäubung
Werte gefunden wurden, die bis zu 60 (Adrenalin) bzw. 20 mal (Noradrenalin) höher lagen
als die Ruhewerte (TRÖGER und WOLTERSDORF 1988, MARAHRENS et al. 1997).
FORSLID und AUGUSTINSSON (1988) stellten 45 Sekunden nach Kohlendioxidbetäubung
eine Steigerung des Adrenalingehaltes um das fünfzehnfache und des Noradrenalingehaltes
um das fünfzigfache fest und vermuteten als Grund hierfür eine starke respiratorische
Acidose.
2.2.4 Cortisol
Die Bestimmung des Cortisolspiegels im Blutplasma als Ausdruck einer Aktivierung der
HHA-Achse ist eine der gebräuchlichsten Methoden, um Belastungssituationen beim Schwein
zu kennzeichnen (FORSLID und AUGUSTINSSON 1988, NYBERG et al. 1988, WARRIS et
al. 1992, KNOWLES et al. 1998). Cortisol ist bei Mensch, Schwein, Rind und Hund das
wichtigste Glucocorticoid (THUN und SCHWARTZ-PORSCHE 1994) und wird neben
Cortison und Corticosteron in der zona fasciculata der Nebennierenrinde gebildet. Die
Synthese verläuft mit Cholesterol als Ausgangssubstanz über Pregnenolon und Progesteron,
das durch Eingreifen spezifischer Enzyme (Desmolasen, Isomerasen, Dehydrogenasen und
Hydroxylasen) und Coenzyme (NADH, NADPH) schließlich zu den Glucocorticoiden
umgewandelt wird und weiterhin Ausgangssubstanz für die Mineralcorticoide und
Sexualsteroide ist (KARLSON 1988). Im Blut sind mehr als 90 % des Cortisols an
Plasmaproteine gebunden (DALIN et al. 1993). Wichtigstes Bindungsprotein ist das
Corticosteroid-bindende–Globulin (CBG, Transcortin), dessen Bindungskapazität tierartliche
Unterschiede aufweist. Wiederkäuer haben die geringste Bindungskapazität und die
niedrigsten Cortisolkonzentrationen, die sich ähnlich wie bei Hund und Schwein während der

32
Trächtigkeit nicht verändern (LINDNER 1964, THUN 1987). Die episodisch erfolgende
Cortisolsekretion zeigt beim Schwein wie bei den meisten Tierarten eine deutliche, eng mit
dem Hell-Dunkel-Wechsel korrelierte Tagesrhythmik mit stärkeren sekretorischen Schüben in
den frühen Morgenstunden und vergleichsweise schwachen Schüben am Nachmittag und
Abend (BECKER et al. 1985b). So zeigt zum Beispiel ein Schwein aus einer Einzelhaltung
mit Stroheinstreu um 19 Uhr mit 10 ng/ml die niedrigste Cortisolkonzentration im
Tagesverlauf, die bis etwa 23 Uhr auf niedrigem Niveau bleibt. Von diesem Zeitpunkt an
steigt die Amplitude der pulsatilen Schübe bis zum Höchstwert von 65 ng/ ml, der um 6 Uhr
früh erreicht wird (SMIDT et al. 1988). Zahlreiche physische und psychische Stressoren
können zu einem Anstieg des Cortisolspiegels im Blutplasma führen (WEEDING et al. 1993,
SHAW und TROUT 1995). So zeigte sich beispielsweise bei Versuchen auf dem Laufband
(HEINZE und MITCHELL 1989, ZHANG et al. 1992), bei Bewegungseinschränkung
(BECKER et al. 1985a), auf dem Transport (SPENCER et al. 1984, BECKER et al. 1985b,
DALIN et al. 1988, GEVERINK et al. 1995) und bei Hitzestress (KLEMCKE et al. 1989)
eine erhöhte Cortisolsekretion beim Schwein. DANTZER und MORMEDE (1981) verglichen
die Cortisolkonzentration im Blut von Schweinen, die einer neuen Umgebung ausgesetzt
waren, mit der von Schweinen, die einige schmerzhafte Elektroschocks erhalten hatten, und
stellten fest, dass beide Stressoren den Cortisolspiegel im Blut nahezu gleich stark ansteigen
ließen (90 ng/ml zu 80 ng/ml). Dies ist auch ein Beispiel für die ausgeprägte Sensitivität der
HHA-Achse auf psychische Stressoren. LADEWIG und von BORELL (1988) fanden, dass
sich Schweine an nicht schmerzhafte Stressoren wie Isolation und Immobilisation innerhalb
von 14 Tagen gewöhnen können und keine erhöhte Nebennierenreaktion mehr zeigen. Beim
stressinduzierten Anstieg des Cortisolspiegels im Blutplasma zeigen sich beim Schwein
erhebliche individuelle Unterschiede (von BORELL und LADEWIG 1989, STEFFENS et al.
1995). Auch findet eine Beeinflussung durch eine Reihe weiterer Faktoren statt. So ist die
Nebennierenrindenfunktion unter anderem von der Haltung abhängig, wobei Schweine aus
Anbindehaltung in der Regel höhere Cortisolkonzentrationen zeigen als solche aus
Gruppenhaltung (von BORELL und LADEWIG 1986). Auch das Verhalten und der
Sozialstatus wirken sich auf den Grad der Aktivierung der HHA-Achse aus: Aktivere
Schweine haben im Verhältnis niedrigere Cortisolspiegel als Tiere, deren motorische
Aktivität gering ist (von BORELL und LADEWIG 1986, BALDI et al. 1989), und im Plasma
von ranghohen Schweinen findet man geringere Konzentrationen als bei den unterlegenen
Tieren (DANTZER und MORMEDE 1981).

33
2.2.5 Lactat
Lactat stellt unterhalb der regulativen Ebene einen biochemischen Indikator des durch
physische und mit Einschränkung emotionelle Stressoren aktivierbaren Energiestoffwechsels
dar, der den Erfolg oder Verzug der Anpassung des Energiestoffwechsels an die
Belastungssituation anzeigen kann (BICKHARDT und WIRTZ 1986, BICKHARDT 1992,
WARRIS et al. 1994).
Im Blutplasma befindliches Lactat stammt beim Warmblüter vor allem aus dem
Muskelstoffwechsel. Neben der Energiegewinnung durch Spaltung von Kreatinphosphat für
kurzzeitige Höchstleistungen liefert vor allem die Glykolyse die zur Muskelkontraktion
notwendige Energie in Form von ATP (BICKHARDT 1992, GREGORY 1996). Der Abbau
von Glucose läuft bis zur Bildung des Pyruvats unter aeroben und anaeroben Bedingungen
vollkommen gleich ab, jedoch unterscheidet sich das Schicksal des Pyruvats. Steht genügend
Sauerstoff zur Verfügung, wird Pyruvat oxidativ decarboxyliert und das Endprodukt Acetyl-
CoA wird im Citratzyklus verbraucht (KARLSON 1988). Diese Art der Energiegewinnung ist
mit einem Gewinn von 36 Mol ATP pro Mol Glucose sehr effizient (SMIDT et al. 1988) und
für Dauerleistungen unentbehrlich, läuft jedoch vergleichsweise langsam ab und erfordert
Training. Bei ungewohnten Belastungen, aber auch in der Initialaphase von Muskelarbeit
unterhalb der Dauerleistungsgrenze, herrschen im Skelettmuskel anaerobe Bedingungen vor,
und das Pyruvat wird unter diesen Umständen durch die Lactat-Dehydrogenase zu Lactat
reduziert, wobei gleichzeitig eine Oxidation des NADH zu NAD+ stattfindet (KARLSON
1988, KIRSCH 1996). Die Energieausbeute ist dabei mit 2 Mol ATP pro Mol Glucose gering,
und der durch die Lactatanhäufung verursachte pH-Abfall hemmt die zur Muskelkontraktion
nötigen Reaktionen, so dass es zu ATP-Verarmung und Ermüdung kommt (SHAW und
TUME 1992, SMIDT et al. 1988). Die Lactat-Bildung ist eine Sackgasse des Stoffwechsels,
und anfallendes Lactat muss unter erhöhtem Sauerstoff-Verbrauch in der auf eine Belastung
folgenden Ruhephase in Leber und Herz verstoffwechselt werden (KIRSCH 1996). Im
Herzmuskel wird Lactat über Pyruvat in den Citratzyklus eingschleust und unter ATP-
Gewinn oxidiert, während es in der Leber zur Gluconeogenese genutzt wird. Bei anhaltender
Arbeit kommt es zur Verschiebung von Glykogen vom Muskel zur Leber und von dort
während der Erholung in Form von Glucose zurück zum Muskel. Dieses Wechselspiel
zwischen Leber- und Muskelglycogen wird Cori-Zyklus genannt (KARLSON 1988). Die Art
der Energiegewinnung ist auch abhängig vom Muskelfasertyp. Im Allgemeinen wird
zwischen den Fasertypen I, IIA und IIB unterschieden (BROOKE und KAISER 1970), wobei
I auch als rote, IIA als intermediäre und IIB als weiße Fasern bezeichnet werden. In roten
Muskelfasern findet vorwiegend die oxydative Energiegewinnung statt, während in weißen

34
IIB-Fasern die anaerobe Glykolyse besonders ausgeprägt ist. Tiere mit einem hohen Anteil an
weißen Fasern, wie Kaltblutpferd, Mastgeflügel und das Schwein, besitzen eine genetische
Disposition zu Muskelerkrankungen, die mit übermäßiger Produktion von Lactat einhergehen.
Der Musculus longissimus dorsi des Hausschweins besteht bespielsweise zu 70% aus
großkalibrigen weißen IIB-Fasern, während der des Wildschweins diese nur zu 40% aufweist
(PLONAIT und BICKHARDT 1988). Weiße Muskelfasern enthalten wenig Mitochondrien,
in denen die oxidative Energiegewinnung abläuft, und ihr großer Durchmesser erschwert die
Diffusion von Sauerstoff, so dass es beim Schwein in Situationen außergewöhnlicher
physischer und/oder psychischer Belastung zu einer extremen Beschleunigung der
Milchsäureproduktion kommen kann (TRÖGER 1990). Als Ruhewert wird beim Schwein ein
Plasma-Lactatgehalt von 1-2 mmol/l angesehen (JENSEN-WAERN und NYBERG 1993,
BICKHARDT 1996, MARAHRENS et al. 1997, NOWAK 1998). Lactat im Plasma wird
beim Schwein hauptsächlich als ein Indikator für motorische Belastungen angesehen
(BROOM 1996, GRANDIN 1997), jedoch führen auch psychische Stressoren
catecholaminvermittelt zu einer vermehrten Lactatproduktion (SHAW und TUME 1992,
GREGORY 1996). Ungewohnte Belastungen, die einen verstärkten anaeroben Abbau der
Muskelglucose bedingen, sind beim Schwein zum Beispiel Deckakt und Geburt (PLONAIT
und BICKHARDT 1988), der Transport (BECKER et al. 1989), Rangordnungskämpfe
(WARRIS und BROWN 1985, BARTON GADE 1997) und die mit der Schlachtung in
Zusammenhang stehenden Vorgänge (van der WAL 1985, TRÖGER 1990).
2.2.6 Verhalten
Das dem Untersucher durch Beobachtung leicht zugängliche Verhalten eines Tieres wird
neben der Erhebung physiologischer Daten allgemein als besonders geeignet zur
Einschätzung von Belastungssituationen angesehen (van PUTTEN 1982, WECHSLER 1993,
TSCHANZ 1995, BUCHHOLTZ 1996, MENCH und MASON 1997).
Hausschweine sind soziale Tiere (GRAUVOGL 1970, PORZIG und SCHEIBE 1982,
BARTON-GADE 1997), deren wildlebende Verwandte, die europäischen Wildschweine (sus
scrofa), in Gruppen von Tieren unterschiedlichen Alters und Geschlechts zusammenleben
(HOSMAN 1972), wobei eine bestimmte Rangordnung besteht. Diese bleibt besonders in
den Positionen der ranghohen und rangniederen Tiere stabil (EWBANK und MEESE 1971),
und es wird vorwiegend ohne Körperkontakt gedroht statt gekämpft (SAMBRAUS 1979).
Das Hausschwein ist durch die Einwirkung des Menschen immer wieder Situationen
ausgesetzt, in denen Gruppen neu zusammengestellt werden und somit eine Klärung der
Dominanzverhältnisse ansteht. Eine neue Rangordnungsformierung, die mit Kämpfen

35
einhergeht, dauert nach RASMUSSEN et al. (1962) und McBRIDE et al. (1964) einige Tage.
Außer im Herkunftsbetrieb kommt es auch auf dem Transport und in der Wartebucht häufig
zu einem Aufeinandertreffen von einander unbekannten Schweinen. BARTON-GADE und
CHRISTENSEN (1998) beobachteten auf dem Transport bei unterschiedlichen Ladedichten
zwar keine Kämpfe, jedoch bei einer hohen Ladedichte von 0,35 m2/100 kg eine deutlich
erhöhte Reizbarkeit der Schweine, die mit aggressivem Verhalten einherging : Einige Tiere
versuchten mit einer Seitwärtsbewegung des Kopfes andere Schweine wegzuschieben. In
einer Untersuchung von MARAHRENS et al. (1995a) waren 10 % der Tiere einer Gruppe
während des Wartestallaufenthaltes in Kämpfe verwickelt. Insgesamt scheint es in größeren
Gruppen nur wenige aggressive Tiere zu geben, die Kämpfe beginnen und so Unruhe in die
gesamte Gruppe bringen (BALDI et al. 1989, HANSEN et al. 1989, 1991, GEVERINK et al.
1996).
Hausschweine verfügen über ein gutes Gehör und einen feinen Geruchssinn (HOSMAN
1972). Sie können ebenfalls sehr gut sehen, wobei ihr Gesichtfeld 310 Grad beträgt (PRINCE
1977). Diese Eigenschaften versetzen Schweine in hohem Maße in die Lage, Veränderungen
ihrer Umwelt und vielfältige Reize wahrzunehmen. Von einigen Autoren wird in diesem
Zusammenhang die außerordentlich leichte Erregbarkeit von Schweinen hervorgehoben
(HOSMAN 1972, DOMEL und LEISTNER 1977, GRANDIN 1991). Die besondere
Empfindlichkeit dieser Tierart auf jegliche Form von Zwang (HEMSWORTH et al. 1987)
und ihre Eigenschaft, darauf mit lauter Vokalisation zu reagieren, führte Pavlov zu folgender
von LIDDELL et al. (1934) zitierten Aussage: „ Es ist schon lange mein fester Glaube, dass
das Schwein das nervöseste aller Tiere ist. Alle Schweine sind hysterisch.“ Schweine gelten
jedoch auch als lernbegabt und „pfiffig“ (HOSMAN et al. 1972). Eine neue Umgebung wird
von Schweinen obligatorisch intensiv erkundet (WOLTERSDORF und LYHS 1988, LAUBE
und SCHULZE 1992). Das Erkunden ist stets eine eigenmotivierte Handlung (TEMBROCK
1982) und geht mit einem latenten Lernen einher. Es dient der raumzeitlichen Zuordnung des
eigenen Körpers und der Merkmalsextraktion zum Zweck der Identifikation von Objekten
und Vorgängen (TEMBROCK 1969, 1982). Das Explorationsverhalten ist nach van PUTTEN
(1981) eine Verhaltensnotwendigkeit wie die Futteraufnahme oder das Sexualverhalten. Das
Anbieten von Beschäftigungsmaterial wie Stroh oder Spielzeug kann ein in einer reizarmen
Umwelt auftretendes umgerichtetes Explorationsverhalten wie das Schwanzbeißen
eindämmen oder verhindern (van PUTTEN 1980, GRANDIN 1988). Das Erkunden der neuen
Umgebung fordert einen nicht unerheblichen Teil der zumeist mit etwa 2 Stunden
(SACKMANN 1988, STEGEN 1993, von HOLLEBEN 1994) vorgegebenen Aufenthaltszeit
im Wartestall. LAUBE (1995) et al. machen den Versuch, die Erkundungszeit durch das
Anbieten von bevorzugt das Ruhen auslösender Reize zu verkürzen. Es stellte sich heraus,

36
dass Schweine zum Ruhen die Buchtenflächen an der vom Ein- und Ausgang abgelegenen
Wandseite bevorzugen und, dass erwärmte Anteile gegenüber nichterwärmten einen
verstärkten Anreiz zum Niederlegen ausüben. Ebenso können geringe Beleuchtungsstärken
und sichtsperrende Buchtenverkleidungen eine das Ruhen fördernde Wirkung haben (LAUBE
und SCHULZE 1992).
2.3 Die Zuführung zur Betäubung
Von den mit der Schlachtung in Zusammenhang stehenden Vorgängen wird die Zuführung
zur Betäubung als besonders große Belastungsquelle angesehen (TRÖGER und
WOLTERSDORF 1988, HOLZER et al. 1992, von WENZLAWOWICZ et al. 1994,
BARTON-GADE 1997, SCHÄFFER et al. 1997). Die früher in kleineren Schlachtbetrieben
übliche gruppenweise Betäubung der Schweine in einer Bucht, bei der die Tiere quasi mit der
Betäubungszange „eingefangen“ werden mussten, gilt als wenig geeignete Methode zur
möglichst belastungsarmen Behandlung von Schlachtschweinen und wurde von
HOENDERKEN (1979) stark kritisierend als „Wildwest-Methode“ bezeichnet. Größere
moderne Betriebe mit Schlachtzahlen von 400 Schweinen pro Stunde und mehr führten zu
Änderungen in der Schlachttechnologie, die eine kontinuierliche und möglichst schnelle
Zuführung zur Betäubung gewährleisten sollen (TRÖGER 1990). Schlachthöfe nutzen hierfür
eine Vielzahl von Systemen (WEEDING 1993), die häufig nicht tiergerecht sind und von den
Schlachtbetrieben nach eigenen Vorstellungen „verbessert“ werden, um die Schlachtzahlen
einhalten zu können (von WENZLAWOWICZ et al. 1994). Häufig anzutreffen sind
Einzeltreibgänge, in denen die Tiere oft das Weitergehen verweigern und dann nur durch den
massiven Einsatz von „Treibhilfen“, wie Schlagstöcken und/oder Elektrotreibern, zur
Betäubungsfalle gelangen (TRÖGER 1990, HOLZER et al. 1992, LAUBE und SCHULZE
1992). Aber nicht nur das Durchlaufen des Ganges, sondern auch die vorausgehende
Vereinzelung stellt eine erhebliche Belastung für die Tiere dar (BARTON-GADE 1997). Das
Ausmaß der häufig beobachteten, tierschutzrelevanten Probleme bei der Zuführung zur
Betäubung ist neben dem Ausbildungsstand und der Sorgfalt des Personals auch in
entscheidender Weise von der baulichen Gestaltung dieses Abschnitts abhängig (TRÖGER
und WOLTERSDORF 1988, REUTER 1992, WEEDING 1993, HUNTER et al. 1994,
WARRIS et al. 1994, von WENZLAWOWICZ et al. 1994). Es wurde daher immer wieder
gefordert, die Systeme dem Verhalten von Schweinen anzupassen und nicht das Schwein den
Systemen (TARRANT 1989, TRÖGER 1990, LAUBE und SCHULZE 1992, BARTON-
GADE 1997, CHRISTENSEN und BARTON-GADE 1997, SCHÄFFER et al. 1997,

37
GRANDIN 1982). Als Alternative zu Einzeltreibgängen werden zum Beispiel
Zwillingseinzeltreibgänge vorgeschlagen, die nebeneinander liegen und in denen sich die
Tiere sehen können, so dass ein arttypisches Neben- und Hintereinanderherlaufen gefördert
wird (TRÖGER und WOLTERSDORF 1988). TARRANT (1989) beschreibt kurvig gestaltete
Gänge, die die Neugier von Schweinen nutzen sollen, während GRANDIN (1981) in der
Verwendung von kurvigen Gängen für Schweine außer in der Vermeidung von
rechtwinkligen Abschnitten keine Vorteile sieht. Um einem Stau im Eingangsbereich von
Treibgängen entgegen zu wirken, werden sogenannte „stair-step“ –Eingänge
(HOENDERKEN 1976, HUTSON und BUTLER 1978, BRAATHEN 1980) benutzt, die den
Schweinen eine Ausweichmöglichkeit geben und so ein problemloseres Überholen erlauben.
GRANDIN (1982) empfiehlt zur Vermeidung von Engpässen, die Tiere zwischen zwei
Eingängen wählen zu lassen. LAUBE und SCHULZE (1992) schlagen zur Lösung von
Problemen beim Treiben und Vereinzeln von Schweinen einen langen Gang (18 m) vor, in
dem der Tierstrom mit Hilfe von hineinragenden dreieckigen Elementen immer weiter
auseinandergezogen wird. CHRISTENSEN und BARTON GADE (1997) beschreiben zur
Zuführung von Schlachtschweinen zur CO2-Betäubung eine labyrinthartige Konstruktion mit
verschiedenen Sektionen, die mittels beweglicher Wände verkleinert werden können und aus
denen die Tiere dann unter Verwendung von sogenannten „push-hoist-gates“ (Schiebe-Hebe-
Gattern) in die CO2-Gondeln gelangen. Eine allmähliche Verkleinerung des Raumes und/oder
Anpassung an verschieden große Tiergruppen gelten als große Vorteile von beweglichen
Wänden, so dass ihre Verwendung sowohl für Betäubungsbuchten (DAYEN 1989, STEGEN
1993) als auch für Vorwartebuchten (SCHÄFFER et al. 1997) empfohlen wird. Detaillierte
Forderungen zu Gestaltung und Management von Buchten und Treibgängen finden sich bei
GRANDIN (1982).
Zusammenfassung der Literatur
Schweine sind im Schlachtbetrieb erheblichen Belastungen ausgesetzt. Dies trifft in
besonderem Maße für den Abschnitt Zuführung zur Betäubung zu. Eine Vielzahl von
Untersuchungen belegt, dass emotionale und physische Stressoren zu einer Aktivierung der
Hypothalamo-Hypophysär-Adrenalen Achse und des Sympatho-Adrenomedullären Systems
bei Schweinen unter diesen Bedingungen führen. Die Belastungsreaktionen des Organismus
sind über biochemische, physiologische und ethologische Parametern erfassbar. Auf diesem
Wege wird versucht, das Wohlbefinden oder die Stressbelastung eines Tieres einzuschätzen.
Neben Verhaltensbeobachtungen gelten unter anderem die „Stresshormone“ Adrenalin,

38
Noradrenalin und Cortisol, die physiologischen Messgrößen Herzfrequenz und
Körpertemperatur und das Lactat im Blut, als Parameter des Energiehaushalts, als geeignete
Belastungsindikatoren. Es gibt eine Reihe von Untersuchungen über die Belastung von
Schlachtschweinen in unterschiedlich gestalteten Zuführungssystemen. Daher wird auch
empfohlen, bei der Konstruktion von Zuführungssystemen das Verhalten von Schweinen zu
berücksichtigen. Bislang liegen jedoch keine vergleichenden Untersuchungen zwischen einem
herkömmlichen System und einem überwiegend nach ethologischen Gesichtspunkten
entwickelten Zuführungssystem unter Praxisbedingungen vor.

39
3 Eigene Untersuchungen
In der vorliegenden Arbeit werden die Belastungsantworten von Schlachtschweinen in zwei
in ihrer baulichen Gestaltung unterschiedlichen Zuführungssystemen verglichen. Die
Zuführung zur Elektrobetäubung kann in die Belastungszonen Vereinzelung und Zutrieb
unterteilt werden, die jeweils eine bauliche Entsprechung besitzen. Die Vereinzelung
erstreckt sich vom Herantreten des Treibers an ein Tier mit der Absicht, es zu separieren bis
zu dessen vollständigem Eintritt in die Zutriebskonstruktion. Dieser Zeitpunkt markiert
gleichzeitig den Beginn des Zutriebs, der mit dem Eintritt in die Betäubungsfalle endet. Ziel
der Untersuchungen war es, den Einfluss des Zuführungssystems auf die Belastungsantwort
und das Wohlbefinden von Schlachtschweinen herauszuarbeiten, so dass es notwendig war,
die Bedingungen vor der Zuführung zur Betäubung möglichst konstant zu halten. Daher
fanden die Untersuchungen nach Art eines „Vorher-Nachher“-Vergleichs im selben
Schlachtbetrieb statt und gliederten sich in zwei Messperioden: In der ersten fanden die
Versuche in dem zu diesem Zeitpunkt bestehenden konventionellen Zuführungssystem,
dessen Zutrieb als Einzeltreibgang konstruiert war, statt. Nach einem Umbau, bei dem ein
vom Institut für Tierhygiene,Tierschutz und Nutztierethologie der Tierärztlichen Hochschule
Hannover und der Firma Vogt GmbH neu entwickeltes, tierfreundliches Zuführungssystem
eingerichtet wurde, fand die zweite Messperiode statt. Die während der ersten Messperiode
vor dem Umbau bestehende Konzeption von Vereinzelung und Zutrieb wird im folgenden als
altes System (System alt), die im zweiten Abschnitt als neues System (System neu)
bezeichnet.
Für die Beurteilung der Belastungsreaktion der Schweine in den beiden unterschiedlichen
Systemen reicht eine isolierte Betrachtung dieses Abschnitts nicht aus. Saisonal und
verfahrenstechnisch bedingte Unterschiede in der Aufzucht, dem Transport und dem
Wartestallaufenthalt können die Belastungsreaktion in den nachfolgenden
Verfahrensabschnitten beeinflussen und sind daher zur weitestmöglichen Standardisierung in
die Untersuchungen einbezogen. Die Belastungsreaktionen der Schlachtschweine werden in
der vorliegenden Arbeit immer zwischen den Systemen alt und neu verglichen. Im folgenden
werden die Systeme näher beschrieben.
Im alten System wurden von August bis Oktober 1997 in 21 Versuchstagen insgesamt Nalt =
266 Tiere untersucht, im neuen System von April bis Juni 1998 in 20 Versuchstagen Nneu =
249 Tiere.

40
3.1 Material, Methoden und Tiere
3.1.1 Tiere
Die in die Untersuchungen einbezogenen Tiere entstammten sämtlich dem BHZP-
Zuchtprogramm. Es handelte sich sowohl um kastrierte männliche als auch um weibliche
Tiere. Um eine ausreichende Tierzahl für zwei Versuchstage pro Woche sicherzustellen,
wurden die Tiere von zwei Herkunftsbetrieben (Betrieb 1 und Betrieb 2), die in beiden
Messperioden identisch waren, geliefert. Pro Versuchstag wurden jeweils nur Tiere aus einem
Betrieb einbezogen.
Betrieb 1 ist etwa 15 km vom Schlachtbetrieb entfernt. Es handelt sich um ein geschlossenes
System mit 125 Sauen- und 850 Mastplätzen. Die Tiere wurden dort mit 10 Wochen von
Flatdecks in den Maststall umgestallt, wo sie bei einer Gruppengrösse von 13 bis 15 Tieren
auf Vollspaltenböden gehalten wurden. Bis zu einem Gewicht von 40 kg wurde ein
Alleinfuttermittel für Schweine mit einem Rohproteingehalt von 17,5 % gefüttert, danach ein
für die Endmast vorgesehenes Alleinfutter mit einem Rohproteingehalt von 17 %. Die
Fütterung erfolgte jeweils am Breifutterautomaten. Die durchschnittliche Tageszunahme lag
bei 800 g. Nach einer gesamten Mastdauer von 26 Wochen wurden pro Versuchstag jeweils
26 Tiere am Vorabend der Schlachtung ausgestallt und auf eine überdachte Verladerampe mit
Vollspaltenboden verbracht. Von den 26 Tieren wurden nach dem Zufallsprinzip 10 bis 13
ausgewählt und in die Untersuchungen einbezogen.
Betrieb 2 ist ebenfalls etwa 15 km vom Schlachtbetrieb entfernt, und auch hier handelt es
sich um ein geschlossenes System. Der Betrieb verfügt über 90 Sauen- und 600 Mastplätze.
Die Tiere wurden dort von der Abferkelbucht bis zur Endmast dreimal umgestallt. Im Vor-
und Endmastbereich wurden sie auf Vollspaltenböden gehalten. In der Vormast erfolgte eine
ad-libitum-Fütterung mit einem Alleinfutter für Mastschweine mit einem Rohproteingehalt
von 16,5 %, in der Endmast wurde ein Alleinfutter mit einem Rohproteingehalt von 13,5 %
zweimal täglich flüssig verfüttert. Die durchschnittliche Tageszunahme lag bei 750 g. Nach
einer Mastdauer von insgesamt 26 Wochen wurde eine Gruppe von 26 Tieren, in der sich
wiederum 10 bis 13 Versuchstiere befanden, ausgestallt und auf eine in Buchten aufgeteilte
Verladerampe ohne Überdachung verbracht. Die Tiere wurden dort auf vier mit Betonboden
ausgekleidete Buchten verteilt.

41
3.1.2 Transport
Alle Fahrten wurden mit demselben, im Besitz des Schlachtbetriebs stehenden LKW an
denselben Wochentagen durchgeführt. Die Ladefläche war mit Sägemehl ausgestreut und
wurde mittels eines Klappgitters in zwei gleich große Abteile geteilt. Die ebenerdig
erfolgende Beladung des LKW mit den 26 Tieren fand stets zwischen 6.30 und 7 Uhr statt.
Hierzu fuhr der LKW rückwärts direkt an die Verladerampe des jeweiligen Betriebs heran,
und die Tiere wurden mittels eines Brettes auf den LKW getrieben, wobei das Trenngitter
heruntergeklappt wurde, nachdem sich 13 Tiere auf der Ladefläche befanden. Es wurde
darauf geachtet, die Versuchstiere gleichmäßig auf beide Hälften der Ladefläche zu verteilen.
Bei Außentemperaturen von unter +5°C wurden sämtliche Lüftungsklappen des Fahrzeugs
geschlossen, bei Temperaturen über +15°C geöffnet. Die etwa 15 km lange Fahrt von Betrieb
1 zum Schlachtbetrieb verlief in ihrer ersten Hälfte kurvenreich und führte über einen
Feldweg und ein kurzes Stück Landstraße durch einen kleinen Ort, nach dessen Passage sich
ein längeres Stück Landstraße anschloss. Die zweite Hälfte der Fahrt verlief kurvenarm über
die Autobahn und breit ausgebaute Straßen eines Industriegebiets. Die Fahrt von Betrieb 2
zum Schlachtbetrieb verlief insgesamt kurvenarm und führte über eine Landstraße und
ebenfalls über die breit ausgebauten Straßen innerhalb eines Industriegebiets. In beiden Fällen
erreichte der Transport nach etwa zwanzigminütiger Fahrt den Schlachtbetrieb, und nach
einer Rangierzeit von etwa drei Minuten wurde mit dem Entladen begonnen.
3.1.3 Schlachtbetriebsmanagement und bauliche Einrichtungen
Bei dem Schlachtbetrieb handelt es sich um ein einer Erzeugergemeinschaft angeschlossenes
mittelständisches Unternehmen mit einer Schlachtzahl von 130 bis 150 Tieren pro Stunde.
Nach der Ankunft des LKW im Schlachtbetrieb wurde sofort mit dem Entladen begonnen.
Hierzu fuhr der LKW rückwärts direkt an die zur Entladung konzipierte, mit Rolltoren
ausgestattete Seite der Wartehalle, die sich auf gleicher Höhe wie die Ladefläche des
Fahrzeugs befand. Die Gruppe von 26 Tieren gelangte vom LKW über einen 6 m langen
Gang in immer dieselben zwei hintereinanderliegenden Wartebuchten mit den Maßen 3,00 x
2,00 m, die durch eine Metallgittertür getrennt waren. In jede der beiden Buchten wurden 13
Tiere eingetrieben. Es wurde darauf geachtet, die in den Versuch einbezogenen Tiere
gleichmäßig zu verteilen. Seitenwände und Türen der Buchten wiesen dieselben
Konstruktionsmerkmale auf und bestanden aus einer Rohrkonstruktion mit eingebauten,
blickdichten Blechen ab einer Höhe von 10 cm bis etwa 60 cm. Pro Abteil befanden sich zwei

42
Nippeltränken und zwei Duschen in der Wartebucht. Bei Außentemperaturen von über 15°C
wurden die Tiere nach dem Eintrieb über etwa 5 Minuten mit einer feintropfigen
Kaltwasserdusche besprüht. Eine obligate Beduschung erfolgte nach einer Ausruhzeit von
etwa 110 + 5 Minuten. Danach wurde die erste und etwa 15 Minuten später die zweite Gruppe
von 13 Tieren über eine Strecke von etwa 20 Metern in die in der Wartehalle gelegene 6 m²
große Vorvereinzelungsbucht getrieben. Sowohl in der Wartebucht als auch in den
Treibgängen und der Vorvereinzelungsbucht bestand der Fußboden aus Beton. Die sich der
Vorvereinzelungsbucht anschließenden Bereiche der Vereinzelung und des Zutriebs waren im
alten und neuen System sowohl in ihrer baulichen Ausführung, als auch in ihrer Lokalisation
unterschiedlich (Abbildung 4a, 4b und 5) und werden im folgenden näher beschrieben.
Vereinzelungs- bucht
Treibgang Betäubungsfalle
Entbluteband
Rücklaufsperren
Wartestallbereich Schlachthalle
Abb. 4a: Schematische Darstellung des Zuführungssystems vor dem Umbau desSchlachtbetriebs (altes System)
4,40m

43
Im alten System gelangten die Tiere aus der Vorvereinzelungsbucht durch Öffnen eines Tores
in die direkt benachbarte, ebenfalls im Wartestallbereich gelegene 4 m² große
Vereinzelungsbucht. Ihre seitlichen Begrenzugen erreichten eine Höhe von 160 cm und
bestanden aus verkleideten Metallrohren. Der mittels einer abgedunkelten Lampe beleuchtete
Eingang in die Zutriebskonstruktion führte durch die zwischen Schlachthalle und
Wartestallbereich gelegene Wand und wies eine Höhe von 80 cm und eine Breite von 40 cm
auf. Von hier gelangten die Tiere zumeist unter massivem Treibhilfeeinsatz in die
Zutriebskonstruktion. Hierbei handelte es sich um einen aus Metallrohren käfigartig
konstruierten Einzeltreibgang, dessen Seiten bis auf das obere Drittel mit Stahlblech
verkleidet waren. Der Gang war aktiv zu durchlaufen und optisch und akustisch nicht von der
Schlachthalle getrennt. Er war 4,40 m lang und wies zwischen Eintrieb und der am Ende
gelegenen, horizontal angeordneten Betäubungsfalle eine Höhendifferenz von 70 cm auf, was
einer Steigung von 10 ° entspricht. Um durch den Gang zu gelangen, mussten die Schweine
zwei mechanische Rücklaufsperren aufdrücken. Die Betäubungsfalle war durch ein
pneumatisch betriebenes Schott vom übrigen Gang getrennt. Da die Tiere den Einzeltreibgang
zumeist nicht ohne Schwierigkeiten durchliefen, wurde zur kontinuierlichen Beschickung der
Betäubungsanlage häufig eine „Bevorratung“ mit bis zu drei Schweinen im Gang betrieben.
Vorvereinzelungs- bucht
Betäubungsfalle
Entbluteband
Wartestallbereich Schlachthalle
automatischerZutrieb
Abb. 4b: Schematische Darstellung des Zuführungssystems nach dem Umbau des Schlachtbetriebs(neues System)
Metallspiegel
Vereinzelungs-bucht
2,30m
Schalldämmung

44
Im neuen System sind sowohl die Vereinzelungsbucht, als auch die Zutriebskonstruktion
durch Einziehen schallgedämmter Zwischenwände optisch und akustisch von Wartehalle und
Schlachthalle getrennt. Aus der in der Wartehalle gelegenen Vorvereinzelungsbucht gelangten
die Schweine in die 4 m² große Vereinzelungsbucht. Von hier aus wurden die Tiere in die 230
cm lange und horizontal verlaufende Zutriebskonstruktion getrieben, in der sie auf einem
Förderband stehend in die Betäubungsfalle transportiert wurden, sich aber auch aktiv vorwärts
bewegen konnten. Die Zutriebskonstruktion wird durch kunststoffverkleidete Seitewände
begrenzt, ist nach oben offen und wird schattenfrei beleuchtet. Im Abstand von 160 cm zur
Eintrittsöffnung befindet sich ein Metallspiegel mit der Funktion eines vertikal beweglichen
Schotts, der die gesamte Konstruktion in Betäubungsfalle und Eintriebsbereich unterteilt
(Abbildung 4b). Die Schweine wurden bei stehendem Transportband und
heruntergefahrenem Metallspiegel eingetrieben, so dass sie auf ihr Spiegelbild zuliefen. Im
Gegensatz zum alten System war der Schlachtbereich mit Betäubungsfalle und Entbluteband
beim Eintrieb nicht sichtbar. Das Förderband aus rutschfestem Gummimaterial setzte sich
Abb. 5: Als Einzeltreibgang konzipierter Zutrieb des alten Systems (linke Bildseite) undautomatische Zutriebseinrichtung des neuen Systems (rechte Bildseite).

45
nach Abschluss des Betäubungsvorgangs beim vorhergehendem Schwein elektronisch
gesteuert in Bewegung und der Metallspiegel fuhr hoch. Ein Zurücklaufen der Schweine
wurde durch ein fingerartig konstruiertes Absperrgitter verhindert, das mittels eines
rechtwinklig zum Förderband angeordneten Bandes hinter dem Schwein hergefahren wurde.
Das Band zum Transport der Schweine und das zur Bewegung des Absperrgitters wurden in
gleicher Geschwindigkeit betrieben. In der Betäubungsfalle wurde zur Erleichterung des
Ansatzes der Betäubungszange die Bewegungsmöglichkeit der Tiere durch einen Restrainer
eingschränkt. Im Zutrieb befand sich immer nur ein Schwein, eine „Bevorratung“ zur
Aufrechterhaltung eines kontinuierlichen Schlachtablaufs erfolgte in der Vereinzelungsbucht,
sie war konstruktionsbedingt im eigentlichen Zutrieb nicht nötig.
3.1.4 Untersuchungsparameter
3.1.4.1 Erfassung der Klimaverhältnisse
Zur Kennzeichnung der Luftsituation auf den Verladerampen der beiden Herkunftsbetiebe,
während des Transports und des Wartestallaufenthalts wurden Geräte (Rologg® Agents,
Rotronic AG, Bassendorf, Schweiz) zur elektronischen Erfassung und Aufzeichnung von
Temperatur und relativer Feuchte der Luft eingesetzt. Die gespeicherten Daten wurden mit
Hilfe einer Ablesestation an einen Computer übertragen.
3.1.4.2 Herzfrequenz
Zur kontinuierlichen Erfassung der Herzfrequenz (HF) wurden Geräte eingesetzt (Polar
Accurex Plus, Fa. Polar® Electro OY, Kempele, Finnland), die üblicherweise in der
Sportmedizin Verwendung finden. Eine Messeinheit besteht aus einem Sender (Elektroden)
und einem Empfänger (Monitor), der in Form und Größe einer Armbanduhr entspricht. Der
Sender hat eine Reichweite von etwa einem Meter, und zur Signalübertragung zum
Empfänger wird ein bestimmter Code benutzt, so dass Interferenzen mit anderen
Messeinheiten verhindert werden. Die Messeinheiten wurden am Morgen des Schlachttags
auf der Verladerampe des jeweiligen Herkunftsbetriebs je 10 Tieren angelegt. Hierzu wurden
unmittelbar vor dem Anlegen auf die Größe der Elektroden zugeschnittene und mit
Elektrodengel (Kochsalz in Methylcellulose und Propylenglycol) getränkte Schwammtücher
mittels Gummibändern auf den Elektroden befestigt. Die so vorbereiteten Sender wurden

46
anschließend mit Hilfe eines schmalen elastischen Gurtes und eines Karabinerhakens den
Tieren angelegt, wobei sie in linker seitlicher Lage auf Höhe des Herzens zu liegen kamen.
Um sie in dieser Position zu fixieren und vor dem Benagen durch andere Schweine zu
schützen, wurden den Tieren 10 cm breite elastische Brustgurte angelegt. Auf diesen waren
verschließbare Taschen angebracht, in denen sich die Empfänger während des Messvorgangs
befanden. Die Herzfrequenz wurde in Intervallen von fünf Sekunden durchgehend vom
Versuchsbeginn bis zum Einsetzen der Elektrobetäubung aufgezeichnet. Nach Abnahme der
Messeinheiten von den Tieren wurden die im Empfänger gespeicherten Einzelsignale mit
Hilfe einer Ablesestation an einen Computer übertragen, so dass individuelle
Herzfrequenzverläufe grafisch dargestellt werden konnten. Die Qualität der Messergebnisse
variierte allerdings erheblich. Besonders im Bereich der Vereinzelung und des Zutriebs kam
es teilweise zu Messausfällen oder unlogischen Werten, die eliminiert werden mussten. Ein
Grund hierfür war, dass die Schweine hier häufig körperlichen Kontakt mit anderen
Schweinen oder mit Teilen der Buchten- oder Treibgangbegrenzung hatten, und es so zu einer
starken mechanischen Beanspruchung oder sogar zu einem Verlust der Messeinheiten kam.
Weiterhin schien sich das starke elektrische Feld im Bereich der Betäubung negativ auf die
Herzfrequenzmessung auszuwirken. Während des Wartestallaufenthaltes konnte in einigen
Fällen trotz intensiver Beobachtung der Tiere ein gelegentliches Benagen und kurzfristiges
Entfernen der Messeinrichtung nicht verhindert werden. Mit Hilfe von Protokollen über den
zeitlichen Ablauf bis zur Ankunft im Wartestall und der ab dort einsetzenden
Videobeobachtung konnten die Herzfrequenzen den verschiedenen Phasen, Ereignissen und
Belastungszonen zugeordnet werden. Dabei wurden zur Auswertung die Phasen „vor
Verladen“ (vV), „Transport“ (T), „Wartestall“ (W), „Wartestall minimum“ (W min),
„Umtrieb“ (U) und „Zuführung“ (Z) unterschieden. Die vom Anlegen der Messeinheiten auf
der Verladerampe bis zum Beginn des Aufladens gespeicherten Herzfrequenzen wurden als
Abschnitt „vor Verladen“ (vV) zusammengefasst. Die während des Aufladens, des
Transports, des Abladens und des durchschnittlich eine Minute dauernden Eintriebs in die
Wartebucht aufgezeichneten Daten wurden „Transport (T)“ zugeordnet. Die während des
Aufenthaltes in der Wartebucht gespeicherten Herzfrequenzen wurden der Phase
„Wartestall“ (W) zugeordnet. Unmittelbar nach dem Eintrieb in die Wartebucht erfolgten
dort Blutentnahmen (BE) und eine Kontrolle und gegebenenfalls Korrektur der Brustgurte
durch zwei Personen. Innerhalb des Wartestallaufenhaltes zeigten die Schweine eine als
„Wartestall minimum“ (W min) bezeichnete Periode größter Ruhe, die bei jedem einzelnen
Tier über einen Zeitraum von 30 Minuten individuell anhand der Herzfrequenzverlaufskurve
festgelegt wurde. Gegen Ende des Aufenthaltes in der Wartebucht wurden wiederum
Blutentnahmen (BE) durchgeführt. Sobald die Tore der Wartebucht geöffnet wurden,
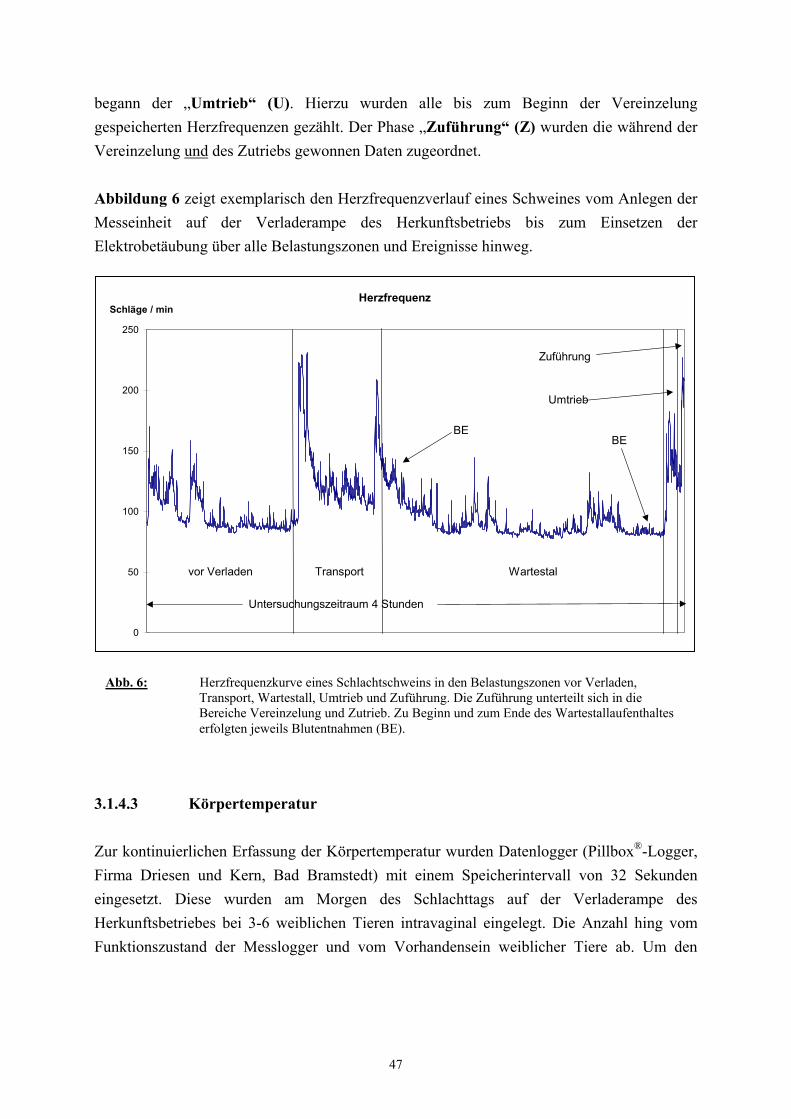
47
begann der „Umtrieb“ (U). Hierzu wurden alle bis zum Beginn der Vereinzelung
gespeicherten Herzfrequenzen gezählt. Der Phase „Zuführung“ (Z) wurden die während der
Vereinzelung und des Zutriebs gewonnen Daten zugeordnet.
Abbildung 6 zeigt exemplarisch den Herzfrequenzverlauf eines Schweines vom Anlegen der
Messeinheit auf der Verladerampe des Herkunftsbetriebs bis zum Einsetzen der
Elektrobetäubung über alle Belastungszonen und Ereignisse hinweg.
3.1.4.3 Körpertemperatur
Zur kontinuierlichen Erfassung der Körpertemperatur wurden Datenlogger (Pillbox®-Logger,
Firma Driesen und Kern, Bad Bramstedt) mit einem Speicherintervall von 32 Sekunden
eingesetzt. Diese wurden am Morgen des Schlachttags auf der Verladerampe des
Herkunftsbetriebes bei 3-6 weiblichen Tieren intravaginal eingelegt. Die Anzahl hing vom
Funktionszustand der Messlogger und vom Vorhandensein weiblicher Tiere ab. Um den
Herzfrequenz
0
50
100
150
200
250
Schläge / min
Transportvor Verladen Wartestal
BEBE
Umtrieb
Zuführung
Untersuchungszeitraum 4 Stunden
Abb. 6: Herzfrequenzkurve eines Schlachtschweins in den Belastungszonen vor Verladen,Transport, Wartestall, Umtrieb und Zuführung. Die Zuführung unterteilt sich in dieBereiche Vereinzelung und Zutrieb. Zu Beginn und zum Ende des Wartestallaufenthalteserfolgten jeweils Blutentnahmen (BE).

48
sicheren Sitz der Temperaturlogger zu gewährleisten, wurden sie mittels Pflaster an
Harnkathetern aus Gummi (20 cm/CH. 22; Fa. Eickemeyer Hannover) befestigt, wobei nach
erfolgter intravaginaler Einlage der Ballon des Harnkatheters mit einer Spritze aufgeblasen
wurde. Die Datenlogger verblieben bis zum Einsetzen der Elektrobetäubung, in einigen Fällen
bis zum Ausweiden, im Tier. Nach Entfernung der Logger wurden die Daten mit Hilfe eines
Interface in einen Computer überspielt, wo sie mit einer Echtzeitskala versehen und grafisch
dargestellt werden konnten. Die Daten wurden entsprechend der Herzfrequenzauswertung den
Phasen „vor Verladen“ (vV), „Transport“ (T), „Wartestall“ (W), „Umtrieb“ (U), und
„Zuführung“ (Z) zugeordnet, wobei die Phase „Zuführung“ (Z) wiederum die während der
Vereinzelung und des Zutriebs gespeicherten Daten enthält. In den Loggern, die erst auf dem
Ausweidetisch entfernt wurden, waren während des Entblute-, Brüh-, und
Entborstungsvorgangs erhobene Daten gespeichert. Diese wurden der Phase „Schlachtband“
(B) zugeordnet.
Abbildung 7 zeigt exemplarisch die Körpertemperaturkurve eines Schweines, vom Einlegen
der Datenlogger auf der Verladerampe des Herkunftsbetriebs bis zur Entfernung des Loggers
auf dem Ausweidetisch.
Körpertemperatur
38,5
38,7
38,9
39,1
39,3
39,5
39,7
39,9
40,1
40,3
40,5
°C
TransportWartestall
Entblute- band
BE BE
Umtrieb
Zuführung
Untersuchungszeitraum 3 Stunden
Abb. 7: Körpertemperaturkurve eines Schlachtschweins in den Zonen Transport, Wartestall,Umtrieb, Zuführung und Entbluteband. Die Zuführung unterteilt sich in die BereicheVereinzelung und Zutrieb. Zu Beginn und zum Ende des Wartestallaufenthaltes erfolgtenjeweils Blutentnahmen (BE).

49
3.1.4.4 Katheterisierung der Schweine
Um fortlaufende, belastungsarme Blutentnahmen für die Untersuchung auf biochemische
Belastungsindikatoren zu ermöglichen, wurden am Vorabend der Schlachtung auf der
Verladerampe des Herkunftsbetriebes jeweils 5 Tiere mit Venenverweilkathetern ausgestattet.
Dabei wurde in Anlehnung an die von GRÜN et al. (1973) und BRÜSSOW et al. (1981)
beschriebene Methode vorgegangen. Die Tiere wurden mittels einer Seil-Oberkieferschlinge
fixiert, die Ohrhaut trocken gereinigt, und die Vena auricularis magna mit einer
Venenverweilkanüle der Stärke 14 G (Vygonüle®, Firma Vygon Medical Produkte, Hamburg)
punktiert. Durch die Venenverweilkanüle wurde anschließend ein 35 cm langer
Venenkatheter (Firma Vygon) eingeführt. Dieser wurde mit 4 ml heparinisierter (150 I.E./ml)
0,9 % Kochsalzlösung gespült und mit einem Schraubverschluss (Firma Vygon) abgedichtet.
Die Fixation des Katheters erfolgte mit Sprühkleber (Leukospray®, Beiersdorf AG,
Hamburg) und wasserfestem Pflaster (Leukoplast® wasserfest, Beiersdorf AG).
3.1.4.5 Gewinnung und Behandlung der Blutproben
Am Morgen des Schlachttages erfolgte vor jeder weiteren Manipulation, wie Anlegen der
Herzfrequenzmessgeräte und Einlegen der Temperaturlogger, die erste Blutentnahme
(Blutprobe 1). Eine Nichtdurchgängigkeit oder ein Verlust konnte dabei bei etwa 10 % der
insgesamt in beiden Messperioden angelegten Venenverweilkatheter festgestellt werden. Bei
der Blutentnahme wurde darauf geachtet, die Tiere möglichst wenig zu beunruhigen, was
durch Maßnahmen, wie minimale Beleuchtung und lärmvermeidendes Verhalten seitens der
Probennehmer, erreicht wurde. So erfolgte die erste Blutentnahme in den meisten Fällen an
liegenden Schweinen. Zunächst wurde dabei der Verschluss des Katheters entfernt und ein 50
cm langer Verlängerungsschlauch (Firma Vygon Medical Produkte, Hamburg) auf den
Katheter aufgeschraubt, um einer Bewegung des Tieres folgen zu können. Mit einer Spritze
wurden zunächst 4 ml Blut aspiriert und verworfen, dann weitere 10 ml Blut aspiriert und in
10 ml-Einmalmonovetten (Fa. Sarstedt, Nümbrecht) gefüllt. Gerinnungshemmende Substanz
in den Monovetten war Kalium-EDTA. Zusätzlich wurden sie unmittelbar vor dem Gebrauch
mit je 0,25 ml EGTA-Glutathion beschickt, um einen vorzeitigen Abbau der sich im Blut
befindlichen Catecholamine zu verhindern. Nach erfolgter Probennahme wurde der Katheter
mit 4 ml heparinisierter Kochsalzlösung gespült, der Verlängerungsschlauch entfernt und der
Verschluss aufgeschraubt. Weitere Blutentnahmen erfolgten in der beschriebenen Weise
unmittelbar nach dem Transport in der Wartebucht des Schlachtbetriebs (Blutprobe 2), in der

50
Wartebucht am Ende der Wartezeit (Blutprobe 3), in der Vereinzelungsbucht nach dem
Umtrieb (Blutprobe 4) und in der Betäubungsfalle nach der Zuführung zur Betäubung
(Blutprobe 5). Im Zeitraum vom Transport bis zur Betäubung fielen noch einmal rund 10 %
der in beiden Messperioden insgesamt angelegten Venenverweilkather wiederum durch
Verlust oder Nichtdurchgängigkeit aus. Von bis zu 10 Tieren pro Untersuchungstag wurden
Stichblutproben gewonnen. Hierzu wurde unmittelbar nach Anbringung des Entblutestichs
austretendes Blut mit einem Plastikbecher aufgefangen und hiervon 10 ml in
Einmalmonovetten gefüllt (Blutprobe 6).
Alle Blutproben (Katheter- und Stichblut) wurden sofort nach ihrer Gewinnung in Eiswasser
gekühlt und nach durchschnittlich 30 Minuten zentrifugiert. Das abzentrifugierte Plasma jeder
10 ml-Blutprobe wurde anschließend in je 4 Eppendorf-Cups (1ml) (Fa. Eppendorf,
Hamburg) pipettiert und noch im Schlachtbetrieb bei –20°C eingefroren. Im Anschluss an den
Transport vom Schlachtbetrieb zum Institut für Tierhygiene,Tierschutz und Nutztierethologie
der Tierärztlichen Hochschule Hannover wurden die Proben bis zur Analyse bei –80°C
gelagert.
3.1.4.6 Biochemische Untersuchungen (Laboruntersuchungen)
Catecholamine
Zur Isolierung der Catecholamine aus den Schweineplasmaproben wurde eine
Flüssigextraktionsmethode angewendet, die von SMEDES et al. (1982) entwickelt und von
PILZ (1998) für die Verwendung kleiner Probenvolumina modifiziert wurde. Die Analysen
fanden im Institut für Neurobiologie der psychiatrischen Klinik der Georg-August-
Universität Göttingen in enger Zusammenarbeit mit der Arbeitsgruppe um Herrn Prof. Hüther
statt.
Von der frisch aufgetauten Probe (Lagerung bei -80 °C) wurden 500 µl in ein 1,5 ml
Eppendorf-Cup gegeben und mit 10 µl internem Standard (Dihydroxybenzylamin, DHBA)
vermischt. Anschließend wurden 150 µl Lösung A und 750 µl Lösung B zugegeben. Die
Proben wurden 2 Minuten von Hand geschüttelt und bei 1200 g für 5 Minuten zentrifugiert.
500 µl des organischen Überstands wurden abgenommen, in ein zweites 1,5 ml
Eppendorfgefäß gegeben und mit 300 µl Lösung C und 60 µl 0,08 M Essigsäure versetzt. Die
Proben wurden nochmals 2 Minuten geschüttelt und anschließend zentrifugiert. Der größte
Teil des organischen Überstands wurde grob entfernt und dem organischen Abfall zugeführt.

51
Von der unteren wässrigen Phase wurden jeweils 40 µl mit einer Hamilton-Spritze
entnommen und in Autosampler-Mikrogefäße überführt, die bis zur automatischen Injektion
bei einer Temperatur von 4 °C und ohne Lichteinfluss in den Autosampler eingelagert
wurden. Das Injektionsvolumen betrug 30 µl Catecholamin-Extrakt, die Retentionszeit je
nach Alter der Säule 13-16 Minuten.
Die Auftrennung der extrahierten Proben erfolgte über
Hochdruckflüssigkeitschromatographie (HPLC) mit anschließender amperometrischer
Detektion. Das HPLC-System bestand aus einem Entgasungsgerät (GT 103), einer
Präzisionspumpe (M 480), einem kühlbaren Injektionsautomat (Autosampler GINA 160),
einem Peltier-Säulenthermostat (STH-585) und einem amperometrischen Detektor (EP 30).
Alle Geräte wurden von der Firma GYNKOTEK, Germering geliefert. Für die
amperometrische Detektion wurde mit Hilfe einer Standard-Potentialreihe ein optimales
Arbeitspotential von 650 mV ermittelt.
Die Trennung erfolgte über eine Reversed-Phase-C 18-Säule (Nucleosil; 3µm; 120 x 3mm),
die von der Fa. BIOMETRA, Göttingen, bezogen wurde. Als Elutionsmittel wurde ein
Citratpuffer folgender Zusammensetzung verwendet:
72 mM Citrat; 36 mM Natriumhydrogenphosphat; 0,255 mM EDTA und 1,39 bis 1,45 mM
Octansulfonsäure als Ionenpaar-Reagenz. Der Puffer enthielt 6% Methanol, sein pH-Wert
betrug 2,41.
Die Trennung erfolgte bei einer Temperatur von 30°C bis 40°C und einer Flussrate von 380
bis 510 µl/min bei einem Druck von 109 bis 155 bar. Durch geringe Veränderungen des
Gehaltes an Ionenpaarreagenz, der Temperatur und der Flussrate konnte eine gute
Trennleistung über 600 bis 800 Proben pro Säule aufrecht erhalten werden.
Zur Berechnung der Catecholaminkonzentration im Schweineplasma wurde täglich mittels
einer Eichreihe die Wiederfindungsrate bestimmt und in die Chromatografie-Software
(Gynkosoft®, Ver. 5.21) zur Kalibrierung der Peaks und der Berechnung des Offsets
eingegeben.
Zur Systemkontrolle wurde nach jeweils 25 Einspritzungen eine Messung mit Standardlösung
durchgeführt. Die Probenaufbereitung wurde über den Zusatz von internem Standard (DHBA)
(10 µl mit Ing Substanz) zu den Plasmaproben kontrolliert. Zur Bestimmung der Intra-und
Interassaypräzision wurden regelmäßig Kontrollplasmen (Aliquots eines selbst hergestellten
Pools aus Schweineplasma) extrahiert und gemessen. Der Intraassay-Variation für Adrenalin
und Noradrenalin betrug 7,6 % bzw. 12,1 % und die Interassay-Variation für Adrenalin und
Noradrenalin 14,2 % und 8,12 %.

52
Verwendete Materialien und Chemikalien:
• Citronensäure-Monohydrat (MERK, Darmstadt)
• Natriumhydrogenphosphat (MERK)
• Titriplex III (MERK)
• 1-Octane-sulfonic-acid (SIGMA, Deisenhofen)
• Tetraoctylamoniumbromid (SIGMA)
• 1-Octanol (SIGMA)
• Methanol (Lichrosolv) (MERK)
• n-Heptane (SIGMA)
• Essigsäure (MERK)
• Diphenylboric Acid 2-Amino-Ethyl Ester (SIGMA)
Standardsubstanzen:
• 3,4-Dihydroxybenzylamine (SIGMA)
• 3-Hydroxytyramine (Dopamin) Hydrochlorid (SIGMA)
• (-)-Arterenol ((-)-Norepinephrine) (SIGMA)
• (-)-Epinephrine (SIGMA)
• H2O bidest.
Cortisol
Die Bestimmung der Cortisolkonzentration im Schweineplasma erfolgte im Labor des
Instituts für Tierzucht und Tierverhalten (Frau Dr. Dr. N. Parvizi) der Forschungsanstalt für
Landwirtschaft in Mariensee. Für die Analyse wurde ein Enzymimmunoassay verwendet. Die
Intraassay-Variation der Cortisolkonzentration der Poolproben (Kontrolle) betrug 8,9 %, die
Variation der Mittelwerte von Poolproben verschiedener Assays (Interassay-Variation) 12,6
% . Die Kreuzreaktionen betrugen für:
Cortisol: 100%
Cortison: 45%
Corticosteron: 15%
Desoxycorticosteron: 8%

53
Progesteron: 8%
Testosteron: 3%
Lactat
Zur Untersuchung des Lactats wurden 500 µl Plasmaprobe mit 1 ml 10%iger
Trichloressigsäure enteiweisst und über 10 Minuten bei 4°C zentrifugiert. Die Bestimmung
erfolgte als enzymatischer UV-Test (STAHLHUT-KLIPP 1975) nach folgendem
Reaktionsschema:
Pyruvat + NADH+ H´ ⇔ Lactat+ NAD´
Die durch Lactat-Dehydrogenase (LDH) katalysierte Reaktion wird mit einem NAD-
Überschuss von rechts nach links durchgeführt, wobei zur vollständigen Umsetzung des
Lactats das gebildete Pyruvat durch Hydrazin abgefangen werden muss. Die aufgrund der
NADH-Bildung erhöhte Absorption bei 340 nm gibt Aufschluss über die ursprüngliche
Lactatkonzentration. Die für die Applikation im COBAS-Mira® Analyseautomaten (Firma
Hoffmann-La Rosche, Prozedur No. 826-UV) erforderlichen Reagenzien wie NAD, LDH,
Glycinpuffer und Standards wurden von der Firma SIGMA (Deisenhofen) bezogen. Die
Intraassay-Variation der Lactatkonzentrationen der Poolproben (Kontrolle) betrug 13,6 %, die
Interassay-Variation 10,7 %.
Die Lactatuntersuchungen wurden in der Klinik für Rinderkrankheiten der Tierärztlichen
Hochschule Hannover (Prof Dr. H. Scholz) durchgeführt.
3.1.4.7 Ethologische Untersuchungen
Zur Registrierung und späteren Auswertung des Verhaltens der Tiere dienten schwarzweisse
Echtzeit-Videoaufnahmen, die mit Hilfe von Sony Hyper HAD Kameras und einem
Panasonic Videorecorder in Verbindung mit einem VT Super Quad HR Bildschirmteiler
erstellt wurden. Zur Aufzeichnung wurden Videokassetten in S-VHS Quälität verwendet. Die
eingesetzten Kameras waren in einem wetterfesten mit einem Heizwiderstand temperierten
Gehäuse untergebracht, wodurch eine Wasserbildung in der Elektronik verhindert werden
soll. Im alten System wurde das Verhalten von 266, im neuen System das von 249 Schweinen
erfasst und ausgewertet. Zur eindeutigen Identifikation wurden die Schweine beim Anlegen

54
der HF-Messgeräte mit Zahlen gekennzeichnet, die mittels dunkelblauen Viehzeichensprays
(Raidex®, Firma Raidex, Dettingen, Erms)) auf Nacken und Rücken aufgebracht wurden.
Im alten und neuen System wurden an jedem Untersuchungstag Aufnahmen im Wartestall, in
der Vereinzelung und im Zutrieb gemacht. Der Umtriebsbereich war in beiden Fällen
aufgrund der baulichen Gegebenheiten nicht komplett mit Kameras einsehbar.
Zur späteren Auswertung wurden im alten und neuen System für die einzelnen
Belastungszonen folgende ethologische Parameter erfasst:
• Im Wartestall: Anteil der Verhaltensweisen Liegen, Sitzen, Stehen und Gehen am
Gesamtverhalten
• In der Vereinzelung: Zeitdauer, Verhaltensweisen, Treibhilfeeinsatz, Anzahl der
Verhaltenswechsel
• Im Zutrieb: Die gleichen Parameter wie in der Vereinzelung
Um eine Aussage bezüglich der körperlichen Belastung der Tiere vor der Zuführung zur
Betäubung machen zu können, kam es bei der Verhaltensuntersuchung im Wartestall vor
allem darauf an festzustellen, wie aktiv die Tiere während dieser Zeit waren.
Hierzu wurde unterschieden, ob die Tiere liegen, sitzen, stehen oder gehen. Dazu wurde das
Verhalten der gesamten Gruppe in Form eines Scan samplings (MARTIN und BATESON
1986) in einem Intervall von 2 Minuten anhand der Videoaufnahmen registriert und
festgestellt, welches Tier welche der 4 unterschiedenen Verhaltensweisen zeigte. Mittels
dieser Daten konnte dann der prozentuale Anteil der einzelnen Verhaltensweisen am
Gesamtverhalten während des Wartestallaufenthaltes für jedes Tier einzeln ermittelt werden.
Der Beginn der Vereinzelung wurde definiert als das Herantreten des Treibers an ein Tier mit
der Intention, es zu separieren und in den Zutrieb zu treiben. In einigen Fällen betrat ein
Schwein ohne Treibereinwirkung den Zutrieb. Hier wurde der Zeitpunkt, zu dem das Schwein
begann, sich eigenmotiviert auf den Eingang des Zutriebs zuzubewegen, als
Vereinzelungsbeginn gewertet. Die Vereinzelung wurde als beendet angesehen, wenn sich das
Schwein mit dem gesamten Körper im jeweiligen Zutrieb befand. Es wurden die Zeitdauern
und Häufigkeiten folgender Verhaltensweisen erhoben:
• Eigenmotiviertes Vorwärtsgehen ohne sichtbaren äußeren Anlass (Vh 1)

55
• Vorwärtsgehen nach Treibhilfeeinsatz (Vh 2)
• Stehen trotz Treibhilfeeinsatz (Vh 3)
• Abdrehen und Ausweichen vor einem äußeren Reiz (Vh 4)
• Rückwärtsbewegung und Rückwärtsdrängen gegen ein Hindernis (Vh 5)
• Stehen ohne Treibdruck betriebstechnisch bedingt. (Vh 6)
Zur Auswertung wurden Verhaltensweisen zusammengefasst, um eine statistische
Bearbeitung zu ermöglichen. So wurden Vh 3, Vh 4 und Vh 5 dem Vermeidungsverhalten
zugeordnet, da sie als im Vereinzelungsprozess nicht angestrebte Verhaltensweisen
angesehen werden können und ihre Ausführung auf eine besondere Belastung der Schweine
hinzuweisen schien. Vh 1 wurde in beiden Systemen so selten ausgeführt, dass es nicht in die
statistische Auswertung gelangte.
In der Vereinzelung wurde weiterhin die Art und die Häufigkeit des Einsatzes von Treibhilfen
untersucht. Dabei erfolgte eine Einteilung nach:
• Treiben mit der Hand ( Th 1)
• Treiben mit beiden Händen oder Knien ( Th 2)
• Treiben unter geringem Stockeinsatz ( Th 3)
• Treiben unter starkem Stockeinsatz ( Th 4)
• Treiben unter Einsatz des Elektrotreibers ( Th 5)
Bei den Treibhilfen Th1-Th4 wurde jede Anwendung auf das beobachtete Tier einzeln
gezählt, beim Einsatz des Elektrotreibers (Th 5) jeder Stromstoss als ein Ereignis erfasst,
wobei eine plötzliche, starke Fluchtbewegung als Indiz für einen erfolgten Stromstoss
angesehen wurde.
Th 1-Th 4 kamen in beiden Systemen, Th 5 dagegen nur im alten System vor.
Da die Belastung eines Tieres durch Treibhilfen nicht nur von der Häufigkeit ihres Einsatzes,
sondern auch von deren Art abhängt, wurden die verschiedenen Treibhilfen unterschiedlich
gewichtet, indem ihnen ein Faktor zugeordnet wurde. Bei der Zuordnung wurde insbeondere
die Intensität der jeweiligen Treibhilfe und der Abstand zum Tier berücksichtigt.
Anschließend wurden die Häufigkeiten der gewichteten Treibhilfen addiert. Der so für jedes
Schwein erhaltene Index kann als ein Maß für die durch den Treibhilfeeinsatz auf ein
Schwein wirkende Belastung angesehen werden (FLOß 1999).

56
Die Aufzählung zeigt die den Treibhilfen zugeordneten Gewichtungsfaktoren und gibt eine
kurze Erklärung für deren Wahl.
• Th1 Faktor 1,0 mittlerer Abstand zum Tier, geringe Belastung
• Th 2 Faktor 1,5 geringer Abstand zum Tier , mittlere Belastung
• Th 3 Faktor 1,0 mittlerer Abstand zum Tier, geringe Belastung
• Th 4 Faktor 3,0 mittlerer Abstand zum Tier, schmerzhaft, große Belastung
• Th 5 Faktor 8,0 mittlerer-geringer Abstand zum Tier, schmerzhaft,
sehr große Belastung
Zwar ist die Wahl der gewählten Faktoren subjektiv, jedoch wurde versucht, durch ständige
aufmerksame Beobachtung der Reaktionen der Tiere eine durchgängige Vergleichbarkeit und
damit relative Objektivität zu gewährleisten.
Neben den einzelnen Verhaltensweisen und dem Treibhilfeeinsatz wurde in der Vereinzelung
auch die Anzahl der Verhaltenswechsel für jedes einzelne Tier erfasst, da auch dies einen
Hinweis auf die Belastung der Tiere gibt. So schienen sowohl bei der direkten Beobachtung,
als auch bei der späteren Auswertung der Videoaufnahmen die Schweine, die ihr Verhalten
häufig wechselten, besonders belastet zu sein. Um die Anzahl der Verhaltenswechsel zu
erhalten, wurden die Häufigkeiten der einzelnen während der Vereinzelung gezeigten
Verhaltensweisen addiert.
In beiden Systemen wurde als Dauer des Zutriebs die Zeitspanne vom Eintritt eines Tieres
mit dem gesamten Körper in die jeweilige Zutriebskonstruktion bis zum Eintritt in die
Betäubungsfalle gewertet.
Für die Untersuchung der Belastung der Tiere wurden im Zutrieb die gleichen ethologischen
Parameter wie in der Vereinzelung herangezogen. Auch gelten hier dieselben Erläuterungen
zu den einzelnen Verhaltensweisen und zum Treibhilfeeinsatz wie in der Vereinzelung.
3.1.5 Statistische Auswertung der Befunde
Die Auswertung der erhobenen Daten erfolgte mit der Version 6.12 des Statistical Analysis
Systems (SAS Institute Inc., Cary, North Carolina, USA). Grundlage der Auswertung waren

57
die Ausführungen von SACHS (1992) und des SAS/STAT User´s Guide (1988). Die
Berechnungen erfolgten im Institut für Biometrie und dem Institut für Tierzucht und
Vererbungsforschung der Tierärztlichen Hochschule Hannover gemäß den Ausführungen von
SCHMIDT (1999).
Die Residuen der Daten aus dem Systemvergleich wurden vor der varianzanalytischen
Berechnung mit der SAS-Prozedur UNIVARIATE (Shapiro-Wilk-Test) auf Normalverteilung
nach Parametern geprüft und als Histogramm dargestellt. Eine angenäherte Normalverteilung
wurde bei Abweichung von W > 0,95 angenommen. Zur Herstellung einer angenäherten
Normalverteilung wurden die Daten von Noradrenalin, Adrenalin und Cortisol sowie die
sämtlicher ethologischer Befunde logarithmiert.
Zur varianzanalytischen Auswertung der physiologischen und ethologischen Daten im
Systemvergleich wurde folgendes gemischtes statistisches Modell verwendet (SAS-Prozedur
GLM, nicht signifikante Interaktionen eliminiert):
Yijkl = µ + systi + betrj + geslk + st(betr syst)lij + betr∙geslik + eijkl
Y = Beobachtungswert des Merkmals
µ = Gesamtmittel
systi = fixer Effekt der Versuchsvariante (Zuführungssystem), i =1 2
betrj = fixer Effekt des Herkunftsbetriebs, j =1 2
geslk = fixer Effekt des Geschlechts der Schweine k =1 2
st(betr syst)lij = zufälliger Effekt des Versuchstags, genestet
innerhalb Herkunftsbetrieb und Zuführungssystem, l =2 42
eijkl = Restvarianz

58
3.2 Befunde
Alle ermittelten Ergebnisse sind als Least Squares Means (LSM) berechnet. Hierbei handelt
es sich um bestmögliche Schätzwerte der wahren Mittelwerte bei Gleichverteilung der
Beobachtungen über alle berücksichtigten Einflussfaktoren. Die Darstellung der Säulengrafik
bezieht sich in allen folgenden Grafiken auf Unterschiede in Bezug auf die Messperiode, das
heisst auf Unterschiede zwischen altem und neuem System. In der unterhalb der Grafik
angeordneten Tabelle ist in der zweiten Zeile (System) die Signifikanz der dargestellten
Unterschiede in den jeweiligen Verfahrensabschnitten aufgeführt, in der ersten die Anzahl der
in beiden Messperioden untersuchten Tiere, bei denen auswertbare Ergebnisse erzielt wurden
(n). Die dritte Zeile (Betrieb) zeigt die Signifikanz der Einflüsse des Herkunftsbetriebes auf
die jeweils dargestellte Belastungsreaktion der Tiere. Dargestellt sind auch
geschlechtsspezifische Unterschiede in der Belastungsreaktion der Schweine (Geschlecht, 4.
Zeile).
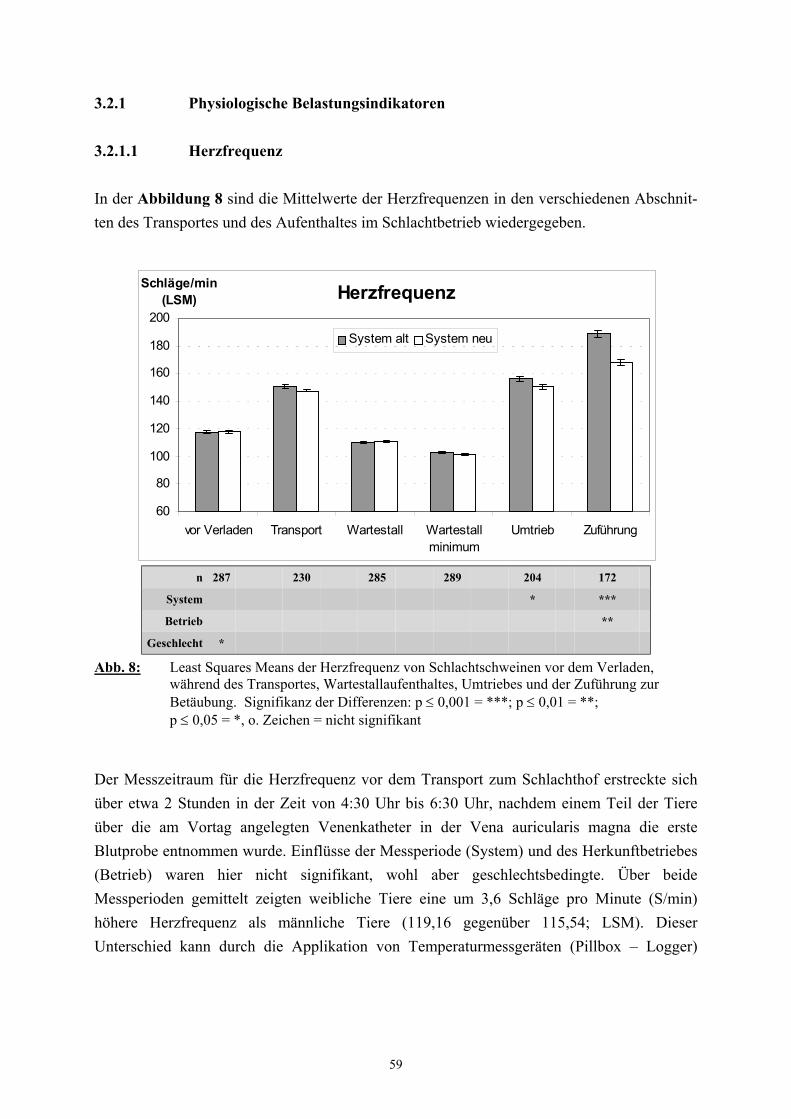
59
3.2.1 Physiologische Belastungsindikatoren
3.2.1.1 Herzfrequenz
In der Abbildung 8 sind die Mittelwerte der Herzfrequenzen in den verschiedenen Abschnit-
ten des Transportes und des Aufenthaltes im Schlachtbetrieb wiedergegeben.
n 287 230 285 289 204 172
System * ***
Betrieb **
Geschlecht *
Abb. 8: Least Squares Means der Herzfrequenz von Schlachtschweinen vor dem Verladen,während des Transportes, Wartestallaufenthaltes, Umtriebes und der Zuführung zurBetäubung. Signifikanz der Differenzen: p ≤ 0,001 = ***; p ≤ 0,01 = **; p ≤ 0,05 = *, o. Zeichen = nicht signifikant
Der Messzeitraum für die Herzfrequenz vor dem Transport zum Schlachthof erstreckte sich
über etwa 2 Stunden in der Zeit von 4:30 Uhr bis 6:30 Uhr, nachdem einem Teil der Tiere
über die am Vortag angelegten Venenkatheter in der Vena auricularis magna die erste
Blutprobe entnommen wurde. Einflüsse der Messperiode (System) und des Herkunftbetriebes
(Betrieb) waren hier nicht signifikant, wohl aber geschlechtsbedingte. Über beide
Messperioden gemittelt zeigten weibliche Tiere eine um 3,6 Schläge pro Minute (S/min)
höhere Herzfrequenz als männliche Tiere (119,16 gegenüber 115,54; LSM). Dieser
Unterschied kann durch die Applikation von Temperaturmessgeräten (Pillbox – Logger)
Herzfrequenz
60
80
100
120
140
160
180
200
vor Verladen Transport Wartestall Wartestallminimum
Umtrieb Zuführung
Schläge/min(LSM)
System alt System neu

60
bedingt sein, die im angegebenen Messzeitraum ausschließlich bei weiblichen Tieren
intravaginal erfolgte. In späteren Verfahrensabschnitten ist dieser Einfluss nicht mehr
vorhanden.
Mit 117 Schlägen pro Minute (Least squares means - LSM) ist die mittlere Herzfrequenz vor
der Verladung („vor Verladen“) der Tiere in die Transportfahrzeuge nicht als Ruhewert
anzusehen, der bei Schweinen bei einem Gewicht von etwa 100 kg mit etwa 80 bis 100
Schlägen pro Minute angegeben wird. Bedingt durch verschiedene Manipulationen an den
Tieren (Anlage der Messgurte, Applikation von Pillbox – Loggern) kam es zu
Aktivitätssteigerungen der Tiere, die sich in diesem Verfahrensabschnitt in erhöhten
Herzfrequenzen niederschlugen. Während des zweistündigen Wartestallaufenthaltes
(„Wartestall“) lagen die mittleren Herzfrequenzen mit 109,8 S/min im alten und 110,4 S/min
im neuen System niedriger als noch im Herkunftsbetrieb vor dem Verladen. Innerhalb des
Wartestallaufenthaltes zeigten die Schweine in einer Periode größter Ruhe, die bei jedem
einzelnen Tier individuell über einen Zeitraum von 30 Minuten festgelegt wurde („Wartestall
minimum“), Herzfrequenzen von im Mittel 102,5 S/min im alten und 101,2 S/min (LSM) im
neuen System.
Durch den Transport („Transport“), der über Kreisstraßen und durch mehrere kleine
Ortschaften führte und etwa 30 Minuten dauerte, stieg die mittlere HF von etwa 117 S/min in
beiden Messperioden nahezu gleich auf etwa 150 S/min an. Im Messzeitraum sind das Be-
und Entladen der Tiere enthalten, wodurch Spitzenwerte von bis zu 190 S/min erreicht
wurden.
Der Umtrieb erfolgte im Schlachthof vom Wartestall bis zur Vorgruppenbucht über eine
Strecke von etwa 20 Metern. Im Umtriebsweg sind 2 rechte Winkel vorhanden, die von den
Tieren zu passieren sind. Die mittleren Herzfrequenzen liegen in der ersten Messperiode (altes
System) bei 156,4 S/min, in der zweiten (neues System) bei 150,7 S/min. Der Unterschied ist
signifikant (p = 0,03).
Während der Zuführung zeigt sich ein hochsignifikanter Unterschied in der
Kreislaufbelastung der Tiere zwischen dem alten und dem neuen System. Die HF sinkt von
im Mittel 189,1 S/min im alten System mit Käfigtreibgang auf im Mittel 168,4 S/min im
neuen System (p < 0,001). Über beide Messperioden gemittelt zeigt sich hier ein Unterschied
in der Kreislaufreaktion der Tiere, der durch ihre Herkunft bedingt ist. Schweine aus Betrieb 1
zeigen eine mittlere HF von 183,4 S/min, während Tiere aus Betrieb 2 im Zutrieb eine
mittlere HF von nur 174 S/min aufweisen (p = 0,007).
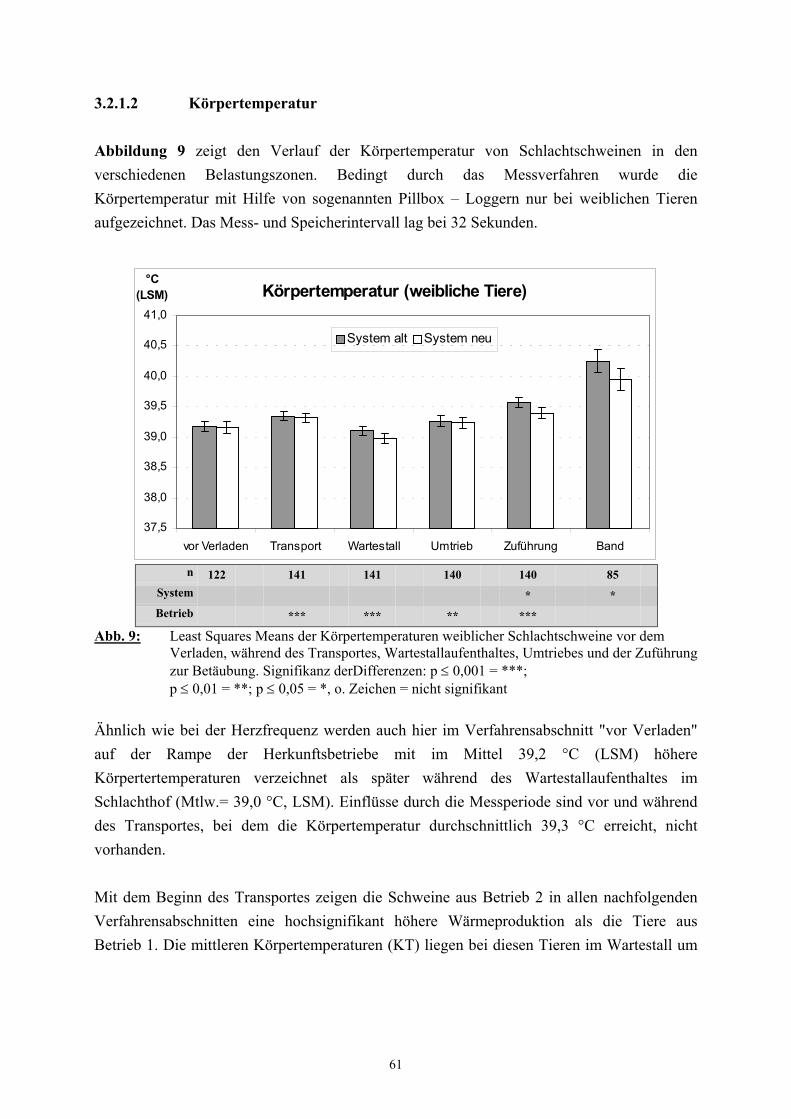
61
3.2.1.2 Körpertemperatur
Abbildung 9 zeigt den Verlauf der Körpertemperatur von Schlachtschweinen in den
verschiedenen Belastungszonen. Bedingt durch das Messverfahren wurde die
Körpertemperatur mit Hilfe von sogenannten Pillbox – Loggern nur bei weiblichen Tieren
aufgezeichnet. Das Mess- und Speicherintervall lag bei 32 Sekunden.
n 122 141 141 140 140 85
System * *
Betrieb *** *** ** ***
Abb. 9: Least Squares Means der Körpertemperaturen weiblicher Schlachtschweine vor demVerladen, während des Transportes, Wartestallaufenthaltes, Umtriebes und der Zuführungzur Betäubung. Signifikanz derDifferenzen: p ≤ 0,001 = ***; p ≤ 0,01 = **; p ≤ 0,05 = *, o. Zeichen = nicht signifikant
Ähnlich wie bei der Herzfrequenz werden auch hier im Verfahrensabschnitt "vor Verladen"
auf der Rampe der Herkunftsbetriebe mit im Mittel 39,2 °C (LSM) höhere
Körpertertemperaturen verzeichnet als später während des Wartestallaufenthaltes im
Schlachthof (Mtlw.= 39,0 °C, LSM). Einflüsse durch die Messperiode sind vor und während
des Transportes, bei dem die Körpertemperatur durchschnittlich 39,3 °C erreicht, nicht
vorhanden.
Mit dem Beginn des Transportes zeigen die Schweine aus Betrieb 2 in allen nachfolgenden
Verfahrensabschnitten eine hochsignifikant höhere Wärmeproduktion als die Tiere aus
Betrieb 1. Die mittleren Körpertemperaturen (KT) liegen bei diesen Tieren im Wartestall um
Körpertemperatur (weibliche Tiere)
37,5
38,0
38,5
39,0
39,5
40,0
40,5
41,0
vor Verladen Transport Wartestall Umtrieb Zuführung Band
°C(LSM)
System alt System neu

62
0,5 °C (39,3° vs. 38,8 °C, LSM), während des Umtriebes um 0,4 °C (39,5° vs. 39,1°C) und
der Zuführung um 0,6 °C (39,8°C vs. 39,2°C) höher. Eine weitere Zunahme des
Unterschiedes ist bis zur Entnahme der Aufzeichnungsgeräte am Schlachtband zu
verzeichnen. Bis zu diesem Zeitpunkt beträgt der Unterschied 0,7 °C (40,5°C vs. 39,8°C). Die
Ursache dieser Reaktionsunterschiede ist nicht bekannt. Es sollte jedoch untersucht werden,
ob sich eine unterschiedliche Gewöhnung der Tiere an Umgebungswechsel und Umtriebe in
Stoffwechsel und Wärmeproduktion niederschlägt.
Der Anstieg der Körpertemperatur weiblicher Schlachtschweine erfolgt während der
Zuführung im neuen System in signifikant geringerem Maße als im alten (39,4° vs. 39,6°).
Besonders deutlich wird dieser Unterschied nach Betäubung und Tötung der Tiere während
der Zeitspanne bis zum Erreichen des Schlachtbandes. Die Körpertemperatur der Schweine,
die im neuen Zuführungssystem zur Betäubung gelangten, ist hier im Mittel um 0,4 °C (39,9°
vs. 40,3°; LSM) geringer, was auf eine Verringerung des postmortalen Glycogenabbaus und
damit einer stoffwechselbedingten Wärmeproduktion hindeutet.
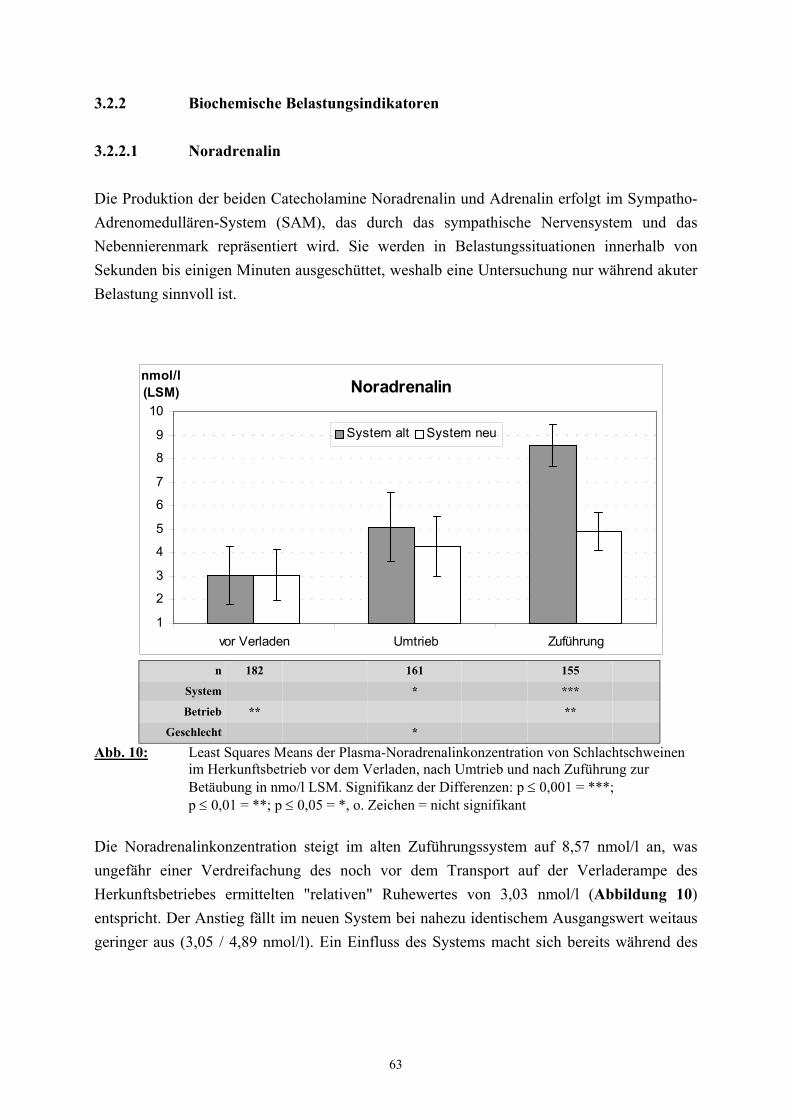
63
3.2.2 Biochemische Belastungsindikatoren
3.2.2.1 Noradrenalin
Die Produktion der beiden Catecholamine Noradrenalin und Adrenalin erfolgt im Sympatho-
Adrenomedullären-System (SAM), das durch das sympathische Nervensystem und das
Nebennierenmark repräsentiert wird. Sie werden in Belastungssituationen innerhalb von
Sekunden bis einigen Minuten ausgeschüttet, weshalb eine Untersuchung nur während akuter
Belastung sinnvoll ist.
n 182 161 155
System * ***
Betrieb ** **
Geschlecht *
Abb. 10: Least Squares Means der Plasma-Noradrenalinkonzentration von Schlachtschweinenim Herkunftsbetrieb vor dem Verladen, nach Umtrieb und nach Zuführung zurBetäubung in nmo/l LSM. Signifikanz der Differenzen: p ≤ 0,001 = ***; p ≤ 0,01 = **; p ≤ 0,05 = *, o. Zeichen = nicht signifikant
Die Noradrenalinkonzentration steigt im alten Zuführungssystem auf 8,57 nmol/l an, was
ungefähr einer Verdreifachung des noch vor dem Transport auf der Verladerampe des
Herkunftsbetriebes ermittelten "relativen" Ruhewertes von 3,03 nmol/l (Abbildung 10)
entspricht. Der Anstieg fällt im neuen System bei nahezu identischem Ausgangswert weitaus
geringer aus (3,05 / 4,89 nmol/l). Ein Einfluss des Systems macht sich bereits während des
Noradrenalin
1
2
3
4
5
6
7
8
9
10
vor Verladen Umtrieb Zuführung
nmol/l(LSM)
System alt System neu

64
Umtriebes bemerkbar: Im alten System steigt die Plasma-Noradrenalinkonzentration auf 5,09
nmol/l, im neuen auf 4,27 nmol/l.
Über alle Tiere beider Herkünfte gemittelt beträgt die NA-Konzentration auf der Rampe der
Herkunftsbetriebe in beiden Versuchsabschnitten 3,03 nmol/l LSM. Es gibt hier jedoch
deutliche Unterschiede bei den beiden Herkünften: Schweine aus Betrieb 1 zeigen
Plasmaspiegel von im Mittel 2,7 nmo/l, während Tiere aus Betrieb 2 im Mittel 3,4 nmol/l
aufweisen (p = 0,002). Diese Unterschiede in der Noradrenalinausschüttung sind auch
während der Zuführung feststellbar, in der Schweine aus Betrieb 1 im Mittel 6,1 nmol/l
gegenüber 7,4 nmol/l bei Tieren aus Betrieb 2 aufweisen (p = 0,01).
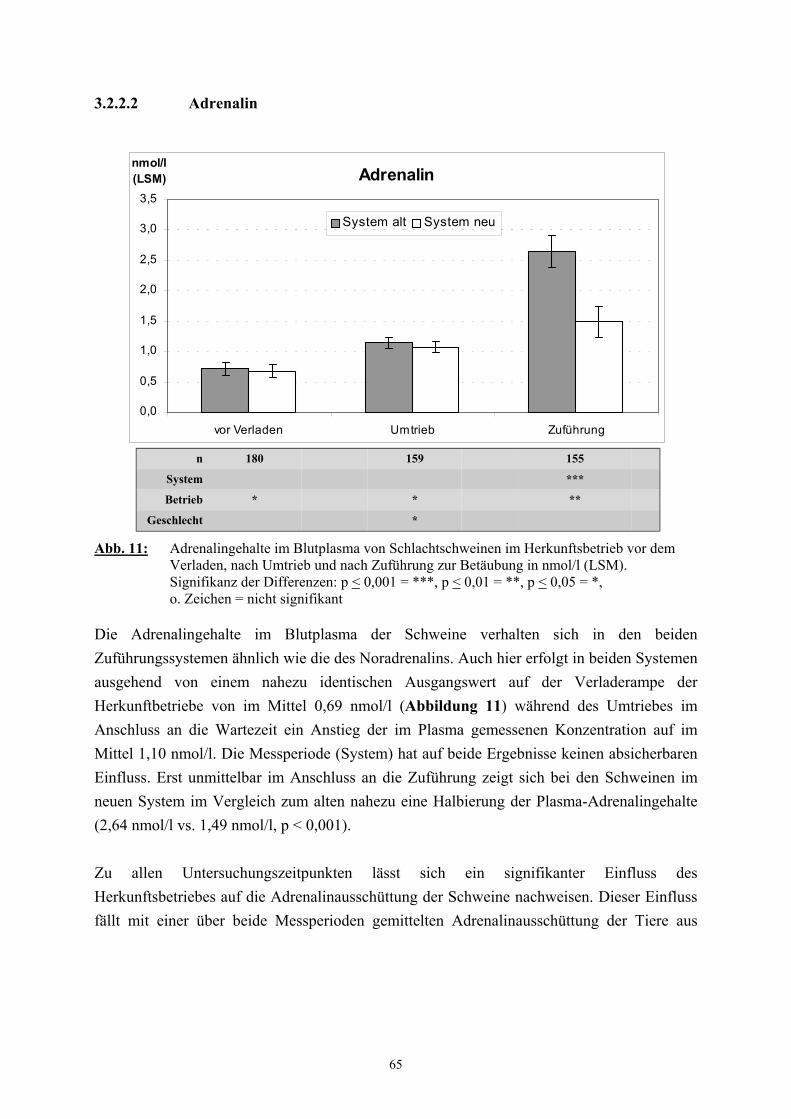
65
3.2.2.2 Adrenalin
n 180 159 155
System ***
Betrieb * * **
Geschlecht *
Abb. 11: Adrenalingehalte im Blutplasma von Schlachtschweinen im Herkunftsbetrieb vor demVerladen, nach Umtrieb und nach Zuführung zur Betäubung in nmol/l (LSM).Signifikanz der Differenzen: p < 0,001 = ***, p < 0,01 = **, p < 0,05 = *, o. Zeichen = nicht signifikant
Die Adrenalingehalte im Blutplasma der Schweine verhalten sich in den beiden
Zuführungssystemen ähnlich wie die des Noradrenalins. Auch hier erfolgt in beiden Systemen
ausgehend von einem nahezu identischen Ausgangswert auf der Verladerampe der
Herkunftbetriebe von im Mittel 0,69 nmol/l (Abbildung 11) während des Umtriebes im
Anschluss an die Wartezeit ein Anstieg der im Plasma gemessenen Konzentration auf im
Mittel 1,10 nmol/l. Die Messperiode (System) hat auf beide Ergebnisse keinen absicherbaren
Einfluss. Erst unmittelbar im Anschluss an die Zuführung zeigt sich bei den Schweinen im
neuen System im Vergleich zum alten nahezu eine Halbierung der Plasma-Adrenalingehalte
(2,64 nmol/l vs. 1,49 nmol/l, p < 0,001).
Zu allen Untersuchungszeitpunkten lässt sich ein signifikanter Einfluss des
Herkunftsbetriebes auf die Adrenalinausschüttung der Schweine nachweisen. Dieser Einfluss
fällt mit einer über beide Messperioden gemittelten Adrenalinausschüttung der Tiere aus
Adrenalin
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
vor Verladen Umtrieb Zuführung
nmol/l(LSM)
System alt System neu

66
Betrieb 2 zum Zeitpunkt des Zutriebes mit 2,43 nmol/l gegenüber den Tieren aus Betrieb 1
mit im Mittel 1,70 nmol/l besonders groß aus.
Die Freisetzung der beiden „Stresshormone“ Noradrenalin und Adrenalin verdeutlicht den
großen Einfluss der Zuführungstechnik auf die Schweine in diesem sensiblen
Verfahrensabschnitt des Schlachtablaufes.
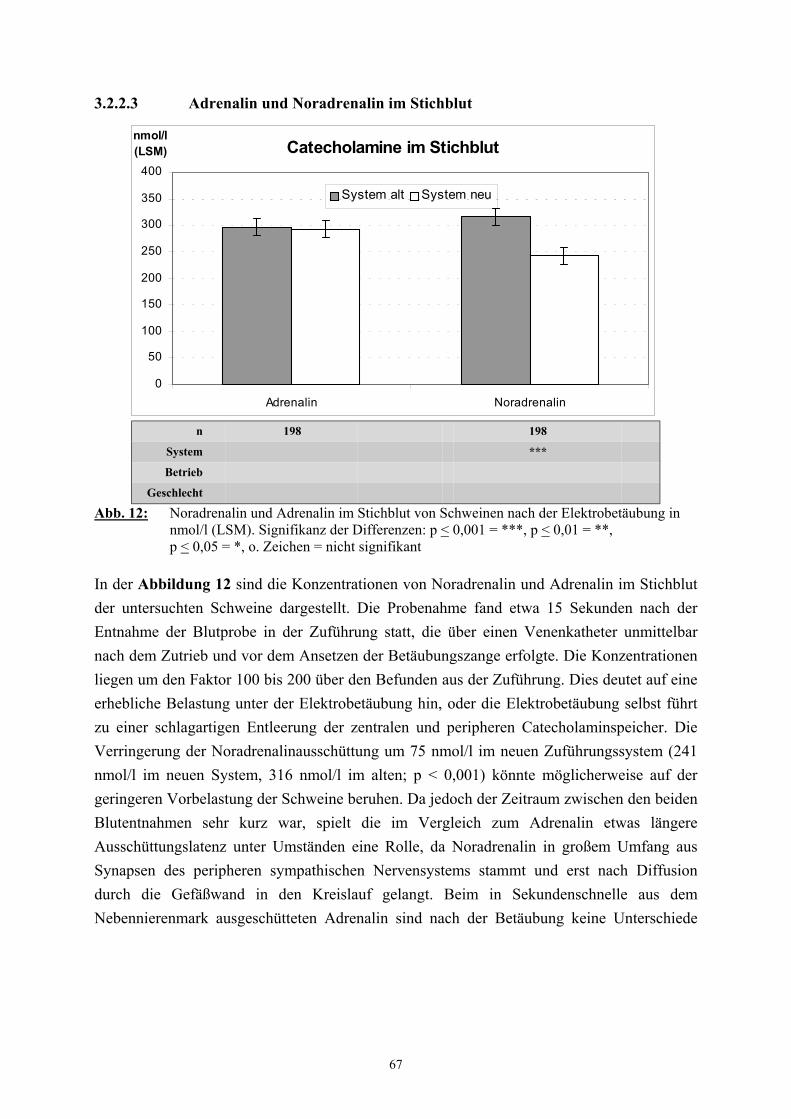
67
3.2.2.3 Adrenalin und Noradrenalin im Stichblut
n 198 198
System ***
Betrieb
Geschlecht
Abb. 12: Noradrenalin und Adrenalin im Stichblut von Schweinen nach der Elektrobetäubung innmol/l (LSM). Signifikanz der Differenzen: p < 0,001 = ***, p < 0,01 = **, p < 0,05 = *, o. Zeichen = nicht signifikant
In der Abbildung 12 sind die Konzentrationen von Noradrenalin und Adrenalin im Stichblut
der untersuchten Schweine dargestellt. Die Probenahme fand etwa 15 Sekunden nach der
Entnahme der Blutprobe in der Zuführung statt, die über einen Venenkatheter unmittelbar
nach dem Zutrieb und vor dem Ansetzen der Betäubungszange erfolgte. Die Konzentrationen
liegen um den Faktor 100 bis 200 über den Befunden aus der Zuführung. Dies deutet auf eine
erhebliche Belastung unter der Elektrobetäubung hin, oder die Elektrobetäubung selbst führt
zu einer schlagartigen Entleerung der zentralen und peripheren Catecholaminspeicher. Die
Verringerung der Noradrenalinausschüttung um 75 nmol/l im neuen Zuführungssystem (241
nmol/l im neuen System, 316 nmol/l im alten; p < 0,001) könnte möglicherweise auf der
geringeren Vorbelastung der Schweine beruhen. Da jedoch der Zeitraum zwischen den beiden
Blutentnahmen sehr kurz war, spielt die im Vergleich zum Adrenalin etwas längere
Ausschüttungslatenz unter Umständen eine Rolle, da Noradrenalin in großem Umfang aus
Synapsen des peripheren sympathischen Nervensystems stammt und erst nach Diffusion
durch die Gefäßwand in den Kreislauf gelangt. Beim in Sekundenschnelle aus dem
Nebennierenmark ausgeschütteten Adrenalin sind nach der Betäubung keine Unterschiede
Catecholamine im Stichblut
0
50
100
150
200
250
300
350
400
Adrenalin Noradrenalin
nmol/l(LSM)
System alt System neu

68
festzustellen, die durch das Zuführungssystem verursacht sein könnten. Hier besteht noch
Klärungsbedarf.
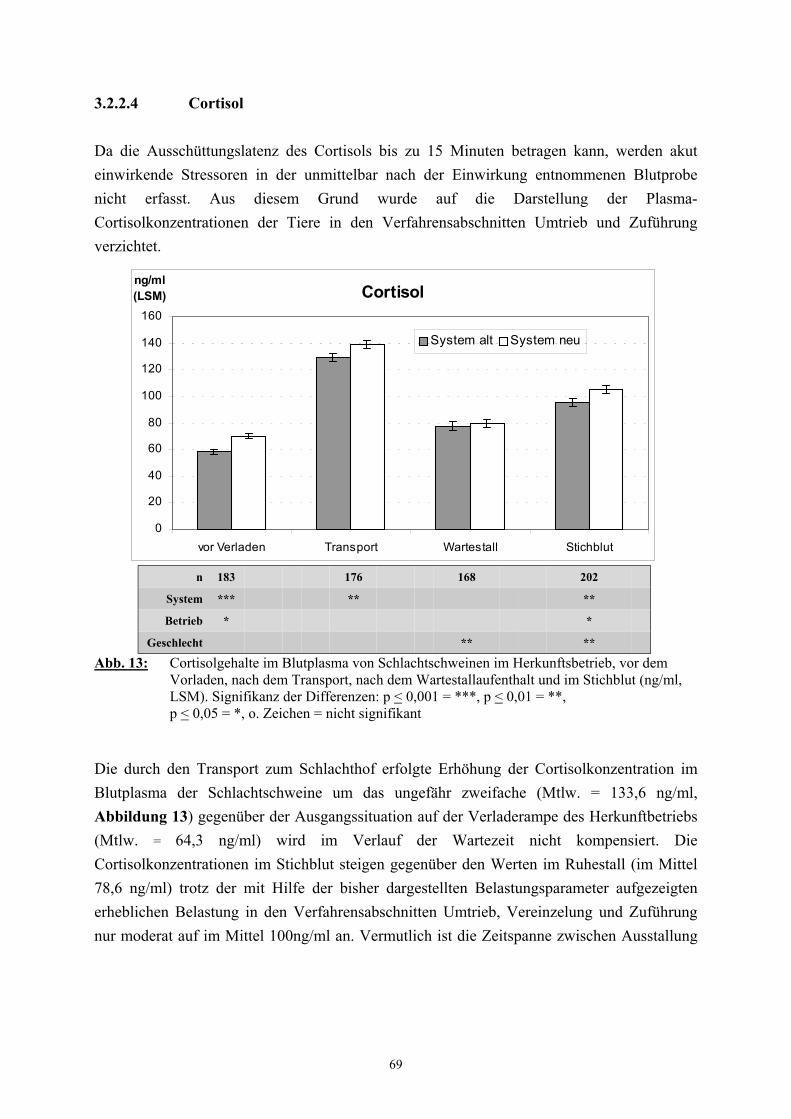
69
3.2.2.4 Cortisol
Da die Ausschüttungslatenz des Cortisols bis zu 15 Minuten betragen kann, werden akut
einwirkende Stressoren in der unmittelbar nach der Einwirkung entnommenen Blutprobe
nicht erfasst. Aus diesem Grund wurde auf die Darstellung der Plasma-
Cortisolkonzentrationen der Tiere in den Verfahrensabschnitten Umtrieb und Zuführung
verzichtet.
n 183 176 168 202
System *** ** **
Betrieb * *
Geschlecht ** **
Abb. 13: Cortisolgehalte im Blutplasma von Schlachtschweinen im Herkunftsbetrieb, vor demVorladen, nach dem Transport, nach dem Wartestallaufenthalt und im Stichblut (ng/ml,LSM). Signifikanz der Differenzen: p < 0,001 = ***, p < 0,01 = **, p < 0,05 = *, o. Zeichen = nicht signifikant
Die durch den Transport zum Schlachthof erfolgte Erhöhung der Cortisolkonzentration im
Blutplasma der Schlachtschweine um das ungefähr zweifache (Mtlw. = 133,6 ng/ml,
Abbildung 13) gegenüber der Ausgangssituation auf der Verladerampe des Herkunftbetriebs
(Mtlw. = 64,3 ng/ml) wird im Verlauf der Wartezeit nicht kompensiert. Die
Cortisolkonzentrationen im Stichblut steigen gegenüber den Werten im Ruhestall (im Mittel
78,6 ng/ml) trotz der mit Hilfe der bisher dargestellten Belastungsparameter aufgezeigten
erheblichen Belastung in den Verfahrensabschnitten Umtrieb, Vereinzelung und Zuführung
nur moderat auf im Mittel 100ng/ml an. Vermutlich ist die Zeitspanne zwischen Ausstallung
Cortisol
0
20
40
60
80
100
120
140
160
vor Verladen Transport Wartestall Stichblut
ng/ml(LSM)
System alt System neu

70
aus der Ruhebucht und dem Setzen des Entblutestiches zu kurz, um höhere
Cortisolkonzentrationen zuzulassen.
Die bei Transport und Wartezeit zu beobachtenden erhöhten Cortisolkonzentrationen in der
zweiten Messperiode (neues System) sind vermutlich durch eine erhöhte Ausschüttung von
Glucocorticoiden infolge des Kältereizes nach der Ausstallung zu erklären, da hier tiefere
Umgebungstemperaturen vorherrschten als während der ersten Messperiode (Tabelle 1).
Diese Unterschiede von etwa 10 bis 15 ng/ml zwischen den Messperioden liegen schon auf
der Verladerampe der Herkunftsbetriebe vor und setzen sich mit Ausnahme des
Wartestallaufenthaltes bis zur Betäubung der Tiere fort. Es treten auch geschlechtsspezifische
Reaktionsunterschiede auf, wobei weibliche Tiere sowohl im Wartestall wie auch im
Stichblut eine um 15 ng/ml höhere Cortisolausschüttung aufweisen als männliche (Wartestall:
85,3 vs. 72,2 ng/ml, Stichblut 107,5 vs. 92,8 ng/ml). Zum Ende des Aufenthaltes im
Schlachtbetrieb zeigen die Schweine in Phasen zunehmender Belastung auch
herkunftsbedingte Reaktionsunterschiede. Wie bei den vorhergehend besprochenen
Belastungsparametern Körpertemperatur, Noradrenalin und Adrenalin zeigen auch beim
Cortisol die Tiere aus Betrieb 2 mit 104,9 ng/ml im Stichblut gegenüber 95,5 ng/ml bei den
Schweinen aus Betrieb 1 die ausgeprägtere Belastungsreaktion (p = 0,03).
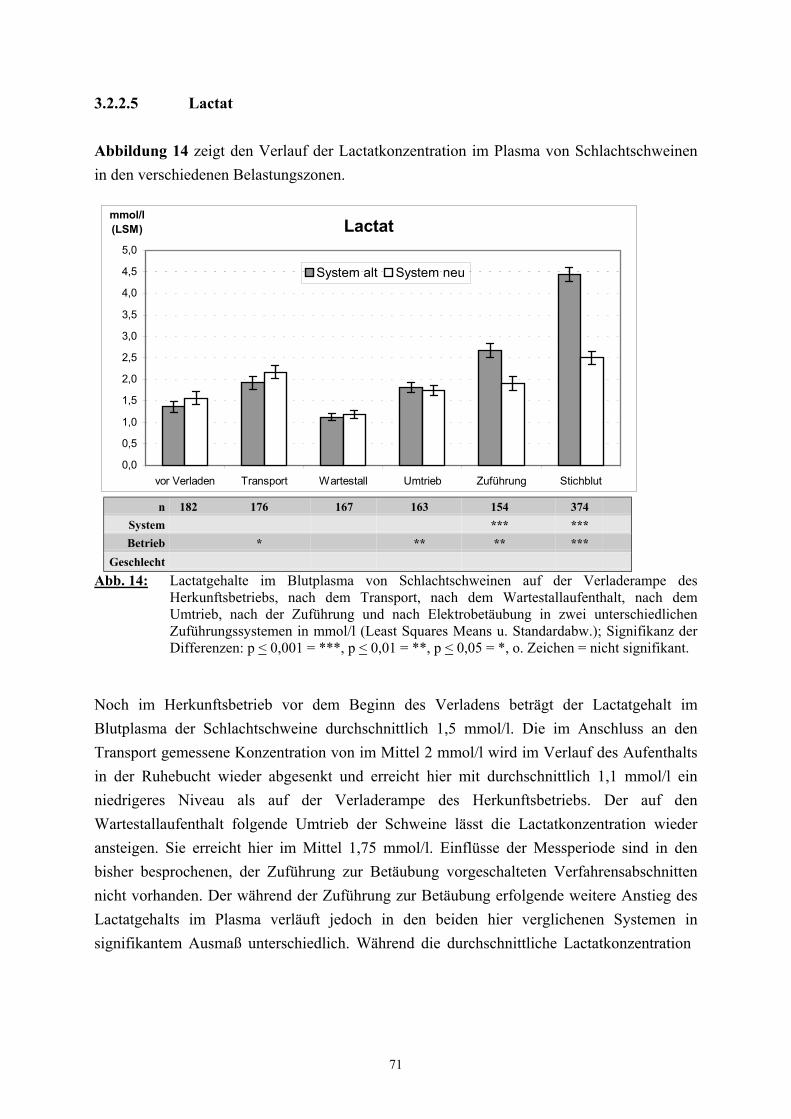
71
3.2.2.5 Lactat
Abbildung 14 zeigt den Verlauf der Lactatkonzentration im Plasma von Schlachtschweinen
in den verschiedenen Belastungszonen.
n 182 176 167 163 154 374
System *** ***
Betrieb * ** ** ***
Geschlecht
Abb. 14: Lactatgehalte im Blutplasma von Schlachtschweinen auf der Verladerampe desHerkunftsbetriebs, nach dem Transport, nach dem Wartestallaufenthalt, nach demUmtrieb, nach der Zuführung und nach Elektrobetäubung in zwei unterschiedlichenZuführungssystemen in mmol/l (Least Squares Means u. Standardabw.); Signifikanz derDifferenzen: p < 0,001 = ***, p < 0,01 = **, p < 0,05 = *, o. Zeichen = nicht signifikant.
Noch im Herkunftsbetrieb vor dem Beginn des Verladens beträgt der Lactatgehalt im
Blutplasma der Schlachtschweine durchschnittlich 1,5 mmol/l. Die im Anschluss an den
Transport gemessene Konzentration von im Mittel 2 mmol/l wird im Verlauf des Aufenthalts
in der Ruhebucht wieder abgesenkt und erreicht hier mit durchschnittlich 1,1 mmol/l ein
niedrigeres Niveau als auf der Verladerampe des Herkunftsbetriebs. Der auf den
Wartestallaufenthalt folgende Umtrieb der Schweine lässt die Lactatkonzentration wieder
ansteigen. Sie erreicht hier im Mittel 1,75 mmol/l. Einflüsse der Messperiode sind in den
bisher besprochenen, der Zuführung zur Betäubung vorgeschalteten Verfahrensabschnitten
nicht vorhanden. Der während der Zuführung zur Betäubung erfolgende weitere Anstieg des
Lactatgehalts im Plasma verläuft jedoch in den beiden hier verglichenen Systemen in
signifikantem Ausmaß unterschiedlich. Während die durchschnittliche Lactatkonzentration
Lactat
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
vor Verladen Transport Wartestall Umtrieb Zuführung Stichblut
mmol/l(LSM)
System alt System neu

72
unmittelbar nach der Passage des Einzeltreibganges im alten System bei 2,66 mmol/l liegt,
beträgt sie nach der automatischen Beförderung im neuen System nur 1,90 mmol/l (p <
0,001). In der etwa 15 Sekunden später entnommenen Stichblutprobe liegen die mittleren
Konzentrationen bei 4,45 mmol/l rsp. 2,49 mmol/l (p < 0,001). Die Lactatspiegel zeigen somit
im Stichblut eine Vervierfachung gegenüber den während der Wartephase erreichten
Ruhewerten, im neuen System lediglich eine Verdopplung.
In den Belastungsphasen Transport, Umtrieb, Zuführung und Betäubung zeigen die Schweine
aus Betrieb 2 um 0,5 bis 1,0 mmol/l höhere Lactatspiegel im Blutplasma als Tiere aus Betrieb
1. Dieses deutet auf ein herkunftsbedingt schlechteres "Training" von Bewegungsapparat und
Kreislauf hin, das mit einer anhand anderer Belastungsparameter nachweisbaren geringeren
emotionalen Belastungsfähigkeit einhergeht.
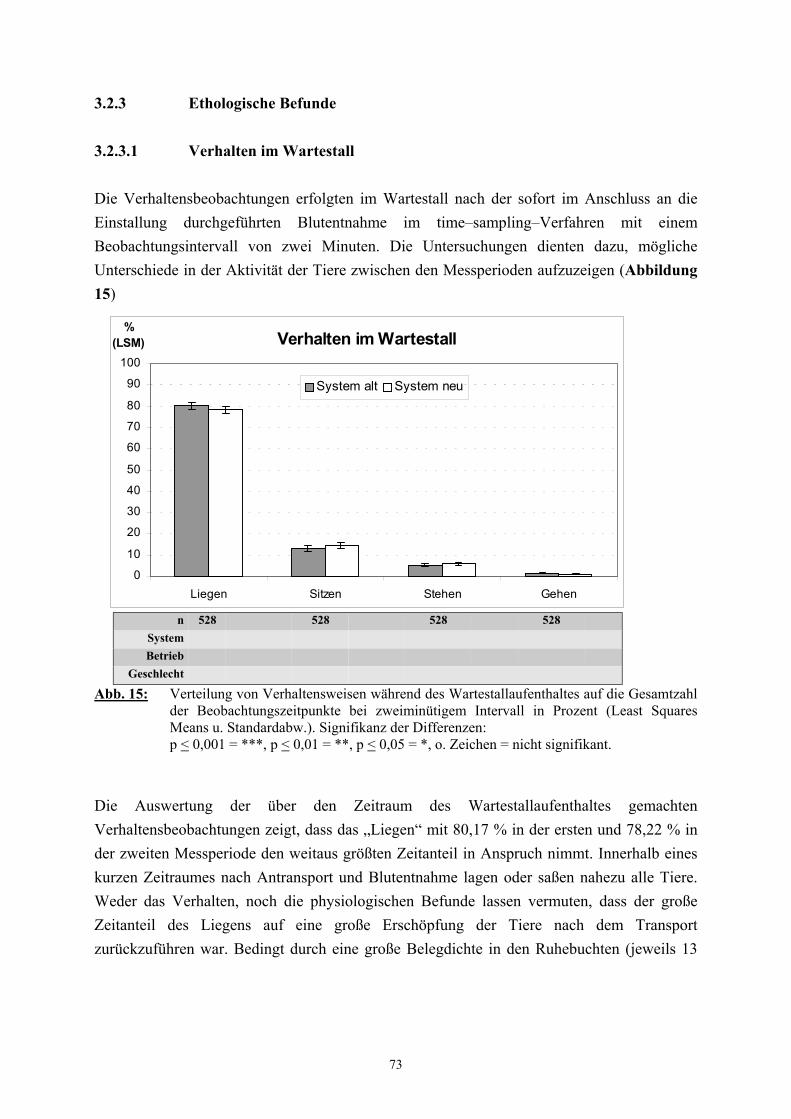
73
3.2.3 Ethologische Befunde
3.2.3.1 Verhalten im Wartestall
Die Verhaltensbeobachtungen erfolgten im Wartestall nach der sofort im Anschluss an die
Einstallung durchgeführten Blutentnahme im time–sampling–Verfahren mit einem
Beobachtungsintervall von zwei Minuten. Die Untersuchungen dienten dazu, mögliche
Unterschiede in der Aktivität der Tiere zwischen den Messperioden aufzuzeigen (Abbildung
15)
n 528 528 528 528
System
Betrieb
Geschlecht
Abb. 15: Verteilung von Verhaltensweisen während des Wartestallaufenthaltes auf die Gesamtzahlder Beobachtungszeitpunkte bei zweiminütigem Intervall in Prozent (Least SquaresMeans u. Standardabw.). Signifikanz der Differenzen: p < 0,001 = ***, p < 0,01 = **, p < 0,05 = *, o. Zeichen = nicht signifikant.
Die Auswertung der über den Zeitraum des Wartestallaufenthaltes gemachten
Verhaltensbeobachtungen zeigt, dass das „Liegen“ mit 80,17 % in der ersten und 78,22 % in
der zweiten Messperiode den weitaus größten Zeitanteil in Anspruch nimmt. Innerhalb eines
kurzen Zeitraumes nach Antransport und Blutentnahme lagen oder saßen nahezu alle Tiere.
Weder das Verhalten, noch die physiologischen Befunde lassen vermuten, dass der große
Zeitanteil des Liegens auf eine große Erschöpfung der Tiere nach dem Transport
zurückzuführen war. Bedingt durch eine große Belegdichte in den Ruhebuchten (jeweils 13
Verhalten im Wartestall
0
10
20
30
40
50
60
70
80
90
100
Liegen Sitzen Stehen Gehen
%(LSM)
System alt System neu

74
Tiere auf 2 x 3 m = etwa 0,46 m2 pro Schwein) machte das „Sitzen“ in beiden Messperioden
mit im Mittel 13,5 % den zweitgrößten Zeitanteil aus. Hier ist wie beim Liegen weder ein
Einfluss durch die Messperiode, noch durch Herkunftsbetrieb oder Geschlecht der Tiere
festzustellen. Dies gilt ebenso für die als aktives Verhalten anzusehenden Verhaltensweisen
„Stehen“ und „Gehen“, die im Mittel 5,6 % bzw. 1,3 % des Zeitbudgets beanspruchten.
Rangauseinandersetzungen wurden nur in sehr seltenen Einzelfällen registriert.
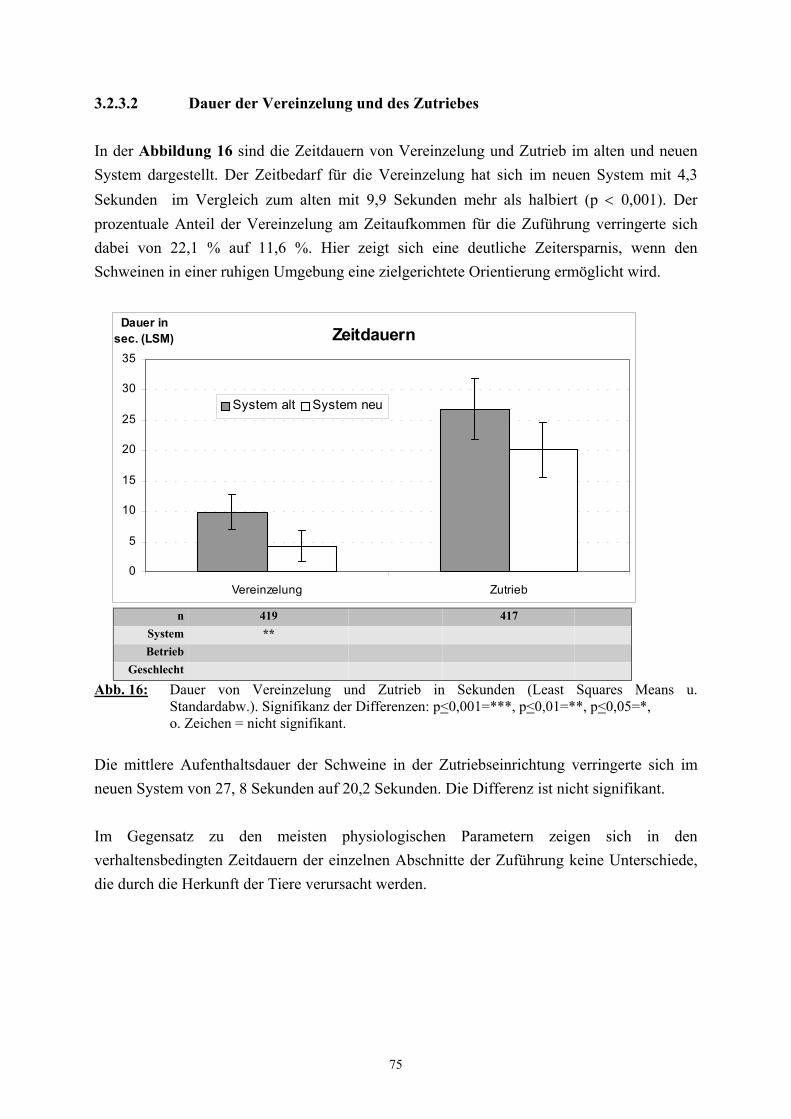
75
3.2.3.2 Dauer der Vereinzelung und des Zutriebes
In der Abbildung 16 sind die Zeitdauern von Vereinzelung und Zutrieb im alten und neuen
System dargestellt. Der Zeitbedarf für die Vereinzelung hat sich im neuen System mit 4,3
Sekunden im Vergleich zum alten mit 9,9 Sekunden mehr als halbiert (p < 0,001). Der
prozentuale Anteil der Vereinzelung am Zeitaufkommen für die Zuführung verringerte sich
dabei von 22,1 % auf 11,6 %. Hier zeigt sich eine deutliche Zeitersparnis, wenn den
Schweinen in einer ruhigen Umgebung eine zielgerichtete Orientierung ermöglicht wird.
n 419 417
System **
Betrieb
Geschlecht
Abb. 16: Dauer von Vereinzelung und Zutrieb in Sekunden (Least Squares Means u.Standardabw.). Signifikanz der Differenzen: p<0,001=***, p<0,01=**, p<0,05=*, o. Zeichen = nicht signifikant.
Die mittlere Aufenthaltsdauer der Schweine in der Zutriebseinrichtung verringerte sich im
neuen System von 27, 8 Sekunden auf 20,2 Sekunden. Die Differenz ist nicht signifikant.
Im Gegensatz zu den meisten physiologischen Parametern zeigen sich in den
verhaltensbedingten Zeitdauern der einzelnen Abschnitte der Zuführung keine Unterschiede,
die durch die Herkunft der Tiere verursacht werden.
Zeitdauern
0
5
10
15
20
25
30
35
Vereinzelung Zutrieb
Dauer in sec. (LSM)
System alt System neu

76
3.2.3.3 Verhalten während der Vereinzelung
In Abbildung 17 sind die Zeitanteile der dominierenden Verhaltensweisen von
Schlachtschweinen im Verfahrensabschnitt Vereinzelung , der im alten System für ein Tier
im Mittel 9,9 Sekunden und im neuen System gemittelt 4,2 Sekunden dauerte, dargestellt.
n 515 515
System * *
Betrieb
Geschlecht
Abb. 17: Dauer dominierender Verhaltensweisen während der Vereinzelung in Sekunden (LeastSquares Means u. Standardabw.). Signifikanz der Differenzen: p<0,001=***, p<0,01=**,p<0,05=*, o. Zeichen = nicht signifikant.
Zur Vereinzelung im alten System wurde nach Treibhilfeeinsatz, der häufig als erste Intention
des Treibers zur Vereinzelung des betreffenden Tieres aufzufassen ist, eine Fortbewegung der
Tiere über etwa 5 Sekunden erreicht. 3,4 Sekunden verbrachten die Schweine mit der
Ausprägung von Vermeidungsverhalten (Stehen trotz Treibdruck, Rückwärtsbewegung,
Ausweichen), so dass bei einer Gesamtdauer der Vereinzelung von 9,9 Sekunden rechnerisch
noch etwa 1,5 Sekunden für eine freiwillige Fortbewegung der Tiere in die
Zutriebseinrichtung hinein verblieben. Auch im neuen System erfolgte die Vereinzelung zum
größten Teil nach dem Einsatz von Treibhilfen, der hier eine Fortbewegung der Tiere über 3,6
Sekunden induzierte. Der Treibhilfeeinsatz erfolgte jedoch zumeist nur als Bewegungsreiz in
Form eines leichten Klapses mit der Hand oder mit Hilfe eines Treibstockes auf den Rücken
der Tiere und diente zum Separieren der Schweine aus der Gruppe. Im Gegensatz zum alten
System verlief der Eintrieb in die Zutriebseinrichtung nahezu problemlos. Dies wird auch
Verhalten während der Vereinzelung
0
1
2
3
4
5
6
7
Vorwärtsgehen nach Treibhilfe Vermeidungsverhalten
Dauer in sec.(LSM)
System alt System neu

77
durch die äußerst geringe Ausprägung von Vermeidungsverhalten deutlich, das im Mittel über
weniger als eine Sekunde (0,3 Sekunden) lang gezeigt wurde. Setzt man das zeitliche
Aufkommen des durch Treibhilfen induzierten Vorwärtsgehens in Relation zu dem des
Vermeidungsverhaltens, so zeigt sich, dass die Tiere während der Vereinzelung im neuen
System die 12-fache Zeit das vermeintlich weniger belastende und im Schlachtablauf
erwünschte Vorwärtsgehen zeigten, während dieser Wert im alten System nur 1,47 beträgt.
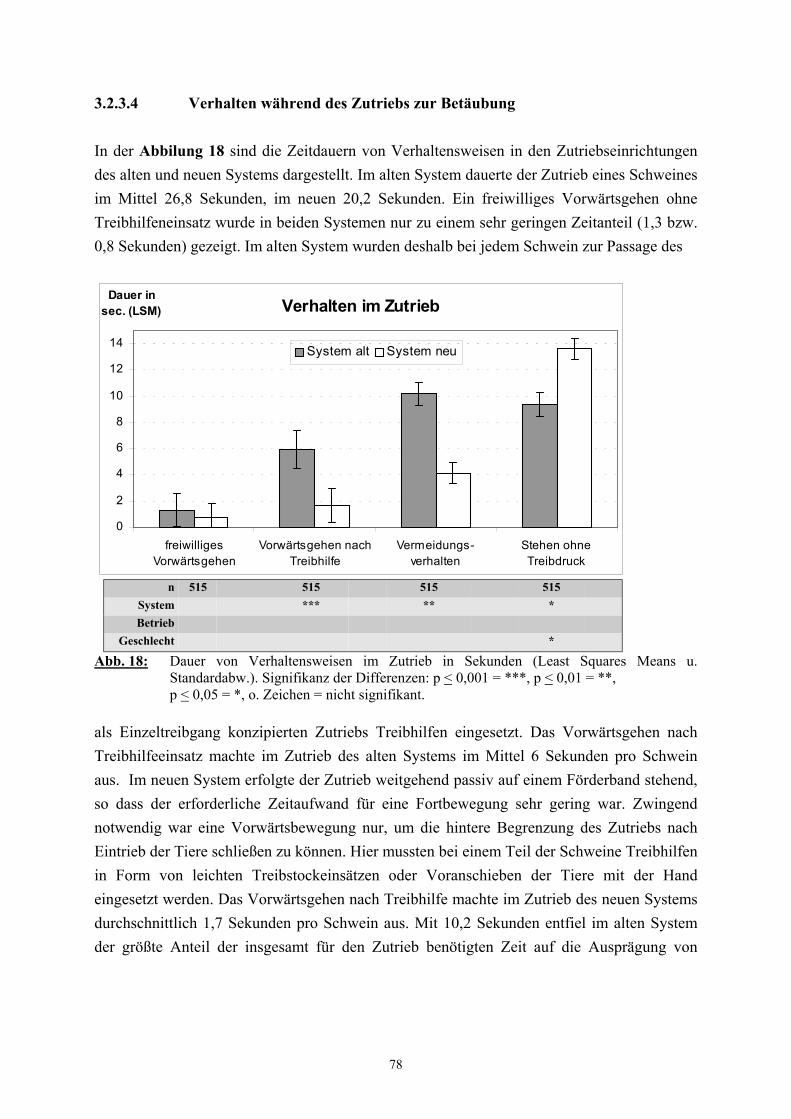
78
3.2.3.4 Verhalten während des Zutriebs zur Betäubung
In der Abbilung 18 sind die Zeitdauern von Verhaltensweisen in den Zutriebseinrichtungen
des alten und neuen Systems dargestellt. Im alten System dauerte der Zutrieb eines Schweines
im Mittel 26,8 Sekunden, im neuen 20,2 Sekunden. Ein freiwilliges Vorwärtsgehen ohne
Treibhilfeneinsatz wurde in beiden Systemen nur zu einem sehr geringen Zeitanteil (1,3 bzw.
0,8 Sekunden) gezeigt. Im alten System wurden deshalb bei jedem Schwein zur Passage des
n 515 515 515 515
System *** ** *
Betrieb
Geschlecht *
Abb. 18: Dauer von Verhaltensweisen im Zutrieb in Sekunden (Least Squares Means u.Standardabw.). Signifikanz der Differenzen: p < 0,001 = ***, p < 0,01 = **, p < 0,05 = *, o. Zeichen = nicht signifikant.
als Einzeltreibgang konzipierten Zutriebs Treibhilfen eingesetzt. Das Vorwärtsgehen nach
Treibhilfeeinsatz machte im Zutrieb des alten Systems im Mittel 6 Sekunden pro Schwein
aus. Im neuen System erfolgte der Zutrieb weitgehend passiv auf einem Förderband stehend,
so dass der erforderliche Zeitaufwand für eine Fortbewegung sehr gering war. Zwingend
notwendig war eine Vorwärtsbewegung nur, um die hintere Begrenzung des Zutriebs nach
Eintrieb der Tiere schließen zu können. Hier mussten bei einem Teil der Schweine Treibhilfen
in Form von leichten Treibstockeinsätzen oder Voranschieben der Tiere mit der Hand
eingesetzt werden. Das Vorwärtsgehen nach Treibhilfe machte im Zutrieb des neuen Systems
durchschnittlich 1,7 Sekunden pro Schwein aus. Mit 10,2 Sekunden entfiel im alten System
der größte Anteil der insgesamt für den Zutrieb benötigten Zeit auf die Ausprägung von
Verhalten im Zutrieb
0
2
4
6
8
10
12
14
freiwilligesVorwärtsgehen
Vorwärtsgehen nachTreibhilfe
Vermeidungs-verhalten
Stehen ohneTreibdruck
Dauer in sec. (LSM)
System alt System neu

79
Vermeidungsverhalten, was die Schwierigkeiten bei der Passage des Einzeltreibganges
verdeutlicht. Dementsprechend kehrt sich das Verhältnis von Vorwärtsbewegung zu
Vermeidungsverhalten im Vergleich zur Vereinzelung um. Betrug es dort noch 1,47
(Vorwärtsgehen nach Treibhilfe) zu 1 (Vermeidungsverhalten), ergibt sich nun ein Verhältnis
von 0,72 (freiwilliges und durch Treibhilfe induziertes Vorwärtsgehen) zu 1
(Vermeidungsverhalten). Anders betrachtet bedeutet das, dass der zeitliche Anteil des
Vermeidungsverhaltens im Zutrieb des alten Systems um den Faktor 1,4 höher ist als der der
Vorwärtsbewegung. Im alten System zeigten die Tiere also zu einem größeren Anteil das
vermeintlich stärker belastende und im Schlachtablauf unerwünschte Vermeidungsverhalten.
Dagegen kam es im Zutrieb des neuen Systems zu einer signifikanten Reduzierung des
Vermeidungsverhaltens, das hier durchschnittlich 4,1 Sekunden lang gezeigt wurde und
hauptsächlich in Form eines Zurücktretens an die Rücklaufsperre auftrat. Einige wenige Tiere
zeigten ein intensives Zurückdrängen. Den größten Zeitanteil verbrachten die Tiere im neuen
System erwartungsgemäß stehend ohne Wirkungseinfluss des Personals. Die
durchschnittliche Dauer der Verhaltensausprägung Stehen ohne Treibdruck machte
dementsprechend im Mittel 13,6 Sekunden pro Schwein aus. Im Zutrieb des alten Systems
nahm das Stehen ohne Treibdruck mit im Mittel 9,4 Sekunden den zweitgrößten Anteil des
gesamten Zeitaufwandes ein und war als indifferentes Warten auf das Öffnen der
Betäubungsfalle aufzufassen. Für den Eintrieb in die Falle mussten in der Regel erneut
Treibhilfen eingesetzt werden, teilweise durch den wiederholten Einsatz des Elektrotreibers.

80
3.2.3.5 Einsatz von Treibhilfen
In der Abbildung 19 ist der Einsatz von Treibhilfen durch das Schlachthofpersonal in den
Verfahrensabschnitten Vereinzelung und Zutrieb dargestellt. Der Einsatz verschiedener
Treibhilfen (Einwirkung per Hand, per Treibstock, Elektrotreiber) wurde mit einem Faktor
versehen, um subjektiv die aversive Wirkung auf die Tiere gewichten zu können. Je nach
Intensität erhielten die Einwirkungen per Hand oder Stock einen Faktor von 1 bis 3, der nur
im Zutrieb des alten Systems verwendete Elektrotreibstab von 8.
n 515 515
System *** ***
Betrieb
Geschlecht
Abb. 19: Summe der gewichteten Treibhilfen (Least Squares Means u. Standardabw.). Signifikanzder Differenzen: p<0,001=***, p<0,01=**, p<0,05=*, o. Zeichen = nicht signifikant.
Sowohl in der Vereinzelung, als auch im Zutrieb wurde der Einsatz von Treibhilfen im neuen
System signifikant reduziert. Während der Vereinzelung im alten System erreichte der
Treibhilfeindex (= Summe der gewichteten Treibhilfen) im Mittel 17,8 Einheiten, während er
im neuen System auf 6,3 Einheiten sank. Der erhebliche Einsatz von Treibhilfen im Zutrieb
des alten Systems wird durch den Index von 76,9 Einheiten verdeutlicht. Im Zutrieb des
neuen Systems betrug er dagegen 28,5 Einheiten. Dennoch erscheint der Treibhilfeindex im
neuen System immer noch zu hoch. Um das hintere Begrenzungstor schließen zu können,
musste eine Anzahl von Schweinen vorwärts getrieben werden, nachdem sie das Förderband
betreten hatten und erkundend stehen blieben. Häufig erfolgte dieser Treibhilfeeinsatz durch
Treibhilfeeinsatz
0
10
20
30
40
50
60
70
80
90
100
Vereinzelung Zutrieb
gewichteter Index (LSM)
System alt System neu

81
einzelne Mitarbeiter des Schlachtbetriebs jedoch zu früh, da nach Schließen des Tores erst
einige Zeit verging, bevor nach Freiwerden der Betäubungsfalle das Förderband in Bewegung
gesetzt werden konnte.
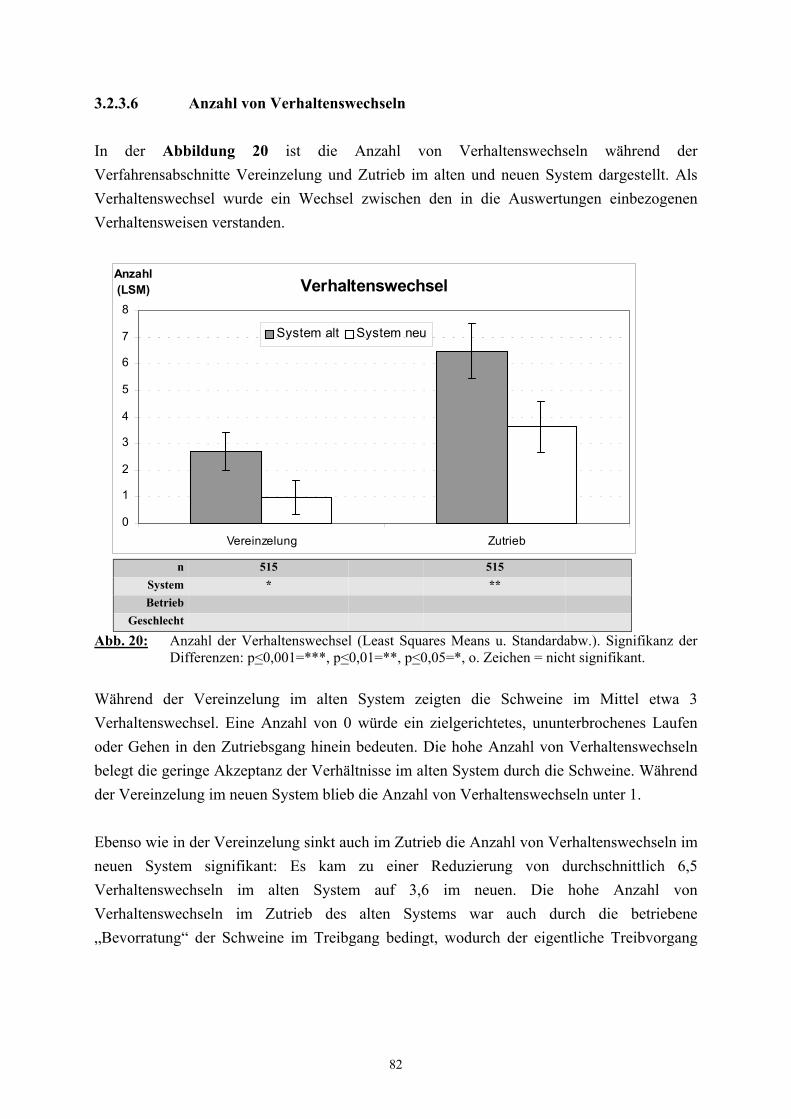
82
3.2.3.6 Anzahl von Verhaltenswechseln
In der Abbildung 20 ist die Anzahl von Verhaltenswechseln während der
Verfahrensabschnitte Vereinzelung und Zutrieb im alten und neuen System dargestellt. Als
Verhaltenswechsel wurde ein Wechsel zwischen den in die Auswertungen einbezogenen
Verhaltensweisen verstanden.
n 515 515
System * **
Betrieb
Geschlecht
Abb. 20: Anzahl der Verhaltenswechsel (Least Squares Means u. Standardabw.). Signifikanz derDifferenzen: p<0,001=***, p<0,01=**, p<0,05=*, o. Zeichen = nicht signifikant.
Während der Vereinzelung im alten System zeigten die Schweine im Mittel etwa 3
Verhaltenswechsel. Eine Anzahl von 0 würde ein zielgerichtetes, ununterbrochenes Laufen
oder Gehen in den Zutriebsgang hinein bedeuten. Die hohe Anzahl von Verhaltenswechseln
belegt die geringe Akzeptanz der Verhältnisse im alten System durch die Schweine. Während
der Vereinzelung im neuen System blieb die Anzahl von Verhaltenswechseln unter 1.
Ebenso wie in der Vereinzelung sinkt auch im Zutrieb die Anzahl von Verhaltenswechseln im
neuen System signifikant: Es kam zu einer Reduzierung von durchschnittlich 6,5
Verhaltenswechseln im alten System auf 3,6 im neuen. Die hohe Anzahl von
Verhaltenswechseln im Zutrieb des alten Systems war auch durch die betriebene
„Bevorratung“ der Schweine im Treibgang bedingt, wodurch der eigentliche Treibvorgang
Verhaltenswechsel
0
1
2
3
4
5
6
7
8
Vereinzelung Zutrieb
Anzahl(LSM)
System alt System neu

83
nach einer Zeit des Wartens erneut begonnen werden musste. Es kam hier besonders zu einem
Wechsel zwischen Vermeidungsverhalten und Vorwärtsgehen. Die geringere Zahl von
Verhaltenswechseln im Zutrieb des neuen Systems erklärt sich dadurch, dass die Schweine
hier ihr Verhalten nur eigenmotiviert ohne Einwirkung des Personals wechselten.

84
4 Diskussion
In den letzten Jahren wurde dem Wohlbefinden von Schlachttieren sowohl gesellschaftlich,
als auch wissenschaftlich immer mehr Aufmerksamkeit gewidmet (DANTZER 1994,
BARTON-GADE 1997). Untersuchungen haben gezeigt, dass für Schlachtschweine die
Zuführung zur Betäubung die wesentlichste Belastungsquelle im Schlachtprozess darstellt
(TRÖGER und WOLTERSDORF 1989, MARAHRENS et al. 1997). Häufig kommt es hier
zu Problemen, die mit Tierschutzanforderungen nicht zu vereinbaren sind. So wird eine
Fortbewegung der Tiere während der Vereinzelung und des nachfolgenden Zutriebs oft nur
durch den massiven Einsatz von Treibhilfen wie Schlagstöcken und/oder Elektrotreibern
erreicht, wodurch das Wohlbefinden der Tiere erheblich beeinträchtigt wird. Zur
Verbesserung der Situation ist eine Überwachung und gründliche Schulung des Personals von
großer Bedeutung, jedoch kann auch hoch qualifiziertes und einfühlsames Personal eine
unzureichende, nicht auf das Verhalten von Schweinen abgestimmte bauliche Gestaltung
kaum ausgleichen (BRIESE et al. 1997a). Daher wird gefordert, Zuführungssysteme
tiergerechter zu gestalten. Zur Reduzierung der Belastung werden insbesondere bauliche
Maßnahmen empfohlen, die eine den Bedürfnissen der Tiere entsprechende schonende
Fortbewegung ermöglichen (CHRISTENSEN und BARTON-GADE 1997).
Ziel der vorliegenden Arbeit ist es, den Einfluss der auf das Verhalten von Schweinen
abgestimmten baulichen Gestaltung eines neuartigen Zuführungssystems auf die
Belastungsreaktion der Tiere während der Zuführung zur Betäubung näher zu kennzeichnen
und zu überprüfen, welche Bedeutung sich daraus für das Wohlbefinden der Tiere ergibt. Zu
diesem Zweck wurden die Belastungsantworten von Schlachtschweinen anhand
physiologischer, hämatologischer und ethologischer Parameter zunächst in einem
herkömmlichen Zuführungssystem mit Einzeltreibgang und nachfolgend in einem im selben
Schlachtbetrieb eingerichteten neuen System, das auf das Verhalten von Schweinen
abgestimmt ist und unter anderem Orientierungshilfen bietet und eine aktive Fortbewegung
der Tiere im Zutrieb durch ein Förderband entbehrlich macht, erfasst. Zur Kennzeichnung von
zumindest relativen Ruhewerten und, um eine Übersicht über die Vorbelastung der Schweine
zu erhalten, wurden jeweils auch die der Zuführung zur Betäubung vorausgehenden Vorgänge
berücksichtigt.

85
4.1 Belastungsreaktion der Schweine vor und nach dem Einbau des neuen
Zuführungssystems anhand physiologischer und hämatologischer
Parameter
In der vorliegenden Arbeit sind in allen dem Verfahrensabschnitt „Zuführung zur Betäubung“
vorgeschalteten, weitgehend standardisierten Belastungszonen von der Verladerampe in den
Herkunftsbetrieben über den Transport und den Wartestallaufenthalt bei der Herzfrequenz
keine saisonal bedingten Unterschiede zwischen den beiden Untersuchungsperioden vor und
nach dem Umbau des Zuführungssystems festzustellen (Abbildung 8). Die nach den
Belastungen während des Transports von den Schweinen im Wartestall des Schlachtbetriebes
über einen Zeitraum von 30 Minuten erreichte minimale HF von etwa 100 Schlägen pro
Minute kann noch als Ruhewert angesehen werden. Trotz einer erheblichen Reduzierung der
motorischen Aktivität während der Ruhephase liegt sie allerdings an der Obergrenze des
Normbereichs für Schweine dieser Gewichtsklasse, was unter anderem darauf zurück geführt
werden kann, dass sich die Schweine nach Ausstallung und Transport erneut in einer für sie
fremden Umgebung befanden. Die hieraus resultierende Erhöhung der Wachsamkeit und
Fluchtbereitschaft führt nach LYHS (1986) und WOLTERSDORF und LYHS (1988) auch zu
einer Erhöhung der Herzfrequenz. Weiterhin könnte auch die hohe Belegdichte in der
Ruhebucht, die mit 0,46 m2 pro Schwein unter den empfohlenen 0,55 bis 0,67 m2 liegt
(WARRISS 1994, BARTON-GADE 1997), einer weitergehenden Reduzierung der
Herzfrequenz während der Wartephase entgegen gewirkt haben.
Der Anstieg der Herzfrequenz während des Umtriebes zur Vereinzelungsbucht über etwa 20
Meter Triebweg, der zwei Richtungswechsel von jeweils 90° enthielt, auf 156 Schläge/min in
der ersten und 150 Schläge/min in der zweiten Messperiode ist mit großer Wahrscheinlichkeit
auf die plötzlich einsetzende motorische Aktivität der Tiere zurückzuführen. In
Untersuchungen von BARTON-GADE (1997) zeigten dänische Schlachtschweine in einem
automatischen Umtriebssystem ohne Einwirkung des Schlachthofpersonals Herzfrequenzen
von 149 bis 155 Schlägen/min. Der in der vorliegenden Arbeit zwischen den
Untersuchungsperioden vorhandene signifikante Unterschied von etwa 6 Schlägen/min
könnte dagegen auf psychische Faktoren zurückzuführen sein. Im alten System lag die
Vereinzelungsbucht im Wartestall und dessen akustischer und optischer Umgebung, während
sie im neuen System von Wartestall und Schlachthalle getrennt in einer ruhigen und hellen
Umgebung untergebracht war. Hierdurch könnte eine Reduzierung der HF verursacht worden
sein, da der Umtrieb erst mit dem Beginn der Vereinzelung endete.

86
Während der Zuführung zur Betäubung (Vereinzelung und Zutrieb) im alten System, dessen
Zutrieb aus einem Einzeltreibgang bestand, lag die Herzfrequenz der Schlachtschweine mit
im Mittel 189 Schlägen/min im präpathologischen Bereich (WIEPKEMA und KOOLHAAS
1993), während sie im neuen System mit automatischem Zutrieb nur 168 Schläge/min betrug.
Eine Differenzierung zwischen Vereinzelung und Zutrieb war messtechnisch nicht möglich,
da die mittlere Vereinzelungsdauer im neuen Zuführungssystem etwa 4 Sekunden dauerte, das
Mess- und Speicherintervall bei der HF-Messung jedoch bei 5 Sekunden lag. Bei gleicher
Ausgangs- oder Reaktionslage zeigt sich eine erhebliche Reduzierung der Kreislaufbelastung
bei den im neuen System der Betäubung zugeführten Tieren. In Untersuchungen von
MARAHRENS et al. (1997) und von FLOß et al. (1997) zeigten Schlachtschweine in
verschiedenen konventionellen Zutriebseinrichtungen Herzfrequenzen von über 200
Schlägen/min. Ähnlich wie in der vorliegenden Untersuchung war die Passage dieser
Zutriebsgänge mit einer massiven Anwendung von elektrischen Treibhilfen verbunden, die
die Herzfrequenz stark ansteigen lassen (GEVERINK et al. 1995). In einem neuen
vollautomatischen Zuführungssystem zur CO2 – Betäubung eines dänischen Schlachtbetriebes
stellten CHRISTENSEN und BARTON-GADE (1997) bei Schlachtschweinen
Herzfrequenzen von 136 bis 145 Schlägen pro Minute fest. Die Tiere verblieben während der
gesamten Zuführung in Gruppen von jeweils 5 Mitgliedern, ohne dass ein Einsatz von
Treibhilfen erfolgte. Auch FLOß et al. (1997) und FLOß (1999) fanden in automatischen,
nach ethologischen Gesichtspunkten eingerichteten Systemen bei Schlachtschweinen
verschiedener Herkünfte Herzfrequenzen von 140 bis 150 Schlägen in der Minute vor. Die
drastische Reduzierung der Herzfrequenz beruht in dem hier vorgestellten neuen
Zuführungssystem einerseits auf einer verminderten motorisch bedingten Kreislaufbelastung
durch den passiven Transport auf Förderbändern, andererseits, jedoch in einem vermutlich
viel stärkerem Maße, auf einem verminderten psychischen Anpassungsdruck, da die Tiere im
Zutrieb nicht zum Ausführen eines bestimmten Verhaltens gezwungen waren und in der
Vereinzelung eine Vorwärtsbewegung durch entsprechende Maßnahmen gefördert wurde.
Auch die Haltung im Herkunftsbetrieb scheint einen Einfluss auf die kreislaufregulative
Belastungsantwort der Schweine während der Zuführung zu besitzen. Schweine aus dem
Herkunftsbetrieb 1 zeigen hier eine um etwa 10 Schläge/min höhere HF als Schweine aus
dem Betrieb 2. Je nach dem, wie im allgemeinen während der Aufzucht mit den Tieren
umgegangen wird, zeigen Schweine, die im Umgang mit dem Menschen positive Erfahrungen
gemacht haben und in einer abwechslungsreichen Haltungsumgebung aufgezogen wurden,
nach GRANDIN (1986), FISCHER (1995), HEMSWORTH et al. (1996), HUNTER et al.

87
(1997) und FLOß (1999) im Schlachtablauf teilweise deutlich geringere Belastungsreaktionen
des Herz-Kreislaufsystems als reizarm gehaltene und den Kontakt mit Menschen kaum
gewöhnte Tiere. Da aber in beiden Betrieben konventionelle Haltungsbedingungen vorlagen
und über einen Umgang mit den Tieren keine genauen Kenntnisse vorhanden sind, kann der
Grund für die festgestellten Unterschiede in der Herzfrequenz nur vermutet werden. In
diesem Zusammenhang könnte ein häufigerer Umtrieb während der Mastperiode in Betrieb 2
in dem Sinne eine Rolle gespielt haben, dass die Tiere hier eher die Möglichkeit hatten,
Copingstrategien für den Wechsel von Umgebungsbedingungen zu entwickeln (GONYOU et
al. 1986, BROOM und JOHNSON 1993, BROOM 1996).
Bezogen auf die Unterschiede zwischen den beiden Herkunftsbetrieben zeigt die
Körpertemperatur weiblicher Schweine ebenso wie auch alle untersuchten hämatologischen
Belastungsindikatoren eine im Vergleich zur Herzfrequenz gegenläufige Tendenz. Die
vaginal kontinuierlich gemessene Körpertemperatur der Tiere aus Betrieb 2 liegt in allen
Belastungszonen (vor Verladen, Transport, Wartestall, Umtrieb, Zuführung) des
Schlachtablaufes höher. Es wird hier offensichtlich, dass die Bedingungen der
Haltungsumwelt im Herkunftsbestand in einer differenzierten Weise verschiedene
Regelkreise der Belastungsantwort wie das Herz-Kreislaufsystem, den Stoffwechsel, die
Thermoregulation oder die psychische Bewältigungskapazität (Coping) beeinflussen. Trotz
einer derzeitigen Zunahme integrierter, auch verschiedene Haltungsfaktoren einbeziehende
Fleischerzeugungsprogramme sind die diesbezüglichen Erkenntnisse eher das Ergebnis
empirischer Erfahrungen als systematischer Untersuchungen (BARTON-GADE 1997).
Die in beiden Messperioden in nahezu gleichem Umfang stattfindende Erhöhung der
Körpertemperatur während des Transports kann besonders auf den Verladevorgang zurück
geführt werden, denn nach AUGUSTINI und FISCHER (1982) steigt die Körpertemperatur
von Schlachtschweinen dabei ebenso wie die Herzfrequenz schnell an. Während des
Wartestallaufenthaltes kam es bei nur gering ausgeprägten, saisonal bedingten Unterschieden
zwischen den Messperioden wieder zu einem Absinken der Körpertemperatur auf Werte, die
mit durchschnittlich 39 °C im physiologischen Bereich liegen (Abbildung 9). Von
MICKWITZ (1982) fand, dass eine minimale Ruhephase von einer Stunde nötig ist, damit die
Körpertemperatur transporterhitzter Schweine wieder Normwerte erreicht. LONG und
TARRANT (1989) ermittelten die Rektaltemperatur von Schweinen zum Zeitpunkt der
Schlachtung und fanden bei Tieren, die während des Wartestallaufenthaltes nicht beduscht
worden waren, je nach Jahreszeit Temperaturen zwischen 39,3 und 39,5 °C. Als Äquivalent
für die Körpertemperatur unmittelbar vor der Betäubung untersuchten SCHÜTTE et al.

88
(1995c) die Temperatur im Stichblut von Schweinen. Abhängig davon, ob eine Beduschung
statt fand oder nicht und, ob diese mit kaltem oder warmen Wasser durchgeführt wurde,
fanden sie Werte zwischen 39,9 und 40,4 °C , die somit die obere Grenze des Normbereichs
deutlich überschritten. Auch in der vorliegenden Untersuchung bewegte sich die
Körpertemperatur während des Umtriebs und der sich anschließenden Zuführung zur
Betäubung wieder über das im Wartestall erreichte Ruheniveau hinaus, jedoch lag die
durchschnittliche Körpertemperatur während der Zuführung zur Betäubung im neuen System
mit 39,4 °C signifikant niedriger als im herkömmlichen mit 39,6 °C. Verschiedene Autoren
fordern, die Körpertemperatur von Schweinen vor der Schlachtung so niedrig wie möglich zu
halten, um verschiedenen Fleischqualitätsmängeln vorzubeugen (AUGUSTINI et al. 1977,
KLONT und LAMBOOY 1995a,b). Zu beachten ist hierbei jedoch, dass negative
Auswirkungen auf die Fleischqualität nicht durch eine reine Beschleunigung von
biochemischen Vorgängen, also durch die erhöhte Temperatur selbst, zustande kommen,
sondern durch die Fortsetzung der im Zuge der Belastungsreaktion ablaufenden
physiologischen und hormonellen Veränderungen, die sich auf den Muskelstoffwechsel
auswirken (MONIN et al. 1995). Der im neuen Zuführungssystem signifikant geringere
Anstieg der Körpertemperatur deutet darauf hin, dass hier der wärmeproduzierende, durch
motorische und psychische (catecholaminvermittelte) Belastungen induzierte Energieumsatz
in einem geringeren Ausmaß provoziert wird.
Die Plasmaspiegel der beiden Catecholamine Noradrenalin und Adrenalin sind im
Abschnitt „vor Verladen“ in beiden Messperioden nahezu identisch (Abbildung 10 und 11)
und entsprechen in etwa den von FORSLID und AUGUSTINSSON (1988) für Katheterblut
angegebenen Ruhewerten, was dafür spricht, dass die Belastung durch den
Blutentnahmevorgang selbst nur gering ausgeprägt und weiterhin in beiden Messperioden
etwa gleich groß war. Die Ausschüttung der Catecholamine verlief in den sich an den
Wartestallaufenthalt anschließenden Verfahrensabschnitten vor und nach dem Einbau des
neuen Zuführungssystems in ähnlicher Weise, wobei sich jeweils auch Noradrenalin und
Adrenalin sehr ähnlich verhielten. Im Anschluss an den Umtrieb kam es in beiden Systemen
zu einer weiteren Erhöhung der Catecholaminspiegel im Plasma, die jedoch im
herkömmlichen Zuführungssystem ein im Vergleich zum neuen System nahezu doppelt so
hohes Niveau erreichten. Bezogen auf die relativen Ruhewerte kam es demnach im alten
System während der Zuführung zur Betäubung durchschnittlich zu einer Verdreifachung der
Plasma-Catecholaminspiegel, während diese im neuen System nur noch etwa um den Faktor
1,5 erhöht waren. Die Ergebnisse entsprechen in der Größenordnung denen von RAABE et al.
(1996), liegen jedoch weit unter denen von TROEGER und WOLTERSDORF (1988), die

89
nach dem Zutrieb Catecholaminspiegel fanden, die bis zu 7 mal höher lagen als die der
Ruheproben. Im Stichblut nach der Betäubung (etwa 15 Sekunden nach der Blutentnahme im
Zutrieb entnommen) setzen sich die beim Vergleich des alten mit dem neuen
Zuführungssystem gefundenen Unterschiede auf einem um den Faktor 60 erhöhten Niveau
nur beim Noradrenalin fort, während die Adrenalinausschüttung in beiden Systemen etwa 300
nmol/l betrug und damit gegenüber der nach dem Zutrieb gemessenen Konzentration um den
Faktor 120 bis 200 erhöht wurde (Abbildung 12). In Untersuchungen von TROEGER und
WOLTERSDORF (1988), MARAHRENS et al. (1997) und NOWAK (1998) lagen die
mittleren Plasmagehalte an Catecholaminen bei Schweinen nach der Elektrobetäubung um die
Faktoren 60 bis 500 höher als im Wartestall oder im Herkunftstall, bevor die Schweine zum
Schlachtbetrieb transportiert wurden. Diese dramatische Erhöhung der
Catecholaminausschüttung während der Elektrobetäubung wurde bisher darauf zurückgeführt,
dass es infolge des Stromflusses zu einer Depolarisation der synaptischen
Nervenzellmembran mit maximaler Entspeicherung der Catecholaminvorräte kommt
(TROEGER und WOLTERSDORF 1988, REUTER 1992, MARAHRENS et al. 1997).
Neuere Untersuchungen zur Betäubung mit CO2 – Gas in Konzentrationen von 85 % konnten
diese Zusammenhänge jedoch nicht bestätigen, da hier im Vergleich zur Elektrobetäubung
ähnlich hohe oder teilweise sogar noch um den Faktor 10 höhere
Catecholaminkonzentrationen gemessen wurden (NOWAK et al. 2000, 2001). Die hohe
Catecholaminausschüttung während der Betäubung scheint daher die tatsächliche Belastung
der Schweine in diesem Abschnitt widerzuspiegeln.
Die durchschnittliche Cortisolkonzentration der noch vor dem Verladen genommenen Proben
entspricht mit 64,3 ng/ml (Abbildung 13) in etwa den Ergebnissen von MARAHRENS et al.
(1997), die im Blutplasma von Schlachtschweinen vor dem Ausstallen und dem
anschließenden Transport zum Schlachtbetrieb einen Cortisolgehalt von im Mittel 60 ng/ml
fanden. Aufgrund der diurnalen Schwankungen und der episodischen Sekretion variiert der
Ruhewert für den Cortisolgehalt des Plasmas relativ stark und bewegt sich beim Schwein
zwischen 20 bis 80 ng/ml (HANNON 1990).
Die durch den Transport zum Schlachthof erfolgte Erhöhung der Cortisolkonzentration im
Blutplasma der Schlachtschweine um etwa 100 % gegenüber der Ausgangssituation im
Herkunftsstall wird im Verlauf der sich anschließenden etwa zweistündigen Wartezeit im
Schlachthof nicht vollständig kompensiert. WARRIS et al. (1992) halten für die Absenkung
der durch den Transport angestiegenen Cortisolkonzentration einen zwei- bis dreistündigen
Wartestallaufenthalt für notwendig. Die Cortisolkonzentrationen im Stichblut sind gegenüber
den Befunden nach der Wartezeit nur in einem geringen Umfang erhöht, obwohl die Tiere

90
durch Umtrieb, Vereinzelung und Zutrieb zur Betäubung erheblichen Belastungen
unterworfen waren, wie durch andere Parameter, wie Herzfrequenz, Catecholamin- und
Lactatgehalt des Blutplasmas, dokumentiert ist. Wahrscheinlich verhindert die
Ausschüttungslatenz des Cortisols ein Ansteigen der Konzentration im Stichblut, weil die
Zeitspanne zwischen Umtrieb und Betäubung maximal 10 Minuten betrug und somit zu kurz
für einen neuerlichen Anstieg ist (GEERS et al. 1994, WARRISS et al. 1995a, LINDNER et
al. 1996, RAABE et al. 1996). Somit kann der Meinung von WARRIS et al. (1994)
zugestimmt werden, dass der Cortisolgehalt im Stichblut nicht die unmittelbar vor der
Betäubung stattfindenden Ereignisse reflektiert und daher nur wenig zur Beurteilung von
unterschiedlichen Zuführungssystemen geeignet ist.
Die in allen Phasen des Schlachtablaufes zu beobachtenden erhöhten Cortisolkonzentrationen
während der zweiten Messperiode (neues System) sind vermutlich mit der vermehrten
Ausschüttung von Glucocorticoiden infolge eines in der zweiten Messperiode stärkeren
Kältereizes nach der Ausstallung (Tabelle 1) zu erklären. Eine metabolische Thermogenese
durch eine entsprechende Aktivierung des Energiestoffwechsels wird durch Cortisol
vermittelt (THUN und SCHWARTZ-PORSCHE 1994). Auch MARAHRENS et al. (1997)
fanden in einer Untersuchung zur Belastung von Schlachtschweinen im Winter in allen
Belastungszonen höhere Cortisolspiegel als im Sommer. Durch die Untersuchung des
Cortisolgehaltes des Plasmas ist somit die Möglichkeit gegeben, auch Begleitfaktoren wie
Klima und Wetter in die physiologischen Erhebungen einzubeziehen.
Der Verlauf der Plasma-Lactatkonzentration weist in den der Zuführung zur Betäubung
vorgeschalteten Abschnitten (vor Verladen, Transport, Wartestall und Umtrieb) keine
signifikanten Unterschiede zwischen den Messperioden auf und ähnelt den Herzfrequenz- und
Körpertemperaturverläufen.
Der durchschnitlliche Lactatgehalt der noch im Herkunftsbetrieb vor dem Verladen
gewonnenen Proben betrug durchschnittlich 1,5 mmol/l (Abbildung 14) und liegt damit
wenig über dem von MARAHRENS et al. (1997) im entsprechenden Abschnitt gefundenen
Wert von 1 mmol/l, jedoch noch innerhalb des von BICKHARDT (1996) für den Plasma-
Lactatgehalt von Schweinen angegebenen Ruherahmens (1 bis 2 mmol/l). KRAFT (1973)
untersuchte Schweine während eines 24-stündigen Wartestallaufenthalts und fand, dass es
innerhalb der ersten Stunde des Aufenthalts zu einem Abfall der Lactatkonzentration im
Blutplasma kam, sich die Konzentration in den folgenden 23 Stunden dann aber nicht mehr
änderte. Auch in der vorliegenden Untersuchung kam es während der etwa zwei Stunden
dauernden Ruhephase im Wartestall wieder zu einem Absinken der im Verlauf des Transports
angestiegenen Lactatkonzentration. Im Gegensatz zu den Ergebnissen von MARAHRENS et

91
al. (1997) wurden hierbei wieder Ruhewerte erreicht, die mit durchschnittlich 1,1 mmol/l
sogar noch unter den Ausgangswerten vor dem Verladen liegen. Dies deutet darauf hin, dass
in beiden Messperioden während des Wartestallaufenthalts ein tatsächliches Ausruhen der
Schweine erfolgte, was durch die direkten Beobachtungen und die Ergebnisse der
ethologischen Untersuchungen (Abbildung 15) bestätigt werden kann. Zusammenhängen
könnte dies interessanterweise mit der hohen Belegdichte in den Wartebuchten, die zwar
einerseits durch die erhebliche Einschränkung der Bewegungsfreiheit zu einer psychischen
Belastung der Tiere führte, andererseits jedoch auch eine starke Reduzierung der motorischen
Aktivität bewirkte. In Untersuchungen zum Transport von Schweinen bei unterschiedlichen
Ladedichten machten CHRISTENSEN und BARTON-GADE (1998) eine ähnliche
Beobachtung. Sie stellten fest, dass hohe Ladedichten mit geringerer Aktivität der Tiere und
tendenziell geringeren Lactatgehalten im Stichblut einhergingen. Der sich an den
Wartestallaufenthalt anschließende Umtrieb der Tiere äußerte sich in beiden hier untersuchten
Messperioden in einer Erhöhung des Plasma-Lactatgehalts auf im Mittel 1,8 mmol/l. Ein
weiterer Anstieg während Vereinzelung und Zutrieb erfolgte im neuen Zuführungssystem, in
dem eine aktive Fortbewegung der Schweine im Zutrieb nicht notwendig war, nur noch um
0,1 mmol/l und damit in einem weitaus geringeren Umfang als im herkömmlichen System mit
Einzeltreibgang, wo sich die Lactatkonzentration im Vergleich zum Umtrieb noch um 0,9
mmol/l erhöhte. Zu einer weiteren Zunahme dieses Unterschieds, der die weitaus geringere
motorische Belastung der Schweine im neuen Zuführungssystem anzeigt, kommt es im
Stichblut. Die im alten Zuführungssystem ermittelten Werte liegen zwar unter den häufig in
herkömmlichen Systemen gefundenen 8 bis 15 mmol/l (HONKAVAARA 1989, WARRIS et
al. 1992, WARRIS et al. 1995b, NEUBERT et al. 1996, SCHÜTTE et al. 1996,
MARAHRENS et al. 1997, BROWN et al. 1998, von WENZLAWOWICZ et al. 1998), sind
mit durchschnittlich 4,45 mmol/l jedoch beinahe doppelt so hoch wie im neuen
Zuführungssystem. Die dort gefundenen 2,5 mmol/l unterschreiten noch die von BARTON-
GADE und CHRISTENSEN (1998) in einem automatischen Zuführungssystem ermittelten
Werte von etwa 4 mmol/l, entsprechen aber exakt der von FLOß (1999) nach Zuführung in
einem System mit automatischem Zutrieb gefundenen Lactatkonzentration im Stichblut.
Dieser vergleichsweise niedrige Lactatgehalt deutet darauf hin, dass sich die physische
Belastung der Schweine während der Zuführung zur Betäubung im neuen System in Grenzen
hielt, die aus der Sicht des Tierschutzes zu vertreten sind.
Zusammenfassend lässt sich noch einmal sagen, dass die im neuen Zuführungssystem im
Vergleich zum alten signifikant niedrigere Herzfrequenz und Körpertemperatur sowie die
signifikant geringeren Konzentrationen von Noradrenalin-, Adrenalin- und Lactat im

92
Blutplasma eine deutliche Reduzierung der physischen und emotionellen Belastung der
Schweine im neuen Zuführungssystem anzeigen, während die Ausschüttungslatenz beim
Cortisol eine eindeutige Zuordnung der Ereignisse erschwert und dazu führt, dass die
Konzentration im Stichblut offensichtlich nicht die Belastung zum Zeitpunkt der Zuführung
zur Betäubung widerspiegelt.

93
4.2 Belastungsreaktion der Schweine vor und nach dem Einbau des neuen
Zuführungssystems anhand ethologischer Parameter
In dem der Vereinzelung und dem eigentlichen Zutrieb vorgeschalteten Verfahrensabschnitt
Wartestallaufenthalt, in dem die Schweine sich von den Belastungen des vorausgegangenen
Transportes zum Schlachtbetrieb erholen sollen, sind zwischen den beiden Messperioden
keine Unterschiede im Verhalten der Tiere zu erkennen (Abbildung 15). Mit etwa 80 %
nimmt das Liegen hier den weitaus größten Zeitanteil in Anspruch, gefolgt von Sitzen (14 %)
und Stehen (5 %). In Untersuchungen von FLOß (1999) zeigten Schweine bei morgendlicher
Anlieferung und einer Ausruhzeit von etwa zwei Stunden in verschiedenen Schlachtbetrieben
Aktivität (alle Verhaltensausprägungen außer Liegen und Sitzen) über 40 bis 70 % des
Wartestallaufenthaltes. LAUBE und SCHULZE (1992) zeigten auf, dass der Zeitpunkt des
Ausruhens nach Anlieferung von der Gruppengröße und der Beleuchtung in der Wartebucht
abhängt. In Gruppengrößen unter 60 Tieren und einer Beleuchtung von nicht über 80 lux
ruhten nach 60 Minuten 87 % der Tiere. Dass insgesamt in kleineren Gruppen während des
Wartestallaufenthaltes weniger Unruhe herrscht, führen CHRISTENSEN und BARTON-
GADE (1997) darauf zurück, dass hier die Chance für die Schweine geringer ist, auf einen
aggressiven Artgenossen zu treffen. Auch HANSEN et al. (1989, 1991) und GEVERINK et
al. (1996) beobachteten, dass es in größeren Gruppen immer nur wenige Tiere gibt, die
aggressive Auseinandersetzungen beginnen und so zu Schwierigkeiten führen. Die
Gruppengröße in der vorliegenden Untersuchung betrug konstant 13 Tiere, wobei die
Zusammensetzung der Transportgruppe beibehalten wurde. Die Beleuchtungsstärke in der
Wartebucht lag bei 50 bis 110 Lux. Beide Faktoren trugen dazu bei, dass etwa 30 Minuten
nach der unmittelbar im Anschluss an die Einstallung vorgenommenen Blutentnahme in der
Wartebucht unabhängig von der Jahreszeit trotz eines begrenzten Liegeflächenangebotes von
im Mittel 0,46 m2 pro Tier (Empfehlung: 0,55 bis 0,67 m2, WARRISS et al. 1994, BARTON-
GADE 1997) nahezu alle Schweine lagen und ruhten.
Wie weiter oben dargelegt, wird die auf physiologischer Ebene ablaufende
Belastungsreaktion von Schlachtschweinen während der Zuführung zur Betäubung erheblich
vom dafür genutzten System beeinflusst. Im Folgenden werden die Auswirkungen der
wichtigsten Konstruktionsmerkmale der beiden Zuführungssysteme auf ethologische und
typische Prozessparameter miteinander verglichen.
Die nicht auf das Verhalten von Schweinen abgestimmten baulichen Gegebenheiten im alten
System bedingten, dass der Verfahrensabschnitt Vereinzelung häufig Schwierigkeiten
bereitete und mit durchschnittlich 10 Sekunden pro Tier mehr als doppelt so lange dauerte wie

94
im neuen System mit 4,3 Sekunden (Abbildung 16). Die von FLOß (1999) in einem
Systemvergleich für die Vereinzelungsdauer ermittelten 23 bzw. 91 Sekunden pro Tier
erscheinen vergleichsweise hoch, es muss jedoch beachtet werden, dass in seiner
Untersuchung zur Vereinzelung die gesamte Aufenthaltsdauer der Schweine in der
Vereinzelungsbucht einschließlich des Eintriebs in den Zutrieb gezählt wurde. Dagegen
beschreibt die Vereinzelungsdauer in der vorliegenden Arbeit jeweils nur den eigentlichen
Treibvorgang aus der Vereinzelungsbucht in den Zutrieb hinein. Eine hiermit vergleichbare
Zeitspanne untersuchten auch SCHÄFFER et al. (1999), die die zum Austrieb einer
Schweinegruppe aus einer Bucht in einen Treibgang benötigte Zeit ermittelten und daraus die
durchschnittliche Austriebszeit pro Tier berechneten. Diese variierte, abhängig von
Gruppengröße und Treiber, zwischen 3,5 und 7,8 Sekunden. Die akustische Umgebung spielt
für das Schwein eine große Rolle (KLEIBS 1992, SPENCLEY und HARTUNG 1994,
WARRIS et al. 1994, LAUBE 1996). Da sich im alten System die Vereinzelungsbucht
innerhalb des Wartestallbereichs befand, waren die Schweine dort nicht nur dem durch den
Vereinzelungsvorgang selbst entstehenden Lärm, sondern auch Fremdgeräuschen, die unter
anderem durch die im selben Raum erfolgende Anlieferung, Rangordnungskämpfe oder
andere Treibprozesse verursacht wurden, ausgesetzt. Hierdurch wurde eine Beruhigung der
durch den Umtrieb von der Wartebucht zur Vereinzelungsbucht aufgeschreckten Schweine
erschwert, und die Tiere wurden beim Erkunden der neuen Umgebung häufig abgelenkt.
Neben der häufig mit Schwierigkeiten verbundenen Separierung eines einzelnen Schweines
aus einer Gruppe (ZERBONI und GRAUVOGL 1984, BARTON-GADE 1997) gilt in vielen
Schlachtbetrieben der Eingangsbereich des Zutriebs als problematisch (GRANDIN 1983,
1988, 1991, 1994, LAUBE 1989, von MICKWITZ 1990, TRÖGER 1990, SEIDEL 1992,
LAUBE et al. 1993, STEGEN 1993, WEEDING et al. 1993, von MICKWITZ und BRIESE
1994, SCHÄFFER et al. 1997). Diese Beobachtung traf in hohem Maße auf den
Zutriebseingang des alten Systems zu, was durch die starke Ausprägung des während der
Vereinzelung gezeigten Vermeidungsverhaltens (Abbildung 17) verdeutlicht wird, das mit
einer durchschnittlichen Dauer von 3,5 Sekunden ein Verhältnis von Vorwärtsgehen zu
Vermeidungsverhalten von nur 1,5 zu 1 bedingte. Der Eingangsbereich des Zutriebs war mit
einer mittleren Beleuchtungsstärke von 30 lux nur sehr unzureichend ausgeleuchtet, wodurch
das Betreten des Zutriebes durch die Schweine erheblich erschwert wurde. Einerseits war
aufgrund der Differenz zwischen der mittleren Beleuchtungsstärke von Wartebucht (50 bis
110 lux) und Zutriebseingang eine Hell-Dunkel Adaptation durch die Schweine erforderlich,
die sehr langsam verläuft (WAIDA 1986), andererseits wirkte der dunkle Eingangsbereich
dem natürlichen Verhalten von Schweinen, bevorzugt von einem weniger beleuchteten Areal
in ein hell erleuchtetes zu laufen (van PUTTEN und ELSHOF 1978, van PUTTEN 1982b),

95
genau entgegen. Neben der Dunkelheit wirkte auch die im Eingangsbereich des Zutriebs
angebrachte erste Rücklaufsperre irritierend und abschreckend auf die Schweine. GRANDIN
(1981) empfiehlt, im Eingangsbereich von Einzeltreibgängen herunterhängende und/oder sich
bewegende Objekte zu vermeiden. Da sie von den Schweinen aufgedrückt werden musste,
wirkte die Rücklaufsperre nicht nur als optische, sondern auch als taktile Barriere beim
Eintrieb. Ungeeignete Rücklaufsperren gehören zu den häufig anzutreffenden Defiziten bei
der Zuführung zur Betäubung (von WENZLAWOWICZ et al. 1994). Den Problemen beim
Eintrieb in den Einzeltreibgang wurde mit dem Einsatz von Treibhilfen begegnet, auf die die
Schweine in vielen Fällen wiederum mit Vermeidungsverhalten reagierten. Angesichts der
beschriebenen Schwierigkeiten ist der mittlere Treibhilfeindex von 17,8 (Abbildung 19) als
moderat zu bezeichnen, was auf die große Sachkenntnis des im Bereich der Vereinzelung
arbeitenden Treibers zurückzuführen ist. Bei einer mittleren Vereinzelungsdauer von 10
Sekunden im alten System wechselten die Schweine durchschnittlich 3 mal ihr Verhalten
(Abbildung 20). In dieser hohen Anzahl von Verhaltenswechseln drückt sich der Konflikt
der Schweine zwischen einer Flucht vor einer Treibhilfe und der Aversion vor dem Betreten
des Einzeltreibganges aus.
Beim Bau des neuen Zuführungssystems wurde die von GRANDIN (1981, 1982) gegebene
Empfehlung umgesetzt, Vereinzelungsbucht und Zutrieb nicht durch die Wand zwischen
Wartestallbereich und Schlachthalle zu trennen, sondern beides hinter die entsprechende
Wand zu platzieren. Hierdurch kann auch der ungünstige Einfluss von Fremdgeräuschen aus
dem Wartestallbereich (SCHÄFFER et al. 1999) vermieden werden. Da beispielsweise durch
die Kratz-, Brüh- und Entborstungsmaschinen ein erheblicher Lärm ausgelöst wird (BRIESE
et al. 1997b), wurden im neuen System Vereinzelung und Zutrieb nicht nur vom
Wartestallbereich, sondern auch von der Schlachthalle räumlich getrennt, wobei
schallabsorbierendes Material verwendet wurde. Außer durch die Reduzierung des
einwirkenden Lärms wurde im neuen System auch durch eine ausreichende Beleuchtung die
Voraussetzung für die Ausübung von Erkundungsverhalten durch die Schweine geschaffen
(BOGNER und GRAUVOGL 1984). Die mittlere Beleuchtungsstärke im Eingangsbereich der
Zutriebseinrichtung lag bei 380 lux. BALDWIN und MEESE (1977) und SCHULZE (1990)
geben für das Schwein eine Lichtpräferenz von 350 lux an. Entsprechend der im Vergleich
zum alten System ruhigen Atmosphäre und der den Tieren ermöglichten visuellen
Orientierung wurde das Dirigieren der Schweine zum Zutriebseingang hin im neuen System
erheblich erleichtert. Das eigentliche Betreten des Zutriebs wurde in hohem Maße durch den
zum Zeitpunkt des Eintriebs sichtbaren Metallspiegel gefördert, da die sonst den
Treibvorgang eher behindernde Neugierde der Schweine (von HOLLEBEN und von
WENZLAWOWICZ 1995) hier dazu führte, dass die Tiere zumeist auf ihr Spiegelbild und

96
somit auf den vermeintlichen Artgenossen im Zutrieb zuliefen. Die im neuen System für
Mensch und Tier wesentliche Erleichterung der Vereinzelung wird besonders dadurch
verdeutlicht, dass hier die zeitliche Ausdehnung des initial durch eine Treibhilfe induzierten
Vorwärtsgehens (Abbildung 17) mit im Mittel 3,6 Sekunden das 12-fache der
durchschnittlich mit der Ausprägung von Vermeidungsverhalten verbrachten Zeit beträgt. Im
Vergleich zum alten System und zu den Ergebnissen von FLOß (1999), der für das Verhältnis
von Vorwärtsbewegung zu Vermeidungsverhalten während der Vereinzelung in einem
System mit automatisch arbeitenden Zutrieb 2,18 zu 1 bzw. 1,88 zu 1 (Abend- bzw. Morgen-
Anlieferung) angibt, erscheint der zeitliche Anteil des Vorwärtsgehens während der
Vereinzelung im neuen System sehr hoch. Dies deckt sich mit der direkten Beobachtung, dass
die Tiere häufig nach einer initialen Treibhilfe durchgängig bis in den Zutrieb hinein vorwärts
gingen. Dementsprechend blieb auch die durchschnitliche Anzahl von Verhaltenswechseln
unter 1 (Abbildung 20), und der Einsatz von Treibhilfen fiel mit einem mittleren Index von
6,3 (Abbildung 19) gering aus. Treibhilfen wurden zumeist nur „sortierend“, das heisst zum
Herauslösen eines Tieres aus der Gruppe, und dirigierend eingesetzt.
Ebenso wie die Vereinzelung des alten Systems führte der schon aufgrund seiner
Konstruktion als Einzeltreibgang als wenig empfehlenswert einzuordnende
(WOLTERSDORF und TRÖGER 1987, TRÖGER und WOLTERSDORF 1988, TARRANT
1989, WEEDING 1993) Zutrieb im alten System zu mitunter tierschutzrelevanten
Problemen. Ausdruck der von den Schweinen eingenommenen Abwehrhaltung ist das
Vermeidungsverhalten, dessen Ausprägung mit durchschnittlich 10,2 Sekunden (Abbildung
18) den größten Anteil der für den Zutrieb benötigten Zeit einnimmt und um den Faktor 1,4
höher ist als der Zeitanteil der insgesamt ausgeführten Vorwärtsbewegung, die sich aus
freiwilligem (1,3 Sekunden) und durch Treibhilfe induziertem Vorwärtsgehen (6 Sekunden)
zusammensetzt. Auch in einem von FLOß (1999) untersuchten Einzeltreibgang nahm das
Vermeidungsverhalten den größten Zeitanteil ein und wurde 1,44 mal länger ausgeführt als
die Vorwärtsbewegung, was in der Größenordnung beinahe exakt dem in der vorliegenden
Arbeit gefundenen Ergebnis entspricht. Stockungen im Zutrieb sollten im alten System durch
eine „Vorratshaltung“ von Schweinen im Einzeltreibgang verhindert, bzw. ausgeglichen
werden, wodurch sich der mit durchschnittlich 9,4 Sekunden hohe zeitliche Anteil des
Stehens im Sinne von Warten erklärt. Während dieser Zeit blieben die Tiere zwar vom
Personal unbehelligt, doch waren sie durch die engen Ausmaße des Ganges erheblich in ihrem
Bewegungsspielraum eingeschränkt und wurden teilweise auch durch die hinter ihnen
befindlichen Tiere belästigt, was insgesamt eine erhebliche psychische Belastung bedeutet
(BRIESE et al. 1997b). Der Eintrieb in die Betäubungsfalle ist häufig ein besonders kritisches
Element des Zutriebs (ELDRIDGE et al. 1992, STEGEN 1993, WARRIS et al. 1994, LAUBE

97
1996), da die Tiere vor dem sackgassenähnlichen Ende des Einzeltreibgangs zurückschrecken
und dort zusätzlich oft Irritationen der Schweine durch bewegliche Teile, Lärmentwicklung
aus der Betäubungsanlage und Treiberbewegungen (GRANDIN 1981, 1982, 1988) ausgelöst
werden. Im Zutrieb des alten Systems scheuten die Tiere beim Eintrieb in die Betäubungsfalle
vor dem sich öffnenden Schott zurück und verweigerten häufig das Weitergehen, wenn sie die
Bewegungen des Treibers wahrnahmen, der sowohl die Beschickung der Betäubungsfalle, als
auch den Betäubungsvorgang selbst ausführte, und deshalb viel hin und her laufen musste.
Der im Zutrieb des alten Systems mit einem Index von 76,9 (Abbildung 19) starke
Treibhilfeeinsatz beruht zu einem großen Teil auf den beschriebenen Schwierigkeiten kurz
vor und beim Eintrieb in die Betäubungsfalle, was die Beobachtung von BRIESE et al.
(1997b) bestätigt, dass es häufig direkt vor der Betäubung zu einem massiven Einsatz des
Elektrotreibers kommt. Insgesamt wurde der Treibvorgang im Zutrieb des alten Systems
durch die im Zuge der Bevorratung entstehenden Wartezeiten und die häufigen
Abwehrreaktionen der Schweine immer wieder unterbrochen, was sich auch durch die
durchschnittlich 6,5 mal ausgeführten Verhaltenswechsel ausdrückt (Abbildung 20), die hier
bezogen auf die Gesamtzutriebsdauer von im Mittel 26,8 Sekunden (Abbildung 16) noch
häufiger stattfanden als in dem von FLOß (1999) untersuchten Einzeltreibgang (12 Wechsel
in 71 Sekunden). Im Zutrieb des neuen Systems konnten die vorher beobachteten Probleme
beim Zutrieb, insbesondere beim Eintrieb in die Betäubungsfalle, durch die automatische
Beförderung der Schweine entschärft werden. Die Tiere wurden dabei nicht wie in
sogenannten Zwangsförderern seitlich erfasst und ohne Bodenkontakt transportiert, was sehr
belastend ist (TRÖGER 1990), sondern sie wurden bei erhaltener Bewegungsfreiheit auf
einem sich langsam bewegenden Förderband stehend bis in die Betäubungsfalle gefahren.
Entsprechend verbrachten sie die durchschnittliche Zutriebsdauer von 20,2 Sekunden
(Abbildung 16) im neuen System mit im Mittel 13,6 Sekunden größtenteils indifferent
stehend (Abbildung 18). Die Zeit bis sich das Transportband nach Abschluss des
vorhergehenden Betäubungsvorgangs in Bewegung setzte, verbrachten die Schweine häufig
mit Erkundungsverhalten, wobei der Metallspiegel sich wiederum als besonders attraktiv
erwies. Ein von FLOß (1999) häufig beobachtetes Hinsetzen der Schweine beim Anfahren
des Transportbandes konnte nicht festgestellt werden, jedoch kam es dabei teilweise zu einem
Zurücktreten der Schweine, was auch den größten Anteil des durchschnittlich 4,1 Sekunden
lang gezeigten Vermeidungsverhaltens ausmacht. Ein Treiben der Schweine im eigentlichen
Sinne fand im Zutrieb des neuen Systems nur in geringem Umfang statt, wie durch die
zeitliche Ausdehnung des durch Treibhilfe induzierten Vorwärtsgehens verdeutlicht wird, das
im Mittel 1,7 Sekunden lang gezeigt wurde. Der Einsatz von Treibhilfen konnte so im
Vergleich zum alten System signifikant reduziert werden, doch erscheint der

98
durchschnittliche Index von 28,5 (Abbildung 19) noch zu hoch. Der Grund hierfür liegt im
Verhalten des ansonsten sachkundigen und geduldigen Personals, das sich noch nicht
genügend auf das neue System eingestellt hatte und teilweise „aus alter Gewohnheit“
Treibhilfen einsetzte, obwohl dies aufgrund der automatischen Beförderung und auch der
beweglichen Rücklaufsperre in den meisten Fällen nicht notwendig war. Dies verdeutlicht die
Notwendigkeit einer auf die Bedingungen im jeweiligen Schlachtbetrieb ausgerichteten,
gründlichen Mitarbeiterschulung (TRÖGER 1990, von WENZLAWOWICZ et al. 1994,
BRIESE et al. 1997a, SCHÄFFER et al. 1997).
Insgesamt bestätigen die Ergebnisse, dass die Nutzung des Erkundungsverhaltens die
wesentliche Voraussetzung dafür darstellt, die tierschutzrelevanten Probleme bei der
Zuführung zur Betäubung zu entschärfen (LAUBE und SCHULZE 1992). So trugen die
ruhige und schattenfrei hell beleuchtete Umgebung und der als Orientierungshilfe eingesetzte
Metallspiegel dazu bei, dass die Schweine während der Vereinzelung im neuen System nur
noch in einem geringen Ausmaß mit Vermeidungsverhalten reagierten und der Zeitbedarf für
die Vereinzelung im Vergleich zum alten System mehr als halbiert werden konnte. Neben den
das Erkundungsverhalten von Schweinen berücksichtigenden Details des neuen
Zuführungssystems wirkt sich auch die automatische Beförderung der Tiere im Zutrieb
belastungsmindernd aus, da die Schweine nicht zur Fortbewegung in Richtung Betäubung
gezwungen werden mussten, wodurch der Treibhilfeeinsatz im Vergleich zum alten System
erheblich reduziert werden konnte.

99
4.3 Bedeutung der baulichen Gestaltung des Zuführungssystems für das
Wohlbefinden der Schweine
Wie weiter oben ausgeführt kann Wohlbefinden nicht direkt gemessen werden, wohl aber ist
über die Erhebung eines breiten Spektrums an physiologischen, hämatologischen und
ethologischen Parametern eine Einschätzung möglich, indem die gewonnenen Daten als
Belastungsindikatoren und konsequenterweise auch als Indikatoren für eingeschränktes
Wohlbefinden herangezogen werden (WIEPKEMA und KOOLHAAS 1993).
Für das Wohlbefinden von Schlachtschweinen ist der Grad der motorischen Belastung von
großer Bedeutung, da die zumeist laufuntrainierten und mit nur eingeschränkten kreislauf-
und thermoregulatorischen Fähigkeiten ausgestatteten Tiere schnell an die Grenzen ihrer
körperlichen Belastbarkeit gelangen (DOMEL und LEISTNER 1977, SACKMANN et al.
1988, TRÖGER 1990, SCHULZE und LAUBE 1991, HOLZER et al. 1992). Im neuen
System waren die Schweine in erster Linie aufgrund der passiven Beförderung im Zutrieb
einer deutlich geringeren physischen Belastung ausgesetzt als im alten System, in dem die
Schweine den als Einzeltreibgang konstruierten Zutrieb aktiv durchlaufen mussten. Dies wird
insbesondere durch die mittleren Plasma-Lactatspiegel während der Zuführung zur Betäubung
und im Stichblut angezeigt, die im neuen System mit 1,90 mmol/l bzw. 2,49 mmol/l
signifikant niedriger lagen als im alten System mit 2,66 mmol/l bzw. 4,45 mmo/l (Abbildung
14). Auch die mittlere Plasma-Noradrenalinkonzentration, die wie das Lactat besonders die
motorische Belastung repräsentiert (SCHEURINK et al. 1989), erreichte im neuen System
während der Zuführung zur Betäubung mit 4,89 nmol/l ein signifikant niedrigeres Niveau als
im alten System mit 8,57 nmol/l (Abbildung 10). Die physische Entlastung der Schweine im
neuen Zuführungssystem spiegelt sich auch in der dort im Vergleich zum alten System um 21
Schläge pro Minute signifikant niedrigeren durchschnittlichen Herzfrequenz (Abbildung 8),
der um 0,2 °C niedrigeren mittleren Körpertemperatur (Abbildung 9) und in der nahezu um
die Hälfte geringeren Plasma-Adrenalinkonzentration (Abbildung 11) wider. Diese
Ergebnisse, insbesondere die Plasma-Adrenalinkonzentration (Akerstedt et al. 1983) zeigen
aber auch eine Reduzierung der psychischen Belastung im neuen Zuführungssystem an. Für
die Beurteilung des Wohlbefindens ist die psychische Komponente von besonderer
Wichtigkeit. Laut DAWKINS (1990) und BROOM (1995) schließt Wohlbefinden auch die
emotionalen Empfindungen eines Tieres mit ein. Eine in Zusammenhang mit der Schlachtung
predominant auftretende, das Wohlbefinden stark beeinträchtigende Empfindung dürfte die
Angst sein (STEPHENS und PERRY 1990, BRIESE et al. 1997a). MURPHY (1978a,b)
unterscheidet zwischen „angstauslösenden-Stimuli“, „Angst-Antworten“ und „sich in einem

100
Zustand der Angst befinden“. SCHÄFFER et al. 1999 gehen davon aus, dass das Treiben von
Schweinen aus einer Bucht in einen Gang stark angstauslösend ist. Ein besonderer Faktor
scheint hierbei neben der ständigen Treiberanwesenheit der Austrieb ins „Unbekannte“ zu
sein. Diese Situation war sowohl bei der Vereinzelung im alten System, in der die Schweine
den dunklen Einzeltreibgang betreten mussten, als auch im Zutrieb selbst gegeben. Geht man
davon aus, dass alles, was mit dem unangenehmen Gefühl der Angst verbunden ist, für Tiere
einen hohen Vermeidwert besitzt (BUCHHOLTZ 1996), kann der im neuen System im
Vergleich zum alten signifikant geringere zeitliche Anteil des Vermeidungsverhaltens
während Vereinzelung (Abbildung 17) und Zutrieb (Abbildung 18) einen Hinweis darauf
geben, dass die bauliche Gestaltung des neuen Zuführungssystems auch über eine
Reduzierung von angstauslösenden Stimuli weniger aversiv auf die Schweine wirkte. Auch
BROOM (1995) misst der von Tieren auf der Verhaltensebene geäußerten Aversion bei der
Beurteilung von psychischen Leiden Bedeutung zu und gibt den Grad des Auftretens von
Vermeidungsverhalten als Indikator für eingeschränktes Wohlbefinden an. Im
Zusammenhang mit dem hohen zeitlichen Aufkommen des Vermeidungsverhaltens muss
auch der außergewöhnlich starke Einsatz von Treibhilfen im alten System gesehen werden.
Der während der Vereinzelung und des Zutriebs durchschnittlich erreichte Index von 17,8
bzw. 76,9 lag signifikant höher als im neuen System mit 6,3 und 28,5 (Abbildung 19). Die
starke Treibbelastung im alten System war nicht nur Folge der nicht auf das Verhalten von
Schweinen abgestimmten baulichen Gestaltung, sondern wiederum selbst geeignet, die
Schweine in den Zustand der Angst zu versetzen (GRAUVOGL 1988). Außerdem kann
davon ausgegangen werden, dass durch die Qualität der im alten Zuführungssystem
eingesetzten Treibhilfen auch Schmerzen ausgelöst wurden. Insbesondere kann dies von der
nur im alten System, dort jedoch in beträchtlichem Umfang, erfolgten Anwendung des
Elektrotreibers angenommen werden, dessen Einsatz äußerst kritisch bis generell ablehnend
gesehen wird (GRANDIN 1983, TRÖGER und WOLTERSDORF 1988, TARRANT 1989,
HOLZER et al. 1992, CHRISTENSEN und BARTON-GADE 1997). Es kann gefolgert
werden, dass durch die signifikante Reduzierung des Treibhilfeeinsatzes im neuen System bei
den Schweinen in einem weitaus geringerem Umfang unangenehme Empfindungen ausgelöst
wurden als im alten System. Befindlichkeiten werden heute als Ausdruck eines
physiologischen Balancesystems verstanden (BUCHHOLTZ 1996). Ziel ist zu jedem
Zeitpunkt das Erreichen der Homöostase. Konfliktsituationen, in denen der Organismus
versuchen muss, über verschiedene Regelsysteme erneut homöostatische Verhältnisse zu
erreichen, entstehen, wenn die Handlungsbereitschaft für nicht miteinander zu vereinbarende
Verhaltensweisen etwa gleich hoch ist. Während der Zuführung zur Betäubung im alten
System kam es häufig zu Situationen, in denen bei den Schweinen gleichzeitig eine hohe

101
Handlungsbereitschaft für das Ausführen von Vermeidungsverhalten und für eine Flucht vor
den eingesetzten Treibhilfen, also für ein Vorwärtsgehen, vorlag. Ein derartiger
Motivationskonflikt drückt sich in der Anzahl von Verhaltenswechseln aus, die mit
durchschnittlich 2,7 Wechseln während der Vereinzelung und 6,5 während des Zutriebs
(Abbildung 20) gemessen an der Gesamtdauer dieser Abschnitte (Abbildung 16) hoch ist.
Kann das während eines Konflikts gestörte Gleichgewicht nicht wieder hergestellt werden,
weil etwa wie bei der Zuführung zur Betäubung eine Flucht nicht oder nur sehr eingeschränkt
möglich ist, resultiert daraus erheblich eingeschränktes Wohlbefinden (BUCHHOLTZ 1993,
1994, WIEPKEMA und KOOLHAAS 1993). Demnach zeigt die im neuen System signifikant
seltenere Ausführung von Verhaltenswechseln während der Vereinzelung und des Zutriebs
auch, dass das Wohlbefinden der Tiere hier durch belastende, eine Gegenregulation
erforderlich machende Konfliktsituationen weniger stark beeinträchtigt wurde.
Angesichts der Definition von Wohlbefinden, nach der es sich um einen Zustand von geistiger
und körperlicher Gesundheit und von Harmonie mit sich selbst und der Umwelt handelt, wird
deutlich, dass sich ein Tier während der Zuführung zur Betäubung nicht im Sinne des Wortes
wohl befinden kann, obwohl Wohlbefinden durchaus differenziert zwischen sehr gutem und
weniger gutem Wohlbefinden eingeteilt werden kann (BROOM 1997). Daher soll
abschließend nicht unerwähnt bleiben, dass die Schweine auch im neuen, nach ethologischen
Gesichtspunkten gestalteten System während der Zuführung zur Betäubung eine erhebliche
Anpassungsleistung vollbringen mussten und eine Beeinträchtigung ihres Wohlbefindens
daher auch hier nicht gänzlich vermieden werden konnte. Die physiologischen,
hämatologischen und ethologischen Belastungsindikatoren zeigen jedoch eine im Vergleich
zum herkömmlichen System deutlich verminderte physische und psychische Belastung der
Schweine im neuen Zuführungssystem an, woraus gefolgert werden kann, dass hier auch die
Beeinträchtigung des Wohlbefindens ein weitaus geringeres Ausmaß erreichte, was aus
Tierschutzsicht von entscheidender Bedeutung ist.

102
5. Schlussfolgerungen
Aus der Diskussion der Befunde zur Belastungsreaktion von Schlachtschweinen in zwei
unterschiedlichen Zuführungssystemen zur Elektrobetäubung lassen sich folgende
Schlussfolgerungen ziehen:
1. Die während der Zuführung zur Betäubung auf physiologischer Ebene erfolgende
Belastungsreaktion von Schlachtschweinen lässt sich durch kontinuierliche Herzfrequenz-
und Körpertemperaturmessungen sowie die Bestimmung der unmittelbar nach Beendigung
der Zuführung im Plasma vorhandenen Noradrenalin-, Adrenalin- und Lactatkonzentration
darstellen. Die dafür erforderliche Gewinnung von Blutproben sollte im Fall der
Catecholamine im Anschluss an den Zutrieb vor dem Einsetzen der Betäubung über einen
Venenverweilkatheter erfolgen, da in Stichblutproben der Einfluss der Zuführung durch die
Belastungen, die bei der Betäubung selbst entstehen, überlagert wird. Zur Untersuchung auf
den Lactatgehalt eignen sich dagegen Stichblutproben.
2. Die Bestimmung der Cortisolkonzentration im Stichblut ist aufgrund der
Ausschüttungslatenz des Cortisols nicht geeignet, die während der Zuführung zur Betäubung
entstehenden Belastungen wiederzugeben.
3. Schweine aus zwei verschiedenen Herkunftsbetrieben, deren Haltungsbedingungen sich
nicht grundsätzlich unterscheiden, zeigen signifikante Unterschiede in der physiologischen
Belastungsreaktion, die sich unabhängig von der baulichen Gestaltung des
Zuführungssystems bis in den Bereich der Zuführung zur Betäubung fortsetzen. Offenbar
beeinflussen die vielfältigen Faktoren der Haltungsumwelt im Herkunftsbestand verschiedene
Regelkreise der Belastungsantwort in differenzierter Weise, so dass sie in künftige
Untersuchungen eingeschlossen werden sollten.
4. Das Ausmaß der während der Zuführung zur Betäubung aktivierten physiologischen
Belastungsreaktion von Schlachtschweinen hängt wesentlich von der baulichen Gestaltung
des Zuführungssystems ab. Dabei reduziert eine das Verhalten von Schweinen
berücksichtigende Gestaltung alle hier untersuchten physiologischen und biochemischen
Indikatoren (Herzfrequenz, Körpertemperatur, Catecholamine und Lactat), die eine durch die
Zuführung zur Betäubung verursachte physische und psychische Belastung anzeigen.

103
5. Die Anwendung von optischen Attraktivreizen, wie zum Beispiel ein Metallspiegel im
Zutriebseingang, in einem die Ausübung von Erkundungsverhalten erleichternden ruhigen
und schattenfrei hell beleuchteten Umfeld reduziert in der Vereinzelung die Ausprägung des
Vermeidungsverhaltens und fördert eine eigenmotivierte Fortbewegung der Tiere in den
Zutrieb. Der Einsatz von Treibhilfen beschränkt sich daher weitgehend auf eine sortierende
Orientierungshilfe, wodurch die Vereinzelung bei gleichzeitiger Reduzierung der Belastung
der Tiere im zeitlichen Ablauf verkürzt und standardisiert wird.
6. Auf den automatischen Transport auf einem sich langsam bewegenden Förderband
reagieren Schweine unter den hier untersuchten Bedingungen ruhig und zeigen nur in sehr
geringem Umfang Vermeidungsverhalten. Die automatische Beförderung von Schweinen im
Zutrieb ermöglicht es dem Personal, weitgehend auf den Einsatz von Treibhilfen,
insbesondere des Elektrotreibers, zu verzichten und trägt so zu einer erheblichen Reduzierung
der Belastung der Schweine bei.
7. Durch den Einbau eines nach ethologischen Gesichtspunkten konstruierten
Zuführungssystems, das im Zutrieb keine „Bevorratung“ von Schweinen für einen
kontinuierlichen Betäubungstakt vorsieht, kommt es nicht zu Verzögerungen im
Schlachtablauf. Durch zeitliche Verkürzung vor allem des Verfahrensabschnitts Vereinzelung
und bei konsequenter Vermeidung von Wartezeiten im Zutrieb wird im Gegenteil die
Möglichkeit geschaffen, die hier untersuchte Schlachtzahl von etwa 150 bis 180 Tieren pro
Stunde weiter zu erhöhen, ohne die Schlachtschweine dabei in einem stärkeren Umfang zu
belasten.
8. Das neue Zuführungssystem, dessen bauliche Gestaltung darauf ausgerichtet ist, sich das
Erkundungsverhalten von Schweinen zu Nutze zu machen und die Tiere möglichst schonend
in die Betäubungsfalle zu befördern, führt gegenüber herkömmlichen Systemen zu einer
geringeren Beeinträchtigung des Wohlbefindens von Schlachtschweinen. Damit wird auch der
Forderung der Tierschutz-Schlachtverordnung (TierSchlV 1997) nach Vermeidung unnötiger
Schmerzen und Leiden eher Rechnung getragen. Die vorgestellten Anregungen zur
Verbesserung des Tierschutzes sollten verstärkt in der Praxis umgesetzt werden, zumal sie zu
keinen Mehrkosten führen müssen.

104
6 Zusammenfassung
Die Zuführung zur Betäubung stellt für Schweine eine der wesentlichsten Belastungsquellen
im Schlachtablauf dar. In vielen Schlachtbetrieben wird ein schonender Umgang des
Personals mit den Tieren während Vereinzelung und Zutrieb durch eine unzureichende
bauliche Gestaltung dieser Bereiche erschwert, so dass es hier häufig zu Problemen kommt,
die das Wohlbefinden der Tiere beeinträchtigen können. In der vorliegenden Arbeit sollte der
Frage nachgegangen werden, inwieweit sich die Belastungen, denen Schweine während der
Zuführung zur Betäubung ausgesetzt sind, durch bauliche Maßnahmen reduzieren lassen.
Dazu wurden ein herkömmliches und ein neuartiges, in Zusammenarbeit mit dem Institut für
Tierhygiene, Tierschutz und Nutztierethologie der Tierärztlichen Hochschule Hannover
entwickeltes Zuführungssystem miteinander verglichen.
Die Untersuchung, in die insgesamt 515 dem Bundeshybridzuchtprogramm (BHZP)
entstammende Schweine einbezogen wurden, gliederte sich in zwei nacheinander im selben
Schlachtbetrieb stattfindende Messperioden. In der ersten Messperiode, in der 266 Tiere
untersucht wurden, verfügte der Schlachtbetrieb über ein herkömmliches Zuführungssystem
(altes System). Dabei gelangten die Schweine aus der im Wartestallbereich gelegenen
Vereinzelungsbucht in einen zur Betäubungsfalle führenden Einzeltreibgang. In der zweiten,
nach einem Umbau des Schlachtbetriebs stattfindenden Messperiode wurden 249 Schweine
untersucht, die nun in dem neuartigen, nach ethologischen Gesichtspunkten entwickelten
System der Betäubung zugeführt wurden (neues System). Im neuen System erfolgt die
Vereinzelung und der Zutrieb optisch und akustisch von Wartestall und Schlachthalle
getrennt, und im Zutrieb werden die Tiere auf einem Förderband stehend automatisch in die
Betäubungsfalle transportiert. Um die Belastung der Schweine einschätzen zu können,
wurden jeweils kontinuierliche Herzfrequenz- (Polar® Accurex Plus) und
Körpertemperaturmessungen (Pillbox® -Logger, intravaginal) vorgenommen, und es wurden
über Venenverweilkatheter und aus dem Stichblut gewonnene Blutproben auf ihren Gehalt an
Noradrenalin, Adrenalin, Cortisol und Lactat untersucht. Weiterhin wurde das Verhalten der
Schweine per Videoüberwachung erfasst und ausgewertet.
Die Ergebnisse zeigen, dass die Zuführung zur Betäubung im neuen System im Vergleich
zum herkömmlichen mit geringeren Belastungen für die Schweine verbunden ist. So lagen
während der Zuführung zur Betäubung Herzfrequenz und Körpertemperatur im neuen
System mit durchschnittlich 168 Schlägen/min und 39,4 °C signifikant (p < 0,001 bzw. p <
0,05) niedriger als im alten System mit 189 Schlägen/min und 39,6 °C. Zudem fand sich im

105
Plasma der unmittelbar im Anschluss an den Zutrieb gewonnenen Katheterblutproben im
neuen System im Vergleich zum alten ein signifikant geringerer Gehalt an hormonellen und
biochemischen Belastungsindikatoren. Der Noradrenalingehalt reduzierte sich von 8,57
nmol/l auf 4,89 nmol/l (p < 0,001), der Adrenalingehalt von 2,64 nmol/l auf 1,49 nmol/l (p <
0,001) und der Lactatgehalt von 2,7 mmol/l auf 1,9 mmol/l (p < 0,001). Die im Plasma des
Stichbluts gemessene Lactatkonzentration verdeutlicht die Unterschiede zwischen altem und
neuem System noch weiter (4,49 mmol/l zu 2,49 mmol/l, p < 0,001). Dagegen repräsentiert
der im Stichblut gemessene Cortisolgehalt von 95,17 ng/ml im alten und 105,21 ng/ml im
neuen System aufgrund der Ausschüttungslatenz des Cortisols offensichtlich nicht die
Belastungen während der Zuführung zur Betäubung. Im Zuge der signifikanten (p < 0,05)
Reduzierung des während der Vereinzelung von den Schweinen gezeigten
Vermeidungsverhaltens von durchschnittlich 3,4 Sekunden im alten System auf rechnerisch
0,3 Sekunden im neuen System verkürzte sich auch die Zeitdauer der Vereinzelung im neuen
System um mehr als die Hälfte (altes System = 10 Sekunden, neues System = 4,3 Sekunden,
p < 0,01), der Einsatz von Treibhilfen (Index) reduzierte sich von 17,8 auf 6,3 (p < 0,001)
und die Anzahl der Verhaltenswechsel sank von durchschnittlich 2,7 auf 1 (p < 0,05). Im
Zutrieb verringerten sich im neuen System signifikant die zeitliche Ausdehnung des
Vermeidungsverhaltens (altes System = 10,2 Sekunden, neues System = 4,1 Sekunden, p <
0,01), der Treibhilfeindex (altes System = 76,9, neues System = 28,5, p < 0,001) und die
Anzahl der Verhaltenswechsel (altes System = 6,5, neues System = 3,6, p < 0,01). Auch die
Zutriebsdauer verkürzte sich im neuen System von 26,8 auf 20,2 Sekunden, dieser
Unterschied ist jedoch nicht signifikant.
Die deutliche Reduzierung der physischen und psychischen Belastung während der
Zuführung zur Betäubung im neuen System ist auf dessen bauliche Gestaltung
zurückzuführen. Das beim Bau des neuen Zuführungssystems umgesetzte Prinzip, sich das
Erkundungsverhalten von Schweinen zu Nutze zu machen, hilft, insbesondere im Bereich der
Vereinzelung, Belastungen zu verringern. So fördert die ruhige und schattenfrei hell
beleuchtete Umgebung die Orientierung zum Zutriebseingang hin und ein als zusätzliche
Orientierungshilfe eingesetzter Metallspiegel, der die Anwesenheit eines Artgenossen im
Zutrieb vortäuscht, motiviert die Schweine über das Setzen eines Erkundungsreizes zum
Betreten des Zutriebs. Hierdurch wird eine schonende Behandlung der Schweine durch das
Personal gefördert. Der automatische Transport im Zutrieb senkt den auf den Schweinen
lastenden Anpassungsdruck und ermöglicht es wiederum dem Personal, weitgehend auf den
Einsatz von Treibhilfen zu verzichten.

106
Insgesamt erscheint die bauliche Gestaltung des neuen Zuführungssystems geeignet, das
Wohlbefinden von Schlachtschweinen im Problembereich Zuführung zur Betäubung im
Vergleich zu herkömmlichen Systemen zu verbessern. Es bleibt zu überlegen, ob der in den
letzten Jahren erfolgte Erkenntnisgewinn zur Senkung der prämortalen Belastung von
Schlachttieren durch Einführung der Definition „nach Stand der Technik“ in den § 3 der
Tierschutz-Schlachtverordnung schneller in die Schlachtpraxis umgesetzt werden könnte.

107
7 Summary
Melanie Außel
Stress slaughter-pigs experience in two different conveyance systems to electrical
stunning and the effects on the animals´ welfare
Moving pigs to the stunning point is one of the most important sources of stress in
slaughtering pigs. In many abattoirs it is difficult for the drivers to handle pigs carefully
during separation and access to the restrainer because these areas are often poorly designed.
This may lead to problems which can impair the animals´ welfare. This dissertation
investigates to what extent constructional measures are able to reduce the stress which occurs
during the conveyance to the stunning point. Therefore a conventional conveyance system
and a new one, which had been developed in cooperation with the Institute for Animal
Hygiene, Animal Welfare and Farm Animal Behaviour of the School of Veterinary Medicine
Hannover (TiHo Hannover), were compared.
The investigation included a total of 515 pigs of a German hybrid breeding programme and
was divided in two measuring-periods which both took place in the same slaughter plant. In
the first period 266 pigs were investigated in the conventional system (old system). In the old
system the pigs were sent one by one through a single file race which led to the stunning
point. The preceding separation of pigs from the group took place in a pen which was located
inside the lairage area. After the slaughter plant had been reconstructed the second measuring
period took place. It included 249 pigs which now were moved to the stunning point in the
new developed conveyance system which design considers ethological aspects (new system).
In the new system separation of pigs from the group and access to the restrainer take place in
a special room which is visually and acoustically separated from lairage and slaughtering
section and the access functions automatically: the pigs are transported to the restrainer by a
conveyor belt on which the pigs are standing. In order to judge the extent of a pre-mortal
strain the pigs´ heart rate (Polar® Accurex Plus) and body temperature (Pillbox®-Logger,
intravaginal) were continuously measured and blood samples were taken from the sticking
blood and via venous catheters and they were analysed for norepinephrine, epinephrine,
cortisol and lactate. Additionally the pigs´ behaviour was recorded on video tape.
The results show that in the new system the stress pigs experience during the conveyance to
the stunning point is lower than in the old system. The pigs´ heart rate and body

108
temperature were significantly (p < 0,001, p < 0,05) lower in the new system than in the old
one (new system = 168,4 bpm and 39,4 °C, old system = 189,1 bpm and 39,6 °C).
Additionally the concentration of hormonal and biochemical stress indicators in the plasma of
blood samples which had been gathered via venous catheters immediately after the access was
significantly lower in the new system compared to the old one. The average norepinephrine-
level declined from 8,57 nmol/l to 4,89 nmol/l (p < 0,001), the concentration of epinephrine
was reduced from 2,64 nmol/l to 1,49 nmol/l (p < 0,001) and those of lactate from 2,7 mmol/l
to 1,9 mmol/l (p < 0,001). The concentration of lactate in the sticking blood (old system =
4,49 mmol/l, new system = 2,49 mmol/l, p < 0,001) illustrates further the differences between
the old and the new system. In contrast to that the concentration of cortisol in the sticking
blood (old system = 95.17 ng/ml, new system = 105,21 ng/ml) does obviously not represent
the strain during the conveyance because circulating cortisol levels respond rather less rapidly
to stressful stimuli than the others. As a result of a significant reduction of the extent to which
avoidance behaviour was shown by the pigs during separation (old system = 3,4 seconds,
new system = 0,3 seconds, p < 0,05) the duration of this procedure halved in the new system
(old system = 10 seconds, new system = 4,3 seconds, p < 0,01), the extent of prodding
(index) during separation declined from 17,8 to 6,3 (p < 0,001) and the pigs changed less
frequent their behaviour (old system = 2,7, new system = 1, p < 0,05). During the access in
the new system the extent to which avoidance behaviour was shown (old system = 10,2
seconds, new system = 4,1 seconds, p < 0,01), the extent of prodding (index) (old system =
76,9, new system = 28,5, p < 0,001) and the quantity of behaviour-changes (old system =
6,5, new system = 3,6, p < 0,01) decreased significantly. The duration of access was also
reduced from 26,8 seconds to 20,2 seconds, but this difference is not significant.
The distinct reduction of physical and emotional stress during the conveyance to the stunning
point in the new system can be explained by its constructional design. The new system´s
design realizes the principle of using pigs´ exploratory behaviour. This helps, particulary
during separation, to reduce the strain. So the calm and brightly illuminated environment
encourages orientation to the entrance of the access and a mirror out of metal, which
simulates the presence of a species companion in the access, motivated the pigs to enter the
access by setting a stimulus for exploratory behaviour. This promotes a gentle treatment of
the pigs. The automatical transport during access reduces the pigs´ distress and enables the
drivers to handle the pigs nearly without prodding them.
All in all the constructional design of the new conveyance system seems suitable to improve
the welfare of pigs during the critical procedure of moving them to the stunning point. It is to
consider if the new knowledge about reducing the pre-mortal strain of slaughter animals,
which has been gained in the last years, could be realized in slaughter practice more quickly if

109
the definition „as latest state of engineering“ would be inserted in § 3 of the German
regulation for the protection of animals during slaughter (Tierschutz-Schlachtverordnung).

110
8 Literaturverzeichnis
AHLQUIST, R.P. (1948):A study of the adrenotropic receptors.Am. J. Physiol. 153, S. 586-600
ALLEN, J.P., C.F. ALLEN, M.A. GREER u. J.J. Jacobs (1973):Stress-induced secretion of ACTH.In: BRODISH, A. u. E.S. REDGATE (Hrsg.): Brain-Pituitary-Adrenal Interrelationships.Verlag Karger, Basel, S. 99-127 ARTHUR, A.Z. (1987):Stress as a state of anticipatory vigilance.Percept. Mot. Skiles 64, S. 75-85
AUGUSTINI, C. (1976):EKG- und Körpertemperaturmessungen an Schweinen während der Mast und auf demTransport.Schlachten und Vermarkten 10, S. 255
AUGUSTINI, C., K. FISCHER u. L. SCHÖN (1977):Welche Informationen können unmittelbar vor der Schlachtung erhobene physiologischeMeßwerte über die zu erwartende Fleischbeschaffenheit geben?Fleischwirtschaft 57, S. 1028-1033
AUGUSTINI, C, und K. FISCHER (1982):Physiological reaction of slaughter animals during transport.In: MOSS, R. (Hrsg.): Transport of animals intended for breeding, production and slaughter.Verlag Martinus Nijhoff, Den Haag, Niederlande, S. 125-135
AXELROD, J., u. T.D. REISINE (1984):Stress hormones: their interaction and regulation.Science 22, S. 452-459
BALDI, A., M. VERGA, M. MAFFIL, E. CANALI, D. CHIARAVIGLIO u. C. FERRARI(1989):Effects of blood sampling procedures, grouping and adrenal stimulation on stress responses inthe growing pig.Reprod. Nutr. Develop. 29, S. 95-103
BALDWIN, B.A., und G.B. MEESE (1977):Sensory reinforcement and illumination preference in the domesticated pig.Anim. Behav. 25, S. 497-507

111
BARTON-GADE, P.A. (1996):Live Handling, Stunning and Slaughter of Red Meat Species.In: TAYLOR, S.A., A. RAIMUNDO, M. SEVERINI u. F.J.M. SMULDERS (Hrsg.):Meat Quality and Meat Packaging, Verlag Eccemast, Utrecht, Niederlande, S. 25-42
BARTON–GADE, P.A. (1997):The effect of preslaughter handling on meat quality in pigs.In: CRANWELL, P.D. (Hrsg.): Manipulating pig production VI, Australasian pig scienceAss. Werribee, S. 100-115
BARTON-GADE, P.A., u. L. CHRISTENSEN (1998):Effect of different stocking densities during transport on welfare and meat quality in Danishslaughter pigs.Meat Science 48, S. 237-247
BECKER, B.A., J.A. NIENABER, R.K. CHRISTENSON, R.C. MANAK, J.A. DeSHAZERu. G.L. HAHN (1985a):Peripheral concentrations of cortisol as an indicator of stress in the pig.Am J. Vet. Res. 46, S. 1034-1039
BECKER, B.A., J.A. NIENABER, J.A. DeSHAZER u. G.L. HAHN (1985b):Effect of transportation on cortisol concentrations and on the circadian rhythm of cortisol ingilts.Am J. Vet. Res. 46, S. 1457-1459
BECKER, B.A., H.F. MAYES, G.L. HAHN, J.A. NIEBAUER, G.W. JESSE, M.E. ANDER-SON, H. HEYMANN u. H.B. HEDRICK (1989): Effect of fasting and transportation on va-rious physiological parameters and meat quality of slaughter hogs.J. Anim. Sci. 37, S. 334 – 341
BEKOFF, M. (1997):Tierliebe in der Wissenschaft.In: SOLISTI, K. u. M. TOBIAS (Hrsg.): Ich spürte die Seele der Tiere.Kosmos Verlag, Stuttgart, S. 28-37
BICKHARDT, K., und A. WIRTZ (1986):Kinetics of L-Lactate in pigs. II. Studies in stress resistant and stress susceptible pigs underdifferent metabolic conditions.J. Vet. Med. A 34, S. 377-389
BICKHARDT, K. (1992):Kompendium der Allgemeinen inneren Medizin und Pathophysiologie für Tierärzte.Verlag P. Parey, Berlin, Hamburg, S. 97-108

112
BICKHARDT, K. (1996):Probleme der haltungs- und zuchtbedingten Erkrankungen von Schweinen.Verwirklichung des Tierschutzes in der „Nutztierhaltung.“ Eine gemeinsame Aufgabe vonLandwirtschaft und Tiermedizin. Tagung vom 1.-3. März 1996 in der EvangelischenAkademie Bad Boll
BLIGH, J. (1985):Temperature regulation.In: YOUSEF. M.K. (Hrsg.): Stress physiology of livestock I: basic principles.CRC Press, Boca Raton, Florida, USA, S. 75-96
BLOOM, F.E., A. LAZERSON u. L. HOFSTADTER (1985):Brain, Mind and Behaviour.Verlag Freeman and Company, New York, USA
BOGNER, H., u. A. GRAUVOGL (1984):Verhalten landwirtschaftlicher Nutztiere.Verlag Eugen Ulmer, Stuttgart
BOHUS, B., R.F. BENUS, D.S. FOKKEMA, J.M. KOOLHAAS, C. NYAKAS, G.A. vanORTMERSSEN, A.J.A. PRINS, A.J.H. DE RUITER, A.J.W. SCHEURINGK u. A.B.STEFFENS (1987):Neuroendocrine states and behavioural and physiological stress response.In: DE KLOET, V.M. WIEGANT u.D. DE WIED (Hrsg.): Progress in brain research 72,Neuropeptids and brain function. Verlag Elsevier, Amsterdam, S. 55-70
BORELL, E. von, u. J. LADEWIG (1986):Möglichkeiten der Erfassung von chronischen Belatungsreaktionen beim Schwein mit Hilfevon Nebennierenrinden-Funktionsprüfungen und ethologischen Merkmalen.In: Aktuelle Arbeiten zur artgemäßen Tierhaltung 1985, KTBL-Schrift 311, S. 30-36
BORELL, E. von, u. J. LADEWIG (1989):Altered adrenocortical response to acute stressors or ACTH in intensively housed pigs.Dom. Anim. Endocr. 64, S. 299-309
BRAATHEN, O.S. (1980):Ideas about methods and equipment for transport, lairage, stunning and debleeding in order toreduce the incidence of PSE-meat.Norwegian Meat Res. Lab. Rep., Oslo, Norway
BRIESE, A., K. von HOLLEBEN u. M. DAYEN (1997a):Der Tierschutzpreis 1996 des Landes Niedersachsen. Ergebnisse des Landeswettbewerbes zurFörderung einer tierschonenden Behandlung von Schweinen am Schlachtbetrieb (Teil 1).Fleischwirtschaft 77 (7), S. 598-603

113
BRIESE, A., K. von HOLLEBEN u. M. DAYEN (1997b):Der Tierschutzpreis 1996 des Landes Niedersachsen. Ergebnisse des Landeswettbewerbes zurFörderung einer tierschonenden Behandlung von Schweinen am Schlachtbetrieb (Teil 2).Fleischwirtschaft 77 (8), S. 721-725
BROOKE, M.H., u. K.K. KAISER (1970):Muscle fiber types: how many and what kind?Arch. Neurol. 23, S. 369
BROOM, D.M. (1991):Animal welfare: Concept and measurement.J. Anim. Sci. 69, S. 4167-4175
BROOM, D.M., u. K.G. JOHNSON (1993)Stress and animal welfare.Verlag Chapman and Hall, London, UK
BROOM, D.M. (1995):Quantifying pig welfare during transport using physiological measures.In: SCHÜTTE, A. (Hrsg.): Proc. EU-Seminar „New information on welfare and meat qualityof pigs as related to handling, transport and lairage conditions.“Mariensee, 29-30.06.1995, S. 3-10
BROOM, D.M. (1996):Animal welfare defined in terms of attempts to cope with the environment.Acta Agric. Scand. Sect. A, Animal Sci. Suppl. 27, S. 22-28
BROOM, D.M. (1997):Animal behaviour as an indicator of animal welfare in different housing and managementsystems.In: Proc. 9th International Congress in Animal Hygiene (ISAH).Helsinki, Finnland, 17.-21.8. 1997, S. 371-378
BROWN, S.N., P.D. WARRISS, G.P. NUTE, J.E. EDWARDS u. T.G. KNOWLES (1998):Meat quality in pigs subjected to minimal preslaughter stress.Meat Science 49, 3, S. 257-265
BRÜSSOW, K.-P., J. BERGFELD u. G. PARCHOW (1981):Über mehrjährige Erfahrungen zur Blutgewinnung durch intravenöse Dauerkatheter beimSchwein.Monatshefte für Veterinärmedizin 36, S. 300-303
BUCHHOLTZ, C. (1993):Das Handlungsbereitschaftsmodell - Ein Konzept zur Beurteilung und Bewertung vonVerhaltensstörungen.In: Leiden und Verhaltensstörungen bei Tieren, Tierhaltung 23.Verlag Birkhäuser, Basel, S. 93-109

114
BUCHHOLTZ, C. (1994):Verhaltensstörungen bei Versuchstieren als Ausdruck schlechter Befindlichkeit.Tierärztliche Umschau 49, S. 532-538
BUCHHOLTZ, C. (1996):Motivation als Regulationsinstanz für Verhalten und Befindlichkeit – Kriterien fürErheblichkeit von Leiden.Tierärztl. Umschau 51, S. 142-146
BÜHLER, H.U., M. DA PRADA, W. HAEFELY u. G.B. PICOTTI (1978):Plasma adrenaline, noradrenaline and dopamine in man and different animal species.J. Physiol. 276, S. 311-320
BURCHFIELD, S.R. (1979):The stress response: A new perspective.Psychsom. Med. 41, S. 661 - 672
CANNON, W.B. (1915):Bodily changes in pain, hunger, fear and rage: an account of recent researches into thefunction of emotional excitement.Verlag Appleton, New York, USA
CANNON, W.B. (1928):Die Notfallfunktion des Sympathiko-Adrenalen Systems.Ergebn. Physiol. 27, S. 380-406
CARLSTEAD, K. (1986):Predictability of feedings: its effects on agonistic behaviour and growth in grower pigs.Applied Animal Behaviour Science 16, S.25-38
CHRISTENSEN, L,. und P. BARTON-GADE (1997): Neue Entwicklungen beim Handling von Schweinen in Schlachtbetrieben. Fleischwirtschaft 77, S. 604-607
CHROUSOS, G.P., u. P.. GOLD (1992):The Concepts of Stress and Stress System Disorders – Overview of Physical and BehaviouralHOMOEOSTASIS.J. Am. Med. Ass., 267, S. 1244-1252
COCKRAM, M.S., u. M.A. MITCHELL (1999):Role of research in the formulation of „rules“ to protect the welfare of farm animals duringroad transportation. In: RUSSEL, A.J.F., C.A. MORGAN, C.J. SAVORY, M.C. APPLEBY u. T.C.J.LAWRENCE (Hrsg.): Farm animal welfare – who write the rules? Occasional Publication 23.British Society of Animal Science 1999.

115
DALIN, A.M., L. NYBERG u. L. ELIASSON (1988):The effect of transportation/relocation on cortisol, CBG and induction of puberty in gilts withdelayed puberty.Acta vet. Scand. 29, S. 207-218
DALIN, A.M., U. MAGNUSSON, J. HÄGGENDAHL u. L. NYBERG (1993):The effect of transport stress on plasma levels of catecholamines, cortisol, corticosteroidbinding globulin, blood cell count, and lymphocyte proliferation in pigs .Acta Vet. Scand. 34, S. 59-68
DANTZER, R., M. ARNONE u. P. MORMEDE (1980):Effects of frustration on behaviour and plasma corticosteroids.Physiology and Behaviour 24, S. 1-4
DANTZER, R., u. P. MORMEDE (1981):Can physiological criteria be used to assess welfare in pigs?In: SYMBESMA, W. (Hrsg.): The welfare of pigs.Verlag Martinus Nijhoff, Den Haag, Niederlande, S. 53-73
DANTZER, R., u. P. MORMEDE (1983):De-arousal properties of stereotyped behaviour: evidence from pituitary-adrenal correlates inpigs.Applied Animal Ethology 10, S. 233-244
DANTZER, R., u. P. MORMEDE (1985):Stress in domestic animals: a psychoneuroendocrine approach.In: MOBERG, G.P. (Hrsg.): Animal stress.Am. Physiol. Soc., Bethesda, Maryland, USA, S. 81-96
DANTZER, R. (1989):Neuroendocrine correlates of control and coping.In: STEPTOE, A. u. A. APPLES (Eds.): Stress, Personal Control and Health.Verlag John Wiley & Sons, Brüssel, Belgien, S. 277-294
DANTZER, R. (1994): Animal welfare methodology and criteria.Rev. sci. tech. Off. int. Epiz., 13, 1, S. 291-302
DAWKINS, M.S. (1990):From an animal´s point of view: motivation, fitness and animal welfare.Behavioural and Brain Sciences 13, S. 1-61
DAYEN, C. (1989):Diskussionsbeitrag zum Referat von Troeger und Woltersdorf :„Die Elektrobetäubung vonSchlachtschweinen“.Dtsch. Tierärztl. Wochenschrift 96, S. 103

116
DeBOER, S.F., R. DeBEUN u. J.L. SLANGEN (1990):Dynamics of plasma catecholamines and corticosterone concentrations during reinforced andextinguished operant behaviour in rats.Physiology and Behaviour 47, S. 691-698
DeROTH, L., L. VERMETTE u. N. LARIVIERE (1989):Blood catecholamines in response to handling in normal and stress-susceptible swine.Appl. Anim. Behav. Sci. 22, S. 255-260
DÖCKE, F. (1994):Hypothalamus-Hypophysensystem.In: F. DÖCKE (Hrsg.): Veterinärmedizinische Endokrinologie, 3. AuflageVerlag Gustav Fischer, Jena, S. 131-175
DÖCKE, F., und A. KEMPER (1994): Nebennierenmark.In: F. DÖCKE (Hrsg.): Veterinärmedizinische Endokrinologie, 3. AuflageVerlag Gustav Fischer, Jena, S. 357-378
DOMEL, G., u. W. LEISTNER (1977):Grundsätzliche Betrachtungen und neue Untersuchungsergebnisse zur Fleischqualität beimSchwein.Monatshefte für Veterinärmedizin 16, S. 407-411
DUNCAN, I.J.H., u. M.S. DAWKINS (1983):The problem of assessing „well-being“ and „suffering“ in farm animals.In: SMIDT, D. (Hrsg.): Indicators Relevant to Farm Animal Welfare.Verlag Martinus Nijhoff, Boston, USA, S. 13-24
ELDRIDGE, G.A., R.D. WARNER, C.L. BALL u. E. NANTHAN (1992):Influence of previous experience and pre-slaughter behaviour on pig meat quality.38th ICOMST, Clermont-Ferrand, Frankreich
ENGELHARDT, W. von (1963):Untersuchungen am Schwein über die Systolen- und Diastolendauer des Herzens und überden Blutdruck in der Ruhe und während der Erholung nach körperlicher Belastung.Zentralbl. Veterinärmedizin A, 10, S. 39-50
EWBANK, R., u. G.B. MEESE (1971):Aggressive behaviour in groups of domesticated pigs on removal and return of individuals.Anim. Prod. 13, S. 685-693
FERRERIA , S.H., u. J.R. VANE (1967):Halflifes of Peptids and Amines in the Circulation.Nature 215, S. 1237-1240

117
FISCHER, K. (1995):Schlachttiertransport: Auswirkungen, Schwachstellen und Maßnahmen.Fleischwirtschaft 75, S. 790-796
FLOß, M., M. MARAHRENS, B. NOWAK, F. FELDHUSEN u. J. HARTUNG (1997):Der Zutrieb von Schlachtschweinen zur Elektrobetäubung – Ein Systemvergleich.Fleischwirtschaft 77, S. 919-922
FLOß, M. (1999):Belastungsantwort von Schlachtschweinen in zwei unterschiedlichen Zuführungssystemen zurElektrobetäubung.Diss. Tierärztliche Hochschule Hannover, Institut für Tierhygiene und Tierschutz
FORSLID, A., u. O. AUGUSTINSSON (1988):Acidosis, hypoxia and stress hormone-release in response to one-minute inhalation of 80 %CO2 in swine.Acta physiol. Scand. 132, S. 223-226
FRASER, D., J.S.D. RITCHIE u. A.F. FRASER (1975):The term stress in a veterinary context.Brit. Vet. J., 131, S. 653-662
GALOSY, R.A., L.K. CLARKE u. J.H. MITCHELL (1979):Cardiac changes during behavioural stress in dogs.Am. J. Physiol. 236, S. H750-H758
GARRICK, N.A., J.L. HILL, F.G. SZELLE, T.P. TOMAL, P.W. GOLD u. D.L. MURPHY(1987):Corticotropin-releasing factor: a marked circadian rhythm in primate cerebrospinal fluidpeaks in the evening and its inversely related to the cortisol circadian rhythm.Endocrinology 121, S. 1329-1334
GEERS, R. H. VILLE, S. JANSSENS, V. GOEDSEELS, K. GOOSSENS, G. PARDUYNS,J. VAN BAEL, L. BOSSCHAERTS und H. HEYLEN (1992):Changes of body temperatures of piglets as related to halothane sensitivity and treatmillexercise.J. Therm. Biol. 17, 2, S. 125-128
GEERS, R., T. VAN SCHIE, H. VILLÉ, H. GERARD, S. JANSSENS, G. NACKAERTS, E.DECUYPERE u. J. JOURQUIN (1994):Transport of pigs different with respect of halothane gene: stress assessment.J. Anim. Sci. 72, S. 2552-2558

118
GEVERINK, N.A., R.H BRADSHAW, E. LAMBOOY u. D.M. BROOM (1995):Handling of slaughter pigs in lairage: Behavioural and physiological effects.in: A. SCHÜTTE (Ed.): Proc. EU-Seminar „New information on welfare and meat quality ofpigs as related to handling, transport and lairage conditions.“ Mariensee, 29.-30.06.1995, S.145 – 152
GEVERINK, N.A., B. ENGEL, E. LAMBOOIJ u. V.M. WIEGANT (1996):Observations on behaviour and skin damage of slaughter pigs and treatment during lairage.Appl. Anim. Behav. Sci. 50, S. 1-13
GOLDSTEIN, D.S. (1987):Stress-induced activation of the sympathetic nervous system.Baillière´s Clin. Endocr. Metab. 1.2, S. 253-278
GONYOU, H.W, P.H. HEMSWORTH und J.L. BARNETT (1986):Effect of frequent interactions with humans on growing pigs.Appl. Anim. Behav. Sci. 16, S. 269-278
GRANDIN, T. (1981):Design of animal handling systems to reduce stress.Sonderdruck
GRANDIN, T. (1982):Pig behavior studies applied to slaughter-plant design. Appl. Anim. Ethol. 9, S. 141-151
GRANDIN, T. (1983):Welfare requirements of handling facilities. In: BAXTER, S.H., M.R. BAXTER u. J.A.D.MacCORMACK (Hrsg.): Farm Animal Housing and WelfareVerl. Mart. Nijhoff, Dordrecht, Niederlande, S. 137-149
GRANDIN, T. (1986):Minimize stress in pig handling.Laboratory Animal 15, S. 17-21
GRANDIN. T. (1988):Hog psychology: an aid in handling.Agic. Practice 9, 4, S. 22-26
GRANDIN. T. (1991):Handling problems caused by excitable pigs.37 th Int. Congr. Meat Sci. Techn., Kulmbach
GRANDIN. T. (1994):Farm animal welfare during handling, transport and slaughter.JAVMA 204, 3, S. 1023-1231

119
GRAUVOGL, A. (1970):Angewandte Ethologie beim Hausschwein.Dt. Tierärztl. Wschr. 77, S. 126-132
GRAUVOGL, A. (1988):Des Schlachtschweins letzte Reise.SUS 36, S. 78-80
GRAY, J.A. (1982):The neuropsychology of anxiety.University Press, New York.
GREGORY, N.G. (1996):Welfare and hygiene during preslaughter handling.Meat Science 43, Supplement, S. S35-S46
GREGORY, N.G. (1998):Animal welfare and meat science.CAB International, Wallingford
GRÜN, E., H. HÜLLER u. H.G. MÖCKEL (1973):Dauerkatheter am Schweineohr.Monatshefte für Veterinärmedizin 28, S. 263-265
HANNON, J.P. (1990):Normal Physiological Values for Conscious Pigs Used in Biomedical Research.Laboratory Animal Science 40, S. 293-298
HANSEN, L., L. BLAABJERG, E. JORGENSEN u. P.A. BARTON-GADE (1989):Effect of group size, floortype and straw on aggressive behaviour, skin damage and meatquality.Proc. 40th Animal Meeting of E.A.A.P., Dublin, Ireland, Vol. I, S. 497-498
HANSEN, L., P.A. BARTON-GADE u. P.P. VORUP (1991):Effect of mixing peaceful or aggressive pigs at abattoirs on behaviour and quality.Proc. Int. Congr. Soc. Vet. Ethol., Edinburgh, Scotland. In: APPLEBY, M.C., R.I. HALL,J.C. Petrick u. S.M. RUTTER (Hrsg.): Applied Animal Behaviour, Universities Federation ofAnimal Welfare, S. 88
HEINZE, P.H., u. G. MITCHELL (1989):Stress resistant and stress susceptible landrace pigs: comparison of blood variables afterexposure to halothane or exercise on a treadmill.Veterinary Record 124, S. 163-179

120
HELLMANN, L., F. NAKADA, J. CURTI, E.D. WEITZMAN, J. KREAM, H.ROFFWARG, S. ELLMAN, D.K. FUKUSHIMA u. T.F. GALLAGHER (1970):Cortisol is secreted episodically by normal man.J. Clin. Endocrinol. 30, S. 411-422
HEMSWORTH, P.H., J.L. BARNETT u. C. HANSEN (1987):The influence of handling by humans on the behaviour, growth and corticosteroids of youngpigs.Appl. Anim. Behav. Sci. 17, S. 245-252
HEMSWORTH, P.H., J. VERGE und G.J. COLEMAN (1996):Conditioned approach-avoidance responses to humans: the ability of pigs to associate feedingand aversive social experiences in the presence of humans with humans.Appl. Anim. Behav. Sci. 50, S. 71-82
HENRY, J.P., u. P.M. STEPHENS (1977):Stress, health and the social environment. A sociobiological approach to medicine.Verlag Springer, Heidelberg
HENRY, J.P., P.M. STEPHENS u. D.L. ELY (1986):Psychosocial hypertension and the defence and defeat reactions.J. Hypertens. 4, S. 687-697
HOENDERKEN, R. (1976):Ein verbessertes System für das Treiben von Schlachtschweinen zum Restrainer.Fleischwirtschaft 56, S. 838-839
HOENDERKEN, R. (1979):Zur Elektro- und Kohlendioxid-Betäubung von Schweinen.Mittbl. BAFF 64, S. 3647-3648
HOLLEBEN, K. von (1994):Anforderungen an das tierschutzgerechte Verladen, Transportieren, Betäuben und Schlachtenvon Rindern und Schweinen.Veto 34, S. 13-16
HOLLEBEN, K. von, u. M. von WENZLAWOWICZ (1995):Anforderungen an das treibende Personal sowie Vorschläge für Vorgehensweisen inkritischen Situationen.RFL 47, S. 148-152
HOLST, D. von (1977):Social stress in tree-shrews problems, results and goals.Journal of Comparative Physiology 120, S. 71-86

121
HOLST, D. von, u. K.R. SCHERER (1988):Stress.In: IMMELMANN, K., K.R. SCHERER, C. VOGEL u. P. SCHMOOCK (Hrsg.):Grundlagen des Verhaltens.Verlag Gustav Fischer, Jena, S. 289-327
HOLZBAUER, M., u. D.F. SHARMAN (1972):The distribution of catecholamines in vertebrates.In: BLASCHKO, H. u. E. MUSCHOLL (Hrsg.): Catecholamines. Handb. Exp. Pharmakol.XX, Springer Verlag, Berlin-Heidelberg-New York, S. 110-185
HOLZER, A., P. MATZKE u. E. LITTMANN (1992):Schweine wollen schonend behandelt werden - Ergebnisse einer Untersuchung über denEinfluß von Bedingungen beim Transport und vor der Schlachtung auf dieFleischbeschaffenheit beim Schwein an sechs Schlachthöfen Bayerns.Akt. Inf. u. Hinw. der BLT Grub 4, S. 1-15
HONKAVAARA, M. (1989):Influence of carcass temperature, glycogenolysis and glycolysis 45 min postmortem on thedevelopment of PSE pork.J. Agric. Sci. Finland 61, S. 433-440
HOSMAN, L. (1972):Das Verhalten von sus scrofa im Vergleich zum Verhalten des Hausschweins (I).Der Tierzüchter 17, S. 496-498
HUETHER, G. (1996):The central adaptation syndrome: psychosocial stress as a trigger for adaptive modificationsof brain structure and brain function.Prog. Neurobiol. 48, S. 569-612
HUNTER, E.J., C.M. WEEDING, H.J. GUISE, T.A. ABBOTT u. R.H.C. PENNY (1994):Pig welfare and carcasse quality: a comparison of the influence of slaughter handling systemsat two abattoirs.Vet. Rec. 135, S. 423-425
HUNTER,E.J., H.L. RICHES, H.J. GUISE u. R.H. PENNY (1997):The behaviour of pigs in lairage in relation to their post-weaning management: Results of apostal survey.Anim. Welf. 6, S. 139-144
HUTSON, G.D., u. M.L. BUTLER (1978):A self-feeding sheep race that works.J. Agric. 76, S. 335-336

122
INGRAM, D.L., u. M.J. DAUNCEY (1985):Thermoregulatory behaviour.In: YOUSEF. M.K. (Hrsg.): Stress physiology of livestock I: basic principles.CRC Press, Boca Raton, Florida, USA, S. 97-108
JENSEN-WEARN, M., und L. NYBERG (1993):Valuable indicators of physical stress in porcine plasma.J. Vet. Med. A 40, S. 321-327JOHANSSON, G., K. OLSSON, J. HÄGGENDAL, L. JÖNSSON, K. THOREN-TOLLING(1982):Effects of stress on myocardial cells and blood levels of catecholamines in normal andamygdalectomized pigs.Can. J. Comp. Med. 46, S. 176-182
KANT, G.J., T. EGGLESTON, L. LANDMAN-ROBERTS, C.C. KENION, G.C. DRIVER u.J.C. MEYERHOFF (1985):Habituation to repeated stress is stressor specific.Pharmacol. Biochem. Behav. 22, S. 631-634
KARLSON, P. (1988):Kurzes Lehrbuch der Biochemie für Mediziner und Naturwissenschaftler.Verlag Georg Thieme, Stuttgart
KEMPER, A. (1977):Organismus-Umwelt- Beziehungen und neuroendokrine Regulation beim Hausschwein.Dissertation B., Humboldt-Universität Berlin
KIRSCH, K. (1996):Leistungsphysiologie.In: KLINKE, R. u. S. SILBERNAGEL (Hrsg.): Lehrbuch der Physiologie.Verlag Georg Thieme, Stuttgart, New York, S. 509-530
KLAWITTER, H.J. (1971):Klinische Beurteilung von Schlachtschweinen bei unterschiedlichen Transportbedingungen.Vet.med. Dissertation, Hannover.
KLEIBS, M. (1992):Das Treiben von Schlachtschweinen bei unterschiedlichen Beleuchtungsstärken bzw.Gangmaßen und die bioindikatorische Nutzung des Lauf- und Erkundungsverhaltens zurartgemäßen Gestaltung und Triftregulierung.Diss. Univ. Leipzig, Agrarw. Fak., WB Haustiergenetik und Züchtungsmethodik
KLEMCKE, H.G., J.A. NIENABER u. G.L. HAHN (1989):Plasma adrenocorticotropic hormone and cortisol in pigs: effects of day on basal and stressor-altered concentrations.Proc. Soc. Exp. Biol. Med. 190, S. 42-53

123
KLONT, R.E., und E. LAMBOOIJ (1995a):Influence of preslaughter muscle temperature on muscle metabolism and meat quality inanesthetized pigs of different halothane genotypes.J. Anim. Sci. 73, S. 96-107
KLONT, R.E., und E. LAMBOOIJ (1995b):Effects of preslaughter muscle exercise on muscle metabolism and meat quality studied inanesthetized pigs of different halothane genotypes.J. Anim. Sci. 73, S. 108-117
KNOWLES, T.G., S.N. BROWN, J.E. EDWARDS u. P.D. WARRIS (1998):Ambient temperature below which pigs should not be continously showered in lairage.Veterinary Record 143, S. 575-578
KONARSAKA, M., R.E. STEWART u. R. Mc CARTY (1990):Predictability of chronic intermittent stress: effects on sympathetic-adrenal medullaryresponses of laboratory rats.Behav. Neurol. Biol. 53, S. 231-243
KOOLHAAS, J.M., u. B. BOHUS (1989):Social control in relation to neuroendocrine and immunological responses.In: STEPTOE, A. u. A. APPLES (Hrsg.): Stress, Personal Control and Health.Verlag John Wiley & Sons, Brüssel, Belgien, S. 295-304
KRAFT, W. (1973):Das Verhalten einiger Plasma-Enzyme und –Elektrolyte sowie von Glukose, unverestertenFettsäuren und Laktat bei Schweinen während der sich an den Transport anschließendenAufstallung.Zentralblatt für Veterinärmedizin A 20, S. 357-369
LADEWIG, J. (1987):Endocrine aspects of stress: evaluation of stress reactions in farm animals.In: Wiepkema, P.R. u. P.W.M. van ADRICHEM (Hrsg.): Biology of stress in farm animals:an integrative approach.Verlag Mart. Nijhoff, Dordrecht, Niederlande, S. 13 – 25
LADEWIG, J., u. E. von BORELL (1988):Ethological methods alone are not sufficient to measure the impact of environment on animalhealth and animal well-being.Proc. Int. Congr. Appl. Ethol., Skara, Schweden, S. 95-102
LADEWIG, J., u. D. SMIDT (1989):Behaviour, episodic secretion of cortisol and adrenocortical reactivity in bulls subjected totethering.Horm. Behav. 23, S. 344-360

124
LADEWIG, J. (1994):Streß. In: F. DÖCKE (Hrsg.): Veterinärmedizinische Endokrinologie, 3. AuflageVerlag Gustav Fischer, Jena, S. 379-398
LAMBOOIJ, E., N.A. GEVERINK, D.M. BROOM u. R.H. BRADSHAW (1995):Quantification of pig welfare by behavioural parameters.In: SCHÜTTE, A. (Hrsg.): Proceedings EU-Seminar „ New information on welfare and meatquality of pigs as related to handling, transport and lairage conditions.“Mariensee, 29.-30.06.1995, S. 13-19
LAUBE, R.-B. (1989):Untersuchungen zur Ethologie des Schweines beim Treiben im erweiterten prämortalenZeitraum.Tierzucht 43, S. 541-543
LAUBE, R.B., J. LIPPMANN u. D. SCHÄFFER (1995):Kann mittels Attraktivreizen bei Schlachtschweinen in Ruhebuchten ein erhöhter Anteilruhender Tiere erreicht werden?In: Aktuelle Arbeiten zur artgemäßen Tierhaltung 1995/1996, KTBL-Schrift, S. 36-47
LAUBE, R.-B. (1996):Ethologische Untersuchungen zur artgerechten prämortalen Haltung und zum Tierschutz beiSchlachtschweinen.Abschlußbericht zum Forschungsauftrag 91 – HS 021, Bundesministerium für Ernährung,Landwirtschaft und Forsten
LAUBE, R.-B., und B. SCHULZE (1992): Erkundungsverhalten von Schlachtschweinen im sozialen Kontext bei der Haltung vor demSchlachten. Aktuelle Arbeiten zur artgemäßen Tierhaltung, KTBL-Schrift 351, S. 188-198
LAUBE, R.-B., J. LIPPMANN, M. KLEIBS u. L. SCHÖBERLEIN (1993):Grenzen der Verhaltensanpassung von Schlachtschweinen auf dem Wege bis zur Betäubung.Koll. "Adaptation bei Nutztieren" und "Neue Aspekte in der Züchtung", Dummersdorf 17./18.Mai 1993
LAZARUS, R.S. (1966):Psychological stress and coping.Verlag Mc Graw Hile, New York, USA
LAZARUS, R.S., u. R. LAUNIER (1978):Stress-related transactions between person and environment.In: PERVIN, L.A. u. M. LEWIS (Hrsg.): Perspectives in international psychology.Verlag Plenum Press, New York, USA, S. 287-337

125
LAZARUS, R.S., u. S. FOLKMAN (1984):Stress appraisal and coping.Verlag Springer, New York, USA
LEVINE, S., C. COE u. S.G. WIENER (1989):Psychoneuroendocrinology of stress: a psychobiological perspective.In: BRUSH, F.R. u. S. LEVINE (Hrsg.): Psychoendocrinology.Verlag Academic Press, San Diego, USA, S. 341-377
LEVINE, S., u. G.D. COOVER (1976):Environmental control of suppression of the pituitary adrenal system.Physiology and Behaviour 17, S. 35-37LIDELL, H.S., W.T. JAMES u. O.D. ANDERSON (1934):The comparative physiology of the conditioned motor reflex. Based on experiments with thepig, dog, sheep, goat and rabbit.Psychol. Monogr. 11, S. 89-91
LINDNER, H.R. (1964):Comparative aspects of cortisol transport: lack of firm binding to plasma proteins in domesticruminants.J. Endocrinol. 28, S. 301-320
LINDNER, K., T. RAABE u. H. GÜRTLER (1996):Einfluss von Belastungssituationen auf dem Transport auf die Plasmakonzentration anCatecholaminen und Stoffwechselparameter im Blutplasma von Schlachtschweinen.Proc. Wiss. Kolloq. "Schlachttierbelastung und Produktsicherheit", Univ. Leipzig, Vet. Med.
LÖSCHER, W. (1994): Sympathikus.In: LÖSCHER, W., F.R. UNGEMACH u. R. KROKER (Hrsg.): Grundlagen derPharmakotherapie bei Haus- und Nutztieren. 2. AuflageVerlag Paul Parey, Berlin und Hamburg
LONG, V.P., u. P.V. TARRANT (1990):The effect of pre-slaughter showering and post-slaughter rapid chilling on meat quality inintact pork sides.Meat Science Vol.27, Nr. 3
LOON, G.R. van, R. KVETNANSKY u. J. AXELROD (1988):Stress: Neurochemical and Humoral Mechanisms.Verlag Gordon & Breach, New York, USA
LORENZ, K. (1953):So kam der Mensch auf den Hund.Verlag Borotha-Schoeler, Wien

126
LORZ, A. (1973):Kommentar. TierschutzgesetzVerlag CH Beck, München
LUMPKIN, M.D. (1987):The regulation of ACTH secretion by IL-1.Science 238, S. 452-454
LYHS, I. (1986):Grundaktivität und Leistung landwirtschaftlicher Nutztiere.Monatshefte für Veterinärmedizin 41, S. 109
MAIER, S.B., u. M.E.P. SELIGMAN (1976):Learned helplessness: theory and evidence.Journal Experimental Psychology (General) 105, S. 3-46
MARAHRENS, M., C. STEFFENS, H. HARTMANN, K.H. HOPPENBROCK,F. FELDHUSEN u. J. HARTUNG (1995a):Auswirkungen von Rangordnungskämpfen im Schlachttierstall auf Herzfrequenzen undFleischqualität bei Schweinen.Dtsch. Veterinärmed. Gesellsch., 21. Kongress zum Leitthema „Aktuelle Forschung“,21. - 24. März 1995, Bad Nauheim, S. 53 - 59
MARAHRENS, C. STEFFENS, M. F. FELDHUSEN u. J. HARTUNG (1995b): Ruhe und Belastungsindikatoren im Wartestall und ihre Bedeutung für die Fleischqualitätbeim Schwein.Proc. 36. Arbeitstagung des Arbeitsgebietes "Lebensmittelhygiene" der DeutschenVeterinärmedizinischen Gesellschaft, Teil I, Garmisch-Partenkirchen, S. 152-160
MARAHRENS, M., B. NOWAK, F. FELDHUSEN u. J. HARTUNG (1997):Belastungen von Schweinen im Wartestall sowie beim Zutrieb zur Elektrobetäubung und ihreAuswirkungen auf die Fleischbeschaffenheit.Fleischwirtschaft 77, 8, S. 717-720
MARTIN, P., u. P. BATESON (1986):Recording methods.In: MARTIN, P. u. P. BATESON (Hrsg.):Measuring behaviour, Verlag Cambridge University Press, Cambridge, New York, USA, S.48-69
MASON, J.W. (1968):A review of psychoendocrine research on the pituitary adrenal cortical system.Psychosom. Med. 30, S. 576-607
MASON, J.W. (1971):A re-evaluation of the concept of „non-specificity“ in stress theory.J. Psych. Res. 8, S. 323-333

127
MAYES, H. F., u. G.W. JESSE (1980):Heart rate data of feeder pigs.Amer. Soc. Agric. Eng., Technical Paper No. 80-4023, St. Joseph, Michigan, USA
McBRIDE, G., J.W. JAMES u. N. HODGENS (1964):Social behaviour of domestic animals.IV.Growing pigs.Anim. Prod. 6, S. 129-139
McCARTY, R., R. KVETNANSKY u. I.J. KOPIN (1981):Plasma catecholamines in rats: daily variations in basal levels and increments in response tostress.Physiol. Behav. 26, S. 27-31
McCARTY, R., K. HORWATT u. M. KONARSAKY (1988):Chronic stress and sympathetic-adrenal medullary responsiveness.Soc. Sci. Med 26, 3, S. 333-341
MENCH, J.A., u. G.J. MASON (1997):Behaviour.In: APPLEBY, M.C. u. B.U. HUGHES (Hrsg.): Animal welfare.CAB International, New York, USA
MEYER, K. (1970):Untersuchungen über Transportschäden und Transportverluste bei mit Lastkraftwagen zurSchlachterei transportierten Schweinen.Vet. med. Dissertation, Hannover.
MICKWITZ, G. von, u. H. GRUND (1970):Telemetrische Verlaufsuntersuchungen umweltbedingter Herzfrequenzänderungen beimSchwein.Dtsch. Tierärztl. Wochenschrift 77, S. 136-138
MICKWITZ, G. von (1982):Various transport conditions and their influence on physiological reactions. In: MOSS, R. (Hrsg.): Transport of animals intended for breeding, production and slaughter.Verlag Martinus Nijhoff, Den Haag, Niederlande, S. 45-54
MICKWITZ, G. von (1990):Anwendung von CO2 zur Betäubung von Schlachtschweinen.Fleischwirtschaft 70, S. 1161-1166
MICKWITZ, G. von, u. A. BRIESE (1994)Ohne Tierschutz keine optimale Fleischqualität.Allg. Fleischerzeitung 5, S. 6-8

128
MONIN, G., E. LAMBOOY und R. KLONT (1995):Influence of temperature variation on the metabolism of pig muscle in situ and after excision.Meat Science 40, S. 149-158MÜLLER, C., J. LADEWIG, H.H. THIELSCHER u. D. SMIDT (1989):Behaviour and heart rate of heifers housed in tether stanchions without straw.Physiol. Behav. 46, S. 751-754
MUNCK, A., P.M. GUYRE u. N.J. HOLBROOK (1984):Physiological functions of glucocorticoids in stress and their relation to pharmacologicalactions.Endocrine Rev. 5, S. 25-44
MURPHY, L.B. (1978a):A review of animal welfare and intensive animal production.Report of Queensland Department of Primary Industries, Poultry Section, Brisbane,Queensland, Australia
MURPHY, L.B. (1978b):The practical problems of recognising and measuring fear and exploration behaviour in thedomestic fowl.Anim. Behaviour 26, S. 422-431
NEUBERT, E., H. GÜRTLER, M. STEINBERG u. K. HÖRÜGEL (1996):Untersuchungen zur Catecholaminkonzentration im Stichblut und zur Fleischqualität beimSchwein.Fleischwirtschaft 76, S. 961-962
NICOL, C.J. (1996):Farm animal cognition.Anim. Sci. 62, S. 375-391
NIEZGODA, J., S. BOBEK, D. WRONSKA-FORTIMA u. E. WIERZCHOS (1993):Response of Sympatho-Adrenal-Axis and Adrenal Cortex to Short-Term Restraint Stress inSheep.J.Vet.Med. A 40, S. 631-638
NOWAK, B. (1998):Auswirkungen von prämortalen Belastungen und Elektrobetäubungsbedingungen auf dasAuftreten von Muskelblutungen beim Schwein.Diss. Tierärztliche Hochschule Hannover, Zentrum für Lebensmittelwissenschaften
NOWAK, B., S. WENZEL u. J. HARTUNG (2000):Einfluß von Streßbelastungen vor der Schlachtung auf die Fleischqualität beim Schwein.Deutsche Veterinärmedizinische Gesellschaft, Fachtagung „Schweinekrankheiten“ am 2./3.März 2000 in Hannover

129
NOWAK, B., S. ELLERBROCK, A. ANDREAE, K.-H. WALDMANN u. J. HARTUNG(2001):Möglichkeiten der EEG-Ableitung beim Schwein während der Betäubung im Schlachtbetrieb.In: Proc. 42. Arbeitstagung des Arbeitsgebietes „Lebensmittelhygiene“ der DeutschenVeterinärmedizinischen Gesellschaft (DVG) am 24.-28.9. 2001 in Garmisch-Partenkirchen,im Druck
NYBERG, L., K. LINDSTRÖM, I. EDFORS-LILJA u. M. RUNDGREN (1988):Effects of transport stress on concentrations of cortisol, corticosteroid-binding globulin andglucocorticoid receptors in pigs with different halothane genotypes.J. Anim. Sci. 66, S. 1201-1211OBRIST, P.A., J.E. LAWLER, J.L. HOWLER, K.W. SMTHSON, P.L. MARTIN, u. J.MANNING (1974):Sympathetic influences on cardiac rate and contractility during acute stress in humans.Psychophysiology 11, S. 405-427
OBRIST, P.A., K.C. LIGHT, A.W. LANGER, A.G. GRIGNDO u. J.P. KOEPKE (1982):Behavioural-cardiovascular interaction.In: SMITH, O.A., R.A. GALOSY u. S.M. WEISS (Hrsg.): Circulation, neurobiology andbehaviour.Verlag Academic Press, Amsterdam, Niederlande
OLIVERIO, A. (1987):Endocrine aspects of stress: Central and peripheral mechanisms.In: WIEPKEMA, P.R. u. P.V.M. van ANDRICHEM (Hrsg.): Biology of stress in FarmAnimals: An integrative Approach.Nijhoff Publishers, S. 3-12
PANKSEPP, (1982):Toward a general psychobiological theory of emotions.The Behavioural and Brain Sciences, 5, S. 407-467
PARROT, R.F., R.H. BRADSHAW, D.M. LLOYD u. J.A. GOODE (1998):Effects of transport and indomethacin on telemetered body temperature and release of cortisoland prolactin in prepuberal pigs.Research in Veterinary Science 64, S. 51-55
PENZLIN, H. (1996):Das periphere vegetative Nervensystem der Wirbeltiere.In: PENZLIN, H. (Hrsg.):Lehrbuch der Tierphysiologie.6.Auflage.Verlag Gustav Fischer, Jena, S. 185-190
PLONAIT, H., u. K. BICKHARDT (1988):Lehrbuch der Schweinekrankheiten.Verlag Paul Parey, Berlin und Hamburg

130
POHORECKY. L.A., u. R.J. WURTMAN (1971):Adrenocortical control of epinephrine synthesis.Pharmacol. Rev. 23, S. 1-35
PORZIG, E., u. K. SCHEIBE (1982):Verhaltensbiologische Grundlagen und ihre Bedeutung für hohe Fortpflanzungsleistungenund Senkung von Verlusten.Tierzucht 37, S. 166-172
PRIBRAM, K.H. (1971):Languages of the Brain.Verlag Prentice-Hall: Englewood Cliffs, N.J., USA
PRINCE, J.H. (1977):The eye and vision.In: SWENSON, M.J. (Hrsg.): Dukes Physiology of Domestic AnimalsVerlag Cornell, New York, USA, S. 696-712
PUTTEN, G. van, u. W.J. ELSHOF (1978):Observations on the effect of transport on the well-being and lean quality of slaughter pigs.Animal Regulation Studies 1, S. 247-271
PUTTEN, G. van (1980):Objective Observations on the Behaviour of fattening pigs.Animal Regulation Studies 3, S. 105-118
PUTTEN, G. van (1981):Using pig behaviour to advantage.Pig Veterinary Society Proceedings 1981, S. 32-37
PUTTEN, G. van (1982a):Zum Messen von Wohlbefinden bei Nutztieren. In: FÖLSCH, D.W. u. A. NABHOLZ (Hrsg.): Ethologische Aussagen zur artgerechtenNutztierhaltung, Tierhaltung 13.Verlag Birkhäuser Basel, S. 78-95
PUTTEN, G. van (1982b):Handling of slaughter pigs prior to loading and during loading on a lorry.In: MOSS, R. (Hrsg.): Transport of animals intended for breeding, production and slaughter.Verlag Martinus Nijhoff, Den Haag, Niederlande, S. 15-25
RAABE, T., K. LINDNER u. H. GÜRTLER (1996):Einfluss von Belastungssituationen auf dem Schlachthof auf die Plasmakonzentrationen anCatecholaminen und Stoffwechselparametern im Blutplasma von Schlachtschweinen.Proc. Wiss. Kolloq. "Schlachttierbelastung und Produktsicherheit", Univ. Leipzig, Vet. Med.

131
RASMUSSEN, O.G., E.M. BANKS, T.H. BERRY u. D.E. BECKER (1962):Social dominance in gilts.J. Anim. Sci. 21, S. 520-522
REMANE, A. (1952):Die Grundlagen des natürlichen Systems der vergleichenden Anatomie und der Phylogenetik.Akademische Verlagsgesellschaft, Leipzig
REUTER, G. (1992):Qualitätssicherung bei der Fleischgewinnung.Technologische Aspekte.Fleischwirtschaft 72, S. 974-980
RIVIER, C., u. W. VALE (1983):Modulation of stress-induced ACTH release by corticotropin-releasing factor, catecholaminesand vasopressin.Nature 305, S. 325-327
ROWAN, A.N. (1988):Animal anxiety and animal suffering.Appl. Anim. Behav. Sci. 20, S. 135-142
RÜGER, U., A.F. BLOMERT, u. W. FÖRSTER (1990):Coping.Verlag Med. Psychol. im Verlag Vandenhoeck & Ruprecht, Göttingen
RUSSEL, A.J.F., C.A. MORGAN, C.J. SAVORY, M.C. APPLEBY u. T.H.J. LAWRENCE(Hrsg.) (1999):Farm animal welfare: who writes the rules? Proceedings of an international symposium.Occ. Publ. Br. Soc. Anim. Sci. No. 23
SACHS, L (1992):Angewandte Statistik.7. Auflage Springer Verlag, Berlin
SACKMANN, G. (1988):Einfluß der Ausruhzeit sowie von Umgebungs- und technologischen Faktoren auf klinischeund fleischhygienische Parameter bei Schlachtschweinen.Diss, Freie Universität Berlin, Fachber. Veterinärmed.
SAMBRAUS, H.H. (1979):Das Sozialverhalten von Sauen in Gruppenhaltung.Züchtungskunde 53, S. 147-150

132
SAMBRAUS, H.H. (1982):Ethologische Grundlagen einer tiergerechten Nutztierhaltung.In: FÖLSCH, D.W. u. A.NABHOLZ (Hrsg.): Ethologische Aussagen zur artgerechtenNutztierhaltung, Tierhaltung 13.Verlag Birkhäuser, Basel, S. 23-41
SAMBRAUS, H.H. (1995):Befindlichkeiten und Analogieschluß.KTBL-Schrift 370, S. 31-39
SAMBRAUS, H.H. (1998):Möglichkeiten und Grenzen des Analogieschlusses.Tagungsbericht Tierschutz u. Nutztierhaltung, 05.-07. März, Nürtingen, S. 67-71
SAMBRAUS, H.H. (1999):Der Tierarzt als berufener Tierschützer: Einfacher gesagt als getan?Vet Impulse 14, S. 1-4
SAPOLSKY, R.M. (1994):Why Zebras don´t get ulcers.Verlag W.H. Freeman, New York, USA, S. 63
SAS/SAST User´s Guide (1988):6.12. Version des Statistical Analysis Systems, Inc., Cary, North Carolina, USA
SCHÄFFER, D., E. von BORELL und R.-B. LAUBE (1997):Handling von Schlachtschweinen in der Vorwartebucht. Eine geeignete Methode zumbelastungsreduzierten Austrieb.Fleischwirtschaft 77, S. 608-610
SCHÄFFER, D., E. von BORELL, R.-B. LAUBE, G. MARX u. A. SCHMIDT (1999):Reduzierung von Lärm beim Austrieb von Schweinen aus der Vorwartebucht.Fleischwirtschaft 1, S. 113-116
SCHEURINK, A.J.W., A.B. STEFFENS, H. BOURITIUS, G. DRETELER, R. BRUNTIK u.J. REMIE ZAAGSMA (1989):Adrenal and sympathetic catecholamines in exercising rats.American Journal of Physiology 256, S. 155-161
SCHMIDT, T. (1999):Persönliche Mitteilung.
SCHOUTEN, W., J. RUSHEN u. A.M. de PASSILE (1991):Stereotypic behaviour and heart rate in pigs.Physiol. Behav. 50, S. 617-624

133
SCHÜTTE, A., M. von WENZLAWOWICZ und G. von MICKWITZ (1994):Tiertransport und Fleischqualität bei Schweinen.Fleischwirtschaft 74, S. 126-132
SCHÜTTE, A., (Hrsg.) (1995): Methods of improving pig wlefare and meat quality byreducing stress and discomfort before slaughter.Proc. EU-Seminar „ New information on welfare and meat quality of pigs as related tohandling, transport and lairage conditions“, Mariensee, 29.-30. Juni 1995
SCHÜTTE, A., D.M. BROOM, E. LAMBOOIJ (1995a):Standard methods of estimating physiological parameters – minimal standards.In: SCHÜTTE, A. (Hrsg.): Proc. EU-Seminar „ New information on welfare and meat qualityof pigs as related to handling, transport and lairage conditions“, Mariensee, 29.-30. Juni 1995,S. 69-80
SCHÜTTE, A., A. MERGENS, U. POTT u. S. VENTHIEN (1995b):Effect of transport conditions (straw; stoppage) and unloading procedures on physiologicaland meat quality parameters.Proc. EU-Seminar „ New information on welfare and meat quality of pigs as related tohandling, transport and lairage conditions“, Mariensee, 29.-30. Juni 1995, S. 117-132
SCHÜTTE, A., A. MERGENS, U. POTT u. S. VENTHIEN (1995c):Effect of different kinds of showering in lairage on physiological and meat qualityparameters, taking climatic circumstances into account.In: SCHÜTTE, A. (Hrsg.): Proc. EU-Seminar „ New information on welfare and meat qualityof pigs as related to handling, transport and lairage conditions“, Mariensee, 29.-30. Juni 1995,S. 163-187
SCHÜTTE, A., M. DRUCKREY u. S. KUR (1996):Laktat im Stichblut von Schlachtschweinen. Ein relevanter Parameter zur Sicherung des Tier-und Verbraucherschutzes?Deutsche Veterinärmed. Gesellsch., Gießen, Tagung der Fachgruppe Epidemiologie undDokumentation vom 4. – 6.9.1996 in Leipzig, S. 248-249
SCHULZE, B. (1990):Ethologische Untersuchungen zur artgerechten Haltung von Schweinen im erweitertenprämortalen Bereich. Diss, Freie Universität Berlin, Fachber. Veterinärmed.
SCHULZE, B., u. R.-B. LAUBE (1991):Neue ethologische Erkenntnisse zum Treiben und zur Vereinzelung von Schlachtschweinen.Leipziger Tierzuchtsymposium 12./13. Dezember 1991
SCHWITZGEBEL, D. (1992):Safety Training: Ein komplexes Verfahren in der Verhaltenstherapie beim Hund.Kleintierpraxis 37, S.241-253

134
SEIDEL, B. (1992): Zum Stand der Entwicklung von Maschinen und Anlagen für dasSchlachten von Rindern und Schweinen.Fleisch 46, S. 211-215
SELIGMAN, M.E.P. (1975):Helplessness: On Depression, Development and Death.Verlag Freeman, San Francisco, USA
SELLER, H. (1996):Neurovegetative Regulation.In: KLINKE, R. u. S. SILBERNAGEL (Hrsg.): Lehrbuch der Physiologie.Verlag Georg Thieme, Stuttgart, New York, S. 675-690
SELYE, H. (1936):A syndrome produced by diverse noxious agents.Nature 136, S. 32
SELYE, H. (1950):The physiology and pathology of exposure to stress.Verlag Acta, Montreal, Canada
SELYE, H. (1981):Geschichte und Grundzüge des Streßkonzeptes. In: NITSCH, J.R. (Hrsg.): Streß. Theorien,Untersuchungen, Maßnahmen.Verlag Hans Huber, Bern, Schweiz, S. 163-187
SHAPIRO, P.A., R.P. SLOAN, E.M. HORN, M.M. MYERS u. J.M. GORMAN (1993):Effect of innervation on heart rate response to mental stress.Arch. Gen. Psych. 50, S. 275-279
SHAW, F.D., u. R.K. TUME (1992):The assessment of pre-slaughter and slaughter treatments of livestock by measurement ofplasma constituents.A review of recent work.Meat Sci. 32, S. 311-329
SHAW, F.D., u. G.R. TROUT (1995):Plasma and muscle cortisol measurements as indicators of meat quality and stress in pigs.Meat Sci. 39, S. 237-246
SINZ, R. (1974)Lernen und Gedächtnis.Verlag Fischer, Stuttgart

135
SLOAN R.P., P.A. SHAPIRO, E. BAGIELLA, J.T. BIGGER, E.S. LO u. J.M. GORMAN(1996): Relationship between circulating catecholamines and low frequency heart periodvariability as indices of cardiac sympathetic activity during mental stress.Psychosom. Med. 58, S. 25-31
SMEDES, F., J.C. KRAAK u. H. POPPE (1982):Simple and fast solvent extraction system for selective and quantitative isolation ofadrenaline, noradrenaline and dopamine from plasma and urine.J, Chromatog. 231, S. 25-39
SMIDT, D., E. KALLWEIT u. J. LADEWIG (1988):Streß, Gesundheit und Leistung beim Schwein.Tierärztl. Prax. Suppl. 3, S. 1-10
SPENCER, G.S.G., L.J. WILKINS u. K.G. HALLET (1984):Hormone and metabolic changes in the blood following loading and during transport and theirpossible relationship with subsequent meat quality.In: Proc. Symp. 30th European Meeting of Meat Res. Workers, Bristol 1984, S. 15-16
SPENCLEY, J., und J. HARTUNG (1994):Einfluß von Geräuschen auf einzeln gehaltene Jungschweine.Proc. Tagung "Hygiene und Tierschutz bei Tiertransporten" in Hannover, 8./9. März 1994Dt. Vet. Med. Gesellsch.,Gießen
STAHLHUT-KLIPP, H. (1975):Zur Methodik der enzymatischen Pyruvat- und Lactatbestimmung in Rohmilch.Milchwissenschaft 30, S. 468-471
STAUFFACHER, M. (1993):Angst bei Tieren – ein zoologisches und forensisches Problem.Dt. tierärztl. Wochenschrift 100, S. 301-340
STEFFENS, C., M. MARAHRENS, J. HARTUNG u. F. FELDHUSEN (1995):Individuelle Reaktionen von Schlachtschweinen beim Transport.Proc. 36. Arbeitstagung des Arbeitsgebietes "Lebensmittelhygiene" der DeutschenVeterinärmedizinischen Gesellschaft, Teil I, Garmisch-Partenkirchen, S. 146-151
STEINHARDT, M. (1966):Beziehung zwischen Temperaturregulation und Kreislauf beim Schwein.Arch. Exp. Vet. med. 20, S. 1091-1100
STEGEMANN, J. (1984):Leistungsphysiologie.Georg Thieme Verlag, Stuttgart, New York

136
STEGEN, D. (1993):Tierschutzgerechte Behandlung von Schlachttieren.Dtsch. tierärztl. Wschr. 100, S. 58-64
STEPHENS, D.B., u. G.C. PERRY (1990):The effect of restraint, handling, simulated and real transport in the pig (with reference to manand other species)Applied Animal Behaviour Science 28, S. 41-55
TARRANT, P.V. (1989):The effects of handling, transport, slaughter and chilling on meat quality and yield in pigs.Ir. J. Food Sci. Technol. 13, 79-93TEMBROCK, G. (1969):Allgemeine Grundlagen des Verhaltens von Haustieren.In: PORZIG, E. (Hrsg.): Das Verhalten landwirtschaftlicher Nutztiere.Deutscher Landwirtschaftsverlag,Berlin
TEMBROCK, G. (1982):Spezielle Verhaltensbiologie der Tiere. Bd. I.Verlag Gustav Fischer, Jena
THOMPSON, G.E. (1977):Physiological effects of cold exposure.International Review of Physiology, Environmental Physiology II 15, S. 30-69
THUN, R., E. EGGENBERGER, K. ZEROBIN, T. LUSCHER u. W. VETTERT (1981):Twenty-four-hour secretory pattern of cortisol in the bull: Evidence of episodic secretion andcircadian rhythm.Endocrinology 109, S. 2208-2212
THUN, R. (1987):Untersuchungen über die Tagesrhythmik von Cortisol beim Rind.Verlag Ferdinand Enke, Stuttgart
THUN, R., und D. SCHWARTZ - PORSCHE (1994):Nebennierenrinde.in: DÖCKE, F (Hrsg.): Veterinärmedizinische EndokrinologieVerlag Gustav Fischer, Jena, 3. Auflage
TOATES, F. (1986):Motivational Systems.Cambridge University Press
TOATES, F. (1995):Stress.Conceptional and biological aspects.Verlag Wiley, Chichester, UK

137
TORTORA, D.F. (1983):Safety Training: The Elimination of Avoidance-Motivated Aggression in Dogs.Journal of Experimental Psychology, 2, S.176-214
TRÖGER, K., und W. WOLTERSDORF (1988):Die Messung der Streßbelastung des Schweines während der Schlachtung. Fleischwirtschaft 68, S. 249-254
TRÖGER, K., u. W. WOLTERSDORF (1989):Measuring stress in pigs during slaughter.Fleischwirtschaft 69, S. 373-376
TROEGER, K. (1990):Schlachten: Tierschutz und Fleischqualität. Gegenwärtige Praxis - Welche Forderungen sindzu stellen?Fleischwirtschaft 70, S. 266-272
TRUNKFIELD, H.R., D.M. BROOM, K. MAATJE, H.K. WIERENGA, E. LAMBOOIJ u. J.KOOIJMAN (1991):Effects of housing on responses of veal calves to handling and transport.In: METZ, J.H.M. u. C.M. GROENENSTEIN (Hrsg.): New Trends in Veal Calf Production.Verl. Pudoc, Wageningen, S. 40-43
TSCHANZ, B. (1982):Verhalten, Bedarfsdeckung und Schadensvermeidung bei Tieren.Nutztierkommission Schweizer Tierschutz/Internationale Gesellschaft für Nutztierhaltung(IGN), S. 9-13
TSCHANZ, B. (1987):Bedarfsdeckung und Schadensvermeidung – ein ethologisches Konzept.In: ZEEB, K. (Hrsg.): Akt. Arb. Zur artgem. Tierhaltung, 1986, KTBL-Schrift 319.Darmstadt, S. 9-17
TSCHANZ, B. (1995):Erfaßbarkeit von Befindlichkeiten bei Tieren.In: Aktuelle Arbeiten zur artgemäßen Tierhaltung, KTBL-Schrift 370, S. 20-30
UNSHELM, I. (1971):Review of papers Section 3: Transport (Physiological and morphological factors inducingstress-susceptibility in pigs).Proc. 2nd int. Symp. Condition Meat Quality Pigs, Zeist Pudoc, Wageningen, 1971, S. 171-179
USDIN, E., R. KVETNANSKY u. I.J. KOPIN (1976):Catecholamines and Stress.Verlag Pergamon Press, Oxford, UK

138
VEITH-FLANIGAN, J., u. C.A. SANDMAN (1985):Neuroendocrine relationships with stress.In: BURCHFIELD, S.R. (Hrsg.): Stress. Psychological and Physiological Interactions.Verlag Hemisphere, Washington, USA, S. 129-161
VERHAGEN, J.M.F., J.J.M. MICHELS und W.G.P. SCHOUTEN (1987):The relation between body temperature, metabolic rate and climatic environment in young-growing pigs.In: VERHAGEN, J.M.F. (Ed.): Acclimation of growing pigs to climatic environment.Diss. Univ. Wageningen, Dept. Anim. Husbandry, Niederlande
VOIGT, K. (1995):Endokrines System.In: KLINKE, R. u. S. SILBERNAGEL (Hrsg.): Lehrbuch der Physiologie.Verlag Georg Thieme, Stuttgart, S. 439-490
WAIDA, S. (1986):Einfluß unterschiedlicher Treibemethoden auf den Schlachtwert von Schweinen.Fleischwirtschaft 66, S. 1271-1272
WAL, P.G. van der, B. ENGEL, G. van ESSEN u. H.G. HULSHOF (1985):Changes in blood acid-base characteristics, haemoglobin and lactate concentrations due toincreasing moderate stress in pigs.I.V.O.-Report B-271, S. 19-22
WARRIS, P.D., u. S.N. BROWN (1985):The physiological responses to fighting in pigs and the consequences for meat quality.Journal of Science of Food in Agriculture 36, S. 87-92
WARRISS, P.D, S.N. BROWN, J.E. EDWARDS, M.H. ANIL u. D.P. FORDHAM (1992):Time in lairage needed by pigs to recover from the stress of transport.Vet. Rec. 131, S. 194-196
WARRISS, P.D., S.N. BROWN, S.J.M. ADAMS u. I.K. CORLETT (1994):Relationships between subjective and objective assessments of stress at slaughter and meatquality in pigs.Meat Sci. 38. S. 329-340
WARRISS, P.D, S.N. BROWN, J.E. EDWARDS, u. TG. KNOWLES (1995a): Effect of lai-rage time on levels of stress and meat quality in pigs. in: A. SCHÜTTE (Ed.): Proc. EU-Seminar „New information on welfare and meat quality ofpigs as related to handling, transport and lairage conditions.“ Mariensee, 29.-30.06.1995, S.145 - 152

139
WARRISS, P.D, S.N. BROWN, G.R. NUTE, TG. KNOWLES, J.E. EDWARDS, A.M.PERRY u. S.P. JOHNSON (1995b):Potential interactions between effects of preslaughter stress and post-mortem electricalstimulation of the carcasses on meat quality in pigs.Meat Science 41, S. 55-68
WECHSLER, B. (1993):Verhaltensstörungen und Wohlbefinden: Ethologische Überlegungen.In: Leiden und Verhaltensstörungen bei Tieren. Tierhaltung 23.Verlag Birkhäuser, Basel, Boston, Berlin, S.50-64
WEEDING, C.M., E.J. HUNTER, H.J. GUISE u. R.H.C. PENNY (1993):Effects of abattoir and slaughter handling systems on stress indicators in pig blood.Vet. Rec. 133, S. 10-13WEINBERG, J., u. S. LEVINE (1980):Psychobiology of coping in animals: the effects of predictability.In: LEVINE, S. u. H. URSIN (Hrsg.): coping and health.Verlag Plenum Press, New York, USA, S. 39-59
WEISS, J.M. (1971):Effects of coping behaviour in different warning signal conditions on stress pathology in rats.J. Comp. Physiol. Psych. 77, S.1-13
WEISS, J.M. (1972):Psychological factors in stress and disease.Scientific American 226, S. 104-113
WENZLAWOWICZ, M. von, K. von HOLLEBEN, A. BRIESE u. L. HEUKING (1994):Tierschutz am Schlachthof.Berl. Münch. Tierärztl. Wschr. 107, S. 237-244
WENZLAWOWICZ, M. von, A. SCHÜTTE, K. von HOLLEBEN, A. von ALTROCK, N.BOSTELMANN u. S. ROEB (1998):Felduntersuchung der MIDAS-Schweinebetäubungsanlage mit Inarco-System hinsichtlichTierschutz und Schlachtkörperqualität. 2. Vorbelastung und Schlachtkörperqualität.Fleischwirtschaft 78, 10, S. 1096-1099
WIEPKEMA, P.R., u. J. M. KOOLHAAS (1992):The emotional brain.Animal welfare 1, S.13-18
WIEPKEMA, P.R., u. J. M. KOOLHAAS (1993):Stress and animal welfare.Animal welfare 2, S. 195-218

140
WOLTERSDORF, B., und L. LYHS (1988):Verhaltenssituationen und Herzfrequenzen beim Schwein.Mh. Vet. Med. 43, S. 880-882
WOLTERSDORF, W., u. K. TROEGER (1987):Schlachttechnik zur Verminderung des PSE – Anteiles beim Schwein. Fleischwirtschaft 67, S. 724-730
YAMAGUCHI, J., u. J.J. KOPIN (1979):Plasma catecholamine and blood pressure responses to sympathetic stimulation in pithed rats.Am. J. Physiol. 237, S. 305-310
ZERBONI, N. von, u. A. GRAUVOGL (1984):Schwein.In: BOGNER, H. u. A. GRAUVOGL (Hrsg.): Verhalten landwirtschaftlicher Nutztiere.Eugen Ulmer Verlag Stuttgart, 1. Auflage, S. 246-282
ZHANG, S.H., D.P. HENNESY, D.E. NOONAN u. H.J. FRANCIS (1992):Physiological responses to exercise in pigs of differing adrenal responsiveness.Comp. Biochem. Physiol. 103, S. 695-703
Gesetze und Verordnungen
1993TierschutzgesetzVom 17. Februar 1993Zuletzt geändert durch Gesetz vom 25. Juni 2001BGBL. I, S. 254-267
1997Tierschutz-Schlachtverordnung (TierSchlV)Vom 3. März 1997Zuletzt geändert durch 1. ÄndVO vom 25. November 1999BGBL. I, S. 405
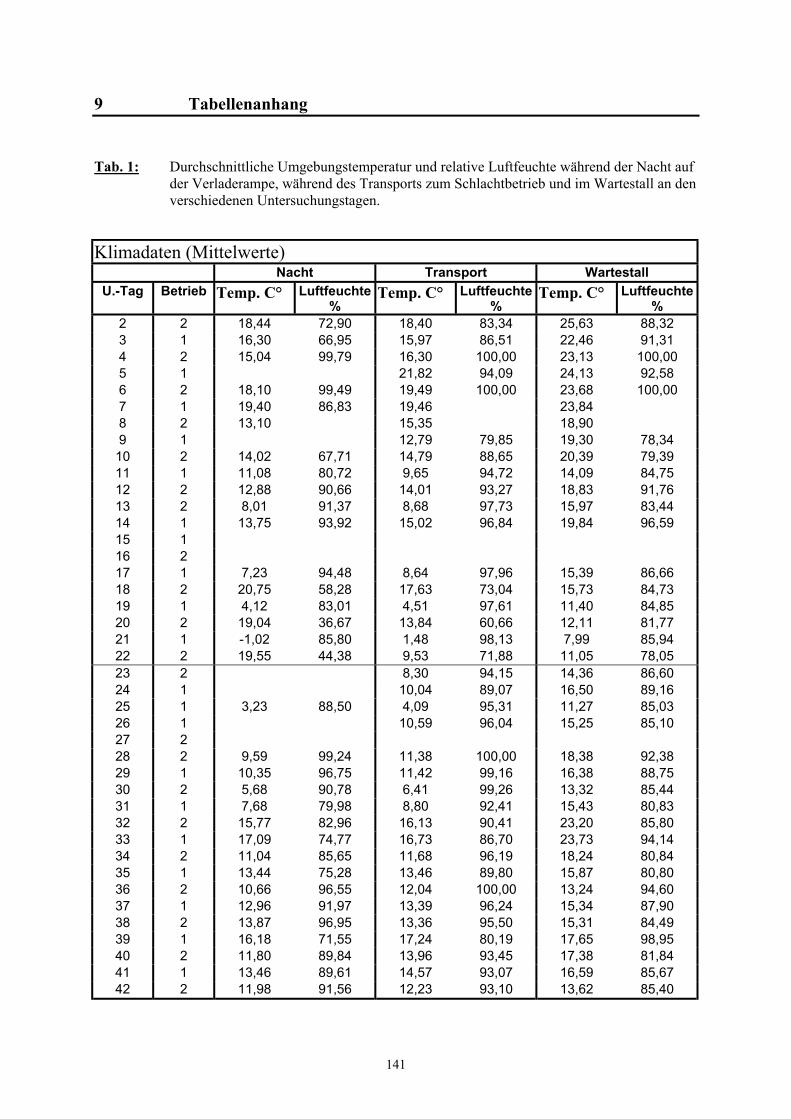
141
9 Tabellenanhang
Tab. 1: Durchschnittliche Umgebungstemperatur und relative Luftfeuchte während der Nacht aufder Verladerampe, während des Transports zum Schlachtbetrieb und im Wartestall an denverschiedenen Untersuchungstagen.
Klimadaten (Mittelwerte)Nacht Transport Wartestall
U.-Tag Betrieb Temp. C° Luftfeuchte%
Temp. C° Luftfeuchte%
Temp. C° Luftfeuchte%
2 2 18,44 72,90 18,40 83,34 25,63 88,323 1 16,30 66,95 15,97 86,51 22,46 91,314 2 15,04 99,79 16,30 100,00 23,13 100,005 1 21,82 94,09 24,13 92,586 2 18,10 99,49 19,49 100,00 23,68 100,007 1 19,40 86,83 19,46 23,848 2 13,10 15,35 18,909 1 12,79 79,85 19,30 78,3410 2 14,02 67,71 14,79 88,65 20,39 79,3911 1 11,08 80,72 9,65 94,72 14,09 84,7512 2 12,88 90,66 14,01 93,27 18,83 91,7613 2 8,01 91,37 8,68 97,73 15,97 83,4414 1 13,75 93,92 15,02 96,84 19,84 96,5915 116 217 1 7,23 94,48 8,64 97,96 15,39 86,6618 2 20,75 58,28 17,63 73,04 15,73 84,7319 1 4,12 83,01 4,51 97,61 11,40 84,8520 2 19,04 36,67 13,84 60,66 12,11 81,7721 1 -1,02 85,80 1,48 98,13 7,99 85,9422 2 19,55 44,38 9,53 71,88 11,05 78,0523 2 8,30 94,15 14,36 86,6024 1 10,04 89,07 16,50 89,1625 1 3,23 88,50 4,09 95,31 11,27 85,0326 1 10,59 96,04 15,25 85,1027 228 2 9,59 99,24 11,38 100,00 18,38 92,3829 1 10,35 96,75 11,42 99,16 16,38 88,7530 2 5,68 90,78 6,41 99,26 13,32 85,4431 1 7,68 79,98 8,80 92,41 15,43 80,8332 2 15,77 82,96 16,13 90,41 23,20 85,8033 1 17,09 74,77 16,73 86,70 23,73 94,1434 2 11,04 85,65 11,68 96,19 18,24 80,8435 1 13,44 75,28 13,46 89,80 15,87 80,8036 2 10,66 96,55 12,04 100,00 13,24 94,6037 1 12,96 91,97 13,39 96,24 15,34 87,9038 2 13,87 96,95 13,36 95,50 15,31 84,4939 1 16,18 71,55 17,24 80,19 17,65 98,9540 2 11,80 89,84 13,96 93,45 17,38 81,8441 1 13,46 89,61 14,57 93,07 16,59 85,6742 2 11,98 91,56 12,23 93,10 13,62 85,40
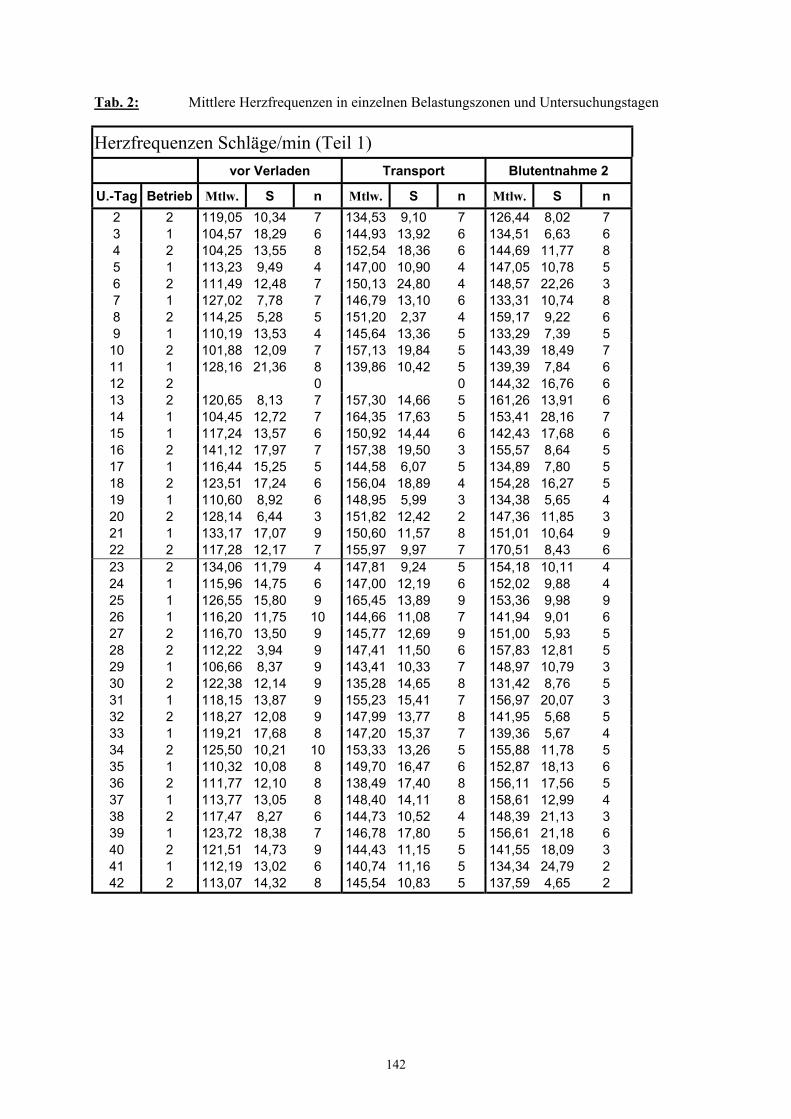
142
Tab. 2: Mittlere Herzfrequenzen in einzelnen Belastungszonen und Untersuchungstagen
Herzfrequenzen Schläge/min (Teil 1)
vor Verladen Transport Blutentnahme 2
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n
2 2 119,05 10,34 7 134,53 9,10 7 126,44 8,02 73 1 104,57 18,29 6 144,93 13,92 6 134,51 6,63 64 2 104,25 13,55 8 152,54 18,36 6 144,69 11,77 85 1 113,23 9,49 4 147,00 10,90 4 147,05 10,78 56 2 111,49 12,48 7 150,13 24,80 4 148,57 22,26 37 1 127,02 7,78 7 146,79 13,10 6 133,31 10,74 88 2 114,25 5,28 5 151,20 2,37 4 159,17 9,22 69 1 110,19 13,53 4 145,64 13,36 5 133,29 7,39 510 2 101,88 12,09 7 157,13 19,84 5 143,39 18,49 711 1 128,16 21,36 8 139,86 10,42 5 139,39 7,84 612 2 0 0 144,32 16,76 613 2 120,65 8,13 7 157,30 14,66 5 161,26 13,91 614 1 104,45 12,72 7 164,35 17,63 5 153,41 28,16 715 1 117,24 13,57 6 150,92 14,44 6 142,43 17,68 616 2 141,12 17,97 7 157,38 19,50 3 155,57 8,64 517 1 116,44 15,25 5 144,58 6,07 5 134,89 7,80 518 2 123,51 17,24 6 156,04 18,89 4 154,28 16,27 519 1 110,60 8,92 6 148,95 5,99 3 134,38 5,65 420 2 128,14 6,44 3 151,82 12,42 2 147,36 11,85 321 1 133,17 17,07 9 150,60 11,57 8 151,01 10,64 922 2 117,28 12,17 7 155,97 9,97 7 170,51 8,43 623 2 134,06 11,79 4 147,81 9,24 5 154,18 10,11 424 1 115,96 14,75 6 147,00 12,19 6 152,02 9,88 425 1 126,55 15,80 9 165,45 13,89 9 153,36 9,98 926 1 116,20 11,75 10 144,66 11,08 7 141,94 9,01 627 2 116,70 13,50 9 145,77 12,69 9 151,00 5,93 528 2 112,22 3,94 9 147,41 11,50 6 157,83 12,81 529 1 106,66 8,37 9 143,41 10,33 7 148,97 10,79 330 2 122,38 12,14 9 135,28 14,65 8 131,42 8,76 531 1 118,15 13,87 9 155,23 15,41 7 156,97 20,07 332 2 118,27 12,08 9 147,99 13,77 8 141,95 5,68 533 1 119,21 17,68 8 147,20 15,37 7 139,36 5,67 434 2 125,50 10,21 10 153,33 13,26 5 155,88 11,78 535 1 110,32 10,08 8 149,70 16,47 6 152,87 18,13 636 2 111,77 12,10 8 138,49 17,40 8 156,11 17,56 537 1 113,77 13,05 8 148,40 14,11 8 158,61 12,99 438 2 117,47 8,27 6 144,73 10,52 4 148,39 21,13 339 1 123,72 18,38 7 146,78 17,80 5 156,61 21,18 640 2 121,51 14,73 9 144,43 11,15 5 141,55 18,09 341 1 112,19 13,02 6 140,74 11,16 5 134,34 24,79 242 2 113,07 14,32 8 145,54 10,83 5 137,59 4,65 2

143
Tab.3: Mittlere Herzfrequenzen in einzelnen Belastungszonen und Untersuchungstagen
Herzfrequenzen Schläge/min (Teil 2)
Wartestall Blutentnahme 3 Umtrieb Zuführung*
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n Mtlw. S n
2 2 105,11 9,04 6 106,48 10,53 6 135,53 15,62 6 165,42 39,61 23 1 99,63 7,65 4 95,87 9,01 3 141,93 7,23 2 204,44 2,55 24 2 108,62 13,40 8 102,03 11,94 7 164,99 24,65 7 191,34 24,18 45 1 112,57 7,34 5 98,60 1,22 3 148,63 17,95 3 186,63 6,61 36 2 107,75 13,56 5 108,45 10,04 6 149,01 9,25 3 192,50 13,90 37 1 104,81 10,26 8 114,03 15,43 6 154,00 26,54 5 196,82 14,39 48 2 109,12 5,83 7 122,42 11,33 5 166,14 17,35 4 192,04 25,43 49 1 106,42 13,49 4 110,60 21,32 3 173,53 7,70 3 184,86 9,55 310 2 100,92 9,97 9 110,47 15,37 8 145,81 11,07 7 193,07 18,41 611 1 108,25 6,16 6 123,36 10,59 5 147,54 18,95 6 184,42 12,06 412 2 113,24 5,98 6 113,45 5,48 5 164,30 12,60 5 176,58 14,54 513 2 113,52 11,04 6 135,46 15,32 5 154,95 19,66 5 187,55 29,32 414 1 113,89 10,99 9 124,40 12,64 5 154,04 13,80 5 205,69 17,68 415 1 113,70 11,50 7 132,49 30,91 5 160,27 34,58 4 182,11 10,22 516 2 113,81 8,18 7 121,80 12,76 4 167,92 14,65 6 194,04 12,78 217 1 108,38 8,82 7 120,68 12,74 7 151,70 9,59 5 195,45 15,49 418 2 117,98 11,30 6 142,65 24,68 5 163,80 28,32 5 172,94 21,76 419 1 102,12 5,24 4 111,62 2,56 4 151,23 20,44 4 181,59 6,37 320 2 115,95 10,50 3 139,57 24,69 3 160,21 9,67 2 195,95 34,01 221 1 116,71 12,32 7 134,45 13,32 6 164,95 14,50 6 204,38 7,63 522 2 116,06 5,45 7 132,48 15,23 5 167,74 11,55 5 191,63 10,68 423 2 114,21 6,89 7 125,26 11,92 4 156,16 13,80 6 187,80 12,87 424 1 108,54 2,32 6 103,11 11,77 4 158,43 8,92 6 165,35 12,81 625 1 111,28 4,49 8 120,75 15,21 7 171,11 14,55 8 197,96 23,89 826 1 111,83 9,60 7 104,51 6,87 7 142,50 8,14 6 163,23 22,23 527 2 110,46 9,80 9 119,99 28,05 5 163,68 14,94 6 172,48 31,42 528 2 117,63 8,73 9 136,06 7,51 4 159,00 15,55 6 166,66 17,41 429 1 107,21 8,06 8 107,46 8,76 4 146,72 11,61 7 177,89 20,92 630 2 103,93 7,65 9 102,44 5,99 4 138,31 8,57 3 157,46 1,77 231 1 113,27 16,15 7 104,41 15,19 5 166,82 31,67 5 180,12 27,32 332 2 112,85 12,09 9 104,68 9,81 4 147,48 14,13 5 162,71 26,64 733 1 113,49 11,71 8 108,52 10,36 5 146,30 9,96 4 162,12 12,63 434 2 117,97 8,26 9 108,84 7,06 5 146,95 10,99 8 154,38 11,37 435 1 105,93 8,04 7 95,20 3,77 4 158,39 17,86 6 174,99 32,70 536 2 110,48 12,61 9 110,47 13,91 4 152,37 15,39 5 163,84 6,93 437 1 110,36 7,90 7 111,81 6,70 4 156,04 16,05 6 173,04 20,39 638 2 109,74 10,10 6 0 143,96 1 155,97 8,56 339 1 112,88 9,31 7 115,27 8,41 6 151,78 27,75 5 179,00 24,65 440 2 109,74 7,77 9 113,90 5,22 4 140,57 9,07 4 169,34 15,18 641 1 102,90 8,29 6 93,28 5,51 2 141,21 22,90 4 168,11 29,24 442 2 104,32 8,87 7 94,93 2,96 2 132,83 11,26 5 152,05 23,62 4
* Zuführung = Vereinzelung und Zutrieb

144
Tab. 4: Mittlere Körpertemperaturen weiblicher Schweine in einzelnen Belastungszonen undUntersuchungstagen
Temperaturen (Teil 1)
vor Verladen Transport Wartestall
U.-Tag Betrieb Mtlw. °C S n Mtlw. °C S n Mtlw. °C S n
2 2 39,40 0,22 2 39,57 0,06 2 39,30 0,25 23 1 39,51 0,10 2 39,50 0,02 2 39,36 0,09 24 2 0 0 05 1 38,48 0,60 2 38,85 0,70 2 38,03 0,21 26 2 0 0 07 1 0 0 08 2 0 0 09 1 0 0 010 2 0 0 011 1 38,90 0,58 4 39,02 0,53 4 38,76 0,46 412 2 0 0 013 2 39,39 0,40 7 39,53 0,57 7 39,17 0,55 714 1 39,28 0,24 5 39,10 0,26 5 38,88 0,41 515 1 38,49 1,28 5 39,36 0,12 5 39,34 0,19 516 2 39,22 0,11 5 39,42 0,20 5 39,07 0,24 517 1 39,29 0,17 5 39,25 0,24 5 39,01 0,25 518 2 39,28 0,23 4 39,43 0,30 4 39,12 0,34 419 1 39,21 0,48 4 39,20 0,36 4 39,02 0,28 420 2 39,69 0,06 4 39,71 0,17 4 39,59 0,38 421 1 39,20 0,20 5 39,29 0,09 5 38,80 0,20 522 2 39,45 0,37 5 39,57 0,32 5 39,30 0,24 523 2 39,07 0,31 3 39,20 0,36 3 38,60 0,35 324 1 39,16 0,16 3 39,36 0,07 3 38,85 0,17 325 1 39,33 0,08 2 39,32 0,08 2 38,83 0,21 226 1 39,04 0,16 2 38,72 0,37 3 38,79 0,26 327 2 39,08 0,38 4 39,26 0,29 4 39,03 0,39 428 2 39,64 0,26 3 39,63 0,36 3 39,35 0,25 329 1 39,03 0,33 2 39,13 0,08 2 38,51 0,06 230 2 0 39,28 0,33 5 39,01 0,36 531 1 39,25 0,13 4 39,24 0,28 4 38,81 0,25 432 2 0 39,49 0,35 5 39,30 0,51 533 1 39,21 0,14 5 39,51 0,27 5 39,10 0,17 534 2 0 39,51 0,23 6 39,43 0,55 635 1 39,03 0,21 6 39,33 0,20 6 38,86 0,29 636 2 39,56 0,23 6 39,41 0,41 6 38,97 0,51 637 1 39,26 0,20 5 39,30 0,21 5 38,85 0,21 538 2 38,97 0,27 4 39,11 0,20 4 38,72 0,37 439 1 39,34 0,23 2 39,45 0,26 2 39,02 0,25 240 2 39,23 0,57 2 39,80 0,43 3 39,30 0,19 341 1 39,29 0,20 5 39,24 0,23 6 38,64 0,23 642 2 39,25 0,21 5 39,46 0,27 5 39,11 0,22 5
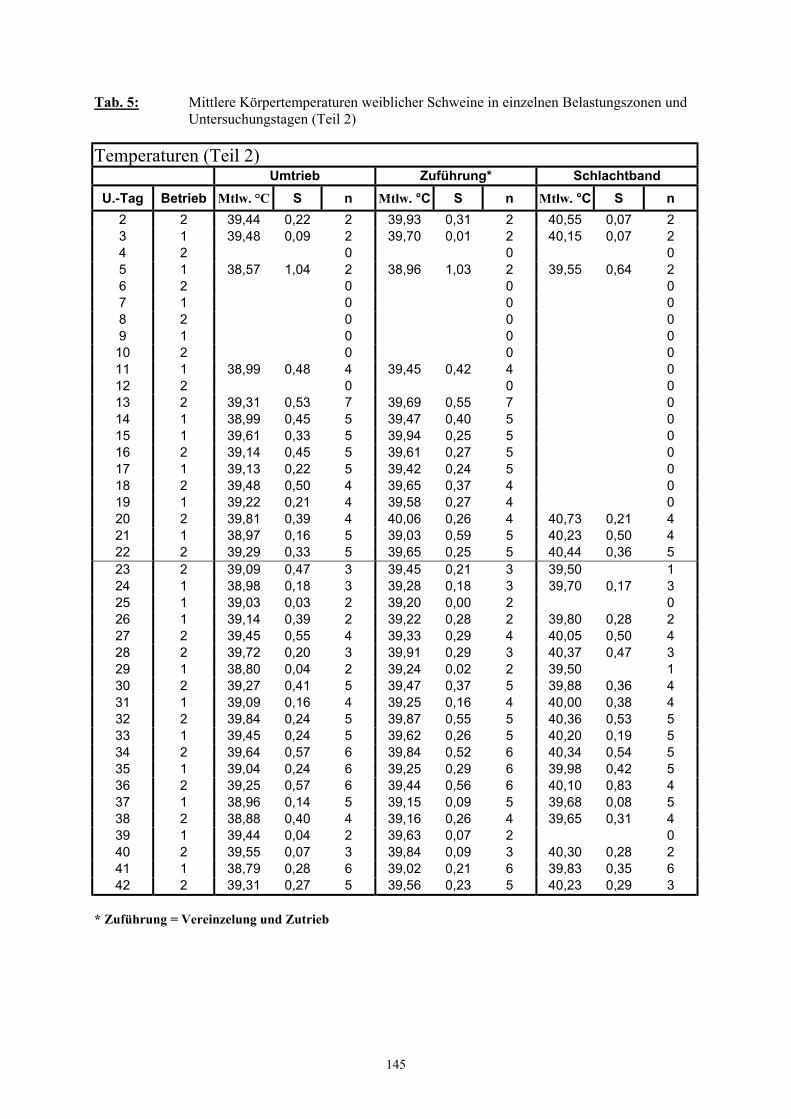
145
Tab. 5: Mittlere Körpertemperaturen weiblicher Schweine in einzelnen Belastungszonen undUntersuchungstagen (Teil 2)
Temperaturen (Teil 2)Umtrieb Zuführung* Schlachtband
U.-Tag Betrieb Mtlw. °C S n Mtlw. °C S n Mtlw. °C S n
2 2 39,44 0,22 2 39,93 0,31 2 40,55 0,07 23 1 39,48 0,09 2 39,70 0,01 2 40,15 0,07 24 2 0 0 05 1 38,57 1,04 2 38,96 1,03 2 39,55 0,64 26 2 0 0 07 1 0 0 08 2 0 0 09 1 0 0 010 2 0 0 011 1 38,99 0,48 4 39,45 0,42 4 012 2 0 0 013 2 39,31 0,53 7 39,69 0,55 7 014 1 38,99 0,45 5 39,47 0,40 5 015 1 39,61 0,33 5 39,94 0,25 5 016 2 39,14 0,45 5 39,61 0,27 5 017 1 39,13 0,22 5 39,42 0,24 5 018 2 39,48 0,50 4 39,65 0,37 4 019 1 39,22 0,21 4 39,58 0,27 4 020 2 39,81 0,39 4 40,06 0,26 4 40,73 0,21 421 1 38,97 0,16 5 39,03 0,59 5 40,23 0,50 422 2 39,29 0,33 5 39,65 0,25 5 40,44 0,36 523 2 39,09 0,47 3 39,45 0,21 3 39,50 124 1 38,98 0,18 3 39,28 0,18 3 39,70 0,17 325 1 39,03 0,03 2 39,20 0,00 2 026 1 39,14 0,39 2 39,22 0,28 2 39,80 0,28 227 2 39,45 0,55 4 39,33 0,29 4 40,05 0,50 428 2 39,72 0,20 3 39,91 0,29 3 40,37 0,47 329 1 38,80 0,04 2 39,24 0,02 2 39,50 130 2 39,27 0,41 5 39,47 0,37 5 39,88 0,36 431 1 39,09 0,16 4 39,25 0,16 4 40,00 0,38 432 2 39,84 0,24 5 39,87 0,55 5 40,36 0,53 533 1 39,45 0,24 5 39,62 0,26 5 40,20 0,19 534 2 39,64 0,57 6 39,84 0,52 6 40,34 0,54 535 1 39,04 0,24 6 39,25 0,29 6 39,98 0,42 536 2 39,25 0,57 6 39,44 0,56 6 40,10 0,83 437 1 38,96 0,14 5 39,15 0,09 5 39,68 0,08 538 2 38,88 0,40 4 39,16 0,26 4 39,65 0,31 439 1 39,44 0,04 2 39,63 0,07 2 040 2 39,55 0,07 3 39,84 0,09 3 40,30 0,28 241 1 38,79 0,28 6 39,02 0,21 6 39,83 0,35 642 2 39,31 0,27 5 39,56 0,23 5 40,23 0,29 3
* Zuführung = Vereinzelung und Zutrieb

146
Tab. 6: Mittlere Plasmakonzentrationen an Noradrenalin von Schlachtschweinen in einzelnenBelastungszonen und Untersuchungstagen
Noradrenalin (nmol/l)
vor Verladen nach Umtrieb nach Zuführung* Stichblut
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n Mtlw. S n
2 2 2,19 1,11 4 2,87 1,00 4 8,29 7,94 4 253,08 129,88 53 1 3,57 1,73 5 4,37 1,76 5 6,76 2,50 5 296,40 111,91 54 2 2,09 0,74 5 2,48 0,92 4 4,41 2,93 3 285,72 97,38 55 1 2,26 0,76 4 4,22 0,69 4 4,93 0,50 2 121,00 27,79 36 2 3,77 1,24 3 6,25 3,00 3 10,65 6,54 3 181,44 108,29 57 1 3,58 1,87 5 4,26 1,68 3 7,44 1,09 3 257,20 59,40 38 2 4,24 2,11 3 4,25 0,51 3 5,27 1,79 2 346,92 129,56 59 1 3,73 1,46 5 5,50 1,35 5 10,34 3,19 5 481,80 218,47 510 2 3,59 0,87 5 7,72 5,32 5 10,57 5,29 5 445,80 83,35 511 1 2,51 0,40 5 5,71 1,70 3 11,38 4,91 3 461,88 198,66 512 2 6,19 2,66 5 6,54 1,42 5 15,76 11,94 5 386,40 187,65 513 2 4,82 1,41 5 11,04 5,62 4 12,97 6,15 5 598,08 232,77 514 1 2,85 1,06 4 6,31 2,27 5 8,44 4,35 4 317,52 195,17 515 1 0,74 0,27 5 2,10 0,99 5 5,03 3,50 5 175,56 129,45 516 2 3,11 4,05 4 0 0 231,60 78,08 517 1 0,95 0,22 5 0,91 0,44 5 1,31 0,76 5 170,55 56,13 418 2 1,57 0,69 3 2,47 0,89 3 10,06 3,84 3 294,72 116,14 519 1 3,13 1,07 4 3,81 1,89 4 5,79 2,94 4 422,16 174,41 520 2 3,35 1,19 5 9,13 4,45 4 13,40 8,03 3 370,08 205,70 521 1 2,58 0,56 2 5,99 4,04 2 9,55 5,64 2 355,92 223,45 522 2 4,01 2,28 5 4,77 1,40 5 7,49 2,48 5 167,64 74,48 523 2 4,70 2,27 3 4,21 1,11 3 8,23 4,31 2 394,56 209,94 524 1 2,57 0,49 4 3,55 0,33 4 3,57 0,40 4 229,32 184,21 525 1 2,69 1,45 5 8,04 5,09 5 9,00 4,26 5 123,60 98,84 526 1 1,92 0,27 4 3,47 2,03 4 3,98 1,38 4 116,52 65,89 527 2 3,11 1,17 4 4,20 0,71 4 4,52 1,29 4 131,64 91,17 528 2 4,73 2,02 5 5,15 1,94 5 6,46 4,92 5 117,90 82,52 429 1 2,38 1,15 5 4,59 2,00 5 6,11 2,09 5 133,52 132,45 530 2 4,16 1,56 5 4,13 1,41 5 4,36 0,97 5 115,80 56,28 531 1 2,85 0,69 5 4,97 2,19 4 5,23 1,20 5 215,28 281,15 532 2 2,77 1,00 5 2,62 1,03 4 3,57 1,52 4 239,76 253,44 533 1 2,32 0,72 5 2,84 0,54 5 2,71 0,28 5 175,20 154,82 534 2 2,45 0,68 5 4,94 3,53 5 3,70 0,79 5 209,52 108,34 535 1 2,12 0,66 3 2,44 0,32 2 2,23 0,20 2 350,16 181,15 536 2 2,38 0,60 5 3,93 1,06 5 3,42 0,66 5 189,00 47,21 537 1 4,67 1,43 4 4,83 0,35 2 6,14 0,00 2 484,44 230,46 538 2 3,05 0,81 5 4,23 2,28 2 4,60 2,83 2 370,20 152,48 539 1 3,48 1,35 5 4,95 4,20 4 4,90 3,03 3 338,85 217,89 440 2 2,30 0,65 5 3,18 1,80 4 4,40 1,74 4 290,76 126,47 541 1 3,20 0,73 5 4,47 2,09 5 5,43 2,21 5 296,40 164,88 542 2 3,58 1,09 4 4,28 1,71 3 4,82 1,33 3 230,28 146,07 5
* Zuführung = Vereinzelung und Zutrieb
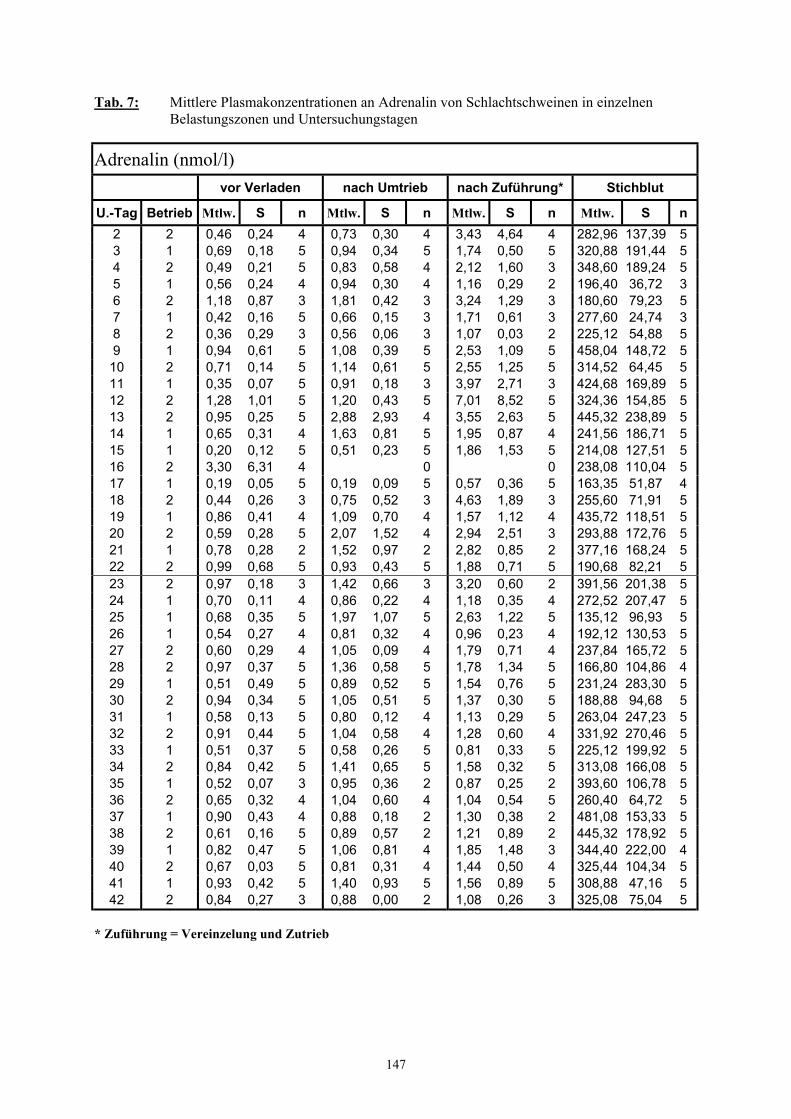
147
Tab. 7: Mittlere Plasmakonzentrationen an Adrenalin von Schlachtschweinen in einzelnenBelastungszonen und Untersuchungstagen
Adrenalin (nmol/l)
vor Verladen nach Umtrieb nach Zuführung* Stichblut
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n Mtlw. S n
2 2 0,46 0,24 4 0,73 0,30 4 3,43 4,64 4 282,96 137,39 53 1 0,69 0,18 5 0,94 0,34 5 1,74 0,50 5 320,88 191,44 54 2 0,49 0,21 5 0,83 0,58 4 2,12 1,60 3 348,60 189,24 55 1 0,56 0,24 4 0,94 0,30 4 1,16 0,29 2 196,40 36,72 36 2 1,18 0,87 3 1,81 0,42 3 3,24 1,29 3 180,60 79,23 57 1 0,42 0,16 5 0,66 0,15 3 1,71 0,61 3 277,60 24,74 38 2 0,36 0,29 3 0,56 0,06 3 1,07 0,03 2 225,12 54,88 59 1 0,94 0,61 5 1,08 0,39 5 2,53 1,09 5 458,04 148,72 510 2 0,71 0,14 5 1,14 0,61 5 2,55 1,25 5 314,52 64,45 511 1 0,35 0,07 5 0,91 0,18 3 3,97 2,71 3 424,68 169,89 512 2 1,28 1,01 5 1,20 0,43 5 7,01 8,52 5 324,36 154,85 513 2 0,95 0,25 5 2,88 2,93 4 3,55 2,63 5 445,32 238,89 514 1 0,65 0,31 4 1,63 0,81 5 1,95 0,87 4 241,56 186,71 515 1 0,20 0,12 5 0,51 0,23 5 1,86 1,53 5 214,08 127,51 516 2 3,30 6,31 4 0 0 238,08 110,04 517 1 0,19 0,05 5 0,19 0,09 5 0,57 0,36 5 163,35 51,87 418 2 0,44 0,26 3 0,75 0,52 3 4,63 1,89 3 255,60 71,91 519 1 0,86 0,41 4 1,09 0,70 4 1,57 1,12 4 435,72 118,51 520 2 0,59 0,28 5 2,07 1,52 4 2,94 2,51 3 293,88 172,76 521 1 0,78 0,28 2 1,52 0,97 2 2,82 0,85 2 377,16 168,24 522 2 0,99 0,68 5 0,93 0,43 5 1,88 0,71 5 190,68 82,21 523 2 0,97 0,18 3 1,42 0,66 3 3,20 0,60 2 391,56 201,38 524 1 0,70 0,11 4 0,86 0,22 4 1,18 0,35 4 272,52 207,47 525 1 0,68 0,35 5 1,97 1,07 5 2,63 1,22 5 135,12 96,93 526 1 0,54 0,27 4 0,81 0,32 4 0,96 0,23 4 192,12 130,53 527 2 0,60 0,29 4 1,05 0,09 4 1,79 0,71 4 237,84 165,72 528 2 0,97 0,37 5 1,36 0,58 5 1,78 1,34 5 166,80 104,86 429 1 0,51 0,49 5 0,89 0,52 5 1,54 0,76 5 231,24 283,30 530 2 0,94 0,34 5 1,05 0,51 5 1,37 0,30 5 188,88 94,68 531 1 0,58 0,13 5 0,80 0,12 4 1,13 0,29 5 263,04 247,23 532 2 0,91 0,44 5 1,04 0,58 4 1,28 0,60 4 331,92 270,46 533 1 0,51 0,37 5 0,58 0,26 5 0,81 0,33 5 225,12 199,92 534 2 0,84 0,42 5 1,41 0,65 5 1,58 0,32 5 313,08 166,08 535 1 0,52 0,07 3 0,95 0,36 2 0,87 0,25 2 393,60 106,78 536 2 0,65 0,32 4 1,04 0,60 4 1,04 0,54 5 260,40 64,72 537 1 0,90 0,43 4 0,88 0,18 2 1,30 0,38 2 481,08 153,33 538 2 0,61 0,16 5 0,89 0,57 2 1,21 0,89 2 445,32 178,92 539 1 0,82 0,47 5 1,06 0,81 4 1,85 1,48 3 344,40 222,00 440 2 0,67 0,03 5 0,81 0,31 4 1,44 0,50 4 325,44 104,34 541 1 0,93 0,42 5 1,40 0,93 5 1,56 0,89 5 308,88 47,16 542 2 0,84 0,27 3 0,88 0,00 2 1,08 0,26 3 325,08 75,04 5
* Zuführung = Vereinzelung und Zutrieb
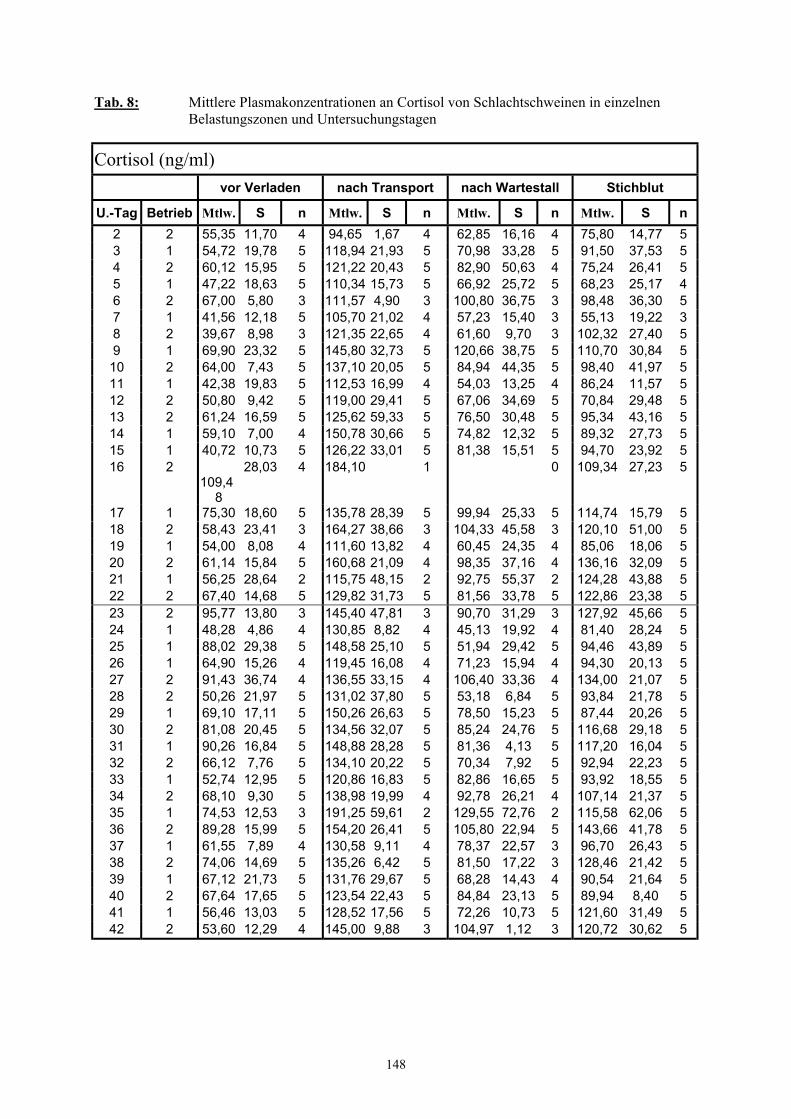
148
Tab. 8: Mittlere Plasmakonzentrationen an Cortisol von Schlachtschweinen in einzelnenBelastungszonen und Untersuchungstagen
Cortisol (ng/ml)
vor Verladen nach Transport nach Wartestall Stichblut
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n Mtlw. S n
2 2 55,35 11,70 4 94,65 1,67 4 62,85 16,16 4 75,80 14,77 53 1 54,72 19,78 5 118,94 21,93 5 70,98 33,28 5 91,50 37,53 54 2 60,12 15,95 5 121,22 20,43 5 82,90 50,63 4 75,24 26,41 55 1 47,22 18,63 5 110,34 15,73 5 66,92 25,72 5 68,23 25,17 46 2 67,00 5,80 3 111,57 4,90 3 100,80 36,75 3 98,48 36,30 57 1 41,56 12,18 5 105,70 21,02 4 57,23 15,40 3 55,13 19,22 38 2 39,67 8,98 3 121,35 22,65 4 61,60 9,70 3 102,32 27,40 59 1 69,90 23,32 5 145,80 32,73 5 120,66 38,75 5 110,70 30,84 510 2 64,00 7,43 5 137,10 20,05 5 84,94 44,35 5 98,40 41,97 511 1 42,38 19,83 5 112,53 16,99 4 54,03 13,25 4 86,24 11,57 512 2 50,80 9,42 5 119,00 29,41 5 67,06 34,69 5 70,84 29,48 513 2 61,24 16,59 5 125,62 59,33 5 76,50 30,48 5 95,34 43,16 514 1 59,10 7,00 4 150,78 30,66 5 74,82 12,32 5 89,32 27,73 515 1 40,72 10,73 5 126,22 33,01 5 81,38 15,51 5 94,70 23,92 516 2
109,48
28,03 4 184,10 1 0 109,34 27,23 5
17 1 75,30 18,60 5 135,78 28,39 5 99,94 25,33 5 114,74 15,79 518 2 58,43 23,41 3 164,27 38,66 3 104,33 45,58 3 120,10 51,00 519 1 54,00 8,08 4 111,60 13,82 4 60,45 24,35 4 85,06 18,06 520 2 61,14 15,84 5 160,68 21,09 4 98,35 37,16 4 136,16 32,09 521 1 56,25 28,64 2 115,75 48,15 2 92,75 55,37 2 124,28 43,88 522 2 67,40 14,68 5 129,82 31,73 5 81,56 33,78 5 122,86 23,38 523 2 95,77 13,80 3 145,40 47,81 3 90,70 31,29 3 127,92 45,66 524 1 48,28 4,86 4 130,85 8,82 4 45,13 19,92 4 81,40 28,24 525 1 88,02 29,38 5 148,58 25,10 5 51,94 29,42 5 94,46 43,89 526 1 64,90 15,26 4 119,45 16,08 4 71,23 15,94 4 94,30 20,13 527 2 91,43 36,74 4 136,55 33,15 4 106,40 33,36 4 134,00 21,07 528 2 50,26 21,97 5 131,02 37,80 5 53,18 6,84 5 93,84 21,78 529 1 69,10 17,11 5 150,26 26,63 5 78,50 15,23 5 87,44 20,26 530 2 81,08 20,45 5 134,56 32,07 5 85,24 24,76 5 116,68 29,18 531 1 90,26 16,84 5 148,88 28,28 5 81,36 4,13 5 117,20 16,04 532 2 66,12 7,76 5 134,10 20,22 5 70,34 7,92 5 92,94 22,23 533 1 52,74 12,95 5 120,86 16,83 5 82,86 16,65 5 93,92 18,55 534 2 68,10 9,30 5 138,98 19,99 4 92,78 26,21 4 107,14 21,37 535 1 74,53 12,53 3 191,25 59,61 2 129,55 72,76 2 115,58 62,06 536 2 89,28 15,99 5 154,20 26,41 5 105,80 22,94 5 143,66 41,78 537 1 61,55 7,89 4 130,58 9,11 4 78,37 22,57 3 96,70 26,43 538 2 74,06 14,69 5 135,26 6,42 5 81,50 17,22 3 128,46 21,42 539 1 67,12 21,73 5 131,76 29,67 5 68,28 14,43 4 90,54 21,64 540 2 67,64 17,65 5 123,54 22,43 5 84,84 23,13 5 89,94 8,40 541 1 56,46 13,03 5 128,52 17,56 5 72,26 10,73 5 121,60 31,49 542 2 53,60 12,29 4 145,00 9,88 3 104,97 1,12 3 120,72 30,62 5

149
Tab.9: Mittlere Plasmakonzentration an Lactat bei Schlachtschweinen in einzelnenBelastungszonen und Untersuchungstagen
Lactat mmol/l (Teil 1)
vor Verladen nach Transport nach Wartestall
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n
2 2 1,45 0,73 4 1,16 0,48 4 0,77 0,14 43 1 1,05 0,48 5 1,57 0,72 5 0,86 0,15 54 2 0,82 0,37 5 1,03 0,32 5 0,89 0,51 45 1 1,85 1,37 5 1,73 0,72 5 1,13 0,07 56 2 1,87 1,29 3 2,01 1,22 3 1,43 1,28 37 1 0,93 0,21 5 0,98 0,20 4 1,00 0,19 38 2 0,88 0,41 3 1,27 0,54 4 0,95 0,17 39 1 0,85 0,19 5 1,93 1,77 5 2,45 3,13 510 2 0,95 0,21 5 2,26 2,47 5 0,96 0,48 511 1 0,79 0,13 5 1,67 0,87 4 1,10 0,23 412 2 1,64 0,88 5 1,50 1,52 5 1,13 0,64 513 2 1,64 0,88 5 1,50 1,52 5 1,13 0,64 514 1 1,16 0,22 4 3,45 2,28 5 1,07 0,25 515 1 1,12 0,37 5 1,56 0,65 5 1,04 0,27 516 2 3,33 4,19 4 3,23 1 017 1 1,05 0,26 5 1,22 0,26 5 1,47 0,84 518 2 2,95 0,70 3 2,97 1,17 3 1,79 1,36 319 1 0,97 0,25 4 0,81 0,26 4 0,71 0,11 420 2 1,06 0,47 5 2,68 1,51 4 0,75 0,30 421 1 0,79 0,31 2 2,21 1,26 2 0,92 0,22 222 2 1,89 1,53 5 1,48 0,66 5 0,85 0,19 523 2 1,38 0,66 2 3,05 1,23 3 1,66 0,67 324 1 1,08 0,35 4 1,15 0,20 4 1,01 0,25 425 1 3,55 4,59 5 3,62 3,67 5 1,08 0,46 526 1 1,10 0,24 4 1,25 0,52 4 1,13 0,33 427 2 0,93 0,61 4 1,50 0,73 4 0,98 0,38 428 2 2,27 0,86 5 2,15 1,41 5 1,20 0,35 529 1 1,22 0,37 5 2,87 1,38 5 1,16 0,41 530 2 1,05 0,17 5 2,49 1,24 5 1,10 0,49 531 1 1,24 0,34 5 1,65 0,55 5 1,36 1,20 532 2 1,80 1,28 5 2,35 0,58 5 1,60 0,44 533 1 1,54 0,59 5 1,97 0,63 5 1,60 0,33 534 2 1,35 0,25 5 2,92 1,68 4 0,98 0,17 435 1 1,10 0,22 3 1,83 0,29 2 1,08 0,01 236 2 0,94 0,23 5 3,34 1,50 5 1,25 0,42 537 1 1,45 0,89 4 1,42 0,72 4 0,92 0,19 338 2 2,16 1,68 5 3,10 1,51 5 0,85 0,12 239 1 2,63 1,84 5 1,29 0,68 5 1,10 0,35 440 2 2,11 0,35 5 1,55 0,34 5 1,32 0,39 541 1 1,67 0,45 5 1,81 0,90 5 1,25 0,16 542 2 1,26 0,83 4 1,73 0,97 3 0,97 0,21 3
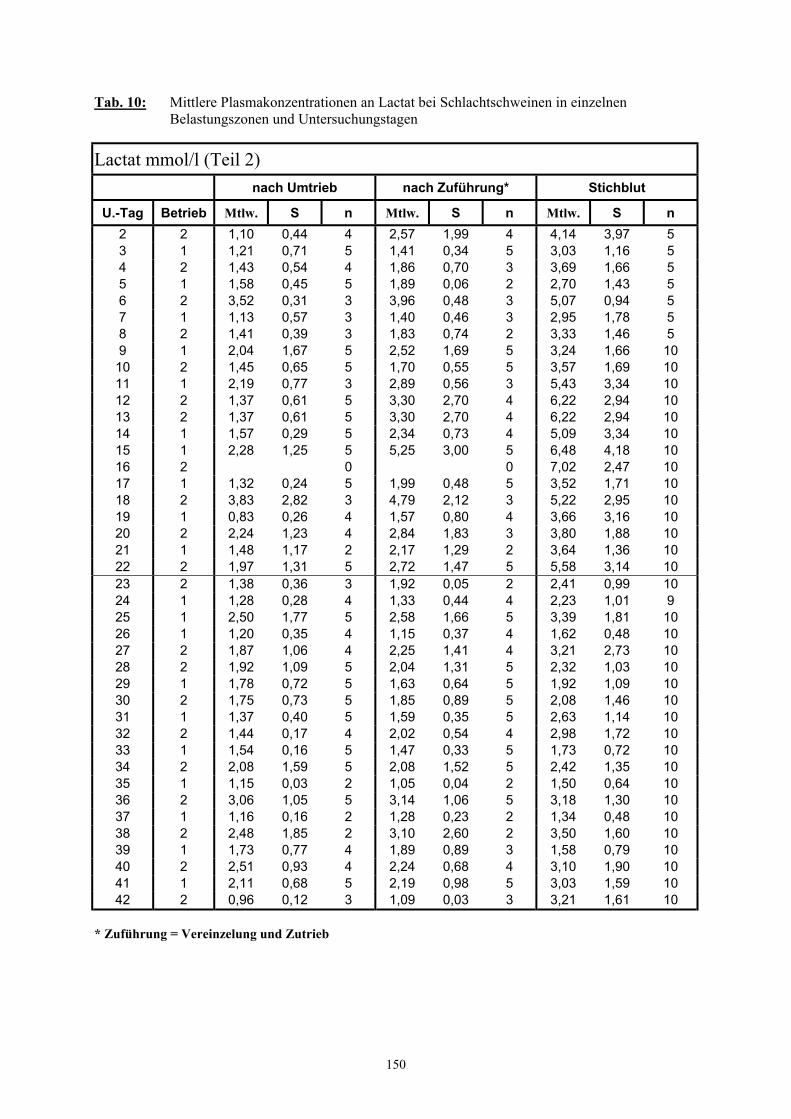
150
Tab. 10: Mittlere Plasmakonzentrationen an Lactat bei Schlachtschweinen in einzelnenBelastungszonen und Untersuchungstagen
Lactat mmol/l (Teil 2)
nach Umtrieb nach Zuführung* Stichblut
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n
2 2 1,10 0,44 4 2,57 1,99 4 4,14 3,97 53 1 1,21 0,71 5 1,41 0,34 5 3,03 1,16 54 2 1,43 0,54 4 1,86 0,70 3 3,69 1,66 55 1 1,58 0,45 5 1,89 0,06 2 2,70 1,43 56 2 3,52 0,31 3 3,96 0,48 3 5,07 0,94 57 1 1,13 0,57 3 1,40 0,46 3 2,95 1,78 58 2 1,41 0,39 3 1,83 0,74 2 3,33 1,46 59 1 2,04 1,67 5 2,52 1,69 5 3,24 1,66 1010 2 1,45 0,65 5 1,70 0,55 5 3,57 1,69 1011 1 2,19 0,77 3 2,89 0,56 3 5,43 3,34 1012 2 1,37 0,61 5 3,30 2,70 4 6,22 2,94 1013 2 1,37 0,61 5 3,30 2,70 4 6,22 2,94 1014 1 1,57 0,29 5 2,34 0,73 4 5,09 3,34 1015 1 2,28 1,25 5 5,25 3,00 5 6,48 4,18 1016 2 0 0 7,02 2,47 1017 1 1,32 0,24 5 1,99 0,48 5 3,52 1,71 1018 2 3,83 2,82 3 4,79 2,12 3 5,22 2,95 1019 1 0,83 0,26 4 1,57 0,80 4 3,66 3,16 1020 2 2,24 1,23 4 2,84 1,83 3 3,80 1,88 1021 1 1,48 1,17 2 2,17 1,29 2 3,64 1,36 1022 2 1,97 1,31 5 2,72 1,47 5 5,58 3,14 1023 2 1,38 0,36 3 1,92 0,05 2 2,41 0,99 1024 1 1,28 0,28 4 1,33 0,44 4 2,23 1,01 925 1 2,50 1,77 5 2,58 1,66 5 3,39 1,81 1026 1 1,20 0,35 4 1,15 0,37 4 1,62 0,48 1027 2 1,87 1,06 4 2,25 1,41 4 3,21 2,73 1028 2 1,92 1,09 5 2,04 1,31 5 2,32 1,03 1029 1 1,78 0,72 5 1,63 0,64 5 1,92 1,09 1030 2 1,75 0,73 5 1,85 0,89 5 2,08 1,46 1031 1 1,37 0,40 5 1,59 0,35 5 2,63 1,14 1032 2 1,44 0,17 4 2,02 0,54 4 2,98 1,72 1033 1 1,54 0,16 5 1,47 0,33 5 1,73 0,72 1034 2 2,08 1,59 5 2,08 1,52 5 2,42 1,35 1035 1 1,15 0,03 2 1,05 0,04 2 1,50 0,64 1036 2 3,06 1,05 5 3,14 1,06 5 3,18 1,30 1037 1 1,16 0,16 2 1,28 0,23 2 1,34 0,48 1038 2 2,48 1,85 2 3,10 2,60 2 3,50 1,60 1039 1 1,73 0,77 4 1,89 0,89 3 1,58 0,79 1040 2 2,51 0,93 4 2,24 0,68 4 3,10 1,90 1041 1 2,11 0,68 5 2,19 0,98 5 3,03 1,59 1042 2 0,96 0,12 3 1,09 0,03 3 3,21 1,61 10
* Zuführung = Vereinzelung und Zutrieb
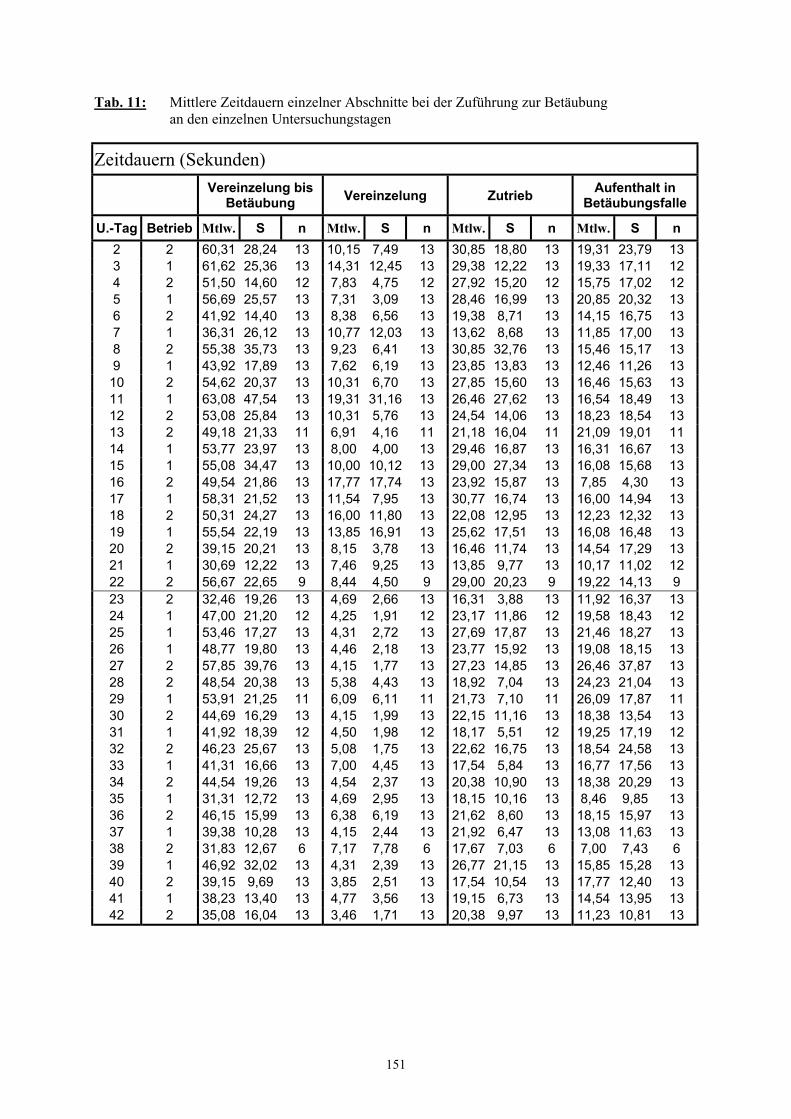
151
Tab. 11: Mittlere Zeitdauern einzelner Abschnitte bei der Zuführung zur Betäubungan den einzelnen Untersuchungstagen
Zeitdauern (Sekunden)
Vereinzelung bisBetäubung Vereinzelung Zutrieb
Aufenthalt inBetäubungsfalle
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n Mtlw. S n
2 2 60,31 28,24 13 10,15 7,49 13 30,85 18,80 13 19,31 23,79 133 1 61,62 25,36 13 14,31 12,45 13 29,38 12,22 13 19,33 17,11 124 2 51,50 14,60 12 7,83 4,75 12 27,92 15,20 12 15,75 17,02 125 1 56,69 25,57 13 7,31 3,09 13 28,46 16,99 13 20,85 20,32 136 2 41,92 14,40 13 8,38 6,56 13 19,38 8,71 13 14,15 16,75 137 1 36,31 26,12 13 10,77 12,03 13 13,62 8,68 13 11,85 17,00 138 2 55,38 35,73 13 9,23 6,41 13 30,85 32,76 13 15,46 15,17 139 1 43,92 17,89 13 7,62 6,19 13 23,85 13,83 13 12,46 11,26 1310 2 54,62 20,37 13 10,31 6,70 13 27,85 15,60 13 16,46 15,63 1311 1 63,08 47,54 13 19,31 31,16 13 26,46 27,62 13 16,54 18,49 1312 2 53,08 25,84 13 10,31 5,76 13 24,54 14,06 13 18,23 18,54 1313 2 49,18 21,33 11 6,91 4,16 11 21,18 16,04 11 21,09 19,01 1114 1 53,77 23,97 13 8,00 4,00 13 29,46 16,87 13 16,31 16,67 1315 1 55,08 34,47 13 10,00 10,12 13 29,00 27,34 13 16,08 15,68 1316 2 49,54 21,86 13 17,77 17,74 13 23,92 15,87 13 7,85 4,30 1317 1 58,31 21,52 13 11,54 7,95 13 30,77 16,74 13 16,00 14,94 1318 2 50,31 24,27 13 16,00 11,80 13 22,08 12,95 13 12,23 12,32 1319 1 55,54 22,19 13 13,85 16,91 13 25,62 17,51 13 16,08 16,48 1320 2 39,15 20,21 13 8,15 3,78 13 16,46 11,74 13 14,54 17,29 1321 1 30,69 12,22 13 7,46 9,25 13 13,85 9,77 13 10,17 11,02 1222 2 56,67 22,65 9 8,44 4,50 9 29,00 20,23 9 19,22 14,13 923 2 32,46 19,26 13 4,69 2,66 13 16,31 3,88 13 11,92 16,37 1324 1 47,00 21,20 12 4,25 1,91 12 23,17 11,86 12 19,58 18,43 1225 1 53,46 17,27 13 4,31 2,72 13 27,69 17,87 13 21,46 18,27 1326 1 48,77 19,80 13 4,46 2,18 13 23,77 15,92 13 19,08 18,15 1327 2 57,85 39,76 13 4,15 1,77 13 27,23 14,85 13 26,46 37,87 1328 2 48,54 20,38 13 5,38 4,43 13 18,92 7,04 13 24,23 21,04 1329 1 53,91 21,25 11 6,09 6,11 11 21,73 7,10 11 26,09 17,87 1130 2 44,69 16,29 13 4,15 1,99 13 22,15 11,16 13 18,38 13,54 1331 1 41,92 18,39 12 4,50 1,98 12 18,17 5,51 12 19,25 17,19 1232 2 46,23 25,67 13 5,08 1,75 13 22,62 16,75 13 18,54 24,58 1333 1 41,31 16,66 13 7,00 4,45 13 17,54 5,84 13 16,77 17,56 1334 2 44,54 19,26 13 4,54 2,37 13 20,38 10,90 13 18,38 20,29 1335 1 31,31 12,72 13 4,69 2,95 13 18,15 10,16 13 8,46 9,85 1336 2 46,15 15,99 13 6,38 6,19 13 21,62 8,60 13 18,15 15,97 1337 1 39,38 10,28 13 4,15 2,44 13 21,92 6,47 13 13,08 11,63 1338 2 31,83 12,67 6 7,17 7,78 6 17,67 7,03 6 7,00 7,43 639 1 46,92 32,02 13 4,31 2,39 13 26,77 21,15 13 15,85 15,28 1340 2 39,15 9,69 13 3,85 2,51 13 17,54 10,54 13 17,77 12,40 1341 1 38,23 13,40 13 4,77 3,56 13 19,15 6,73 13 14,54 13,95 1342 2 35,08 16,04 13 3,46 1,71 13 20,38 9,97 13 11,23 10,81 13

152
Tab. 12: Mittlere Zeitdauern einzelner Verhaltensausprägungen von Schlachtschweinen währendder Vereinzelung bei der Zuführung zur Betäubung
Verhalten während der Vereinzelung (Sekunden) (Teil 1)
freiwilligesVorwärtsgehen
Vorwärtsgehen nachTreibhilfe Vermeidungsverhalten
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n
2 2 0,00 0,00 13 6,54 3,69 13 3,62 5,22 133 1 0,46 1,13 13 7,08 2,06 13 6,00 10,83 134 2 0,00 0,00 12 5,00 1,86 12 2,83 3,38 125 1 0,08 0,28 13 5,23 2,45 13 2,00 2,04 136 2 0,23 0,60 13 5,00 2,24 13 3,15 6,19 137 1 0,08 0,28 13 5,31 3,50 13 4,77 8,95 138 2 0,46 1,39 13 4,77 2,42 13 4,00 5,52 139 1 0,38 1,12 13 4,92 2,02 13 1,38 2,29 1310 2 0,77 1,54 13 4,15 2,48 13 5,31 5,88 1311 1 1,00 1,41 13 5,31 2,53 13 5,54 7,78 1312 2 1,15 2,03 13 5,77 3,09 13 2,92 3,25 1313 2 0,18 0,60 11 5,64 2,87 11 0,55 1,51 1114 1 0,62 1,26 13 6,15 2,88 13 1,08 2,90 1315 1 0,46 1,39 13 5,54 2,76 13 2,23 3,98 1316 2 1,62 2,75 13 6,08 3,86 13 10,00 13,86 1317 1 1,85 4,18 13 4,69 2,36 13 4,15 7,21 1318 2 0,92 1,93 13 6,92 3,73 13 5,00 6,08 1319 1 0,00 0,00 13 6,77 2,35 13 7,08 16,30 1320 2 0,00 0,00 13 6,23 2,17 13 1,38 1,76 1321 1 0,77 1,92 13 4,08 2,60 13 2,54 8,00 1322 2 1,11 2,42 9 5,22 2,28 9 2,11 3,26 923 2 0,31 0,85 13 3,69 1,84 13 0,69 1,97 1324 1 0,33 1,15 12 2,58 1,31 12 0,75 1,42 1225 1 0,23 0,83 13 3,15 1,82 13 0,92 1,80 1326 1 0,38 0,96 13 3,15 1,52 13 0,92 2,29 1327 2 0,00 0,00 13 3,69 1,32 13 0,46 0,78 1328 2 0,00 0,00 13 3,92 1,55 13 1,46 4,16 1329 1 0,00 0,00 11 3,45 1,21 11 2,64 5,94 1130 2 0,23 0,83 13 3,46 1,71 13 0,46 1,66 1331 1 0,50 1,24 12 3,00 1,60 12 1,00 1,54 1232 2 0,85 1,72 13 3,62 2,33 13 0,38 0,96 1333 1 0,23 0,83 13 4,69 2,90 13 0,62 1,45 1334 2 0,15 0,55 13 3,62 1,71 13 0,54 1,45 1335 1 0,23 0,83 13 3,62 2,06 13 0,38 0,96 1336 2 0,38 0,96 13 3,08 1,80 13 2,92 5,69 1337 1 0,31 0,75 13 3,08 1,66 13 0,77 2,24 1338 2 0,00 0,00 6 4,50 1,38 6 2,67 6,53 639 1 0,23 0,83 13 3,15 1,57 13 0,69 1,70 1340 2 0,31 0,75 13 2,77 1,88 13 0,77 2,05 1341 1 0,31 1,11 13 3,54 1,56 13 0,92 3,33 1342 2 1,15 1,77 13 1,77 1,79 13 0,54 1,45 13
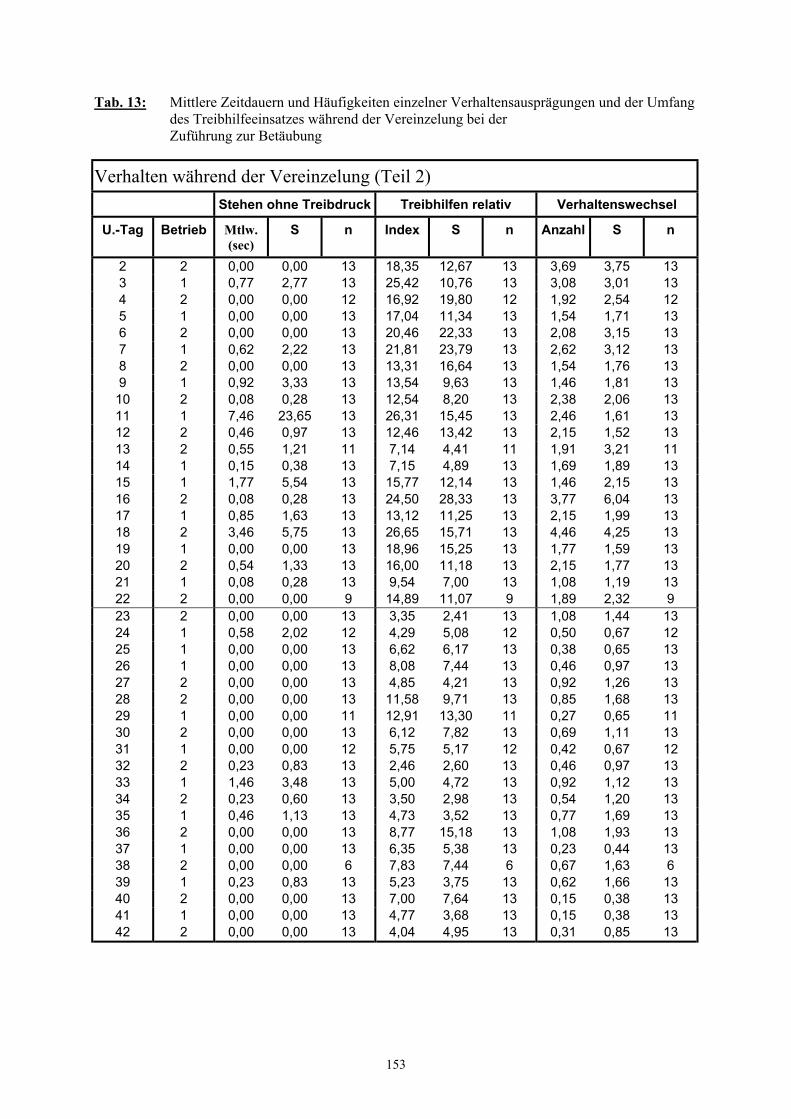
153
Tab. 13: Mittlere Zeitdauern und Häufigkeiten einzelner Verhaltensausprägungen und der Umfangdes Treibhilfeeinsatzes während der Vereinzelung bei derZuführung zur Betäubung
Verhalten während der Vereinzelung (Teil 2)
Stehen ohne Treibdruck Treibhilfen relativ Verhaltenswechsel
U.-Tag Betrieb Mtlw.(sec)
S n Index S n Anzahl S n
2 2 0,00 0,00 13 18,35 12,67 13 3,69 3,75 133 1 0,77 2,77 13 25,42 10,76 13 3,08 3,01 134 2 0,00 0,00 12 16,92 19,80 12 1,92 2,54 125 1 0,00 0,00 13 17,04 11,34 13 1,54 1,71 136 2 0,00 0,00 13 20,46 22,33 13 2,08 3,15 137 1 0,62 2,22 13 21,81 23,79 13 2,62 3,12 138 2 0,00 0,00 13 13,31 16,64 13 1,54 1,76 139 1 0,92 3,33 13 13,54 9,63 13 1,46 1,81 1310 2 0,08 0,28 13 12,54 8,20 13 2,38 2,06 1311 1 7,46 23,65 13 26,31 15,45 13 2,46 1,61 1312 2 0,46 0,97 13 12,46 13,42 13 2,15 1,52 1313 2 0,55 1,21 11 7,14 4,41 11 1,91 3,21 1114 1 0,15 0,38 13 7,15 4,89 13 1,69 1,89 1315 1 1,77 5,54 13 15,77 12,14 13 1,46 2,15 1316 2 0,08 0,28 13 24,50 28,33 13 3,77 6,04 1317 1 0,85 1,63 13 13,12 11,25 13 2,15 1,99 1318 2 3,46 5,75 13 26,65 15,71 13 4,46 4,25 1319 1 0,00 0,00 13 18,96 15,25 13 1,77 1,59 1320 2 0,54 1,33 13 16,00 11,18 13 2,15 1,77 1321 1 0,08 0,28 13 9,54 7,00 13 1,08 1,19 1322 2 0,00 0,00 9 14,89 11,07 9 1,89 2,32 923 2 0,00 0,00 13 3,35 2,41 13 1,08 1,44 1324 1 0,58 2,02 12 4,29 5,08 12 0,50 0,67 1225 1 0,00 0,00 13 6,62 6,17 13 0,38 0,65 1326 1 0,00 0,00 13 8,08 7,44 13 0,46 0,97 1327 2 0,00 0,00 13 4,85 4,21 13 0,92 1,26 1328 2 0,00 0,00 13 11,58 9,71 13 0,85 1,68 1329 1 0,00 0,00 11 12,91 13,30 11 0,27 0,65 1130 2 0,00 0,00 13 6,12 7,82 13 0,69 1,11 1331 1 0,00 0,00 12 5,75 5,17 12 0,42 0,67 1232 2 0,23 0,83 13 2,46 2,60 13 0,46 0,97 1333 1 1,46 3,48 13 5,00 4,72 13 0,92 1,12 1334 2 0,23 0,60 13 3,50 2,98 13 0,54 1,20 1335 1 0,46 1,13 13 4,73 3,52 13 0,77 1,69 1336 2 0,00 0,00 13 8,77 15,18 13 1,08 1,93 1337 1 0,00 0,00 13 6,35 5,38 13 0,23 0,44 1338 2 0,00 0,00 6 7,83 7,44 6 0,67 1,63 639 1 0,23 0,83 13 5,23 3,75 13 0,62 1,66 1340 2 0,00 0,00 13 7,00 7,64 13 0,15 0,38 1341 1 0,00 0,00 13 4,77 3,68 13 0,15 0,38 1342 2 0,00 0,00 13 4,04 4,95 13 0,31 0,85 13
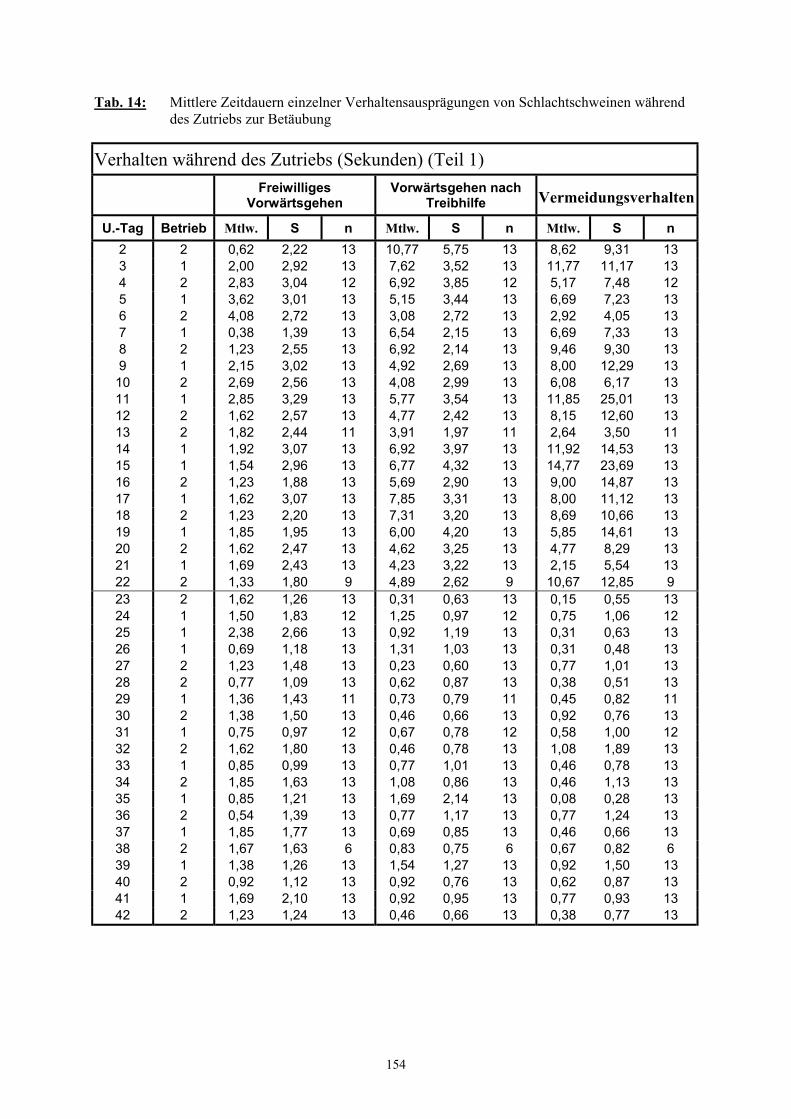
154
Tab. 14: Mittlere Zeitdauern einzelner Verhaltensausprägungen von Schlachtschweinen währenddes Zutriebs zur Betäubung
Verhalten während des Zutriebs (Sekunden) (Teil 1)
FreiwilligesVorwärtsgehen
Vorwärtsgehen nachTreibhilfe Vermeidungsverhalten
U.-Tag Betrieb Mtlw. S n Mtlw. S n Mtlw. S n
2 2 0,62 2,22 13 10,77 5,75 13 8,62 9,31 133 1 2,00 2,92 13 7,62 3,52 13 11,77 11,17 134 2 2,83 3,04 12 6,92 3,85 12 5,17 7,48 125 1 3,62 3,01 13 5,15 3,44 13 6,69 7,23 136 2 4,08 2,72 13 3,08 2,72 13 2,92 4,05 137 1 0,38 1,39 13 6,54 2,15 13 6,69 7,33 138 2 1,23 2,55 13 6,92 2,14 13 9,46 9,30 139 1 2,15 3,02 13 4,92 2,69 13 8,00 12,29 1310 2 2,69 2,56 13 4,08 2,99 13 6,08 6,17 1311 1 2,85 3,29 13 5,77 3,54 13 11,85 25,01 1312 2 1,62 2,57 13 4,77 2,42 13 8,15 12,60 1313 2 1,82 2,44 11 3,91 1,97 11 2,64 3,50 1114 1 1,92 3,07 13 6,92 3,97 13 11,92 14,53 1315 1 1,54 2,96 13 6,77 4,32 13 14,77 23,69 1316 2 1,23 1,88 13 5,69 2,90 13 9,00 14,87 1317 1 1,62 3,07 13 7,85 3,31 13 8,00 11,12 1318 2 1,23 2,20 13 7,31 3,20 13 8,69 10,66 1319 1 1,85 1,95 13 6,00 4,20 13 5,85 14,61 1320 2 1,62 2,47 13 4,62 3,25 13 4,77 8,29 1321 1 1,69 2,43 13 4,23 3,22 13 2,15 5,54 1322 2 1,33 1,80 9 4,89 2,62 9 10,67 12,85 923 2 1,62 1,26 13 0,31 0,63 13 0,15 0,55 1324 1 1,50 1,83 12 1,25 0,97 12 0,75 1,06 1225 1 2,38 2,66 13 0,92 1,19 13 0,31 0,63 1326 1 0,69 1,18 13 1,31 1,03 13 0,31 0,48 1327 2 1,23 1,48 13 0,23 0,60 13 0,77 1,01 1328 2 0,77 1,09 13 0,62 0,87 13 0,38 0,51 1329 1 1,36 1,43 11 0,73 0,79 11 0,45 0,82 1130 2 1,38 1,50 13 0,46 0,66 13 0,92 0,76 1331 1 0,75 0,97 12 0,67 0,78 12 0,58 1,00 1232 2 1,62 1,80 13 0,46 0,78 13 1,08 1,89 1333 1 0,85 0,99 13 0,77 1,01 13 0,46 0,78 1334 2 1,85 1,63 13 1,08 0,86 13 0,46 1,13 1335 1 0,85 1,21 13 1,69 2,14 13 0,08 0,28 1336 2 0,54 1,39 13 0,77 1,17 13 0,77 1,24 1337 1 1,85 1,77 13 0,69 0,85 13 0,46 0,66 1338 2 1,67 1,63 6 0,83 0,75 6 0,67 0,82 639 1 1,38 1,26 13 1,54 1,27 13 0,92 1,50 1340 2 0,92 1,12 13 0,92 0,76 13 0,62 0,87 1341 1 1,69 2,10 13 0,92 0,95 13 0,77 0,93 1342 2 1,23 1,24 13 0,46 0,66 13 0,38 0,77 13

155
Tab. 15: Mittlere Zeitdauern und Häufigkeiten einzelner Verhaltensausprägungen und der Umfangdes Treibhilfeeinsatzes während des Zutriebs zur Betäubung
Verhalten während des Zutriebs (Teil 2)
Stehen ohne Treibdruck Treibhilfen Verhaltenswechsel
U.-Tag Betrieb Mtlw.(sec)
S n Index S n Anzahl S n
2 2 10,85 9,47 13 71,46 67,68 13 8,46 5,19 133 1 8,00 11,12 13 72,69 80,12 13 5,46 2,44 134 2 13,00 12,22 12 35,08 46,44 12 5,17 4,95 125 1 13,00 12,38 13 55,85 54,05 13 5,92 3,59 136 2 9,31 6,60 13 30,08 48,30 13 6,00 3,39 137 1 0,00 0,00 13 49,69 27,81 13 4,77 2,24 138 2 13,23 30,51 13 83,92 89,57 13 5,38 2,66 139 1 8,77 9,64 13 45,62 38,09 13 5,00 2,27 1310 2 15,00 17,66 13 61,15 75,10 13 6,62 2,57 1311 1 6,00 8,65 13 113,31 183,77 13 7,38 8,19 1312 2 10,00 12,12 13 69,19 89,71 13 5,38 4,07 1313 2 12,82 14,78 11 17,27 14,26 11 4,73 2,37 1114 1 8,69 6,80 13 102,77 129,06 13 6,62 4,11 1315 1 5,92 7,38 13 87,00 157,71 13 4,85 4,47 1316 2 8,00 6,72 13 55,00 69,12 13 5,15 3,13 1317 1 13,31 16,23 13 58,23 50,19 13 7,54 2,88 1318 2 4,85 5,83 13 71,85 47,00 13 5,85 3,46 1319 1 11,92 11,75 13 45,62 57,10 13 6,31 4,91 1320 2 5,46 5,70 13 47,31 52,22 13 4,31 2,02 1321 1 5,77 7,77 13 26,31 31,89 13 3,77 3,52 1322 2 12,11 14,23 9 47,89 42,27 9 5,44 2,40 923 2 14,23 4,60 13 0,00 0,00 13 2,54 1,39 1324 1 19,67 12,06 12 0,00 0,00 12 4,25 3,05 1225 1 24,08 17,26 13 0,23 0,83 13 3,23 2,35 1326 1 21,46 15,61 13 0,38 0,96 13 2,38 1,56 1327 2 25,00 14,02 13 0,15 0,38 13 2,92 1,93 1328 2 17,15 7,82 13 0,50 1,22 13 2,46 1,66 1329 1 19,18 7,59 11 0,00 0,00 11 3,36 1,96 1130 2 19,38 10,78 13 0,23 0,83 13 3,69 1,38 1331 1 16,17 6,26 12 0,42 1,44 12 2,33 1,72 1232 2 19,46 14,54 13 0,00 0,00 13 3,15 3,26 1333 1 15,46 5,64 13 0,00 0,00 13 2,31 1,44 1334 2 17,00 11,37 13 0,38 0,87 13 3,31 2,02 1335 1 15,54 9,49 13 0,00 0,00 13 2,15 1,91 1336 2 19,54 8,09 13 0,85 1,68 13 2,31 1,93 1337 1 18,92 7,23 13 0,46 1,13 13 3,08 1,50 1338 2 14,50 6,06 6 0,50 1,22 6 3,50 1,87 639 1 22,92 21,80 13 0,46 1,20 13 3,08 2,06 1340 2 15,08 10,87 13 0,08 0,28 13 3,00 1,35 1341 1 15,77 6,87 13 0,69 1,70 13 3,31 2,29 1342 2 18,31 10,52 13 0,85 2,51 13 2,69 1,84 13


Danksagung
Herrn Prof. Dr. J. Hartung danke ich herzlich für die Überlassung des Themas und diefreundliche Unterstützung bei der Abfassung der Arbeit.
Den Mitarbeitern des Instituts danke ich für ihre freundliche Unterstützung. Mein besondererDank gilt Herrn Dr. Michael Marahrens für die prompte und sehr gute Hilfestellung, die ermir in allen Belangen meiner Arbeit leistete. Hervorheben möchte ich ebenfalls HaraldUlbrich, der mir immer tatkräftig zur Seite stand.
Für ihre Hilfe und Geduld bei den Laboranalysen danke ich den Mitarbeitern der Rinderklinikder TiHo Hannover und den anderen beteiligten Instituten.
Herrn Dr. Thomas Schmidt danke ich herzlich für seine hilfreiche Unterstützung bei derstatistischen Bearbeitung des Datenmaterials.
Den Mitarbeitern des Schlachthofs der EGO schulde ich Dank dafür, dass sie dieDurchführung der Versuche ermöglichten und optimal unterstützten. Ebenso möchte ich michfür die Bereitstellung der Versuchsschweine und die gewährte Hilfe recht herzlich bei denLandwirten Martin Uthoff und Georg Dütemeyer und ihren Familien bedanken.
Bedanken möchte ich mich auch bei meinen Freunden und den Mitgliedern derRettungshundestaffel Münsterland für die mentale Unterstützung und die Hilfe beiComputerproblemen jeglicher Art.
Bei meiner Oma, meinen Eltern und meiner Schwiegermutter möchte ich mich herzlich fürdie Unterstützung, die sie mir gewährt haben, bedanken.
Mein ganz besonderer Dank gilt meinem Mann Helmuth für seine tatkräftige Hilfe in derEndphase der Arbeit. Weiterhin möchte ich mich bei ihm und unserem Sohn Ben für diebewunderswerte Geduld, die sie für mich aufgebracht haben, bedanken.





























