Embed Size (px)
Citation preview

This article was downloaded by: [Missouri S & T]On: 13 January 2014, At: 22:19Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
International Journal ofPhytoremediationPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/bijp20
Pilot Study of Bioaccumulation andDistribution of Cesium, Potassium,Sodium and Calcium in King OysterMushroom (Pleurotus Eryngii) GrownUnder Controlled ConditionsGrażyna Bystrzejewska-Piotrowska a , Dariusz Pianka a , Michał A.
Bazała a , Romuald Stęborowski a , José L. Manjón b & Pawel L.
Urban aa Isotope Laboratory, Faculty of Biology , University of Warsaw ,Warsaw, Polandb Departamento de Biología Vegetal, Facultad de Biología ,Universidad de Alcalá , Alcalá de Henares (Madrid), SpainPublished online: 20 Nov 2008.
To cite this article: Grażyna Bystrzejewska-Piotrowska , Dariusz Pianka , Michał A. Bazała ,Romuald Stęborowski , José L. Manjón & Pawel L. Urban (2008) Pilot Study of Bioaccumulation andDistribution of Cesium, Potassium, Sodium and Calcium in King Oyster Mushroom (Pleurotus Eryngii)Grown Under Controlled Conditions, International Journal of Phytoremediation, 10:6, 503-514, DOI:10.1080/15226510802114987
To link to this article: http://dx.doi.org/10.1080/15226510802114987
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.

This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

International Journal of Phytoremediation, 10:503–514, 2008Copyright C© Taylor & Francis Group, LLCISSN: 1522-6514 print / 1549-7879 onlineDOI: 10.1080/15226510802114987
PILOT STUDY OF BIOACCUMULATION ANDDISTRIBUTION OF CESIUM, POTASSIUM, SODIUMAND CALCIUM IN KING OYSTER MUSHROOM(PLEUROTUS ERYNGII) GROWN UNDER CONTROLLEDCONDITIONS
Grazyna Bystrzejewska-Piotrowska,1 Dariusz Pianka,1
Michał A. Bazała,1 Romuald St↪eborowski,1 Jose L. Manjon,2
and Pawel L. Urban1
1Isotope Laboratory, Faculty of Biology, University of Warsaw, Warsaw, Poland2Departamento de Biologıa Vegetal, Facultad de Biologıa, Universidad de Alcala,Alcala de Henares (Madrid), Spain
This pilot study presents preliminary results on interrelations between alkali and alkalineearth elements during their transfer to mycelium and fruitbodies of saprophytic fungi. Theaccumulation and distribution of four elements (cesium, potassium, sodium, and calcium)was evaluated in king oyster mushroom (Pleurotus eryngii) cultivated under controlledconditions. Elemental composition of caps, stipes, and the substrate was analyzed by atomicabsorption/emission spectroscopy to evaluate discrimination, concentration, and transferfactors. The transfer factors determined for all the investigated elements were different andcan be put in the following order: Cs > K > Na > Ca. There has been a higher accumulationof cesium in caps than in stipes. Distribution of cesium in fruitbodies depended on the pres-ence of other ions in the substrate. The addition of Ca2+ limited the transport of cesium andpotassium from stipes to caps. Sodium and calcium were mainly accumulated in the stipes.In a control experiment, without supplementation with K+, Na+, and Ca2+, ∼ 62% of thecesium present in the substrate was extracted by mycelium and transported to the fruitbodies.Possible applications of fruiting saprophytic fungi in bioremediation are discussed.
KEY WORDS bioremediation, cesium hyperaccumulation, discrimination factor (DF),mycoextraction, transfer factor (TF)
INTRODUCTION
Mycoextraction (Manjon, Urban, and Bystrzejewska-Piotrowska, 2004) can bedefined as a process leading to the uptake and accumulation of chemical species in cells,mycelium, and fruitbodies of fungi. In principle, fungi can take up and decompose variousorganic and inorganic substances present in the substrate media (Kalac and Svoboda, 2000;Kalac, 2001; Stamets, 2000; Gadd, 2001; Falandysz et al., 2004; Singh, 2007). Chemicalelements and their complexes, as well as naturallyoccurring and xenobiotic compounds,
Address correspondence to Grazyna Bystrzejewska-Piotrowska, Isotope Laboratory, Faculty of Biology,University of Warsaw, 02-096, Warsaw, Miecznikowa 1, Poland. E-mail: [email protected]
503
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

504 G. BYSTRZEJEWSKA-PIOTROWSKA ET AL.
can be withdrawn from the medium and, in the case of some chemical species, metabolizedby mycelium (van Elteren, Woroniecka, and Kroon, 1998; Gadd, 2001, 2007). Some fungalspecies possess the ability to hyperaccumulate environmental pollutants—for example,radionuclides (Bazała, Bystrzejewska-Piotrowska, and Cipakova, 2005)—feature that couldbe utilized in bioremediation and biorestoration mediated by fungi (Pletsch, de Araujo, andCharlwood, 1999; Gadd, 2001; Stamets, 2005; Singh, 2007). These two processes have alsobeen referred to as mycoremediation and mycorestoration, respectively (Stamets, 2005).
In numerous studies conducted on environmental specimens, it has become apparentthat fungi possess a great ability to take up and accumulate cesium (Mietelski et al., 1994;Yoshida and Muramatsu, 1998; Bystrzejewska-Piotrowska, Urban, and St
↪eborowski, 2003;
Baeza, Guillen, and Bernedo, 2005; Cipakova, 2005; Kuwahara et al., 2005; Karadeniz andYaprak, 2007). Radiocesium (134Cs and 137Cs), mainly originating from the accident in thenuclear power plant near Chernobyl (Ukraine) in 1986, is one of the most importantradioactive contaminants of European soils (Peplow, 2006). While most studies wereconducted on fruiting basidiomycetes collected at different locations (Mietelski et al.,1994; Bystrzejewska-Piotrowska et al., 2003; Bazała et al., 2005; Cipakova, 2005), scarcestudies considered the physiological abilities of macromycetes, which are responsible forthe transfer of cesium to mushrooms (Terada et al., 1998; van Elteren et al., 1998; Baezaet al., 2000; Kalac, 2001; Bystrzejewska-Piotrowska et al., 2005; Baeza et al., 2006; Bazała,Golda, and Bystrzejewska-Piotrowska, 2008; Bystrzejewska-Piotrowska and Bazała, 2008).
Fruitbodies of higher fungi (mushrooms) are characterised by fast and intensivegrowth, which is also accompanied by the uptake of vast amounts of water containingdissolved elements (Griffin, 1994). Therefore, the physiological condition and degree ofmaturity is also important for the uptake and accumulation of cesium in mushrooms (Baezaet al., 2006). The level of accumulation may also depend on the age of the mycelium(Svoboda, Zimmermannova, and Kalac, 2000): the older it is, more of an element it hasaccumulated. In the environment, many other factors can affect the accumulation of cesiumand other elements in the fruitbodies of macromycetes. In environmental studies, soilcharacteristics have a huge influence on the bioavailability of cesium. For example, theuptake of cesium can be suppressed by complexing agents that are present in the soil(Wauters et al., 1996). In soils with a lower potassium concentration, cesium is taken upto a greater extent (Kruyts and Delvaux, 2002). The presence of microorganisms can alsoincrease the bioavailability of radiocesium (Sanchez et al., 2000). The influence of allthese factors can be eliminated in the laboratory-based studies aiming to investigate thephysiological processes that are responsible for the uptake, transport, and accumulation ofcesium and other chemical species in higher fungi.
Interactions between cesium and other cations during their transfer from the envi-ronment to fungi as well as plants are still a subject of debate (Yoshida and Muramatsu,1998; Rodrıguez-Navarro, 2000; Gadd, 2001; Urban and Bystrzejewska-Piotrowska, 2003;Bystrzejewska-Piotrowska and Urban, 2004; Le Lay et al., 2006). Since all mushrooms growon different types of soil and in distinct climate conditions, the observations made cannotbe generalized and the influence of other elements on the uptake of cesium is obscured.For this reason, it has been difficult to describe, in an unbiased way, the influence of othernaturally occurring elements, such as potassium, sodium, and calcium, on the accumulationof the contaminant cesium. In order to study the physiological mechanisms of uptake andtransport of cesium in fruiting fungi, including edible basidiomycetes, it is essential to carryout experiments in well-defined and controlled conditions. Experimental cultures shouldexclude influence of any indeterminate growth media, e.g., soil. Standardization should
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

BIOACCUMULATION AND DISTRIBUTION IN KING OYSTER MUSHROOM 505
include parameters such as illumination, temperature, and humidity (Manjon et al., 2004).Unlike when studying responses of plants to environmental stress, it is extremely difficultto carry out experiments on “pure” cultures of fungi, which would exclude contact withsoil or any other complex growth matrices. In order to overcome this problem, and tofacilitate physiological studies on fruiting saprophytic fungi, a simple protocol using kingoyster mushroom (Pleurotus eryngii) as a model species has been developed (Manjonet al., 2004). The present study follows previous reports in which the accumulationand distribution of different radionuclides were described for the same species (Baezaet al., 2000; Bystrzejewska-Piotrowska et al., 2005; Urban et al., 2005; Baeza et al.,2006). This species also exhibits a high ability to accumulate cesium and radiocesium(Bystrzejewska-Piotrowska et al., 2005; Bazała et al., 2008; Bystrzejewska-Piotrowskaand Bazała, 2008).
The main goal of the present study was to assess the ability of the model saprophyticbasidiomycete to take up, transport, and accumulate cesium in the presence of otherelements added to the growth medium. Considering conclusions of the previous studieson the accumulation of cesium in plants (Urban and Bystrzejewska-Piotrowska, 2003;Bystrzejewska-Piotrowska and Urban, 2004) and the above-cited reports on fungi, wehypothesise that alkali and alkaline earth elements may influence physiological processessuch as the ionic uptake, transport, and distribution of cesium, leading to mycoextractionof this element from growth media. All of these fundamental processes need to be studiedcarefully before higher fungi can be utilized in bioremediation protocols. In contrast to ourprevious study using a 137Cs radiotracer, in the present work we aimed to analyze the contentof all the elements of interest using a non-radiochemical technique. This approach enablesstudy of the interdependence of transport and accumulation of cesium and other elements.
MATERIALS AND METHODS
In the present work, we have re-analyzed samples obtained in the previous radiotracer-aided study (Bystrzejewska-Piotrowska et al., 2005), this time using the atomic absorptionspectroscopy (AAS) method, so as to obtain information on the bulk content of differentelements in the parts of fruitbodies and the growth medium (substrate/mycelium).
Briefly, in order to prepare medium for the growth of fungal mycelium, barley seeds(Plant Breeding and Acclimatisation Institute—IHAR, Radzikow, Poland) were submergedin water for 24 h. The hydrated seeds were then packed into plastic containers (Duchefa,Haarlem, the Netherlands), covered, and sterilized in an autoclave at 120◦C for 2 h. Thissubstrate did not contain cesium as confirmed using AAS; it contained potassium (2.3 mgg−1 d.w.), sodium (2.1 mg g−1 d.w.), and calcium (0.135 mg g−1 d.w.).
Each container was inoculated with ∼16 g of the inoculum. The utilized strain ofking oyster mushroom, Pleurotus eryngii, had previously been isolated from a fruitbodyfound growing on the remains of roots of an Eryngium campestre plant found in thelocality of Camarma de Esteruelas (3◦ 22 45 W, 40◦ 33′ 15′′ N, 30T 4684489 UTM; MadridProvince, Spain) (Manjon et al., 2004; Bystrzejewska-Piotrowska et al., 2005; Urban et al.,2005). The culture protocol described by Manjon et al. (2004) was followed with somemodifications. Mycelium was developing during ∼30 d in darkness at 25◦C until thewhole substrate block was coated with hyphae forming compact spawn. In order to induceformation of fruitbodies, the containers were transferred to a growth chamber with thethermostat temperature set to 18◦C and the controlled relative humidity of 80%. Thephotoperiod was set to a 16-h day (8-h night).
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

506 G. BYSTRZEJEWSKA-PIOTROWSKA ET AL.
All the salts used in the culture experiment (KCl, NaCl, CaCl2, and CsCl) wereanalytical-grade chemicals (POCh, Gliwice, Poland). The radioactive tracer, ∼100 kBqml−1 137CsCl, was purchased from Polatom (Otwock, Poland). All the test solutionscontained 137Cs radiotracer and, optionally, stable cesium (0.1 mM) and appropriate salts(10 or 100 mM). Two control experiments were carried out, using either CsCl suppliedin trace amounts (137CsCl radiotracer only) or 0.1 mM CsCl with radiotracer. Cesiumconcentration in the test solution containing the 137CsCl radiotracer only was ∼1.13 ×10-4 mM (further referred to as "trace Cs”). Twenty milliliters out of a 50-mL aliquot ofeach test solution was used to moisten the spawn block in each replicate container. Then,the spawn blocks were covered with ∼15 g of sterile peat soil. The residue 30 mL of thetest solution was split into three aliquots (10 mL each), subsequently introduced to thespawn/container interface three times with 24-h intervals. The fruitbodies were collectedafter ∼14 d following the induction of fructification. Only the first harvest of fruitbodieswas used in this study. Three containers were prepared for each experimental variant andtwo samples were collected from each container. Bulk samples of caps and stipes wereprepared using material from several fruitbodies. This was necessary due to the apparentvariability of specimens in the culture.
The collected samples were dried at 105◦C for 24 h and homogenized as describedpreviously (Bystrzejewska-Piotrowska et al., 2005). Before further analysis, they weresubjected to additional drying at 85◦C. Two hundred milligrams of mushroom and mediumsamples were mineralized in a microwave oven Ethos Plus (Milestone, Shelton, CT, USA)with a 65:35 (v/v) mixture of HNO3 and H2O2 (Merck, Darmstadt, Germany). Analysis ofthe samples was carried out using an atomic absorption/emission spectrometer, Solar M6(Thermo Fisher Scientific, Waltham, MA, USA), with background correction and using theQuadline and HCL lamps (TJA Solutions, Franklin, MA, USA). Potassium, sodium, andcalcium were determined using the absorption mode and the wavelengths: 766, 589, and 423nm, respectively. Cesium was analyzed in the emission mode at 852 nm. Cesium chloride,used for preparation of the standard solutions, was from Riedel-de Haen (Seelze, Germany),whereas potassium nitrate, sodium nitrate, and calcium nitrate were from Merck. All thesamples and standard solutions were analyzed in triplicate. Quality control procedures werefollowed: the blank and one standard solution of each analyte were run every 10 measure-ments and the standard deviation was always below 10%. The calibration plot was madefor five standard solutions and concentrations of the analytes were determined using thesoftware accompanying the AAS instrument. The obtained values were scaled against dryweight of the initial samples. Unlike the previous study (Bystrzejewska-Piotrowska et al.,2005), no further radiometric measurements were performed.
The data treatment was carried out using MS Excel 2000 software. Transfer factors(TFs), concentration factors (CFs), and discrimination factors (DFs) for different elementssupplied to the mycelium were determined as in our previous studies (Bystrzejewska-Piotrowska et al., 2003, 2005):
TF = Cupper
Csubstrate(1)
DF =(
C1upper
C2upper
)(
C1lowerC2lower
) (2)
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

BIOACCUMULATION AND DISTRIBUTION IN KING OYSTER MUSHROOM 507
CF = Cupper
Clower(3)
where C (C1 and C2) are concentrations of elements (1 and 2) measured in the upper (capsor stipes) and the lower parts (stipes or substrate) of the studied system. Mean and spreadvalues were normally calculated for two independent results.
RESULTS AND DISCUSSION
The culture protocol using Pleurotus eryngii as a model macromycete seems to bevery helpful in studies of ecotoxicology and radioecology, since use of complex growthmedia can easily be limited (Manjon et al., 2004). In the previous studies, carried out onthe same species (Bystrzejewska-Piotrowska et al., 2005; Bystrzejewska-Piotrowska andBazała, 2008), the influence of potassium, sodium, and calcium on the accumulation ofradioactive cesium-137 was investigated. The present study is based on the analysis of totalconcentration of cesium and other elements using a non-radiometric method.
Cesium concentrations in the substrate contaminated with 0.1 mM CsCl, measured atthe beginning of the culture and after the harvest of fruitbodies, were 4.67 and 1.78 µg g−1
d.w., respectively. Therefore, 62% of the cesium supplied to the culture was extracted by themycelium and transported to fruitbodies, which shows a high efficiency of mycoextractionin the tested system. The measured value is similar to the value calculated from the resultsobtained in the radiotracer-aided study: 64% (Bystrzejewska-Piotrowska et al., 2005). Thiscomparison shows that either stable or radioactive isotopes can readily be used to evaluateefficiency of fungal bioremediation.
The concentration of stable cesium reported in other work for the samples of wildmushrooms was in the range 0.044 × 10−3−20 × 10−3 mg g−1 d.w. (Yoshida andMuramatsu, 1998). In our work, cesium concentration measured in the caps was one orderof magnitude higher than that observed in wild mushrooms: the values were 0.030 mg g−1
d.w. (after treatment with 0.1 mM CsCl) and 0.002 mg g−1 d.w., respectively. Potassiumconcentration, measured in the caps of mushrooms cultured in the present study, was atthe same level as mean potassium concentration measured in wild mushrooms. This can beexplained with existence of mechanisms regulating uptake of minerals in mushrooms: evenwhen availability of macroelements is limited by composition of the growth medium (barleyseeds or soil), mycelium can still maintain appropriate concentration of these elements infruitbodies at the level observed in the specimens collected in the wild.
TFs
The TF determined for cesium was the highest among all the investigated elementsand amounted to ∼17.1 in the variant with 0.1 mM Cs+ (Table 1). Differences in theTFs estimated for cesium in the present and previous studies (Bystrzejewska-Piotrowskaet al., 2005) are explained with different methodology used to determine the TFs. Inthe present study, cesium concentration measured in fruitbodies was divided by cesiumconcentration in the bulk growth medium determined after termination of the culture, i.e.,following the final harvest of fruitbodies. During the culture of P. eryngii and formation offruitbodies, a significant part of cesium initially present in the spawn block was depleted.Therefore, cesium concentration in the medium after the experiment is lower than before
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014
508 G. BYSTRZEJEWSKA-PIOTROWSKA ET AL.
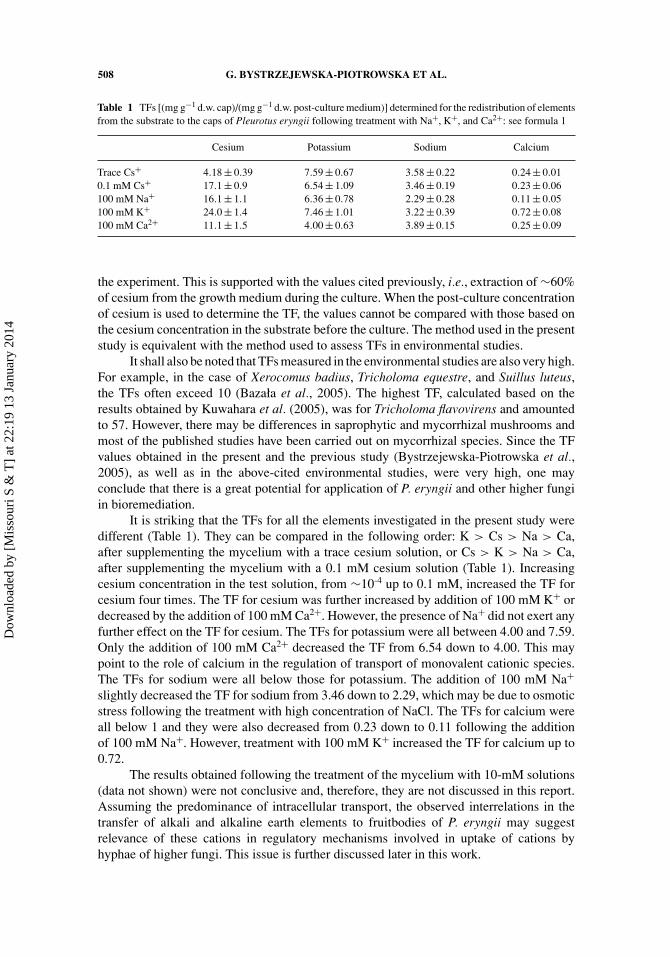
Table 1 TFs [(mg g−1 d.w. cap)/(mg g−1 d.w. post-culture medium)] determined for the redistribution of elementsfrom the substrate to the caps of Pleurotus eryngii following treatment with Na+, K+, and Ca2+: see formula 1
Cesium Potassium Sodium Calcium
Trace Cs+ 4.18 ± 0.39 7.59 ± 0.67 3.58 ± 0.22 0.24 ± 0.010.1 mM Cs+ 17.1 ± 0.9 6.54 ± 1.09 3.46 ± 0.19 0.23 ± 0.06100 mM Na+ 16.1 ± 1.1 6.36 ± 0.78 2.29 ± 0.28 0.11 ± 0.05100 mM K+ 24.0 ± 1.4 7.46 ± 1.01 3.22 ± 0.39 0.72 ± 0.08100 mM Ca2+ 11.1 ± 1.5 4.00 ± 0.63 3.89 ± 0.15 0.25 ± 0.09
the experiment. This is supported with the values cited previously, i.e., extraction of ∼60%of cesium from the growth medium during the culture. When the post-culture concentrationof cesium is used to determine the TF, the values cannot be compared with those based onthe cesium concentration in the substrate before the culture. The method used in the presentstudy is equivalent with the method used to assess TFs in environmental studies.
It shall also be noted that TFs measured in the environmental studies are also very high.For example, in the case of Xerocomus badius, Tricholoma equestre, and Suillus luteus,the TFs often exceed 10 (Bazała et al., 2005). The highest TF, calculated based on theresults obtained by Kuwahara et al. (2005), was for Tricholoma flavovirens and amountedto 57. However, there may be differences in saprophytic and mycorrhizal mushrooms andmost of the published studies have been carried out on mycorrhizal species. Since the TFvalues obtained in the present and the previous study (Bystrzejewska-Piotrowska et al.,2005), as well as in the above-cited environmental studies, were very high, one mayconclude that there is a great potential for application of P. eryngii and other higher fungiin bioremediation.
It is striking that the TFs for all the elements investigated in the present study weredifferent (Table 1). They can be compared in the following order: K > Cs > Na > Ca,after supplementing the mycelium with a trace cesium solution, or Cs > K > Na > Ca,after supplementing the mycelium with a 0.1 mM cesium solution (Table 1). Increasingcesium concentration in the test solution, from ∼10-4 up to 0.1 mM, increased the TF forcesium four times. The TF for cesium was further increased by addition of 100 mM K+ ordecreased by the addition of 100 mM Ca2+. However, the presence of Na+ did not exert anyfurther effect on the TF for cesium. The TFs for potassium were all between 4.00 and 7.59.Only the addition of 100 mM Ca2+ decreased the TF from 6.54 down to 4.00. This maypoint to the role of calcium in the regulation of transport of monovalent cationic species.The TFs for sodium were all below those for potassium. The addition of 100 mM Na+
slightly decreased the TF for sodium from 3.46 down to 2.29, which may be due to osmoticstress following the treatment with high concentration of NaCl. The TFs for calcium wereall below 1 and they were also decreased from 0.23 down to 0.11 following the additionof 100 mM Na+. However, treatment with 100 mM K+ increased the TF for calcium up to0.72.
The results obtained following the treatment of the mycelium with 10-mM solutions(data not shown) were not conclusive and, therefore, they are not discussed in this report.Assuming the predominance of intracellular transport, the observed interrelations in thetransfer of alkali and alkaline earth elements to fruitbodies of P. eryngii may suggestrelevance of these cations in regulatory mechanisms involved in uptake of cations byhyphae of higher fungi. This issue is further discussed later in this work.
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

BIOACCUMULATION AND DISTRIBUTION IN KING OYSTER MUSHROOM 509
Figure 1 DFs following treatment of mycelium with solutions of Cs+, K+, Na+, and Ca2+: Cesium/potassiumDF, cesium/sodium DF, cesium/calcium DF. See formula 2. Error bars signify spread of the experimental datapoints.
In general, analysis of the transfer factors showed a greater bioavailability of cesium,potassium, sodium, and calcium for the mycelium of P. eryngii grown on barley seeds (Table1) than it was observed in a study of wild mushrooms growing in a pine forest (Yoshidaand Muramatsu, 1998). In the case of cesium and calcium, this difference amounts to oneorder of magnitude and, in the case of sodium, two orders of magnitude. Also, the TF forpotassium was higher in the present study than the TFs based on data presented in theabove-cited environmental study (Yoshida and Muramatsu, 1998).
DFs
Discrimination of the uptake, transport, and accumulation of one ion by another ioncan be assessed by analysis of DFs, which are calculated according to equation 2. Forexample, DFs can be determined for the transfer of cesium and potassium from substrate tofruitbodies or from stipes to caps (Bystrzejewska-Piotrowska et al., 2003). The DF values(cesium/potassium) lower than 1 indicate that the transport of potassium prevails over thatof cesium.
Except for the control experiment (trace Cs, no stable cesium added), all the calculatedDF values (cesium/potassium, cesium/sodium, cesium/calcium) were higher than 1 (Figure1). Increasing cesium concentration from the trace level up to 0.1 mM (in the solutionsupplemented to the substrate medium) eliminated discrimination of cesium transport fromthe substrate to the cap and from the substrate to the stipe by potassium (Figure 1). A similareffect was observed following supplementation of the medium with 100 mM K+, Na+, andCa2+. The increase of cesium level in the substrate medium eliminated discrimination ofcesium by sodium even to a greater extent than was the case in the discrimination ofcesium by potassium (Figure 1). Discrimination of cesium by sodium, in the transport tothe cap and to the stipe, was limited by the addition of 100 mM K+ to a greater extent thanby any other cations (Figure 1). This observation shows that transport of sodium to thefruitbodies is strongly discriminated by cesium in the presence of potassium. Increasingcesium concentration up to 0.1 mM favored discrimination of calcium by cesium (Figure 1).The strongest discrimination of calcium by cesium was observed following supplementation
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

510 G. BYSTRZEJEWSKA-PIOTROWSKA ET AL.
with 100 mM Na+. It should be noted that in almost all cases, excluding the variant withadded cesium in trace levels, there was no discrimination of cesium observed. These wereother ions, i.e., Na+, K+ and Ca2+, which were discriminated by Cs+ in the most of theexperiments carried out (Figure 1).
It must be noted that, when analyzing environmental samples, one can observe a highvariability of DFs (Bystrzejewska-Piotrowska et al., 2003; Bazała et al., 2005; Kuwaharaet al., 2005). For example, the DFs calculated using the data obtained by Kuwahara et al.(2005) for wild mushrooms were in the range of 0.348–2.58 and there was no correlationbetween the DFs and the concentrations of stable cesium and potassium in the soil. Thisvariability of DFs may be due to heterogeneity of the environment and, in particular,differences in chemical composition of the soil that also influence the bioavailabilityof elements. However, in the present study, such variability was practically eliminatedby using standardized growth medium and controlling other experimental conditions.Therefore, we suggest that the discrimination factors could more likely be used to studyphysiological properties of model species grown under controlled conditions and usingsubstrates supplemented with different elements. Unlike in the environmental conditions,in the present study bioavailability of minerals could not be readily modified by mycelium,for instance, by secretion of H+ from the hyphae (bioweathering) (Gadd, 2007).
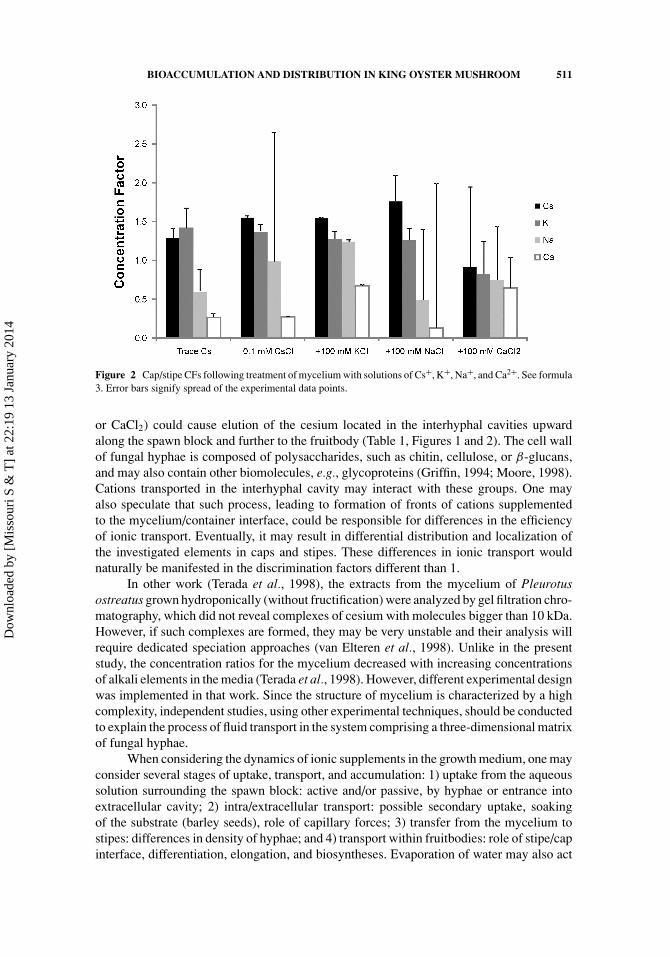
CFs
An analysis of CFs shows the preference of accumulation of the investigated elements(cesium, potassium, sodium, and calcium) within the fruitbodies. Distribution of theseelements was investigated in different parts of fruitbodies (caps and stipes). Similar toprevious studies (Baeza et al., 2006, Bystrzejewska-Piotrowska et al., 2005), there hasbeen higher accumulation of cesium in the caps than in the stipes (Figure 2). Increasing theconcentration of Cs+, K+, and Na+ did not change this effect. A similar effect was observedin the case of the distribution of potassium within fruitbodies (Figure 2). The addition of100 mM K+ increased the accumulation of sodium in the caps. On the other hand, calciumwas mainly accumulated in the hyphae of the stipes (Figure 2): all the CF values were lowerthan 1. The CF for calcium was only increased after supplementation of the medium with100 mM K+ and Ca2+.
Potassium plays an important role in the maintenance of enzyme activity, carbohy-drate metabolism, and ionic balance (Griffin, 1994), processes that are essential for growth(biosynthesis) and differentiation of cap and gills. Therefore, there may exist translocationof potassium and similar alkali metal ions to the upper part of the fruitbody. In fact, it hasalready been suggested that transport of cesium and potassium in plants and fungi may bemediated by similar mechanisms, including channels and other carriers present in cellularmembranes (Rodrıguez-Navarro, 2000; Bystrzejewska-Piotrowska et al., 2003; Urbanet al., 2003; Bystrzejewska-Piotrowska and Bazała, 2008).
Ionic interactions observed in the transport of different chemical species may beexplained by competitive binding to the transporters and differences in their affinities tothese transporters (Bystrzejewska-Piotrowska and Bazała, 2008). However, the observeddifferences in transport and accumulation of different alkali and alkaline earth elementscan also be explained in terms of passive extracellular transport based on capillary forcesdriving the aqueous solutions of dissociated salts upward, i.e., from the spawn hyphae tothe fruitbodies. The application of solutions with high ionic strength (100 mM KCl, NaCl,
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014
BIOACCUMULATION AND DISTRIBUTION IN KING OYSTER MUSHROOM 511
Figure 2 Cap/stipe CFs following treatment of mycelium with solutions of Cs+, K+, Na+, and Ca2+. See formula3. Error bars signify spread of the experimental data points.
or CaCl2) could cause elution of the cesium located in the interhyphal cavities upwardalong the spawn block and further to the fruitbody (Table 1, Figures 1 and 2). The cell wallof fungal hyphae is composed of polysaccharides, such as chitin, cellulose, or β-glucans,and may also contain other biomolecules, e.g., glycoproteins (Griffin, 1994; Moore, 1998).Cations transported in the interhyphal cavity may interact with these groups. One mayalso speculate that such process, leading to formation of fronts of cations supplementedto the mycelium/container interface, could be responsible for differences in the efficiencyof ionic transport. Eventually, it may result in differential distribution and localization ofthe investigated elements in caps and stipes. These differences in ionic transport wouldnaturally be manifested in the discrimination factors different than 1.
In other work (Terada et al., 1998), the extracts from the mycelium of Pleurotusostreatus grown hydroponically (without fructification) were analyzed by gel filtration chro-matography, which did not reveal complexes of cesium with molecules bigger than 10 kDa.However, if such complexes are formed, they may be very unstable and their analysis willrequire dedicated speciation approaches (van Elteren et al., 1998). Unlike in the presentstudy, the concentration ratios for the mycelium decreased with increasing concentrationsof alkali elements in the media (Terada et al., 1998). However, different experimental designwas implemented in that work. Since the structure of mycelium is characterized by a highcomplexity, independent studies, using other experimental techniques, should be conductedto explain the process of fluid transport in the system comprising a three-dimensional matrixof fungal hyphae.
When considering the dynamics of ionic supplements in the growth medium, one mayconsider several stages of uptake, transport, and accumulation: 1) uptake from the aqueoussolution surrounding the spawn block: active and/or passive, by hyphae or entrance intoextracellular cavity; 2) intra/extracellular transport: possible secondary uptake, soakingof the substrate (barley seeds), role of capillary forces; 3) transfer from the mycelium tostipes: differences in density of hyphae; and 4) transport within fruitbodies: role of stipe/capinterface, differentiation, elongation, and biosyntheses. Evaporation of water may also act
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

512 G. BYSTRZEJEWSKA-PIOTROWSKA ET AL.
as a pumping force for the transport of water and solutes in the mycelia and the fruitbodies.This shows that although macromycetes are supposed to be simpler, less-differentiatedbiological organisms than plants, yet interpretation of ionic transport in fruiting fungi maybe complex and require further experimental efforts.
CONCLUDING REMARKS
The results obtained in this study demonstrate that P. eryngii can selectively extractcesium from the substrate medium with a high efficiency. This conclusion is supportedby the result of depletion of cesium in the growth substrate caused by mycoextraction andaccumulation of cesium in fruitbodies of P. eryngii. The study has confirmed our assumptionthat there exists a strong influence of alkali and alkaline earth elements on the uptake,transport, and distribution of cesium among the mycelium, fruitbodies, and their parts. Thetransfer factors, showing characteristics of the fruitbodies in terms of accumulation of theinvestigated elements, were in agreement with the order of affinity of the ions transferred viapotassium carriers, i.e., K+ > Cs+ > Na+ > Ca2+ (trace Cs concentration). Increasing thecesium concentration affected preference of transport toward cesium: it increased the TFs.The observation of discrimination of potassium by cesium may also point to the increaseof the passive extracellular (interhyphal) transport when the medium is supplemented witha solution of high ionic strength.
It is clear that the distribution of cesium in the fruitbodies depends on other ionicspecies added to the medium. Therefore, there is enough evidence to conclude thatthe culture conditions, particularly chemical composition of the medium, can affect theefficiency of mycoextraction of cesium and other ionic species. This information willespecially be relevant when saprophytic basidiomycetes, such as P. eryngii, start to be usedin bioremediation protocols. However, much effort still has to be dedicated to bioengineeringaspects of mycoremediation in order to transfer the fundamental research findings into thenatural environment.
It is necessary to note that the conclusions presented in this work are derived fromthe outcomes of small-scale laboratory experiments; therefore, they can only be consideredpreliminary results of essentially pilot studies. Further experiments including a large numberof replicates should verify these initial findings. Future studies should also focus on furthermechanistic explanation of the process of ionic transport within the mycelium and the ionicdiscrimination. An especially interesting question is to what extent the ions are transportedthrough the extracellular pathway, i.e., through the cavity between the hyphae of themycelium. Another goal is to study the behavior of multivalent elements, including heavymetals and their competition with naturallyoccurring elements. The application of advancedmethods such as chemical speciation, metallomics, metabolomics, and implementation ofimaging techniques in the research on fungal physiology will bring a deeper insight intothe physiological and biochemical pathways of xenobiotic and naturallyoccurring elementsin macromycetes.
ACKNOWLEDGEMENTS
This project was funded by KBN (Committee for Scientific Research, Poland) underGrant 6P04G 12928.
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

BIOACCUMULATION AND DISTRIBUTION IN KING OYSTER MUSHROOM 513
REFERENCES
Baeza, A., Guillen, F.J., and Bernedo, J.M. 2005. Soil-fungi transfer coefficient: Importance of thelocation of mycelium in soil and of the differential availability of radionuclides in soil fractions.J. Environ. Radioact. 81, 89–106.
Baeza, A., Guillen, F.J., Salas, A., and Manjon, J.L. 2006. Distribution of radionuclides in differentparts of a mushroom: Influence of the degree of maturity. Sci. Tot. Environ. 359, 255–266.
Baeza, A., Guillen, J., Paniagua, J.M., Hernandez, S., Martın, J.L., Dıez, J., Manjon, J.L., and Moreno,G. 2000. Radiocaesium and radiostrontium uptake by fruit bodies of Pleurotus eryngii viamycelium, soil and aerial absorption. Appl. Radiat. Isot. 53, 455–462.
Bazała, M.A., Bystrzejewska-Piotrowska, G., and Cipakova, A. 2005. Bioaccumulation of 137Cs inwild mushrooms collected in Poland and Slovakia. Nukleonika 50, S15–S19.
Bazała, M.A., Gołda, K., and Bystrzejewska-Piotrowska, G. 2008. Transport of radiocesium inmycelium and its translocation to fruitbodies of a saprophytic macromycete. J. Environ.Radioact. 99, 1200–1202.
Bystrzejewska-Piotrowska, G. and Bazała, M.A. 2008. A study of mechanisms responsible forincorporation of cesium and radiocesium into fruitbodies of king oyster mushroom (Pleurotuseryngii). J. Environ. Radioact. 99, 1185–1191.
Bystrzejewska-Piotrowska, G., Manjon, J.L., Pianka, D., Bazała, M.A., and Urban, P.L. 2005. Cationicinteractions in caesium uptake by king oyster mushroom (Pleurotus eryngii). Nukleonika 50,S9–S13.
Bystrzejewska-Piotrowska, G. and Urban, P.L. 2004. Accumulation and translocation of cesium-137in onion plants (Allium cepa). Environ. Exp. Bot. 51, 3–7.
Bystrzejewska-Piotrowska, G., Urban, P.L., and St↪eborowski, R. 2003. Discrimination between 137Cs
and 40K in the fruiting body of wild edible mushrooms. Nukleonika 48, 155–157.Cipakova, A. 2005. Migration of radiocaesium in individual parts of the environment. Nukleonika
50, S19–S25.Falandysz, J., J
↪edrusiak, A., Lipka, K., Kannan, K., Kawano, M., Gucia, M., Brzostowski, A., and
Dadej, M. 2004. Mercury in wild mushrooms and underlying soil substrate from Koszalin,North-central Poland. Chemosphere 54, 461–466.
Gadd, G.M. (Ed.) 2001. Fungi in Bioremediation. Cambridge, UK, British Mycological Society,Cambridge University Press.
Gadd, G.M. 2007. Geomycology: Biogeochemical transformations of rocks, minerals, metals andradionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 111, 3–49.
Griffin, D.H. 1994. Fungal Physiology. New York, Wiley.Kalac, P. and Svoboda, L. 2000. A review of trace element concentrations in edible mushrooms. Food.
Chem. 69, 273–281.Kalac, P. 2001. A review of edible mushroom radioactivity. Food. Chem. 75, 29–35.Karadeniz, O. and Yaprak, G. 2007. Dynamic equilibrium of radiocesium with stable cesium within
the soil-mushroom system in Turkish pine forest. Environ. Pollut. 148, 316–324.Kruyts, N. and Delvaux, B. 2002. Soil organic horizons as a major source for radiocesium biorecycling
in forest ecosystems. J. Environ. Radioact. 58, 175–190.Kuwahara, C., Fukumoto, A., Ohsone, A., Furuya, N., Shibata, H., Sugiyama, H., and Kato, F. 2005.
Accumulation of radiocesium in wild mushrooms collected from a Japanese forest and cesiumuptake by microorganisms isolated from the mushroom-growing soils. Sci. Tot. Environ. 345,165–173.
Le Lay, P., Isaure, M.P., Sarry, J.E., Kuhn, L., Fayard, B., Le Bail, J.L., Bastien, O., Garin, J., Roby, C.,and Bourguignon, J. 2006. Metabolomic, proteomic and biophysical analyses of Arabidopsisthaliana cells exposed to a caesium stress. Influence of potassium supply. Biochimie 88,1533–1547.
Dow
nloa
ded
by [
Mis
sour
i S &
T]
at 2
2:19
13
Janu
ary
2014

514 G. BYSTRZEJEWSKA-PIOTROWSKA ET AL.
Manjon, J.L., Urban, P.L., and Bystrzejewska-Piotrowska, G. 2004. A simple and quick model tostudy uptake and transfer of radionuclides and heavy metals from mycelium to the fruitbodyof saprophytic edible fungi. Nukleonika 49, S21–S24.
Mietelski, J.W., Jasinska, M., Kubica, B., Kozak, K., and Macharski, P. 1994. Radioactive contami-nation of Polish mushrooms. Sci. Tot. Environ. 157, 217–226.
Moore, D. 1998. Fungal Morphogenesis. New York, Cambridge University Press.Peplow, M. 2006. Counting the dead. Nature 440, 982–983.Pletsch, M., de Araujo, B.S., and Charlwood, B.V. 1999. Novel biotechnological approaches in
environmental remediation research. Biotechnol. Adv. 17, 679–687.Rodrıguez-Navarro, A. 2000. Potassium transport in fungi and plants. Biochim. Biophys. Acta 1469,
1–30.Sanchez, A.L., Parekh, N.R., Dodd, B.A., and Ineson, P. 2000. Microbial component of radiocaesium
retention in highly organic soils. Soil Biol. Biochem. 32, 2091–2094.Singh, H. 2007. Mycoremediation. Fungal Bioremediation. Hoboken, NJ, Wiley.Stamets, P. 2000. Growing Gourmet and Medicinal Mushrooms. Berkeley, CA, Ten Speed.Stamets, P. 2005. Mycelium Running. How Mushrooms Can Help Save the World. Berkeley, CA, Ten
Speed.Svoboda, L., Zimmermannova, K., and Kalac, P. 2000. Concentrations of mercury, cadmium, lead
and copper in fruiting bodies of edible mushrooms in an emission area of a copper smelter anda mercury smelter. Sci. Tot. Environ. 246, 61–67.
Terada, H., Shibata, H., Kato, F., and Sugiyama, H. 1998. Influence of alkali elements on theaccumulation of radiocesium by mushrooms. J. Radioanal. Nucl. Chem. 235, 195–200.
Urban, P.L., Bazała, M.A., Asztemborska, M., Manjon, J.L., Kowalska, J., Bystrzejewska-Piotrowska,G., Pianka, D., St
↪eborowski, R., and Kuthan, R.T. 2005. Preliminary study of platinum
accumulation in the fruitbodies of a model fungal species: King oyster mushroom (Pleurotuseryngii). Nukleonika 50, S63–S67.
Urban, P.Ł. and Bystrzejewska-Piotrowska, G. 2003. Comparative analysis of cesium and potassiumuptake in onion Allium cepa L. Czech. J. Phys. 53, A91–A96.
van Elteren, J.T., Woroniecka, U.D., and Kroon, K.J. 1998. Accumulation and distribution ofselenium and cesium in the cultivated mushroom Agaricus bisporus—A radiotracer-aidedstudy. Chemosphere 36, 1787–1798.
Wauters, J., Vidal, M., Elsen, A., and Cremers, A. 1996. Prediction of solid/liquid distributioncoefficients of radiocaesium in soils and sediments. Part two: A new procedure for solid phasespeciation of radiocaesium. Appl. Geochem. 11, 595–599.
Yoshida, S. and Muramatsu, Y. 1998. Concentrations of alkali and alkaline earth elements inmushrooms and plants collected in a Japanese pine forest, and their relationship with 137Cs. J.Environ. Radioact. 41, 183–205.D
ownl
oade
d by
[M
isso
uri S
& T
] at
22:
19 1
3 Ja
nuar
y 20
14