Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 22, Issue of June 3, pp. 15512-15519, 1994 Printed in U.S.A.
Binding of Sugar Ligands to Ca2+-dependent Animal Lectins 11. GENERATION OF HIGH-AFFINITY GALACTOSE BINDING BY SITE-DIRECTED MUTAGENESIS*
(Received for publication, February 8, 1994, and in revised form, March 31, 1994)
Susanne T. IobstS and Kurt DrickamerO From the Department of Biochemistry and Molecular Biophysics, Columbia University, New York, New York 10032
Changes have been introduced into the Ca2+-depend- ent carbohydrate-recognition domain (CRD) of rat se- rum mannose-binding protein by site-directed mutagen- esis to model the binding sites of homologous galactose- binding CRDs. Binding assays reveal that galactose- binding activity nearly identical to that of the CRD from the asialoglycoprotein receptor can be introduced into the mannose-binding site by 3 single amino acid changes and insertion of a segment of 6 amino acids. Separate changes are required to establish high-affinity binding to galactose and create high selectivity by exclusion of mannose from the binding site. The mutagenesis studies and NMR analysis of sugar-CRD titrations demonstrate that an important component of high-affinity galactose binding is interaction between the B face of the sugar and tryptophan. The binding properties of the C-type CRD from the cartilage proteoglycan, aggrecan, can also be modeled based on the mannose-binding CRD frame- work. This lower affinity binding site involves stacking of a phenylalanine residue against the sugar ligand.
The Ca2+-dependent (C-type) animal lectins bind a variety of different sugar residues through homologous 115-130 amino acid carbohydrate-recognition domains (CRDs)‘ (1). Based on the crystal structure of the C-type CRD from rat serum man- nose-binding protein (2), it seems likely that all the CRDs have similar overall three-dimensional folds, since the 32 conserved amino acid residues characteristic of all of these CRDs form two disulfide bonds, Ca2+-binding sites, and the hydrophobic core which together determine the structure of the CRD. This sug- gestion is supported by the recent determination of the struc- ture of the C-type CRD in human E-selectin (3), which re- sembles closely the CRD from mannose-binding protein in overall fold.
In spite of their overall similarity in structure, the C-type CRDs have distinct sugar binding characteristics. The man- nose-binding protein and other collectins (C-type lectins in which CRDs are juxtaposed with collagenous domains) gener- ally bind ligands with equatorial 3 and 4 hydroxyl groups such as mannose and N-acetylglucosamine, as well as fucose, which contains equatorial 2 and 3 hydroxyl groups than can be su-
* This work was supported in part by Grant GM42628 from the Na- tional Institutes of Health and Fogarty Senior International Fellowship TW01925. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisenent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Supported by Unilever Research US Inc. 5 Recipient of a Faculty Salary Award from the American Cancer
Society. To whom correspondence should be addressed: Dept. of Bio- chemistry and Molecular Biophysics, Columbia University, 630 West 168th St., New York, NY 10032. Tel.: 212-305-6453; Fax: 212-305-4131.
The abbreviations used are: CRD, carbohydrate-recognition domain; BSA, bovine serum albumin. All sugars are in the D configuration except L-fucose.
perimposed on the 3 and 4 hydroxyl groups of mannose (4). However, the relative affinities of the different collectins for these sugars varies widely (5, 6). In contrast, the asialoglyco- protein receptor and related lectins preferentially bind galac- tose and N-acetylgalactosamine (7). The mechanism of sugar binding by these lectins has not been elucidated, but sequence comparisons reveal that all of the mammalian lectins that bind galactose with high-affinity contain a few common differences from the mannose-binding sequences. Introduction of the changes G~U’’~ + Gln and + Asp into the mannose- binding sequence allows preferential binding of the modified domain to galactose, although the absolute affinity appears to be weak (8). Studies with a variety of sugar analogs indicate that the 3 and 4 hydroxyl groups of galactose and N-acetyl- galactosamine are the major aspects of the sugar ligand recog- nized by this subclass of C-type CRDs (9).
Additional mutations have now been introduced into the mannose-binding CRD to generate a binding site with binding characteristics essentially identical to those of natural galac- tose-binding proteins. Distinct changes lead to high-affinity binding of galactose and high selectivity in exclusion of man- nose. The importance of interaction between galactose and a tryptophan in the galactose-binding CRDs is demonstrated.
EXPERIMENTAL PROCEDURES Materials-Gal-BSAwas obtained from E-Y Laboratories. Galactose-
Sepharose was prepared by the divinyl sulphone method (10). Sources for other reagents were as indicated in the accompanying manuscript (11).
Mutagenesis Procedures-Synthetic oligonucleotides and the restric- tion sites at which they were inserted are shown in Fig. 1. Procedures for construction of modified expression vectors have been described (11).
Protein Production-Mutant versions of the mannose-binding CRD were prepared as described in the accompanying manuscript (11), ex- cept that galactose-Sepharose was substituted for mannose-Sepharose. The CRD from the major subunit of the rat liver asialoglycoprotein receptor was prepared using a bacterial expression vector containing the T7 promotor.’ The CRD from aggrecan was prepared by published procedures (12).
formed as described (11, 131, using ‘251-Gal,,-BSA as test ligand. Each Binding Assays-Direct binding and competition assays were per-
assay was performed in duplicate. Values reported are the average * S.D. for three separate assays. NMR procedures were exactly as for mannose-binding mutants (11).
RESULTS AND DISCUSSION Sequence Comparisons-The sequences of several mannose-
and galactose-binding CRDs in the region of the domains be- lieved to be involved in contacting sugar are compared in Fig. 2. It has been previously noted that 3 of the 5 ligands for Ca2+ number 2 in the mannose-binding CRD are conserved, but PO- sitions 185 and 187 are invariably Glu and Asn in CRDs that preferentially bind mannose, and are changed to Gln and h p in CRDs that preferentially bind galactose (8). Incorporation of
K. Drickamer, unpublished observations.
15512

Galactose Binding to C-type Animal Lectins 15513
A W T A T I O N S I N QPD BACKGROUND
A M I S e r J A I B s t X I 5-CAGCMCTGGAAAAAGGATCAGCCCGATGACCATGGCTCT 3-GTCGTTGACCTTTTTCCTAGTCGGGCTACTGGTACCGAGA
SerAsnTrplysLysAspGl~r~s~spHisGl~er 180 190
- Bbs I AccI GGGGAAGACTGTGTCACTATAGTAGACAAC-3 CCCCTTCTGACACAGTGATATCATCTGTTG-5
GlyGluAspCysVa~ThrIleValAspAsn
-
200
T rP 5-GATCAGCCCGATGACTGGGGCT-3
3-TCGGGCTACTGACC-5
TrpTyrGLyHisGLyLeuGlyCly 5-GATCAGCCCGATGACTGGTACGGCCATGGCCTAGGCGGTGGTGAAGACTG-3
3-TCGGGCTACTGACCATGCCGGTACCGGATCCGCCACCACTTCTGACACAG-5
B WTATIOWS I N QPDUG BACKGROUND
A M I S e l J A I R s a I N c o I A v r I I B b S I AccI 5-CAGCMCTGGAAAAAGCATCAGCCCGATGACTGGTACGGCCATGGCCTAGGCGGTGGT~GACTGTGTCACTATAGTAGACMC-3 3-GTCGTTGACCTTTTTCCTACTCGGGCTACTGACCATGCCGGTACCGGATCCGCCACCACTTCTGACACAGTGATATCATCTGTTG-5
-
SerAsnTrplysLysAspGl~r~spAspTrpTyrGlyHisGlyLeuGlyCLyClyGluAAspCysValThrI LeValAspAsn 180 190 A B C D E 200
A l a 5-ACGGCGCGGGC-3 3-TGCCGCGCCCGGATC-5
T Y r 5-ACGGCTATGGC-3 3-TGCCGATACCGGATC-5
G l U 5-ACGGCGAAGGC-3 3-TGCCGCTTCCGGATC-5
G l n 5-ACGGCCAAGGC-3 3-TGCCGGTTCCGGATC-5
G l n 5-CTGGMAAAGGATCAGCCCGACGACTGGCAAGGC-3
3-GTTGACCTTTTTCCTAGTCGGGCTGCTGACCGTTCCGGTAC-5
A l a 5-CTGGAAAAAGGATCAGCCCGACGACTGGGCTGGC-3
3-GTTGACCTTTTTCCTAGTCGGGCTGCTGACCCGACCGGTAC-5
H i s G L y L e u G l y G l y G l u A s p 5-CATGGCCTAGGCGGT GAAGACTG-3
3-CGGATCCGCCA CTTCTGACACAG-5 FIG. 1. Oligonucleotides used for site-directed mutagenesis. The sequences of oligonucleotide pairs are indicated, along with the restriction
sites at which these fragments were inserted.
the double change GlulS5 ”-* Gln, A d s 7 ”-* Asp into the man- nose-binding CRD establishes weak preferential binding for galactose.
The low affinity of the Glu185 + Gln, Am”’ + Asp mutant for galactose indicates that other residues must contribute to ga- lactose binding by the asialoglycoprotein receptor and other natural galactose-binding C-type animal lectins. Examination of the sequences in Fig. 2 suggests other consistent changes that might be associated with establishment of a high-affinity galactose-binding site. Given their proximity to the mannose- binding site, certain of these changes were selected as candi- dates for incorporation into the mannose-binding CRD. Changes that have been made are summarized in Fig. 3.
Creation of a High-affinity Galactose-selective Binding Site- The mammalian asialoglycoprotein receptors were used as the primary model for a relatively high-affinity galactose-binding site. Following the double change GlulB5 + Gln, + Asp to make mutant QPD, two additional changes were added to cre-
ate a higher affinity galactose-binding site. First, incorporation of the change His’’’ + Trp (mutant QPDW) was suggested by the presence of tryptophan at this position in all of the high affinity, galactose-binding CRDs shown in Fig. 2. Direct bind- ing assays with 1251-Gal,,-BSA reveal that the absolute affinity of this mutant CRD is comparable to the affinity of the major subunit of the rat liver asialoglycoprotein receptor (Table I), while the affinity of the QPD mutant was too weak to be meas- ured in this assay. However, competition assays with monosac- charides show that selectivity of the QPDW mutant for galac- tose over mannose is not comparable to the natural galactose- selective CRD, and is not significantly different from the QPD mutant.
High selectivity for galactose, approaching that of the asia- loglycoprotein receptor, was obtained by incorporation of an additional glycine-rich sequence found in the galactose-selec- tive CRDs. This glycine-rich stretch represents an insertion of 5 amino acid residues relative to the mannose-binding CRD.

15514
MOT1 F C8 #2 LIGANDS
MANNOSE RESIDUES:
r A (Rat ) MANNOSE I A (Mouse) BINDING I (Human) PROTEIN I C (Rat )
L C (House) ~ C R ~ H A G E MANNOSE RECEPTOR CRD-4 (Hunen)
PLACENTAL MANNOSE RECEPTOR (Hunan)
GALACTOSE RESIDUES 1:
~ C R ~ G A G E r (Rat ) GAL RECEPTOR L (Mouse)
r 1 (Rat ) ASIALO 1 1 (Mouse) GLYCO- 1 1 (Human) PROTEIN 1 2 (Ret ) RECEPTOR I 2 (Mouse)
L 2 (Human)
GALACTOSE RESIDUES 2:
r (Human) AGGRECAN I ( R e t )
Galactose Binding to C-type Animal Leetins
#we o n G n u ZP E O C ~ ~ GUND c n c
1 60 1 70 180 190 200 210 220 1 1 1 i t
E N H
FLGITDEVTEGPFMYVTGGR---LTYSNUKKDEPNDHGS-----GEDCVTIV--DNGLUNDISCQASHTAVCEFPA FLGITDEAfEGPFMYVTGGR---LTYSNUKKDEPNNHGS-----GEDCVIIL--DNGLUNDISCQASFKAVCEFPA FLGITDEKTEGPF#LTGNR---LTYTNUNEGEPNNAGS-----DEDCVLLL--KN~VWD~CSTSHLAVCEFPI FLGITDQRTENVFEDLTGNR---VRYTNUNEGEPNNVGS-----GENCWLK--TNGI(YWDVPCSDSFLWCEFSD YLGiTDVRVEGSFEDLTGNR---VRYTNVWDGEPNNTGD-----GEDCWIL--GNGI<UWDVPCSDSFLAICEFSD
UIGLTYGSPSEGFTUSDGSP---VSYENUAYGEPNNY~N-----VEYCGELK~PTHSUNDINCEHLNNUIC~1
UWGLSDLNQEGTWPWVDGSPLLPSFKQYUNRGEPNNVGE------ EDCAEFS---GNGUNDDKCN-LAWICKKSAA
Q D UYGHGLGG
UIGLTDQ--NGPWRWVDGTD-YEKGFTHUAPKQ~NUYGHGLGGGEDCAHFT--SDGRUWDDVCQRPYRWVCELAKDS UIGLTOQ--NGPURUVDGTD-FEKGFKNUAPLQPDNUFGHGLGGGEDCAHIT--TGGPUWDDVCQRTFRWlCEMKLAKES UfGLTDQ--NGPUI<WMGTD-YETGFKN~PGQPDDUYGHGLGGGEDCAHFT--TDGH~DDVCRRPYRWCETELGICI\N U1GLTDQ--NGPUKWVDGTD-YETGFQNURPEQPDNUYCHGLGGGEDCAHFT--TDGRUNDDVCRRPYRVVCETKLDKAN UWGLHDQ--NGPUKWVOGTD-YETGFKNW(PEQPDDWTGHGLGGGEDCAHFT--DDGRUNDDVCQRPYRUVCETELDKAS- UIGLTDK--DGSUKWVDGTE-YRSNFKNUAFTQPDNUPGHEEGGSEDCAEIL--SDGLUNDNFCQQVNRUACERKRDITY UIGLTDR--DGSUKWVDGTD-YRSNYRN~FTQPDN~GHEQGGGEDCAEIL--SDGHUNDNFCQQVNRWVCEKRRNITH UIGLTDS--DGSUKWVDGTD-YRHNYKNUAVTQPDFIUHGHELGGS~CVEVQ--PDGRUWDDFCLQWRUVCKRRNATGE-
Q D F
UfGLNDRTIEGDFRWSDGHP--”QFENURPNQPDNFFAA----GEDCWHIUHEKGEUNDVPCNYHLPFTCKKGTAT- UfGLNDRTIEGDFRWSDGHS---LQFEKURPNQPDNFFAT----GEDCWHIUHERGEUNDVPCNY~LPFTCKKGlVA-
(Chicken) UIGLSDRAVENDFRUSDGHS---LQFENURPNQPDNFFFA----GEDCWHIUHE~GEUNDVPCNYHLPFTCKKGTVA- VERSICAN (Human) UIGLNDKHFEHDFRUTDGST---LQYENURPNQPDSFFSA----GEDCWIIUHENGQUNDVPCNYHLTYTCKKGTVA- NEUROCAN ( R e t 1 UIGLNDRTVERDFQUTDNTG---LQYEN~EKQPDNFFAG----GEDCWHVAHENGRUNDVPCNYNLPYVCKKGTVL-
FIG. 2. Comparison of sequences near binding-site residues in mannose- and galactose-binding CRDs. Motif, residues characteristic of all C-type CRDs indicated with one-letter amino acid code and cb, aromatic residues; 8, aliphatic residues; a, aromatic or aliphatic residues; 0, amino acid residues with oxygen-con~ining side chains. Ligands for Ca2+ number 2 are indicated with arrows. Residue numbers are for rat m~nose-binding protein A. ~ u n n o s e residues, residues in rat mannose-binding p r o t e ~ Ashown by mu~genesis to be involved in mannose binding (4). Other ma~ose-hinding CRDs are shown for comparison. Only the single CRD of the macrophage mannose receptor known to display high amnity binding to mannose is shown. Galactose residues 1 and 2, residues common to high- and low-affinity galactose-binding proteins, respec- tively. References for sequences are available in Refs. 14-19.
RAT MANNOSE-BINDING PROTEIN A: ( 181) UKKDEPNDHGS”---GEDCVTIV--DNGLUNDISC f 209)
high-affinity galactose-binding activ- MUTANT QpD: ity into the mannose-binding CRD. wTANT QpDu: Seauence of rat man~ose-bind in^ Drotein
FIG. 3. Changes made to introduce Q D 1 1
Q D U , i
A i i shown at the top. Changesmade to ~~UTANT ppDm: Q D UYGHGLGG mimic the asialog~ycoprotein receptor are shown in the rnidae, while changes made RAT ASIALOGLYC~ROTEIN RECEPTOR: ( 23s) ~PGPPDD~GHGLGGGED~HFT--TDGHUNDDVC ( 254) to resemble the cartilage proteoglycan, aggrecan, are shown at the bottom. Num- bers indicate positions, in the intact pro- ANT Q ~ Q F : teins, of the first and last residues shown.
1 Q D F
1 MUTANT QPDFG: Q D FYGHGLGG
HUHAN AGGRECAN: (2248) URPNQPDNFFAA- ---GEDCWMIUHEKGEUNDVPC (2263)
The resulting mutant, designated QPDWG, shows no further closely resemble the asialoglycoprotein receptor from perito- increase in absolute affinity for neoglycop~tein ligand, but neal macrophages, which binds galactose and N-acetylgalac- now binds galactose with the same degree of selectivity as the tosamine with roughly equal affinities (20). asia~oglycoprotein receptor CRD. In spite of the strong binding ~ ~ c t o s e - s e ~ e ~ t i v e Binding to ~ ~ o ~ o g l y c u ~ CRDs-The to galactose, the still higher affinity of the hepatic asialoglyco- QPDW mutant has somewhat the same binding characteristics protein receptor for N-acetylgalactosamine (9) is not observed. as the CRD found in the cartilage proteoglycan, aggrecan (12). In this respect, the properties of the QFDWG mutant most Comparison of the sequences of various proteoglycan CRDS
Galactose Binding to C-type Animal Lectins 15515
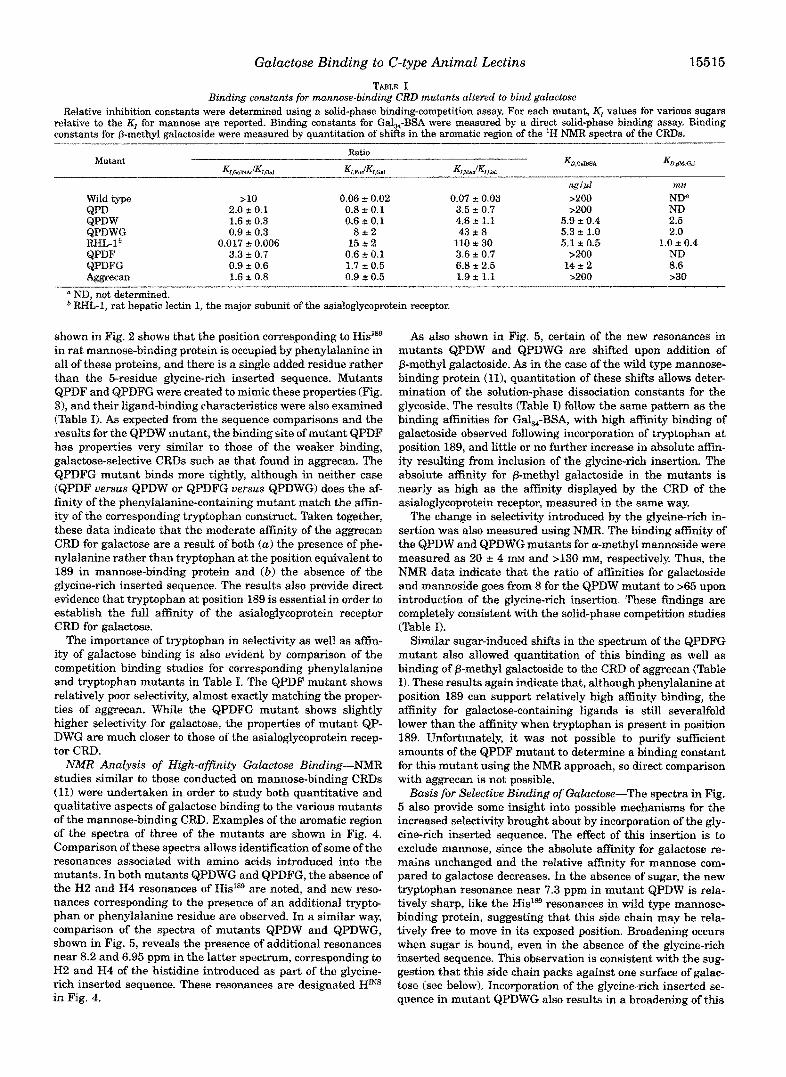
Binding constants for mannose-binding CRD mutants altered to bind galactose TABLE I
relative to the KI for mannose are reported. Binding constants for Gal,-BSA were measured by a direct solid-phase binding assay. Binding Relative inhibition constants were determined using a sofid-phase binding-competition assay. For each mutant, Kl values for various sugars
constants for @-methyl galactoside were measured by quantitation of shiRs in the aromatic region of the 'H NMR spectra of the CRDs.
Mutant
Wild type QPD QPDW QPDWG RHL-1* QPDF QPDFG & ? m a n
Ratio
K ~ , ~ l ~ ~ K ~ , G ~ l K ~ , ~ " ~ K ~ * ~ ~ K ~ , ~ ~ ~ f K ~ , ~
>10 0.06 t 0.02 0.07 t 0.03
"
2.0 t 0.1 0.8 t 0.1 3.5 2 0.7 1.6 t 0.3 0.6 t 0.1 4.6 2 1.1 0.9 t 0.3
0.017 t 0.006 8 2 2
15 t 2 110 * 30 3.3 ~t 0.7 0.6 t 0.1 3.6 t 0.7 0.9 t 0.6 1.7 t 0.5 6.8 2 2.5 1.6 t 0.8 0.9 t 0.5 1.9 f 1.1
43 t 8
Kl2,GdBSA
>200 >200
5.9 t 0.4 5.3 f 1.0 5.1 t 0.5
>200 1 4 t 2 >200
mbf ND" ND 2.5 2.0
1.0 f 0.4 ND 8.6 >30
a ND, not determined. RHL-1, rat hepatic lectin I, the major subunit of the asialog~ycoprotein receptor.
shown in Fig. 2 shows that the position co~esponding to His'ss in rat mannose-binding protein is occupied by phenyla~anine in all of these proteins, and there is a single added residue rather than the 5-residue g~ycine-rich inserted sequence. Mutants QPDF and QPDFG were created to mimic these properties (Fig. 3), and their ligand-binding characteristics were also examined (Table I). As expected from the sequence comparisons and the results for the QPDW mutant, the binding site of mutant QPDF has properties very similar to those of the weaker binding, galactose-selective CRDs such as that found in aggrecan. The QPDFG mutant binds more tightly, although in neither case (QPDF versus QPDW or QPDFG versus QPDWG) does the af- finity of the phenylalanine-containing mutant match the affin- ity of the corresponding t r ~ t o p h a n construct. Taken together, these data indicate that the moderate affinity of the aggrecan CRD for galactose are a result of both (a ) the presence of phe- nylalanine rather than tryptophan at the position equivalent to 189 in mannose-binding protein and ( b ) the absence of the glycine-rich inserted sequence. The results also provide direct evidence that try~tophan at position 189 is essential in order to establish the full asni ty of the asialoglycoprotein receptor CRD for galactose.
The importance of t ~ t o p h ~ in selectivity as well as affin- ity of galactose binding i s also evident by comparison of the competition binding studies for corresponding phenylalanine and tryptophan mutants in Table I. The QPDF mutant shows relatively poor selectivity, almost exactly matching the proper- ties of aggrecan. While the QPDFG mutant shows slightly higher selectivity for galactose, the properties of mutant QP- DWG are much closer to those of the asialoglycoprotein recep- tor CRD.
NMR Analysis of ~ l g ~ - a f ~ n i t y Galactose Binding-NMR studies similar to those conducted on mannose-binding CRDs (11) were undertaken in order to study both quantitative and qualitative aspects of galactose binding to the various mutants of the mannose-binding CRD. Examples of the aromatic region of the spectra of three of the mutants are shown in Fig. 4. Comparison of these spectra allows identification of some of the resonances associated with amino acids introduced into the mutants. In both mutants QPDWG and QPDFG, the absence of the H2 and H4 resonances of Hislag are noted, and new reso- nances co~esponding to the presence of an additional trypto- phan or phenylalanine residue are observed. In a similar way, comparison of the spectra of mutants QPDW and QPDWG, shown in Fig. 5, reveals the presence of additional resonances near 8.2 and 6.95 ppm in the latter spectrum, corresponding to H2 and H4 of the histidine introduced as part of the glycine- rich inserted sequence. These resonances are designated HmS in Fig. 4.
As also shown in Fig. 5, certain of the new resonances in mutants QPDW and QPDWG are shifted upon addition of @-methyl galactoside. As in the case of the wild type mannose- binding protein (11), ~ u ~ t i t a t i o n of these shifts allows deter- mination of the solution-phase dissociation constants for the glycoside. The results (Table I) follow the same pattern as the binding affinities for Gal,,-BSA, with high a n i t y binding of galactoside observed following incorporation of tryptophan at position 189, and little or no further increase in absolute affin- ity resulting from inclusion o f the glycine-rich insertion. The absolute affinity for @-methyl galactoside in the mutants is nearly as high as the affinity displayed by the CRD of the asialoglycoprotein receptor, measured in the same way.
The change in selectivity introduced by the glycine-rich in- sertion was also measured using NMR. The binding affinity of the QPDW and QPDWG mutants for a-methyl mannoside were measured as 20 r 4 mM and >130 mM, respectively. Thus, the NMR data indicate that the ratio of asnities for galactoside and mannoside goes from 8 for the QPDW mutant to >65 upon introduction of the glycine-rich insertion. These findings are completeIy ~ n s i s t e n t with the so~id-phase competition studies (Table I).
Similar sugar-induced shifts in the spectrum of the QPDFG m u ~ n t also allowed quantitation of this binding as well as binding of @-methyl galactoside to the CRD of aggrecan (Table I). These results again indicate that, although phenylalanine at position 189 can support relatively high affinity binding, the affinity for galactose-containing ligands is still severalfold lower than the affinity when tryptophan is present in position 189. Unfortunately, it was not possible to purify sufficient amounts of the QPDF mutant to determine a binding constant for this mutant using the NMR approach, so direct comparison with aggrecan is not possible.
Basis for Selective Binding of Galactose-The spectra in Fig. 5 also provide some insight into possible mechanisms for the increased selectivity brought about by incorporation of the gly- cine-rich inserted sequence. The effect of this insertion is to exclude mannose, since the absolute aflEinity for galactose re- mains unchanged and the relative affinity for mannose com- pared to galactose decreases. In the absence of sugar, the new t r ~ t o p h a n resonance near 7.3 pprn in mutant QPDW is rela- tively sharp, like the Hislag resonances in wild type mannose- binding protein, suggesting that this side chain may be rela- tively free to move in its exposed position. Broadening occurs when sugar is bound, even in the absence of the glycine-rich inserted sequence. This observation is consistent with the sug- gestion that this side chain packs against one surface of galac- tose (see below). Incorporation of the glycine-rich inserted se- quence in mutant QPDWG also results in a broadening of this
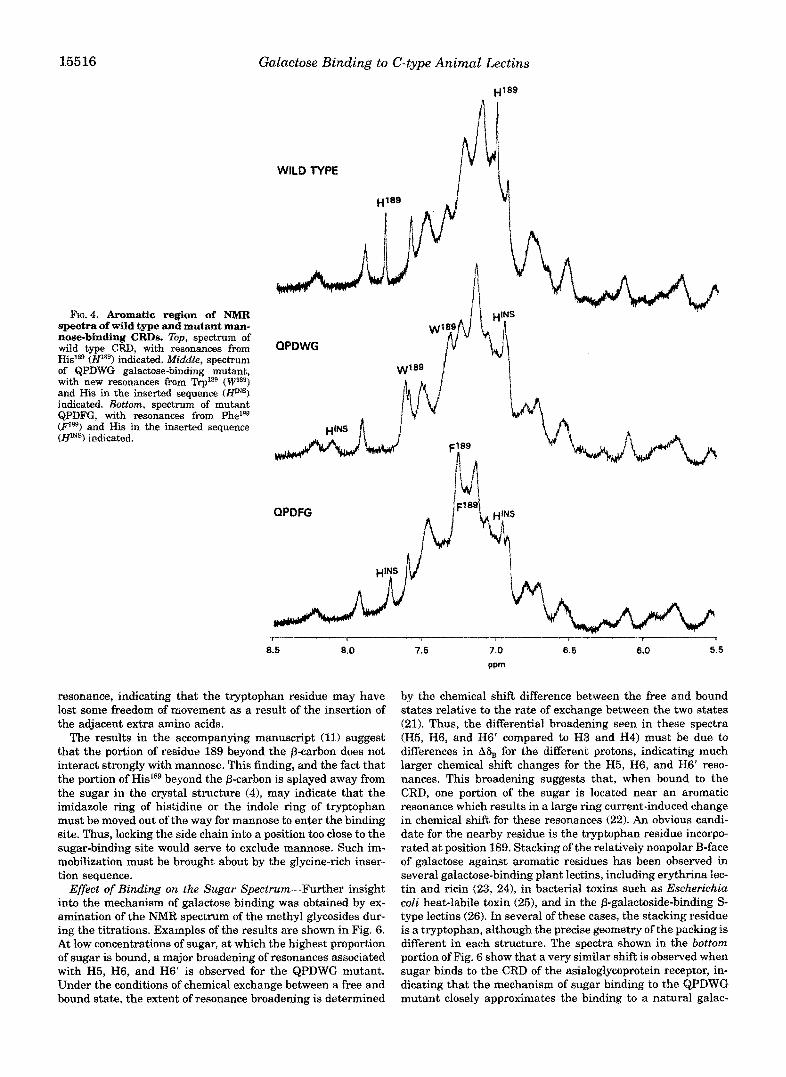
15516 Galactose Binding to C-type Animal Lectins
FIG.^. Aromatic region of NMR spectra of wild type and mutant man- nose-binding CRDs. Zbp, spectrum of
His18e (H1*@) indicated, Middle, spectrum wild type CRD, with resonances from
of QPDWG galactose-binding mutant, with new resonances from Trp1*9 (Wla9) and His in the inserted sequence (WINS) indicated. Bottom, spectrum of mutant QPDFG, with resonances from Phelss (Ps9) and His in the inserted sequence (Hms) indicated.
WILD TYPE
QPDWG
QPDFG
1 i I
8.5 8.0 7.5 7.0 6.5 6.0 5.5
ppm
resonance, indicating that the tryptophan residue may have lost some freedom of movement as a result of the insertion of the adjacent extra amino acids.
The results in the accompanying manuscript (11) suggest that the portion of residue 189 beyond the p-carbon does not interact strongly with mannose. This finding, and the fact that the portion of His'39 beyond the p-carbon is splayed away from the sugar in the crystal structure (41, may indicate that the imidazole ring of histidine or the indole ring of tryptophan must be moved out of the way for mannose to enter the binding site. Thus, locking the side chain into a position too close to the sugar-binding site would serve to exclude mannose. Such im- mobilization must be brought about by the glycine-rich inser- tion sequence.
Egect of Binding on the Sugar Spectru~-Further insight into the mechanism of galactose binding was obtained by ex- amination of the NMR spectrum of the methyl glycosides dur- ing the titrations. Examples of the results are shown in Fig. 6. At low concentrations of sugar, at which the highest proportion of sugar is bound, a major broadening of resonances associated with H5, H6, and H6' is observed for the QPDWG mutant. Under the conditions of chemical exchange between a free and bound state, the extent of resonance broadening is determined
by the chemical shift difference between the free and bound states relative to the rate of exchange between the two states (21). Thus, the differential broadening seen in these spectra (H5, H6, and H6' compared to H3 and H4) must be due to differences in Ah for the different protons, indicating much larger chemical shift changes for the H5, H6, and H6' reso- nances. This broadening suggests that, when bound to the CRD, one portion of the sugar is located near an aromatic resonance which results in a large ring current-induced change in chemical shift for these resonances (22). An obvious candi- date for the nearby residue is the tryptophan residue incorpo- rated at position 189. Stacking of the relatively nonpolar B-face of galactose against aromatic residues has been observed in several galactose-binding plant lectins, including erythrina lec- tin and ricin (23, 241, in bacterial toxins such as ~ s c ~ e r i c ~ i a coli heat-labile toxin (251, and in the ~-galactoside-binding S- type lectins (26). In several o f these cases, the stacking residue i s a tryptophan, although the precise geometry of the packing is different in each structure. The spectra shown in the bottom portion of Fig. 6 show that a very similar shift is observed when sugar binds to the CRD of the asialoglycoprotein receptor, in- dicating that the mechanism of sugar binding to the QPDWG mutant closely approximates the binding to a natural galac-
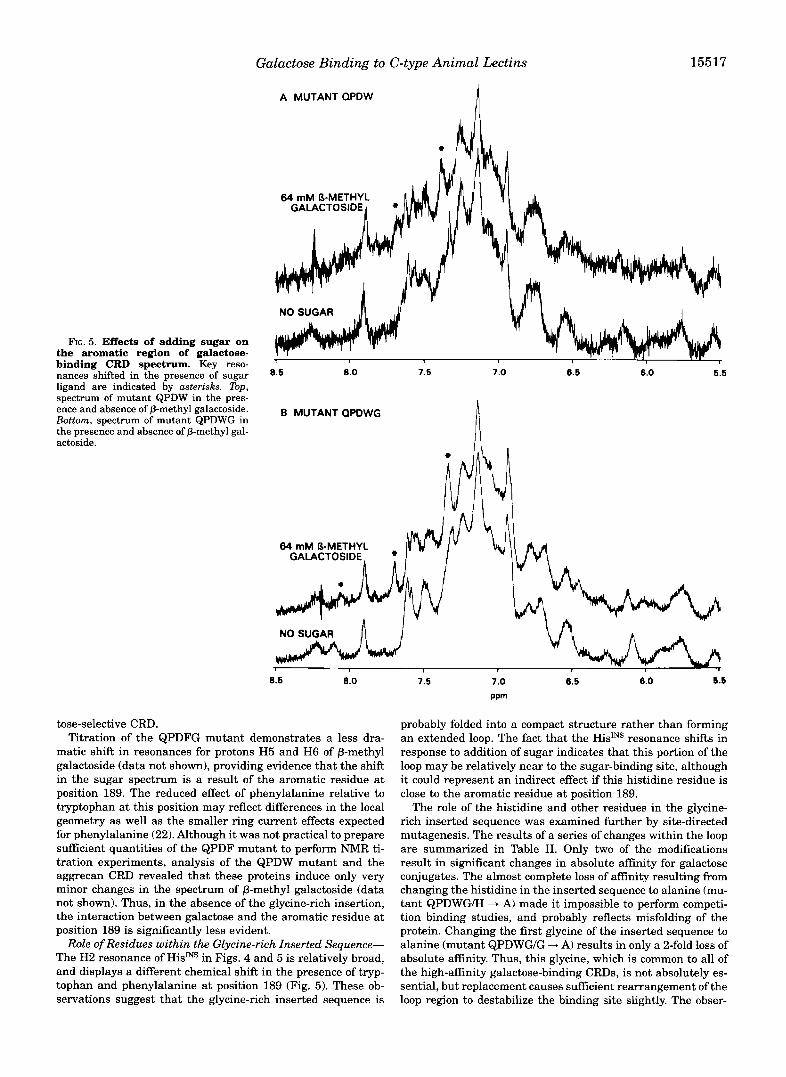
Galactose Binding to C-type Animal Lectins
A MUTANT QPDW n 15517
GALACTOSIDE
FIG. 5. Effects of adding sugar on
binding CRD spectrum. Key reso- the aromatic region of galactose- . .
nances shifted in the presence of sugar ligand are indicated by asterisks. Top, spectrum of mutant QPDW in the pres- ence and absence of P-methyl galactoside. Bottom, spectrum of mutant QPDWG in the presence and absence of P-methyl gal- actoside.
1 I
8.0 7.5 7.0 6.5 6.0 5.5
MUTANT QPDWG
8.6 8.0
tose-selective CRD. Titration of the QPDFG mutant demonstrates a less dra-
matic shift in resonances for protons H5 and H6 of P-methyl galactoside (data not shown), providing evidence that the shift in the sugar spectrum is a result of the aromatic residue at position 189. The reduced effect of phenylalanine relative to tryptophan at this position may reflect differences in the local geometry as well as the smaller ring current effects expected for phenylalanine (22). Although it was not practical to prepare s a c i e n t quantities of the QPDF mutant to perform NMR ti- tration experiments, analysis of the QPDW mutant and the aggrecan CRD revealed that these proteins induce only very minor changes in the spectrum of P-methyl galactoside (data not shown). Thus, in the absence of the glycine-rich insertion, the interaction between galactose and the aromatic residue at position 189 is significantly less evident.
Role of Residues within the Glycine-rich Inserted Sequence- The H2 resonance of HisJNS in Figs. 4 and 5 is relatively broad, and displays a different chemical shift in the presence of tryp- tophan and phenylalanine at position 189 (Fig. 5 ) . These ob- servations suggest that the glycine-rich inserted sequence is
7.5 7.0 6.5 8.0 6.6
ppm
probably folded into a compact structure rather than forming an extended loop. The fact that the HidNS resonance shifts in response to addition of sugar indicates that this portion of the loop may be relatively near to the sugar-binding site, although it could represent an indirect effect if this histidine residue is close to the aromatic residue at position 189.
The role of the histidine and other residues in the glycine- rich inserted sequence was examined further by site-directed mutagenesis. The results of a series of changes within the loop are summarized in Table 11. Only two of the modifications result in significant changes in absolute affinity for galactose conjugates. The almost complete loss of affinity resulting from changing the histidine in the inserted sequence to alanine (mu- tant QPDWG/H + A) made it impossible to perform competi- tion binding studies, and probably reflects misfolding of the protein. Changing the first glycine of the inserted sequence to alanine (mutant QPDWG/G + A) results in only a 2-fold loss of absolute affinity. Thus, this glycine, which is common to all of the high-affinity galactose-binding CRDs, is not absolutely es- sential, but replacement causes sufficient rearrangement of the loop region to destabilize the binding site slightly. The obser-
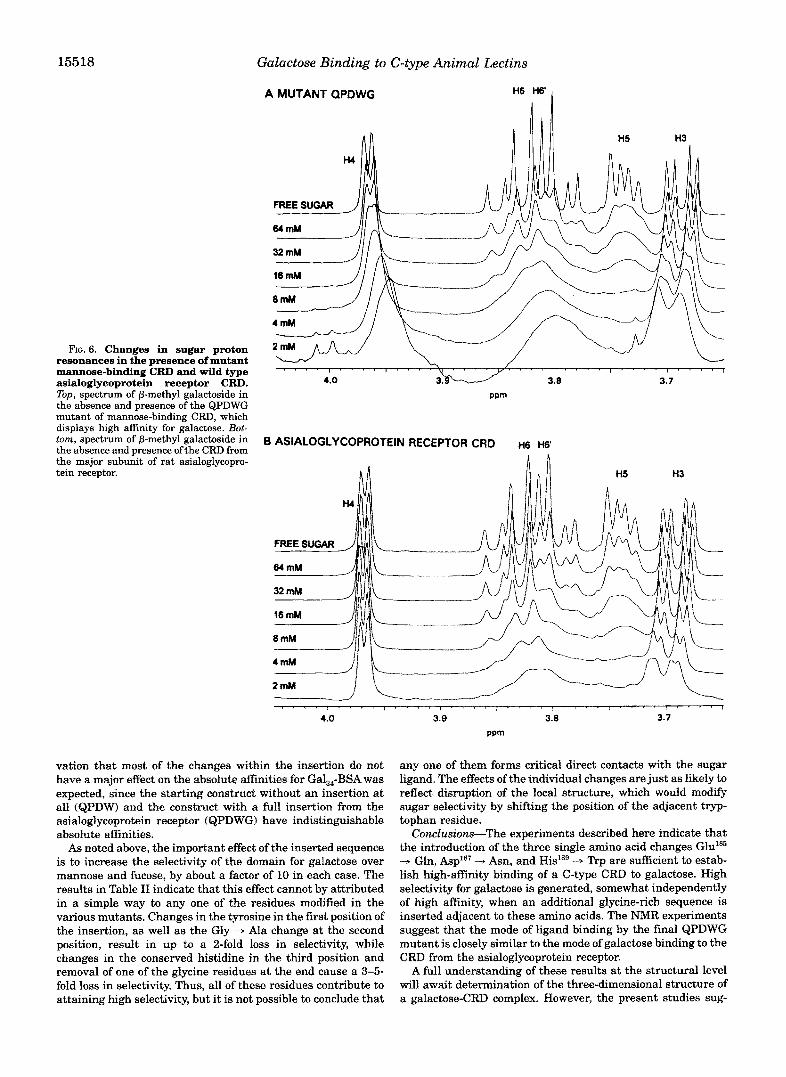
15518 Galactose Binding to C-type Animal Lectins
A MUTANT QPDWG H6 H6‘ 1
FIG. 6. Changes in sugar proton resonances in the presence of mutant mannose-binding CRD and wild type asialoglycoprotein receptor CRD. Top, spectrum of P-methyl galactoside in the absence and presence of the QPDWG mutant of mannose-binding CRD, which displays high affinity for galactose. Bot- tom, spectrum of P-methyl galactoside in the absence and presence of the CRD from the major subunit of rat asialoglycopro- tein receptor.
8 ASIALOGLYCOPROTEIN RECEPTOR CRD n6 ~ 6 ‘
4.0
vation that most of the changes within the insertion do not have a major effect on the absolute affinities for Gal,4-BSA was expected, since the starting construct without an insertion at all (QPDW) and the construct with a full insertion from the asialoglycoprotein receptor (QPDWG) have indistinguishable absolute affinities.
As noted above, the important effect of the inserted sequence is to increase the selectivity of the domain for galactose over mannose and fucose, by about a factor of 10 in each case. The results in Table I1 indicate that this effect cannot by attributed in a simple way to any one of the residues modified in the various mutants. Changes in the tyrosine in the first position of the insertion, as well as the Gly - Ala change at the second position, result in up to a 2-fold loss in selectivity, while changes in the conserved histidine in the third position and removal of one of the glycine residues at the end cause a 3-5- fold loss in selectivity. Thus, all of these residues contribute to attaining high selectivity, but it is not possible to conclude that
3.9 3.8 3.7 ppm
any one of them forms critical direct contacts with the sugar ligand. The effects of the individual changes are just as likely to reflect disruption of the local structure, which would modify sugar selectivity by shiRing the position of the adjacent tryp- tophan residue.
Conclusions-The experiments described here indicate that the introduction of the three single amino acid changes Gluls5 -+ Gln, Asp’87 ”+ Asn, and His’89 -3 Trp are sufficient to estab- lish high-aEnity binding of a C-type CRD to galactose. High selectivity for galactose is generated, somewhat independently of high affinity, when an additional glycine-rich sequence is inserted adjacent to these amino acids. The NMR experiments suggest that the mode of ligand binding by the final QPDWG mutant is closely similar to the mode of galactose binding to the CRD from the asialoglycoprotein receptor.
A full understanding of these results at the structural level will await determination of the three-dimensional structure of a galactose-CRD complex. However, the present studies sug-

Galactose Binding to C-type Animal Lectins 15519 TABLE I1
Binding constants for galactose-binding CRD mutants in glycine-rich inserted sequence All values were measured using solid-phase binding (K,,,) and binding competition (K,) assays. For each mutant, KI values for various sugars
relative to the KI for mannose are reported. Ratio
Mutant Sequence KD,,,,,,, K I , O d N A ~ K I , G . d K I F * e l K f , G d K l , M a ” 4 G s l
ngial QPDWG WYGHGLGGG 0.9 t 0.3 8 t 2 4338 5.3 t 1.0
WAGHGLGGG 2.0 e 0.4 4.9 t 0.6 29 -c 8 5.1 t 1.5 QPDWGN-Q WQGHGLGGG 2.0 c 1.0 20 -c 10 6.9 t 1.2 QPDWGICLA
3 2 1 WYmGLGGG 2.1 2 0.6 522 3028 10.9 t 1.5 WYGAGLGGG ND” ND ND >loo
QPDWGIH-Q WYGQGLGGG 1.9 .c 0.5 QPDWGIH-E WYGEGLGGG
1.7 t 0.5 14 4 6.7 2 2.3
QPDWGIH-Y 1.3 0.3 1.8 t 0.4 13 * 3 6.9 2 0.8
WYGXGLGGG 1.6 +. 0.5 1.4 c 0.4 1224 WYGQGL-GG
7.1 t 1.2 2.0 c 0.3 1.9 t 0.4
QPDW W 13 t 2 7.4 t 2.1
1.6 c 0.3 0.6 t 0.1 5*1 5.9 * 0.4
QPDWGN-*A
QPDWGIHhA
QPDWGI-G
ND, not determined.
gest two important aspects of the binding site. First, the galac- tose-binding site probably resembles the sites seen in other plant, bacterial, and animal lectins, in which the B face of galactose is stacked against an aromatic residue 127). Second, high selectivity for galactose seems to be achieved by restrict- ing the mobility of the critical aromatic residue by interaction with the adjacent glycine-rich inserted sequence. This sequence probably forms a folded structure in which the conserved his- tidine residue may be positioned quite close to the sugar ligand.
It is interesting to note that natural C-type CRDs have evolved to exploit the range of different possibilities of affinity and selectivity. Based on analysis of protein and gene struc- tures, it has been suggested that several groups of galactose- binding C-type CRDs evolved from mannose-binding CRDs in- dependently (15). Thus, the high affinity, high selectivity binding site found in the asialoglycoprotein receptor, resulting from the presence of “rp’sg and the glycine-rich insertion, came about independently of the lower affinity, lower selectivity site in proteoglycan created by PheXs9 alone. The relatively modest nature of the changes needed to bring about these specificity changes starting from a mannose-binding CRD is consistent with the proposal that galactose-binding sites have evolved several different times.
Acknowledgments-We thank Raymond Dwek for providing NMR time at the Oxford Glycobiology Institute; Nydia Ruiz, Geoffrey Guile,
tion and other experimental procedures; and Bill Weis, Mark Wonnald, and Maureen Taylor for assisting with and expediting protein prepara-
Maureen Taylor, and Raymond Dwek for discussing and commenting on the manuscript.
REFERENCES 1. Drickamer, K., and Taylor, M. E. 11993) Annu. Reu. CelE Biol. 9,237-264 2. Weis, W. I., Kahn, R., Fourme, R., Drickamer, K., and Hendrickson, W. A.
3. Graves, B. J., Cmwther, R. L., Chandran, C., Rumberger, J. M., Li, S., Huang, 11991) Science 251,1608-1615
K.-S., Presky, D. H., Familletti, P. C., Wolitzky, B. A., and Burns, D. K. (1994) Nature 367,532438
4. Weis, W. I., Drickamer, K., and Hendrickson, W. A. (19921 Nature 360, 127-134
5. Lee, R. T., Ichikawa, Y., Fay, M., Drickamer, K., Shao, M.-C., and Lee, Y. C. (1991) J. Bid. Chem. 266,481M815
6. Bolmskov, U., Teisner, B., Willis, A. C., Ried, K. B. M., and Jensenius, J. C . (1993) J. Biol. Chem. 268,1012~10125
7. Connolly, D. T., Tbwnsend, R. R., Kawaguchi, K., Bell, W. R., and Lee, Y. C . (1982) J. Bzol. Chem. 257, 939-945
8. Drickamer, K. (1992) Nature 360, 183-186 9. Lee, R. T. (1982) B i ~ h m ~ t ~ 21,104&1050
10. Fornstedt, N., and Porath, J. (1975) FEBS Lett. 57,187-191 11. Iobst, S. T., Wormald, M. R., Weis, W. I., Dwek, R. A., and Drickamer, K. 11994)
12. Saleque, S., Ruiz, N., and Drickamer, K. 11993) GZ-wbioZogy 3,185-190 13. Quesenberry, M. S., and Drickamer, K. 11992) J. Biof. C k m . 267,10831-10841 14. Bezouska, K., Crichlow, G. V.. Rose, J. M., Taylor, M. E., and Drickamer, K.
15. Drickamer, K. (1993) Prog. Nucleic Acid Res. Mol. Biol. 46,207-232 16. Sastry, K., Zahedi, K., Lelias, J.-M., Whitehead, A. S., and Ezekowitz, R. A. B.
17. Takezawa, R., Shinzawa, K., Watanabe, Y., and Akaike, T. (1993) Biochim.
18. Sanford, J. P., and Doyle, D. (19901 Biochim. Bwphys. Acta 1087,259-261 19. Rauch, U., Karthikeyan, L., Maurel, E?, Margolis, R. U., and Margolis, R. K.
20. Kawasaki, T., Ii, M., Kozutsumi, Y., and Yamashina, I. (1986) C ~ ~ b o h y d ~ Res.
21. Dwek, R. A. (1973) Nuclear Mugentic Resonance in Biochemistry:Applications
23. Shaanan, B., Lis, H., and Sharon, N. (1991) Science 254,862-866 22. Pekins, S. J., and Dwek, R. A. (1980) Biochemistry 19,245-258
24. Montfort, W., Villafranca, J. E., Monzingo, A. F., Emst, S. R., Katzin, B.,
Chem. 262,5398-5403 Rutenber, E., Xuong, N., Hamlin, R., and Robertus, J. D. (1987) J. Biol.
25. Sixma, T. K., Pronk, S. E., Kalk, K. H., Wartna, E. S., van Zanten, B. A. M., Witholt, B., and Hol, W. G. J. (1991) Nature 351, 371-377
26. Lobsanov, Y. D., Gitt, M. A., Leffler, H., Barondes, S. H., and Rini, J. M. (1993) J. Bid. Chem. 268, 27034-27038
27. Vyas, N. K. (1991) Cum Opin. Struct. Biol. 1, 732-740
J. Biol. Chem. 269,1550&15511
(1991) J. Biol. C h m . 266, 11604-11609
(19911 J. Immunol. 147,692-697
Biophys. Acta 1172,220-222
(1992) J. Biol. Chem. 267, 1953619547
151, 197-206
to Enzyme Systems, Clarendon Press, Oxford
![Legume Lectins: Proteins with Diverse Applications · Legume Lectins: Proteins with Diverse Applications Irlanda Lagarda-Diaz 1, ... [30]. The specificity of legume lectins for some](https://img.pdfslide.net/doc/110x75/5fc6b4c426138432574b638e/legume-lectins-proteins-with-diverse-applications-legume-lectins-proteins-with.jpg)




























