Embed Size (px)
Citation preview

J BIOCHEM MOLECULAR TOXICOLOGYVolume 24, Number 2, 2010
Biochanin A Induction of Sulfotransferases in RatsYue Chen,1 Chaoqun Huang,1 Tianyan Zhou,2 Shunfen Zhang,1 and Guangping Chen1
1Department of Physiological Sciences, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK 74078, USA;E-mail: [email protected] of Pharmaceutics, School of Pharmaceutical Sciences, Peking University Health Science Center, Beijing 100083,People’s Republic of China
Received 3 February 2009; revised 22 June 2009; accepted 10 July 2009
ABSTRACT: Biochanin A (BCA) is a dietary isoflavonepresent in red clover (Trifoliumn pretense) and manyherbal products. BCA has been reported to havechemopreventive actions against various cancers in-cluding prostate, breast, colon cancer, and so on.Sulfotransferases are a family of phase II drug-metabolizing enzymes, which are important for xeno-biotic detoxification and regulation of biological sig-naling molecule biological activities. Sulfotransferasegene expressions are regulated by different hormonesand xenobiotics. Improper regulation of sulfotrans-ferases leads to improper functions of biological sig-naling molecules, which in turn can cause cancer orother diseases. BCA inhibits the enzyme activitiesof the phase I drug-metabolizing enzymes CYP1A1and CYP1B1 in Chinese hamster ovary cells and in-duces the phase II drug-metabolizing enzymes UDP-glucuronosyltransferases in human prostate cancercells. BCA induction of sulfotransferases has not beenstudied. This investigation evaluates the in vivo regu-lation of sulfotransferases at protein and mRNA lev-els in the liver and intestine of Sprague-Dawley ratstreated with BCA (0, 2, 10, and 50 mg/kg/day) for 7 days.Our experimental results demonstrate for the first timethat chronic BCA treatment can significantly inducethe expression of rat sulfotransferase 1A1 (rSULT1A1,AST-IV), sulfotransferase 2A1 (rSULT2A1, STa), andrat estrogen sulfotransferase (rSULT1E1, EST) in ratliver and intestine. Our Western blot results are ingood agreement with real-time RT-PCR data, suggest-ing that BCA induction of sulfotransferases occurs at
Correspondence to: Guangping Chen.Contract Grant Sponsor: NIH.Contract Grant Number: GM078606.Contract Grant Sponsor: American Cancer Society.Contract Grant Number: RSG-07-028-01-CNE.Contract Grant Sponsor: USDA.Contract Grant Number: 2006-35200-17137.Contract Grant Sponsor: Oklahoma Center for the Advance-
ment of Science and Technology (OCAST).Contract Grant Number: HR05-015.
c© 2010 Wiley Periodicals, Inc.
the transcriptional level. C© 2010 Wiley Periodicals, Inc.J Biochem Mol Toxicol 24:102–114, 2010; Published on-line in Wiley InterScience (www.interscience.wiley.com).DOI 10:1002/jbt.20318
KEYWORDS: Biochanin A; Rat sulfotransferase; Induc-tion; Gene regulation
INTRODUCTION
Biochanin A (BCA) is a dietary isoflavone, which isrich in red clover (Trifoliumn pretense) and many herbaldietary supplements. Herbal supplements used to al-leviate menopausal symptoms contain high amountsof BCA. It has been reported that red clover extract(PureWorld Botanicals, South Hackensack, NJ) contain14.5% biochanin A (by dry weight) [1]. BCA reportedlyhas several beneficial health effects: cardiovascularprotection, chemopreventive effects, antioxidant, anti-inflammatory, and anticarcinogenic effects [2]. At theconcentration of 5 μM, BCA has been reported to inhibitthe enzyme activities of the phase I drug-metabolizingenzymes CYP1A1 and CYP1B1 in Chinese hamsterovary cells [3] and to induce phase II drug-metabolizingenzymes UDP-glucuronosyltransferases in a humanprostate cancer cell line [4]. The regulation of drug-metabolizing enzymes by BCA is very important forhuman health since upregulation of detoxifying en-zymes may reduce the activity of carcinogenic com-pounds [5]. In addition, BCA may also interact withtherapeutic drugs by inducing or inhibiting theirmetabolism.
Cytosolic sulfotransferases (SULTs) catalyzethe sulfate conjugation of many hormones,neurotransmitters, and xenobiotic compounds [6,7].They are involved in the phase II detoxification ofxenobiotics as well as in the inactivation of endogenouscompounds such as steroid and thyroid hormones,
102

Volume 24, Number 2, 2010 BIOCHANIN A INDUCES RAT SULFOTRANSFERASES 103
neurotransmitters, catecholamines, and bile acids[6,8]. There are two major subfamilies of SULTs: thephenol SULTs (SULT1) and hydroxysteroid (alcohol)SULTs (SULT2), based on their substrate specificity.Hepatic phenol and alcohol SULTs show a distinctsex dimorphism in rats [9]. Higher immunoreactivityfor rSULT1A1 (AST-IV) and rSULT2A1 (STa) proteinshas been shown in the livers of male and female rats,respectively [10,11]. In 2007, Moon et al. reportedthat BCA can significantly upregulate the expressionof seven human drug metabolism enzyme genes,including CYP2A6, CYP2B6, CYP2C9, CYP2F1, MDR1,thromboxane A synthase 1 (TBXAS1), and SULT1A2in human hepatocytes as well as in human colonadenocarcinoma cells (LS-180) using microarray andreal-time RT-PCR methods [5]. To the best of ourknowledge, BCA induction of SULTs in rats has notbeen reported.
The objective of this study is to evaluate the in vivoregulation of SULT isoforms in the liver and intestine ofSprague-Dawley rats. The results of this study are im-portant for understanding drug–drug interactions andbiological functions of isoflavones. These results mayalso provide information on the anticancer activities ofisoflavones.
MATERIALS AND METHODS
Materials
BCA was purchased from Sigma-Aldrich (St. Louis,MO). Sodium dodecyl sulfate polyacrylamide gel elec-trophoresis (SDS-PAGE) reagents were obtained fromBio-Rad (Hercules, CA). Western blot chemilumines-cence reagent kits (Super Signal West Pico Stable Per-oxide and Super Signal West Pico Luminol/Enhancersolutions) were purchased from Pierce Chemical(Rockford, IL). Nitrocellulose membranes (Immobilon-P; Millipore Corporation, Bedford, MA) used for West-ern blots were purchased from Fisher Scientific (FairLawn, NJ). Total RNA extraction kit (TRIZOL reagent)was purchased from Molecular Research Center, Inc.(Cincinnati, OH). SuperScriptII Reverse Transcriptasewas from Invitrogen Corporation (Carlsbad, CA). Real-time PCR kits (qPCR MasterMix Plus for SYBR GreenI dNTP) were from EUROGENTEC (San Diego, CA).Protein assay reagent was obtained from Bio-Rad(Hercules, CA). All other reagents and chemicals wereof the highest analytical grade available.
Animals and Drug Treatments
Male and female Sprague-Dawley rats (Harlan,Indianapolis, IN), 10–11 weeks old and 200–300 g in
body weight, were used in this investigation. Rats werehoused in a temperature- and humidity-controlledroom and supplied with rodent chow and water for atleast 1 week before the experiments. Rats of both sexeswere divided into four groups, with three to four ratsin each group. BCA was dissolved in sterilized 75%DMSO/water and administered through intraperi-toneal (i.p.) injections (0.2 mL each time) at 2.0, 10,and 50 mg/kg/day for 1 week. The correspondinggroup of control rats of either sex received only vehicle(75% DMSO/water) injections. The animals weresacrificed 24 h after the final drug treatment. Liverswere collected, washed with sterile, ice-cold NaCl(0.9%, w/v) solution, and frozen in a liquid nitrogenbath. Intestinal lumens were carefully washed withsterile ice-cold NaCl (0.9%, w/v) solution. Luminalcavities were opened, and fat particles and smallblood vessels were trimmed out. The mucosal cellsfrom the small intestine and colon were collectedseparately by scraping and were frozen immediately ina liquid nitrogen bath. Samples were stored at −80◦Cuntil use.
Cytosolic Sample Preparation
Rat liver homogenates were prepared with 50 mMTris buffer containing 0.25 M sucrose (pH 7.5). Smallintestinal and colon mucosal homogenates were pre-pared with the same buffer containing 0.01 mg/mLtrypsin inhibitor and 10 μg/mL phenylmethylsul-fonyl fluoride. All homogenates were centrifuged at100,000×g for 1 h at 4◦C. Cytosol aliquots were col-lected in 1.5-mL microcentrifuge tubes and preservedat −80◦C for enzyme assays and Western blot assay.
Enzyme Assays
Rat SULT activity in the cytosol was determinedusing a radioactive assay method previously described[12]. Briefly, 50 μg of protein from rat liver cytosol and100 μg of protein from rat small intestinal and colonmucosal cytosol were used as the enzyme source foreach assay. For radioactive 2-naphthol sulfation activ-ity, [14C]2-naphthol (4.7 mCi/mmol; 0.1 mM final con-centration) was used. To determine DHEA sulfation ac-tivity, [3H]-DHEA (diluted to 0.4 Ci/mmol; 2 μM finalconcentration) was used. For all assays, 20 μM PAPSwas used. All enzymatic assays were performed in atotal reaction volume of 250 μL. After 30 min of in-cubation at 37◦C in a shaking water bath, the reactionwas stopped by adding 250 μL stop buffer (0.25 M Tris,pH 8.7). Extraction was performed twice by adding0.5 mL of water-saturated chloroform. After the finalextraction, 100 μL of the aqueous phase was used for
J Biochem Molecular Toxicology DOI 10:1002/jbt

104 CHEN ET AL. Volume 24, Number 2, 2010
scintillation counting. Assays were run in duplicate. Alldata represented the average of the results from threeexperiments.
Western Blot Analysis
Antibodies against rat sulfotransferases,rSULT1A1 (AST-IV) and rSULT2A1 (STa), were pro-vided by Dr. Michael W. Duffel (Division of Medicinaland Natural Products Chemistry, College of Pharmacy,The University of Iowa, Iowa city, IA). The primaryantibody against rSULT1E1 (EST) was purchased fromBiovision, Inc. (Boston, MA). The specific Westernblot procedure has been described previously [12–14].Briefly, cytosolic proteins from liver (10 μg) and smallintestine and colon (50 μg) were separated in a 12%polyacrylamide gel in an electrophoresis system (Bio-Rad, Hercules, CA). After running at 200 V, the proteinbands were transferred overnight at 35 V onto a PVDFnitrocellulose membrane at 4◦C. Membranes wereblocked for 1 h with 5% nonfat dry milk in Tris-bufferedsaline (TBS). For rat liver cytosol proteins, membraneswere incubated with either rabbit anti-rSULT1A1, rab-bit anti-rSULT2A1, or rabbit anti-rSULT1E1 primaryantibody (1:5000) in TBS containing 0.05% Tween 20(TBS-T) for 2 h on a shaker at room temperature.The membranes carrying intestinal samples wereincubated in the same antibodies (1:1000) overnightat 4◦C. After incubation, all membranes were washedwith TBS-T for 3 × 10 min and then incubated insecond antibody (horseradish peroxidase-conjugatedImmunopure goat anti-rabbit IgG, H + L) at 1:5000in 5% TBS-T, containing 5% nonfat dry milk for 1 h.Then the membranes were washed with TBS-T for 4 ×10 min. Fluorescent bands were developed with 3 mLof substrate, containing the same volume of each SuperSignal West Pico Luminol Enhancer solution and SuperSignal West Pico Stable Peroxidase solution at roomtemperature for 5 min. The fluorescent band signalswere recorded with a Bio-Rad Versa Doc 5000 ImageSystem, and densitometry analysis was performedwith Quantity One software (Bio-Rad, Hercules, CA).
Quantitative Real-Time RT-PCR
Total RNA was prepared from rat liver andintestinal cells using Trizol reagent according tothe manufacturer’s protocol. RNA samples wereincubated with RQ1 DNase at 37◦C for 30 min andthen inactivated at 65◦C for 10 min. Superscript II(Invitrogen) reverse transcriptase with 1 μg of totalRNA was used to synthesize first strand cDNA, and5 μL of 5× diluted reverse-transcribed cDNA product
served as the template in polymerase chain reactions.Real-time PCR was performed using qPCR MasterMixPlus for SYBR Green I dNTP kit (EUROGENTEC)according to the manufacturer’s instructions. Primerswere designed with primer express as follows—ratACTB-F320: 5′-AGGCCCCTCTGAACCCTAAG-3′, ratACTB-R435: 5′-AGAGGCATACAGGGACAACACA-3′ (GenBank accession no. 56227); rSULT1A1-F530:5′-AGCTGAGACACACTCACCCTGTT-3′, rSULT1A1-R651: 5′-ATCCACAGTCTCCTCGGGTAGA-3′
(GenBank accession no. X52883); rSULT2A1-F496:5′-ATCCGTGCCTGGCTGTCTAT-3′, rSULT2A1-R642: 5′-GAGGACCAAATCCAGCTCATCT-3′
(GenBank accession no. M33329); rSULT1E1-F706:5′-CCATGCACCAATTATTCAATGC-3′, rSULT1E1-R815: 5′-AGGGCTTCAGGGAAGTGGTT-3′ (GenBankaccession no. M86758). Real-time PCR was performedon an ABI 7500 Fast Real Time PCR System (AppliedBiosystems, Foster City, CA). Initially, regular PCRproduct DNA was purified with GENECLEAN Turbo(Qbiogene, Carlsbad, CA) for constructing standardcurves (103–108 copies gene). A standard curve wasplotted: threshold cycle (CT) vs. logarithmic valueof the gene copy number. The gene copy number ofunknown samples was generated directly from thestandard curve by Sequence Detector 1.7 software. Atleast two duplications were run for each experiment,and each experiment was repeated at least three times.All gene mRNA copy numbers were normalized to ratβ-actin mRNA (rSULT mRNA/β-actin mRNA).
Statistical Analysis
One-way ANOVA followed by the Dunnett’s testwas used to calculate the statistical significance ofthe difference between the control means and BCA-treatment means. In all cases, *, p < 0.05 was consideredsignificant; **, p < 0.01 was considered very significant.Data presented in the figures are means ± SD (standarddeviation) of the data collected separately from at leastthree individual animals.
RESULTS
Western Blot Analysis of Rat LiverrSULT1A1 and rSULT2A1 Proteins
Figure 1 shows Western blot results for the effectsof BCA on male rat liver rSULT1A1, rSULT2A1, andrSULT1E1 proteins. The densitometry data were ana-lyzed to semiquantify the relative amount of each pro-tein. These data demonstrate that, in male rat liver,rSULT1A1 protein levels increased about 2.0- and
J Biochem Molecular Toxicology DOI 10:1002/jbt

Volume 24, Number 2, 2010 BIOCHANIN A INDUCES RAT SULFOTRANSFERASES 105
FIGURE 1. Representative Western blot and densitometry analyses of rSULT1A1, rSULT2A1, and rSULT1E1 in the liver of male rats treatedwith different doses of BCA for 7 days. Division factors are expressed and plotted as relative densities. BCA = Biochanin A. ∗ p < 0.05, ∗∗ p < 0.01compared with the control group.
1.8-fold, respectively, in response to 10 and50 mg/kg/day BCA treatment; and rSULT2A1 proteinlevels increased about 2.2- and 2.0-fold, respectively,in response to 2 and 10 mg/kg/day BCA treatment(Figure 1). We also found that rSULT1E1 protein levelsincreased significantly at all doses of BCA.
In female rat liver, rSULT1A1 protein levels in-creased about 1.7- and 2.0-fold, respectively, in re-sponse to 2 and 10 mg/kg/day BCA treatment for7 days (Figure 2). rSULT2A1 protein levels increasedabout 1.2- and 1.3-fold, respectively, in response to2 and 10 mg/kg/day BCA treatment for 7 days(Figure 2). We found, however, that in female rat liverthe 50 mg/kg/day BCA treatment did not signifi-cantly change rSULT1A1 and rSULT2A1 protein levels(Figure 2).
The Western blot results from male and female ratliver cells suggest that the BCA treatment can signifi-cantly induce rSULT1A1 and rSULT2A1 protein expres-sion in a dose-dependent manner in male and femalerat liver and can significantly induce rSULT1E1 proteinin male rat liver.
Western Blot Analysis of Rat IntestinerSULT1A1 and rSULT2A1 Proteins
The Western blot and densitometry analysis inFigure 3 shows that, in male rat small intestinal mu-cosal cells, rSULT1A1 protein expression levels in-creased about 2.2-, 3.0-, and 3.2-fold, respectively, inresponse to 2, 10, and 50 mg/kg/day BCA treatment.rSULT2A1 protein levels also increased about 2.1-,2.4-, and 2.2-fold, respectively, in response to 2, 10,and 50 mg/kg/day BCA treatment. The induction ofrSULT1A1and rSULT2A1 proteins by BCA was dosedependent (Figure 3).
Western blot results shown in Figure 4 demon-strate that, in female rat small intestinal mucosal cells,rSULT1A1 protein levels increased 2.5-, 2.1-, and 2.3-fold, respectively, in response to a 7-day BCA treatmentof 2, 10, and 50 mg/kg/day; whereas rSULT2A1 pro-tein levels increased 2.0-, 2.2- and 4-fold, respectively.The induction of rSULT2A1 proteins by BCA was dosedependent (Figure 4).
The Western blot results and densitometry analysisof male and female rat small intestinal mucosal cells
J Biochem Molecular Toxicology DOI 10:1002/jbt

106 CHEN ET AL. Volume 24, Number 2, 2010
FIGURE 2. Representative Western blot and densitometry analyses of rSULT1A1 and rSULT2A1 in the liver of female rats treated with differentdoses of BCA for 7 days. Division factors are expressed and plotted as relative densities. BCA = Biochanin A. ∗ p < 0.05, ∗∗ p < 0.01 comparedwith the control group.
demonstrate that BCA treatment significantly inducedrSULT1A1 and rSULT2A1 protein expression in a dose-dependent manner in rat small intestinal mucosal cells.
Western Blot Analysis of Rat ColonrSULT1A1 and rSULT2A1 Proteins
In male colon mucosal cells, rSULT1A1 protein ex-pression levels increased 3.0- and 4.0-fold, respectively,
in response to 2 and 10 mg/kg/day BCA treatment(Figure 5). However, 50 mg/kg/day BCA treatment didnot significantly change rSULT1A1 protein expressionlevels in male colon mucosal cells. In female colon mu-cosal cells, rSULT2A1 protein levels increased 2.5-foldin response to 10 mg/kg/day BCA treatment (Figure 5),and rSULT2A1 protein levels decreased in response to50 mg/kg/day BCA treatment.
The Western blot results and densitometry analysisof male and female rat colon mucosal cells demonstrate
FIGURE 3. Representative Western blot and densitometry analyses of rSULT1A1 and rSULT2A1 in the small intestinal mucosa cells of malerats treated with different doses of BCA for 7 days. Division factors are expressed and plotted as relative densities. BCA = Biochanin A. ∗ p <
0.05, ∗∗ p < 0.01, ∗∗∗ p < 0.001 compared with the control group.
J Biochem Molecular Toxicology DOI 10:1002/jbt

Volume 24, Number 2, 2010 BIOCHANIN A INDUCES RAT SULFOTRANSFERASES 107
FIGURE 4. Representative Western blot and densitometry analyses of rSULT1A1 and rSULT2A1 in the small intestinal mucosa cells of femalerats treated with different doses of BCA for 7 days. Division factors are expressed and plotted as relative densities. BCA = Biochanin A. ∗ p <
0.05, ∗∗ p < 0.01 compared with the control group.
that 10 mg/kg/day BCA treatment can significantlyinduce rSULT1A1 protein expression in male colon andrSULT2A1 protein expression in female rat colon.
Real-Time RT-PCR Analysis of Rat LiverSulfotransferases
To determine whether BCA’s effects on rat SULTsoccur at the translational (protein) or transcrip-
tional (mRNA) level, we designed gene-specific real-time PCR primers against rat rSULT1A1, rSULT2A1,and rSULT1E1 (see the section Material and Meth-ods). Results and data analysis of real-time RT-PCR experiments (Figure 6) demonstrated that BCAinduced the transcriptional activity of rSULT1A1gene (Figure 6A), rSULT2A1 gene (Figure 6B), andrSULT1E1 gene (Figure 6C) in male rat liver in adose-dependent manner. In female liver, the transcrip-tional activity of rSULT1A1 gene (Figure 6D) and
FIGURE 5. Representative Western blot and densitometry analyses of rSULT1A1 and rSULT2A1 in colorectal mucosa cells of male and femalerats treated with different doses of BCA for 7 days. Division factors are expressed and plotted as relative densities. BCA = Biochanin A. ∗ p <
0.05, ∗∗ p < 0.01 compared with the control group.
J Biochem Molecular Toxicology DOI 10:1002/jbt

108 CHEN ET AL. Volume 24, Number 2, 2010
FIGURE 6. Real-time RT-PCR results showing the levels of rSULT1A1, rSULT2A1, and rSULT1E1 mRNA expression in the livers of male(A, SULT1A1; B, rSULT2A1; C, rSULT1E1) and female (D, rSULT1A1; E, rSULT2A1) rats treated with different doses of BCA for 7 days. AllmRNA gene copy numbers were normalized to rat β-actin mRNA. BCA = Biochanin A. ∗ p < 0.05 compared with the control group.
rSULT2A1 gene (Figure 6E) were significantly inducedby 10 mg/kg/day BCA treatment. This type of gender-specific transcriptional regulation of rat SULTs is ingood agreement with our rat liver Western blot re-sults (Figures 1 and 2). These data suggest that BCA’sregulation of rat SULTs occurs at the transcriptionallevel.
Real-Time RT-PCR Analysis of Rat SmallIntestinal Mucosal Cell Sulfotransferases
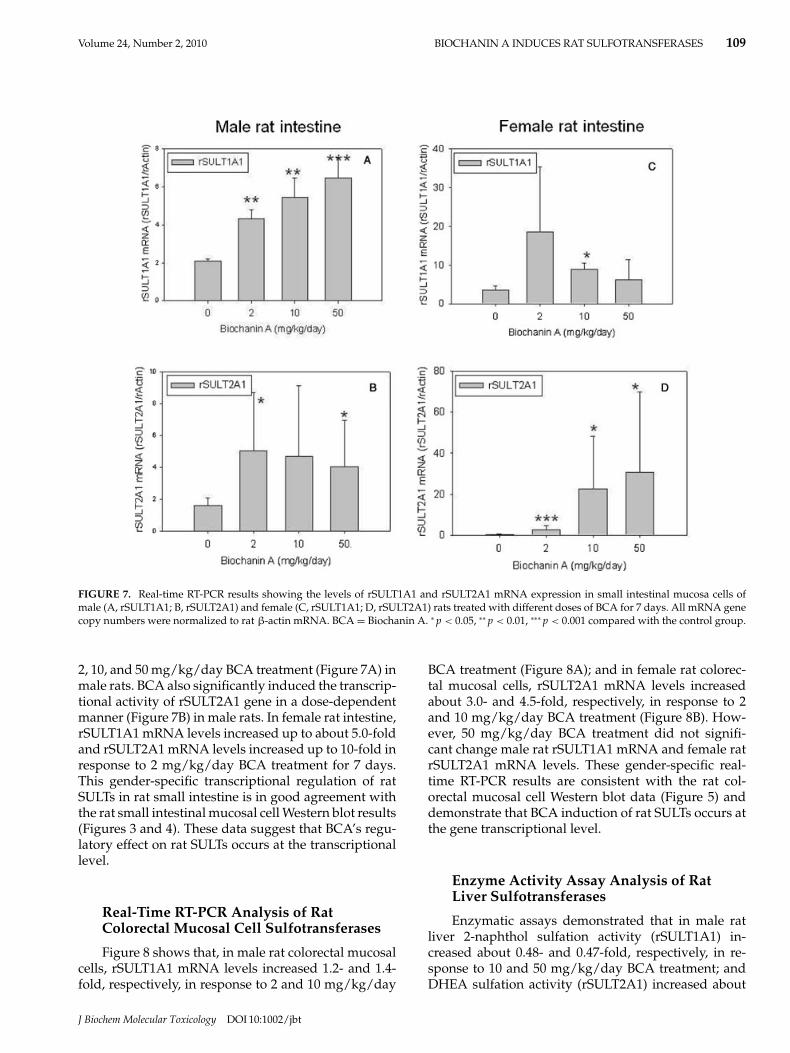
The results of real-time RT-PCR experiments on ratsmall intestinal mucosal cells shown in Figure 7 demon-strate that BCA induced rSULT1A1 gene transcrip-tion 2.1-, 2.5-, and 3.0-fold, respectively, in response to
J Biochem Molecular Toxicology DOI 10:1002/jbt
Volume 24, Number 2, 2010 BIOCHANIN A INDUCES RAT SULFOTRANSFERASES 109
FIGURE 7. Real-time RT-PCR results showing the levels of rSULT1A1 and rSULT2A1 mRNA expression in small intestinal mucosa cells ofmale (A, rSULT1A1; B, rSULT2A1) and female (C, rSULT1A1; D, rSULT2A1) rats treated with different doses of BCA for 7 days. All mRNA genecopy numbers were normalized to rat β-actin mRNA. BCA = Biochanin A. ∗ p < 0.05, ∗∗ p < 0.01, ∗∗∗ p < 0.001 compared with the control group.
2, 10, and 50 mg/kg/day BCA treatment (Figure 7A) inmale rats. BCA also significantly induced the transcrip-tional activity of rSULT2A1 gene in a dose-dependentmanner (Figure 7B) in male rats. In female rat intestine,rSULT1A1 mRNA levels increased up to about 5.0-foldand rSULT2A1 mRNA levels increased up to 10-fold inresponse to 2 mg/kg/day BCA treatment for 7 days.This gender-specific transcriptional regulation of ratSULTs in rat small intestine is in good agreement withthe rat small intestinal mucosal cell Western blot results(Figures 3 and 4). These data suggest that BCA’s regu-latory effect on rat SULTs occurs at the transcriptionallevel.
Real-Time RT-PCR Analysis of RatColorectal Mucosal Cell Sulfotransferases
Figure 8 shows that, in male rat colorectal mucosalcells, rSULT1A1 mRNA levels increased 1.2- and 1.4-fold, respectively, in response to 2 and 10 mg/kg/day
BCA treatment (Figure 8A); and in female rat colorec-tal mucosal cells, rSULT2A1 mRNA levels increasedabout 3.0- and 4.5-fold, respectively, in response to 2and 10 mg/kg/day BCA treatment (Figure 8B). How-ever, 50 mg/kg/day BCA treatment did not signifi-cant change male rat rSULT1A1 mRNA and female ratrSULT2A1 mRNA levels. These gender-specific real-time RT-PCR results are consistent with the rat col-orectal mucosal cell Western blot data (Figure 5) anddemonstrate that BCA induction of rat SULTs occurs atthe gene transcriptional level.
Enzyme Activity Assay Analysis of RatLiver Sulfotransferases
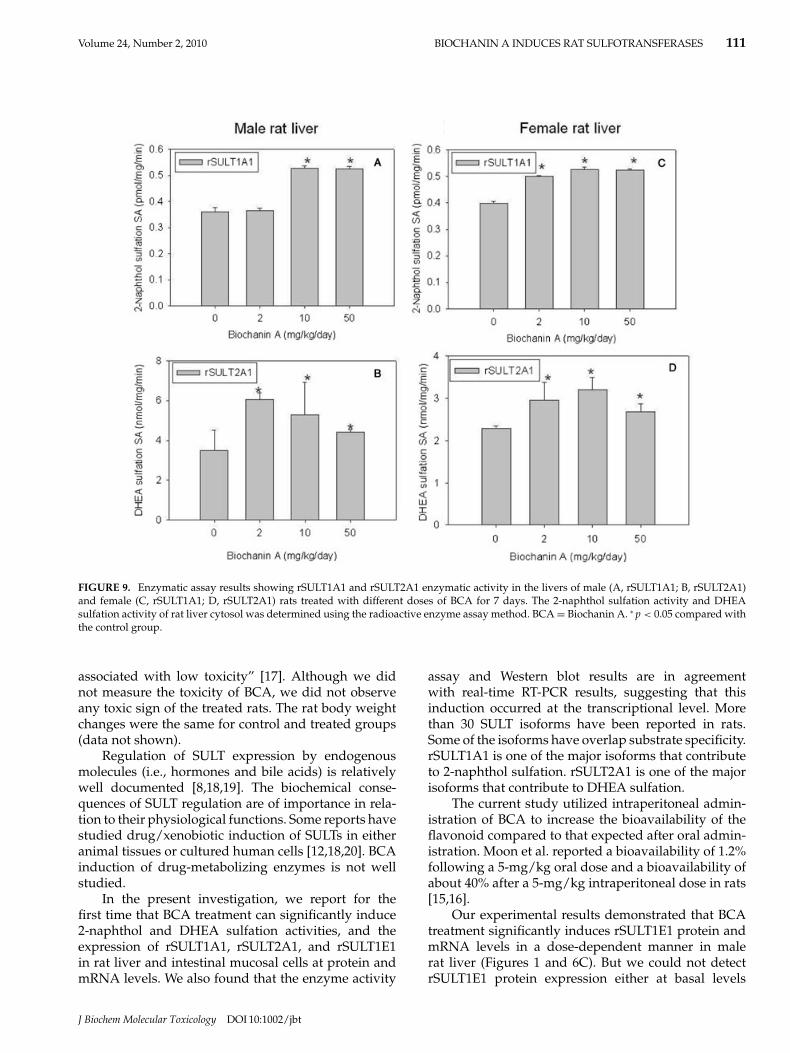
Enzymatic assays demonstrated that in male ratliver 2-naphthol sulfation activity (rSULT1A1) in-creased about 0.48- and 0.47-fold, respectively, in re-sponse to 10 and 50 mg/kg/day BCA treatment; andDHEA sulfation activity (rSULT2A1) increased about
J Biochem Molecular Toxicology DOI 10:1002/jbt

110 CHEN ET AL. Volume 24, Number 2, 2010
Biochanin A (mg/kg/day)
0 2 10 50
cDN
A n
um
ber
of A
ST
-IV
gene
0.0
0.2
0.4
0.6
0.8
Biochanin A (mg/kg/day)
0 2 10 50
cDN
A n
um
ber
of S
Tag
ene
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
A B
Male rat colon
**
*
Female rat colon
FIGURE 8. Real-time RT-PCR results showing the levels of rSULT1A1and rSULT2A1 mRNA expression in colorectal mucosa cells of male (A,rSULT1A1) and female (B, rSULT2A1) rats treated with different doses of BCA for 7 days. All mRNA gene copy numbers were normalized torat β-actin mRNA. BCA = Biochanin A. ∗ p < 0.05 compared with the control group.
0.73-, 0.51-, and 0.26-fold, respectively, in response to2, 10, and 50 mg/kg/day BCA treatment. In femalerat liver, 2-naphthol sulfation activity increased about0.26-, 0.32-, and 0.32-fold, respectively, in response to2, 10, and 50 mg/kg/day BCA treatment for 7 days.DHEA sulfation activity increased about 0.29- and 0.40-fold, respectively, in response to 2 and 10 mg/kg/dayBCA treatment for 7 days (Figure 9D) in female rat liver.The enzyme activity assay results are basically in agree-ment with Western blot and real-time RT-PCR resultsfor rSULT1A1 and rSULT2A1.
Enzyme Activity Assay Analysis of RatSmall Intestinal Mucosal CellSulfotransferases
In Figure 10, the enzymatic activity assay resultsshow that, in male rat small intestinal mucosal cells, 2-naphthol sulfation activity (rSULT1A1) increased about0.20- and 0.21-fold, respectively, in response to 2 and10 mg/kg/day BCA treatment. DHEA sulfation ac-tivity (rSULT2A1) also increased about 0.35-, 0.38-,and 0.35-fold, respectively, in response to 2, 10, and50 mg/kg/day BCA.
In the female rat intestinal mucosal cells, 2-naphthol sulfation activity increased about 0.37-and 0.25-fold, respectively, in response to 2 and10 mg/kg/day BCA treatment for 7 days (Figure 10C).DHEA sulfation activity increased about 0.20-, 0.21-,and 0.33-fold, respectively, in response to 2, 10, and50 mg/kg/day BCA treatment for 7 days (Figure 10D).The induction of 2-naphthol and DHEA sulfation ac-tivities by BCA was dose dependent.
Enzyme Activity Assay Analysis of RatColorectal Mucosal Cell Sulfotransferases
In male rat colorectal mucosal cells, 2-naphtholsulfation activity significantly increased 0.35-fold after10 mg/kg/day BCA treatment for 7 days. In female ratcolorectal mucosal cells, DHEA sulfation activity sig-nificantly increased 0.44- and 0.48-fold, respectively, inresponse to 10 and 50 mg/kg/day BCA treatment for7 days (Figure 11B). The upregulation of DHEA sul-fation activity was in a dose-dependent manner. Theenzyme activity assay results of rat colorectal mucosalcells are basically in agreement with Western blot andreal-time RT-PCR results.
DISCUSSION
BCA, 4′-O-methyl derivative of genistein (GEN), isthe major isoflavone in red clover (Trifolium pratense).BCA has been reported to have chemopreventive prop-erties; its metabolites include GEN, BCA conjugates,and GEN conjugates [15,16]. These metabolites maycontribute to the chemopreventive effects of BCA [16].People who have different dietary habit (vegetablesand beverages), or those who take dietary supplements,may intake very high amount of BCA or their metabolicderivatives. It has been reported that red clover extract(PureWorld Botanicals, South Hackensack, NJ) contain14.5% biochanin A (by dry weight) [1]. Pharmacokinet-ics and bioavailability of BCA in rats by different ad-ministration routes, at the doses of 1, 5, and 50 mg/kg,had been studied [15]. In this study, we used 2, 10, and50 mg/kg body weight per day for the investigation.It is reported that flavonoids “are extremely safe and
J Biochem Molecular Toxicology DOI 10:1002/jbt
Volume 24, Number 2, 2010 BIOCHANIN A INDUCES RAT SULFOTRANSFERASES 111
FIGURE 9. Enzymatic assay results showing rSULT1A1 and rSULT2A1 enzymatic activity in the livers of male (A, rSULT1A1; B, rSULT2A1)and female (C, rSULT1A1; D, rSULT2A1) rats treated with different doses of BCA for 7 days. The 2-naphthol sulfation activity and DHEAsulfation activity of rat liver cytosol was determined using the radioactive enzyme assay method. BCA = Biochanin A. ∗ p < 0.05 compared withthe control group.
associated with low toxicity” [17]. Although we didnot measure the toxicity of BCA, we did not observeany toxic sign of the treated rats. The rat body weightchanges were the same for control and treated groups(data not shown).
Regulation of SULT expression by endogenousmolecules (i.e., hormones and bile acids) is relativelywell documented [8,18,19]. The biochemical conse-quences of SULT regulation are of importance in rela-tion to their physiological functions. Some reports havestudied drug/xenobiotic induction of SULTs in eitheranimal tissues or cultured human cells [12,18,20]. BCAinduction of drug-metabolizing enzymes is not wellstudied.
In the present investigation, we report for thefirst time that BCA treatment can significantly induce2-naphthol and DHEA sulfation activities, and theexpression of rSULT1A1, rSULT2A1, and rSULT1E1in rat liver and intestinal mucosal cells at protein andmRNA levels. We also found that the enzyme activity
assay and Western blot results are in agreementwith real-time RT-PCR results, suggesting that thisinduction occurred at the transcriptional level. Morethan 30 SULT isoforms have been reported in rats.Some of the isoforms have overlap substrate specificity.rSULT1A1 is one of the major isoforms that contributeto 2-naphthol sulfation. rSULT2A1 is one of the majorisoforms that contribute to DHEA sulfation.
The current study utilized intraperitoneal admin-istration of BCA to increase the bioavailability of theflavonoid compared to that expected after oral admin-istration. Moon et al. reported a bioavailability of 1.2%following a 5-mg/kg oral dose and a bioavailability ofabout 40% after a 5-mg/kg intraperitoneal dose in rats[15,16].
Our experimental results demonstrated that BCAtreatment significantly induces rSULT1E1 protein andmRNA levels in a dose-dependent manner in malerat liver (Figures 1 and 6C). But we could not detectrSULT1E1 protein expression either at basal levels
J Biochem Molecular Toxicology DOI 10:1002/jbt

112 CHEN ET AL. Volume 24, Number 2, 2010
FIGURE 10. Enzymatic assay results showing rSULT1A1 and rSULT2A1 enzymatic activity in small intestinal mucosa cells of male(A, rSULT1A1; B, rSULT2A1) and female (C, rSULT1A1; D, rSULT2A1) rats treated with different doses of BCA for 7 days. The 2-naphtholsulfation activity and DHEA sulfation activity of rat small intestinal cytosol was determined using the radioactive enzyme assay method. BCA= Biochanin A. ∗ p < 0.05 compared with the control group.
FIGURE 11. Enzymatic assay results showing BCA induction of rSULT1A1 and rSULT2A1 enzymatic activity in colorectal mucosa cells ofmale (A, rSULT1A1) and female (B, rSULT2A1) rats treated with different doses of BCA for 7 days. The 2-naphthol sulfation activity and DHEAsulfation activity of rat colorectal cytosol was determined using the radioactive enzyme assay method. BCA = Biochanin A. ∗ p < 0.05 comparedwith the control group.
J Biochem Molecular Toxicology DOI 10:1002/jbt

Volume 24, Number 2, 2010 BIOCHANIN A INDUCES RAT SULFOTRANSFERASES 113
or after BCA treatment in female rat liver (data notshown). Hellriegel et al. reported that rSULT1E1 wasinduced in male rats after tamoxifen (TAM) treatment[11]. However, they also did not determine rSULT1E1protein levels before and after TAM treatment infemale rats. Unlike rSULT2A1 induction in rats, BCA-mediated rSULT1E1 induction exerts its beneficialeffects against estrogen-dependent carcinogenesis inhumans. rSULT1E1 catalyzes the sulfation of estro-gens to make them biologically inactive. Moon et al.reported that BCA treatment can inhibit breast cancertumor growth in a murine xenograft model either aloneor in combination with other flavonoids [21]. Anotherrecent study also showed that BCA protects against7,12-dimethylbenz(a)anthracene induced mammarytumors in a prepubertal female rat model and has amodulatory effect on phase II antioxidative enzymes,GSH content, and lipid peroxidation in prepubertalrat liver and mammary gland [22]. Although both epi-demiological and in vivo studies have demonstratedthe protective role of the phytoestrogen BCA againstbreast cancer, the molecular mechanism of BCA cancerchemoprevention has not been elucidated. One ofthe proposed mechanisms for cellular protection isthe induction of phase II detoxification enzymes andantioxidant enzymes [22].
Here, we investigated the effect of BCA treatmenton rSULT1A1 and rSULT2A1 protein and mRNA ex-pression in rat liver and intestinal mucosal cells. Ourresults demonstrated that BCA treatment increasedrSULT1A1and rSULT2A1 protein levels in the liver ofmale rats up to 2.0- (p < 0.01) and 2.2-fold (p < 0.01),respectively. In female rat liver, the highest increase inrSULT1A1 and rSULT2A1 protein levels was 2.0- (p <
0.01) and 1.3-fold (p < 0.05), respectively. Our previousstudy showed that GEN, a metabolite of BCA, signif-icantly induced human phenol-catalyzing sulfotrans-ferase (hSULT1A1) and human dehydroepiandros-terone sulfotransferase (hSULT2A1) in human HepG2and Caco-2 cells [14]. Moon et al. also reported that20 μM BCA upregulates human hSULT1A2 gene ex-pression in human hepatocytes [23]. SULT1A2 is themost efficient enzyme in sulfating some aromatic hy-droxylamines [24]. It has been reported that humanphenol-sulfating form of sulfotransferase, hSULT1A1(the counterpart of rSULT1A1), human hSULT1E1 (thecounterpart of rSULT1E1), and human hSULT2A1 (thecounterpart of rSULT2A1) also have GEN-sulfating ac-tivities at higher substrate concentrations [25,26]. Theseresults will be helpful for evaluating the metabolic na-ture of exogenously administered phytoestrogens orisoflavones in liver and intestinal mucosal cells.
Our previous study confirmed that human smallintestinal SULTs vary highly among different individ-
uals compared with human liver SULTs [13]. Intestinalenzymes are different from enzymes of other tissuesin that intestinal enzymes are mainly exposed to ex-ogenous xenobiotics as they first pass through the in-testines. SULTs in rat intestinal mucosal cells may playan important role in drug metabolism and xenobioticdetoxification. To our best knowledge, the induction ofintestinal SULTs by BCA is basically unknown. In thepresent investigation, we report for the first time thatBCA treatment can induce rSULT1A1 and rSULT2A1expression significantly at mRNA and protein levelsin male and female rat small intestinal and colorectalmucosal cells. The up to 3-fold increase in rSULT1A1(male) mRNA levels and the up to 10-fold increase inrSULT2A1 (female) mRNA levels in intestinal (smallintestine and colon) mucosal cells suggest that theseSULT genes are much more inducible in intestinal mu-cosal cells than in liver after BCA treatment. Moreover,the induction of SULT protein and mRNA expressionin rat intestinal mucosal cells are gender dependent. Itis known that in rat liver rSULT1A1 levels are higherin males than in females, whereas rSULT2A1 levels arehigher in females than in males [10].
BCA was reported to significantly induce UDP-glucuronosyltransferase activity at a concentrationrange of 0.5–50 μM in an androgen-responsive prostatecancer cell line [4]. BCA is a well-established activa-tor of the estrogen receptor (EC50 = 0.99 μM) and hasrecently been shown to be potent activator of PPARα
and PPARγ (EC50 < 1.0 μM) [27]. Han et al. suggestedthat BCA might be a ligand of the aryl hydrocarbonreceptor (AhR) [28]. Recently, Medjakovic and Jung-bauer demonstrated that BCA is a potent agonist ofAhRs with EC50 values of 0.25 μM [29]. We hypothe-size that BCA induction of SULT genes in rat occurs atthe transcriptional level and that the induction is likelyto be mediated through nuclear receptors. SULT in-duction mechanisms by isoflavones are unknown. Weare currently investigating nuclear receptor-mediatedSULT isoflavone induction mechanisms.
In summary, this is the first study to demonstratethat BCA, a dietary component present in food andherbal products, can significantly induce SULT enzymeactivities and gene expressions in rat liver and in-testines after chronic exposure (7 days). Our enzymeactivity assay and Western blot results are in agree-ment with real-time RT-PCR data, which suggest thatthe BCA induction of SULTs expression occurs at thetranscriptional level in both rat liver and intestinal mu-cosal cells. Drug-metabolizing enzyme induction caninfluence drug metabolism and xenobiotic detoxifica-tion. This induction can also cause carcinogenesis be-cause SULT induction can increase the bioactivation ofprocarcinogens and promutagens. Induction of SULTs
J Biochem Molecular Toxicology DOI 10:1002/jbt

114 CHEN ET AL. Volume 24, Number 2, 2010
can also interrupt their normal biological functions in-cluding the regulation of the functions and activities ofbiosignaling molecules.
REFERENCES
1. Booth NL, Nikolic D, van Breemen RB, Geller SE,Banuvar S, Shulman LP, Farnsworth NR. Confusionregarding anticoagulant coumarins in dietary supple-ments. Clin Pharmacol Ther 2004;76(6):511–516.
2. Cassileth BR, Vickers AJ. Soy: an anticancer agent inwide use despite some troubling data. Cancer Invest2003;21(5):817–818.
3. Chan HY, Wang H, Leung LK. The red clover (Trifoliumpratense) isoflavone biochanin A modulates the biotrans-formation pathways of 7,12-dimethylbenz[a]anthracene.Br J Nutr 2003;90(1):87–92.
4. Sun XY, Plouzek CA, Henry JP, Wang TT, Phang JM.Increased UDP-glucuronosyltransferase activity and de-creased prostate specific antigen production by biochaninA in prostate cancer cells. Cancer Res 1998;58(11):2379–2384.
5. Moon YJ, Zhang S, Brazeau DA, Morris ME. Effects ofthe flavonoid biochanin A on gene expression in primaryhuman hepatocytes and human intestinal cells. Mol NutrFood Res 2007;51(3):317–323.
6. Coughtrie MW, Sharp S, Maxwell K, Innes NP. Biologyand function of the reversible sulfation pathway catal-ysed by human sulfotransferases and sulfatases. ChemBiol Interact. 1998;109(1–3):3–25.
7. Klaassen C, Boles JW. Sulfation and sulfotrans-ferases 5: the importance of 3′-phosphoadenosine 5′-phosphosulfate (PAPS) in the regulation of sulfation.FASEB J 1997;11(6):404–418.
8. Klaassen C, Liu L, Dunn RT, II. Regulation of sulfotrans-ferase mRNA expression in male and female rats of var-ious ages. Chem Biol Interact 1998;109: 299–313.
9. Rajkowski KM, Robel P, Baulieu EE. Hydroxysteroid sul-fotransferase activity in the rat brain and liver as a func-tion of age and sex. Steroids 1997;62(5):427–436.
10. Chen G, Baron J, Duffel MW. Enzyme- and sex-specificdifferences in the intralobular localizations and distri-butions of aryl sulfotransferase IV (tyrosine-ester sul-fotransferase) and alcohol (hydroxysteroid) sulfotrans-ferase a in rat liver. Drug Metab Dispos 1995;23:1346–1353.
11. Hellriegel ET, Matwyshyn GA, Fei P, Dragnev KH, NimsRW, Lubet RA, Kong AN. Regulation of gene expres-sion of various phase I and phase II drug-metabolizingenzymes by tamoxifen in rat liver. Biochem Pharmacol1996;52(10):1561–1568.
12. Maiti S, Chen G. Methotrexate is a novel inducer of ratliver and intestinal sulfotransferases. Arch Biochem Bio-phys 2003a;418(2):161–168.
13. Chen G, Zhang Daqing, Jing Nin, Yin Shuhua, FalanyCharles N, Radominska-Pandya Anna. Human gastroin-testinal sulfotransferases: identification and distribution.Toxicol Appl Pharmacol 2003;187:186–197.
14. Chen Y, Huang C, Zhou T, Chen G. Genistein inductionof human sulfotransferases in HepG2 and Caco-2 cells.Basic Clin Pharmacol Toxicol 2008;103(6):553–559.
15. Moon YJ, Sagawa K, Frederick K, Zhang S, Morris ME.Pharmacokinetics and bioavailability of the isoflavonebiochanin A in rats. AAPS J 2006;8(3):E433–E442.
16. Moon YJ, Morris ME. Pharmacokinetics and bioavailabil-ity of the bioflavonoid biochanin A: effects of quercetinand EGCG on biochanin A disposition in rats. Mol Pharm2007;4(6):865–872.
17. Moon YJ, Wang X, Morris ME. Dietary flavonoids: ef-fects on xenobiotic and carcinogen metabolism. ToxicolIn Vitro 2006;20(2):187–210.
18. Runge-Morris M, Rose K, Falany CN, Kocarek TA. Differ-ential regulation of individual sulfotransferase isoformsby phenobarbital in male rat liver. Drug Metab Dispos1998;26(8):795–801.
19. Dunn RT, Klaassen CD. Thyroid hormone modulationof rat sulfotransferase mRNA expression. Xenobiotica2000;30:345–357.
20. Maiti S, Chen G. Tamoxifen induction of aryl sulfo-transferase and hydroxysteroid sulfotransferase in maleand female rat liver and intestine. Drug Metab Dispos2003;31(5):637–644.
21. Moon YJ, Shin BS, An G, Morris ME. Biochanin A inhibitsbreast cancer tumor growth in a murine xenograft model.Pharm Res 2008;25(9):2158–2163.
22. Mishra P, Kale RK, Kar A. Chemoprevention of mam-mary tumorigenesis and chemomodulation of the antiox-idative enzymes and peroxidative damage in prepubertalSprague Dawley rats by Biochanin A. Mol Cell Biochem2008;312(1–2):1–9.
23. Moon YJ, Brazeau DA, Morris ME. Effects of flavonoidsgenistein and biochanin a on gene expression and theirmetabolism in human mammary cells. Nutr Cancer2007;57(1):48–58.
24. Meinl W, Meerman JH, Glatt H. Differential activation ofpromutagens by alloenzymes of human sulfotransferase1A2 expressed in Salmonella typhimurium. Pharmacoge-netics 2002;12:677–689.
25. Doerge D, Chang HC, Churchwell MI, Holder CL. Anal-ysis of soy isoflavone conjugation in vitro and in humanblood using liquid chromatography-mass spectrometry.Drug Metab Dispos. 2000;3:298–307.
26. Gauguly T, Krasnykh V, Falany CN. Bacterial expressionand kinetic characteriztion of the human monoamine-sulfating form of phenol sulfotransferase. Drug MetabDispos 1995;23:945–950.
27. Shen P, Liu MH, Ng TY, Chan YH, Yong EL. Differen-tial effects of isoflavones, from Astragalus membranaceusand Pueraria thomsonii, on the activation of PPARalpha,PPARgamma, and adipocyte differentiation in vitro. JNutr 2006;136(4):899–905.
28. Han EH, Kim JY, Jeong HG. Effect of biochanin A on thearyl hydrocarbon receptor and cytochrome P450 1A1 inMCF-7 human breast carcinoma cells. Arch Pharm Res2006;29(7):570–576.
29. Medjakovic S, Jungbauer A. Red clover isoflavonesbiochanin A and formononetin are potent ligands of thehuman aryl hydrocarbon receptor. J Steroid Biochem MolBiol 2008;108(1–2):171–177.
J Biochem Molecular Toxicology DOI 10:1002/jbt





























