Embed Size (px)
Citation preview
Biochemistry 432/832
September 03September 03
Chapter 23 G&GChapter 23 G&G
GluconeogenesisGluconeogenesis
Glycogen metabolismGlycogen metabolism

Announcements:
-
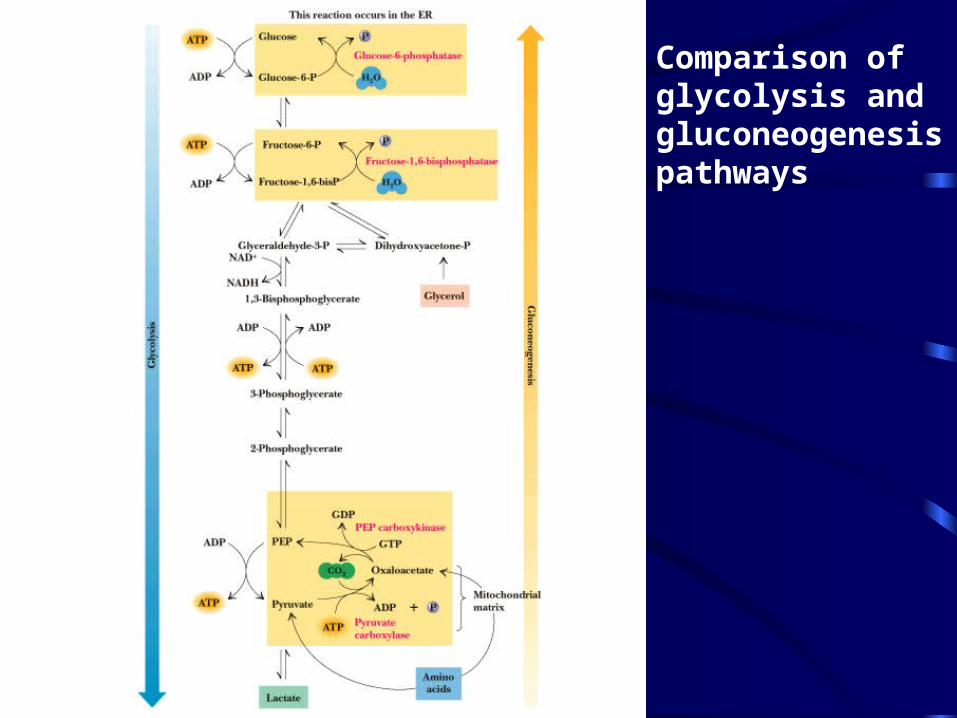
Comparison of glycolysis and gluconeogenesis pathways
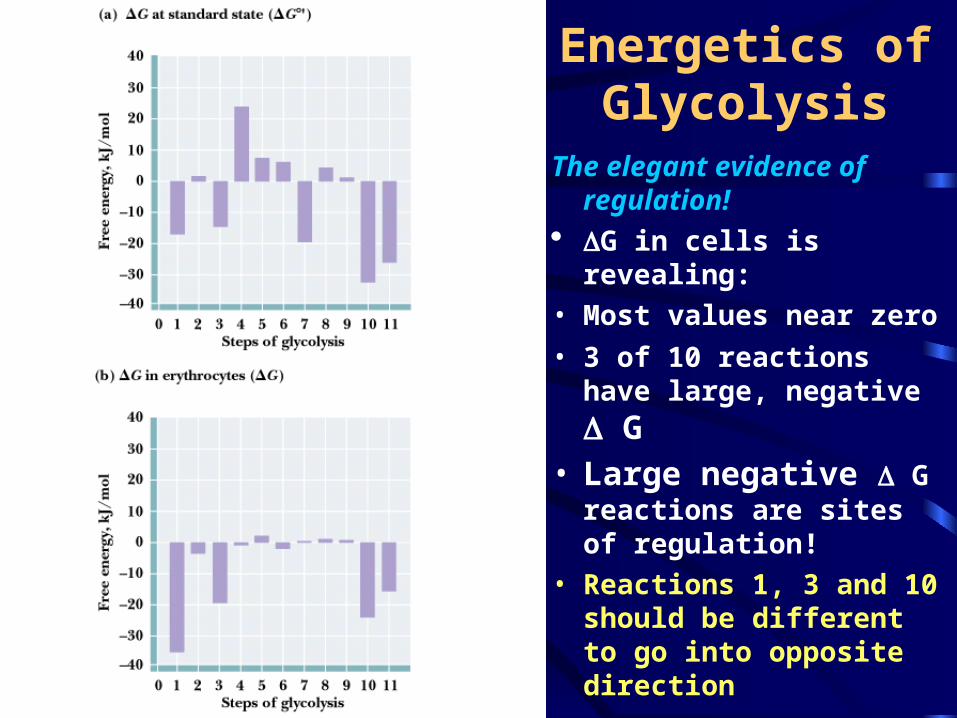
Energetics of Glycolysis
The elegant evidence of regulation!
G in cells is revealing:
• Most values near zero
• 3 of 10 reactions have large, negative G
• Large negative G reactions are sites of regulation!
• Reactions 1, 3 and 10 should be different to go into opposite direction

Gluconeogenesis Something Borrowed, Something New
• Seven steps of glycolysis are retained:– Steps 2 and 4-9
• Three steps are replaced:– Steps 1, 3, and 10 (the regulated steps!)
• The new reactions provide for a spontaneous pathway (G negative in the direction of sugar synthesis), and they provide new mechanisms of regulation

Pyruvate Carboxylase Pyruvate is converted to oxaloacetate
• The reaction requires ATP and bicarbonate as substrates • Biotin-dependent• Biotin is covalently linked to an active site lysine • Acetyl-CoA is an allosteric activator • Regulation: when ATP or acetyl-CoA are high, pyruvate
enters gluconeogenesis • The "conversion problem" in mitochondria
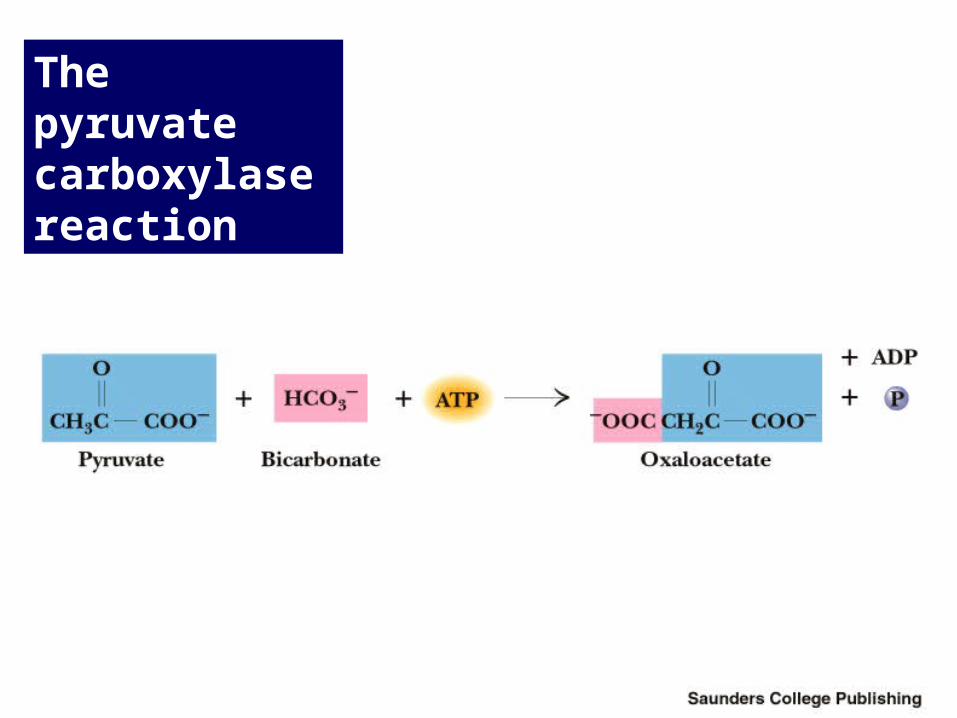
The pyruvate carboxylase reaction
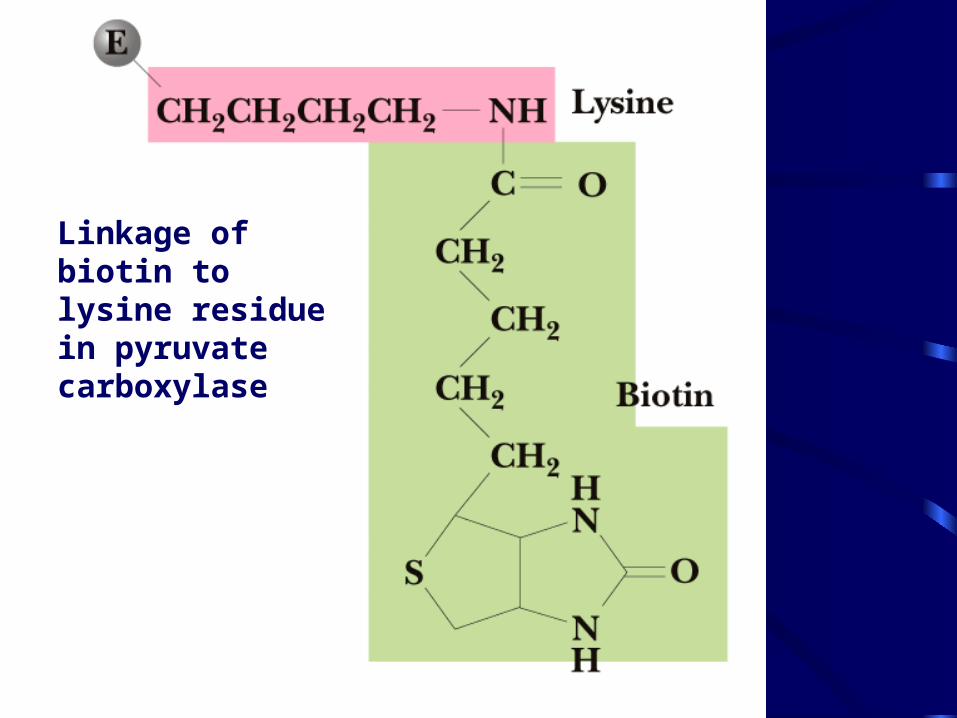
Linkage of biotin to lysine residue in pyruvate carboxylase
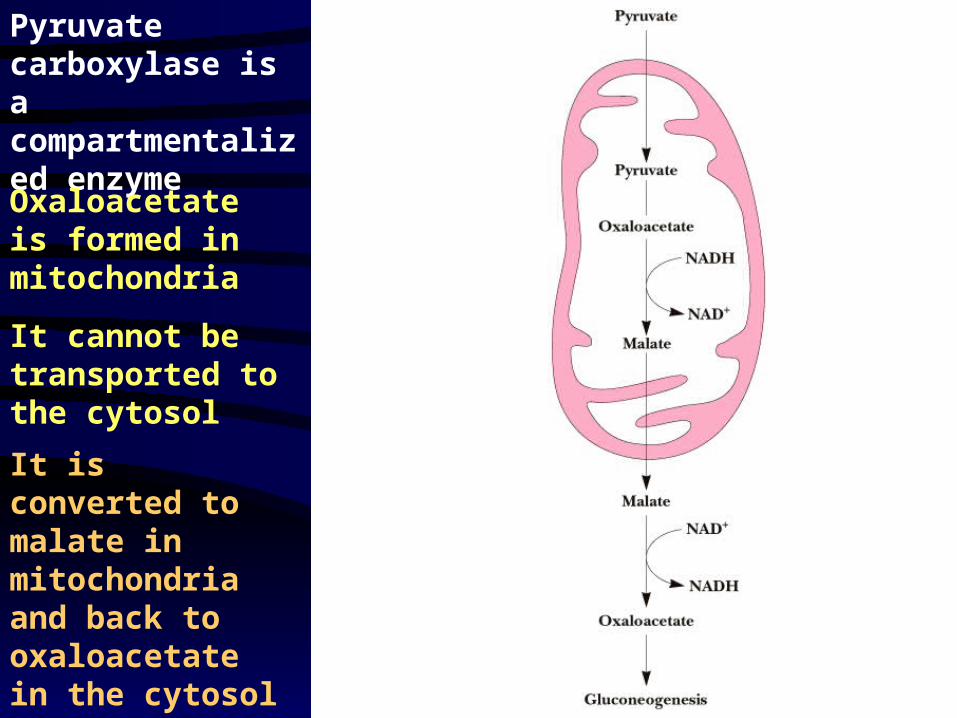
Pyruvate carboxylase is a compartmentalized enzyme
Oxaloacetate is formed in mitochondria
It cannot be transported to the cytosol
It is converted to malate in mitochondria and back to oxaloacetate in the cytosol

PEP Carboxykinase
Conversion of oxaloacetate to PEP
• Lots of energy needed to drive this reaction!
• Energy is provided in 2 ways:– Decarboxylation is a favorable reaction
– GTP is hydrolyzed
• GTP used here is equivalent to an ATP
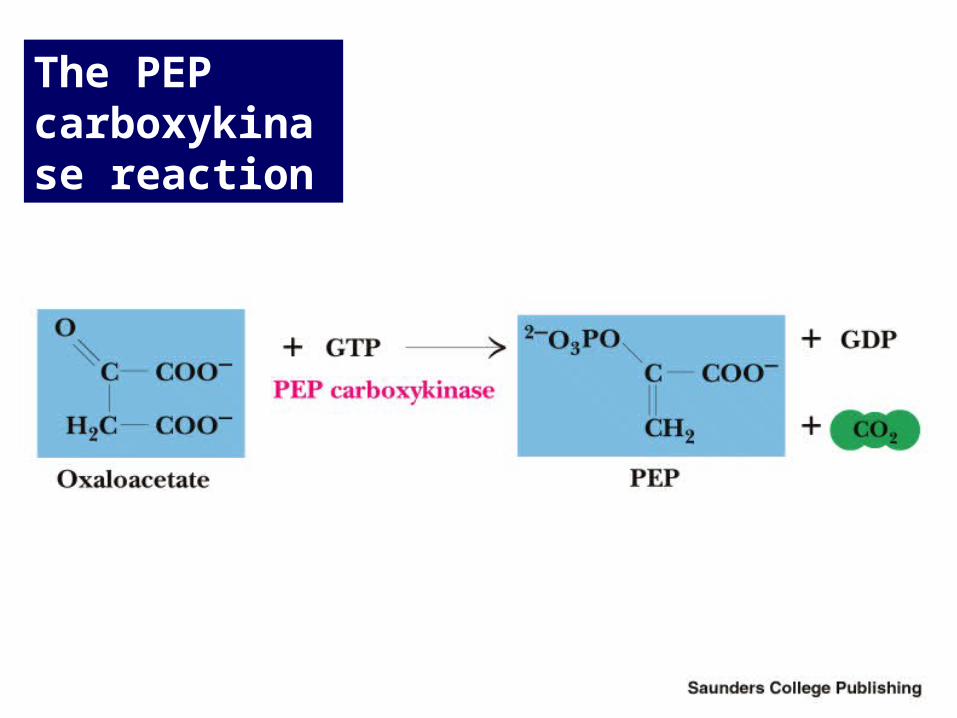
The PEP carboxykinase reaction

Fructose-1,6-bisphosphatase
Hydrolysis of F-1,6-P to F-6-P
• Thermodynamically favorable - G in liver is -8.6 kJ/mol
• Allosteric regulation:– citrate stimulates
– fructose-2,6-bisphosphate inhibits
– AMP inhibits
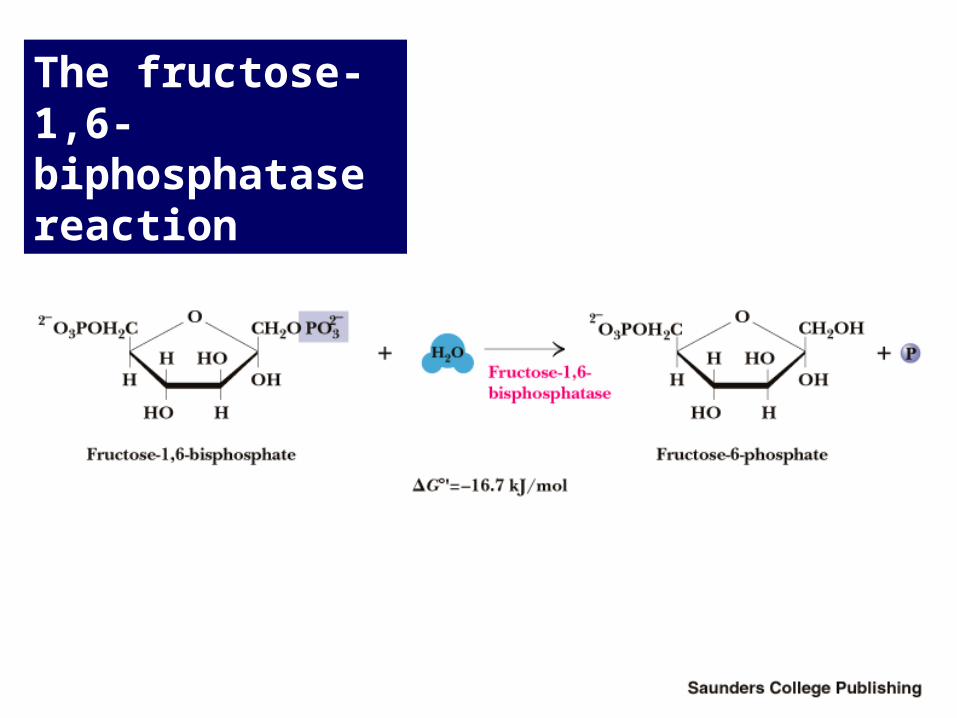
The fructose-1,6-biphosphatase reaction

Glucose-6-Phosphatase
Conversion of Glucose-6-P to Glucose
• Presence of G-6-Pase in ER of liver and kidney cells makes gluconeogenesis possible
• Muscle and brain do not do gluconeogenesis
• G-6-P is hydrolyzed as it passes into the ER
• ER vesicles filled with glucose diffuse to the plasma membrane, fuse with it and open, releasing glucose into the bloodstream
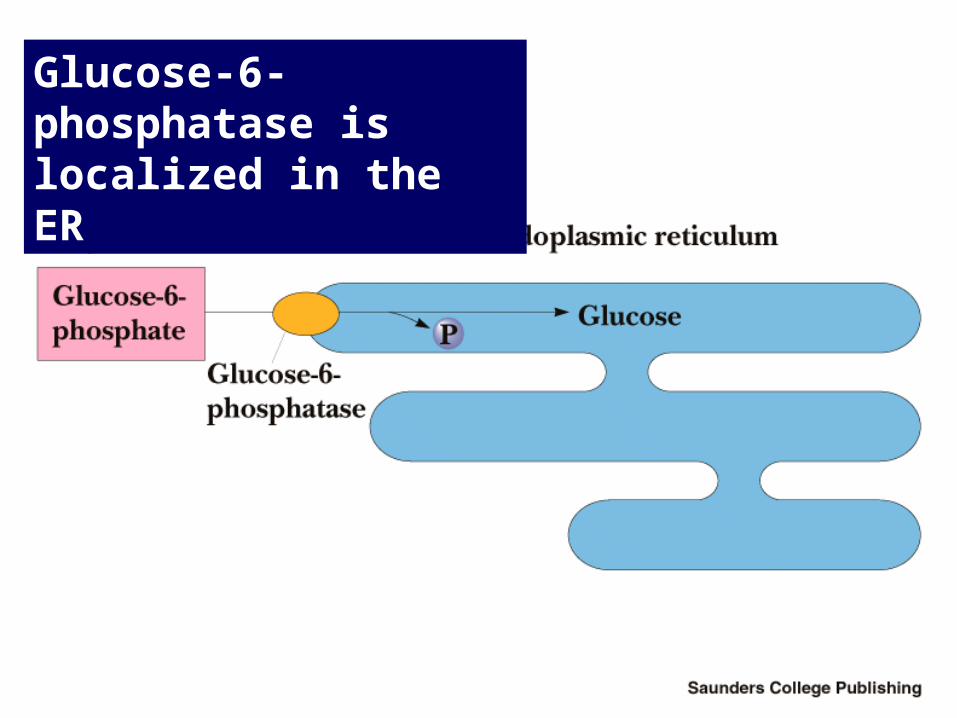
Glucose-6-phosphatase is localized in the ER

Lactate Recycling How your liver helps you during exercise....
• Vigorous exercise can lead to a buildup of lactate and NADH, due to oxygen shortage and the need for more glycolysis
• NADH can be reoxidized during the reduction of pyruvate to lactate
• Lactate is then returned to the liver, where it can be reoxidized to pyruvate by liver LDH
• Liver provides glucose to muscle for exercise and then reprocesses lactate into new glucose
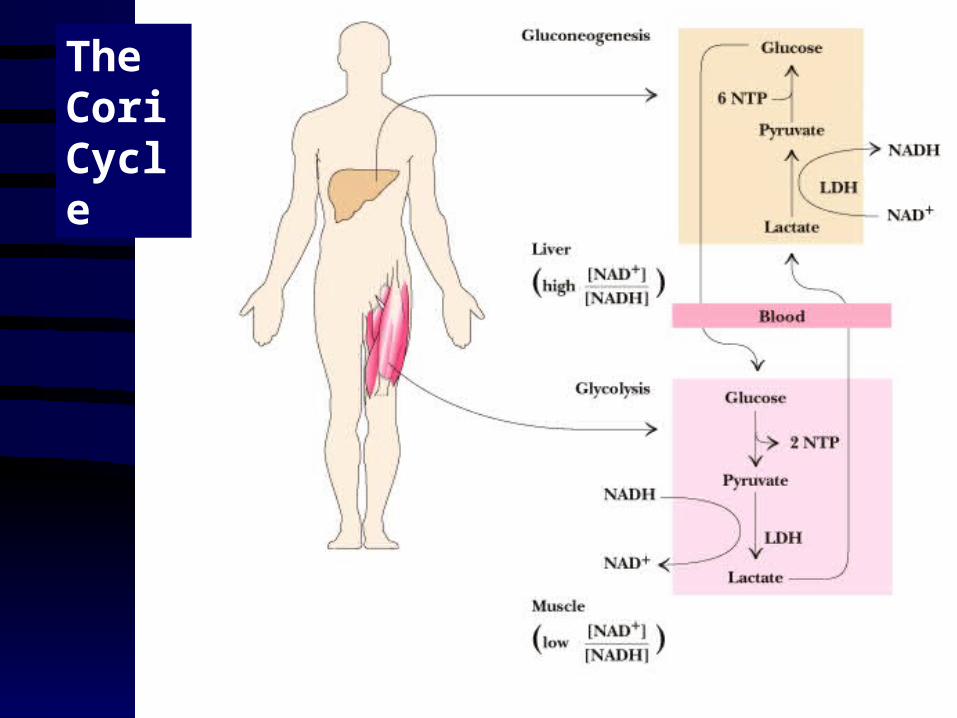
The Cori Cycle

Gerty and Carl Cori
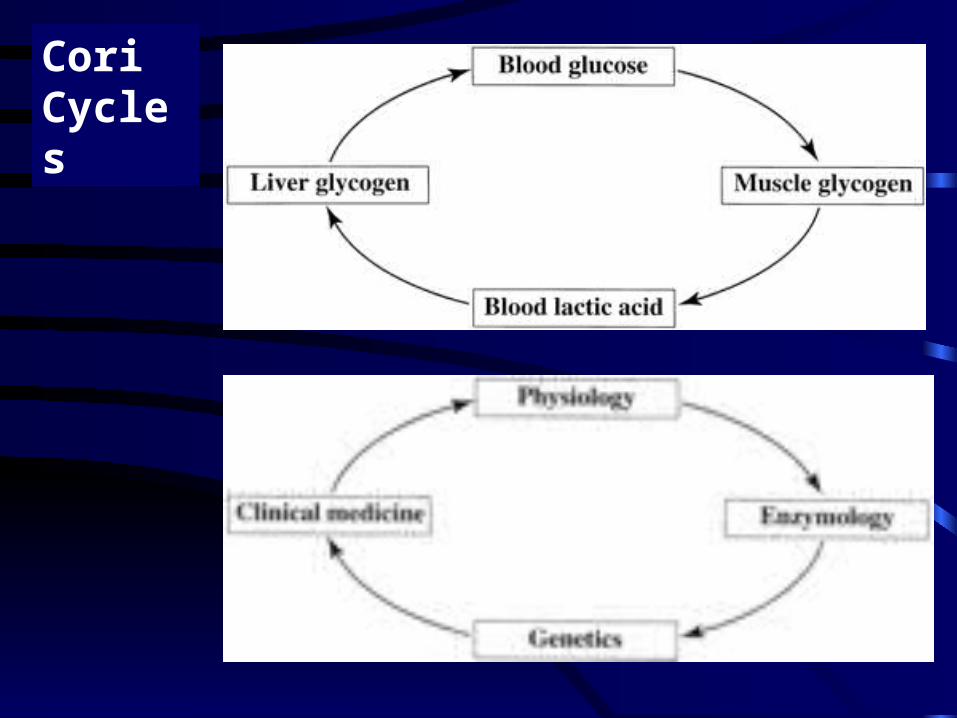
Cori Cycles

Regulation of Gluconeogenesis Reciprocal control with glycolysis
• When glycolysis is turned on, gluconeogenesis should be turned off
• When energy status of cell is high, glycolysis should be off and pyruvate, etc., should be used for synthesis and storage of glucose
• When energy status is low, glucose should be rapidly degraded to provide energy
• The regulated steps of glycolysis are the very steps that are regulated in the reverse direction!
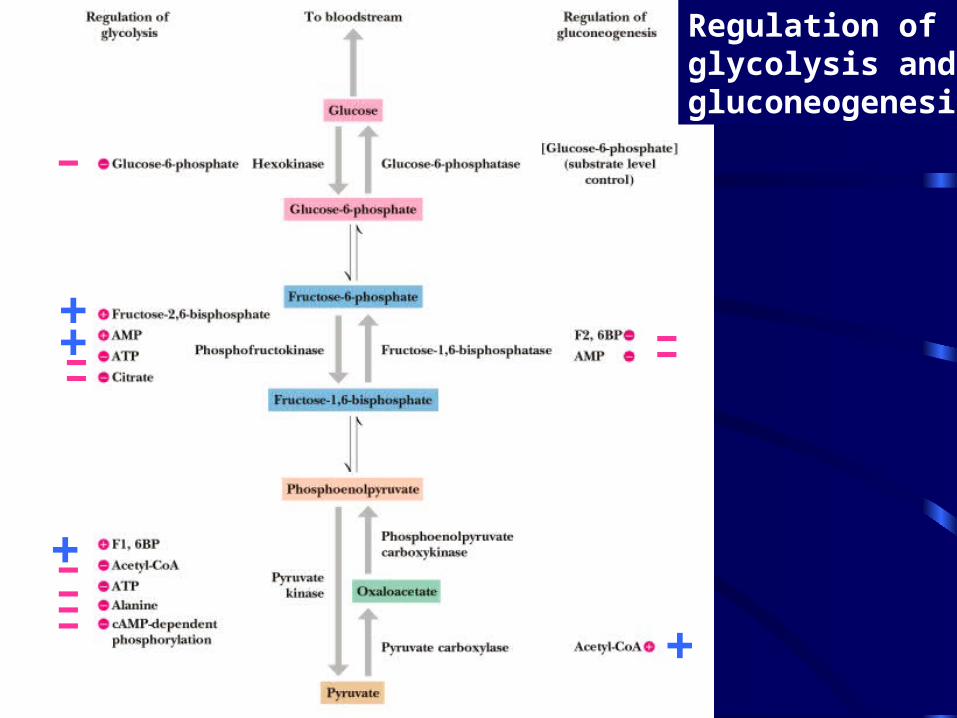
Regulation of glycolysis and gluconeogenesis
-
-
----
---
+
+
++

Gluconeogenesis Regulation II Allosteric and Substrate-Level Control
• Glucose-6-phosphatase is under substrate-level control, not allosteric control
• The fate of pyruvate depends on acetyl-CoA
• F-1,6-bisPase is inhibited by AMP, activated by citrate - the reverse of glycolysis
• Fructose-2,6-bisP is an allosteric inhibitor of F-1,6-bisPase
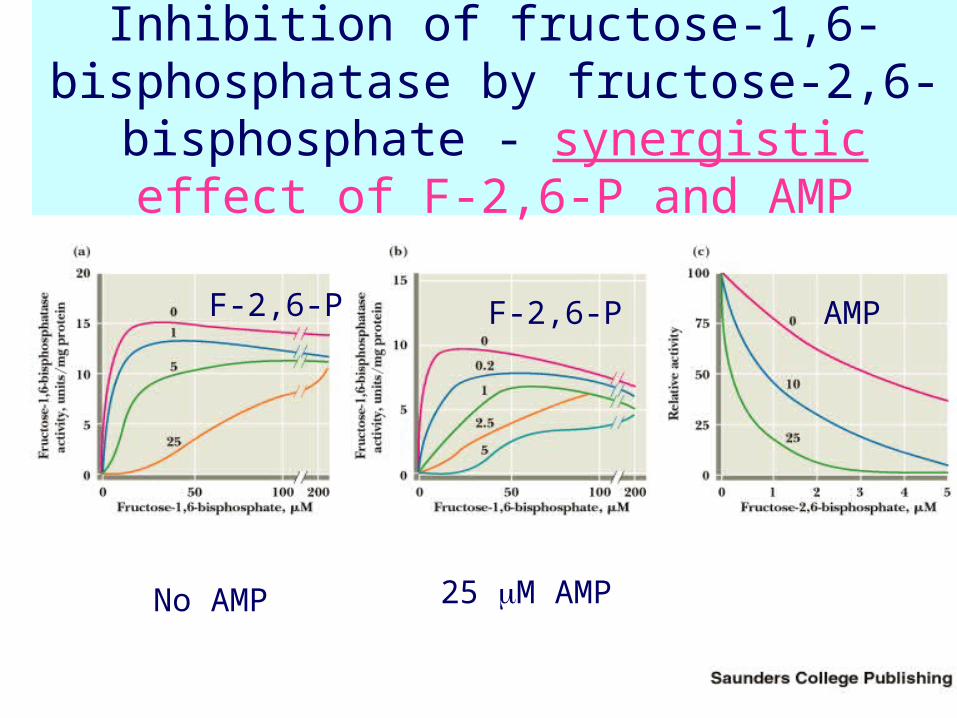
Inhibition of fructose-1,6-bisphosphatase by fructose-2,6-bisphosphate - synergistic
effect of F-2,6-P and AMP
No AMP 25 M AMP
F-2,6-P AMPF-2,6-P
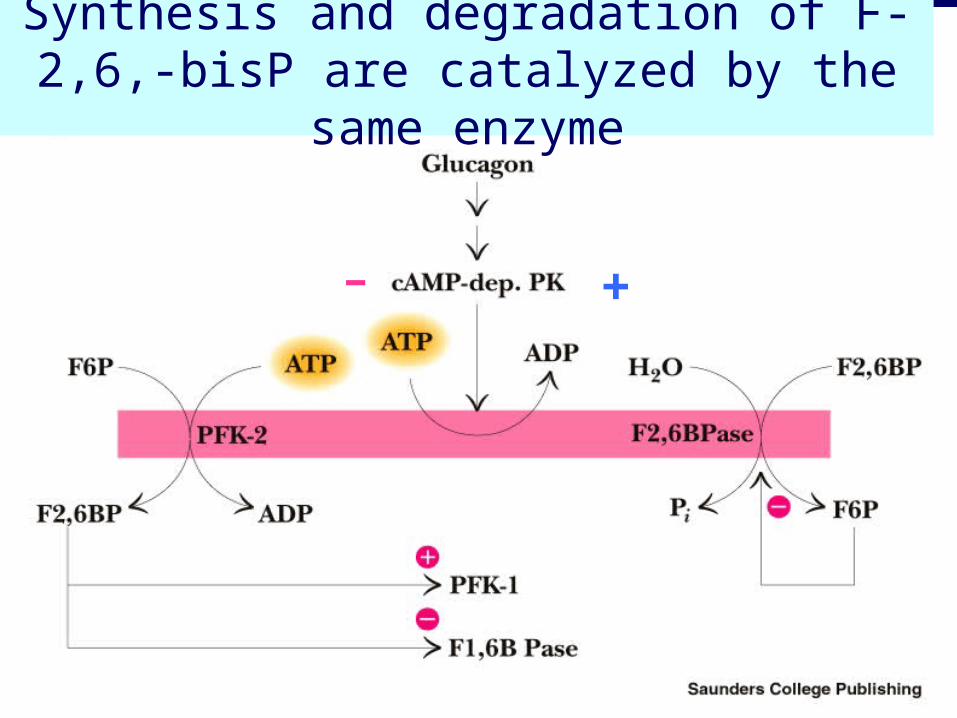
Synthesis and degradation of F-2,6,-bisP are catalyzed by the same enzyme
+-
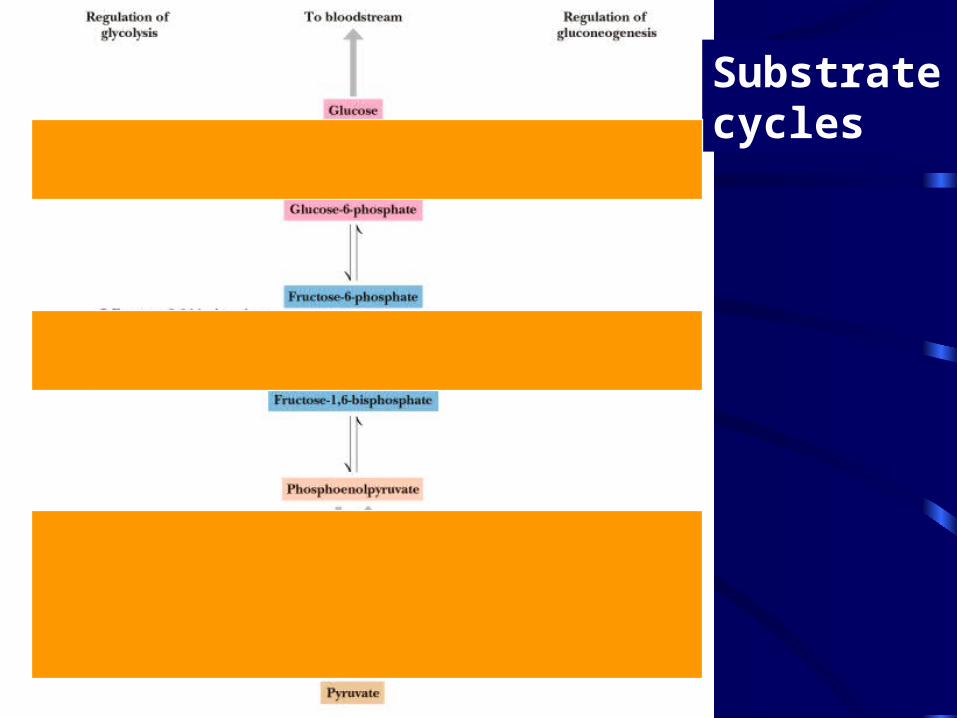
Substrate cycles
-

Substrate cycles• Simultaneous activity of Phosphofructokinase
(glycolysis) and F-1,6-bisPase (gluconeogenesis) yields a substrate cycle
• Reverse reaction decreases steady state flux through the pathway
• Could explain how 10% change in ATP concentration results in ~90-fold increase in the flux through glycolysis
• Synergistic Fructose-2,6-bisP / AMP / ATP / citrate (NAD+/NADH ratio, glucose-6-P, pyruvate, etc.) regulation provide alternative explanation

Substrate cycles• Three potential substrate cycles in glycolysis and
gluconeogenesis
• Example of Phosphofructokinase (glycolysis) and F-1,6-bisPase (gluconeogenesis)
• Reciprocal regulation does not work at high [F-1,6-P]
• Perhaps substrate cycling occurs only at high concentrations of F-1,6-P (PFK product) - - this prevents accumulation of excessively high levels of F-1,6-P
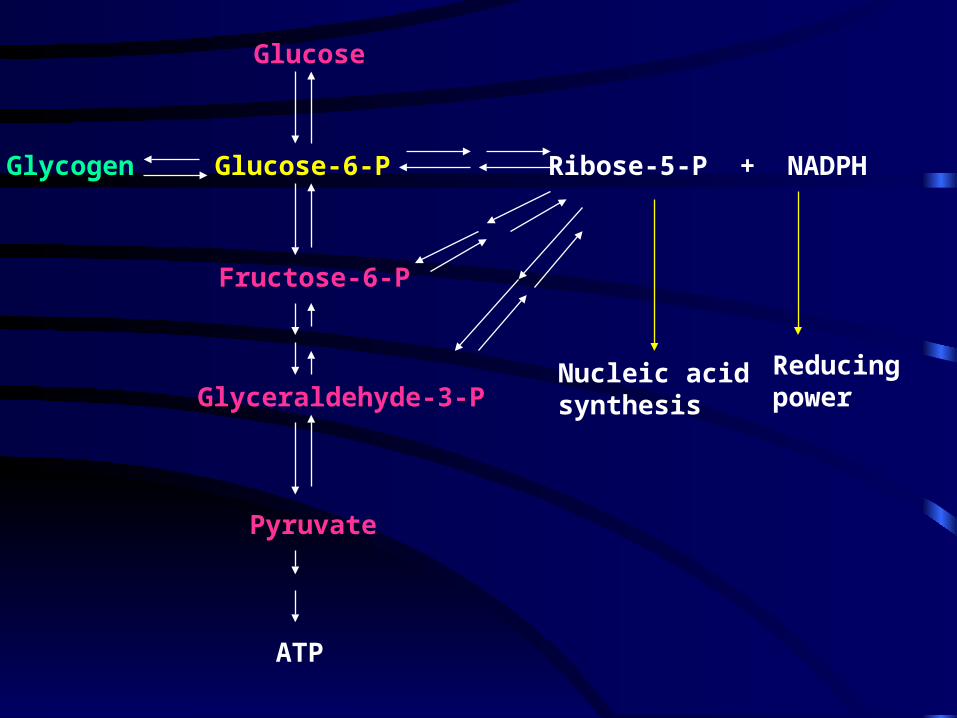
Glucose
Fructose-6-P
Glucose-6-P
Glyceraldehyde-3-P
Pyruvate
ATP
Glycogen Ribose-5-P + NADPH
Nucleic acidsynthesis
Reducingpower

23.3 Glycogen Catabolism Getting glucose from storage (or diet)
· Glycogen is a storage form of glucose -Amylase is an endoglycosidase
• It cleaves amylopectin or glycogen to maltose, maltotriose and other small oligosaccharides
• It is active on either side of a branch point, but activity is reduced near the branch points
• Debranching enzyme cleaves "limit dextrins"
• The 2 activities of the debranching enzyme
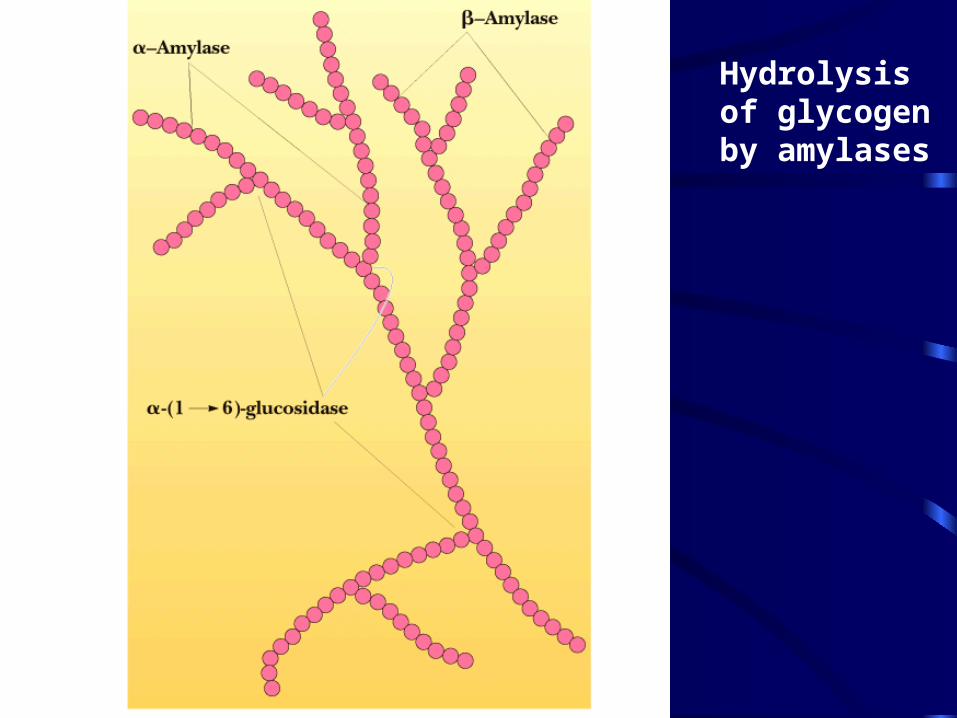
Hydrolysis of glycogen by amylases
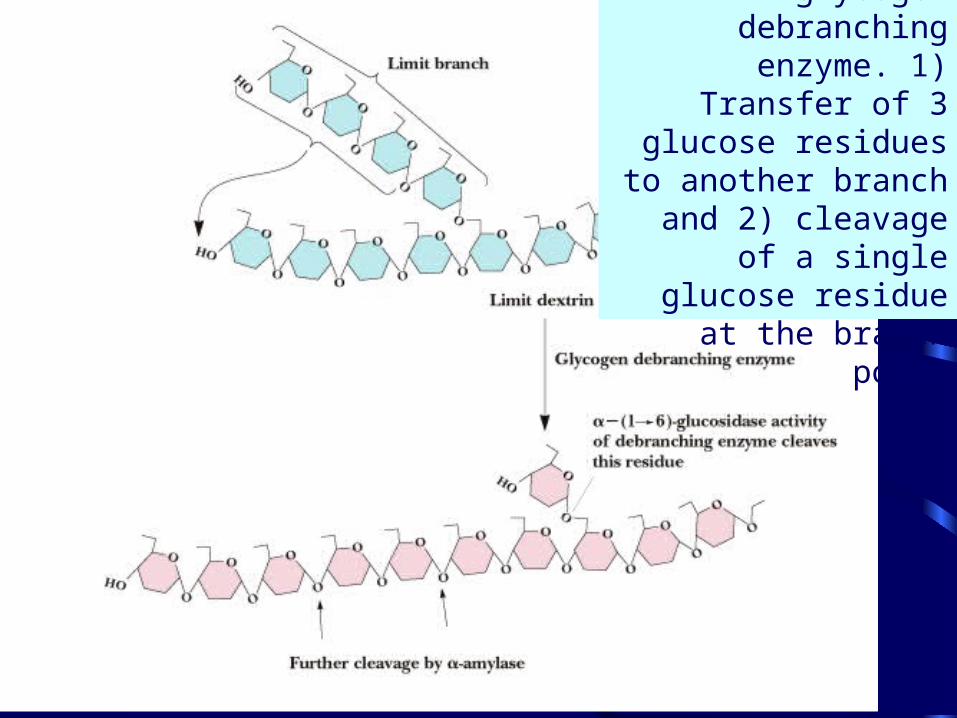
The reactions of glycogen debranching
enzyme. 1) Transfer of 3 glucose residues to
another branch and 2) cleavage of a single
glucose residue at the branch point

Metabolism of Tissue Glycogen Digestive breakdown is unregulated - 100%!
• But tissue glycogen is an important energy reservoir - its breakdown is carefully controlled
• Glycogen consists of "granules" of high MW
• Glycogen phosphorylase cleaves glucose from ends of glycogen molecules
• This is a phosphorolysis, not a hydrolysis
• Metabolic advantage: product is a sugar-P - a "sort-of" glycolysis substrate
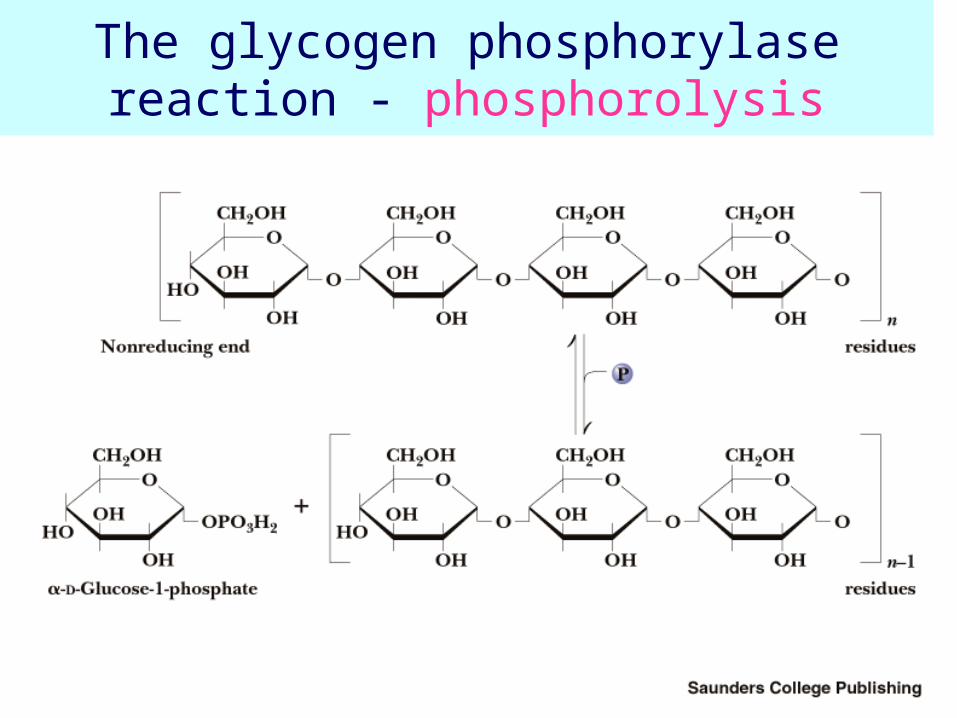
The glycogen phosphorylase reaction - phosphorolysis

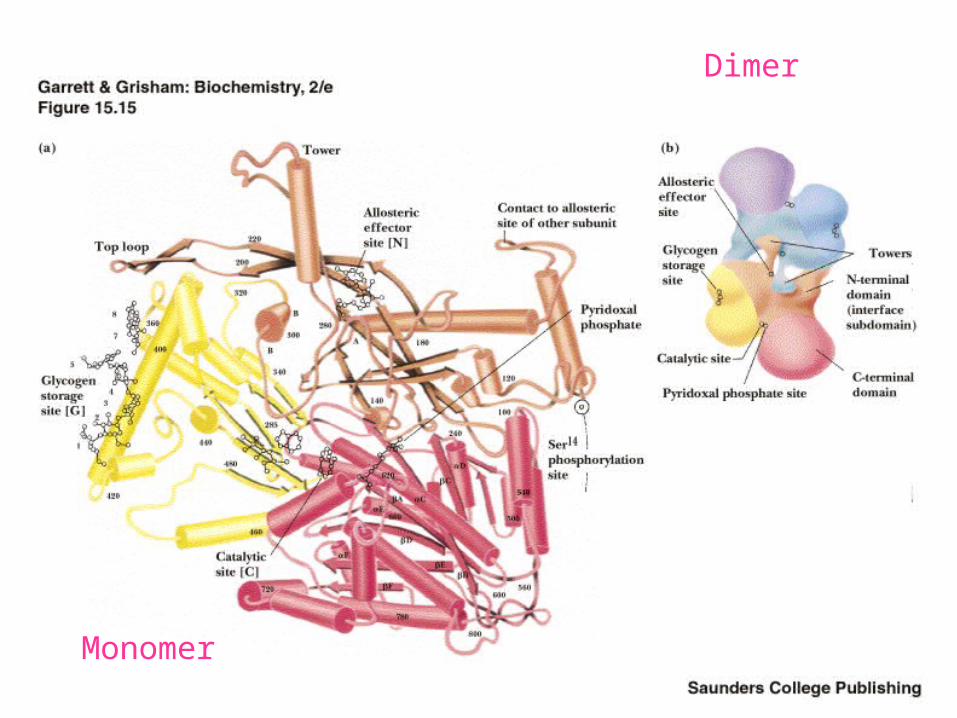
Glycogen Phosphorylase
A beautiful protein structure!
• A dimer of identical subunits (842 res. each)
• Each subunit contains a PLP, which participates in phosphorolysis
• Chapter 15
Dimer
Monomer