Embed Size (px)
Citation preview

BIOLOGICAL CONTROL OF BLACKLINE DISEASE OF ENGLISHWALNUT
GeorgeBruening,StephenD. Dauben, S.M. Mircetich,MindyBrooks, CharlesLeslie, GaleH.McGranahan,and AdibRowhani
ABSTRACT
Blacklineis an imponantdiseaseof graftedorchardwalnuttrees in California. The causalagent is cherry leafrollvirus (CLRV). The criticaldeleteriouseffectsof this pollen-bournevirus areinducednecrosisof cambiumand formationof cankersat the graft union. Generalintroductionintoorchardsof CLRV-resistantEnglishwalnutscions, if available,wouldprovidean excellentlongtermsolutionto the problemof blacklinediseasebecauseCLRVno longer wouldbe present in walnutpollen. Alternatively,rootstockresistantagainst,or tolerantof, CLRV will allow eveninfectedtreesto continuein production. Weare pursuingtwo coursesfor obtainingwalnutlines that possessgeneor geneswith the desiredphenotypes. The first approachgrew out of our observationthat selectionsof Paradox, designatedP 5/15, P 5/13, and P 7/34, demonstratedtoleranceto CLRVintroducedfromits Englishwalnutscion in a 10year test of two Tehemaand one Sunlandtree. New resultsshow thegeneralityof this toleranceto CLRV introducedfrom other Englishcultivars,and experiinentsinprogress are designedto test a new strategyfor the applicationof P 5/15, P 5/13, and P 7/34 tocontrollingblacklinedisease. In the secondapproach,potentialengineeredresistancegeneswereconstructedusing nucleotidesequencesfrom RNA 2 of CLRV. RNA 2 encodesthe coatprotein geneand a protein that probablyfacilitatesCLRVmovementin the host plant. RNA 2 also has a long(1.5 kb) 3'-untranslatedregion (3'-UTR). Two plasmids,one designedto expressthe coatproteinand pan of the movementprotein and a seconddesignedto generatethe complement("antisenseRNAIt)of the 3'-UTR, were found to be effectivein reducingthe accumulationof CLRVininoculatedprotoplasts,as comparedto resultswith controlplasmidsnot bearingthe CLRVRNA 2sequences. These constructionsare beingused to transformwalnutembryosfor productionof walnutshoots for test challengeby CLRV.
Obiective
Cherry leafrollvirus (CLRV),the causalagentof blacklinediseaseof graftedEnglishwalnut,is pollenborne and infectspistillateflowersof the scion. Spreadof CLRV in the infectedscion is ata rate of a few tens of cm'per year, and the infectionhas little or no apparenteffecton theappearanceor productivityof the scion. Blackand Paradoxrootstocksare resistantto CLRV.However, this resistanceis expressedin pan by a hypersensitivereaction(HR), i.e. by deathofinoculatedcells and necrosisat the graft union. As CLRVfrom the scion reaches rootstockcambiumcells, the resultingHR causesgirdlingof the tree in th~advancedstagesof the fatal blacklinedisease.
Our goal is to develoJ}control measures for blackline disease of grafted English walnutorchard trees through identification, improvement andlor engineering of
(i) English scions resistant against CLRV andlor
288- -- - - - --

(ii) rootstock tolerant of, or resistant against, CLRV, but without significantHR.
Becauseof the knownpropensityof virusesto mutateand overcomethe resistanceof potentialhosts, it is prudent to attemptto obtain morethan one geneticsourceof resistanceagainstanyeconomicallyimportantvirus.
In the "Procedure"and "ResultsandConclusion"sectionsbelowwe presentmaterialonengineeringresistanceagainstCLRV and thenon the exploitationof selectionsof Paradoxrootstockthat are tolerantof CLRV.
Procedure
Transient Expression of CLRV Coat Protein Gene and Complement of 3' UntranslatedRegion in protoplasts and demonstration of interference with CLRV
Back~round. Researchwith manyplantviruseshas shownthat the coat proteingene, whenengineeredto be part of the plant nuclearDNA, confersresistanceon the plant. That is, theproductionof virus coat protein in each cellof the plant, before those cells encounterthe virus,prevents or greatlydelaysvirus increaseafter inoculationof the usual virus dose. In other, lessextensiveresearch, includinga study in this laboratoryon tobaccoringspotvirus, the unttanslated3'portion (3'-UTR) of the virus RNA, whenappliedas an independentRNA molecule,was able tointerfere with virus replication. We report our investigationsdemonstratingthat a constructionexpectedto expressthe coat proteirigene and anotherdesignedto generateRNA complementarytothe 3'-UTR of CLRVinterferedwith CLRVincreasein tobaccoprotoplasts.
CLRVhas a genomecomposedof two large RNA molecules,RNA 1 of about7000nucleotideresidues (7 kilobases,Le., 7 kb) andRNA 2 of about6 kb. Analogywith other virusesstronglysuggeststhat the coatprotein gene will be encodedin RNA 2 and that the 3'-UTRs of thetwo genomicRNA moleculeswill be very similar in sequence,and our unpublishedresults confirmthis suggestion. The coat proteingene is not expectedto be expressedindependently. GenomicRNAs of membersof the nepovirusgroup, to whichCLRV is very similar, are translatedintopolyproteins,and it is the specificproteolyticcleavageof the polyproteinthat releasesfunctionalvirusproteins duringthe infectionprocess. Thus, the aminoacid sequenceof the CLRVcoat protein isexpectedto be synthesizedas part of a precursor"polyprotein.As reportedlast year, we determinedthe sequenceof bases in more than half of CLRVRNA 2 in order to deducethe aminoacid sequenceof a portion of the polyproteinand to revealhow far the 3'-UTR extendsfrom the 3' end of themolecule. We have not yet determinedthe exact locationof the coat protein aminoacid sequencewithin the polyproteinand the precise cleavagesites but have demonstratedwhatregionof RNA 2encodesthe coat proteingene. In the approachwe have taken, knowledgeof the precise boundariesof the coat protein gene is not needed. We express in protoplasts(and in future experimentsinwalnutshoots)the coat proteingene as a part of the carboxylterminalportionof the CLRVRNA 2fusionprotein. It is reasonableto expectthat subsequentinfectionby CLRV willgeneratethe CLRVproteinasethat is responsiblefor releasingcoat proteinfrom the completepolyprotein,therebygeneratingauthenticCLRV coatprotein from the CLRVRNA 2 sequencestransferredto the walnutgenome.
289

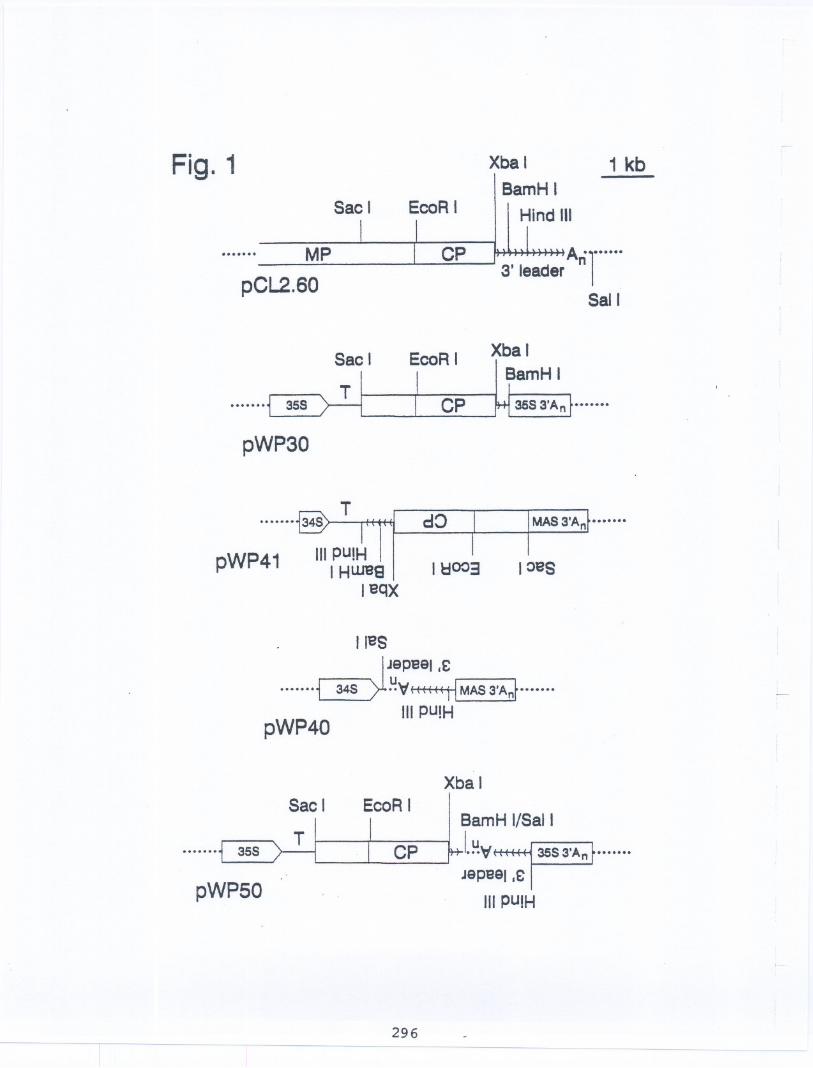
Constructionof Dlasmids. As reportedpreviously,we obtaineda 6.0 kb, nearlyfull lengthcopyDNA cloneof RNA 2 of CLRV,designatedpCL2.6O(Fig. 1). Nucleotidesequencesand invitro translationof transcriptsof truncatedcloneslocatedthe 3' end and probable5' end of the coatprotein gene on RNA 2. Our clone is designatedpCL2.6O(Fig. 1). Fig. 1 providesdiagramsof theplasmidsdesignedto providetransientexpressionof CLRV-targettedcoat protein and antisenseconstructionsbased on our clonedCLRVRNA 2 nucleotidesequence. The 34S promoterplasmidwas described, and comparisonsof the 34S promoterof figwort mosaicvirus was comparedwith the35S promoter of cauliflowermosaicvirus by Sangeret al., 1990. The plasmidpWP3O-Bis anAgrobacteriumplasmidused for transformationof walnutembryos. pWP30-Bhas the promoter,translationsignal, CLRVRNA 2 insert and 3' polyadenylationsignalof pWP30as well as genes forkanamycinresistanceand beta-glucuronidase(GUS). pWP50was constructedby subcloningthe3'-UTR region of pWP40so as to introducea BamHI site adjacentto the Sal I site. Cuttingat thisBamHI site and the BamHI site of pWP30allowedthe two plasmidsto be fused. The fusionplasmidwas the source of the insertof pWP50, whichwas releasedby digestionwith Sac I fortransfer to the Sac I site of plasmidvector pRTL to give pWP50.
Plasmidtransientexpressionand CLRVreplicationin protoplasts. Dr. WilliamJ. Lucas .
(UCD Botany) supplied the inoculum of Nicotiana tabacum suspension cell cultures. The protocolsfor tobacco protoplasts, as presented below, was extensively modified by Margaret Sanger and MindyBrooks from a protocol provided by Dr. Lucas, which in turn is based on a protocol for maizeprotoplasts from Virginia Walbot of Stanford University. All solutions were filter steriliZed.Suspension cell cultures were maintained in a rapidly growing steady state by transfer twice weekly inKCMS medium prepared from Murashige and Skoog minimal organics medium (Gibco-BRL no.510-1118) supplemented with 1.5 niM KHZP04' 0.2 mM 2,4-dichlorophenoxyacetic acid, 0.1 mMkinetin and 0.5 mg/l each of nicotinic acid, pyridoxine-HCI and thiamin-HCI, adjusted to pH 5.8
To prepareprotoplasts,cells were collectedby centrifugationand suspendedin a volumecorrespondingto 20 times the volumeof the suspensioncell cultureof 1% Cellulysin(Calbiochem,Inc., San Diego), 0.6% CellulaseRS (YakultHonshaCo., Tokyo), 0.4% MaceraseR-IO(Yakult)in370 mM mannitol, 12.5 mM sodiumacetate, 5 mM CaCl2(adjustedto pH 5.8 aceticacid; measuredosmolality0.45). The suspemionwas incubatedat 40 oscillationsper min at room temperaturefor 6to 7.5 hr. Protoplastswere filteredthrough 60 JJ. nylonmesh. An equalvolumeof wash solution(420 mM mannitol, 12.5 mM sodiumacetate, 5 mM CaClz,adjustedto pH 5.8 with aceticacid)wasadded, and protoplastswere collectedby low speedcentrifugationaridwashedtwicewith washsolution, all at room temperature. After the secondwash, protoplastswere countedin ahemocytometer. To the suspendedprotoplastswas addedan equalvolumeof cold electroporationbuffer (200 mM mannitol, 120mM KCI, 10 mM NaCI, 4 mM CaClz, 10mM sodiumHEPES,pH 7.2), and the protoplastswere collectedby centrifugation. Protoplastswere suspendedin ice-coldelectroporationbuffer at 1.5 x 1Q6/ml.0.8 ml of this solutionwas transferredto a HoefferPG 235-54 mrn electroporationcell, and 5 JJ.gof CLRV virionsor RNA and 10JJ.gof plasmidwere added.Electroporationwasfor 8 msecat 300V from760JJ.Fdcapacitors. Protoplastswere maintainedonice for 10 min, collected,and then suspendedin 10ml of protoplastmedium(0.60 vol of KCMS,0.20 vol conditionedmediumfrom suspensionculture, 0.05 vol coconutwater, additionalmannitoltoa final concentrationof 260 mM and KCMSto volume. Cephaloridinewas includedat 0.5 mg/ml.Protoplastswere maintainedat room temperaturewithoutshaking. In some experimentsprotoplastswere electroporatedat "minus 12hr" and at "zero time." For the secondelectroporation,protoplastswere collected, suspendedin 0.8 ml of cold electroporationbuffer, and electroporatedas describedabove.
290
---- - ------

At the times indicated,cells were collectedby centrifugation,suspendedin a volumeof0.50 M mannitolequal to 1000X the packedprotoplastvolume, and collectedagain. Cellsweresuspendedin 200 Itl TE (10 mM Tris-HCl, 1 mM N~EDTA, pH 7.4) and mixedwith 200 ILlof100mM Tris-HCl, 20 mg/mi sodiumdodecylsulfate, 2 mM N¥DTA and 10ILgof tRNA. Thesuspensionwas passedthrougha 21G syringeneedle6 times to reduceviscosity. ProteinaseK wasaddedto 100Itg/mI, and the suspensionwas incubatedat 37° for 30 min. Extractionwithphenol(equilibratedwith TE) and then with CHCl3followed. Sodiumacetatewas addedto 2SOmM, andnucleicacids were precipitatedby additionof 3 vol of ethanol. Nucleicacidswere dissolvedin waterand were treated with glyoxalaccordingto Maniatiset al. (1982). Electrophoresiswas through 1%agarosegel in 20 mM sodiumHEPES buffer, 1 mM N~EDTA. Blottingto HybondN+ nylonmembranewas with 50 mM NaOHover a periodof 3-4 hr. The membranewas rinsed with 300 mMNaCI, 30 mM sodiumcitrate, pH 7, and dried. Hybridizationwas accordingto a modificationof themethodof Church and Gilbert (1984).
Transformationof walnutembryosfor CLRVcoat orotein expression. The insertof pWP30(Fig. 1), including35Spromoter and polyadenylationsignal, was integratedintoan Agrobacteriumtumefacienstransformationplasmidto generatepWP30-B. Agrobacreriumvector pWP3O-B,carryinga gene for the CLRV coat protein, was used to inoculatesmall (1-5 mm) whitewalnut somaticembryosof the repetitivelyembryogenicline SU2. The Agrobacteriumstrain was grownovernightin523 mediumsupplementedwith 50 ILMgentimycin(TreatmentsA and B) or withoutgentimycin(TreatmentC). Bacteriawere then centrifugedto removethe growth mediumand resuspendedin oneof three incubationmedia:
a. DKW basal, 100 ItM acetosyringone,1 mM betainephosphate,pH 5.2
b. DKW basal, 100ItM acetosyringone,pH 5.2
c. 60% DKWbasal/40%523, 100ItM acetosyringone,pH 6.0.
Embryoswere incubatedin theseAgrobacteriumsolutionsfor 4 hours in 6 well multiwellsona rotary shaker at 100rpm. They were then removedfrom the liquid, blottedlightly to removeexcess inoculum,and plated to DKW basal mediumcontaining100 ItM acetosyringone. After 48hours, half the embryosfrom each treatmentwere transferredto DKW basal mediumsupplementedwith 100 mg/l kanamycinand half were transferredto mediumcontaining200 mg/l kanamycin.Table 1 summarizesthese treatments.
To select transformants,inoculatedEoembryoswere transferredweeklyon the twokanamycinmedia. E1embryoswhichemergedwere isolatedand culturedseparatelyon the same twomedia. Ezembryoswhich emergedfrom the El embryoswere then testedfor GUS activityusing theX-gluehistochemicalassay. Those that showedGUSactivitywere retainedand multipliedfor coatprotein detectionassays. Germinationand developmentof shoot culturesfrom each of theseEz linesis now in progress.
291
r--- --_ - --

Continued Testing of CLRV-Tolerant Rootstock, Selectionout of Paradox
Background. In earlier research, TehemaEnglishwalnutcultivarhad beengrafted ontoseveral selectionsof Paradoxand subsequentlyinoculatedwith CLRVby meansof bark patchesfromtrees knownto be infectedwith the walnut-8strain of the virus. Althoughmost of the graftunionsofthis set of trees beganto fail withintwo years of inoculation,due to blackline,the graft unionof thetree with ParadoxselectiondesignatedP 5/15, P 5/13, and P 7/34, as rootstocks,remainedfunctional10 years later. Onlya narrow, sharp hairlinehas developedat the graft union, with no indicationofcanker developmentin the Paradox5/15, P 5/13, or P 7/34 rootstock. The existenceof the narrow,hair-likeblack line is itself encouragingbecauseit suggestsa hypersensitivetype of resistantreactionagainstCLRV. This notion is supportedby ELISAof cambiumscrapings. Althoughthe TehemaandSunlandscion samplesgave strong ELISAsignalsusing anti-CLRVserum, the P 5/15, P 5/13 andP 7/34 samplesdid not give a detectedsignal.
Research in progress is designed to evaluate both the generality and the utility of theapplication of the three selections out of Paradox. The generality of the observed tolerant reactions ofP 5/152, P 5/13 and P 7/34 in graft unions with varieties of English walnut other than Tehema is .
unknown. Current technologydoes not allowroutine rootingof Paradoxselections,and vegetativeincreaseof the three selectionsfor use directlyas a rootstockwith unalteredgeneticcharacteristicsisproblematical. However,our experiencesuggeststhat variousParadoxselectionsreadilyarepropagatedby buddingon other Paradoxrootstockgerminatedfrom seed. Weare testingtheapplicationof P 5/15, P 5/13 and P 7/34 as interstockto protect rootstocksfrom the mostdevastatingeffects of CLRV.
Bark patch inoculations and evaluation. In June of 1988, sets of three bark patches wereapplied to two Chandler and three Ashly English walnut trees. In our report January, 1991, thearrangement of bark patches is indicated in Fig. lA of that report. The central patch of the set isfrom an English walnut tree infected with walnut-8 CLRV; the other two patches are from Paradoxselections: P 5/15, the selection under test, and P 3/27, a control known to develop blackline cankerswhen its English scion is infected by CLRV. Each set of the three bark patches was applied at threesites on each tree for a total of 15 replications of the CLRV inoculation. It is known from previousexperiments that CLRV will migrate both upward and downward from the infected bark patch withequal facility, so that the P 3/27 and P 5/15 patches can be expected to be exposed to approximatelyequal challenges of CLRV. .
Fig. IB of our January, 1991, report illustratesour interstockstrategyfor protectingrootstockgrafts from the effectsof CLRV infectionof the scion, in experimentsbegun in 1988. This strategy,if successful,will make the rootingof P 5/15 unnecessary. In 1989,more than 20 Paradoxseedlingrootstockswere buddedwith P 5/15 or, as a control, with P 3/27. In Augustof 1990the P 3/27 orP 5/15 stemof each tree was buddedwithTehema, Vinia, or ChandlerEnglishwalnutat a suitablesite 30 em to 60 em abovethe union withthe Paradoxseedlingrootstock. The result is theintroductionof P 5/15 (or P 3/27) as an interstockof 30 to 60 cm betweenParadoxseedlingrootstockand the Englishwalnutscion. The scionsof Tehema, Vinia, and Chandlerwere graft inoculatedwithbark patchesof CLRV-infected.walnuttrees in August, 1991. The graft unionsat the junctionsof theP 5/15 interstockand the Englishscionswill be examinedfor the presenceand severityof girdlingbyblacklinein the summerof 1992. The scions, interstocksand rootstockswill be assayedby ELISAfor the presenceand relativetiter of walnutstrain CLRV.
292-------.- -

Results and Conc1usions
Coat protein and 3'-UTR antisense plasmids capable of reducing the accumuJationofCLRV in inoculated protoplasts
The first three plasmidconstructionsof Fig. I, pWP30,pWP41,and pWP40,have beencompletedand tested in protoplastsderived from tobaccosuspensioncell cu1ture. PlasmidpWP50,which is to have a combinationof the inserts from plasmidspWP30andpWP40, is underconstruction. Our protoplastexperiments.place the RNA or proteinproductdirectedby a plasmidincompetitionwith a virus. The plasmidof necessitycan be expressedtransientlyonly, becausetheplasmiddoes not replicateand graduallywill be degradedin the electroporatedprotoplasts. CLRVwas inoculatedat a relativelyhigh concentrationto assureinfectionof a significantfractionof theprotoplasts, and, in contrastto the plasmid,CLRVincreaseswith time by replication. Thus even asmall interferenceby plasmidproductswith the replicationof CLRV is likely to be significantandcapableof exploitationusinga more robust expressionsystem.
Table 1 summarizesthe results obtainedwhenplasmidsand CLRVwere electroporatedintoprotoplasts. PlasmidpWP41is designedto be transcribedintoRNA complementaryto the coatprotein gene and surroundingsequences. This antisenseconstruction(Fig. 1) gaveno reductioninCLRV accumulationrelativeto the resultsobtainedwith the controlplasmidpRTL. Therefore,expressingany arbitraryCLRVRNA sequenceis not sufficientto obtaina reducedCLRVtiter.However, significantreductionsin CLRVaccumulationwere obtained,relativeto controlplasmids,for both pWP30, designedto expreSscoat protein, and pWP40, an antisenseconstructionof3'-untranslatedleader sequence(3'-UTR). An experimentwithpWP30is presentedin Fig. 2. Noindependenttest has beenmade to demonstratethat coat proteinor complementof the 3'-UTRactuallyare producedin the electroporatedprotoplasts. However, the vectorswe employhaveprovedto be effectivein this regardusing more easilyassayedreporter genes,both in our laboratoryand inthe laboratoriesof others. Checkson expressionwill be madefor constructionsexpressedin walnutembryosand shoots.
Thus we have two very promisingconstructions,pW30 andpW40. Currentlythe insertsfrom these effectiveplasmidsare being incorporatedintoone plasmid,pWP50 (Fig. I), and we areinitiatingexperimentswith a more powerfulexpressionsystemnow in use in the laboratory. As isindicatedbelow, the coat protein-expressingconstructionpWP30alreadyis beingmovedto walnutembryosand shoots, and we expectto have tests of this and other constructionswithinthe comingyear.
Transformation of walnut embryos for expression of CLRV coat protein
~ embryosthat grew on kanamycinand exhibiteda positiveGUSreactionwere recoveredfrom 45 separate~ embryos. Therefore, the 76 ~ linescurrently in cultureshouldcontainaminimumof 45 individualtransformationevents. In caseswhereduplicate~ lines derivedfrom asingle Eoembryohave been retained, additionalindependenttransformationeventsmay be present.
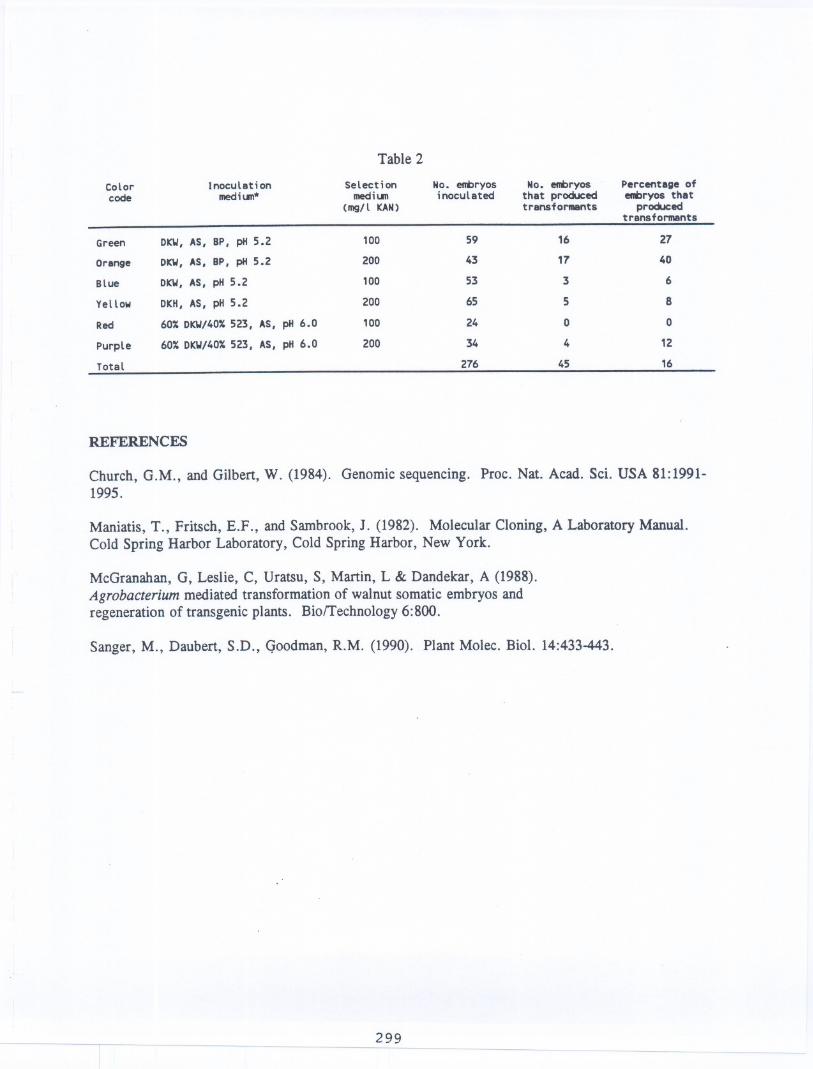
Inoculationin mediumcontainingImM betainephosphateappearedto greatly increasethetransformationfrequency(Table2). In all cases, more transformantswere recoveredwhenembryoswere selectedon 200 mg/l kanamycinthan whenselectedon 100mg/l kanamycin.
293
- --- - - - -

To date, embryoshavebeen germinatedand shoot culturesestablishedfrom Ezlinesderivedfrom 30 of the 45 transgeneratingEoembryos.
Evaluation of CLRV-Tolerant Rootstock
In our report of last year, results from one Ashlyand one Chandlertree (a total of 6 P 5/15and 6 P 3/27 patches)were reported. The necessarilydestructiveexaminationof patch graft unionsrevealedfailuredue to black line and cankerformationfor all P 3/27 patchesbut only blackhairlinesfor P 5/15 graft unions. Examinationof the remainingthree Ashleyand three Chandlertreesrevealedthat all P 3/27 bark patcheshad been lost to typicalblacldinecankerswhereasthe P 5/15bark patchesdevelopedonly hairline-likeblacldine. ApparentlyP 5/15 selectionout of Paradoxcontinuedto show a high degreeof toleranceto blacldinegirdling. Our results to date are veryencouraging with regard to the possibleapplication of P 5/15 as a practical solution to theproblem of blackline diseaseof grafted walnut cultivars.
Continuingon the plan presented in last year's report, we preparedinterstocktrees fromParadox rootstockplanted in December, 1988,buddedwith P 5/15 to generateinterstockin August,1989, and buddedto scionsof Tehema, Viniaor Chandlerin August, 1990. In Augustof 1991weinoculated20 trees of each sciontype with walnut8 strain of CLRVby meansof bark patches.Resultsfrom least some of the trees are likelyto be knownby late summer, 1992.
Encouragedby the results to date with P 5/15, we propagated320 P 5/15 on Paradoxseedlingsin April of 1991. In Augustof 1991P 5/15-on-Paradoxtrees were buddedapproximately12 inchesabovethe graft union, using 50 buds each from Chandler,Vinia, Tehema, HartleyandSunlandcultivars. These trees will be plantedout at permanentsites in the winterof 1992. TheEnglishscionswill be graft inoculatedwith the walnutblacklineCLRV in the summerof 1992. Thepreliminaryresults from this experimentare likely to be availablein the summerof 1993.
294
- - - --

Fig. 1. Derivativesof CLRV RNA2 clonednucleotidesequencetestedfor abilityto interferewith CLRV increasein inoculatedtobaccoprotoplasts. PlasmidpCL2.60 is a 6 kilobase(kb) clonethat has as its insertmost of the CLRVRNA 2 nucleotidesequence. The scale for 1 kb is indicatedin the upper right hand comer. Vector sequencesare indicatedby the dottedlines at eitherend of thediagram. CP =coat protein-encodingsequence. MP = sequenceof the putativeCLRVmovementprotein. The long, 1.5 kb 3'-untranslated(3'-UTR)sequenceis indicatedby multiplearrows. It isterminatedby the polyadenylate(AJ sequencethat formsthe 3' endof CLRVRNA 2. Locationsareindicatedfor cleavagesites of restrictionendonucleasesSacI, EcoRI, Xba I, BamHI, Hind m (allin the CLRVRNA 2 sequence)and Sal I (in the vector sequence). In the representationsof plasmidspWP41, pWP40and pWP50, sequencesinsertedin reverseorientation,in order to allowtranscriptionof RNA with polaritycomplemenaryto that of CLRVRNA 2, are presentedinvertedto emphasizereverse orientation.
PlasmidpWP30 is designedto express in plantcells the CLRVcoat protein fusedat its aminoterminal end to part of the movementprotein. pWP30has the Sac I to BamHI restrictionfragment from pCL2.60 inserteddownstreamfrom the translationalinitiationsite of tobaccoetch virus (T) and the cauliflowermosaicvirus 35Spromoterof plasmidvectorpRTL. Thecauliflowermosaicvirus polyadenylationsignal(35S3'AJ is providedat the 3' end of theinsen for pWP30. The controlplasmid,with all featuresexceptthe insened CLRVsequence,is pRTL,providedby Dr. JamesCarringtonof TexaxA&MUniversity. .
PlasmidpWP41 is intendedto direct in plant cells the synthesisof RNA that iscomplementary("antisense")to the coat proteingene of CLRVRNA 2 and surroundingsequences. pWP41has Sac I to Hind ill fragmentof pCL2.60 inserted intoplasmidpMS34Ain the reverseorientationto the orientationof the pWP30 insert. The vector and controlplasmid, pMS34A,was constructedby Dr. MargaretSanger. This plasmidhas a short (0.56kb) versionof the 34Sfigwon mosaicvirus promoter.
PlasmidpWP40 is designedto direct in plant cells the synthesisof RNA that iscomplementaryto the 3' leadersequence. pWP40has the Hind m to Sal I fragmentfrompCL2.60 inserted in invened configurationunder controlof the figwortmosaicvirus 34Spromoter, with downstreamMAS3' polyadenylationsignal. The control plasmid,pMS34B,has a long, 1.1 kb versionof the 34S promoter.
PlasmidpWP50 has the CLRV-derivedinsen of pWP40placedjust upstreamof the 35S3'polyadenylationsignal ofpWP30. pWP50is designedto take both the coat protein and 3'-antisenseapproachestoward interferingwith CLRVreplication.
295
pWP30
d~ I IMAS3'An~"..".
pWP41IIIPU!H
I HumSI I '=fOO3leqx
pWP40
CP
Xbal
BamH I/Sall
I.~\f~.".""Jepeel.€
III PU!H
Sac 1 EcoRI
pWP50
296- ---.-- -- -- -
Xbal 1 kbFig. 1 I
BamHISac I EcoR I Hind III
MP CP
3' leaderAnr....
.......
pCL2.60 Sail
EcoRI XbalSac I IBamHI
ILCP

123456789101112
I pRTL I pWP301
Mock . CLRV
== ---
o 12 24 0 12 24 0 12 24 0 12 24 hr .
Fig. 2. Analysisof CLRV RNAsin total nucleicacid extractsfrom inoculatedtobaccoprotoplastsand effectsof transientexpressionfrom plasmidpWP30. Aliquotsof protoplastsuspensionswere electroporatedat -12 hr with either controlplasmidpRTL (lanes7-9) or CLRV coatprotein expressionplasmidpWP30 (lanes 10-12),at 10 ILgof plasmidper 1.2 x 1()'Sprotoplasts. Ato time, protoplastsuspensionswere electroporatedwith CLRVvirions (lanes4-12), at 5 JLgof virionsper 1.2 x 1Q6protoplasts,or were electroporatedwithoutaddition(mockinoculation;lanes 1-3).Aliquotsfor extractionof nucleicacidswere taken immediately(lanes 1,4,7,10), at 12he(lanes2,5,8,11)andat 24hr (lanes3,6,9,12). Nucleicacidswerederivatizedandanalyzedbyagarosegel electrophoresis. The gel wasblottedand hybridizedwith a complementaryRNAprobetranscribedfrom the sequencethat correspondsto the HindIII-SalI restrictionfragmentof plasmidpCL2.60 (Fig. 1). This probe shoulddetect CLRVRNA 1 (upperband) and RNA 2 (lowerband)with equalefficiency. However, the probe does not have the CLRVRNA 2 sequencesthat are clonedin pWP30and thereforewill not detectpWP30or its transcripts.
297- - r- -- _ _ _ - ----- --- --

Table 1
Reducedaccumulationof CLRV in protoplaststransientlyexpressingCLRVRNA 2 sequences"
Plasmid StrategyTime after inoculation~
12 hours 24 hours
pWP40 antisense, 69%*3' leader
64%*
pWP30 coat protein 50%* 56%*
pWP4l antisense, 118%coat gene
102%
8Tobaccosuspensioncell protoplastswere electroporatedwith CLRVand with eitherthe indicatedplasmidbeaririga CLRVRNA 2 sequenceor the correspondingcontrolplasmid indicatedin the Fig. 1 legend. The amountsof CLRVRNA 1 and 2, asdetectedby hybridizationprobe and were assessedby quantitative,two-dimensionalbeta-particlescanningof the hybridizationblot. Backgroundradioactivity(fromthepositionsof RNA 1 and RNA 2 in lanesfrom uninoculatedprotoplasts)were no morethan 1.5% of the experimentalvaluesand were neglected. The radioactivitiesfromRNA 1 and 2 zoneswere summed. Resultsfor protoplastsreceivingthe experimentaland controlplasmidswere expressedas a ratio whichappearsas a percentagein thetable. * indicatesaverageof two experiments,the rangebeing from 10%to 20%.
b For pWP40andpWP4l,CLRVvirionsandplasmidwereco-electroporated.ForWP30, protoplastswere electroporatedwith experimentalor controlplasmid 12hrbefore electroporationof CLRVvirions at zero time.
- - -- -- --
REFERENCES
Church, G.M., and Gilbert,W. (1984). Genomicsequencing. Proc. Nat. Acad. Sci. USA 81:1991-1995.
Maniatis,T., Fritsch, E.F., and Sambrook,J. (1982). MolecularCloning,A LaboratoryManual.Cold Spring Harbor Laboratory,Cold SpringHarbor, New York.
McGranahan,G, Leslie, C, Uratsu, S, Martin, L & Dandekar, A (1988).Agrobacteriummediatedtransformationof walnutsomaticembryosandregenerationof transgenicplants. Bio/Technology6:800.
Sanger, M., Daubert, S.D., Goodman,R.M. (1990). Plant Molec.BioI. 14:433-443.
- - --- 299
Table 2
Color Inoculation Selection No. entIryos No. entIryos Percentage ofcode meetiun* meetiun inoculated that produced enmryos that
(mg/l KAN) transformants producedtransformants
Green DKW,AS, BP, pH 5.2 100 59 16 27
Orange DKW,AS, BP, pH 5.2 2DO 43 17 40
Blue DKW,AS, pH 5.2 100 53 3 6
Yellow DKH,AS, pH 5.2 200 65 5 8
Reel 60X DKW/40X523, AS, pH 6.0 100 24 0 0
Purple 60X DKW/40X523, AS, pH 6.0 200 34 4 12
Total 276 45 16





























