Embed Size (px)
Citation preview

Biologické účinky
ionizujícího záření
Zpracoval
Mgr. David J. Zoul

1
Veličiny používané v radiační ochraně
Podíl střední sdělené energie d̅ , kterou předalo ionizující záření látce v
daném objemovém elementu a hmotnosti dm tohoto elementu:
Nazýváme absorbovanou dávkou.
Tato veličina popisuje fyzikální efekty v dané látce, nelze ji tedy oddělit od
látky, ve které je stanovována.
Nelze tedy hovořit o dávce jako takové, musí být vţdy uvedena společně s
materiálem, např. dávka v měkké tkáni, dávka ve vzduchu atp.
Jednotkou dávky je Jkg-1
.
Tato jednotka dostala speciální název gray (Gy).
Ekvivalentní dávkou v orgánu či tkáni nazýváme veličinu:
kde DT,R je střední absorbovaná dávka v T-tém orgánu či tkáni způsobená
zářením druhu R a wR je tzv. radiační váhový faktor - viz tabulka 1
Druh a energetický rozsah záření wR
Fotony všech energií 1
Elektrony a miony všech energií 1
Neutrony s energií < 10 keV 5
10 keV aţ 100 keV 10
100 keV aţ 2 MeV 20
2 MeV aţ 20 MeV 10
> 20 MeV 5
Protony (s výjimkou odraţených), energie 2 MeV 5
Částice , štěpné fragmenty, těţká jádra 20
Tab. 1 - Radiační váhové faktory.
Protoţe radiační váhový faktor je bezrozměrný, je jednotkou ekvivalentní dávky
opět Jkg-1
.
)1(,dm
dD
)2(,, R
RTRT DwH

2
Aby se tato jednotka odlišila od jednotky dávky, pouţívá se pro ni speciální
název sievert (Sv).
Další důleţitou veličinou radiační ochrany je efektivní dávka.
Tvoří ji součet ekvivalentních dávek v jednotlivých orgánech či tkáních,
váţených tkáňovým váhovým faktorem wT, vyjadřujícím míru citlivosti
daného orgánu při celotělovém ozáření:
Hodnoty wT dle doporučení ICRP 60 jsou shrnuty v tabulce 2, přičemţ musí
platit:
Tkáň nebo orgán wT
Gonády 0,08
Červená kostní dřeň 0,12
Tlusté střevo 0,12
Plíce 0,12
Ţaludek 0,12
Močový měchýř 0,04
Mléčná ţláza 0,12
Játra 0,04
Jícen 0,04
Štítná ţláza 0,04
Kůţe 0,01
Povrch kostí 0,01
Mozek 0,01
Ostatní orgány a tkáně 0,12
Tab. 2 - Tkáňové váhové faktory.
Jednotkou efektivní dávky je opět Sv.
Limit efektivní dávky je primárním limitem v radiační ochraně.
Pro některé účely je uţitečné uvaţovat celkovou ekvivalentní či efektivní dávku
na vymezenou skupinu populace nebo na celou populaci.
)3(. T
TT HwE
)4(.1T
Tw

3
Příslušné veličiny se nazývají kolektivní ekvivalentní dávka ve tkáni či
orgánu - ST, resp. kolektivní efektivní dávka - S.
Jednoduše je lze stanovit jako součet ekvivalentních resp. efektivních dávek na
všechny jednotlivce v dané skupině.
Někdy můţe být zajímavé znát i její rozdělení podle dávky obdrţené
jednotlivcem.
Tak např. pro velké skupiny populace můţeme nahradit diskrétní rozdělení
efektivních dávek spojitou funkcí.
Pak bude (dN/dE)dE vyjadřovat počet jedinců, kteří obdrţeli efektivní dávku
mezi E a E+dE, a kolektivní efektivní dávku S dostaneme jako:
Dávka se vztahuje k předávání energie v daném místě, tj. z nabitých částic na
částice látky.
Jsou-li však primární částice nenabité, prvním krokem jejich interakce s látkou
je předání energie na nabitou částici.
Tento krok popisuje veličina zvaná kerma (Kinetic Energy Released in
MAtter).
Je definována coby podíl součtu počátečních kinetických energií dWk všech
nabitých částic uvolněných nenabitými částicemi ionizujícího záření v elementu
látky o hmotnosti dm a této hmotnosti:
Kerma je tedy opět definována ve vztahu k látce.
Z definice je zřejmé, ţe je pouţitelná pouze pro nenabité částice.
Jednotkou je stejně jako u dávky Gy.
Diferenciál dWk zahrnuje počáteční kinetické energie všech nabitých částic, to
znamená, ţe je v něm obsaţena i energie, která se později vyzáří ve formě
brzdného záření.
Interakce ionizujícího záření s živou hmotou
Absorpce energie záření X ve hmotě má kvantový charakter.
Pro energie primárních fotonů pouţívané v radiodiagnostice (24 – 140 keV)
připadají v úvahu celkem dva procesy, jimiţ můţe záření předávat svoji energii
látce.
Těmito procesy jsou fotoelektrický jev (fotoefekt) a Comptonův rozptyl
(Comptonefekt).
)5(.0
dEdE
dNES
)6(.dm
dWK k

4
Pro energie fotonů pouţívané v radioterapii (10 keV – 50 MeV) se začínají
uplatňovat i tvorba elektron-pozitronových párů a fotojaderné reakce.
Lineární přenos energie (LET) udává mnoţství energie předané
prostřednictvím výše zmíněných procesů látce na jednotku délky stopy částice.
LET určuje vztah mezi fluencí částic F a dávkou D záření:
LETD 16,0 [Gy] ( 7 )
Stopa částice je tvořena ionizacemi a excitacemi, které jsou produkovány
jednak primárním fotonem (tvoří jádro stopy), a jednak tzv. -elektrony,
vznikajícími při kolizi primární částice s obalem atomů a molekul látky,
majícími energii dostatečnou pro vznik dalších ionizací a excitací.
Tyto sekundární ionizace a excitace tvoří tzv. polostín stopy.
Tloušťka polostínu je dána doletem -elektronů R , který souvisí s energií
primárního fotonu [MeV] vztahem
4
7
40 ER [nm]. ( 8 )
Střední lokální dávka D ve stopě je nepřímo úměrná čtverci vzdálenosti od
trajektorie primární stopy.
Kvantový charakter přenosu energie záření hmotě vede k tomu, ţe i v
homogenně ozářeném vzorku lze nalézt mikroskopické objemy (objekty, místa)
s větším počtem ionizací a excitací, nebo naopak místa, kde k ţádnému předání
energie nedošlo, Mluvíme o fluktuacích energie deponované v
mikroskopickém objemu. Při náhodném rozdělení ionizací a excitací v
ozařovaném vzorku jsou fluktuace energie popsány Poissonovým rozdělením.
Tak např. pravděpodobnost vzniku k ionizací v mikroskopickém objemu, P(k),
je při střední hodnotě v ionizací v tomto objemu rovna
)exp(!
)( vk
vkP
k
( 9 )
a pravděpodobnost, ţe nevznikne ţádná ionizace, bude P(0) = exp(-v). Velikost
v bude očividně úměrná dávce záření. Tento vzorec má pro radiobiologii zásadní
význam, neboť vyjadřuje úbytek nezasaţených, tj. přeţívajících
mikroskopických objektů (buněk) v závislosti na dávce záření a často se v této
souvislosti mluví o tzv. zásahovém principu. Tento princip říká, ţe k dosaţení
biologického efektu je nutno zářením zasáhnout (jednou nebo několikrát)
určitou citlivou strukturu. Taková citlivá struktura v biologických objektech
vskutku existuje - je to molekula DNA. Zásah nemusí odpovídat jedné excitaci
nebo ionizaci, můţe to být také shluk ionizací, průchod částice citlivou
strukturou apod.
Řada teorií vychází ze struktury stopy částice. První model byl zkonstruován v

5
roce 1967 pro malé biologické objekty (např. bakterie, viry apod.), u nichţ bylo
moţné předpokládat, ţe jsou podstatně menší v porovnání se stopou částice.
O těchto objektech se předpokládalo, ţe mohou být v libovolném místě stopy,
přičemţ v intervalu vzdáleností od trajektorie (r, r+dr) se budou objekty
nacházet s pravděpodobností 2r.dr R-2
, kde R je poloměr stopy částice. V této
vzdálenosti od trajektorie částice bude střední lokální dávka deponovaná
-elektrony (D) úměrná přibliţně
.2
2
rE
ZD
( 10 )
Předpokládá se, ţe biologický účinek je zhruba stejný jako pro -záření při
stejné dávce, tj. pouţije se křivka přeţití pro řídce ionizující záření S(D).
Přeţití objektu zasaţeného stopou bude v průměru dáno integrálem
.2)( 2
0
drRrDS
R
( 11 )
Tato teorie dobře popisuje závislost biologického účinku záření na LET částic
pro malé a citlivé biologické objekty (biomolekuly, viry, citlivé bakterie).
Neplatí však pro rezistentní typy bakterií a savčí buňky. Aby se platnost rozšířila
na tyto objekty, zavedli autoři dva typy biologického účinku urychlených částic,
z nichţ první se vztahuje k jádru stopy částice a druhý k -elektronům. Efekt
můţe nastat buď zásahem biologického objektu jádrem stopy, nebo polostínem
stopy. Pro oba jevy se pouţívá jiná křivka přeţití a jiná parametrizace.
Na základě znalostí o struktuře stopy částice a s vyuţitím mikrodozimetrických
poznatků byly vytvořeny další teoretické přístupy, které interpretují nejen
letální, ale také mutagenní účinky pro různé druhy záření.
Účinky ionizujícího záření na celulární a subcelulární systémy
Biologické systémy obvykle obsahují vysoké procento vody, ve které se tedy
absorbuje značná část záření.
Radiolýzou vody vznikají vysoce reaktivní produkty, jako je hydroxilový radikál
OH, vodíkový radikál H
, hydratovaný elektron eaq a v přítomnosti kyslíku téţ
značné mnoţství peroxidu vodíku H2O2.
Všechny tyto produkty vyvolávají sekundární reakce s biologickými molekulami
včetně DNA, čímţ způsobují jejich poškození.
Tehdy hovoříme o Nepřímém účinku záření.
V lidských buňkách je informace obsaţená v DNA rozdělena do 23 párů
chromozomů.
Celý lidský genom obsahuje okolo 35 000 genů o celkové kapacitě 3109 BP.

6
Přímé působení stopy ionizující částice je na rozdíl od chemických
mutagenních činitelů charakteristické tím, ţe způsobuje ve vysoké míře zlomy
DNA, a to jak jednoduché – SSB (poškozeno je pouze jedno vlákno DNA), tak i
dvojité – DSB (poškozena je celá báze).
Kromě zlomů můţe docházet téţ k poškození např. chromatinového proteinu,
lokální denaturaci DNA apod.
Četnost poškozených bází můţe být dokonce ještě vyšší, neţ je počet
jednoduchých zlomů.
S rostoucí hustotou ionizace záření klesá výtěţek SSB a vzrůstá výtěţek DSB.
Četnost však ještě neukazuje na velikost biologického účinku daného radiačního
poškození.
V buněčném jádře totiţ fungují účinné reparační systémy.
Poškození na jednom řetězci DNA se poměrně snadno a s vysokou spolehlivostí
reparují s vyuţitím komplementární informace na řetězci druhém.
Také DSB se mohou reparovat s vyuţitím homologních chromosomů pomocí
tzv. rekombinační reparace.
Reparace DSB však probíhá podstatně pomaleji neţ reparace SSB.
Proporce nereparovaných DSB se zvětšuje při vyšším LET záření, z čehoţ
plyne, ţe kvalita DSB se můţe lišit.
V poslední době je stále více pozornosti věnováno tzv. komplexním
poškozením DNA.
Jedná se o poškození vzniklá jako následek vydělení většího mnoţství energie
záření ve formě clusteru.
Přesto, ţe četnost těchto jevů je nízká, mohou významně přispívat k výslednému
radiobiologickému efektu (jejich modifikovatelnost na fyzikálně – chemické
úrovni, jakoţ i reparovatelnost enzymatickými systémy je menší).
Vznik zlomů DNA je na chemické úrovni výsledkem konverze kyslíku na
peroxylový radikál, jenţ pak reaguje nevratně s radikálem DNA.
Dvojité zlomy DNA představují pro buňku závaţná poškození, která mohou vést
ke vzniku chromosomových aberací.
V interfázním jádře ozářených buněk lze pozorovat nejčastěji translokace a
fragmenty chromozomů.
Při obarvení určitého chromozomu se tyto aberace jeví jako tři skvrny (namísto
dvou) v jádře buňky.
V mitóze můţeme pozorovat mnoho různých typů aberací.
Vzhledem k tomu, ţe mitotické chromosomy obsahují vţdy dvě chromatidy, lze
rozlišit chromosomové a chromatidové aberace.
Nejčastějšími typy aberací po ozáření buněk ionizujícím zářením jsou zlomy
chromosomů, interchromosomální a intrachromosomální výměny (dicentrické
aberace, translokace, inverze apod.).
Chromosomové aberace jsou povaţovány za hlavní příčinu letálního účinku
záření a ve většině případů vedou k zániku buňky.
Závisí např. na přítomnosti kyslíku, dávce, fázi buněčného cyklu, druhu záření
apod.

7
Frekvence vzniku chromosomových aberací závisí pro záření na dávce
kvadraticky.
Velmi důleţitým typem chromosomálních aberací, zejména z hlediska radiačně
indukovaných onkologických onemocnění, jsou tzv. reciproké translokace.
Zlomy DNA a následné chromosomální aberace jsou většinou neslučitelné
s ţivotními funkcemi buňky.
Nejčastěji dochází k zániku buňky při pokusu o mitózu, kdy neproběhne zdárně
separace chromosomů do dceřiných buněk.
Zůstane tedy jedna buňka, která následně zahyne.
Některé typy aberací, jako jsou právě reciporké translokace však zachovávají
v buňce všechny geny ve funkčním stavu (s výjimkou místa zlomu).
Takové buňky se pak mohou dále dělit, i kdyţ jejich vlastnosti byly pozměněny.
Buňky s pozměněnými vlastnostmi představují pro organismus potenciální
nebezpečí, neboť poškození v oblasti některého protoonkogenu či
suporesorického genu můţe vésti k nádorovému bujení.
Z tohoto hlediska platí pravidlo „mrtvá buňka – dobrá buňka“.
Pokud vycházíme z poškození DNA a chromosomálních aberací, je evidentní, ţe
lineární dávková závislost počtu závaţných poškození náhodně rozdělených po
buněčné populaci povede k exponenciální (lineární v semilogaritmické bázi)
křivce přeţití.
Sebemenší nelinearita v dávkových závislostech v produkci chromosomálních
aberací bude mít za následek nelinearitu v počtu přeţivších buněk.
Zlomy DNA jsou produkovány lineárně s dávkou a jejich vzájemnou interakcí
vznikají chromosomální aberace.
Proto je závislost počtu aberantních buněk na dávce nelineární a křivka přeţití
vykazuje rameno.
Tento mechanismus tzv. reprodukční smrti buňky není však zdaleka jediný a
výsledná křivka přeţití je vţdy superpozicí mnoha různých procesů, jeţ se
odehrály v jednotlivých buňkách.
Kromě reprodukční smrti buňky se často setkáváme s pojmem „programovaná
smrt“ čili apoptóza.
Tento typ smrti se od nekrózy liší koordinovaným působením určitých enzymů
na počátku procesu.
Dochází ke shlukování chromatinu a štěpení DNA na fragmenty určité typické
délky působením endonukleáz.
Přestoţe apoptické buňky lze ojediněle nalézt i v normální buněčné populaci,
ozářením lze vyvolat v buněčné populaci masivní apoptózu.
Apoptóza indukovaná zářením je závislá na přítomnosti funkčního genu p53,
který nemusí být potřebný pro vyvolání apoptózy indukované jinými prostředky.
Programovaná smrt buňky je zřejmě součástí ochrany ţivého organismu před
nádorovým bujením, neboť podnětem k apoptóze je určitý typ poškození DNA.
Jestliţe buňka nedokáţe včas opravit vzniklé poškození, stává se pro organismus
rizikovou a je vhodnější tuto buňku odstranit a nahradit ze zdravé populace.

8
U nádorových buněk je právě gen p53 velmi často defektní, coţ je příčinou
vzniku nádoru.
Kromě letálních účinků způsobuje záření rovněţ vznik dědičných změn v ţivých
systémech, tj. změn v genetickém kódu, jeţ jsou předávány do následující
generace.
Velkou část těchto genových mutací tvoří u savčích buněk rozsáhlé deplece
genetického materiálu.
Změny vyvolané zářením v buňkách jsou často potenciálně reverzibilní.
Uţ na úrovni prvotních radikálů dochází k rekombinaci, která je tím
intenzívnější, čím vyšší je hustota ionizace.
Také vodní radikály se mohou rekombinovat, čímţ se sniţuje nepřímý účinek
záření.
Poškození nukleotidických bází a určité typy jednoduchých zlomů DNA mohou
být opraveny tzv. excizní reparací.
V průběhu této reparace dochází k rozpoznání a vyštěpení poškozeného
fragmentu DNA endo- a exonukleázou.
Chybějící úsek pak resyntetizuje DNA-polymeráza.
Sloţitější poškození buňky indukují aktivaci tzv. SOS-systému.
Důleţitou úlohu při indukci SOS-systému plní RecA-protein, jenţ má několik
aktivních center a je důleţitým reparačním enzymem.
Při radiačním poškození DNA se RecA-protein mění na RecA-proteázu, která
štěpí LexA-protein, který je represorem genů SOS-systému, RecA-proteinu a
sebe sama.
Rozštěpením LexA represoru se spustí transkripce SOS-genů a následná syntéza
celé řady důleţitých proteinů, umoţňujících poškozené buňce přeţít.
Dochází k obnovení syntézy DNA zablokované poškozením, tím, ţe se v místě
poškození vkládá do řetězce DNA nejpravděpodobnější chybějící nukleotid či
dokonce sekvence nukleotidů.
Indukce SOS-reparace vede k většímu objemu reparovaných úseků, ale za cenu
niţší spolehlivosti, a tím i většího počtu mutací.
Proto se také někdy hovoří o mutagenní reparaci.
Pravděpodobně nejproblematičtější je pro buňku reparace DSB, neboť je zde
ztracena informace na obou řetězcích DNA.
I tyto zlomy se však reparují, a to mechanismy zahrnujícími tzv. rekombinační
procesy.
K uskutečnění této reparace je za potřebí homologní sekvence DNA, která se
pouţije jako templát pro rekombinaci.
Důleţitou roli při reparaci mají tzv. kontrolní body buněčného cyklu, v nichţ
se progrese v cyklu při poškození DNA zastaví, a tím je poskytnut čas k reparaci
poškození.
Tak např. je známo, ţe záření zablokuje buněčnou populaci v G2/M-fázi.
Zpoţdění vstupu do mitózy, které tím vznikne, vzrůstá s dávkou a je podstatně
větší pro záření s vysokou hustotou ionizace.

9
Poškození lidského organismu ionizujícím zářením
Pochopení patogenetických mechanismů uplatňujících se při vzniku a rozvoji
somatického a genetického poškození člověka vlivem působení ionizujícího
záření vychází z poznatků o účincích záření na savčí buňku, popsaných výše.
Lidský organismus je ovšem sloţitým souborem tkání s často velmi
rozmanitými biologickými vlastnostmi.
Regulační systémy nervové i humorální představují na rozdíl od pouhých
buněčných kultur kvalitativně novou úroveň.
Lidský organismus prochází během ţivota různými fázemi svého růstu a vývoje,
během kterých se můţe odezva na ozáření měnit.
U člověka se mohou projevit účinky ozáření aţ po několika desetiletích, na
druhé straně existují jiné typy radiačního poškození, jeţ se mohou v průběhu
delší doby reparovat mechanismy vyvinutými aţ u vyšších organismů.
Zářením modifikovaná cytogenetická informace (mutace) můţe i při zachované
schopnosti dělení zasaţených buněk představovat selektivní nevýhodu, jeţ se
projeví aţ v průběhu více generací eliminací zasaţeného genu z populace.
Vedle toho můţe ozáření vésti k neschopnosti některých buněk realizovat své
potomstvo, tj. projít opakovanými mitózami.
Např. po jednorázovém ozáření červené kostní dřeně odpovědné za krvetvorbu
dávkou 1Gy přeţívá 37 % z původního počtu buněk.
V důsledku deplece proliferujících kompartmentů je přísun nových buněk do
periferní krve omezen nebo zastaven.
Funkční erytrocyty v periferii však nejsou v principu poškozeny.
Důsledky blokády dělení prekursorů se tedy projeví aţ po delší době latence,
kdy absence náhrady přirozeně ubývajících erytrocytů vyvolá klinicky patrné
důsledky.
Konečný výsledek jednorázového ozáření není přitom určen jen velikostí
bezprostředních buněčných ztrát v důsledku expozice, ale i následným
uplatněním reparačních procesů.
Tzv. reparace Elkindova typu se projevuje obnovením ramene na křivce
přeţití, coţ lze interpretovat tak, ţe buňky identifikované bezprostředně po
ozáření jako reprodukčně mrtvé znovu získaly schopnost proliferace.
Dalším významným mechanismem nápravy je obnova buněčné populace cestou
proliferace ze zachovalé frakce kmenových buněk.
Buněčnou proliferaci vyvolávají či zastavují extracelulární regulátory působící
prostřednictvím specifických receptorů na buněčných membránách.
Patří k nim cytokiny peptidické povahy s převáţně parakrinním či autokrinním
působením.
Vedle zevních regulátorů se stimulace buněčného cyklu realizuje
intracelulárními mechanismy, např. působením cyklinů.
V modelu radiačního poškození červené krvetvorby je průběh regenerace
progenitorů této buněčné řady ukázán na obr.1.

10
Bezprostředně po ozáření dávkou 2 Gy se fond těchto buněk zmenšuje asi na
10 % a poté působením extracelulárních regulátorů opět stoupá.
Kolem 10. dne dosahuje původní velikosti kompartmentu.
Pro nedostatečné zpětnovazebné tlumení regenerace přestřelí, a teprve poté se
velikost kompartmentu stabilizuje.
Analogické modely lze konstruovat také pro sebeobnovu dalších elementů
periferní krve, tj. leukocyty a trombocyty, které se v rozvoji akutní nemoci
z ozáření především uplatňují.
K dalším sebeobnovným buněčným populacím, které jsou zachovány po celou
dobu ţivota jedince, patří slizniční výstelky, pokoţka s adnexy a v období
pohlavní dospělosti rovněţ gonády.
Tyto populace se liší délkou buněčného cyklu a dobou průchodu od
nediferencované mateřské buňky k buňce s plnou funkcí.
Nejkratší je tato doba u epitelu tenkého střeva, kde činí asi 4 – 5 dnů, delší
v buňkách bílé krvetvorby s efektivní dobou průchodu 10 – 20 dnů a u
spermiogeneze je rozloţena na dobu 6 – 8 týdnů.
Tyto časy jsou relevantní ve vztahu ke klinické manifestaci projevů po ozáření.
V některých buněčných systémech se buňky proliferujících kompartmentů
nalézají v klidovém stavu, ale jsou schopny v mimořádných situacích přejít do
buněčného cyklu.
Patří sem např. jaterní tkáň, parenchym ledvin či kloubní chrupavky.
V jiných buněčných systémech je sebeobnovná buněčná populace přítomna jen
do období fetálního, popřípadě novorozeneckého a v pozdějším období
reprezentují daný systém jiţ jen buňky funkční.
To se týká např. neuronů centrálního nervového systému, jeţ jsou v důsledku
této své charakteristiky málo citlivé k působení ionizujícího záření.

11
Funkční stav centrálního nervového systému je však podmíněn zachováním i
dalších struktur, jako např. cévního řečiště.
Vyšší vnímavost endotheliálních buněk na ozáření můţe pak být pro konečný
výsledek ozáření rozhodující.
Poněkud odlišná je situace ţenských zárodečných buněk.
Zrání oocytů se zastavuje u novorozenců na úrovni profáze prvního zracího
dělení a vytváří se primární folikul.
U dospělé ţeny se v průběhu menstruačních cyklů dokončuje zrání primárních
folikulů a uvolňuje se zralé vajíčko.
V době dospívání má ţena asi 300 000 – 400 000 primárních folikulů.
Jejich počet klesá přirozeným zánikem do 30 let asi na 20 %, do 45 let na cca.
2 %.
Zánik primárních folikulů způsobený ozářením nemůţe být nijak nahrazen.
Dávky okolo 3 Gy způsobují v závislosti na typu frakcionace u mladších ţen
sterilitu v 60 – 70 %, u starších ţen ve 100 %.
Pro komparativní posuzování účinků ionizujícího záření na biologické systémy
je zaveden termín radiosenzitivita.
V experimentální radiobiologii se radiosenzitivitou rozumí strmost křivky přeţití
buněčných systémů, popisovaná klasicky hodnotou střední letální dávky D0 .
Buněčné systémy, vykazující strmě klesající křivky přeţití a charakterizované
niţší hodnotou D0 mají vyšší radiosenzitivitu neţ systémy vykazující pozvolný
pokles křivky přeţití se stoupající dávkou.
Pokud hovoříme o radiosenzitivitě jednotlivých tkání lidského těla, máme tím na
mysli zpravidla vnímavost k vyvolání akutních klinických příznaků.
Ke vnímavým systémům patří z tohoto hlediska lymfatická tkáň, červená kostní
dřeň, epitel tenkého střeva i některých jiných orgánů a muţský pohlavní epitel.
Velmi citlivým orgánem je i oční čočka, ale patogeneze rozvoje oční katarakty
(šedého zákalu čočky) vyvolané zářením je sloţitější a projevy se manifestují aţ
po delší době latence.
Vztah dávky a účinku
Ozáření člověka můţe vyvolat některé chorobné změny projevující se v průběhu
dnů aţ týdnů, jiné v průběhu roků a desetiletí.
To vedlo k zavedení třídění opírajícího se o dva základní typy vztahu dávky a
účinku.
Poznání tohoto vztahu a jeho kvantitativní popis je hlavním cílem biologických
a medicínských studií zaměřených na ochranu před ionizujícím zářením.
Na základě těchto znalostí lze pro činnosti probíhající za kontrolovaných
podmínek vymezit cíle a kritéria radiační ochrany a pro případy mimořádných
situací spojených s ozářením lidí odhadnout moţné následky i úkoly
zdravotnické pomoci.

12
Účinky ionizujícího záření se z hlediska integrovaného savčího organismu dělí
na deterministické, kdy při dosaţení určité dávky ionizujícího záření efekt
zákonitě nastává, a stochastické, kdy se stoupající dávkou stoupá
pravděpodobnost poškození.
Zatímco deterministické účinky jsou podmíněny především buněčnými ztrátami
v důleţitých buněčných populacích, pro stochastické účinky je patogenetickým
podkladem na buněčné úrovni mutace genu.
Zatímco stochastické účinky jsou zřejmě bezprahové, (libovolně malá dávka
můţe vyvolat jí úměrné poškození na genetické úrovni), pro deterministické
účinky existuje vţdy práh manifestace poškození, pod nímţ se účinky ozáření
navenek nijak neprojeví, tj. neovlivní klinický obraz ozářené osoby.
Je třeba zdůraznit, ţe kategorie deterministických účinků předpokládá existenci
projevů zjistitelných makroskopickými pozorováními, jeţ se u člověka kryjí
s pojmem klinické příznaky.
Na úrovni buněčné a molekulární se ovšem i u těchto účinků uplatňuje moment
stochastických zákonitostí (náhodnosti a statistické pravděpodobnosti).
Souvislost obou jevů je znázorněna na obrázku 2.

13
1) Deterministické účinky záření
Se stoupající dávkou ionizujícího záření stoupá počet inaktivovaných elementů
v buněčné populaci, který je relevantní pro daný deterministický efekt.
Malé buněčné ztráty však organismus toleruje, tj. odpovídající funkce není
porušena.
Úbytku buněk v populaci přísluší na základě nestejné radiosenzitivity určité
statistické rozpětí, takţe i dosaţení prahu deterministického efektu vyjádřeného
horizontální přímkou v dolní části grafu 2 je u jednotlivců rozloţeno v určitém
rozpětí dávek.
Klinické projevy deterministických účinků u člověka jsou velmi rozmanité,
neboť proces deplece buněčných populací se realizuje v závislosti na geometrii
ozáření v různých částech těla, tj. v různých orgánech a tkáních či jejich částech,
a to v různé kvantitativní míře v závislosti na dávce a její prostorové distribuci.
Pokud je dávka rozloţena na delší časový interval, uplatňují se vedle toho ještě
reparační mechanismy.
Existence reparačních mechanismů je důvodem toho, ţe vzhledem
k deterministickým účinkům nelze dávky rozloţené do delšího časového období
prostě sčítat.
Vychází-li se z teoretické a experimentálně doloţené představy (např. ve vztahu
k indukci chromosomálních aberací), ţe vztah dávky a účinku je pro řídce
ionizující záření v zásadě lineárně-kvadratický, potom dávky vyvolávající
deterministický efekt spadají do oblasti, kde se výrazně uplatňuje kvadratická
sloţka.
Pro malé dávky a malé dávkové příkony se předpokládá v zásadě lineární
průběh vztahu dávky a účinku.
a) Akutní nemoc z ozáření
Akutní nemoc z ozáření vzniká typicky po jednorázovém celotělovém ozáření
vyšší dávkou pronikavého záření.
U člověka se ke kvantifikaci míry deterministických účinků ozáření pouţívá
komplexního ukazatele LD 50/60, coţ je letální dávka, při níţ zemře 50 %
ozářených jedinců v období do 60 dnů po ozáření.
Ve správě UNSCEAR 1993 se uvádí hodnota LD 50/60 pro ozáření člověka ve
střední čáře pro řídce ionizující záření v rozpětí 2,1 Gy aţ 5,25 Gy se střední
hodnotou 3,5 Gy.
Klinický obraz akutní nemoci z ozáření závisí na velikosti celotělové dávky.
Podle její stoupající výše převládají nejprve příznaky podmíněné poruchou
funkce sliznic, poté krvetvorných orgánů, při vyšších dávkách příznaky z oblasti
trávicího ústrojí a konečně důsledky poruch centrálního nervového systému a
metabolismu.

14
1) Orofaryngeální syndrom se projevuje jiţ při jednorázovém ozáření dávkou
2 – 3 Gy.
Je charakterizován změnami na sliznici dutiny ústní a v hltanu, kde jsou
postiţeny zejména tkáně bohaté na lymfatické elementy, jako jsou tonzily a
kořen jazyka.
Postiţeny jsou i dásně, sliznice tváří a měkkého patra.
Vedle krvácení a povrchových ulcerací mohou vznikat i hluboké nekrózy.
Jde o projevy jiţ vcelku závaţné, provázené bolestí a hromaděním lepkavých
hlenovitých hmot, které ucpávají dýchací cesty a ztěţují dýchání.
Neméně významné můţe být poškození plicní tkáně, neboť pro její relativně
nízkou denzitu je tkáňová dávka z fyzikálních důvodů vyšší neţ v okolních
hutnějších strukturách.
Terčovými elementy jsou zde buňky alveolů, případně i jemné cévy.
V průběhu několika málo měsíců po ozáření se můţe vyvinout radiační
pneumonitis, který můţe vést i ke smrti.
2) Hematologická forma se svými hraničními příznaky můţe projevit jiţ při
jednorázovém celotělovém ozáření dávkou okolo 1 – 2 Gy, avšak typický obraz
odpovídá dávkám nad 3 Gy.
V klinickém průběhu lze rozlišit několik období.
Během prvních 48 hodin po ozáření se rozvíjejí přechodné úvodní příznaky,
které jsou důsledkem odezvy regulačních systémů organismu, především
autonomního nervového systému.
Jsou reprezentovány projevy gastrointestinálními a neuromuskulárními.
K první skupině patří nechutenství, nausea, zvracení, průjem, salivace a
dehydratace organismu.
Ke druhé pak únava, apatie, pocení, bolesti hlavy, pokles krevního tlaku a
zvýšená teplota.
Zvracení bývá dominujícím příznakem.
Je důsledkem podráţdění dvou center v prodlouţené míše nízkomolekulárními
peptidy.
Jedním z center je chemorecepční spouštěcí zóna, druhým vlastní centrum
zvracení.
Doba nástupu zvracení a jeho frekvence můţe býti vodítkem k předběţnému
odhadu závaţnosti ozáření.
Po odeznění úvodních obtíţí nastává několikadenní období latence, kdy
postiţený nemá výrazné příznaky.
Plný rozvoj nemoci z ozáření je charakterizován výrazným zhoršením celkového
stavu.
Nemocný trpí horečkami, za příznaků mikrobiálního rozsevu a zhroucení
imunitních ochranných struktur, krvácením ze sliznic, popř. koţními projevy
krvácení.

15
Tyto příznaky vrcholí v době nejhlubšího poklesu neutrofilních leukocytů a
krevních destiček, který nastupuje při dávkách 4 Gy okolo 20. dne po ozáření,
při dávkách 6 Gy asi 7. den po ozáření.
Jednotlivé třídy elementů periferní krve prodělávají změny vyplývající ze zániku
radiosenzitivních buněk.
Z leukocytů jsou nejcitlivější lymfocyty, které mohou uţ při dávkách 1 – 2 Gy
poklesnout v průběhu 48 hodin na 50 % původní hodnoty, neboť podléhají
interfázové smrti.
Neutrofilní leukocyty vykazují zprvu krátkodobé zvýšení počtu, coţ je
výsledkem nastartování regulačních mechanismů.
Později však rovněţ nastává jejich pokles v důsledku blokády mitotického cyklu
v kompartmentu prekursorů.
Při dávkách 2 – 5 Gy je patrný kolem 10. dne přechodný vzestup této třídy
bílých krvinek připisovaný náznaku obnovy z kompartmentu komitovaných
progenitorů, která však nemůţe pokračovat pro deficit kmenových buněk.
Trombocyty v periferní krvi prodělávají analogické změny, abortivní vzestup se
však u nich nepozoruje.
Pokud celotělová dávka záření nepřekročí 3 Gy, mohou se někdy objevit, po 6 –
8 týdnech, známky pomalého postupného zlepšování zdravotního stavu.
Ze zachovalé frakce kmenových buněk proběhne v krvetvorných orgánech nová
repopulace.
Pro dávky 3 – 6 Gy se průměrná doba přeţití postiţené osoby pohybuje okolo
jednoho měsíce.
3) Gastrointestinální forma akutní nemoci z ozáření se projevuje po
jednorázové celotělové dávce 6 – 10 Gy, kdy rozvinuté příznaky nastupují mezi
4. a 7. dnem po ozáření.
Spočívají v krutých průjmech provázených ztrátou tekutin, minerálním
rozvratem a krvácením ze střev.
Patogeneticky jde o projevy nekrózy buněk střevního epitelu s obnaţením
povrchu střeva.
V pozdějších stádiích dochází k rupturám střevní stěny s následnou sepsí.
Časový nástup změn souvisí s větší rychlostí obratu buněk střevního epitelu tj.
dobou přechodu od kmenových buněk ve střevních kryptách k funkčním
buňkám klků tenkého střeva.
Vyšší prahová dávka zase souvisí s relativní radiorezistencí těchto buněk ve
srovnání s kmenovými buňkami krvetvorby.
Přeţije-li nemocný 7 dnů, coţ bývá spíše výjimkou, projeví se u něho ovšem
v plné míře i příznaky poškození krvetvorných orgánů.
4) Neuropsychická forma akutní nemoci z ozáření se projevuje při jednorázové
celotělové dávce nad 10 Gy. Je důsledkem zejména radiačně indukovaného
narušení krevního zásobení centrálního nervového systému.

16
Během několika minut se objevuje celková zmatenost a dezorientace, nejpozději
za několik hodin upadá postiţený do těţkého komatu, z něhoţ se jiţ neprobere.
Během 6 hodin zcela vymizí lymfocyty v periferní krvi.
Smrt se dostavuje v průběhu několika málo desítek hodin po expozici.
5) Kardiovaskulární a toxemický syndrom nastává po jednorázovém
celotělovém ozáření dávkou v řádu několika desítek Gy.
Poté co postiţený upadne po několika minutách, či desítkách minut do komatu,
dochází k totálnímu metabolickému rozvratu v celém organismu a během
několika hodin nasává srdeční selhání.
b) Člověk jako indikátor míry ozáření
Nejcitlivější a prakticky dobře propracovanou metodou k posouzení významu
celotělového ozáření je stanovení četnosti chromosomálních aberací
v lymfocytech periferní krve.
K hodnocení účinků ionizujícího záření se hodí nejlépe pro svoji tvarovou
jednoznačnost dicentrické chromosomy, vzniklé spojením větších zbytků dvou
zlomených chromosomů.
Laboratoře mají k dispozici kalibrační křivky pro rozdílné energie fotonů, které
jim umoţňují posoudit efektivní odezvu, pro kterou byl zaveden termín
efektivní celotělová dávka (ECD).
V době ozáření jsou v závislosti na rozloţení dávky v těle zasaţeny lymfocyty
cirkulující v periferní krvi nebo procházející orgány (názorné představě pomáhá
přirovnání lymfocytů k cirkulujícím dozimetrům).
Při dostatečném počtu hodnocených buněk lze detekovat ECD řádu stovek mGy.
Desítky procent dicentrických chromozomů se zjišťují po ozáření v oblasti
jednotek Gy.
Horní mez spolehlivosti představuje dávka asi 4 Gy, která vyvolá natolik značné
poškození buněk, ţe výskyt mnohočetných aberací znemoţňuje spolehlivou
aplikaci kalibrační křivky.
Léčba akutní nemoci z ozáření vyţaduje izolaci postiţeného z důvodu ochrany
před mikrobiálními infekcemi a podávání antibiotik.
Nejnáročnějším opatřením v tomto směru můţe být tzv. reverzní izolace
v zařízení typu „life-island“.
c) Akutní radiační poškození kůže
Z lokálních časných účinků je v popředí zájmu radiační ochrany akutní
poškození kůţe.
Je to nejčastější typ poškození při nehodách se zdroji zevního záření.
Pokud se při nehodě ocitl zářič v blízkosti povrchu těla, nebo přišel dokonce do
přímého kontaktu s ním, je vzhledem ke strmosti gradientu dávky podstatně
vyšší dávka v kůţi, neţ v kterékoli jiné tkáni.

17
Odezva koţního orgánu je závislá nejen na velikosti dávky, ale významně i na
druhu záření, energii částic, velikosti pole a lokalizaci terčové oblasti v těle.
Podle závaţnosti projevů se rozlišují tři stupně akutních koţních popálenin
vyvolaných ionizujícím zářením.
1) Erytematózní dermatitis je nejlehčí formou akutního poškození kůţe
charakterizovanou zarudnutím.
Jako typické podmínky pro vyvolání 50 % případů do 30 dnů se uvádí např.
záření X o energii 200 keV v dávce 5,7 Gy na vnitřní plochu předloktí o
rozměrech 1010 cm.
Stejný efekt lze vyvolat u 80 % zasaţených při energii 50 – 100 keV, a dávce
3 – 4 Gy.
Pro fotony o energii 200 – 400 keV činí prahová erytémová dávka asi
dvojnásobek a pro energie okolo 1 MeV dokonce trojnásobek.
Bezpříznakové období latence trvá podle velikosti dávky 2 – 4 týdny.
Potom se projeví zarudnutí spojené se zánětlivou exsudací v koritu.
Drobné tepénky a kapiláry se tromboticky uzavírají a způsobují rozšíření
kolaterál.
Vzhledem k vyšší citlivosti buněk vlasových míšků a mazových ţlázek na
ozáření, zastavuje se buněčné dělení těchto struktur jiţ v dávkových úrovních
odpovídajících této formě radiační dermatitidy a objeví se vypadávání vlasů a
ochlupení, tj. epilace.
Přechodná epilace vzniká po dávce 3 Gy a projeví se kolem třetího týdne.
Po dávce 6 Gy můţe být epilace trvalá.
Nejvnímavější k vyvolání epilace jsou ty partie kůţe, kde se vlas rychle
obnovuje, jako je tomu na vlasaté části hlavy a ovousené části obličeje u muţů.
Zjištění míst postiţených ztrátou ochlupení je významným ukazatelem rozloţení
dávky na povrchu těla.
2) Deskvamativní dermatitis představuje střední stupeň radiačního poškození
kůţe.
Vzniká po ozáření dávkami okolo 12 – 20 Gy.
Bývá uváděna časným erytémem, který se objevuje v průběhu prvních dvou dnů
po ozáření, často však jiţ v prvních hodinách.
Bývá nevýrazný, takţe můţe snadno uniknout pozornosti, a trvá okolo 24 hodin.
Jde o rozšíření kapilár vlivem látek podobných histaminu, které se uvolňují
z ozářené tkáně.
Při větších dávkách se prchavý erytém můţe objevit i v několika vlnách.
Doba latence do rozvoje plných příznaků bývá 2 – 3 týdny.
Zánětlivý exsudát se hromadí v úrovni poškozené vrstvy bazálních buněk a
epidermolýzou vznikají puchýře.
Odlučováním puchýřků a jejich infekcí se stav dále komplikuje - vyvíjí se
plošné mokvání.
18
V příznivějším případě nastává po dalších 2 – 3 týdnech obnova pokoţky
z okrajů defektu a z přeţívajících kmenových buněk ve vlasových míšcích.
3) Nekrotická forma radiační dermatitidy je důsledkem postiţení hlubších
vrstev kůţe, popř. podkoţí.
Je zpravidla následkem patologických cévních změn a komplikující infekce, kdy
postiţený okrsek tkáně odumírá, odlučuje se a vytváří vřed.
Hlubší defekty se velmi obtíţně a dlouhou dobu hojí z důvodu špatného
krevního zásobování v důsledku cévní degradace v okolí postiţeného místa.
Zahojí-li se přeci jen defekt, je další osud postiţeného okrsku více neţ nejistý.
Nová pokoţka, často velmi tenká a křehká, nemá dostatečně spolehlivou
podkladovou vyţivující vrstvu, a proto špatně odolává mechanickým,
chemickým a mikrobiálním faktorům.
Po několika letech se proto můţe objevit sekundární vřed vyţadující zpravidla
zákrok plastického chirurga.
Klinický obraz je výrazně modifikován větším rozsahem lézí.
Těţký stav spočívá v celkových příznacích vyplývajících z toxémie a vyţaduje
aplikaci náročných postupů popáleninové medicíny.
Hodnocení prognózy koţních změn se opírá jak o dostupná dozimetrická data,
tak i o posouzení doby vzniku a charakteru lokálních změn.
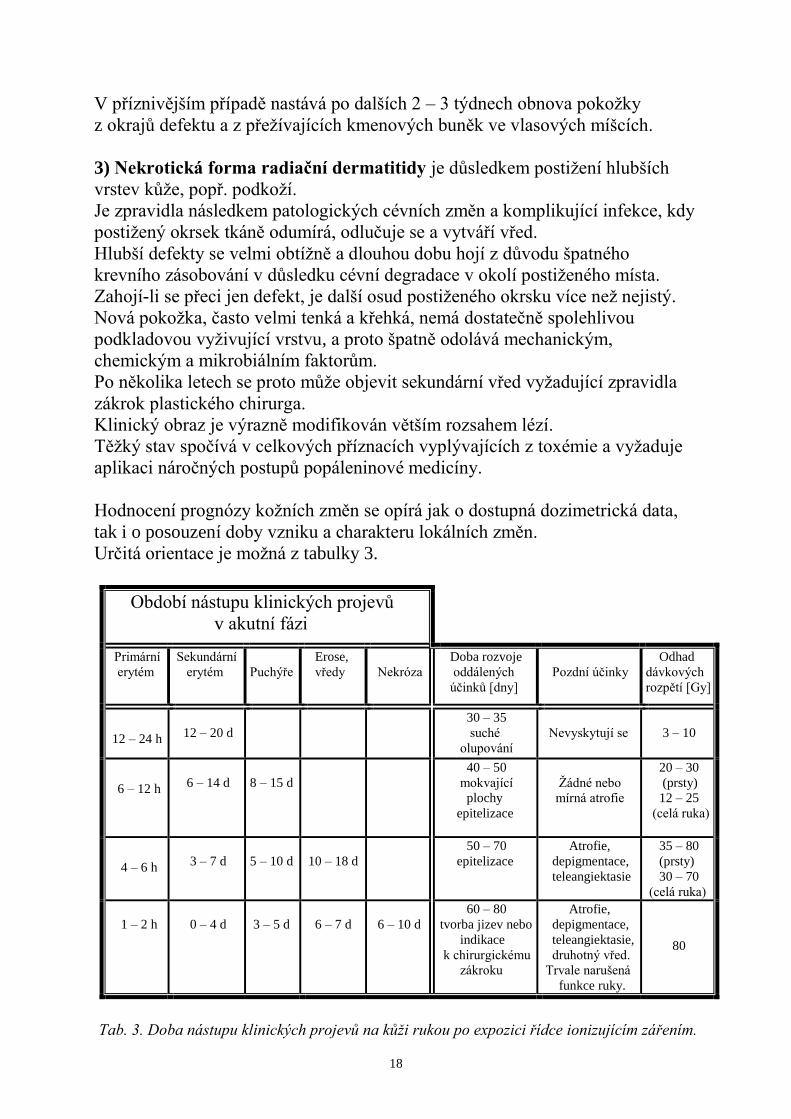
Určitá orientace je moţná z tabulky 3.
Období nástupu klinických projevů
v akutní fázi
Primární
erytém
Sekundární
erytém
Puchýře
Erose,
vředy
Nekróza
Doba rozvoje
oddálených
účinků [dny]
Pozdní účinky
Odhad
dávkových
rozpětí [Gy]
12 – 24 h
12 – 20 d
30 – 35
suché
olupování
Nevyskytují se
3 – 10
6 – 12 h
6 – 14 d
8 – 15 d 40 – 50
mokvající
plochy
epitelizace
Ţádné nebo
mírná atrofie
20 – 30
(prsty)
12 – 25
(celá ruka)
4 – 6 h
3 – 7 d
5 – 10 d
10 – 18 d 50 – 70
epitelizace
Atrofie,
depigmentace,
teleangiektasie
35 – 80
(prsty)
30 – 70
(celá ruka)
1 – 2 h
0 – 4 d
3 – 5 d
6 – 7 d
6 – 10 d
60 – 80
tvorba jizev nebo
indikace
k chirurgickému
zákroku
Atrofie,
depigmentace,
teleangiektasie,
druhotný vřed.
Trvale narušená
funkce ruky.
80
Tab. 3. Doba nástupu klinických projevů na kůži rukou po expozici řídce ionizujícím zářením.

19
Biofyzikální přístup k dávce v kůţi musí respektovat vzrůstový faktor a
nestejnou relativní biologickou účinnost částic ve vztahu k jejich energii.
Jak vyplývá z veličin zavedených pro radiační ochranu, je dávka do hloubky
0,07 mm pro biologický efekt nevýznamná, neboť postihuje převáţně
neţivotnou zrohovatělou vrstvu buněk v pokoţce.
Dávkou v kůţi se často rozumí dávka maximální, která je např. pro 60
Co (1,17-
1,33 MeV) v hloubce 4,5 mm.
Důleţitá je také konvence o průměrování dávky v ploše.
Limity ozáření kůţe v našich předpisech se vztahují k průměrné dávce na ploše
1 cm2.
Dalším důleţitým faktorem je časové rozloţení dávky.
Je-li ozáření kůţe rozloţeno na delší období, je účinek menší.
K dosaţení téhoţ účinku, jaký způsobí jednorázová dávka, je třeba při aplikaci
15 – 20 frakcí v průběhu jednoho měsíce dávky asi trojnásobné.
Jiným faktorem je vliv velikosti ozářené plochy kůţe.
Táţ plošná dávka na menším poli vyvolá menší účinek, coţ je důsledkem
příznivého působení zdravého okolí (migrací buněk a působků) na vývoj a
průběh změn.
Odezva kůţe je podmíněna i nestejnou odolností různých částí povrchu těla.
Nejcitlivější je kůţe na přední části krku, v loketní a záloketní jamce.
d) Poškození zárodečného epitelu
Sterilizační účinek ionizujícího záření na muţské a ţenské pohlavní ţlázy je
třeba hodnotit odděleně.
Jejich anatomické uloţení vytváří odlišné podmínky pro realizaci dávky
v zárodečném epitelu.
Značně odlišná je ovšem i buněčná kinetika muţských a ţenských zárodečných
buněk.
Také metodický přístup k hodnocení poklesu fertility se podle pohlaví liší.
U muţů je hlavní metodou hodnocení spermiogramu, u ţen je hodnocení
zaloţeno na údajích o menstruaci, o historii gravidit a na vyšetření hladiny
hormonů.
Muţi jsou z hlediska vyvolání poruch fertility ionizujícím zářením vnímavější
neţ ţeny.
Při běţném terapeutickém frakcionovaném reţimu (10 – 35 frakcí v rozpětí 2 –
7 týdnů) byla přechodná oligospermie zjištěna jiţ po dávkách 100 mGy.
Za obdobných podmínek vyvolá dávka nad 500 mGy přechodnou aspermii u
100 % jedinců.
Počet spermií poklesne během 8 – 20 týdnů, regenerace proběhne v horizontu 1
– 3 let.
Dávky nad 3 Gy obvykle vedou k aspermii trvalé.
U ţen do 40 let věku frakcionované dávky do úrovně 1,5 Gy nevedou ke
zřetelné odezvě.

20
Jak však bylo uvedeno výše, s věkem počet ovariálních folikulů klesá a
v souvislosti s tím se vnímavost na ozáření zvyšuje.
Zářením vyvolaný zánik folikulů se jiţ nenahradí.
K trvalé sterilitě u ţen mohou vést jiţ dávky okolo 2 Gy.
Z hlediska společenského uplatnění jedince je významná i otázka ovlivnění
druhotných pohlavních znaků.
U muţů zůstávají buňky produkující pohlavní hormony díky své relativní
radiorezistenci obvykle nedotčeny.
Sekundární pohlavní znaky, potence a libido tím zůstávají zachovány.
U ţen je produkce pohlavních hormonů vázána na zrající folikuly a s jejich
zánikem mizí i hormonální funkce a navozuje se předčasné klimakterium.
e) Účinky záření na vývoj lidského zárodku a plodu
Předpokladem úspěšného zahájení vývoje jedince je bezchybný průběh
progeneze.
V progenezi se dokončí vývoj a zrání zárodečných buněk, tj. muţské a ţenské
gamety, které splynutím při koncepci dají vznik zygotě.
Ta se dále vyvíjí v procesu blastogeneze (1. – 2. Týden), kdy probíhá rýhování
oplozeného vajíčka, jeho zanořování do děloţní sliznice a zakládají se zárodečné
listy.
Z tohoto základu vzniká zárodek (embryo, od 3. týdne) a posléze plod (fetus, od
3. Měsíce).
Základní buněčný mechanismus účinku záření působícího v období
nitroděloţního vývoje je usmrcení buněk, tj. jejich vyřazení z další proliferace.
Důsledky závisejí na zmíněných stadiích vývoje.
Důsledky ozáření v období preimplantace a blastogeneze se řídí pravidlem
„vše nebo nic“.
Ozářená zygota nebo základ blastocysty buď zaniknou jako celek, nebo
v případě ztráty ojedinělé buňky vývoj pokračuje bez dalších důsledků, neboť
okolní buňky se širokou prospektivní potencí tuto ztrátu plně nahradí.
Pokud nastane takováto situace u lidského zárodku, nevyplynou z ní ţádné
zdravotní či sociální důsledky.
Z hlediska vlivu na embrio je kritickým časovým úsekem období tzv. velké
organogeneze, které spadá do přibliţně 4. – 8. týdne po oplození vajíčka.
Buněčné ztráty způsobené ozářením v tomto období mohou vést k uhynutí
zárodku, sníţené hmotnosti embria či jednotlivých orgánů nebo k tvarovým
změnám v důsledku narušení morfogeneze organismu.
Tak můţe docházet i k postiţení vývoje centrálního nervového systému a jeho
kosterního obalu, které se projeví např. zakrněním mozku (mikrocefalie), nebo
oka (mikroftalmie), rozštěpy patra aj.
Prahové dávky pro vznik těchto nepříznivých efektů se odhadují na 50 mGy.

21
Ozářením v časném fetálním období je výrazně ohroţeno vyzrávání
centrálního nervového systému, ve kterém zejména mezi 8. – 15. týdnem vrcholí
funkčně významné translace buněk a tvorba synaptických spojení.
Ozáření v tomto kritickém období se projeví v dětství mentální retardací, která
je charakterizována celkovou zaostalostí jedince provázenou neschopností péče
o sebe a nezvládnutím poţadavků gramotnosti.
Pravděpodobnost tohoto poškození je vysoká, jak je patrno z koeficientu rizika
410-1
Sv-1
připisovanému tomuto účinku.
Ozáření 1 Sv způsobí pokles IQ z hodnoty 100 na cca. 70.
Směrem k niţším dávkám se extrapolace lineárního vztahu neuplatňuje.
Usuzuje se spíše na prahovou dávku v úrovni 120 mSv.
Pro období 16. – 25. týdne po početí klesá riziko tohoto efektu ve srovnání
s předchozím obdobím asi na čtvrtinu.
V pozdním fetálním období je hlavním rizikem vnímavost na indukci leukémie
a jiných malignit projevujících se v dětství (kritériem bývá období do deseti let
věku).
Tato vnímavost se týká i dřívějších období vývoje zárodku a plodu, kdy je však
její význam překryt riziky dalšími.
Riziko malignity vedoucí u dítěte k smrti se odhaduje na 1,1210-1
Sv-1
po celé
období gravidity.
Indukce dětských nádorů ozářením in utero je reálná i při velmi malých
dávkách, jak vyplývá mj. ze studií prováděných u dětí, jejichţ matky byly
rentgenologicky vyšetřovány z porodnické indikace v období pokročilé
gravidity.
Při ozáření plodu dávkou niţší neţ 20 mSv neexistuje významnější riziko pro
další nepříznivý vývoj dítěte.
Dávka 20 – 50 mSv jiţ vyţaduje patřičnou pozornost s vyuţitím spolupráce se
sloţkami SÚJB popř. SÚRO pro maximální upřesnění jejího odhadu.
Nepředstavuje však ještě bezpodmínečné poškození plodu.
U dávek 50 – 100 mSv lze jiţ předpokládat vliv ozáření na další vývoj plodu.
Také celkové riziko gravidity se tím přibliţně zdvojnásobí ve srovnání
s přirozenými riziky těhotenství.
Dávky 100 – 200 mSv (které jsou ovšem u radiodiagnostických vyšetření spíše
výjimečné), zejména pokud k ozáření dojde v posledních měsících prenatálního
vývoje, nejsou rozhodně zanedbatelné, neboť při nich mnohonásobně vzrůstá
riziko indukce zhoubných nádorů v období po narození dítěte ve srovnání
s rizikem indukce karcinomu u dospělého člověka ozářeného stejnou dávkou.
Indikace k interrupci je však při těchto dávkách na místě pouze tehdy, pokud
k ozáření došlo v době, kdy ještě neuplynula doba stanovená zákonem pro
moţnost tohoto typu zákroku.

22
f) Pozdní projevy radiačního poškození deterministického typu
1) Chronická radiační dermatitida byla v první polovině 20. století poměrně
častým nálezem u rentgenujících lékařů, kteří si neuvědomovali moţné důsledky
manipulace rukama v primárním svazku rentgenky.
Méně rozvinuté formy byly zjišťovány ještě u generace lékařů začínajících svou
kariéru v padesátých letech 20. Století.
U současných lékařů a zdravotnických pracovníků, zejména pouţívajících
metod invazivní radiologie, bývají někdy zjišťovány náznaky trofických změn
na kůţi rukou.
Takový nález je důvodem k důkladnému prověření pracovních podmínek včetně
technického zajištění ochrany a organizace práce.
Atrofický typ tohoto onemocnění se projevuje suchou, tenkou, hladkou
pokoţkou.
Mohou být patrné teleangiektasie, poruchy pigmentace, lomivost nehtů.
Křehkost takové kůţe vede k trhlinám a pozdním vředovitým defektům.
Hypertrofický typ má pokoţku zhrubělou, koţní záhyby výraznější.
Loţiskové hyperkeratózy přecházejí nejednou ve spinocelulární karcinom.
Jizevnaté a degenerativní změny kůže se mohou zjišťovat po velkých dávkách
záření, jak tomu bývá po nehodách, či nekorektně provedených
radioterapeutických zákrocích.
U osob postiţených chronickými koţními změnami je třeba předpokládat
celkovou lokální dávku alespoň 30 Gy.
2) Zákal oční čočky (katarakta) postihuje útvar, který nemá přímou výţivu ani
vlastní buněčný substrát.
To je příčinou dlouhé doby latence mezi ozářením a rozvojem choroby.
Přitom oční čočka je na ozáření velmi citlivá.
Při jednorázovém ozáření řídce ionizujícím zářením činí prahová dávka 1,5 Gy,
ve směsném poli hustě a řídce ionizujícího záření dokonce jen 1 Gy.
Doba latence do vzniku prvních oftalmologickými metodami zjistitelných
zákalů na zadním pólu čočky činí alespoň 6 měsíců, klinicky závaţný stupeň se
vyvíjí v období let.
V podmínkách dlouhodobé profesionální expozice by bylo třeba počítat
s prahem 3 – 4 Gy a s dobou latence nejméně 2 roky.
3) Chronická nemoc z ozáření je koncept zavedený v bývalém Sovětském
svazu, který zprvu nenašel obecného ohlasu, neboť v jiných zemích nebyly
klinické obrazy popisované sovětskými autory pozorovány.
Po pádu ţelezné opony a odtajnění informací se ukázalo, ţe v jaderných
vojenských zařízeních Sovětského svazu činily roční dávky u některých
pracovníků aţ 1 Gy.

23
Za těchto okolností se výskyt neurovegetativních příznaků, jeţ byly uváděny
jako typické pro první stádium nemoci, jakoţ i deterministicky podmíněných
hraničních krevních odchylek ukázal jako moţný.
4) Urychlené stárnutí a zkrácení střední doby života bylo zjištěno
v některých studiích po ozáření vyšší dávkou.
Patogeneticky se alespoň v části pozorování mohlo uplatnit postiţení
parenchymatózních orgánů v důsledku poruch trofiky, způsobených hyalinním
zesílením stěn kapilár a poruchou jejich permeability.
Pro dlouhodobé ozáření malými dávkami by zkrácení střední doby ţivota u
člověka mohlo vyplývat pouze ze zvýšeného výskytu maligních mutací.
V publikaci ICRP 60 je proveden odhad střední doby ztráty ţivota v důsledku
radiačně indukovaného fatálního nádoru s rozpětím 9 – 31 let pro nádory
různého typu, se střední hodnotou 15 let.
Není dostatek důvodů předpokládat u člověka za podmínek nízkých dávek a
nízkých dávkových příkonů vliv ještě jiného patogenetického mechanismu
působícího zkrácení délky ţivota.
2) Stochastické účinky záření
O cytogenetickém charakteru mutací a s nimi souvisejících maligních
transformací buněk bylo pojednáno ve třetí kapitole.
Tyto molekulární a buněčné mechanismy jsou patogenetickým podkladem dvou
kategorií pozdních účinků, charakterizovaných některými společnými rysy.
Patří sem nádory, které mají vztah k mutacím somatickým a dědičné poruchy
vyvolané zářením, které mají vztah k mutacím gametickým.
Pro obě kategorie stochastických účinků je společné, ţe i v referenční neozářené
populaci se nádory a hereditární nepříznivé projevy spontánně vyskytují.
Jak dále ukáţeme, existuje dostatek důkazů o tom, ţe ionizující záření zvyšuje
pravděpodobnost výskytu těchto projevů.
Přitom jejich klinický obraz ani další charakteristiky nedovolují v individuálním
případě rozlišit, zda se jedná o poruchu v rámci spontánního výskytu nebo o
následek ozáření.
Zatímco u deterministických účinků jsme uvedli, ţe dávky za delší období nelze
prostě sčítat v důsledku uplatňování reparačních mechanismů, je ve vztahu ke
stochastickým účinkům takovéto sčítání v prvním přiblíţení přiměřené, neboť se
vztahuje především k malým dávkám a malým dávkovým příkonům, kde platí
přibliţně lineární závislost účinku na dávce.

24
a) Radiačně indukované maligní nádory
Pokrok v objasnění mechanismů kancerogeneze přinesly objevy šedesátých a
sedmdesátých let 20. století, kdy byla nalezena řada tzv. onkogenů u
experimentálních zvířat i u člověka.
Vedle promotorů buněčného dělení, tzv. protoonkogenů, byly objeveny téţ
supresorické geny které buněčné dělení naopak zastavují.
Mutace obou dvou druhů genů můţe vést ke ztrátě schopnosti regulace
buněčného dělení a tedy i k moţnosti vzniku nádorového bujení.
Tak např. mutace supresorického genu p53 byly nalezeny u nádorů tlustého
střeva, močového měchýře, jater, mléčné ţlázy, aj.
Nové poznatky jsou podnětem ke konstrukci modelových shémat, kde se
vícestupňový proces rozvoje rakoviny interpretuje jako důsledek sledu mutací,
který se nakonec projeví klinicky manifestovanou malignitou.
Na rozvoj a růst nádoru působí však nepochybně i jiné regulační a integrační
systémy organismu, z nichţ některé (např. imunitní) mohou k eliminaci
atypických buněk přispívat, jiné (např. hormonální) růst nádoru podporují.
Specifické podmínky uplatnění ionizujícího záření v procesu kancerogeneze
nejsou zatím známy, současné modely však připouštějí představu, ţe ionizující
záření můţe působit (především indukcí mutací) na více stupních rozvoje ná-
dorů.
Pro rozbor a hodnocení rozvoje nádorů po ozáření je významný časový faktor.
Doba mezi ozářením a manifestací nádoru, zvlášť dobře určitelná po
jednorázovém ozáření, tedy doba latence, trvá několik let. Pro leukémie byl
stanoven medián doby latence na 8 let, nejkratší latence okolo dvou let byla
nalezena u akutní myeloidní leukémie a u kostního sarkomu vyvolaného 224Ra.
U řady solidních nádorů činí medián doby latence 15 - 25 let. Ve studii o
následcích jaderných útoků na japonská města je zjišťován pro některé solidní
nádory zvýšený výskyt i po 45 letech.
Poznatky o vlivu ionizujícího záření na vznik nádorů u člověka a podklady k
odvození kvantitativních ukazatelů takového rizika se získávají ve skupinových
šetřeních (epidemiologických studiích) jak odpovídá stochastickému charakteru
těchto účinků.
b) Dědičné důsledky ozáření rodičů
Ovlivnění dědičných znaků zářením bylo prokázáno na banánových muškách
(Drosophilách melanogaster) uţ v roce 1927, kdy americký badatel a pozdější
nositel Nobelovy ceny H. J. Miller získal první jejich mutanty po ozáření
paprsky X. Později byla provedena řada studií na velikých skupinách myší a
menších skupinách dalších savců včetně primátů. Z nich byly odvozeny některé
obecné závěry, např. větší vnímavost prekursorů zárodečných buněk samců ve
srovnání se samicemi, oprávněnost aproximovat vztah dávky a účinku v oblasti
malých dávek přímkou a poţadavek váţit při dlouhodobém ozařování zjištěnou

25
pravděpodobnost indukovaných mutací dobou trvání té které fáze vývoje gamet.
Řada epidemiologických studií se pokoušela o přímý důkaz účinků ionizujícího
záření na dědičně podmíněné proměnné u člověka.
K provedení této extrapolace ze zvířat na člověka je třeba znalostí o molekulární
a buněčné povaze mutací, o pravidlech přenosu dědičných znaků u člověka, o
spontánní frekvenci geneticky podmíněných znaků v populaci aj.
Je třeba odlišit dva druhy radiačně indukovaného genetického poškození:
genové mutace spočívající v alteraci elementární jednotky dedičnosti, tj. genu,
realizovaného molekulou DNA nebo její podstatnou částí, a hrubé
chromosomové aberace vyjádřené změnou struktury nebo počtu chromosomů.
Mutace můţe být dominantní - kdyţ se důsledky projeví v prvé generaci
potomstva (také "přenos od jednoho z rodičů", neboť stačí jeden mutovaný gen
v jejich párovém seskupení).
Efekt recesivních mutací se projeví, je-li táţ genová mutace zděděna od
kaţdého z rodičů (párové seskupení téţe mutace). Pohlavně vázané mutace se
vyskytují na pohlavním chromosomu X a projevují se důsledky u muţských
potomků, kteří nesou pohlavní chromosomy XY. Mutace na jiných
chromosomech neţ X se označují jako autosomální mutace.
Při hodnocení genetického rizika je třeba uváţit, ţe důsledky mutací se
manifestují velmi různorodým způsobem. Genetické poškození můţe vést k
hrubému narušení vývoje plodu a jeho obalů a způsobit potrat, jak o tom svědčí
častý nález chromosomových aberací v buňkách potraceného plodu (asi u 20 -
50 % spontánních abortů). Genetická komponenta se můţe podílet na zvýšení
prenatální a kojenecké úmrtnosti. Zvláštní kategorií jsou vrozené malformace
charakterizované zjevnými anatomickými odchylkami. Indikátorem zvýšené
frekvence mutací můţe být i vyšší výskyt specifických dědičných znaků
(specific variables), které nemusí znamenat zdravotní postiţení. Předpokládá se,
ţe i řada konstitučních a degenerativních chorob (obezita, hypertenze, cukrovka
aj.) má svou genetickou komponentu a ţe zvýšená frekvence mutací vede k
vyššímu výskytu těchto chorob.
U dětí ozářených rodičů byly sledovány čtyři indikátory genetického poškození.
Pro ţádný z indikátorů neprokázali autoři statisticky významný rozdíl mezi
ozářenou a kontrolní populací. Při znalosti dávek, jimţ byli rodiče vystaveni, lze
odhadnout, ţe minimální hodnota zdvojující dávky nemohla být niţší neţ 1,56
Gy. Volba zdvojující dávky 1 Gy i pro lidskou populaci je tedy dostatečně
konzervativní.
Dalším souborem dat nutných pro aplikaci nepřímé metody je znalost frekvence
spontánních mutací v populaci. Podle ICRP jsou pro frekvenci mutací v západní
populaci typická tato data: autosomálně dominantní a pohlavně vázané 1 %,
autosomálně recesivní 0,25 %, chromosomálně podmíněné 0,38 %, kongenitální
malformace 6 %, multifaktoriálně podmíněné 65 %. Vysoké hodnotě
multifaktoriálních poruch, vztahujících se ke konstitučním a degenerativním
onemocněním, je třeba rozumět tak, ţe jedinec můţe být nositelem i více neţ
jedné z nich. U multifaktoriálních poruch se odhaduje jejich genetická

26
komponenta tj. podíl genetických mechanismů v souboru všech, tedy zejména
zevních příčin nemoci. Pro výpočty se vychází z genetické komponenty 5 %.
Aplikace zdvojující dávky 1 Gy na údaje o spontánním výskytu vede k tomuto
odhadu vlivu ozáření rodičovské generace na potomky v projekci na všechny
příští generace (např. recesivně podmíněné znaky se mohou projevit v rozmezí
aţ desítek generací):
. váţné genetické poruchy 1,210-2
Sv-1
,
. multifaktoriální poruchy 4,710-2
Sv-1
.
Pro konečný odhad genetického rizika na jednotku ekvivalentní dávky
je třeba provést několik dalších korekcí. Rodičovská populace (konvenčně se
počítá se středním rokem rodičovství 30 let) činí jen 25 % celkové populace (0 -
75 let) a 40 % pracující části populace (18 aţ 65 let). Jako korekce na menší
zdravotní závaţnost multifaktoriálních poruch je zvolena hodnota 0,33. Pro
srovnání odvozených koeficientů genetických poruch s následky indukce nádorů
je přihlédnuto k tomu, ţe genetické poruchy zkracují ţivot v průměru více (o 20
let) neţ fatální zhoubné nádory (o 15 let), odtud korekční faktor 1,33.
Konečné hodnoty odhadu genetických následků v ozářené populaci jsou tedy
stanoveny odlišně pro průměrného pracovníka a průměrného (statistického)
obyvatele:
. pro pracovníka 0,810-2
Sv-1
,
. pro obyvatele 1,3310-2
Sv-1
.
c) Objektivní zdravotní újma
V doporučeních ICRP bylo přihlédnuto k souhrnu poznatků o radiační
kancerogenezi a za základ dalších úvah a výpočtů byla zvolena data o úmrtích
na zhoubný nádor u obyvatel, kteří přeţili jaderné bombardování japonských
měst, pocházející ze studie Life Span Study (LSS).
Kohortu tvoří více neţ 100 000 osob.
Individuální dávky na orgány byly odhadnuty u 85 % osob.
V závislosti na poloze a podmínkách stínění v době výbuchu se dávky u
přeţivších pohybovaly od hodnot menších neţ 1 mGy aţ k hodnotám
převyšujícím 4 Gy.
80 % kohorty přitom obdrţelo dávky niţší neţ 100 mGy.
Odhady rizika pocházející z této studie se staly základem většiny doporučení
radiační ochrany.
V současné době jsou k dispozici výsledky týkající se incidence rakoviny
v letech 1958 – 1987 a úmrtnost v letech 1950 – 1990.
Jedním ze závaţných závěrů této studie je zjištění, ţe pro solidní nádory (tj.
zhoubná onemocnění mimo leukémie) je pro ozáření pod 150 mSv doloţena
oprávněnost lineární aproximace vztahu dávky a účinku.
Některé důleţité údaje z této studie jsou zobrazeny v tabulce 4 a na obrázku 3.

27
Incidence Relativní riziko Mortalita Relativní riziko
Počet případů 1958 - 1987 Počet případů 1950 - 1990
Rakoviny celkem 9014 1,07 7827 1,06
Solidní tumory
8613 1,06 7578 1,05
Leukémie 401 1,16 249 1,54
Tab. 4. Výskyt rakoviny ve studii LLS.
Tato vstupní data byla potom dále podrobena analytickým procedurám a
zjednodušující formalizaci. Nejprve byla provedena korekce z jednorázového
ozáření na ozáření nízkým dávkovým příkonem charakteristickým pro ozařování
pracovníků a obyvatelstva. Pouţil se k tomu převodní faktor DDREF (Dose and
Dose-Rate Effectiveness Factor) v hodnotě 2, neboť návrhy na pouţití vyšších
hodnot (pro některé jiné indikátory nalezen DDREF = 5 - 10) se zdály
nezdůvodněné. Odhad celkového počtu očekávaných nádorů v celoţivotní
projekci závisí na volbě modelu radiační kancerogeneze (extrémem jsou čistý
aditivní a čistý multiplikativní model).
Data z japonských studií byla analyzována pomocí tří modelů, a protoţe model
relativního rizika vede k odlišným výsledkům v závislosti na spontánním
výskytu v příslušné etnické skupině, byly dále modely aplikovány na pět
různých populací (Japonsko, Spojené státy, Anglie, Portoriko, Čína).
28
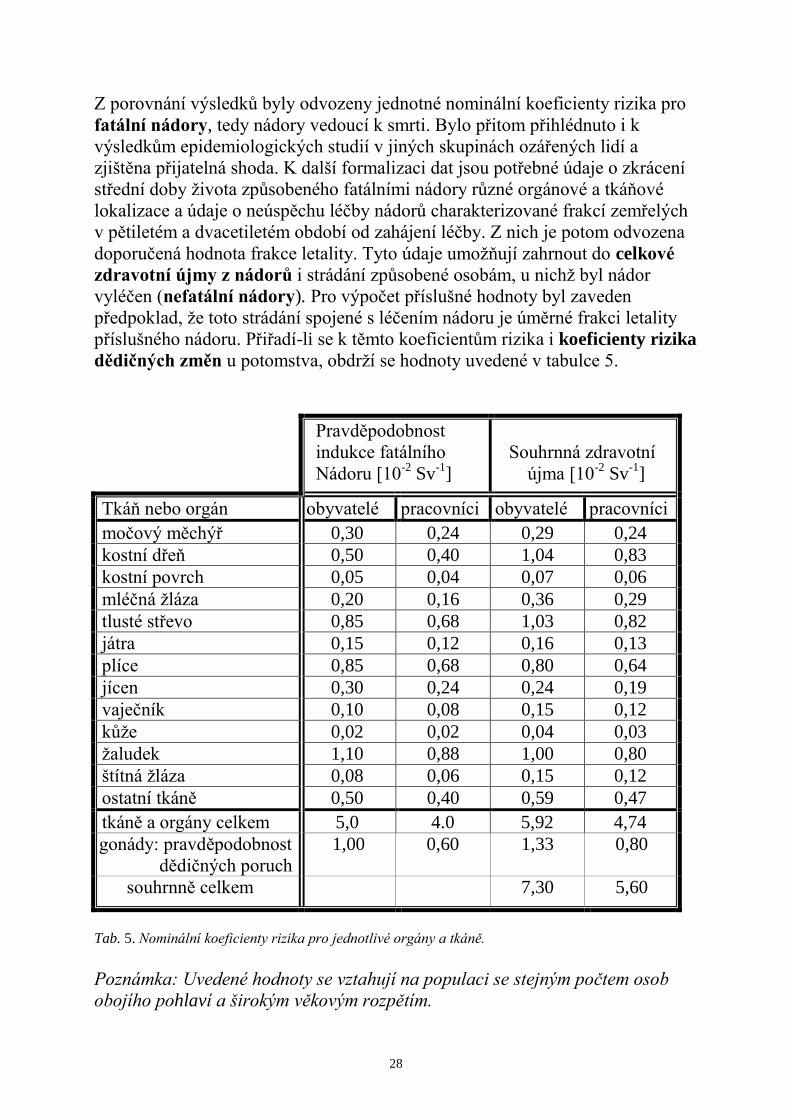
Z porovnání výsledků byly odvozeny jednotné nominální koeficienty rizika pro
fatální nádory, tedy nádory vedoucí k smrti. Bylo přitom přihlédnuto i k
výsledkům epidemiologických studií v jiných skupinách ozářených lidí a
zjištěna přijatelná shoda. K další formalizaci dat jsou potřebné údaje o zkrácení
střední doby ţivota způsobeného fatálními nádory různé orgánové a tkáňové
lokalizace a údaje o neúspěchu léčby nádorů charakterizované frakcí zemřelých
v pětiletém a dvacetiletém období od zahájení léčby. Z nich je potom odvozena
doporučená hodnota frakce letality. Tyto údaje umoţňují zahrnout do celkové
zdravotní újmy z nádorů i strádání způsobené osobám, u nichţ byl nádor
vyléčen (nefatální nádory). Pro výpočet příslušné hodnoty byl zaveden
předpoklad, ţe toto strádání spojené s léčením nádoru je úměrné frakci letality
příslušného nádoru. Přiřadí-li se k těmto koeficientům rizika i koeficienty rizika
dědičných změn u potomstva, obdrţí se hodnoty uvedené v tabulce 5.
Pravděpodobnost
indukce fatálního
Nádoru [10-2
Sv-1
]
Souhrnná zdravotní
újma [10-2
Sv-1
]
Tkáň nebo orgán obyvatelé pracovníci obyvatelé pracovníci
močový měchýř 0,30 0,24 0,29 0,24
kostní dřeň 0,50 0,40 1,04 0,83
kostní povrch 0,05 0,04 0,07 0,06
mléčná ţláza 0,20 0,16 0,36 0,29
tlusté střevo 0,85 0,68 1,03 0,82
játra 0,15 0,12 0,16 0,13
plíce 0,85 0,68 0,80 0,64
jícen 0,30 0,24 0,24 0,19
vaječník 0,10 0,08 0,15 0,12
kůţe 0,02 0,02 0,04 0,03
ţaludek 1,10 0,88 1,00 0,80
štítná ţláza 0,08 0,06 0,15 0,12
ostatní tkáně 0,50 0,40 0,59 0,47
tkáně a orgány celkem 5,0 4.0 5,92 4,74
gonády: pravděpodobnost
dědičných poruch
1,00 0,60 1,33 0,80
souhrnně celkem 7,30 5,60
Tab. 5. Nominální koeficienty rizika pro jednotlivé orgány a tkáně.
Poznámka: Uvedené hodnoty se vztahují na populaci se stejným počtem osob
obojího pohlaví a širokým věkovým rozpětím.

29
V této tabulce jsou uvedeny zvlášť hodnoty pro obyvatele a pracovníky. Rozdíly
jsou způsobeny odlišnou věkovou strukturou těchto dvou kategorií. Postup
výpočtu pro genetické účinky diferencovaně pro pracovníky a obyvatelstvo byl
vysvětlen výše. Pokud jde o nádory, je niţší pravděpodobnost fatálního nádoru u
pracovníků způsobena především odečtením podílu dětí a mladistvých na riziku
obyvatelstva, neboť koeficienty rizika jsou v nízkých věkových kategoriích
výrazně vyšší. Toto konstatování o věkové závislosti platí především pro
koeficienty relativního rizika, zatímco koeficienty absolutního rizika mohou
vykazovat i opačný trend. Je to patrné i z tabulek v publikaci [UNSCEAR
1994]. Analýza věkové závislosti je přitom komplikována i tím, ţe je třeba
respektovat i další proměnnou, jíţ je doba od expozice.
Uvedený souhrn dat je i podkladem pro odvození zaokrouhlených hodnot
tkáňových váhových faktorů wT, které se vztahují k referenční populaci se
shodným počtem příslušníků obojího pohlaví a širokým věkovým rozpětím.
Vyuţití těchto faktorů k výpočtu efektivní dávky je vyloţeno v první kapitole.
Vztah dávky a účinku – závěrečné shrnutí poznatků
Je nyní na místě shrnout a zobecnit soudobé poznatky o vztahu mezi dávkou a
biologickým účinkem, vzhledem ke zdravotnímu vlivu záření na člověka s cílem
vyuţít je k plánování a řízení radiační ochrany. Vedle rekapitulace hlavních
charakteristik účinků deterministických a stochastických je třeba poukázat na
některá omezení tohoto schematického dělení. Vztah dávky a účinku pro
projevy deterministické a stochastické je schématicky znázorněn na obrázku 4.

30
Pro deterministické účinky je charakteristická prahová závislost na dávce.
Kvantitativním ukazatelem, pomocí něhoţ lze odhadnout moţné následky
ozáření, je prahová dávka pro ten který účinek, popřípadě hodnota 50 %
efektivity vztaţená ke zvolené referenční době po ozáření, tedy např. LD 50/30.
Důleţitým rysem této skupiny účinků je měnící se klinický obraz se stoupající
dávkou, jinak řečeno, intenzita projevů a jejich zdravotní závaţnost je závislá na
dávce. Jednotlivé stupně postiţení mohou být popsány svou vlastní esovitou
křivkou. Klinické obrazy jsou do značné míry charakteristické, tzn. nekopírují
stavy obvyklé u jiných nemocí, odlišují se zejména svým časovým průběhem,
prostorovou distribucí postiţení a svéráznou kombinací jednotlivých příznaků.
Základním patogenetickým mechanismem je sníţení počtu buněk v terčové
populaci. Z frakce kmenových buněk, které zůstaly ţivotaschopné, je moţná
repopulace postiţené tkáně. Esovitý tvar křivky a existence dávkového prahu
jsou odrazem skutečnosti, ţe v terčové buněčné populaci je poměrně značná
funkční rezerva, tzn. pokles buněčnosti se stoupající dávkou zprvu nezpůsobí
ţádné funkční důsledky a hodnocený efekt nastane aţ při překročení dávek
prahových. Většina tkání v těle poţadavek funkční rezervy splňuje. Výjimkou je
muţský pohlavní epitel, kde odchylky směrem k niţší buněčnosti nejsou
výjimkou, jak o tom svědčí určitá frekvence muţské sterility v populaci. Za této
situace není hodnota prahu v populaci ostrá, práh je neurčitý, jak je ukázáno i na
obrázku 4. Bývá kladena také otázka, např. v soudních sporech, zda je moţno
připustit v populaci existenci zvláště vnímavých jedinců, zda existuje něco jako
alergie na ionizující záření. Takovou představu nelze na úrovni
deterministických účinků záření podpořit. Jsou sice známy velmi vzácné
vrozené chorobné stavy jako je tzv. ataxia-teleangiectasia, závaţné
onemocnění projevující se uţ v dětském věku neurologickými příznaky a
rozšířením cévek na spojivce a v obličeji, vzácná Fanconiho anemie a v dalším
pořadí i Downův syndrom. U těchto stavů je prokázána závaţná porucha
reparačních mechanismů na molekulární úrovni, a s tím související vyšší
radiosenzitivita. Jde o nemocné osoby zpravidla s mentálními defekty, které
běţně nejsou způsobilé k soustavné výdělečné práci, takţe profesionální
expozice záření, a s tím související ohroţení zdraví u nich nepřipadá v úvahu.
Pro stochastické účinky je vztah dávky a účinku odvozován z
epidemiologických studií. Bylo ukázáno, ţe pro oblast dávek, které jsou
předmětem zájmu radiační ochrany, je na podkladě těchto dat moţné formulovat
hypotézu o bezprahovém a lineárním vztahu mezi dávkou a účinkem. Kaţdé
zvýšení dávky je spojeno s úměrným zvýšením pravděpodobnosti pozdních
změn vázaných na ozářenou tkáň nebo orgán a tato závislost platí i pro oblast
dávek nejniţších. Za předpokladu platnosti této hypotézy jsou vhodnými
ukazateli pro odhad závaţnosti důsledků ozáření koeficienty rizika pro
jednotlivé orgány a tkáně i souhrnně pro celotělové ozáření. Stochastický
charakter účinků spočívá v tom, ţe lze předpovědět jen vzestup výskytu těchto
chorobných projevů v ozářené populaci, nikoliv však onemocnění u konkrétního

31
jedince. Nejde také rozpoznat u ţádného jednotlivce postiţeného nádorem či
vrozenou afekcí, zda právě v jeho případě jde o důsledek ozáření. Nádory a
genetické projevy vyvolané ozářením se totiţ neliší od obdobných poruch zdraví
vznikajících spontánně v obecné populaci. U stochastických účinků není
vyjádřena závislost intenzity projevů na dávce, tj. stoupající dávky nevyvolávají
vznik agresivnějších malignit či výrazněji deformujících malformací. Také pro
tuto kategorii radiačního poškození je třeba posoudit otázku individuální
vnímavosti. Při nedostatku vědeckých podkladů nebyl v minulosti důvod - např.
ve znaleckých posudcích k soudním sporům - přihlíţet k moţnosti
individuálních rozdílů, pokud jde o vnímavost ke vzniku nádorů po ozáření a na
otázku soudu kladenou v tomto směru nebylo moţno dát určitou odpověď. V
posledních desítiletích s objevem onkogenů a získáním dalších poznatků je třeba
připustit, ţe v genomu člověka existují geny, či skupiny genů (clusters), které k
rozvoji zhoubného nádoru predisponují (genetické rizikové faktory). Distribuce
těchto genetických faktorů v prostoru a čase je řízena biologickými a sociálními
procesy souvisejícími s reprodukcí (biologické rizikové faktory). V nedávno
publikované studii se uvádí, ţe bylo nalezeno 641 genetických odchylek
souvisejících s nádory, z nichţ bylo 495 lokalizováno do 120 shluků v lidském
genomu [Doerffer 1998]. Jak také tito autoři uvádějí, princip indukce maligního
onemocnění spočívá v rozšíření heterozygotního zastoupení příslušného genu na
zastoupení homozygotní. Tj. zatímco u predisponovaného jedince je kritický gen
jenom jednou zastoupen v páru genů (heterozygotní zastoupení), u postiţeného
nádorem jsou oba elementy páru tvořeny kritickým genem (homozygotní
zastoupení). Je známo několik subcelulárních mechanismů vedoucích cestou
mutace k rozšíření heterozygotního zastoupení, všechny z nich mohou být
ovlivněny ionizujícím zářením. U heterozygotů nejsou zpravidla přítomny
zjevné projevy této dispozice (u některých geneticky podmíněných chorob se
uplatňují tito nositelé jako přenašeči choroby) a epidemiologické studie se
mohou opírat jen o odkrytí homozygotů v populaci, tj. osob se zjevnými projevy
zdravotní poruchy. To by platilo i pro maligní nádory pokud máme na mysli
jejich genetický podklad. Sledování frekvence nosičů heterozygotně
zastoupených genů se opírá o molekulárně genetické a cytologické metody. Tak
by se mohla připravit půda pro molekulární epidemiologii rizika nádorů. Jako
další spekulativní krok se konstruují modely genetických nádorových rizikových
faktorů s cílem zmenšit nejistoty v hodnocení vnímavosti na ozáření a přispět k
vyššímu standardu radiační ochrany. Při současných poznatcích jsou nejistoty
vyplývající z biologických rizikových faktorů asi patnáctkrát větší neţ nejistoty
dozimetrického hodnocení. Vychází-li se z 20 % chyby dozimetrických dat,
znamenalo by to 300 % chybu odhadu vlivu predisponujících genetických
faktorů. Pro hodnocení rizika radiogenních nádorů u jednotlivců je tedy zatím
tento přístup nepouţitelný.
Hypotetický charakter lineárního a bezprahového vztahu je důvodem
opakovaného úsilí o precizování zákonitostí týkajících se oblasti nízkých dávek.
Výsledky takových studií a úvah jsou nejen předmětem odborného zájmu

32
biofyziků, ale jsou očekávány s mimořádnou pozorností plánovači a realizátory
rozsáhlých jaderných programů a s nimi svázanými ekonomickými a
politickými kruhy. Popření bezprahové závislosti a příklon k představě o
neškodnosti malých dávek ozáření by vedl k úsporám sotva vyčíslitelným a ke
zcela nové argumentaci v diskusích o ochraně ţivotního prostředí. Polemika k
těmto otázkám, zastřešená jako problematika malých dávek ionizujícího
záření, se tématizuje do několika okruhů, věcně spolu souvisejících:
. příznivý účinek malých dávek (radiační hormeze),
. existence či neexistence prahu pro stochastické účinky.
Radiační hormeze není charakterizována jednotně. Někdy se jí rozumí přiznivý
účinek záření, jindy spíše stimulační účinek (coţ není totéţ). Pro některé
fenomeny platí, ţe jde o účinek opačného směru, neţ je působení dávek velkých.
Je nesporné, ţe řada experimentálních pozorování ukázala jevy, které lze do této
skupiny zařadit. Byla prokázána vyšší snůška vajec po ozáření, zvětšila se
velikost bramborových hlíz, plodnost u některých ţivočišných druhů, u člověka
byl prokázán příznivý vliv malých dávek záření např. na zmírnění problémů
spojených s kloubní artritídou. U této kategorie jevů je "příznivý" efekt
zjišťován za určitých podmínek ozáření a v určitém časovém intervalu po
ozáření. Hormezi je moţné pokládat za vedlejší produkt regulačních
mechanismů vyvolaných ozářením. Tento jev není v rozporu s představou, ţe
záření můţe způsobovat s určitou pravděpodobností vţdy jen inaktivaci určité
struktury (a s ní spojené funkce). Je-li vyřazen regulátor typu supresoru, je
výsledkem odbrzdění daného směru reakce. Hormeze není fenoménem
univerzálním, platným pro všechny účinky a pro většinu pozorování. U radiačně
indukovaných nádorů by projevem hormeze byl pokles jejich výskytu pod
úroveň frekvence spontánní. Byly publikovány studie, které se pokoušejí
takovou závislost doloţit, nejsou však obecně pokládány za průkazné. Tento
okruh otázek vedl v posledních letech k diskusím, které byly v roce 1994
zhodnoceny souhrnně ve zprávě Výboru Spojených národů pro účinky
atomového záření [UNSCEAR 1994] pod záhlavím "Adaptivní odezva na
ozáření buněk a organismů".
Adaptivní odpověď je souhrnný název uţívaný pro popis výsledků
experimentů, ve kterých malé dávky ionizujícího záření navodily reparační
procesy nebo stimulaci buněčného dělení. U dělících se buněčných populací je
přesvědčivě doloţeno, ţe jednorázové ozáření řídce ionizujícím zářením v
dávkách 1 - 3 Gy vede k niţšímu výskytu chromosomových aberací a mutací v
případě, kdyţ v intervalu několika hodin předcházelo ozáření v dávkách
jednotek aţ desítek miligrayů. Existují doklady o tom, ţe po ozáření takovými
dávkami probíhá v buňkách řada změn, ke kterým patří aktivace několika skupin
genů, včetně těch, které kódují syntézu enzymů angaţovaných v řízení
buněčného cyklu, proliferace a reparačních procesů. Adaptivní odezva buněk se
chápe jako vícestupňový děj. Ozáření vyvolá zpomalení buněčného cyklu, takţe

33
před vstupem do další mitózy se některá poškození mohou opravit. Byly
navrţeny i alternativní mechanismy adaptace. Tak např. se uvaţuje o detoxikaci
reaktivních radikálů, o aktivaci membránových receptorů stimulujících
proliferaci, nebo snad i o účasti imunitních mechanismů. Jedním z důsledků
reparace DNA by mohlo být i sníţení spontánní incidence nádorů, nebo sníţení
přídatného výskytu nádorů po ozáření. Většina epidemiologických studií o
výskytu nádorů v důsledku malých dávek však má malou sílu statistické
výpovědi, takţe je velmi obtíţné dospět k závěrům o přítomnosti nebo
nepřítomnosti adaptivní odpovědi. Očekává se, ţe teprve další bádání jak
molekulární a cytologická, tak i epidemiologická mohou otázku blíţe objasnit.
Zatím je předčasné přijmout názor, ţe buněčná adaptivní odpověď způsobuje
takový příznivý efekt, který by převáţil nad nepříznivými účinky ozáření
malými dávkami řídce ionizujícího záření.
Kritika bezprahové závislosti stochastických účinků na dávce je součástí širší
otázky platnosti hypotézy linearity a bezprahovosti. Přitom oprávněnost
předpokladu o linearitě v oblasti dávek, pro niţ jsou k dispozici
experimentální podklady, je celkem zřejmá. Výsledky pozorování ať uţ
experimentálních, nebo epidemiologických se vyjadřují body v systému
souřadnic, vůči nimţ lze testovat přiléhavost předem zvolených funkcí. Pro
většinu pozorování je nalezena dobrá shoda pro předpoklad přímé úměrnosti
mezi dávkou a účinkem. Existují ovšem pozorování, kde směrem k niţším
hodnotám experimentální body spadají pod proloţenou přímku; pro tyto situace
představuje linearita přecenění skutečného rizika. Naproti tomu jsou i velmi
ojedinělá pozorování, kde experimentální body směrem k niţším dávkám se
dostávají nad přímku; zde hypotéza linearity skutečnou situaci podceňuje. Je
třeba zdůraznit, ţe hypotézu linearity a bezprahovosti nelze chápat jako nějakou
radiobiologickou zákonitost. Byla formulována pro potřeby radiační ochrany
jako nejlepší v současné době moţné zobecnění dostupných dat a v souladu s
názorem, ţe její aplikace je spojena v oblasti malých dávek spíše s
nadhodnocením reálné situace a že přitom zůstává jen malá pravděpodobnost
možného podhodnocení. Při rozboru jednotlivých radiobiologických či
epidemiologických studií bývá zjišťován při větším rozsahu dávek i vztah
kvadratický, lineárně-kvadratický, popřípadě vykazující při dávkách nejvyšších i
pokles efektu vlivem letálního účinku na buňky.
Oprávněnost extrapolace lineárního vztahu směrem k dávkám nejmenším
vyvolává problémy, které směřují aţ ke zpochybňování bezprahové závislosti
pro stochastické účinky. Vztah mezi dávkou a účinkem, který je v určitém
pásmu dávek zřejmý a statisticky významný, se směrem k niţším dávkám
rozmazává v důsledku narůstajících fluktuací nejistot, a konečně se dospěje k
bodu, kde účinek, i kdyby existoval, není statisticky prokazatelný. Lze ukázat,
ţe počet jedinců v populaci potřebný k průkazu statistické významnosti roste se
čtvercem průměrné individuální dávky potřebné k vyvolání efektu. Kdyby jev
vyţadující průměrnou dávku 1 Gy byl prokazatelný v kolektivu několika tisíců
lidí, pak účinek podmíněný dávkou 0,1 Gy by vyţadoval populaci statisícovou.

34
Z hlediska biofyzikálních zákonitostí je koncept bezprahovosti v dobré shodě s
teoretickými představami. Při malých dávkách či malých dávkových příkonech
je velmi nepravděpodobné, ţe se uskuteční více neţ jedna ionizační událost v
příslušném úseku DNA v době, neţ proběhne reparační děj. Přihlédne-li se k
vlastnostem Poissonovy distribuce ionizačních událostí, jejímu malému
exponentu a malé frakci neúspěšných reparací, očekávaný průběh vztahu bude
lineární. Pro vysoké dávky vede obdobná úvaha ke vztahu kvadratickému. Dále
je třeba v argumentaci přihlédnout k tomu, ţe sama existence přírodního
radiačního pozadí relativizuje význam vztahu dávky a účinku pro dávky blíţící
se nule. Vůči zastáncům existence prahové závislosti i pro stochastické účinky je
třeba čelit námitce, ţe extrapolace je operací nevědeckou. Naopak sama tato
námitka je nevědecká, vţdyť např. objev některých planet je důsledkem
modelování na podkladě jiných pozorování [Beninson 1996]. Zajímavý
argument ve prospěch oprávněnosti aplikace bezprahové závislosti v radiační
ochraně, navazující na úvahy o genetické vnímavosti na vznik nádorů, přinesl
[Gentner 1997]. Je známo, ţe vyšší výskyt onkogenů v genomu má vztah ke
vzniku nádorů. Předpokládá se, ţe velká část spontánních nádorů je důsledkem
nakupení takových genů. Lze také očekávat, ţe riziko jak spontánního, tak i
radiogenního nádoru je v populaci rozděleno nerovnoměrně. Jedinec s
hypotetickým průměrným rizikem vlastně neexistuje. Autor dospívá k názoru,
ţe i kdyţ riziko nádoru můţe klesat u některých jednotlivců k nule, budou
existovat vţdy také jedinci výrazně predisponovaní ať uţ ke spontánní, nebo
radiačně indukované rakovině. Znamená to, ţe vţdy bude existovat kategorie
vnímavých jedinců, ať uţ dávka, která je můţe ohrozit, bude jakkoliv nízká, a to
je třeba při řízení radiační ochrany respektovat. U heterogenních populací bylo
teoreticky vyvozeno, ţe biologická variabilita vede k lineárnímu vztahu dávky a
účinku.
Z výše uvedeného je zřejmé, ţe hypotéza lineární a bezprahové závislosti na
dávce představuje pro radiační ochranu v současné době jedinou praktickou
alternativu, která je s vysokou pravděpodobností dostatečně bezpečná a při
správné aplikaci dovoluje vyhnout se iracionálním poţadavkům na ochranu. Je
známo, ţe uţ řadu let patří jaderná zařízení a jiné zdroje ionizujícího záření k
provozům mimořádně bezpečným. I odborníci radiační ochrany jsou si vědomi,
ţe usměrnění ekonomických zdrojů do jiných sociálních a zdravotních programů
by zachránilo daleko víc ţivotů a zlepšilo výrazněji kvalitu ţivota lidí, neţ
prostředky neúčelně vkládané do nevhodně směrovaných opatření v radiační
ochraně. Nelze v této souvislosti odhlédnout od tlaku veřejného mínění, v němţ
se fixovalo povědomí, ţe i sebemenší dávce je třeba za kaţdou cenu zabránit a
které si vysoký standard radiační ochrany vynucuje. Potom se i do
matematických vztahů k posouzení rovnováhy mezi přínosy a náklady
vynakládanými na opatření v radiační ochraně k objektivní zdravotní újmě musí
přiřadit také újma subjektivní, psychologická, emotivní, která můţe být oceněna
sumou téměř bezbřehou. V diskusích o skutečném zdravotním významu
nejniţších dávek se můţe vyuţít i argument, ţe variabilitu přírodního radiačního

35
pozadí v rámci geografických podmínek nikdo ve vztahu k moţnému ohroţení
zdraví nevnímá. Potom ani dávky v úrovni jednotek procent přírodního pozadí
nemohou vyţadovat nepřiměřeně nákladnou regulaci. Lze tedy uzavřít, ţe
aplikace hypotézy linearity a bezprahovosti v radiační ochraně neznamená
automaticky další zvyšování nákladů na ochranu.













![ORIGINAL RESEARCH Open Access Estimation of absorbed …dosimetry of 177Lu-octreotate and of relevant radiobio-logical and toxicity factors [4,5,11]. Detailed dosimetric calculations](https://img.pdfslide.net/doc/110x75/612254fae33dd419c37869d7/original-research-open-access-estimation-of-absorbed-dosimetry-of-177lu-octreotate.jpg)















