Embed Size (px)
Citation preview

PART IVOther Topics

28 Membrane Potential andDonnan Potential
28.1 INTRODUCTION
When two electrolyte solutions at different concentrations are separated by an ion-
-permeable membrane, a potential difference is generally established between the
two solutions. This potential difference, known as membrane potential, plays an
important role in electrochemical phenomena observed in various biomembrane
systems. In the stationary state, the membrane potential arises from both the diffu-
sion potential [1,2] and the membrane boundary potential [3–6]. To calculate the
membrane potential, one must simultaneously solve the Nernst–Planck equation
and the Poisson equation. Analytic formulas for the membrane potential can be de-
rived only if the electric field within the membrane is assumed to be constant [1,2].
In this chapter, we remove this constant field assumption and numerically solve the
above-mentioned nonlinear equations to calculate the membrane potential [7].
28.2 MEMBRANE POTENTIAL AND DONNAN POTENTIAL
Imagine the stationary flow of electrolyte ions across a membrane separating solu-
tions I and II that contain uni–uni symmetrical electrolyte at different concentrations
nI and nII, respectively. We employ a membrane model [7] in which each side of the
membrane core of thickness dc is covered by a surface charge layer of thickness ds.In this layer, membrane-fixed negative charges are assumed to be distributed at a
uniform density N. We take the x-axis perpendicular to the membrane with its origin
at the boundary between the left surface charge layer and the membrane core
(Fig. 28.1). The potential in the bulk phase of solution II is set equal to zero.
The potential in bulk solution I is thus equal to the membrane potential, which
we denote by Em. First, consider the regions x< 0 and x> dc. We assume that the
distribution of electrolyte ions in the surface charge layer and in solutions I and II is
scarcely affected by ionic flows so that it is in practice at thermodynamic equili-
Biophysical Chemistry of Biointerfaces By Hiroyuki OhshimaCopyright# 2010 by John Wiley & Sons, Inc.
535

brium. Accordingly, the potential c(x) in these regions is given by the Poisson–
Boltzmann equations, namely,
d2cdx2
¼ 2enIereo
sinheðc� EmÞ
kT
� �; x < �ds ð28:1Þ
d2cdx2
¼ 2enIereo
sinheðc� EmÞ
kT
� �þ eN
ereo; � ds < x < 0 ð28:2Þ
d2cdx2
¼ 2enIIereo
sinheckT
� �þ eN
ereo; dc þ ds < x < dc ð28:3Þ
d2cdx2
¼ 2enIIereo
sinheckT
� �; x > dc þ ds ð28:4Þ
where er is the relative permittivity of solutions I and II. Next, consider the region
0< x< dc (i.e., the membrane core). We regard this region as a different phase from
the surrounding solution phase and introduce the ionic partition coefficients
between the membrane core and the solution phase. We denote by b+ and b�,respectively, the partition coefficients for cations and anions. The electric potential
c(x) in this region is given by the following Poisson equation:
d2cdx2
¼ e
e0reonþðxÞ � n�ðxÞf g; 0 < x < dc ð28:5Þ
FIGURE 28.1 Model for a charged membrane. The membrane core (of thickness dc) iscovered by a surface charge layer (of thickness ds). The membrane-fixed charges are assumed
to be negative and are encircled.
536 MEMBRANE POTENTIAL AND DONNAN POTENTIAL

where nþ(x) and n�(x) are the respective concentrations of cations and anions at
position x (0< x< dc) and e0r is the relative permittivity of the membrane core. The
flow of cations, denoted by Jþ, can be expressed as the product of the number den-
sity nþ(x) and velocity vþ of cations. Similarly, the flow J� of anions can be
expressed as the product of the number density n�(x) and velocity v� of anions.
That is,
J� ¼ n�v� ð28:6Þ
The ionic velocities v� are given by
v� ¼ � 1
l�
dm�dx
ð28:7Þ
Here, l� are the ionic drag coefficients of cations and anions, respectively, and
m� are the electrochemical potentials of cations and anions, respectively, which are
m� ¼ mo� þ kT lnn�ðxÞ � zecðxÞ ð28:8Þ
From Eqs. (28.6)–(28.8), we obtain
J� ¼ �D�dn�dx
� n�ekT
dcdx
� �ð28:9Þ
with
D� ¼ kT
l�ð28:10Þ
where D+ and D� are the diffusion coefficients of cations and anions, respectively,
in the membrane core. In the stationary state, div J�=dJ�/dx¼ 0 and the net elec-
tric current must be zero in the absence of external field so that
Jþ ¼ J� ¼ constant ðindependent of xÞ ð28:11Þ
The boundary conditions for c(x) are
cðxÞ ! 0 as x ! þ1 ð28:12Þ
cðxÞ ! Em as x ! �1 ð28:13Þ
The other boundary conditions for c(x) are continuity conditions of c(x) and the
electric displacement at x¼�ds, 0, dc, and dcþ ds. The solution of coupled equa-
tions, Eqs. (28.1)–(28.5), (28.9) and (28.11) determines the whole potential profile
across a membrane.
MEMBRANE POTENTIAL AND DONNAN POTENTIAL 537
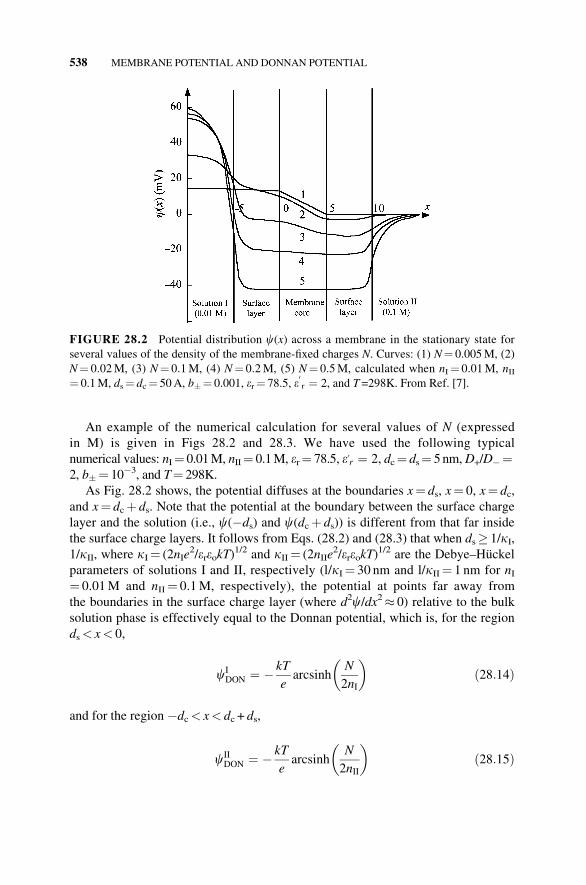
An example of the numerical calculation for several values of N (expressed
in M) is given in Figs 28.2 and 28.3. We have used the following typical
numerical values: nI¼ 0.01M, nII¼ 0.1M, er¼ 78.5, e0r ¼ 2, dc¼ ds¼ 5nm,D+/D�¼2, b�¼ 10�3, and T¼ 298K.
As Fig. 28.2 shows, the potential diffuses at the boundaries x¼ ds, x¼ 0, x¼ dc,and x¼ dcþ ds. Note that the potential at the boundary between the surface charge
layer and the solution (i.e., c(�ds) and c(dcþ ds)) is different from that far inside
the surface charge layers. It follows from Eqs. (28.2) and (28.3) that when ds� 1/kI,1/kII, where kI¼ (2nIe
2/ereokT)1/2 and kII¼ (2nIIe
2/ereokT)1/2 are the Debye–Huckel
parameters of solutions I and II, respectively (l/kI¼ 30 nm and l/kII¼ 1 nm for nI¼ 0.01M and nII¼ 0.1M, respectively), the potential at points far away from
the boundaries in the surface charge layer (where d2c/dx2� 0) relative to the bulk
solution phase is effectively equal to the Donnan potential, which is, for the region
ds< x< 0,
cIDON ¼ � kT
earcsinh
N
2nI
� �ð28:14Þ
and for the region �dc< x< dc + ds,
cIIDON ¼ � kT
earcsinh
N
2nII
� �ð28:15Þ
FIGURE 28.2 Potential distribution c(x) across a membrane in the stationary state for
several values of the density of the membrane-fixed charges N. Curves: (1) N¼ 0.005M, (2)
N¼ 0.02M, (3) N¼ 0.1M, (4) N¼ 0.2M, (5) N¼ 0.5M, calculated when nI¼ 0.01M, nII¼ 0.1M, ds¼ dc¼ 50A, b�¼ 0.001, er¼ 78.5, e
0r ¼ 2, and T =298K. From Ref. [7].
538 MEMBRANE POTENTIAL AND DONNAN POTENTIAL

In this case, by integrating Eqs. (28.1)–(28.4) once and using the appropriate
boundary conditions, we have
cð�dsÞ ¼ cIDON � kT
etanh
ecIDON
2kT
� �þ Em ð28:16Þ
cðdc þ dsÞ ¼ cIIDON � kT
etanh
ecIIDON
2kT
� �ð28:17Þ
which relate the membrane surface potentials c(�ds) and c(dc + ds) to the Donnan
potentials cIDON and cI
DON. When the potential far inside the surface charge layer is
effectively equal to the Donnan potential, the membrane potential can be expressed as
Em ¼ fcð0Þ � cðdcÞg þ fcIIDON � cI
DONg þ ½fcIDON þ Em � cð0Þg
�fcIIDON � cðdcÞg� ð28:18Þ
where the first term enclosed by brackets on the right-hand side corresponds to
the diffusion potential, the second to the contribution from the Donnan potentials in
the surface charge layers, and the third to that from deviation in the boundary poten-
tials at x¼ 0 and x¼ dc due to the ionic flows. The contribution from the third term
is normally very small so that the diffusion potential E can practically be regarded as
arising from the diffusion potential and the Donnan potential. Note that the
FIGURE 28.3 Membrane potential Em, contributions from the diffusion potential Ed
and the Donnan potential difference DcDON ¼ cIIDON � cI
DON as functions of the density of
membrane-fixed charges N. The values of the parameters used in the calculation are the same
as those in Fig. 28.2. The dashed line is the approximate result for Em (Eq. (28.21)). As
N!1, Em tends to the Nernst potential for cations (59mV in the present case). From Ref. [7].
MEMBRANE POTENTIAL AND DONNAN POTENTIAL 539

Donnan potentials (Eqs. (28.14) and (28.15)) are independent of the surface layer
thickness ds and therefore the membrane potential depends little on ds, provided that
ds� 1/kI, 1/kII.Figure 28.2 (in which the potential far inside the surface layer is almost equal to
the Donnan potential) shows how the contributions from the diffusion potential and
from the Donnan potential change with density N of the membrane-fixed charges.
For very low N, the potential in the surface charge layer differs little from that in
the bulk solution phase (the Donnan potential itself is very small) and the mem-
brane potential is mostly due to the potential gradient in the membrane core. In
other words, the membrane potential is almost equal to the diffusion potential. As
N is increased, however, the potential gradient in the membrane core becomes
small, which means that the contribution from the diffusion potential decreases. In-
stead, the potential difference between the surface charge layer and the surrounding
solution becomes appreciable and the membrane potential for very large N is deter-
mined almost solely by the potential difference in each surface charge layer. As
stated before, the potential difference is in practice equal to the Donnan potential in
each surface charge layer if ds� 1/kI, 1/kII so that the membrane potential Em for
large N is expressed as
Em ¼ cIIDON � cI
DON
¼ kT
eln
ðN=2nIÞ þ fðN=2nIÞ2 þ 1g1=2
ðN=2nIIÞ þ fðN=2nIIÞ2 þ 1g1=2" #
þ cIIs � cI
s
ð28:19Þ
To illustrate more clearly the above-mentioned N dependence, we have dis-
played in Fig. 28.3 the contributions from the diffusion and Donnan potentials to
the membrane potential as a function of N.The decrease of the contribution from the diffusion potential with increasing N
can be explained as follows. For large N, that is, for the case where the surface
layers are highly negatively charged, coions (i.e., anions) are repelled by the mem-
brane so that the membrane is in practice impermeable to coions and their flow
within the membrane, J�, decreases. Since the net electric current must be zero (Jþ� J�¼ 0), the flow of counterions (i.e., cations), Jþ, must decrease to the same
extent as J�. That is, all ionic flows decrease, resulting in a smaller diffusion poten-
tial. In the limit of N!1, the membrane becomes fully impermeable to anions and
an equilibrium is reached with respect to cations so that the membrane potential
tends to the Nernst potential for cations, namely,
Em ! kT
eln
nIInI
� �ð28:20Þ
which is obtained by taking the limit N!1 in Eq. (28.19). In this limit, the mem-
brane behaves like a semipermeable membrane (permeable to cations only). Note
that Eq. (28.20) also holds when b�! 0, irrespective of the value of N.
540 MEMBRANE POTENTIAL AND DONNAN POTENTIAL

As shown in Fig. 28.2, the potential gradient in the membrane core is slightly
concave (see curve 3, in particular). However, deviation from linearity is seen to be
small in Fig. 28.2. This means that the usual assumption of constant field within the
membrane is not a bad approximation. If we employ this approximation, Eq. (28.9)
can easily be integrated in the following two limiting cases. When ds� 1/kI, 1/kII,in which case the potential far inside the membrane surface layer is in practice the
Donnan potential, the constant field assumption yields [3]
Em ¼ kT
eln
PþnII expð�ecIIDON=kTÞ þ P�nI expðecI
DON=kTÞPþnI expð�ecI
DON=kTÞ þ P�nII expðecIIDON=kTÞ
" #þ cII
DON � cIDON
ð28:21Þ
with
P� ¼ b�D�=dc ð28:22Þ
where Pþ and P� are the permeabilities of cations and anions, respectively. The
approximate result calculated from Eq. (28.21) is plotted in Fig. 28.3 in comparison
with the exact numerical results. A good agreement is obtained: the relative error is
less than a few percent. On the other hand, in the limit ds! 0 with Nds kept con-stant, we can then obtain the following formula [5,6]:
Em ¼ kT
eln
PþnII expð�ecIIs =kTÞ þ P�nI expðecI
s=kTÞPþnI expð�ecI
s=kTÞ þ P�nII expðecIIs =kTÞ
" #þ cII
s � cIs ð28:23Þ
where
cIs ¼
2kT
earcsinh
s
ð8nIereokTÞ1=2" #
ð28:24Þ
cIIs ¼ 2kT
earcsinh
s
ð8nIIereokTÞ1=2" #
ð28:25Þ
Equations (28.24) and (28.25) are the Gouy–Chapman double-layer potential of
the membrane surface with a surface charge density s¼�eNds.
REFERENCES
1. D. E. Goldman, J. Gen. Physiol. 27 (1943) 37.
2. A. L. Hodgkin and B. G. Katz, J. Physiol. 108 (1949) 37.
REFERENCES 541

3. M. J. Polissar, in: F. H. Johnson, H. Eyring, and M. J. Polissar (Eds.), Kinetic Basis ofMolecular Biology, Wiley, New York, 1954, p. 515.
4. S. Ohki, in: J. F. Danielli (Ed.), Progress in Surface and Membrane Science, Vol. 10,Academic Press, New York, 1976, p. 117.
5. S. Ohki, Phys. Lett. 75A (1979) 149.
6. S. Ohki, Physiol. Chem. Phys. 13 (1981) 195.
7. H. Ohshima and T. Kondo, Biophys. Chem. 29 (1988) 277.
542 MEMBRANE POTENTIAL AND DONNAN POTENTIAL
![Colloids and Surfaces B: Biointerfaces Colloids Surfaces B... · Colloids and Surfaces B: Biointerfaces 116 (2014) ... antibiotics [3–6]. Their broad ... Alamethicin is most effective](https://img.pdfslide.net/doc/110x75/5a94ecce7f8b9a9c5b8c50e4/colloids-and-surfaces-b-colloids-surfaces-bcolloids-and-surfaces-b-biointerfaces.jpg)



![Colloids and Surfaces B: Biointerfaces - CAS · Colloids and Surfaces B: Biointerfaces 88 (2011) ... such as medicine/pharmacy [1–3], chemical engineering ... styrene as co-monomer](https://img.pdfslide.net/doc/110x75/5b2550217f8b9af0278b4666/colloids-and-surfaces-b-biointerfaces-colloids-and-surfaces-b-biointerfaces.jpg)
























