Embed Size (px)
Citation preview

SEMINARIO DE BIOTECNOLOGÍA Y
BIOSEGURIDAD DE OGM’S
BASES DEL MEJORAMIENTO
GENÉTICO DE PLANTAS
CCC - CIBIOGEM
Dr. Netzahualcoyotl Mayek-Pérez Centro de Biotecnología Genómica - IPN

Fuentes: Elaboración propia a partir, principalmente, del documento
intitulado ‘MONOGRAFÍA DEL CURSO DE GENOTECNIA’ del DR.
FERNANDO RAMOS GOURCY, Profesor Titular de la UNIVERSIDAD AUTÓNOMA DE AGUASCALIENTES. Disponible en el sitio electrónico
http://www.fernandoramos.net/genotecnia/genotecnia.pdf
Documentos adicionales:
Márquez-Sánchez F. 1985. Genotecnia Vegetal. AGT Editor. México.
Molina-Galán, JD. 1992. Introducción a la Genética de Poblaciones y
Cuantitativa. AGT Editor. México.
Brauer-Herrera, O. 1987. Fitogenética Aplicada. LIMUSA. México.

FITOMEJORAMIENTO
Técnicas y prácticas basadas en genética para mejorar la expresión de caracteres heredables desarrollados por la genética vegetal aplicada. 1) Diseñar el arquetipo deseado. 2) Reunir la variabilidad genética posible de la especie. 3) Si no existe, producirla artificialmente mediante mutagénesis, transgénesis, especies cercanas, etc. 4) Aplicar el método mejoramiento que maximice la respuesta a la selección. 5) Generar y evaluar el germoplasma recombinante. 6) Registro y liberación.

genético Importancia del mejoramiento
• Incremento de producción y productividad. • Incremento de calidad, capacidad nutricia, etc. • Mayor Resistencia a factores adversos. • Menor uso de contaminantes (agroquímicos). • Siembra en sitios y/o fechas nuevos. • Etc.

Rendimiento promedio (lb) en EUA
Periodo de Trigo Maíz Avena 10 años
Soya Tabaco
1915-1924 1925-1934 1935-1944
833 1,488 825 1,354 910 1,595
998 884 977
-- 805 1,085
788 779 952
1945-1954 1,027 2,103
1955-1964 1,427 3,102
1,091
1,321
1,199
1,398
1,236
1,712

Rendimientos promedio (kg/ha) de cultivos en
México
Cultivo 1960 1978 Diferencia % de
incremento
Arroz 2,297 3,286 989 43.0
Frijol 398 595 197 49.5
Maíz
Trigo
Soya
975 1,518
1,417 3,483
1,230 1,543
543
2,065
313
55.7
145.8
25.5

genético
Importancia del mejoramiento
En México, desde mediados del siglo XX se han obtenido cientos de variedades de plantas con características mejoradas de rendimiento, calidad y adaptación a diferentes regiones y con resistencia a plagas y enfermedades.
Los trigos enanos y semienanos de alta
productividad, resisten a las royas o chahuixtles y relativamente insensibles al fotoperiodo, han sido una contribución de gran importancia a la agricultura nacional y mundial.
Variedades de papa desarrolladas en México y seleccionadas por su resistencia al tizón tardío, han permitido la utilización de extensas superficies.

Mayor producción

Extensión de superficies

Mayor calidad

Resistencia a factores bióticos y abióticos, etc.

Bosquejo histórico
Thomas Andrew Knight (1759 - 1835) generó
los primeros híbridos.
• Luego Darwin publica El origen y la evolución de
las especies (1859).
• Mendel (1865) publica sus Experimentos de
Hibridación en plantas.

Bosquejo histórico
g) El estudio de la herencia de la resistencia a enfermedades. Los estudios de trigo iniciados en 1902 por Biffen, que culminaron con la formación de variedades de trigo resistentes a royas o chahuistles.
h) La formación de un cereal hecho por el hombre. En 1917, Mc. Fadden describe el híbrido entre dos géneros: Triticum x Secale (trigo x centeno), originando el nuevo cereal conocido como triticale (cruza reportada en 1875 por el escocés Wilson como híbrido estéril y en 1891 por el alemán Rimpau como fértil).

Bosquejo histórico
i) El descubrimiento en maíz de la esterilidad masculina citoplásmica, en 1933 por Rhoades, y en trigo por Kihara en 1951. Trabajos de esterilidad y restauración de fertilidad en maíz, sorgo y trigo hicieron posible la producción de semilla híbrida en escala comercial para aprovechar la heterosis.
j) Híbridos obtenidos entre maíz y sorgo en el CIMMyT.

Centro de origen de los cultivos
El ruso Nikolai Ivanovitch Vavilov, publicó un trabajo en 1926 titulado “Estudio sobre el origen de las plantas cultivadas”.
En estos trabajos, producto de una
colección mundial de plantas llevada a cabo por botánicos y agrónomos rusos dirigidos por Vavilov, se destaca la importancia de las plantas nativas de un lugar y en la adaptación de nuevas especies.

Centro de origen de los cultivos
De interés para los programas de mejora y para la agricultura en general, fue el trazo de un mapa con los 11 centros mundiales de origen de las plantas cultivadas, los cuáles son:
China: Mijo, soja, cebolla, lechuga, chabacano, mandarina.
India: Arroz, garbanzo, pimienta, sorgo, naranja, limón, caña de azúcar, mango.
Península y archipiélago Malayo: Palma de coco, clavo, plátano.
Asia Central: Chícharo, lenteja, haba, espinaca, ajonjolí, cártamo, almendra.

Centro de origen de los cultivos
Cercano oriente: Trigo, cebada, alfalfa, uva.
Mediterráneo: Avena, olivo, col, trigo duro.
Abisinia (Etiopía): Ajonjolí, café, higuerilla, sorgo.
Sur de México y Centroamérica: Fríjol, maíz, papaya, chile, camote, henequén, nopal.
Sudamérica (Perú, Ecuador, Bolivia): Papa, tomate.
Chile: Papa.
Brasil - Paraguay: Árbol de caucho, cacahuete, piña.

Centro de origen de los cultivos

Centro de origen de los cultivos
El centro de origen: área que contiene la mayor concentración de genotipos diferentes de la planta.
Las fuerzas selectivas ambientales operan de la misma manera a lo largo de toda la historia evolutiva de una especie; + tiempo de una entidad En un área dada, + acumulación de mutantes.

Centro de origen de los cultivos
Los centros de origen son también las áreas donde los agentes patógenos y las plagas que atacan a las plantas y que evolucionaron con ellas a través de los tiempos, se encuentran en mayor abundancia, y donde por consiguiente existe un número más elevado de biotipos de unas y otras.
En los centros de origen se va dando un proceso de selección natural que conduce, en el huésped, a la aparición de varios tipos de resistencia, y en el parásito, a la formación de diversos grados de virulencia.

Colecta, caracterización y
conservación de recursos genéticos

Centros de domesticación
La domesticación vegetal, la transformación de una especie silvestre en cultivada por la acción del hombre, ha sido el origen de todas las especies agrícolas.
Donde mayor interés tiene la domesticación es a nivel génico, es decir, la domesticación de genes de especies silvestres para incluirlos en las cultivadas.

Centros de domesticación
Ejemplos:
La resistencia al mosaico de la caña de azúcar (Saccharum officinarum), que se vio amenazada de extinción. Se obtuvieron genes de resistencia de una caña silvestre (Saccharum
spontaneoum).
La papa, sobre todo en el norte de Europa, se vio seriamente amenazada por los ataques de mildiu (Phytophtora infestans). Se obtuvieron genes de resistencia en la planta silvestre Solanum demissum.

Centros de domesticación
El orden de domesticación de diferentes cultivos ha sido:
1.- Cereales
2.- Leguminosas de grano
3.- Hortalizas de ciclo corto
4.- Oleaginosas herbáceas
5.- Raíces
6.- Frutales herbáceos
7.- Textiles
8.- Frutes arbóreos 9.- Industriales.

Centros de domesticación
¿Qué plantas han sido domesticadas?
Cerca de 230 cultivos domesticados,
representando:
180 géneros ...de 3000 géneros de angisopermas – son relativamente pocos.
64 familias ...de 300 – amplio rango, debido a:
Diversidad de sitios ecológicos, diversidad de necesidades humanas, algunas familias han aportado muchos cultivos, por ejemplo: gramíneas, leguminosos, crucíferas, solanáceas.

Centros de domesticación
¿Cómo evolucionaron los cultivos?
algunos mecanismos como la selección natural Selección natural y humana
Selección, ocurre por: reproducción diferencial: algunos individuos tienen
más descendencia que otros en la siguiente generación. Selección natural: ocurre cuando un cultivo es genéticamente heterogéneo.

Domesticación de cultivos
Clase de cambios en selección natural:
• Pérdida de dehiscencia
• Pérdida de dormancia
• Etc.

Domesticación de cultivos
Selección artificial: resulta de decisiones hechas por el humano para mantener la progenie de ciertos parentales.
Clase de cambios: pérdida de estructuras defensivas (espinas,
vellosidad, etc).
Mejora la palatibidad
Reduce la toxicidad
etc.

Diversidad genética
a) Tipos de caracteres
Habitos de crecimiento
Resistencia a plagas y enfermedades
Tolerancia a stress abiótico
calidad
Indices de cosecha y rendimiento
b) Tipos de plantas
Pools o acervos genéticos
Progenitores silvestres, cultivares obsoletos, líneas avanzadas de mejora, cultivares avanzados.

Diversidad genética
¿Cómo se mide la diversidad genética?
a) Caracteres morfológicos (fenotipos), usualmente de importancia agronómica:
Hábitos de crecimiento, tamaño de semilla, color de fruto, fenología, etc.
problema: Expresión de genes, efecto ambiental.
b) Marcadores moleculares:
RFLP, RAPD
problema: Técnicas costosas

Diversidad genética
Efecto potencial por la carencia de diversidad genética en los cultivos:
En 1970, hubo una epidemia en USA, porduciendo 15% de pérdidas en el rendimiento de maíz.
Se inició en Florida y se desplazó hacia las zonas productoras del norte (faja maicera).
La enfermedad en la hoja fue causada por el hongo Helmintosporium maydis
Afectó solo al maíz híbrido que tenía el citoplasma T (androesterilidad)

Variabilidad genética en maíz

Crop Acreage
(millions)
Value
(millions)
Total
no.
var.
Major
var.
Acreage
(%)
Beans,
dry 1.4 143 25 2 60
Beans,
snap 0.3 99 70 3 76
Cotton 11.2 120 50 3 53
Maize 66.3 5200 197 6 71
Peanut 1.4 312 15 9 95
Soybean 42.4 2500 62 6 56
Wheat 44.3 1800 269 9 50
Uniformidad genética de los cultivos

Estructuras florales

Estructuras florales

Formas de reproducción de las plantas
Reproducción asexual o vegetativa
No hay fusión de gametos. La planta se
basa en sí misma para producir
descendencia.
Hay plantas en las que cualquier porción
vegetativa es capaz de dar origen a una
nueva planta.

a) Multiplicación vegetativa Puede ser por:
Hijuelos, estaca, esqueje (cogollo), estolón, injerto,
acodo aéreo o subterráneo, rizoma, tubérculo y bulbo.

Ejemplos de multiplicación
Zacates Estolones
Plátano Rizoma
Piña Hijuelos
Papa Tubérculo
Caña de azúcar Estaca, cogollo
Fresa Estolones
Cebolla Bulbo
Frutales Injerto
Plantas de ornato Acodo

b) Apomixis
Formación de una “pseudo semilla” sin que ocurra la fusión
de gametos: partenogénesis, apogamia, aposporia. El embrión se desarrolla a partir de células del saco
embrionario o de células somáticas del óvulo (nucela).

Formas de reproducción
Reproducción sexual
Requiere la intervención de dos células especializadas llamadas gametos, que al
fusionarse dan lugar a una célula conocida como cigoto.
Los gametos pueden ser producidos en el mismo individuo o en diferentes. Los gametos femeninos se producen en el gineceo y los masculinos en el androceo.
El gameto femenino es fijo, rico en citoplasma y grande en volumen.

Reproducción sexual
El gameto masculino, es móvil, escaso en
citoplasma y más pequeño que el femenino.
Generalmente los gametos son iguales en el
número de cromosomas.
El número de gametos masculino, es mayor que el
número de gametos femenino.
El gameto femenino se forma por meiosis, se
encuentra en el saco embrionario y se conoce con
el nombre de oosfera o huevo; el gameto
masculino se encuentra en el grano de polen.

Reproducción sexual

Reproducción sexual
Clasificación de las plantas, en atención al lugar en
donde se forman los gametos de ambos sexos: 1.- Plantas dioicas: Los gametos se forman en plantas diferentes. Hay plantas que forman
únicamente los gametos hembra y hay plantas que
únicamente producen los gametos machos.
Los sexos están separados, por ejemplo: cáñamo,
espinaca, espárrago, palma datilera y papaya.

Reproducción sexual 2.- Plantas monoicas: Los gametos se forman en la misma planta. Hay especies en que los gametos masculino y femenino se forman en la misma flor (perfectas), y se conocen como plantas monoicas con flores hermafroditas; ejemplos: soya, fríjol, alfalfa, trigo, cebada, sorgo, algodón, tomate,
centeno y muchos frutales. En el grupo de plantas monoicas hay especies cuyas flores son imperfectas, únicamente tienen gineceo o androceo. Las flores únicamente producirán granos de polen o saco embrionario, por ejemplo: maíz y cucurbitáceas.

Autofecundación (autogamia)
Cuando los gametos que se fusionan para
formar el cigoto provienen de la misma planta, se dice que la planta se
autofecunda.
Las especies en que ocurre la
autofecundación se llaman autógamas;
ejemplos: soya, fríjol, chícharo, tomate,
tabaco, arroz, trigo, cebada, avena.

Fecundación cruzada (alogamia) La fecundación se llama cruzada cuando los gametos que se fusionan provienen de plantas diferentes. Las especies de fecundación cruzada se llaman
alógamas; ejemplos: las plantas dioicas, frutales, maíz, centeno, cucurbitáceas, tréboles, zacates. Hay especies que se pueden autofecundar y ser también de fecundación cruzada; éstas se conocen como especies de fecundación mixta; ejemplos: alfalfa, sorgo, algodón. Hay plantas que siendo monoicas hermafroditas son alógamas como el caso de frutales, centeno, tréboles, etc.

Esterilidad Incapacidad de una planta para producir gametos o cigotos viables. No es posible ni la autofecundación ni el cruzamiento. Existen diferentes grados de esterilidad y varias son las causas, siendo las de mayor importancia:
a) Aberraciones cromosómicas: fenómenos de
poliploidía, heteroploidía, inversiones, translocaciones.
b) Falta de homología entre los genomios.
c) Genes que afectan a los órganos reproductores.
d) Esterilidad que afecta únicamente al grano de polen (esterilidad masculina), debido a: acción de genes; acción del citoplasma o la interacción núcleo- citoplasma.

Esterilidad masculina
Hay androesterilidad o esterilidad
masculina cuando los órganos
reproductores masculinos (gametos)
de las plantas se encuentran mal
desarrollados o abortados de tal
manera que no se forma polen viable.
Aparece como consecuencia de:

a)
Esterilidad masculina
Genes mutantes (recesivos)
b) Factores citoplásmicos
c) Efectos combinados de ambos.
Es muy útil para los mejoradores de
plantas, porque proporciona un medio muy
eficaz para simplificar la formación de
híbridos (evita emasculación manual).

Androesterilidad genética
Controlada por un gen localizado en los cromosomas (núcleo), recesivo y de herencia simple. Es necesario que se presente la
homocigosis (msms) para que se exprese la androesterilidad.
Es importante identificar los individuos
androestériles por medio de genes marcadores a fin de eliminar los machos fértiles de la línea y evitar posibles fecundaciones. Se ha encontrado en maíz, cebolla, remolacha, trigo, etc.

Androesterilidad citoplásmica
Este tipo de androesterilidad predomina en plantas
alógamas. Existen plantas con un citoplasma especial que son
androestériles; sin embargo, pueden producir semillas si
se polinizan con plantas fértiles. La F1 producirán solo plantas androestériles, ya que el citoplasma se deriva del gameto femenino. El mantenimiento de estas líneas es muy fácil, pues solo se cruza una línea androestéril y una fértil con el mismo genotipo, excepto que el macho fértil tiene el citoplasma normal (N).

Androesterilidad citoplásmica
Es útil para la producción de híbridos simples y dobles en cebolla, remolacha, trigo, sorgo, maíz.
Fuentes de esterilidad citoplásmica en maíz:
a) Fuente (S) USDA.
b) Fuente (T) Texas.

Androesterilidad genética - citoplásmica
El gen F es
dominante
sobre el gen S.
El citoplasma es
de herencia
materna.

Ventajas y desventajas Ventajas:
1.- Producción comercial de semilla híbrida (evita emasculación).
2.- Mejora las posibles formas y proporciones de recombinación en plantas autógamas.
Desventajas:
1.-La incorporación del gen o genes de esterilidad es un proceso lento.
2.- Problemas para el mantenimiento de progenitores.
3.- La utilización de un solo gen extensivamente puede
ocasionar desequilibrios ecológicos (enfermedades Helminthosporium maydis)

Tipos de acción génica
Variación:
Término usado en genética para indicar las diferencias fenotípicas que se presentan entre los individuos que integran una población de plantas de una misma especie, raza, variedad o familia; como consecuencia de la acción de:
a) Factores genéticos
b) Influencia del medio
c) Interacción genotipo por ambiente
d) Efecto de una mutación
VARIACIÓN CONTINUA Y DISCONTINUA

1.- Para fines de mejoramiento de plantas por selección, la acción de genes de aditividad es la de mayor interés porque el fenotipo refleja el genotipo.
2.- La acción de genes en condición de dominancia o sobredominancia tiene relativa importancia para el mejoramiento por selección.
3.- Es de gran importancia investigar cual es el tipo de acción génica.
4.- Cualquiera que sea el tipo de acción génica, el fenotipo que manifiesta un carácter cuantitativo es influido por el efecto acumulativo de los genes.

- Cantidad expresada en porcentaje debido a la acción de los genes que intervienen en un carácter.
- Relación que existe entre un valor genotípico y un fenotípico para un carácter, expresando dichos valores como varianzas que estiman la variación genética y la variación fenotípica en una muestra al azar de una población respecto a un carácter en estudio, pudiendo también asignar la variabilidad con respecto a la unidad.
Heredabilidad

H = Varianza genética aditiva / Varianza fenotípica
Ha = (A2 / F2) * 100
Heredabilidad
La heredabilidad en el sentido más estrecho (genética) es la relación de la varianza genética aditiva, expresada en porcentaje, y la variación fenotípica observada:

Heredabilidad La heredabilidad en el sentido más amplio (genotípica, porque
incluye los diferentes tipos de acción génica) se define como la
relación entre la varianza genotípica y la varianza observada en
una población de plantas: H = Varianza genotípica/Varianza fenotípica
H = G2 / G2 + E2 + G*E2
H = A2 + D2 + I2 / G2 + E2 + G*E2

Heredabilidad
Es común simbolizar la heredabilidad por h2.
La anterior relación suele representarse en tanto por ciento de la
varianza genotípica con respecto a la variación total o fenotípica
observada.
H = (G2 / G2 + E2) * 100

Endogamia
Forma de apareamiento entre individuos más o menos emparentados (consanguinidad en animales). En las plantas monoicas compatibles la endogamia es máxima cuando ocurre la autofecundación, pero pueden presentarse diferentes grados de endogamia en atención al parentesco entre el conjunto de progenitores o al número de ellos.
En las especies autógamas la endogamia es la forma natural de realizarse y en las plantas alógamas se practica la endogamia en forma artificial, controlando la polinización.

Endogamia
El cruzamiento entre progenitores e hijos, entre hermanos, etc., se llama cruza consanguínea y la descendencia puede estar relacionada, usándose los términos cruzas fraternales, cruzas de medios
hermanos, cruzas entre primos, etc. Existe una tendencia en asociar los efectos biológicos desfavorables con la endogamia, debido a que se han observado efectos contrastados cuando se comparan los efectos de la endogamia máxima
(autofecundación), las cruzas consanguíneas y el cruzamiento entre individuos no relacionados.

Endogamia
La consanguinidad suele ir acompañada de:
1.- Reducción de tamaño
2.- Disminución del vigor
3.- Pérdida parcial o total de la fecundidad
4.- Plantas deformes, albinas, susceptibles al acame, etc.
5.- Debilitamiento general de la población.

Endogamia Coeficiente de endogamia:
En una población el coeficiente de endogamia indica
la proporción en que desaparecen los heterocigotos;
se simboliza por F y es un valor cuantitativo que mide
la intensidad de la endogamia y varía de:
F = 1, para completa homocigosis
F = 0, para una población panmíctica.

Endogamia Si se autofecunda un individuo sucesivamente, se
obtiene el máximo valor de endogamia.
F = ½ (1 + F’)
F’ = Coeficiente de endogamia de la generación
anterior.
La Ley de Hardy – Weinberg, considerando el valor de
F, se expresa de la siguiente forma:

Endogamia 1.- Disminución del vigor y rendimiento
2.- Aparición de individuos notables por su
uniformidad o por anormalidades que originan
problemas de supervivencia.

Endogamia Utilidad de la endogamia en el mejoramiento de las
plantas:
1.- Producción de plantas uniformes y genéticamente
homocigotas.
2.- Purificar una variedad de taras y anormalidades,
sea por selección natural o por selección que hace el
mejorador en el proceso de endogamia eliminando a
los homocigotos no deseables.

Endogamia 3.- La disminución del vigor puede restaurarse por el
cruzamiento entre líneas puras seleccionadas.
4.- Los híbridos resultantes entre líneas
autofecundadas, suelen ser muy uniformes y de
mayor vigor que las variedades progenitoras de las
líneas, debido a la heterosis, la cual puede explotarse
en la generación F1 o perpetuarse por vía asexual.

Heterosis
Es el fenómeno en virtud del cual la cruza (F1) entre dos razas, dos variedades, dos líneas, etc., produce un híbrido que es superior en: tamaño, rendimiento o vigor general.
Se habla de heterosis cuando la F1 es superior en vigor al promedio de los progenitores. Se considera también como una manifestación de la heterosis
cuando la F1 es superior al vigor del progenitor más vigoroso y tiene por consiguiente mayor importancia económica.

Heterosis
El término heterosis se debe a G.H. Shull (1914), que lo usó como una contracción de la expresión “estímulo de la heterocigosis”. Se utiliza como sinónimo de “vigor híbrido” por el efecto que se manifiesta en la
generación F1 al presentarse un estímulo general en el híbrido.
Esta manifestación consiste en un incremento en la producción, en la altura, en la resistencia de plagas, a sequía, a enfermedades, etc., o cualquier otra característica que expresa mayor vigor que el que manifiesta el promedio de los progenitores o el progenitor más vigoroso.

Heterosis
Manifestación de la heterosis:
Se produce un estímulo general en la progenie o en el híbrido y afecta a las variedades de diferentes maneras. En general se manifiesta por:
1.- Mayor rendimiento de grano, forraje o frutos.
2.- Madurez más temprana
3.- Mayor resistencia a plagas o enfermedades
4.- Plantas más altas

Heterosis
5.- Aumento en el tamaño o número de ciertas partes u órganos de la planta. 6.- Incremento de algunas características internas de la planta. 7.- Mayor eficacia metabólica (mayor actividad y más rápida).
8.- Mayor eficiencia biológica (mayor actividad reproductiva y mayor capacidad para sobrevivir).

220
Heterosis

- -
Importancia de la heterosis Utilización del vigor híbrido en plantas:
Mayor producción de los cultivos, para satisfacer a una población mayor y más demandante de
alimentos.

Selección
Es un proceso de mejora genética por medio del cual se eligen como progenitores de una generación a los individuos de fenotipo, suponiendo también de
genotipo, más favorable para el carácter ideal y previamente diseñado. Para que la selección sea efectiva es necesaria la aplicación de ciertos principios, información y técnicas que faciliten el proceso, tales como: - Conocer las plantas y el medio ecológico.
- Obtener información sobre la herencia y heredabilidad del carácter.

Selección
- Determinar si es un carácter cuantitativo o cualitativo.
- Conocer el tipo de acción génica.
- Usar técnicas que reduzcan la acción del medio.
- Conocer la forma de reproducción de la especie (autogamia y alogamia).
- Trabajar con las manos, el cerebro y el corazón.

Selección
Selección diferencial: El término indica la diferencia entre la media de la muestra seleccionada como progenitora de la siguiente generación y la media de la población original en donde se practica la selección de los fenotipos. Esta diferencia se expresa en valores de la desviación estándar de la población original.
Selección diferencial = (x - ) = ó >
Este valor mide la presión o intensidad de selección.
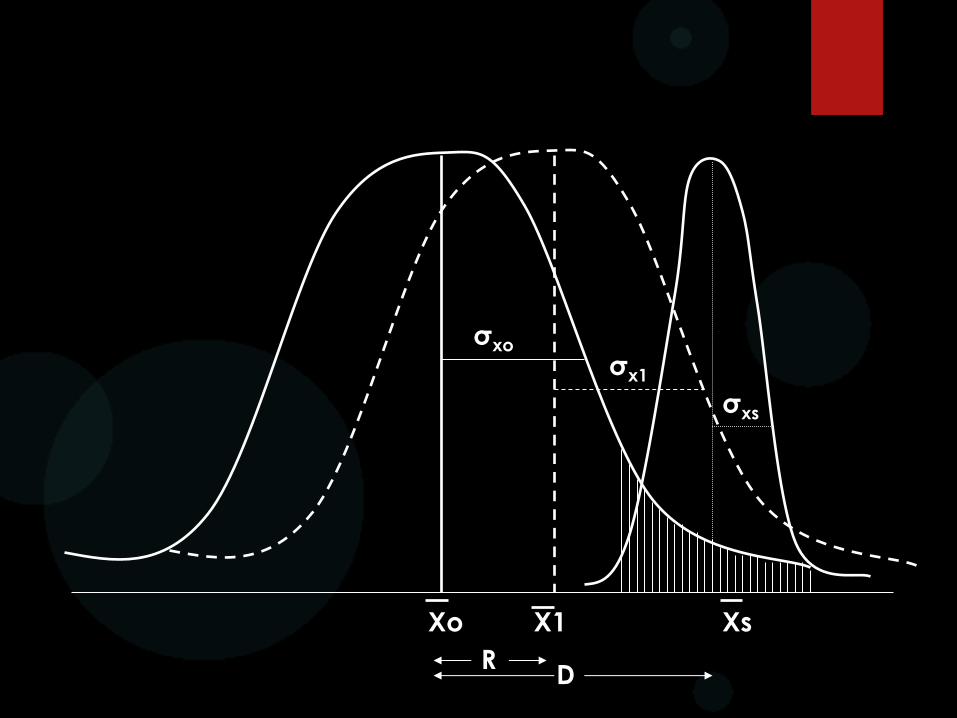
σxo
σx1
Xo Xs X1
σxs
R D

Selección y frecuencias
génicas La selección altera las frecuencias génicas de la
población seleccionada y la progenie originada a partir
de ella (media y varianza).
Frecuentemente, el objetivo es “fijar” uno de los dos alelos de cada locus (dominante o recesivo) en la población.
Por ello, la selección “acelera” un proceso natural que podría ocurrir en varias generaciones de apareamiento
preferencial o aleatorio pues rompemos el supuesto del
equilibrio HWC.

RESPUESTA A LA SELECCIÓN
El avance genético o R (∆G) mide el éxito de la selección mediante la
comparación de la población original y la progenie.
El avance genético puede ser predicho si conocemos Xo, Xs, b y σ2xo.
La R depende de la relación lineal entre la frecuencia del gen favorable
en la unidad de selección y su valor fenotípico.
X = Valor fenotípico
Y = Pu = Frecuencia del gen favorable
po + ∆p
∆p
Xo Xs
po
θ

Avance genético por ciclo de
selección: ∆G = (Xs-Xo) σ
2a/σ
2x = Sh2
Los componentes de la ecuación pueden calcularse antes de la
selección y el efecto de la varianza aditiva se puede estimar con
algún diseño de apareamiento (diseños genéticos o diseños
dialélicos).
S solo va a conocerse después de aplicar selección, pues hasta
entonces conoceremos Xs. S depende de la presión de
selección y la varianza fenotípica.
Cuando se aumenta S se reduce el porcentaje de individuos
seleccionados en la PO; p.e. la S de 1% en 1000 individuos (10)
es mayor a S = 5 % (50).

Respuesta a la selección de acuerdo a la
metodología de selección:
Selección masal.
R = k* ½ σ2A/σx
Selección familial de medios hermanos (maternos o paternos).
R = k* ¼ σ2A/σfMH
Selección familial de hermanos completos.
R = k* ½ σ2A/σfHC
Selección de autohermanos.
R = k* σ2A/σfAH k* [2pq(p-1/2) (αd)]/ σfAH
Función del grado de
dominancia

Mejoramiento de plantas
alógamas

Selección Masal
Método de selección recurrente, que permite concentrar genes favorables para un carácter
deseable. Se inició el método desde que el hombre necesitó domesticar las plantas y se ha venido modificando a través del tiempo a medida que han avanzado la ciencia y la tecnología. Consiste en sembrar una población de plantas, elegir los fenotipos deseables, cosechar la semilla, mezclar las semillas de las plantas seleccionadas y esta mezcla o “masa”, sirve como semilla para la siguiente siembra con la finalidad de que se recombinen o concentren los genes favorables para el carácter bajo selección.

Selección Masal
Para formar la mezcla es conveniente usar el mismo número de semillas de cada planta seleccionada, por lo cual se dice que se forma un “compuesto balanceado”.
El proceso sucesivo de: siembra selección
mezcla siembra, recibe el nombre de ciclo de selección.
En la selección masal únicamente se tiene
información del progenitor femenino y se ignora el masculino mezclado entre los posibles en la población de plantas del lote de selección.

Selección Masal
La selección masal es eficiente para aquellos caracteres y condiciones siguientes:
a) Caracteres cualitativos de herencia simple como son: acame, algunas enfermedades, color y textura del grano.
b) Caracteres cuantitativos de alta heredabilidad, tales como: precocidad y altura de planta.
c) Caracteres cuantitativos en variedades con alto valor para la varianza aditiva.


Selección Masal Estratificada
Esta técnica del 1960 se fundamenta en la
información experimental y teórica siguiente: - La varianza genética aditiva está presente en un gran número de variedades de polinización libre. - Hay cambio en la frecuencia génica, si la muestra es grande y la selección diferencial es tal que incluye a los genotipos deseables. - Las cruzas dialélicas estiman la varianza genética aditiva con un valor altamente confiable.

Selección Masal Estratificada
- La población básica inicial es heterogénea y
heterocigótica.
- Cada planta de la población tiene una aptitud combinatoria general diferente. Las plantas individuales son heterocigotas y se les simboliza por So (plantas con cero autofecundaciones).
- La técnica experimental en el proceso de selección elimina parcial o totalmente el efecto ecológico.
- La recombinación de genes favorables para el rendimiento se van concentrando en cada ciclo de selección cuando el compuesto balanceado se multiplica en un lote aislado.

Selección Masal Estratificada
Para ajustar la variable, se puede utilizar la siguiente expresión:
Yij = Xij – xi + xg
Yij = Valor ajustado de la variable de la planta j en la parcela i.
Xij = Valor observado de la variable en la planta j de la parcela i.
xi = Valor promedio por planta en la parcela i.
xg= Valor promedio de la variable en el lote



Selección Familiar
En este grupo de métodos de selección, las unidades de selección no son plantas individuales, sino familias.
Familia: Es la progenie obtenida de un apareamiento no aleatorio, cuya descendencia guarda una relación más o menos estrecha y de acuerdo a este relación se agrupan.
El esquema general sigue tres pasos:
1) Crear la estructura de la familia.
2) Evaluar y seleccionar las mejores familias a través de las progenies.
3) Recombinar las familias seleccionadas.

255

Familias de medios hermanos
Cada mazorca que proviene de polinización libre (PL) origina una familia de medios hermanos maternos (FMHM), al tener en común a la madre y diferentes padres.
macho,

Familias de hermanos completos
Si una planta macho poliniza a cuatro o seis plantas hembra diferentes y la semilla de cada una de ellas se siembra en surcos individuales, las plantas dentro de cada surco son hermanos completos (HC); porque tienen el mismo padre y madre, así, las plantas del surco uno son MH de las plantas del surco dos; éstos a su vez son MH de las del surco tres y así sucesivamente, ya que tienen un padre en común (macho) pero diferente madre. Si la semilla de las plantas hembra polinizadas por un macho común se mezcla en cantidades iguales (CB), se dará origen a una familia de medios hermanos paternos (MHP).


Familias de hermanos completos
Otra forma de obtener familias de hermanos
completos, consiste en llevar a cabo el apareamiento entre dos individuos o cruza planta a planta (PaP). La familia se puede derivar de la semilla proveniente de la cruza directa ó de la cruza recíproca. Las familias de MH están más relacionadas entre sí que aquellas de HC, debido a que las de MH se originan al cruzar una hembra con varios machos o viceversa, es decir, tienen un padre en común. Mientras las familias de HC que se originan al cruzar pares de plantas no lo tienen, todos son diferentes (cuando la hembra y el macho provienen de una población de entrecruzamientos al azar).

261

Familias de autohermanos (FAH)
La familia de autohermanos o líneas S1, es la progenie obtenida por autofecundación de una planta.

Selección Recurrente
J. L. Jenkins (1940) propuso este tipo de selección con la finalidad de desarrollar variedades sintéticas a partir de líneas de pocas autofecundaciones. Los pasos para la operación son lo siguientes: 1.- Se autopolinizan las plantas de una población heterocigótica y a la vez se valoran por algún carácter o caracteres deseables.
2.- Se desechan las plantas que tienen un comportamiento inferior en cuanto al carácter o caracteres que se desea mejorar.

Selección Recurrente
3.- Se realizan a mano todos los cruzamientos posibles entre los descendientes superiores o, si esto es impracticable, los cruzamientos se hacen por polinización libre entre los descendientes
seleccionados. La población resultante de los cruzamientos sirve como punto de partida para ciclos posteriores de selección o entrecruzamiento.

Selección recurrente simple
En este tipo de selección, se dividen las plantas en dos grupos, uno que se desechará y otro que se propagará después, considerando las puntuaciones dadas según el fenotipo de las plantas individuales o de sus descendencias por autofecundación. Ejemplo. Sprague, Miller y Brimhall (1952), utilizaron la variedad sintética de maíz stiff stalk, para estudios del carácter contenido de aceite. Los resultados del proceso de selección recurrente se presentan en la siguiente figura.



Selección recurrente recíproca
Comstock, Robinson y Havey (1949), propusieron este método de selección. Este procedimiento es una modificación de la selección recurrente; la diferencia consiste en utilizar material básico de dos fuentes, A y B. El material original puede incluir dos o más variedades de polinización libre o sintéticas. La investigación
genética del material o la divergencia deberán ser tan amplias como sea posible, pero deben combinar entre si, ya que los híbridos que se desarrollarán comprenderán el cruzamiento entre las líneas puras derivadas de las dos fuentes.



Producción de
variedades híbridas

275
Autofecundación
276
Cruza o hibridación

277
Cruza natural

278
Cruza fraternal

279
Cruza recíproca

280
Mestizo

281
Cruza simple

282
Cruza triple

283
Cruza doble

284
Cruza doble

285
Cruza doble críptica

286
Cruzamiento múltiple

• Una o Varias Poblaciones a Mejorar
Aptitud Combinatoria General
Cruza de Padres con Aptitud Combinatoria Específica Superior
Variedades de Polinización libre
Variedades Sintéticas
Variedades Híbridas

Población Base
Obtención y desarrollo de
líneas endocriadas
Selección de Líneas
Evaluación de Aptitud Combinatoria
Evaluación de Híbridos experimentales
Híbrido
Mejoramiento
de Líneas

Aspectos en el desarrollo de
Híbridos
• Utilización de la endogamia para
aumentar la variabilidad genética
• Selección de líneas endocriadas
superiores
• Explotación de la Heterosis

Desarrollo de Líneas
Endogámicas
• Fuentes de Material
• Obtención de Líneas Endogámicas
• Evaluación de la Aptitud
Combinatoria
• Aptitud Combinatoria General
• Aptitud Combinatoria Específica

•
•
•
•
Fuentes de Material
Variedades de polinización libre
Híbridos (F2)
Variedades sintéticas
Líneas de segundo ciclo

Porcentaje de esfuerzo para el desarrollo de líneas
endogámicas de acuerdo a diferentes fuentes de
germoplasma en USA
Fuente de Germoplasma
Poblaciones de base amplia
Poblaciones de base estrecha
Poblaciones con endogamia
Cruza entre líneas elite
Poblaciones derivadas de retrocruzas
% de Esfuerzo
15
16
14
39
17

Evaluación de Líneas
• Aptitud Combinatoria General
• Aptitud Combinatoria Específica
• Identificar líneas que en ciertas combina-
Ciones produzcan híbridos superiores.

Evaluación de la Aptitud
Combinatoria General
• Consiste en la evaluación del comportamiento de líneas en cruzas con un probador (Tester) común
• Las F1 se prueban en distintos ambientes
• Evalúa la varianza genética aditiva de cada línea
• Permite descartar líneas de pobre comportamiento en cruzas en etapas tempranas de la endogamia (S2; S3)

Obtención de cruzas para ACG
Líneas S2-S3
(despanojadas) Tester
ECR con las F1 (línea x tester)
en 2 -4 localidades
Selección de líneas con superior ACG
1° Año
2°-3° Años

Germoplasma a utilizar como probador
o tester
•
•
•
•
Variedad de polinización libre
Compuesto de amplia base
Variedad sintética
Generaciones avanzadas de un híbrido

Evaluación de la Aptitud
Combinatoria Específica
• Consiste en la evaluación del comportamiento de líneas endocriadas (S6...Sn) en todas la combinaciones posibles (Dialélico)
• Las F1 se prueban en distintos ambientes
• Evalúa la varianza genética de dominancia
• Permite cuantificar la heterosis en las distintas cruzas
• La mejores cruzas constituyen los híbridos experimentales

Obtención de Cruzas para ACE
A B C D E
A * AxB AxC AxD AxE
B * BxC BxD BxE
C * CxD CxE
* D
E
DxE
*

Número de cruzas simples y dobles de acuerdo
al número de líneas
Cruza
Simple
Líneas 5
10
20 100 n
Top Cross
5
10
20 100 n
(Híbrido Simple)
10
45
190 4950 n(n-1)/2
Cruza Doble (Híbrido Doble)
15
630
14535 11763625 3n(n-1)(n-2)(n-3)/24

Híbridos modificados
• Los híbridos modificados consisten en cruzamientos en
donde uno o ambos progenitores son líneas emparentadas
(líneas hermanas o que poseen un progenitor común en su
ascendencia), con un grado de parentesco variable.
• Se han utilizado principalmente por problemas de
producción de semilla, ya que los progenitores son más
vigorosos que las líneas endogámicas debido ala
manifestación de cierto vigor híbrido en la cruza.

Comportamiento de las
líneas
• Correlación entre caracteres exhibidos
por las líneas y en los híbridos.
• En general las correlaciones son no
significativas y/o de baja magnitud,
sobre todo para productividad.

El valor real de una línea debe ser
demostrada en los híbridos en
que participa
• La baja correlación entre líneas e
híbridos obliga a mantener una gran
cantidad de líneas para aumentar la
probabilidad de obtener híbridos
superiores.

Rendimiento de híbridos simples y
promedio de sus líneas parentales HS
LP

Híbrido Pedigree
TopCross LíneaxPoblación
HíbridoSimple AxB
Diferentes clases de híbridos
HíbridoSimple
Modificado
(AxA’)xB
(AxA’)x(BxB’)
HíbridoTriple (AxB)xC
HíbridoTriple
Modificado
(AxB)x(CxC’)
HíbridoDoble (AxB)x(CxD)

Líneas endogámicas Híbrido

Requerimientos para la
producción de semilla híbrida
• Manifestación de Heterosis en al F1
• Eliminación de polen fértil de la línea
madre
• Transferencia del polen del macho a la
hembra
• Producción económica de semilla híbrida

Relación entre líneas para la
producción de semilla híbrida
Cultivo Línea Línea
productora de polinizadora
semilla
Sorgo
Maíz
Girasol
Trigo
3
2/4
2/7
1/3
1
1
1
1

287
Obtención de híbridos

Mejoramiento de plantas
autógamas

Mejoramiento de plantas autógamas
Las poblaciones de plantas autógamas consisten generalmente en una mezcla de multitud de líneas homocigotas bastante relacionadas que, no obstante que crecen próximas permanecen más o menos
independientes entre si en la reproducción. Las plantas individuales de tales poblaciones son probablemente homocigotos vigorosos.
En determinadas especies autógamas (algodón, sorgo, y trébol) existen cruzamientos naturales que oscilan de 5 al 50%. En otras especies (cebada, avena, arroz, lechuga y tomate), el cruzamiento en raras ocasiones supera el 1%, independientemente de la variedad de que se trate. En fríjol lima el cruzamiento puede oscilar del 25 al 100%.

a) b) c) d) e)
Métodos de selección
Método genealógico o pedigrí.
Método de progenie de una semilla por planta (PSP y/o SSD).
Selección Masal
Selección por retrocruza.
Selección recurrente.

Genealógico
o
Pedigrí


Método masivo

Selección
masal

Retrocruza

Descendiente de
Una sola semilla
(SSD)

autógamas Selección Recurrente en
En plantas autógamas, en principio, el mejoramiento puede hacerse en forma similar a las alógamas, siempre y cuando su aparato reproductivo permita en alguna forma la polinización cruzada más o menos masiva a fin de poder llevar a cabo, en el caso de la selección recurrente, la recombinación genética de las unidades seleccionadas.
El uso de genes de androesterilidad ms permiten la alogamia en sorgo, de manera que es posible realizar selección recurrente en varias formas de selección masal y familiar.
Existe en las plantas autógamas el potencial operativo para llevar a cabo la recombinación genética (limitante).

autógamas Selección Recurrente en
En general la selección recurrente en autógamas se ha llevado a cabo usando familias. Por lo tanto, las etapas que se sobrellevan son las mismas que en la selección familiar observadas, y consisten en:
1.- Polinización (para recombinación) y prueba
2.- Selección, ya que en la recombinación está implícita la formación de las nuevas familias.
En autógamas, se pueden hacer varias generaciones de autofecundación en las cruzas simples que provienen de la recombinación a fin de obtener líneas homocigotas y aprovechar el doble de la varianza aditiva existente.

Recombinación con polinización
Hay plantas en que la polinización manual es tan laboriosa, difícil e ineficiente, que las probabilidades de llevar a cabo la recombinación genética en la selección recurrente son sumamente bajas. Se deben hacer grandes cantidades de polinización manual a efecto de garantizar y obtener el número real (necesario) de polinizaciones efectivas.

Recombinación con androesterilidad
Brim y Stuber (1973) propusieron un esquema general de selección recurrente usando androesterilidad, el cual se resume a
continuación:
En general, cada ciclo de selección recurrente consiste en tres etapas o fases: recombinación, autofecundación y prueba y selección, que estos autores denominaron generaciones. Previo al inicio del programa se supone que se ha sintetizado la población base, usando cruzas dobles, retrocruzas o las cruzas posibles de un diseño dialélico, pudiendo minimizarse la contribución del progenitor donante del gen estéril por retrocruzamiento al progenitor recurrente.

La población base será una mezcla mecánica de los tipos de cruzas mencionados anteriormente; éstas se han hecho utilizando plantas fértiles de las variedades y sus contrapartes estériles habiendo adquirido el gen de esterilidad de una línea dominante, y habiendo retrocruzado 2 o 3 veces hacia la variedad recurrente para recobrar el genotipo. A continuación se ilustran las tres generaciones o fases de las selección recurrente en plantas autógamas.

Selección Recurrente en autógamas

Mejoramiento de
variedades clonales

Variedad clonal:
Material genéticamente
uniforme derivado de un
solo individuo y que se
propaga sólo por
métodos vegetativos o
asexuales.

Variabilidad genética en
especies clonales:
Mutación: naranjos, pomelos,
manzano.
Hibridación
Infecciones virales: variegados.

Métodos:
Selección clonal
Hibridación y selección
clonal
Mejoramiento por
apomixis.

CLON 1 CLON 2
Población con alta variabilidad
Selección de plantas
OBTENCION DE CLONES
COMPARACION Y
SELECCIÓN DE
CLONES Parcelas clonales
Selección entre clones
E.C.R. MULTIPLICACION
Clones Selectos

ClonA x Clon B
Selección de las mejores plántulas
Evaluación
De clones
2 – 3 años
Clones selectos
Multiplicación

Mejoramiento por apomixis





















![[Day 2] Center Presentation: CIMMYT](https://img.pdfslide.net/doc/110x75/55502cfbb4c9059f318b4ba9/day-2-center-presentation-cimmyt.jpg)







