Embed Size (px)
Citation preview

i
Biosorption of Copper by Nepenthes
Ampullaria-Associated-Endophytic
Fungi
by Wong Changi
Thesis submitted in partial fulfilment of the requirements for the
degree of
Master of Science (by research)
Faculty of Engineering, Computing and Science
Swinburne University of Technology
2015

ii
Abstract
In recent years, environmental pollution by heavy metals has caused increasing ecological
damage and led to global public health concerns. Biosorption is one of the ways to deal
with heavy metal pollution. In this study, endophytic fungi were (a) isolated from the
carnivorous plant Nepenthes ampullaria (b) assessed for their resistance against heavy
metal copper and (c) evaluated for their biosorption capacity. In total, 147 fungal isolates
were isolated from Nepenthes ampullaria and only 11 (7.5%) of the total isolates were
capable to resist copper concentration up to 1000 ppm. The 11 fungal isolates were
identified through molecular method, and grouped as members of the Phomopsis,
Diaporthe, Nigrospora, and Xylaria. The fungal isolate NA40 related to Xylaria sp.
achieved the highest biosorption capacity of of 73.26 mg/g using live biomass, thus
chosen for study of proteome expression in response to copper. Three different copper
concentrations (0, 300, 500 ppm) were used in the study. Results show that there are 11
protein spots being up-regulated and 1 protein spot down-regulated in response to copper.
The protein spots were identified to be related to the enzymes involved in heat shock
protein, DNA repair and antioxidant reaction. This study on Xylaria serves as a baseline
study for the response of the fungus to copper.

iii
Acknowledgement
“I will give you every place where you set your foot, as I promised Moses. – (Joshua 1:3)”
First and foremost, thanks be to God for the blessing throughout my life and last forever.
A special word of gratitude to my darling, Julia Wee, for all her love and support.
My family, especially my parents and grandparents who provide me with everything I need, love, support and encouragement.
A person who offers his unreserved help and guidance, who I must offer my profoundest
gratitude - my research supervisor, Dr. Moritz Müller. Instead of being like a supervisor,
I feel more like a friend, who is extremely enthusiastic about any kind of research! I am
looking forward for the next round of the researches!
My co-supervisor, Dr. Daniel Tan and Dr. Samuel Lihan, who provided me with help and
guidance throughout the project experiment.
A Big thanks goes to my lab mates and friends, who share and discuss the knowledge of
different fields with me, and also the laboratory officers, who provided me with help and
guidance throughout my bench work period.
Deepest appreciation to all. The simple phrase “thank you” cannot present how much I
feel thankful to you. Without you, this research as well as the dissertation would not have
been possible.
May God bless you and your family, abundantly.

iv
Declaration
I, Wong Changi, hereby declare that this research project entitled “Biosorption of Heavy
Metal (Copper) and Proteomics Study on Nepenthes ampullaria Associated Endophytic
Fungi” is original and contains no material which has been accepted for the award to the
candidate of any other degree or diploma, except where due reference is made in the text
of the examinable outcome; to the best of the candidate’s knowledge contains no material
previously published or written by another person except where due reference is made in
the text of the examinable outcome; and where the work is based on joint research or
publications, discloses the relative contributions of the respective workers or authors.
(WONG CHANGI)
Date: 1st May 2015

v
Publication Arising from this Thesis
The work described in this thesis has been submitted as described in the following:
Wong, C, Tan, D, Lihan, S, Mujahid, A & Müller, M "Biosorption of Copper (Cu) by Endophytic Fungi Isolated from Nepenthes ampullaria", Applied Microbiology and Biotechnology (Manuscript under consideration)

vi
Table of Contents
Page
List of Figures ix
List of Tables xi
1 Introduction 1
1.1 Heavy Metal 1
1.1.1 Copper Pollution 1
1.1.2 Copper Toxicity 5
1.2 Current Technologies for Heavy Metal Removal 7
1.2.1 Chemical Precipitation 7
1.2.2 Ion Exchange 8
1.2.3 Electrodialysis 8
1.2.4 Semiconductor Photocatalysis 9
1.2.5 Membrane Filtration 10
1.2.6 Phytoremediation 11
1.3 Biosorption 13
1.3.1 Biosorbents 15
1.4 Fungal-plant Symbiotic Interaction 17
1.4.1 Endophytic Fungi 17
1.4.2 Heavy Metal Tolerance of Endophytic Fungi 20
1.4.3 Biosorption of Heavy Metal using Endophytic Fungi 21
1.5 Pitcher plants (Nepenthes) as Source of Endophytic Fungi 22
1.5.1 Distribution 23
1.5.2 Habitat 24
1.6 Proteomics - Regulation of Fungi Proteins in Response to
Heavy Metal Stress
25
1.7 Aims of the Present Study and Dissertation Outline 26
2 Methodology 28
2.1 Sampling Sites 28
2.1.1 Ulu Mentawai 30
2.1.2 Kota Samarahan Roadside 31
2.2 Nepenthes ampullaria Associated Endophytic Fungi Isolation 32
2.2.1 Ulu Mentawai 32

vii
2.2.2 Kota Samarahan Roadside 33
2.2.3 Endophytic Fungi Isolation 33
2.2.4 Endophytic Fungi Purification 35
2.2.5 Short Term Storage of the Isolated Fungi 37
2.2.6 Long Term Storage of the Isolated Fungi 37
2.3 Preliminary Screening of the Resistance Isolated Fungi Against
the Heavy Metal Copper
38
2.4 Molecular Identification of the Chosen (11) Fungal Isolates 39
2.5 Evaluation of Biosorption Capacity of the Chosen Fungal
Isolates
43
2.5.1 Heavy Metal Copper Biosorption by Live Fungal
Biomass
43
2.5.2 Heavy Metal Copper Biosorption by Dead Fungal
Biomass
46
2.6 Proteomic Analysis of the Best Fungal Strain (NA40) on Heavy
Metal Copper
48
2.6.1 Fungal Proteome Preparation 49
2.6.2 Total Protein Measurement by Bradford Assay 51
2.6.3 Two-dimensional Gel Electrophoresis (2-DE) 51
2.6.4 Silver staining for SDS-PAGE 54
2.6.5 Protein Identification and Database Search 56
3 Biosorption of Copper (Cu) by Endophytic Fungi Isolated from
Nepenthes ampullaria
60
3.1 Introduction 61
3.2 Methodology 63
3.2.1 Endophyte Isolation and Purification 63
3.2.2 Preliminary Screening of Heavy Metal Copper
Tolerance Fungi
63
3.2.3 Molecular Identification 63
3.2.4 Biosorption of Copper by Living Fungal Biomass 64
3.2.5 Biosorption of Copper by Dead Fungal Biomass 65
3.3 Results and Discussion 66
3.4 Conclusion 76

viii
4 Proteomics analysis of the Nepenthes ampullaria associated
endophytic fungus, Xylaria sp.
78
4.1 Introduction 79
4.2 Methodology 81
4.2.1 Culture Conditions 81
4.2.2 Protein Extraction 81
4.2.3 2-DE and Image Analysis of Protein Spots 81
4.2.4 Protein Identification and Database Search 82
4.3 Results and Discussion 83
4.4 Conclusion 91
5 Summary, Conclusion and Future Work 92
5.1 Summary 92
5.2 Future Work 93
References 94

ix
List of Figures
Figure Page
1.1 Basic structure of Nepenthes pitcher. 22
1.2 Distribution map of Nepenthes sp., taken from Carnivorous Plants /
Insectivorous Plants in the Wilderness.
24
2.1 Ulu Mentawai (sampling sites), located at northern part of Gunung Mulu
National Park, indicated by red point (Source: Google Map).
29
2.2 Kuching Kota Samarahan roadsite (sampling sites), indicated by red
point (Source: Google Map).
29
2.3 Nepenthes ampullaria in situ, photographed on site (Mentawai jungle). 30
2.4 Author collecting plant samples collecting at in Mentawai jungle. 31
2.5 Kuching Kota Samarahan Roadside, the area where the Nepenthes
ampullaria was collected.
32
2.6 A self-made plastic box - I was doing the endophytic fungi isolation at
the site.
33
2.7 An overview in form of the isolation of endophytic fungi. 35
2.8 Endophytic fungi were observed growing out from the surface sterilized
plant tissue.
36
2.9 Purified fungal strains. 36
2.10 A schematic view of short term storage of isolated fungi. 37
2.11 A schematic view of long term storage of isolated fungi. 37
2.12 A schematic overview of preliminary screening of the resistance isolated
fungi against the heavy metal copper.
39
2.13 Polymerase Chain Reaction (PCR) results – gel bands. 41
2.14 A schematic overview of molecular identification of the chosen (11)
fungal isolates.
42
2.15 Fungal isolates were growing in the potato dextrose broth supplied with
500ppm of copper.
44
2.16 A schematic overview in form of a flowchart of the heavy metal copper
biosorption by Live fungal biomass.
45
2.17 A schematic overview in form of a flowchart of the heavy metal copper
biosorption by Dead fungal biomass.
47

x
2.18 An overview in form of a flowchart of the fungal incubation in PDB
with three different concentration of heavy metal copper concentration.
48
2.19 An overview in form of a flowchart of the fungal proteome preparation. 50
2.20 An overview in form of a flowchart of Isoelectric focusing. 52
2.21 An overview in form of a flowchart of two-dimensional gel
electrophoresis (2-DE).
53
2.22 An overview in form of a flowchart of silver staining for SDS-PAGE. 54
2.23 Silver stained gel image taken using Cannon digital camera, at the bench
of the lab.
55
2.24 Protein spots produced by the fungal isolate NA40 in the PDB with no
heavy metal copper, gel image was taken using image Scanner (GS800
Desitometer (Biorad)).
57
2.25 Protein spots produced by the fungal isolate NA40 in the PDB supplied
with 300ppm of heavy metal copper, gel image was taken using image
Scanner (GS800 Desitometer (Biorad)).
58
2.26 Protein spots produced by the fungal isolate NA40 in the PDB supplied
with 500ppm of heavy metal copper, gel image was taken using image
Scanner (GS800 Desitometer (Biorad)).
58
2.27 An overview in form of the in-gel digestion of the protein spots. 59
3.1 ITS gene-based phylogenetic tree representing fungal sequences
conserved within the internal transcribed spacer region. The
phylogenetic tree was constructed using Mega 6 with distance method
and sequence distances were calculated using maximum likelihood
method. Boot strap values of 2000 are shown and the cut off value of
50%.
70
4.1 Up-regulated protein spots in response to copper. 90
4.2 Down-regulated protein spot in response to copper. 90

xi
List of Tables
Table Page
1.1 Acid mine drainage and some other industrial activities impact towards
the environment and human.
3
1.2 Malaysia local heavy metal contaminated area and the source of pollution. 4
1.3 Effects of acute and chronic copper poisoning on human organ. 6
1.4 Overview of advantages and disadvantages of each of the five
phytoremediation techniques /approaches.
13
1.5 Overview of biosorbents and their uptake efficiencies for selected metals. 15
1.6 Overview of endophytic fungi and their host plant interaction. 18
1.7 Overview of endophytic fungi and their resistance against heavy metal. 20
1.8 Overview of endophytic fungi and their capability of biosorpt heavy
metal.
21
2.1 GPS coordination for the Nepenthes ampullaria plant samples collected. 28
3.1 Fungal isolates that manage to survive up to 1000ppm of heavy metal
copper concentration.
67
3.2 ITS phylogenetic results for the 11 isolated fungi strains. 69
3.3 Biosorption capacity (mg/g) of the Live biomass of the isolated fungal on
heavy metal copper biosorption.
72
3.4 Biosorption capacity (mg/g) of the Dead biomass of the isolated fungal on
heavy metal copper biosorption.
74
4.1 List of identified proteins produced (upregulated and downregulated) in
response of heavy metal copper.
88
4.2 Up-regulated proteins in response to copper. 89
4.3 Down-regulated protein in response to copper. 89

- 1 -
Chapter 1
Introduction
1.1 Heavy Metal
In recent years, environmental pollution by heavy metals has caused increasing ecological
damage and led to global public health concerns (Tchounwou et al. 2012). Heavy metals
are naturally occurring elements that can be found throughout the earth’s crust and can
be considered as trace elements when present at trace concentrations (mgkg-1 or less) in
agro-ecosystems (He et al. 2005). However, most of the heavy metal have been released
into our environment from anthropogenic activities such as smelting operations, industrial
production, agricultural, mining activities, sewage treatment plant, and domestic garbage
dumps (Yunus et al. 2011). These activities result in increasing concentrations of heavy
metals in our environment and lead to heavy metal pollution (Tchounwou et al. 2012).
Heavy metals are catergorised as severe pollutants due to their toxicity, bioaccumulation
and persistent properties (Tam & Wong 2000). They are neither chemically nor
biologically degraded, and therefore, the pollutants may persist in the environment for
long period of time. For instance, mangrove forest of Tanjung Lumpur are known to be
polluted by several heavy metal such as lead, copper and Manganese due to the
terigeneous and antropogenic activities. The copper and lead are know to be most concern
metals due to their accumulation in aquatic organism consumed by humans (Luoma 1990;
MacFarlane & Burchett 2000).
1.1.1 Copper Pollution
According to the Agency of Toxic Substances and Disease Registry (ATSDR 2004),
approximately 640,000,000 kg of the heavy metal copper (Cu) were released into the
environment by industries in the year of 2000. Cu was released into the environment from
phosphate fertilizer production, agriculture and mining activity, metal and wood
production, and metal waste dumps (Nriagu & Pacynat 1988; ATSDR 2004). High
concentrations of Cu can often be found near waste disposal sites, smelters, mines,
landfills, and industrial settings (ATSDR 2004). Cu does not break down easily in the

- 2 -
environment (ATSDR 2004), thus potentially causing pollution and negative impacts to
the environment.
Acid mine drainage (AMD) is often referred to as acid rock drainage (ARD), caused by
acid drainage from the mine waste rock, tailings and mine structures (pits and
underground workings; U.S. Environmental Protection Agency 1994). The generation of
acid by oxidation of sulphur and the precipitation of ferric iron occurs when the sulphide
and elemental sulphur containing minerals are exposed to the weathering effects of water
and oxygen (Price & Errington 1998). This will result in water acidity, thus causing the
elevated leaching of metals, such as silver, cadmium, arsenic, zinc and copper (U.S.
Environmental Protection Agency 1994), due to high metal solubility and sulphide
weathering rate under acidic conditions (Price & Errington 1998). Metals can travel long
distances when dissolved in water, resulting in the contamination of streams and
groundwater and therefore causing significant environmental impact and threaten the
water sources on which we all depend. AMD has been described as the largest
environmental problem facing the U.S. mining industry (USDA Forest Service 1993;
Ferguson & Erickson 1988; Lapakko 1993). More than 7,000 kilometres of streams
affected by acid drainage from coal mines in the Eastern U.S. have been reported by Kim
et al. (1982). Besides that, according to USDA Forest Service (1993) there are between
20,000 and 50,000 mines currently generating acid, resulting in an acid drainage impact
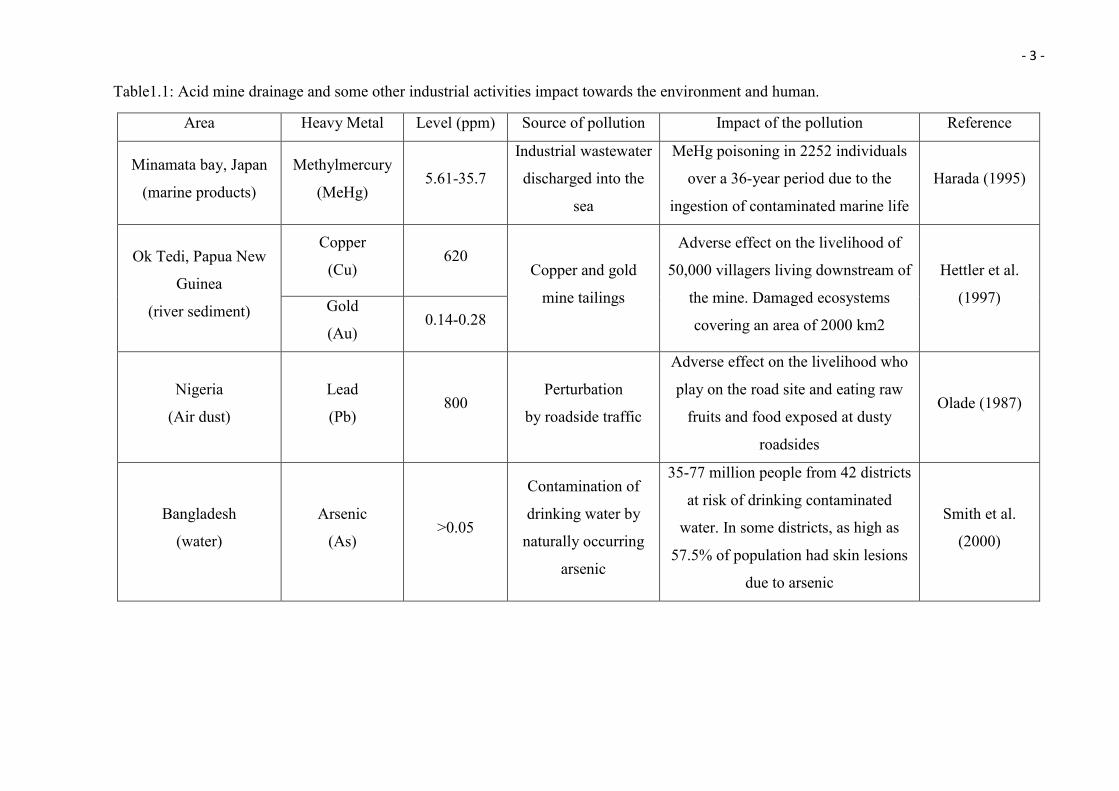
on 8,000 to 16,000 km of streams (in the Western U.S.). See Table 1.1 for examples of
AMD and some other industrial activities impacting the environment and humans.
- 3 -
Table1.1: Acid mine drainage and some other industrial activities impact towards the environment and human.
Area Heavy Metal Level (ppm) Source of pollution Impact of the pollution Reference
Minamata bay, Japan
(marine products)
Methylmercury
(MeHg) 5.61-35.7
Industrial wastewater
discharged into the
sea
MeHg poisoning in 2252 individuals
over a 36-year period due to the
ingestion of contaminated marine life
Harada (1995)
Ok Tedi, Papua New
Guinea
(river sediment)
Copper
(Cu) 620
Copper and gold
mine tailings
Adverse effect on the livelihood of
50,000 villagers living downstream of
the mine. Damaged ecosystems
covering an area of 2000 km2
Hettler et al.
(1997) Gold
(Au) 0.14-0.28
Nigeria
(Air dust)
Lead
(Pb) 800
Perturbation
by roadside traffic
Adverse effect on the livelihood who
play on the road site and eating raw
fruits and food exposed at dusty
roadsides
Olade (1987)
Bangladesh
(water)
Arsenic
(As) >0.05
Contamination of
drinking water by
naturally occurring
arsenic
35-77 million people from 42 districts
at risk of drinking contaminated
water. In some districts, as high as
57.5% of population had skin lesions
due to arsenic
Smith et al.
(2000)
- 4 -
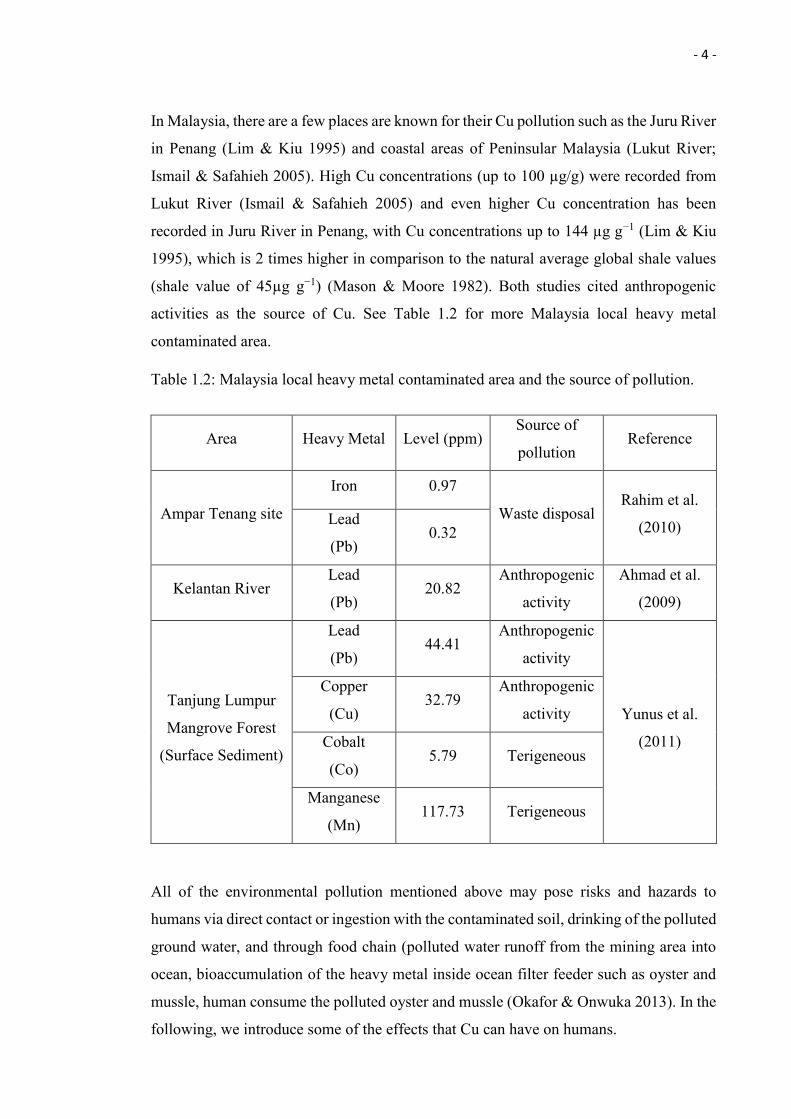
In Malaysia, there are a few places are known for their Cu pollution such as the Juru River
in Penang (Lim & Kiu 1995) and coastal areas of Peninsular Malaysia (Lukut River;
Ismail & Safahieh 2005). High Cu concentrations (up to 100 µg/g) were recorded from
Lukut River (Ismail & Safahieh 2005) and even higher Cu concentration has been
recorded in Juru River in Penang, with Cu concentrations up to 144 µg g−1 (Lim & Kiu
1995), which is 2 times higher in comparison to the natural average global shale values
(shale value of 45µg g−1) (Mason & Moore 1982). Both studies cited anthropogenic
activities as the source of Cu. See Table 1.2 for more Malaysia local heavy metal
contaminated area.
Table 1.2: Malaysia local heavy metal contaminated area and the source of pollution.
All of the environmental pollution mentioned above may pose risks and hazards to
humans via direct contact or ingestion with the contaminated soil, drinking of the polluted
ground water, and through food chain (polluted water runoff from the mining area into
ocean, bioaccumulation of the heavy metal inside ocean filter feeder such as oyster and
mussle, human consume the polluted oyster and mussle (Okafor & Onwuka 2013). In the
following, we introduce some of the effects that Cu can have on humans.
Area Heavy Metal Level (ppm) Source of
pollution Reference
Ampar Tenang site
Iron 0.97
Waste disposal Rahim et al.
(2010) Lead
(Pb) 0.32
Kelantan River Lead
(Pb) 20.82
Anthropogenic
activity
Ahmad et al.
(2009)
Tanjung Lumpur
Mangrove Forest
(Surface Sediment)
Lead
(Pb) 44.41
Anthropogenic
activity
Yunus et al.
(2011)
Copper
(Cu) 32.79
Anthropogenic
activity
Cobalt
(Co) 5.79 Terigeneous
Manganese
(Mn) 117.73 Terigeneous

- 5 -
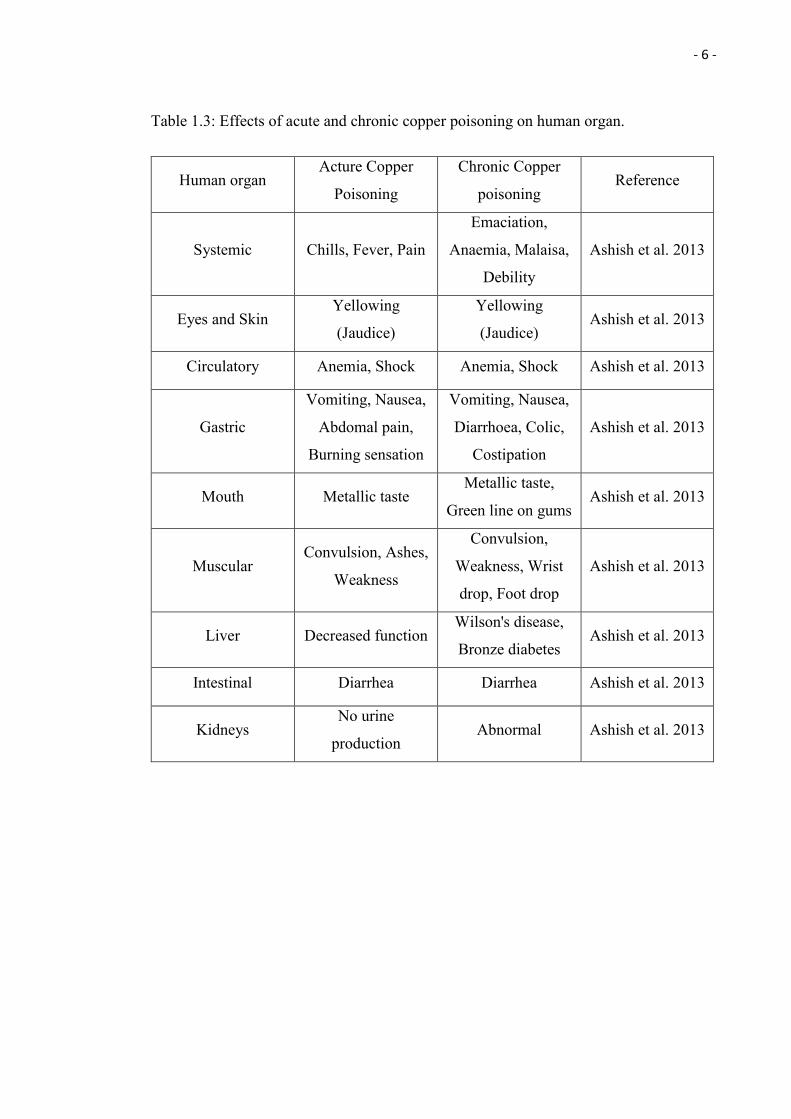
1.1.2 Copper Toxicity
Although Cu is one of the essential micronutrients for humans, it can be extremely toxic
if taken in high concentrations. According to Lester (1987), the daily requirement of
copper for an adult is only 0.03 mg/kg and reports of Cu toxicity are becoming more
common these days (Ashish et al. 2013). Symptoms such as vomiting, abdominal pain,
and nausea may occur when one is exposed to acute copper poisoning (Stern et al. 2007;
Ashish et al. 2013). Acute poisoning is rarely seen (Gamakaranage et al. 2011), however,
chronic poisoning can be found happening at those countries, which copper plumbing is
common. For instance, in Germany, all of the patients who suffered from a series of
severe systemic diseases such as copper induced liver cirrhosis and chronic copper
poisoning induced gastrointestinal diseases were found to have copper plumbing that
released copper into the tap water, in their houses. Besides that, the patients were also
recorded to be suffered from nausea, vomiting, colic, and diarrhoea (Eife et al.
1999). Moreover, Ashish et al. (2013) also reported that hepatic necrosis, anaemia,
hypotension, proteinuria, acute renal tubular failure, tachycardia, vascular collapse,
haemoglobinuria, and death may occur when long term exposure to Cu. Table 1.3 below
summarises the impacts of Cu on humans.
Their toxicity depends on:
• Bioavailability: absorption in the gastrointestinal tract, transport to other cell types,
uptake by them.
• Solubility under physiological conditions
• Chemical species: elements are not generally used by cells in their elemental form.
They are used in ionic, complex or organo-metallic forms.
Copper can be found widely distributed in biological tissues and is involved in important
physiological functions in the human body (World Health Organization 1996). It acts as
cofactor for several enzymes in our body such as cytochrome oxidase, ceruloplasmin,
superoxide dismutase, tyrosinase, and dopamine β-hydroxylase (Ashish et al. 2013).
Besides that, it also helps to transmit electrical signals and facilitates the absorption of
iron in our body (Ashish et al. 2013). Insufficient intake of copper may lead to blood
circulation problems, anaemia and growth inhibition (Jennings & Sneed 1996).
- 6 -
Table 1.3: Effects of acute and chronic copper poisoning on human organ.
Human organ Acture Copper
Poisoning
Chronic Copper
poisoning Reference
Systemic Chills, Fever, Pain
Emaciation,
Anaemia, Malaisa,
Debility
Ashish et al. 2013
Eyes and Skin Yellowing
(Jaudice)
Yellowing
(Jaudice) Ashish et al. 2013
Circulatory Anemia, Shock Anemia, Shock Ashish et al. 2013
Gastric
Vomiting, Nausea,
Abdomal pain,
Burning sensation
Vomiting, Nausea,
Diarrhoea, Colic,
Costipation
Ashish et al. 2013
Mouth Metallic taste Metallic taste,
Green line on gums Ashish et al. 2013
Muscular Convulsion, Ashes,
Weakness
Convulsion,
Weakness, Wrist
drop, Foot drop
Ashish et al. 2013
Liver Decreased function Wilson's disease,
Bronze diabetes Ashish et al. 2013
Intestinal Diarrhea Diarrhea Ashish et al. 2013
Kidneys No urine
production Abnormal Ashish et al. 2013

- 7 -
1.2 Current Technologies for Heavy Metal Removal
Unlike organic pollutants, which can be broken down by microbial communities, heavy
metals do not undergo chemical or microbial degradation (Kirpichtchikova et al. 2006).
Besides that, presence of heavy metals can also negatively affect microbial degradation
of organic contaminants (Maslin & Maier 2000). Therefore, other solutions have to be
found to remove heavy metal contaminants.
In recent years, several technologies have been introduced to remove metal contaminants
such as chemical precipitation, phytoremediation (Vaajasaari & Joutti 2006; Wenzel
2009), membrane filtration, ion exchange (Kurniawana et al 2006), electrodialysis
(Mohammadi et al. 2005), photocatalysis (Barakat et al. 2004; Kajitvichyanukula et al.
2005), reverse osmosis (Rich & Cherry 1987), and biosorption (Fourest & Roux 1992).
1.2.1 Chemical Precipitation
Precipitation is one of the common methods that have been used by almost 75% of the
plating companies to extract heavy metals from solutions (Cushnie 1985). The method
works by converting soluble metal salts from the solution into insoluble salts; thereby
precipitating the heavy metal out of the solution. This step is followed by settling
(clarification) and filtration to remove the metal precipitates from the treated solution
effluent (Seneviratne 2007). It was recorded to be simple to operate, cost effective
(Kurniawan et al. 2006), and has proven to effectively remove heavy metals from
electroplating waste water. For instance, Abrego (1997) reported the use of precipitation
and flotation to remove a wide variety of metal ions, such as toxic metal ions nickel (II),
aluminium (III), lead (II), iron (II) and tin (II), within a wide range of pH (3-12) from
contaminated samples. The metal ions were able to be removed below levels required by
anti-pollution regulations (Abrego 1997).
Although the cost for the treatment is known to be low and the operation is simple, it will
result in large volumes of toxic sludge generation, which requires proper disposal;
however, sludge disposal is cost intensive. Besides that, it is difficult to precipitate the
metals held in solution by complexing agents such as EDTA and cyanide. Moreover, the

- 8 -
addition of the reagents has to be carefully controlled in order to avoid unacceptable
concentrations in the treatment effluent (Seneviratne 2007).
1.2.2 Ion Exchange
Ion exchange is one of the common methods that have been used successfully for metal
ion removal from industry effluents. The treatment uses insoluble polymers (resins) that
contain acidic or basic functional groups to exchange the counter-ions from the
surrounding solution. Da̧browski et al. (2004) and Gold et al. (1987) have proven the
simplicity of the method and the efficiency to remove metal ions such as chromium
(III,VI), cadmium (II), mercury (II), vanadium (IV,V), nickel (II), zinc (II), copper(II)
and lead (II), from industrial wastewaters and contaminated electroless. Gold et al. (1987)
also proved that the resins can be high selective. Amberlite IRC-718 resin, for example,
is more selective to copper (94%) than nickel and is relatively selective to zinc and lead
(50%). Duolite ES- 467 resin is on the other hand more selective to zinc and lead (89%)
and relatively selective to copper (75%).
Although ion exchange has been proven to be simple to operate Da̧browski et al. (2004),
efficient in metal removal Gold et al. (1987), low in maintenance cost and requires only
little energy (Griffin 2011), there are several disadvantages that need to be noted. It is pH
sensitive and not able to handle highly concentrated metal solutions (Baysal et al. 2013).
Besides that, it is also easy to be blocked by organics matter and other solids present in
the wastewater such as calcium sulphate and iron (Griffin 2011).
1.2.3 Electrodialysis
Electrodialysis is a process that uses semipermeable ion-selective membranes (Cho et al.
2010) and an electric potential difference to separate the ionic component from a solution
and other uncharged components (Strathmann 1992). This separation process is able to
maintain the metal ions in low concentrations within the anodizing bath solution. The
metal ions are selectively transported through the selective membrane with the electrical
current induced flow (Cho et al. 2010). This method has been used by Pedersen (2003) to
remove cadmium from wood fly ash, with the assisting agent a mixture of ammonium

- 9 -
citrate (0.25 M) and ammonia (1.25%). Besides that, it has also been recorded to be able
to recover silver, copper, nickel, lead, gold, tin, and zinc from cyanide bath rinse solution
(U.S. Environmental Protection Agency 2013).
This electodialysis method is known to have the advantages of high selectivity and
recovery of the metal ions from the solution (Barakat 2011). However, there are several
disadvantages of the method that need to be noted such as the clogging of the membrane
by metal hydroxide formation (Ahalya 2003), the requirement for periodic maintenance,
and technically challenging operation and handling (Barakat 2011).
1.2.4 Semiconductor Photocatalysis
Semiconductor photocatalysis is one of the latest methods used to remove and recover
metal ions from wastewater. It is a process based on reduction by photo generated
electrons and uses ultraviolet (UV) light and semiconductor particles, such as CdS, CeO2,
ZnO, ZnS and TiO2, as catalyst. Electron–hole pairs (e−/h+) are formed in the conduction
and the valence band of the semiconductor, respectively, once the semiconductor particles
are illuminated by UV light with the energy greater than the semiconductor band gap
energy (Herrmann 1999). The charge carriers will then migrate to the surface of the
semiconductor and conduct the reducing/ oxidizing reaction on the component (metal
ions) within the solution. The method has been shown by Barakat et al. (2004) to be able
to remove 78% of free cyanide (10−3 M) within 4 hours of illumination, and free copper
(10−2 M) within an even shorter time (3 h).
The photocatalysis method is known to remove organic and metal pollutants
simultaneously with less harmful by-products. However, it does possess severe
limitations such as a poor overlap of the solar spectrum with the absorption spectrum of
TiO2 (less than 5%). This can be improved by doping the TiO2 with metal ion, which will,
however, significantly increase the cost of the photocatalyst (Malato et al. 2014). Besides
that, there are others limitations such as a limited range of applications and a long duration
time (Barakat 2011). Deactivation of the photocatalyst by strongly adsorbing end

- 10 -
products onto the surface of the photocatalysis can be another limiting factor (Pichat
2013).
1.2.5 Membrane Filtration
Membrane filtration is gaining attention in many industries for its ability to remove
inorganic contaminants, such as heavy metals, from wastewater. Besides that, it is also
known to be capable of removing suspended solid and organic compounds. Various
membrane filtration types can be used for heavy metal removal from wastewater
(depending on the size of the particles), such as ultrafiltration, nanofiltration and reverse
osmosis. (Vigneswaran et al. 2005; Dyson et al. 2008; Wang et al. 2009).
Ultrafiltration (UF) is the use of a permeable membrane to separate suspended solids,
macromolecules, and heavy metals from inorganic solutions. The separation is based on
the basis of the molecular weight (1000-100,000 Da) and pore size (5-20 nm) of the
separating compounds (Vigneswaran et al. 2005). These will allow the small molecule
such as water, which has the size of 0.38 nm (Ngai 2011), to pass through the membrane,
and at the same time, retain the other molecules that have a size larger than the pore size
of the membrane (Sablani et al. 2001).
Nanofiltration (NF), with a membrane pore size of 0.5 – 5 nm (Dyson et al. 2008), uses
separation mechanisms that involve electrical (Donnan) and steric (sieving) effects. A
Donnan potential is created between the charged anion within the membrane and the co-
ions within the effluent, and therefore it creates the conditions to reject the latter ones
(Wang et al. 2009). More specifically, this technique uses the membrane’s small pore size
and surface charges to reject and prevent the charged solutes smaller than the membrane
pore with the bigger neutral ones to pass though the membrane. This technique has been
proven to be able to remove more than 90% of copper ions from industrial feed water
(Qdais & Moussa 2004).

- 11 -
Reverse Osmosis (RO), with a membrane pore size lower than 0.5 nm (Dyson et al. 2008),
is known to be more efficient than UF and NF for heavy metal removal from inorganic
solutions (Wang et al. 2009). The percentage of heavy metal rejection is up to 97% at
metal concentrations between 20-200 mg/L. The technique uses pressure on the heavy
metal solution to force the fluid to pass through the membrane. Therefore, the heavy metal
and purified water will be separated, retained and accumulated on two different sides of
the membrane (Wang et al. 2009). For instance, Benito and Ruíz (2002) were able to
recover up po 95% of the clean water from the polluted water. Qdais and Moussa (2004)
were able to remove 98% of copper and 99% of cadmium from industrial wastewater.
Although the membrane filtration technologies mentioned above shows high recovery of
metal ions and clean water. However, the high operational costs and sensitivity to oxidant,
pH and chlorine, liable fouling, compaction, scaling and limited life of the membrane are
major disadvantages of these technologies (Kurniawan et al. 2006; Wang et al. 2009;
Wang et al. 2010).
1.2.6 Phytoremediation
Phytoremediation is one of the green technologies that uses plants to remove organic and
inorganic pollutants from the environment (Erakhrumen 2007). This technique has been
previously recorded to be used for heavy metal removal from metal contaminated land
(Pulford & Watson 2003) and wastewater (Singh et al. 2012). There are five different
types of phytoremediation that utilise different ways of removing pollutants from the
environment (Wang et al. 2008; Mirsal 2013).
Phytoextraction: A technology that uses hyper-accumulating plants to transport, and to
accumulate contaminants from the soil into the plant roots and aboveground shoots. The
contaminants can be removed by harvesting the plants (Wang et al. 2008; Jadia & Fulekar
2009; Mirsal 2013).

- 12 -
Phytodegradation: A technology that uses plants to degrade or breakdown organic
contaminants mainly through enzymatic reactions (Mirsal 2013).
Rhizofiltration: A technology that uses plant fibrous root systems to absorb, accumulate
and precipitate the contaminants from wastewater (Mirsal 2013; Wang et al. 2008). It is
similar to the phytoextraction technology, however, this technology is mostly used in
aquatic enviroments (Jadia & Fulekar 2009; Mirsal 2013).
Phytostablisation: A technology that uses highly tolerant plants to limit the mobility and
bioavailability of the contaminant in the soil by complexation, precipitation, or sorption
(Wang et al. 2008).
Phytovolatilisation: A technology using plants (mainly trees (Mirsal 2013)) to uptake,
transform and evaporate the contaminant into the atmosphere (Jadia & Fulekar 2009).
Please refer to Table 1.4 for the advantages and disadvantages of each of the five above
mentioned techniques.
Although phytoremediation technology is known to be cost effective and displays high
efficiency in removing metal contaminants, it is pH sensitive, time consuming, has a high
cost and long term maintenance is needed. Besides that, the consumption of the
contaminants polluted plant biomass by herbivores might cause the contaminants to enter
the food chain (Tangahu et al. 2011).

- 13 -
Table 1.4: Overview of advantages and disadvantages of each of the five
phytoremediation techniques /approaches.
Technologies Advantages Disadvantages
Phytoextraction Cost effective, permanent
removal of the contaminant
from soil (Prasad 2004; Wang
et al. 2008)
Time consuming due to slow
growing plants (Prasad 2004;
Wang et al. 2008)
Phytodegradation Able to degrade a wide range
of contaminants (Pivetz 2001)
Possibility of toxic
intermediates or products
formation (Gaspard & Ncibi
2013)
Rhizofiltration Relocation of the system
(plant) is easy (Prasad 2004)
pH sensitive, pre-grown plant
is needed (Prasad 2004)
Phytostabilization Cost effective (Prasad 2004) Long-term maintenance is
required, Contaminants still
remain in the environment
(Prasad 2004; Wang et al.
2008)
Phytovolatilization Converting toxic
contaminants into less-toxic
form, Rapid natural
degradation process (Prasad
2004)
Accumulation of the
contaminant in the vegetation
(Prasad 2004)
1.3 Biosorption
The technologies mentioned above (chemical precipitation, ion exchange, electrodialysis,
photocatalysis, membrane filtration, reverse osmosis, and phytoremediation) are either
expensive, pH sensitive, or easily get fouled Volesky (1990). Besides that, most of those
them may create secondary problems such as toxic sludge formation, and one of them
(phytoremediation) may even cause the contaminant to enter food chains (Tangahu et al.
2011). Therefore, an alternative technology, which involves a biological process between
a solid phase (biological material) and a liquid phase (solvent) containing a dissolved

- 14 -
component to be sorbed (contaminants), has been extensively studied for the past decades
(Volesky 1990). This technology, called biosorption, is known to be highly efficient in
contaminant removal, possesses high contaminant uptake rates, is environmentally
friendly, cost effective, active in a wide range of pH (Volesky 1990; Lee 2014), and
highly selective (Volesky 1990). These advantages have made biosorption so attractive
for the removal of toxic heavy metal contaminants from polluted wastewater. Besides that,
it is also known to be able to regenerate the biosorbent easily (Kratochvil & Volesky 1998)
and the metal can be recovered (Volesky 1990).
Biosorption was defined as the concentration and accumulation of pollutants from
aqueous solutions by the use of biological materials (biosorbent) and therefore allows the
recovery and (or) environmentally acceptable disposal of the pollutants (Dönmez et al.
1999). Biosorption consist of several mechanisms, such as crystallization adsorption,
chelation, precipitation and ion exchange, followed by ion entrapment in intrafi and inter-
brillar capillaries, diffusion through the cell wall and membranes, and spaces of the
polysaccharide material, which vary depending on the origin of the biomass, the
processing steps and the species used (Singh 2006). Biosorption can be divided into two
categories, which are metabolism dependent (active uptake) and metabolism independent
(passive uptake). Metabolism dependent (in which the contaminant will be taken up and
transported across cell membrane) is also known as active biosorption or bioaccumulation
and is associated with cell metabolic activities. On the other hand, metabolism
independent, which is also known as passive biosorption, does not depend on the cell
metabolic activity. It depends on the functions of the chemical composition of the cell
wall. The metabolism-independent biosorption takes place in both live and dead microbial
cells, while metabolism-dependent biosorption can only occur within alive microbial cells
(Lee 2014).
- 15 -
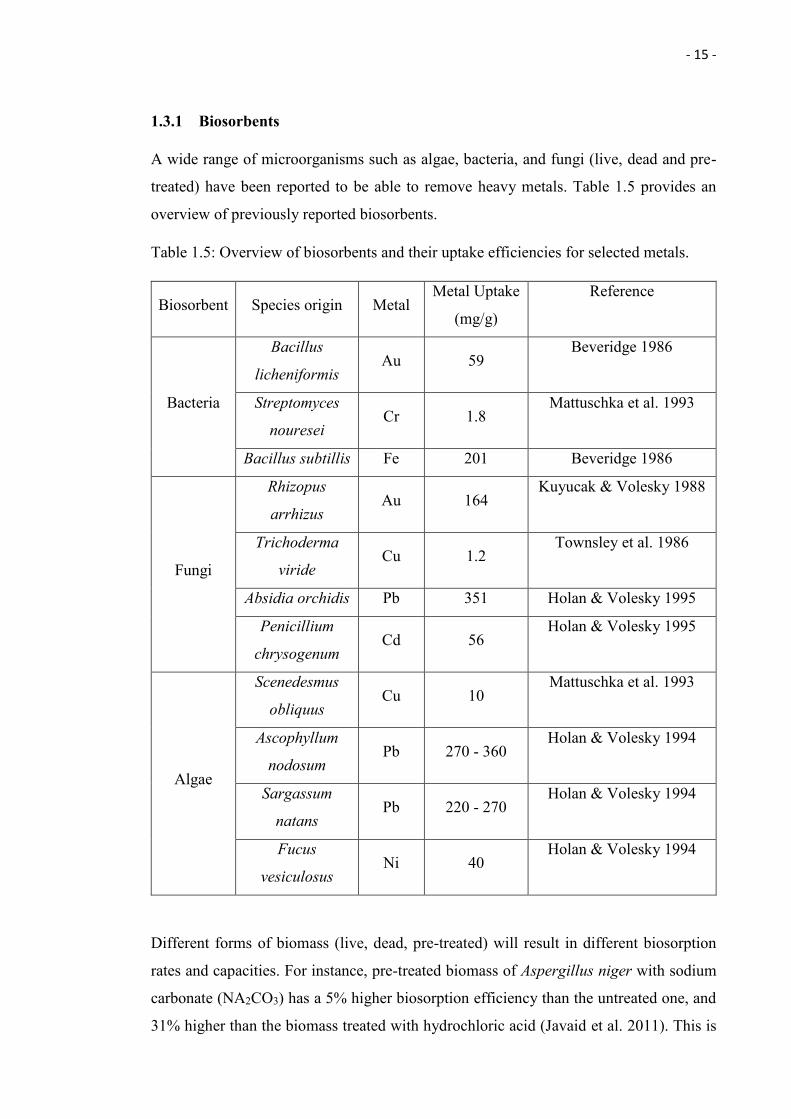
1.3.1 Biosorbents
A wide range of microorganisms such as algae, bacteria, and fungi (live, dead and pre-
treated) have been reported to be able to remove heavy metals. Table 1.5 provides an
overview of previously reported biosorbents.
Table 1.5: Overview of biosorbents and their uptake efficiencies for selected metals.
Biosorbent Species origin Metal Metal Uptake
(mg/g)
Reference
Bacteria
Bacillus
licheniformis Au 59
Beveridge 1986
Streptomyces
nouresei Cr 1.8
Mattuschka et al. 1993
Bacillus subtillis Fe 201 Beveridge 1986
Fungi
Rhizopus
arrhizus Au 164
Kuyucak & Volesky 1988
Trichoderma
viride Cu 1.2
Townsley et al. 1986
Absidia orchidis Pb 351 Holan & Volesky 1995
Penicillium
chrysogenum Cd 56
Holan & Volesky 1995
Algae
Scenedesmus
obliquus Cu 10
Mattuschka et al. 1993
Ascophyllum
nodosum Pb 270 - 360
Holan & Volesky 1994
Sargassum
natans Pb 220 - 270
Holan & Volesky 1994
Fucus
vesiculosus Ni 40
Holan & Volesky 1994
Different forms of biomass (live, dead, pre-treated) will result in different biosorption
rates and capacities. For instance, pre-treated biomass of Aspergillus niger with sodium
carbonate (NA2CO3) has a 5% higher biosorption efficiency than the untreated one, and
31% higher than the biomass treated with hydrochloric acid (Javaid et al. 2011). This is

- 16 -
due to chemical modification of the binding site of the fungal biomass, and thus increased
numbers of active binding sites on the surface area.
Other than microorganisms, plant biomass and mammalian polymers also showed to be
able to biosorb certain metals. For instance, Elifantz and Tel-Or (2002) showed that the
biomass of the Macrophyte, Ludwigia stolonifera, can be used to biosorb- heavy metal
cadmium (Cd) and nickel (Ni). Besides that, Ratnakumari and Sobha (2012) highlighted
the capability of animal polymers, chick and duck feathers, to biosorb the heavy metal
copper (Cu). However, biosorption using microorganisms have been used by most of the
researches (Javaid et al. 2011; Beveridge 1986; Mattuschka et al. 1993; Kuyucak &
Volesky 1988a, Townsley et al. 1986; Holan & Volesky 1995; Holan & Volesky 1994).
Fungal biomass seems to be getting more attention and has been studied more extensively
(Wang et al. 2010). This is mainly due to the reason that fungi possess a wide
morphological variety and can be manipulated morphologically and genetically. Fungal
cell wall is composed of chitosans, glucans, and chitin (Singh 2006). Besides that, it also
contains proteins, lipids, and other polysaccharides. Fungi generate biomass fast due to
short multiplication cycles and can be cultured easily using unsophisticated fermentation
technique (Fulekar 2012; Lee 2014), yielding large quantities of fungal biomass and
derivatives (Lee 2014). Moreover, fungal biomass contains high amounts of cell wall
material such as chitin and chitosan which possess excellent metal-binding properties
(Lee 2014; Gadd 2004). Bishnoi and Garima (2005) and Ahmad et al. (2011) showed that
fungal biomass has better biosorption capacity of heavy metal compared to the other
conventional absorbents such as activated carbon (Nuchar SA) and algae. Fungal biomass
performs better in terms of biosorption of heavy metals compared to ion-exchange resins
which only contain monofunctional groups. This is due to the fact that fungal biomass
contains a much higher variety of functional sites such as sulfate, carboxyl, hydroxyl,
sulfonate, amino, phosphate, imino, sulfydryl, thioether, carbonyl, and imidazole groups
(Singh 2006).
The focus of this thesis is on fungi, in particular endophytic fungi and the following
provides an introduction to the world of endophytes.

- 17 -
1.4 Fungal-plant Symbiotic Interaction
Fossil records indicate that symbiotic interactions between fungi and plants have taken
place since at least 400 million years ago (Krings et al. 2007). A successful fungal-plant
symbiosis involves three different stages:
(a) penetration of the fungus into plant tissues,
(b) colonization of the the host plant tissue by the fungus,
(c) expression of the fungal symbiotic lifestyle (Singh et al. 2011).
There are a few different outcomes of symbiotic interaction as defined by the fitness
benefits realized by both of fungi and the host plant (Lewis 1985). In fungal-plant
symbiosis, the benefits to fungal symbionts can be positive (parasitism, commensalism
and mutualism), neutral (neutralism and amensalism) or negative (competition), while for
the host plant it can also be positive (mutualism), neutral (neutralism and commensalism)
or negative (parasitism, amensalism and competition; Rodriguez et al. 2008).
1.4.1 Endophytic Fungi
An endophytic fungi is a fungus that lives symbiotically with a host plant without showing
any apparent symptoms. Endophytic fungi protect their host plant from biotic and abiotic
stress such as increasing their stress tolerance (for example against drought, salinity, and
heavy metals). They can enhance the growth of their host plants by reducing the infection
rate of nematodes and defending the plant from diseases (Tadych & White 2009; Sikora
et al. 2008; Varma et al. 1999; Redman et al. 2002). In return, they will acquire nutrients
from their host plant (Tadych & White 2009). There are two different mechanisms
involved in the endophytic fungi-conferred stress tolerance, which are (a) rapid activation
of host stress response systems after stress exposure of the symbiotic host plants (Redman
et al. 1999) and (b) synthesis of anti-stress biochemicals in the host plant, either through
endophytic fungi induction or by the endophytic fungi itself (Bacon & Hill 1996).
However, the details of how the endophytic fungi activate their host stress tolerance
/response still remains a mystery (Rodriguez et al. 2004). Many studies conducted all
around the world, have proven that endophytic fungi significantly contribute to or are
responsible for the adaptation of their host plant towards environmental stresses such as
drought, extreme temperature, high salinity, heavy metal toxicity, and oxidative stress
- 18 -
(Malinowski et al. 1997; Redman et al. 2001; Rodriguez et al. 2004; Rodriguez et al. 2008;
Soleimani et al. 2010; Monnet et al. 2001; Ren et al. 2011; Rodriguez & Redman 2008).
The endophytic fungus Penicillium minioluteum LHL09, isolated from soyabean plant
(glycine max. L.) was shown to be able to protect the host plant from abiotic salinity stress
(Khan et al. 2011). Besides that, Ren et al. (2011) and Soleimani et al. (2010) also
demonstrated the capability of endophytic fungi to increase the resistance of their host
plants against heavy metal cadmium. Moreover, Monnet et al. (2001) have shown the
capability of the endophytic fungus Neotyphodium lolii to increase zinc tolerance in
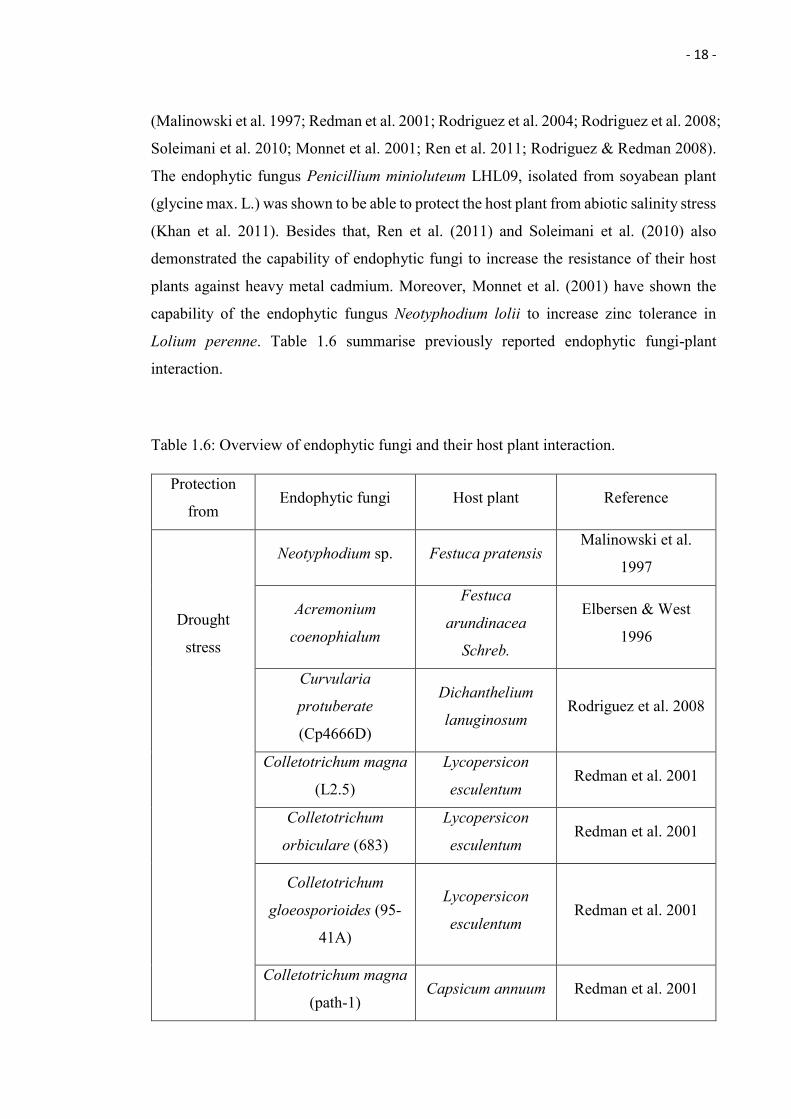
Lolium perenne. Table 1.6 summarise previously reported endophytic fungi-plant
interaction.
Table 1.6: Overview of endophytic fungi and their host plant interaction.
Protection
from Endophytic fungi Host plant Reference
Drought
stress
Neotyphodium sp. Festuca pratensis Malinowski et al.
1997
Acremonium
coenophialum
Festuca
arundinacea
Schreb.
Elbersen & West
1996
Curvularia
protuberate
(Cp4666D)
Dichanthelium
lanuginosum Rodriguez et al. 2008
Colletotrichum magna
(L2.5)
Lycopersicon
esculentum Redman et al. 2001
Colletotrichum
orbiculare (683)
Lycopersicon
esculentum Redman et al. 2001
Colletotrichum
gloeosporioides (95-
41A)
Lycopersicon
esculentum Redman et al. 2001
Colletotrichum magna
(path-1) Capsicum annuum Redman et al. 2001
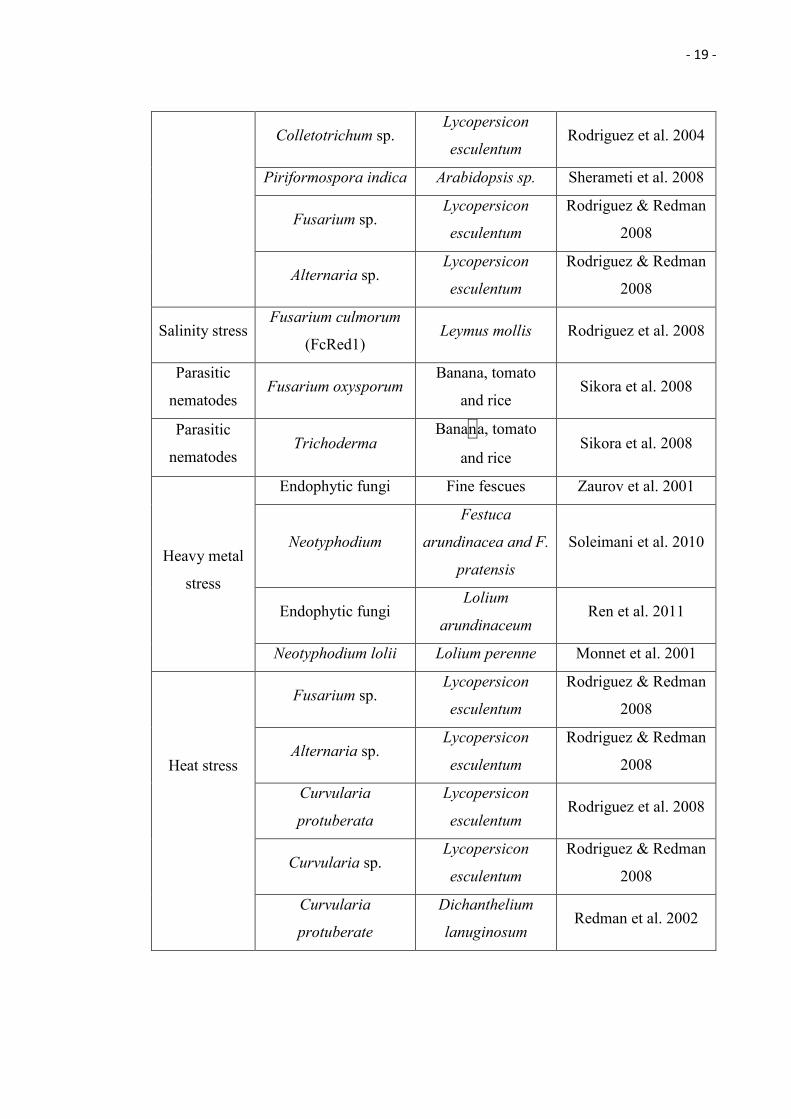
- 19 -
Colletotrichum sp. Lycopersicon
esculentum Rodriguez et al. 2004
Piriformospora indica Arabidopsis sp. Sherameti et al. 2008
Fusarium sp. Lycopersicon
esculentum
Rodriguez & Redman
2008
Alternaria sp. Lycopersicon
esculentum
Rodriguez & Redman
2008
Salinity stress Fusarium culmorum
(FcRed1) Leymus mollis Rodriguez et al. 2008
Parasitic
nematodes Fusarium oxysporum
Banana, tomato
and rice Sikora et al. 2008
Parasitic
nematodes Trichoderma
Banana, tomato
and rice Sikora et al. 2008
Heavy metal
stress
Endophytic fungi Fine fescues Zaurov et al. 2001
Neotyphodium
Festuca
arundinacea and F.
pratensis
Soleimani et al. 2010
Endophytic fungi Lolium
arundinaceum Ren et al. 2011
Neotyphodium lolii Lolium perenne Monnet et al. 2001
Heat stress
Fusarium sp. Lycopersicon
esculentum
Rodriguez & Redman
2008
Alternaria sp. Lycopersicon
esculentum
Rodriguez & Redman
2008
Curvularia
protuberata
Lycopersicon
esculentum Rodriguez et al. 2008
Curvularia sp. Lycopersicon
esculentum
Rodriguez & Redman
2008
Curvularia
protuberate
Dichanthelium
lanuginosum Redman et al. 2002
- 20 -
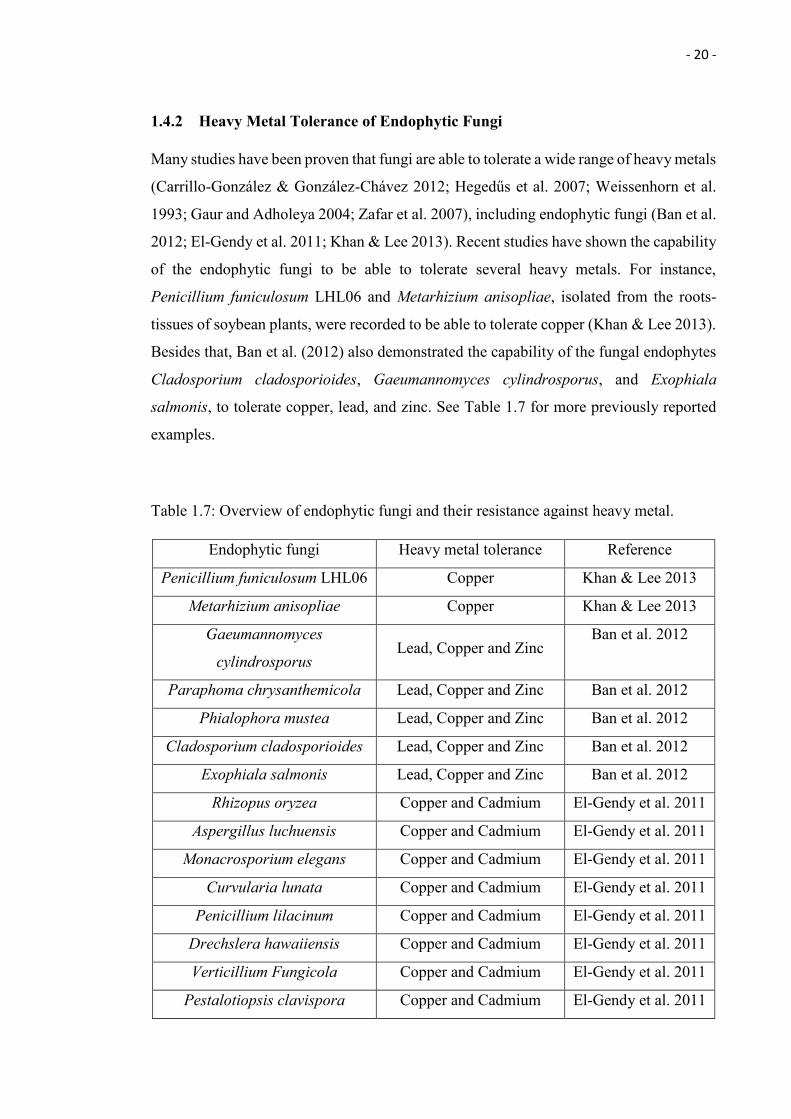
1.4.2 Heavy Metal Tolerance of Endophytic Fungi
Many studies have been proven that fungi are able to tolerate a wide range of heavy metals
(Carrillo-González & González-Chávez 2012; Hegedűs et al. 2007; Weissenhorn et al.
1993; Gaur and Adholeya 2004; Zafar et al. 2007), including endophytic fungi (Ban et al.
2012; El-Gendy et al. 2011; Khan & Lee 2013). Recent studies have shown the capability
of the endophytic fungi to be able to tolerate several heavy metals. For instance,
Penicillium funiculosum LHL06 and Metarhizium anisopliae, isolated from the roots-
tissues of soybean plants, were recorded to be able to tolerate copper (Khan & Lee 2013).
Besides that, Ban et al. (2012) also demonstrated the capability of the fungal endophytes
Cladosporium cladosporioides, Gaeumannomyces cylindrosporus, and Exophiala
salmonis, to tolerate copper, lead, and zinc. See Table 1.7 for more previously reported
examples.
Table 1.7: Overview of endophytic fungi and their resistance against heavy metal.
Endophytic fungi Heavy metal tolerance Reference
Penicillium funiculosum LHL06 Copper Khan & Lee 2013
Metarhizium anisopliae Copper Khan & Lee 2013
Gaeumannomyces
cylindrosporus Lead, Copper and Zinc
Ban et al. 2012
Paraphoma chrysanthemicola Lead, Copper and Zinc Ban et al. 2012
Phialophora mustea Lead, Copper and Zinc Ban et al. 2012
Cladosporium cladosporioides Lead, Copper and Zinc Ban et al. 2012
Exophiala salmonis Lead, Copper and Zinc Ban et al. 2012
Rhizopus oryzea Copper and Cadmium El-Gendy et al. 2011
Aspergillus luchuensis Copper and Cadmium El-Gendy et al. 2011
Monacrosporium elegans Copper and Cadmium El-Gendy et al. 2011
Curvularia lunata Copper and Cadmium El-Gendy et al. 2011
Penicillium lilacinum Copper and Cadmium El-Gendy et al. 2011
Drechslera hawaiiensis Copper and Cadmium El-Gendy et al. 2011
Verticillium Fungicola Copper and Cadmium El-Gendy et al. 2011
Pestalotiopsis clavispora Copper and Cadmium El-Gendy et al. 2011
- 21 -
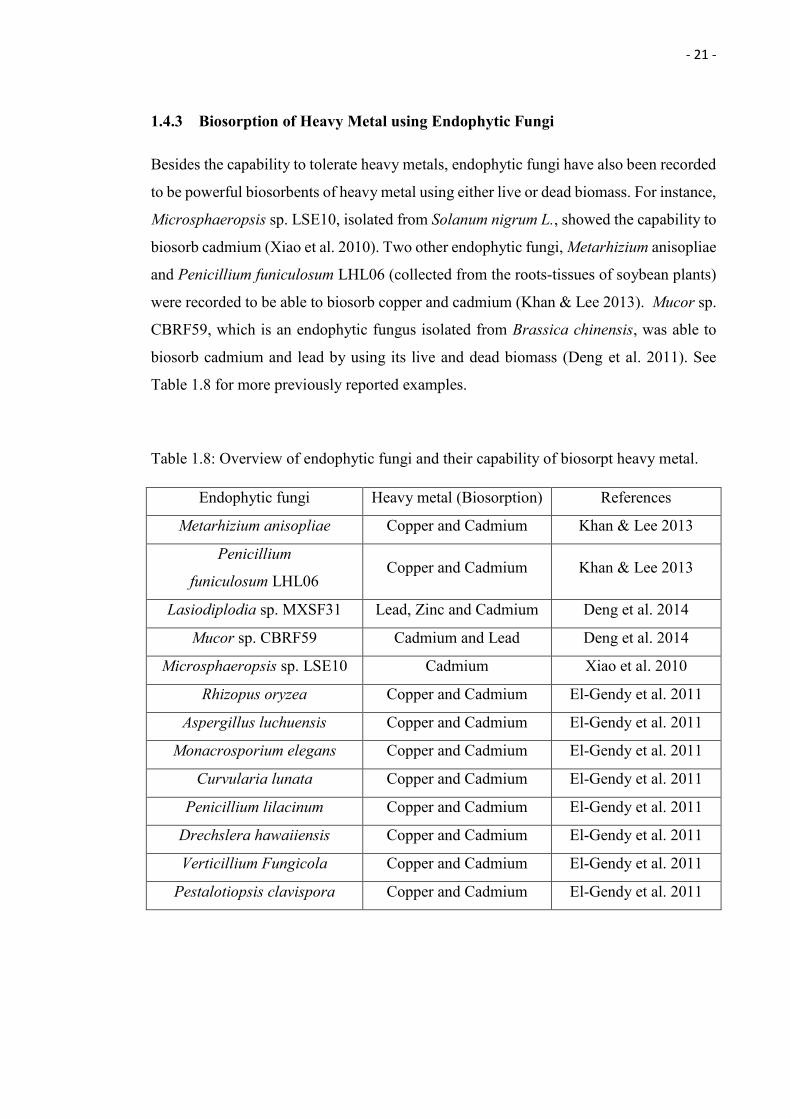
1.4.3 Biosorption of Heavy Metal using Endophytic Fungi
Besides the capability to tolerate heavy metals, endophytic fungi have also been recorded
to be powerful biosorbents of heavy metal using either live or dead biomass. For instance,
Microsphaeropsis sp. LSE10, isolated from Solanum nigrum L., showed the capability to
biosorb cadmium (Xiao et al. 2010). Two other endophytic fungi, Metarhizium anisopliae
and Penicillium funiculosum LHL06 (collected from the roots-tissues of soybean plants)
were recorded to be able to biosorb copper and cadmium (Khan & Lee 2013). Mucor sp.
CBRF59, which is an endophytic fungus isolated from Brassica chinensis, was able to
biosorb cadmium and lead by using its live and dead biomass (Deng et al. 2011). See
Table 1.8 for more previously reported examples.
Table 1.8: Overview of endophytic fungi and their capability of biosorpt heavy metal.
Endophytic fungi Heavy metal (Biosorption) References
Metarhizium anisopliae Copper and Cadmium Khan & Lee 2013
Penicillium
funiculosum LHL06 Copper and Cadmium Khan & Lee 2013
Lasiodiplodia sp. MXSF31 Lead, Zinc and Cadmium Deng et al. 2014
Mucor sp. CBRF59 Cadmium and Lead Deng et al. 2014
Microsphaeropsis sp. LSE10 Cadmium Xiao et al. 2010
Rhizopus oryzea Copper and Cadmium El-Gendy et al. 2011
Aspergillus luchuensis Copper and Cadmium El-Gendy et al. 2011
Monacrosporium elegans Copper and Cadmium El-Gendy et al. 2011
Curvularia lunata Copper and Cadmium El-Gendy et al. 2011
Penicillium lilacinum Copper and Cadmium El-Gendy et al. 2011
Drechslera hawaiiensis Copper and Cadmium El-Gendy et al. 2011
Verticillium Fungicola Copper and Cadmium El-Gendy et al. 2011
Pestalotiopsis clavispora Copper and Cadmium El-Gendy et al. 2011

- 22 -
1.5 Pitcher plants (Nepenthes) as Source of Endophytic Fungi
Nepenthes, also known as pitcher plant or monkey cup, is a type of carnivorous plant
under the family of Nepenthaceae (Adam 1997). Hundreds of species can be found
(McPherson 2009) and several studies have addressed the enzymatic properties of the
digestive fluid, trapping mechanism, and geological distribution (Adam et al. 1992;
Merbach et al. 2001; Mithöfer 2011; Slack & Gate 2000; Adam 1997; Lambers & Colmer
2006). There are, however, only very limited studies regarding endophytic fungi in
Nepenthes and one of the main aims of this thesis is to provide some baseline information
on their occurrence. Chapter 3 provides a more detailed introduction to the topic and in
the following, a short introduction to Nepenthes is provided.
Nepenthes are known to have a remarkable modified leaf that consists of a tube-shaped
cup (pitcher) at the end of the tendril. The pitcher can be divided into three different zones,
which are:
(a) Peristome, which is known to be involving in attracting and trapping prey (Figure 1.1
(Clarke 1997); Mithöfer 2011).
(b) Slippery and waxy inner zone, which used for to trap and prevent the prey from
escaping (Figure 1.1 (Clarke 1997); Gaume et al. 2002).
(c) Digestive zone, which is fully covered with glands and filled with digestive fluid
(Figure 1.1 (Clarke 1997); Gorb et al. 2004; Mithöfer 2011).
Figure 1.1: Basic structure of Nepenthes pitcher.

- 23 -
1.5.1 Distribution
Arthropods, especially insects, are attracted by the extrafloral nectar produced by
nectaries located at peristome of the pitcher (Figure 1.1; Merbach et al. 2001). A study
carried out by Kurup and the team (2013) showed that the peristome of Nepenthes
khasiana emits a distinct blue fluorescence under UV (366 nm), and it is believed that the
fluorescence is used to attract the prey. Reduction of the prey capture in the Nepenthes
khasina pitchers was observed when the blue emissions of the peristome were masked.
Once the prey in trapped and drowned within the pitcher, the digestive glands at the inner
wall of the pitcher will secrete digestive fluid to breakdown the prey. A wide variation of
the enzymes were found within the digestive fluid, such as lipase, ribonuclease, acid
phosphatase protease, and esterase (Slack & Gate 2000).
Nepenthes are widely distributed in Borneo, Peninsular Malaysia, and Sumatra, such as
Nepenthes gracilis, Nepenthes mirabilis, and Nepenthes ampullaria and several species
(Nepenthes rajah, Nepenthes xalisaputrana, Nepenthes villosa; Adam et al. 1992; see
Figure 1.2).
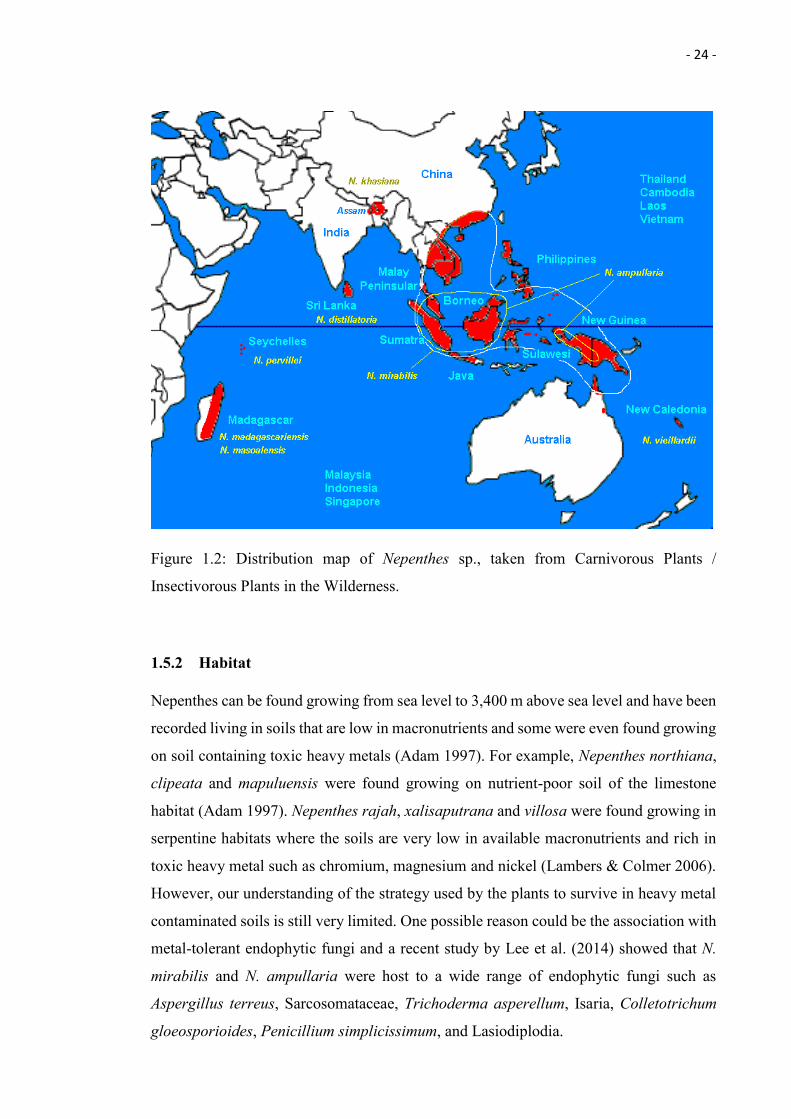
- 24 -
Figure 1.2: Distribution map of Nepenthes sp., taken from Carnivorous Plants /
Insectivorous Plants in the Wilderness.
1.5.2 Habitat
Nepenthes can be found growing from sea level to 3,400 m above sea level and have been
recorded living in soils that are low in macronutrients and some were even found growing
on soil containing toxic heavy metals (Adam 1997). For example, Nepenthes northiana,
clipeata and mapuluensis were found growing on nutrient-poor soil of the limestone
habitat (Adam 1997). Nepenthes rajah, xalisaputrana and villosa were found growing in
serpentine habitats where the soils are very low in available macronutrients and rich in
toxic heavy metal such as chromium, magnesium and nickel (Lambers & Colmer 2006).
However, our understanding of the strategy used by the plants to survive in heavy metal
contaminated soils is still very limited. One possible reason could be the association with
metal-tolerant endophytic fungi and a recent study by Lee et al. (2014) showed that N.
mirabilis and N. ampullaria were host to a wide range of endophytic fungi such as
Aspergillus terreus, Sarcosomataceae, Trichoderma asperellum, Isaria, Colletotrichum
gloeosporioides, Penicillium simplicissimum, and Lasiodiplodia.

- 25 -
Chapter 4 studied the ability of Nepenthes-associated endophytic fungi.
1.6 Proteomics - Regulation of Fungi Proteins in Response to Heavy Metal Stress
The proteome, defined as a complete set of the proteins expressed by a genome (Wilkins
et al. 1996) and “the proteins present in one sample (tissue, organism, and cell culture) at
a certain point in time” (Ravi et al. 2013), is dynamic and different from the genome,
which is relatively static. Every organism has only one unique genome, the proteome,
however, can be varied and even result in different phenotypes. For example, three
different stages of beetle life cycle (larve, pupa, and beetle) share one common genome
but vary in proteomes. The proteome often undergoes changes in response to the extra-
and intracellular environmental signals (Rastogi et al. 2006; Ravi et al. 2013), and all of
the changes can be studied through proteomics, the study of the proteome (Pandey &
Mann 2000).
Heavy metal induced oxidative stress will cause cellular damages to proteins (Letelier et
al. 2005), lipids (Zhao et al. 2014), and nucleic acids (Linder 2012) within an organism.
In order to protect itself from the damages and survive through the stress conditions, fungi
are known to regulate certain types of proteins within the cell which are involved in cell
protection (Yιldιrιm et al. 2011). This regulation of the proteins can be studied and
understood by using the proteomics approach (Jensen 2006). Some of the common
proteins and enzymes involved in cell protection are:
(a) Antioxidant enzyme - helps to protect against oxidative cellular damage (Angelova et
al. 2005).
(b) Heat shock protein - serves as molecular chaperones that play an important role in
protein-protein interactions and prevent against protein mis-folding and aggregation
(Borges and Ramos 2005; Csermely & Yahara 2003).
(c) DNA repairing enzyme - repairs errors occurring during DNA recombination and
replication (Mol et al. 1995).

- 26 -
The proteins mentioned above (a, b, c) are examples of proteins known to be upregulated
when exposed to heavy metal stress. For example, a white rot fungus, Phanerochaete
chrysosporium, is known to produce heat shock protein and DNA repairing enzyme in
response to lead exposure. Besides that, (Azevedo et al. 2007) also demonstrated the
activation of the antioxidant defence system in two aquatic fungi, Heliscus submersus and
Varicosporium elodeae, in response to zinc and copper stress.
Proteomics allows quantitative and quanlitative measurements of the fungal proteins and
the information obtained is important for our understanding of proteins involved in
cellular processes. This method allows an accurate analysis of cellular system changes in
response to different copper concentration.
1.7 Aims of the Present Study and Dissertation Outline
The first aim of the present study is to assess the Nepenthes ampullaria associated
endophytic fungi tolerance against the heavy metal copper and to evaluate for their
biosorption capacity.
The second aim is to compare the two groups of endophytic fungi isolated from Nepenthes
ampullaria plants collected from undisturbed and anthropogenically affected areas
(Mentawai Jungle and Kota Samarahan roadside, Kuching) on their heavy metal
resistance and biosorption capacity of removing heavy metal (Cu) from solution using
their Live and Dead biomass.
The third aim is to study and to understand the differentially expressed proteins of the
best fungal isolate (NA40; achieved the highest biosorption capacity using its live
biomass) in response to treatments with 3 different concentration of copper (0, 300, and
500 ppm).

- 27 -
The objectives of this study are:
Chapter 3
i. Isolation of endophytic fungi from Nepenthes ampullaria collected from
undisturbed and anthropogenically affected areas; Mentawai Jungle and Kota
Samarahan roadside, Kuching.
ii. Preliminary screening of the isolated fungal against their heavy metal copper
tolerance.
iii. Identification of the top 11 fungal isolates that were able to survive up to 1,000
ppm of heavy metal copper
iv. Evaluation of the chosen fungi (11) for their biosorption capacity on heavy metal
copper by using the Live and Dead biomass
Chapter 4
v. Proteomics analysis of the best fungal isolate (NA40; achieved the highest
biosorption capacity using its live biomass) on its differentially expressed proteins
in response to heavy metal copper.

- 28 -
Chapter 2
Methodology
2.1 Sampling Sites
Nepenthes ampullaria plant samples were collected at two different sites (see Table 2.1
for Global Positioning System (GPS) coordinates):
1. Ulu Mentawai (located at northern part of Gunung Mulu National Park)
2. Kota Samarahan road site, Kuching, Sarawak, Malaysia.
Table 2.1: GPS coordination for the Nepenthes ampullaria plant samples collected.
Nepenthes ampullaria Global Positioning System (GPS)
Mentawai Jungle N04o 14' 39.3'' E 114o 52' 04.0''
Mentawai Jungle N04 o 14' 39.4'' E 114o 52' 03.9''
Mentawai Jungle N04o 14' 39.0'' E 114o 52' 04.3''
Mentawai Jungle N04o 14' 39.1'' E 114o 52' 04.4''
Kuching Kota Samarahan Roadside 1.501992, 110.392635
Figure 2.1 and 2.2 show the location maps were the Nepenthes ampullaria plants were collected.
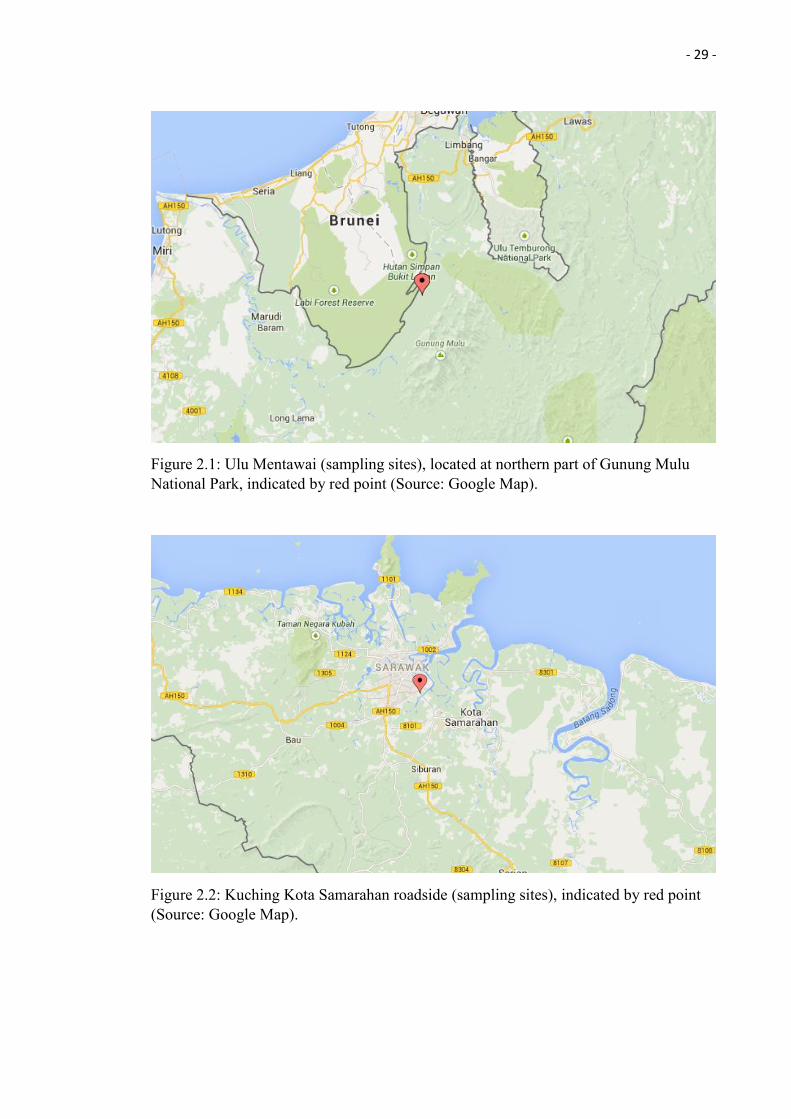
- 29 -
Figure 2.1: Ulu Mentawai (sampling sites), located at northern part of Gunung Mulu National Park, indicated by red point (Source: Google Map).
Figure 2.2: Kuching Kota Samarahan roadside (sampling sites), indicated by red point (Source: Google Map).

- 30 -
2.1.1 Ulu Mentawai
In the year of 2012, the Heart of Borneo Initiative, which is a NGO-supported and
government-led programme, organised a scientific expedition to Ulu Mentawai, located
at northern part of Gunung Mulu National Park (Figure 2.1). The area is well-known as
world heritage site that consist mainly of karangas forest and lowland mixed dipterocarp
forest.
The Nepenthes ampullaria plant samples (Figure 2.3) were collected (Figure 2.4) during
the expedition.
Figure 2.3: Nepenthes ampullaria, photographed on site (Mentawai jungle).

- 31 -
Figure 2.4: Author collecting plant samples collecting at in Mentawai jungle.
The Mentawai expedition was held from 5-17 September 2012, in total there are 5 plants
collected from the jungle: 3 Nepenthes ampullaria and 2 Nepenthes rafflesiana
(identification with the kind help of staff from Sarawak Forestry Department). However,
only the endophytic fungi isolated from the Nepenthes ampullaria were used in the
experiments.
2.1.2 Kota Samarahan Roadside
Another Nepenthes ampullaria collection was carried out at the main roadside of Kota
Samarahan (Figure 2.5), located at south east of Kuching. The plants were growing
besides the main road and very closely to the peat swamp. The plant sampling at Kota
Samarahan roadside was done on 2nd January 2013. Only 1 Nepenthes ampullaria plant
(identification done base on the plant and pitcher morphology; Lloyd 1942) was collected
for endophytic fungi isolation.

- 32 -
Figure 2.5: Kuching Kota Samarahan Roadside, the area where the Nepenthes ampullaria
was collected.
2.2 Nepenthes ampullaria Associated Endophytic Fungi Isolation
2.2.1 Ulu Mentawai
The endophytic fungi isolation for the plant samples collected during the scientific
expedition was conducted on site. The isolation of endophytic fungi was done on the same
day (less than 3 hours) as the plants were collected, right after return from the forest, to
ensure the freshness of the plant samples. Stone et al. (2011) suggested the importance of
the endophytic fungi isolation to be carried out as quickly as possible after the collection
of the sample plant, usually within 2 days’ time. The sampling site was remote, had no
electricity, and could only be accessed by small wooden long boats, thereby limiting the
availability of common laboratory equipment. To avoid contamination, the isolation was
carried out using a self-made lamina flow (Figure 2.6). The self-made lamina flow is
made of a plastic box with a plastic paper covered at the mouth of the box. Two small
hand size holes were made on the plastic cover. The inner and outer parts were sterilized

- 33 -
using 75% ethanol and all the autoclaved water (bottles) and autoclaved beakers were
surface wiped using the 75% ethanol before putting into the box. Additionally, control
plates were prepared and potential contaminants removed from the collection (see 2.2.3
for more details on the procedure).
Figure 2.6: A self-made plastic box - I was doing the endophytic fungi isolation at the
site.
2.2.2 Kota Samarahan Roadside
The plants were collected in the morning and stored at 4oC for 3 hours before carrying
out the endophytic fungi isolation. The isolation was carried out in a biosafety cabinet,
located at Swinburne University of Technology, Sarawak.
2.2.3 Endophytic Fungi Isolation
In order to isolate the endophytes, the plant samples have to be surface sterilized to kill
off all the microorganism that live on the surface of the plant such as epiphytic
microorganisms. A few different chemical solution were used for the surface sterilization
of plant samples such as ethanol (Petrini & Dreyfuss 1981), formaldehyde (Kreisel &

- 34 -
Schauer 1987), and sodium hypochlorite solution (Clark et al. 1983). Different surface
sterilization methods with different concentration of the chemical solution and different
surface sterilization timing will yield different endophytic fungi species (Schulz et al.
1993).
The collected plant samples were cut into small pieces (approximately 1cm3) using sterile
surgical blades and surface sterilized using 75% ethanol for 15-30 seconds. After that, the
plant tissue was dipped into autoclaved distilled water to stop the sterilization process and
surface dried by using autoclaved tissue paper. The plant tissue was then placed on Yeast
Extract Glucose Chloramphenicol Agar (YGCA) plates, which contains 1% of the
antibiotic chloramphenicol which will inhibits the growth of bacteria (Kohanski et al.
2010). A control plate was made using the autoclaved distilled water that used for plant
sample dipping. The plates were incubated at ambient temperature (Mentawai plant
samples) and 25oC (Kota Samarahan roadside plant samples). The isolation protocol is
modified from Strobel and Daisy (2003). Please refer to the Figure 2.7 for an overview
in form of a flow chart.

- 35 -
Figure 2.7: An overview in form of the isolation of endophytic fungi.
2.2.4 Endophytic Fungi Purification
After a week of the incubation, endophytic fungi were observed growing out from the
surface sterilized plant tissue, on the agar plates (Figure 2.8). The endophytic fungi were
then isolated out from the agar plate and placed on a fresh Potato Dextrose Agar (PDA)
plate by using autoclaved plastic straw. The isolates were sub-cultured until pure fungal
strains were obtained (Figure 2.9).
Plant samples
Cut into small pieces (Approx. 1 cm)
Surface sterilization with 75% ethanol for 15-30 seconds
Washed with autoclaved distilled water
Surface dried using autoclaved paper towel
Place it on yeast extract glucose chloramphenicol agar (YGCA) plates
Incubated at ambient temperature for mentaiwai plant samples and 25oc for kuching kota samarahan roadside
plant samples

- 36 -
Figure 2.8: Endophytic fungi were observed growing out from the surface sterilized plant
tissue.
Figure 2.9: Purified fungal strains.

- 37 -
2.2.5 Short Term Storage of the Isolated Fungi
The isolated fungi were grown on a PDA plate at 25oC for few days until the fungal
hyphae covered 2/3 of the plate. The fungal plates were then kept at 4oC until further use.
This way of fungal storage only last up to 6 months before the next sub-culturing into
new PDA plates (Nakasone et al. 2004). Please refer to the Figure 2.10 for more details.
Figure 2.10: A schematic view of short term storage of isolated fungi.
2.2.6 Long Term Storage of the Isolated Fungi
The isolated fungi was transferred into universal bottle that contains pure barley grains
(the barley was autoclaved for 3 times to ensure the barley is fully autoclaved- clean).
The fungi were grown in the media and stored at 4oC until all the barley in the bottle have
been covered by the fungi hyphae. This method was recorded to be able to store the fungal
up to 5 years in -20oC for Rhizoctonia solani with less than 1% loss in viability (Webb et
al. 2011). Please refer to the Figure 2.11 for more details.
Figure 2.11: A schematic view of long term storage of isolated fungi.
Autoclave barley grains in a universal bottle for 3 times
Transfer the fungal isolates onto the barley and incubate at 25oC until the fungal hyphae fully covered the barley
Store at 4oC for further use
Fungal isolated grown on Potato Dextrose Agar plate at 25CC until the fungal hyphae covered
2/3 of the plate
Keep the plate at 4oC for further use

- 38 -
2.3 Preliminary Screening of the Resistance Isolated Fungi Against the Heavy Metal Copper
The screening started with the roadside isolates (100 – 1,000 ppm), followed by the
Mentawai isolates (800 – 1,000 ppm). The testing was done using the direct transfer
method instead of adapting method during which the isolates were directly transferred
from a fungal plate (containing no heavy metal copper) to a new media that contains
different concentrations of copper.
The fungal isolates were grown on PDA plate for a week before been transferred to the
testing plates containing different copper concentration. Potato Dextrose Agar plates
containing 100 – 1,000 ppm copper (copper (II) sulfate salt) were prepared. An agar block
of the actively growing fungal hyphae from the PDA plates (containing no heavy metal
copper) was transferred onto the PDA plates that contain copper, by using sterilized
plastic straw. All plates were then incubated at 25oC for a week, all the results were
recorded and the fungal isolates that were able to grow on the PDA plates that contained
1,000 ppm of heavy metal copper concentration were chosen and utilised for copper
biosorption experiments. All the preliminary screening was undertaken in triplicates. The
protocol is modified from Iskandar et al. (2011). See Figure 2.12 for an overview of the
modified method.
A total of eleven (11) isolates were chosen for subsequent biosorption experiments (see
section 2.5) and identified using molecular methods (see below).

- 39 -
Figure 2.12: A schematic overview of preliminary screening of the resistance isolated fungi against the heavy metal copper.
2.4 Molecular Identification of the Chosen (11) Fungal Isolates
Traditional endophytic fungi identification is based on morphological characteristics
which heavily rely on the reproductive structure/ sporulation of the fungi. However, most
endophytic fungi do not produce the reproductive structure/ sporulation (Jones & Pang
2012). Besides that, morphological identification of the fungi requires an extensive
taxonomical knowledge (Gherbawy & Voigt 2010). Therefore, a molecular technique
based on the fungi rDNA sequence, the Internal Transcribed Spacer (ITS) region, is often
used for fungi identification (Arnold 2007). In this research, the chosen (11) fungi were
identified by using the molecular technique.
All the fungi were identified using the molecular technique. The fungal isolates were
cultured in a PDA plate for 3 days, and actively growing mycelia was transferred (using
a sterile toothpick) into 30 μl sterile lysis solution (10mM Tris-HCL, 1 mM EDTA, pH8.0;
Weising et al. 1994) in a 1.5 ml microcentrifuge tube. The tube was then kept in -80oC
overnight. A control tube (contains lysis solution without fungal mycelia) was prepared.
On the next day, the mixture was thawed at room temperature and 1µl of the supernatant
used for Polymerase Chain Reaction. The rest of the crude extract was stored at -20°C
The Fungal Isolates were cultured on Potato Destrose Agar Plate for a Week
Potato Dextrose Agar Plates that contains 100 – 1000 ppm of Heavy Metal Copper were Prepared
An Agar Block of the Actively Growing Fungal Hyphae was transferred onto the Heavy Metal Containing Agar plates
Incubate the Plates at 25oC for a week

- 40 -
until further usage. The fungal DNA extraction protocol is modified from Huhndorf et al.
(2004).
The universal fungal forward and reverse primers, ITS 4 {5’-
TCCTCCGCTTATTGATATGC-3’} and ITS5 {5’-
GGAAGTAAAAGTCGTAACAAGG-3’}, were used in the fungal DNA amplification.
Twenty two (22) μl of the pcr reaction master mix (BIOLINE) were transferred into a
sterile 0.3 ml PCR tube together with 1 μl each of forward and reverse primers and 1 μl
of the genomic DNA. A negative control (PCR mixture with 1 μl supernatant from the
control tube) was prepared.
The Polymerase Chain Reaction (PCR) consisted of an initial denaturing step of 5 minutes
at 94°C followed by 35 cycles (50 seconds at 94°C, 50 seconds at 54°C and 50 seconds
at 72°C), followed by a final extension step at 72°C for 10 minutes. The PCR products
were resolved by electrophoresis through 1% agarose gels in TAE and visualized by
staining with ethidium bromide for 10 minutes and distaining for 15 minutes. There is no
band observed from the control, which indicates the works is clean (Figure 2.13). The
PCR products were then purified and sent for sequencing to the Beijing Genome Institute
(BGI). The sequences obtained were analysed using the National Center for
Biotechnology Information (NCBI - USA) database and a phylogenetic tree was
constructed from genetic distance and bootstrap values calculated using MEGA 6
(Tamura et al. 2013). Please refer to the Figure 2.14 for an overview in form of a flowchart.
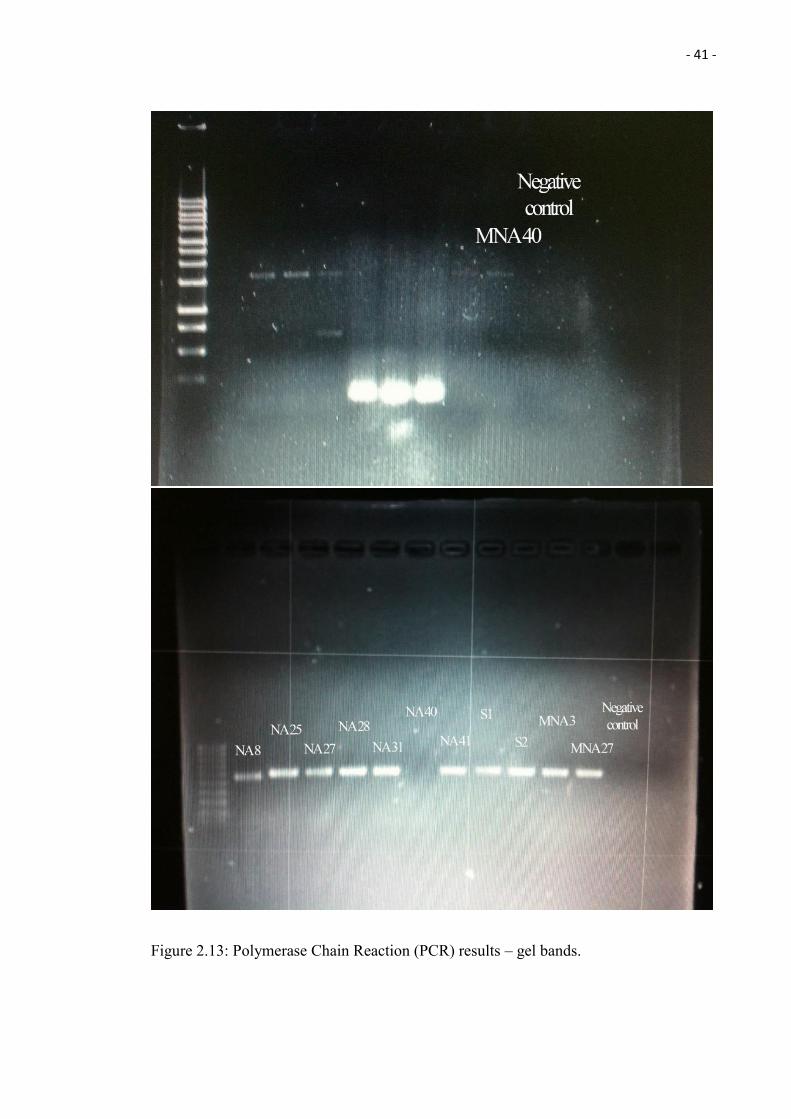
- 41 -
Figure 2.13: Polymerase Chain Reaction (PCR) results – gel bands.
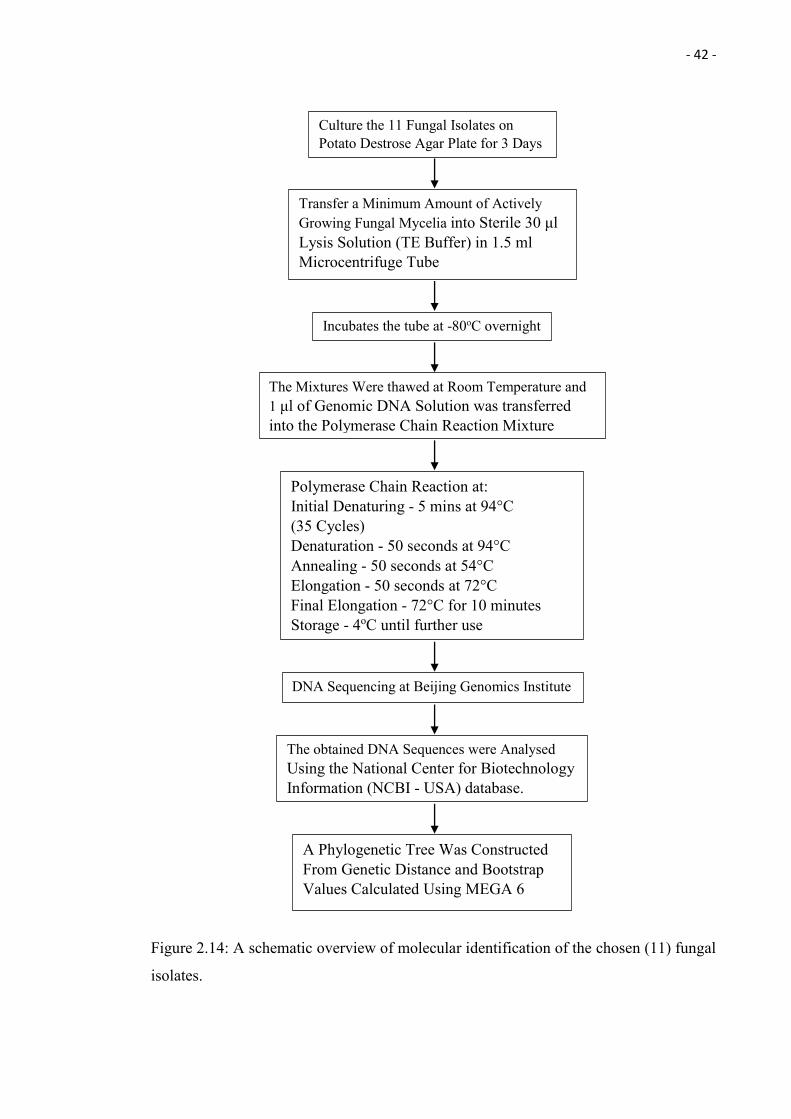
- 42 -
Figure 2.14: A schematic overview of molecular identification of the chosen (11) fungal
isolates.
Culture the 11 Fungal Isolates on Potato Destrose Agar Plate for 3 Days
Incubates the tube at -80oC overnight
Transfer a Minimum Amount of Actively Growing Fungal Mycelia into Sterile 30 μl Lysis Solution (TE Buffer) in 1.5 ml Microcentrifuge Tube
The Mixtures Were thawed at Room Temperature and 1 μl of Genomic DNA Solution was transferred into the Polymerase Chain Reaction Mixture
DNA Sequencing at Beijing Genomics Institute
Polymerase Chain Reaction at: Initial Denaturing - 5 mins at 94°C (35 Cycles) Denaturation - 50 seconds at 94°C Annealing - 50 seconds at 54°C Elongation - 50 seconds at 72°C Final Elongation - 72°C for 10 minutes Storage - 4oC until further use
A Phylogenetic Tree Was Constructed From Genetic Distance and Bootstrap Values Calculated Using MEGA 6
The obtained DNA Sequences were Analysed Using the National Center for Biotechnology Information (NCBI - USA) database.

- 43 -
2.5. Evaluation of Biosorption Capacity of the Chosen Fungal Isolates
2.5.1 Heavy Metal Copper Biosorption by Live Fungal Biomass
Potato Dextrose Broth (PDB) supplied with 500 ppm copper (copper (II) sulfate salt) was
prepared. Fungal isolates were grown on PDA plates (containing no heavy metal copper)
for a week before transferal into Potato Dextrose Broth (PDB) containing 500 ppm copper.
Three cylindrical agar plugs of the actively growing fungal hyphae from the PDA plates
were transferred into the PDB containing 500 ppm copper using sterilized plastic straws.
The mixtures were then incubated at 25oC, under static condition for 2 months.
After the 2 months of incubation (Figure 2.15), the fungal biomass were filtered
usingfilter paper (Whatman A1) and dried at 70oC. The weight of the dried fungal biomass
were measured and recorded. The final concentration of heavy metal was measured using
Atomic Absorption Spectrometer (AAS; Xplor AA (Serial No. A6945)). Please refer to
the Figure 2.16 for more details.
The biosorption capability of 1 gram living fungal biomass was calculated using the
following formula (Zafar et al. 2007):
Q [mg/g] = (Ci – Cf [mg/L] / M [g]) V [L]
where Q is mg of metal ion absorbed per gram of fungal biomass [mg/g], Ci and Cf are
the initial and final concentrations of the metal in the solution [mg/L]. M is the amount
of the added (bio)sorbent to the reaction mixture [g] and V is the volume reaction mixture
[L].

- 44 -
Figure 2.15: Fungal isolates were growing in the potato dextrose broth supplied with 500ppm of copper.
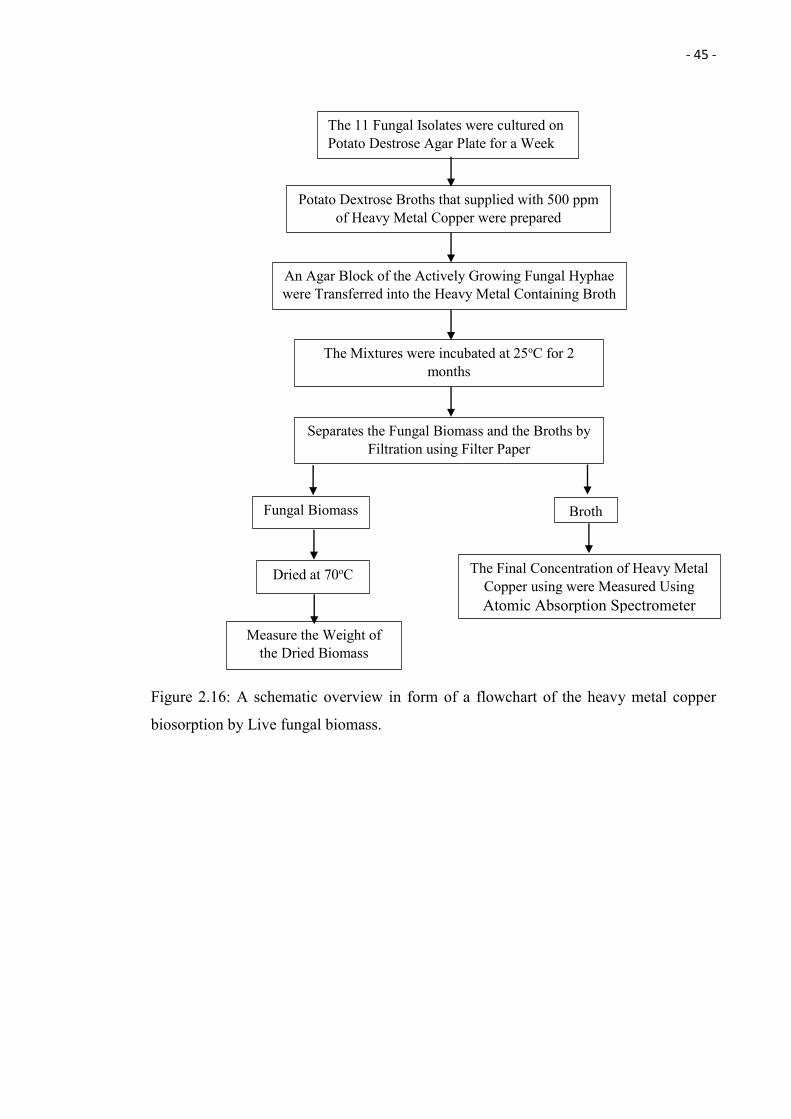
- 45 -
Figure 2.16: A schematic overview in form of a flowchart of the heavy metal copper
biosorption by Live fungal biomass.
The 11 Fungal Isolates were cultured on Potato Destrose Agar Plate for a Week
Potato Dextrose Broths that supplied with 500 ppm of Heavy Metal Copper were prepared
An Agar Block of the Actively Growing Fungal Hyphae were Transferred into the Heavy Metal Containing Broth
plates
Separates the Fungal Biomass and the Broths by Filtration using Filter Paper
Fungal Biomass
Dried at 70oC
Measure the Weight of the Dried Biomass
Broth
The Final Concentration of Heavy Metal Copper using were Measured Using Atomic Absorption Spectrometer
The Mixtures were incubated at 25oC for 2 months

- 46 -
2.5.2 Heavy Metal Copper Biosorption by Dead Fungal Biomass
A single cylindrical agar plug of 5 day old fungal cultures was inoculated into 200 ml of
potato dextrose broth (PDB) and incubated for 2 months at 25oC, under static conditions.
After 2 months of incubation, the fungal biomass was filtered using filter paper, dried and
killed at 70oC. The dried biomass was then ground into powder by using pestle and mortar
and the powdered fungal biomass stored in 1.5 ml centrifuge tubes for further use.
Autoclaved distilled water supplied with 500 ppm copper (Copper(II) sulfate salt) was
prepared. The powdered fungal biomass was added into 10 ml of autoclaved distilled
water containing 500 ppm copper, and incubated for 2 months at 25oC. After 2 months of
incubation, the dead fungal biomass was filtered out and the final concentration of the
heavy metal copper was measured using Atomic Absorption Spectrometer (AAS; Xplor
AA (Serial No. A6945)). The protocol is modified from Martínez-Juárez et al. (2012).
Please refer to the Figure 2.17 for an overview in form of a flowchart.
The biosorption capability of 1 gram dead fungal biomass was again calculated as follows
(Zafar et al. 2007):
Q [mg/g] = (Ci – Cf [mg/L] / M [g] ) V [L]
where Q is mg of metal ion absorbed per gram of fungal biomass [mg/g], Ci and Cf are
the initial and final concentrations of the metal in the solution [mg/L]. M is the amount
of the added (bio)sorbent to the reaction mixture [g] and V is the volume reaction mixture
[L].
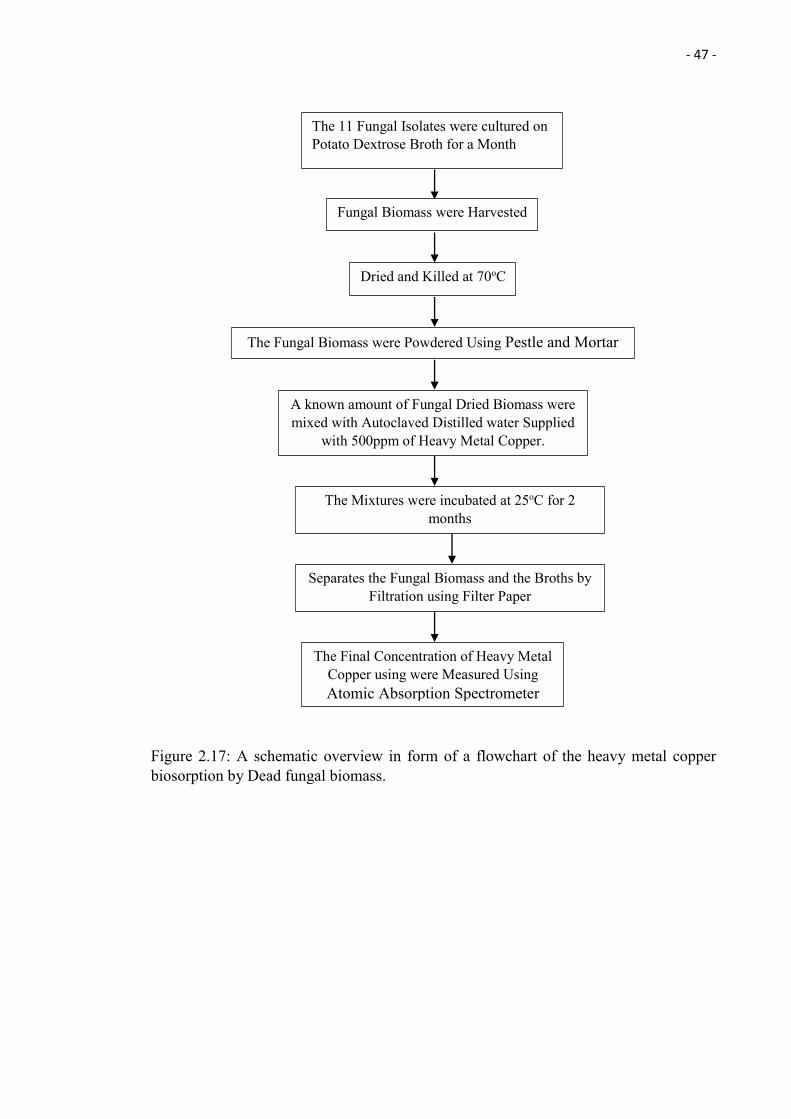
- 47 -
Figure 2.17: A schematic overview in form of a flowchart of the heavy metal copper biosorption by Dead fungal biomass.
The 11 Fungal Isolates were cultured on Potato Dextrose Broth for a Month
Separates the Fungal Biomass and the Broths by Filtration using Filter Paper
The Final Concentration of Heavy Metal Copper using were Measured Using Atomic Absorption Spectrometer
The Mixtures were incubated at 25oC for 2 months
Fungal Biomass were Harvested
Dried and Killed at 70oC
A known amount of Fungal Dried Biomass were mixed with Autoclaved Distilled water Supplied
with 500ppm of Heavy Metal Copper.
The Fungal Biomass were Powdered Using Pestle and Mortar

- 48 -
2.6 Proteomic Analysis of the Best Fungal Strain (NA40) on Heavy Metal Copper
Fungi are known to produce or overexpress certain enzymes in response to heavy metal
induced oxidative stress, and proteomics provides a way for the stress response to be
studied (Washburn and Yates III 2000; Rabilloud et al. 2005).
Fungal isolate NA40 achieved the best biosorption capacity (live biomass) of heavy metal,
thus it was chosen to perform this analysis. In this study (chapter 4), the isolates NA40
was cultivated in 3 different conditions – potato dextrose broth (PDB) without copper,
PBD with 300 ppm and 500 ppm copper concentration. The chosen fungal isolate NA40
was inoculated into the prepared PDB solutions and incubated at 25oC for 3 weeks, under
static condition. After 3 weeks of incubation, the fungal biomass was stored at 4oC before
it was brought to Agrobiotechnology Institute Malaysia (ABI) to perform the protein
extraction and analysis.
Please refer to the Figure 2.18 for an overview in form of a flowchart and to chapter 4 for
more details.
Figure 2.18: An overview in form of a flowchart of the fungal incubation in PDB with three different concentration of heavy metal copper concentration.
The Mixture were incubated for 3 weeks at 25oC under static condition
The Fungal Biomass were stored at 4oC before bringing to Agrobiotechnology Institute for
Protein Analysis
The Fungal Isolate (NA40) were Cultured in Potato Dextrose Broth supplied with 0, 300 and
500 ppm of Heavy Metal Copper.

- 49 -
2.6.1 Fungal Proteome Preparation
The fungal biomass was harvested by spinning down using a temperature controlled
centrifuge at 10,000 g at 4oC, for 10 minutes. After that the supernatants were discarded
and the fungal biomass rinsed with deionised water and again span down using a
temperature controlled centrifuge at 10,000 g at 4oC, for 10 minutes. This rinsing step
was repeated for 2 times. The fungal biomass was grounded into fine powder using mortar
and pestle in the presence of liquid nitrogen. TCA-acetone extraction was performed by
mixing each of the 1g of the powdered fungal biomass with 1.8 ml of 10% trichloroacetic
acid in cold acetone containing 0.07% β-mercaptoethanol and vortex at the temperature
of 4oC. After that, the mixture was incubated in 20oC overnight. On the next day, the
mixture was centrifuged at 10,000 g at 4oC for 15 minutes. The supernatant was discarded
and the pellet was re-suspended in rising solution (each of the 1 g with 1.8 ml of rinsing
solution) that contains 0.07% β-mercaptoethanol in cold acetone, which was then
incubated at -20°C for 1 h (mixed every 15 min intervals) and re-centrifuged at 10,000g
at 4oC for 15 minutes. The supernatant was discarded and this rising steps was repeated
for 2 times. The pellet was then vacuum-dried and re-suspended with lysis buffer and
stored at -80oC. The protocol is modified base on the paper written by Pavoković et al.
(2012). Please refer to the Figure 2.19 for more details.

- 50 -
Figure 2.19: An overview in form of a flowchart of the fungal proteome preparation.
Vortexed the Mixtures at 4oC for 10 minutes
One (1) gram of the Powdered Fungal Biomass was Transferred into 1.5ml Microcentrifuge Tube
The Fungal Biomass were Transferred into clean 50 ml falcon tube
The supernatant were discarded
The amount of 1.8 ml of of 10% Trichloroacetic Acid in Cold Acetone Containing 0.07% β-mercaptoethanol was added to each of the tubes
Powered the Fungal Biomass by Using Mortar and Pestle with Liquid Nitrogen
Incubated at 20oC Overnight
Spin down the Fungal Biomass at 10,000g at 4oC
Centrifuged at 10000 g at 4oC for 15 minutes
Twenty Five (25) ml of Deionised Water were added into the tubes
Spin down the Fungal Biomass using using Temperature controlled Centrifuge at 10,000g at 4oC
The Supernatant were Discarded
The Supernatant were Discarded
Resuspened with Lysis Buffer
The pellets were vacuum-dried
The pellet was Resuspended in Rinsing Solution
Incubated at 4oC for 1 hour (mixed every 15 min interval)
Centrifuged at 10,000g at 4oC for 15 minutes
The Supernatant were Discarded
Stored at -80oC for Further Usage
Rep
eate
d Tw
ice
Rep
eate
d Tw
ice

- 51 -
2.6.2 Total Protein Measurement by Bradford Assay
Bradford Assay, introduced by Bradford in the year of 1976, is a protein determination
method that has been used widely for determination of protein concentration. All the
proteins extracted from fungal biomass, collected from the three different solutions, were
analysed for their total protein concentration using the Bradford Assay kit (Biorad
Bradford Reagent assay). The total concentrations of the extracted proteins were
measured and recorded.
2.6.3 Two-dimensional Gel Electrophoresis (2-DE)
Two-dimensional gel electrophoresis (2-DE) is a gel-based proteomics technique that
have been widely used for the separation, detection and analysis of proteome from
complex biological sources which was 1st introduced by O'Farrell (1975). By using this
technique, proteins are separated based on the different properties. During the 1st
dimension, protein will be separated base on their different isoelectic point in the gel
matrix by Isoelectric focusing (IEF). The separated protein will then be re-separated again
in second dimension, based on their different molecular weight, in Polyacrylamide gel by
sodium dodecyl sulfate Polyacrylamide gel electrophoresis (SDS-PAGE).
2D gels of the control and each treatment were run in triplicates. Isoelectric focusing (IEF)
was performed using 13 cm Nonlinear IPG-strips (pH range 3-10). The IPG-strips were
initially rehydrated for 12 hours in the presence of 70 μg of protein. IEF was performed
using Biorad Protean i12 with standard protocol based on Biorad Handbook (IEF
Protocol), at 20 °C in a stepwise manner: 500 V (2 h), 1.0 kV (1 h), 8.0 kV (1 h), 8.0 kV
(28000 VhS) and finally 750 V (hold). Please refer to Figure 2.20 for more details. The
strips were equilibrated in equilibration buffer (based on GE Healthcare 2D SDS PAGE
Handbook) containing 50 mM Tris–HCl pH 8.8, 6 M urea, 30% (v/v) glycerol, 2% (w/v)
SDS, 0.002% (w/v) bromophenol blue and 1% (w/v) dithiothreitol (DTT) for 15 minutes,
followed by equilibrated in the same equilibration buffer containing 2.5% (w/v)
iodoacetamide instead of DTT for another 15 minutes. The second dimension separation
was performed in 12% polyacrylamide gels, at 20oC, using SE 600 Ruby system (Hoefer
SE 600 Ruby (Amersham Biosciences)), with the running buffer contains 25 mM Tris–
HCl, 192 mM glycine, 0.1% (w/v) SDS, at 10mA/gel (15min) and 20mA/ gel (3h 30min).

- 52 -
The gels were stained with silver staining (Shevchenko et al., 1996; see section 2.6.4).
Please refer to Figure 2.21 for more details.
Figure 2.20: An overview in form of a flowchart of Isoelectric focusing.
The mixture was pipetted into the rehydration tray as a streak slightly shorter than the strip to be rehydrated (Bubble formation was prevented)
The IPG-strip was allowed to rehydrated for 12 hours and IEF for 4-5 hours
Rehydration and IEF parameter were set
IPG-strip was placed into a slot with the dried gel side down
Protective film was removed from the IPG-stips from the acidic (+) end
Approx. 1 ml of cover fluid (mineral oil) were overlaid onto the strip
The Protein Samples was mixed with rehydration buffer to make the total volume of 200 μl with 70 μg protein concentration

- 53 -
Figure 2.21: An overview in form of a flowchart of two-dimensional gel electrophoresis (2-DE).
Equilibrated the strips with SDS equilibration 1 solution (DTT) for 15 minutes
Rinsed the IPG-strips with running buffer
Poured away the SDS equilibration 2 solution
Placed the IPG-strips into the strip holders
Rinsed the IPG-strips with running buffer
Poured away the SDS equilibration 1 solution
Equilibrated the strips with SDS equilibration 2 solution (IAA) for 15 minutes
IPG-strips removed from the slots
Inserted the strips into the gel
Inserted the protein marker
Overlay the strips with agarose sealing solution
Transferred the glass plate into the tank
The gels were allowed to run for 3 hours 45 minutes with the set parameter

- 54 -
2.6.4 Silver staining for SDS-PAGE
The silver staining protocol was modified from Shevchenko et al., (1996). The gel was
fixed with fixation solution for 30 minutes, followed by 30 minutes in sensitizing solution.
After that the gel was rinsed with Millipore water for 5 mins (3 times). After rinsing, the
gel was submerged in silver nitrate solution for 20 minutes. After the incubation, the silver
nitrate was discarded and staining solution was added onto the gel. The gel was incubated
in the staining solution until the protein spots were started to appear (5-10 minutes). The
staining solution was then discarded and stopping solution was added onto the gel to stop
the staining process. The gel was leaved in the stopping solution for 10 minutes and rinsed
with Millipore water for 3 times. After rinsing the gel was stored in conserving solution
until the further use. Please refer to Figure 2.22 for more details and Figure 2.23 for the
gel image after silver staining.
Figure 2.22: An overview in form of a flowchart of silver staining for SDS-PAGE.
Fixed the gel with the fixation solution for 30 minutes
Rinsed the gel using Millipore water for 5 minutes
Rinsed with Millipore water for 10 minutes
Sensitized the gel with the sensitizing solution for 30 minutes
Submerged in silver nitrate solution for 20 minutes
Stained the gel using staining solution unti the protein spots were started to appear (5-10 minutes)
Stopped the staining using stopping solution (incubated for 10 minutes)
Rep
eate
d Th
ree
times
Conserved the gel in conserving solution until further use

- 55 -
Figure 2.23: Silver stained gel image taken using Cannon digital camera, at the bench of
the lab.

- 56 -
2.6.5 Protein Identification and Database Search
Stained gels were digitized by using image Scanner (GS800 Desitometer (Biorad)) and
the protein spots analysis were performed using Progenesis Samespots samespot software,
with the Max fold change ≥ 2, and Anova p-value ≤ 0.05. Figure 2.24, 2.25 and 2.26
show the images scanned using the scanner, with blue spots indicates the protein spots of
interest. The chosen proteins spots were then manually excised from the stained 2D gels
and destained followed by in gel digestion using trypsin overnight at 25oC (Shevchenko
et al. 2007). The peptides were extracted from the gel pieces by using 50% of acetonitrile
and 100% acetonitrile for the second time extraction. The solution is then vaccum dried
and stored for further identification. Please refer to Figure 2.27 for more details.
Protein identification were done by Norasfaliza Rahmad which was accomplished by
mass spectrometry. Peptide Mass Fingerprinting (PMF) data search was performed using
Swiss-Prot database. The obtained protein ID is then further analysed and studied.

- 57 -
Figure 2.24: Protein spots produced by the fungal isolate NA40 in the PDB with no heavy
metal copper, gel image was taken using image Scanner (GS800 Desitometer (Biorad)).

- 58 -
Figure 2.25: Protein spots produced by the fungal isolate NA40 in the PDB supplied with
300ppm of heavy metal copper, gel image was taken using image Scanner (GS800
Desitometer (Biorad)).
Figure 2.26: Protein spots produced by the fungal isolate NA40 in the PDB supplied with
500ppm of heavy metal copper, gel image was taken using image Scanner (GS800
Desitometer (Biorad)).

- 59 -
Figure 2.27: An overview in form of the in-gel digestion of the protein spots.
Cut the gel spot into small pieces (1-2 mm)
A volume of 50 μl of 100% ACN was added into the tube
A volume of 150 μl of 100 mM NA4(HCO3) was added into the tube
Washed the gel with solution (50% ACN in 100 mM of NA4(HCO3)) for 20 minutes
Removed the solution
Incubated the tube for 20 minutes in the dark
Placed the gel pieces into 1.5 ml microcentrifuge tube
Washed the gel for 10 minutes
Removed the NA4(HCO3)
Alkylated the protein by adding 150 μl of 55 mM IAA in 100 mM of NA4(HCO3)
Incubated the tube for 15 minutes at room temperature
The gel was dried by speed vacuum for 15 minutes
A volume of 25 μl of 7 ng/μl trypsin solution was added into the tube
A volume of 50 μl of 100% ACN was added into the tube (second extraction)
Collected the solution into a new 1.5 ml microcentrifuge tube
Incubated the tube in the waterbath at 25oC overnight
Removed the tube from waterbath
A volume of 25 μl of 50% ACN was added into the tube
Incubated the tube for 15 minutes
Collected the solution into the 1.5 ml microcentrifuge tube
Stored at -80oC before proceed to protein identification
Dried the solution by speed vacuum
Rep
eate
d Th
ree
times
R
epea
ted
Twic
e

- 60 -
Chapter 3
Biosorption of Copper (Cu) by Endophytic
Fungi Isolated from Nepenthes ampullaria
Changi Wong1*, Daniel Tan1, Samuel Lihan2, Aazani Mujahid2, and Moritz Müller1
1 Faculty of Engineering, Computing and Science, Swinburne University of Technology
Sarawak, 93350 Kuching, Malaysia.
2 Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 93400
Kota Samarahan, Sarawak, Malaysia.
*Corresponding author
Email: [email protected]
Phone number: +60168716911
ABSTRACT
Biosorption using biological materials is one of the ways to deal with heavy metal
pollution. In this study, endophytic fungi were (a) isolated from the carnivorous plant
Nepenthes ampullaria (collected from undisturbed and anthropogenically affected areas;
Mentawai Jungle and Kota Samarahan roadside, Kuching); (b) assessed for their
resistance against the heavy metal copper; and (c) evaluated for their biosorption capacity
(live and dead biomass). In total, 147 fungal isolates were isolated from Nepenthes
ampullaria and only 7.5% (11) of the total isolates were capable to resist copper
concentration up to 1000 ppm. The 11 fungal isolates were identified through molecular
method, and all of them were grouped with members of the Phomopsis, Diaporthe,
Nigrospora, and Xylaria. The fungal isolate NA40 that was related to Xylaria sp. achieved
the highest biosorption capacity of of 73.26 mg/g using live biomass, while the isolates
NA41 that was related to Phomopsis sp. had the highest biosorption capacity of 59.33
mg/g using dead biomass.
Keywords: Endophytic fungi, Biosorption, Heavy metal copper, Nepenthes ampullaria

- 61 -
3.1 Introduction
Nepenthes, also known as pitcher plant or monkey cup, is a carnivorous plant that consists
of a tube shaped cup at the end of the tendril filled with digestive fluid to trap and digest
other living organisms and to absorb their nutrients (Hua & Li 2005). Nepenthes (N.) are
widely distributed in Borneo, Peninsular Malaysia and Sumatra, such as N. gracilis, N.
mirabilis, and N. ampullaria and several species (N. rajah, N. xalisaputrana, N. villosa)
have been recorded living in soils low in macronutrients and containing toxic heavy
metals such as chromium (Cr), magnesium (Mg) and nickel (Ni) (Adlassnig et al. 2005).
However, our understanding of the strategy used by the plants to survive in heavy metal
contaminated soils is still very limited. Soleimani et al. (2010) studied the effect of
Neotyphodium, an endophytic fungus, on cadmium (Cd) tolerance of two grass species
(Festuca arundinacea and Festuca pratensis) and showed that endophyte-infected plants
had a higher Cd tolerance compared to non-infected plants. Ren et al. (2011) also
demonstrated that endophytic fungus-infected tall fescue (Lolium arundinaceum) had a
higher Cd tolerance compared to the non-infected one. It seems likely that the presence
of the endophytic fungi in the plants – the symbiosis between endophyte and plant – could
be the reason behind the observed tolerance of the carnivorous plants towards the heavy
metal.
Copper is one of the essential micronutrients for human as it helps to transmit electrical
signals and facilitates the absorption of iron in our body (Ashish et al. 2013). However,
it can be extremely toxic in high concentration. Symptoms such as abdomen pain, purging
and vomiting will be observed when taken in excess (Ashish et al. 2013). Copper (Cu)
can be found naturally occurring in the environment. It was also found to be released into
the environment by human activities such as phosphate fertilizer production, metal and
wood production, agriculture and mining activity. Copper does not break down easily in
the environment, thus causing the environmental pollution such as acid mine drainage
(AMD). The leaching of metal (Cu is one of the metals present) by AMD at mining area
into the water and sentiment heavily affects the environment, rendering the heavily
polluted areas unsustainable to life (ATSDR 2004; McCarthy 2011; Johnson & Hallberg
2005). In Malaysia, few copper pollutions have been reported such as the copper pollution
of the Juru River (located in Penang) that was caused by pig farm discharges and wastes,
with Cu concentrations up to 144 µg g−1 (Lim & Kiu 1995), which is 2 times higher in

- 62 -
comparison to the natural average global shale values (shale value of 45µg g−1) (Mason
& Moore, 1982). Besides that, high Cu concentrations (37 - 100 µg/g) have also been
recorded in coastal areas of Peninsular Malaysia (Lukut River), with up to 60% of the
copper the results of previous anthropogenic activities (Ismail & Safahieh 2005).
In recent years, several procedures have been designed to deal with heavy metal
contamination of rivers and streams, for example chemical precipitation,
phytoremediation (Vaajasaari & Joutti 2006; Wenzel 2009), ion exchange, reverse
osmosis, solvent extraction (Iyer 1990) and biosorption (Fourest & Roux, 1992).
Biosorption uses biological materials such as bacteria, algae, yeast and fungi (Volesky
1986) to accumulate heavy metals from wastewater through physico-chemical or
metabolically mediated pathways of uptake (Fourest & Roux 1992). There are several
advantages compared to other approaches such as high efficiency, cost effectiveness, the
possibility of recovering the metal of interest and regeneration of the biosorbent
(Kratochvil & Volesky 1998). Recent studies have shown the capability of using
endophytic fungi as biosorbent to bioabsorb or to remove heavy metal. The endophytic
Microsphaeropsis sp. LSE10 isolated from Solanum nigrum L. plant is capable to biosorb
heavy metal cadmium (Xiao et al. 2010), and another endophytic Mucor sp. CBRF59
isolated from Brassica chinensis plant collected from metal-contaminated soil is able to
biosorb heavy metal cadmium and lead by using its live and dead biomass (Deng et al.
2011).
In this study, endophytic fungi were (a) isolated from of the carnivorous plant Nepenthes
ampullaria (collected from undisturbed and anthropogenically affected areas; Mentawai
Jungle and Kota Samarahan roadside, Kuching); (b) assessed for their resistance against
the heavy metal copper; and (c) their biosorption capacity (live and dead biomass)
evaluated.

- 63 -
3.2 Methodology
3.2.1 Endophyte Isolation and Purification
Nepenthes ampullaria were collected in the Mentawai Jungle (Miri, Sarawak, Malaysia)
during the Heart of Borneo Initiative; Mentawai expedition 2013, and from roadsides at
Kota Samarahan (Kuching, Sarawak, Malaysia). Isolation of endophytic fungi followed
the modified procedures from Strobel and Daisy (2003). In summary, different parts of
the plants (leaves, pitcher, and roots) were cut into small pieces of 1 cm2 in size and
surface-sterilized by immersion in 70% ethanol for 5 - 15 seconds. After that, the sample
was immersed in sterile distilled water (twice) to stop the sterilization. The sample was
then dried using a sterile cotton cloth and placed on a Yeast Extract Glucose
Chloramphenicol Agar (YGCA) plate, which contains chloramphenicol to suppress the
growth of bacteria. One millilitre of the sterile distilled water that was used to clean the
sample was taken out and poured on another YCGA plate as negative control. The agar
dish was then sealed with parafilm, labelled and incubated at 25°C for 7 days. The growth
of hyphae was observed after 7 days and all of the hyphae were isolated using sterile
plastic straws and placed on fresh YCGA dishes. This step was repeated until pure fungal
colonies were obtained. Purified fungi were cultured on Potato Dextrose Agar (PDA) and
incubated at 25°C.
3.2.2 Preliminary Screening of Heavy Metal Copper Tolerance Fungi
Preliminary screening followed procedures modified from Iskandar et al. (2011). In
summary, a single cylindrical block (agar plug) of 5 day old fungal cultures were placed
on Potato Dextrose Agar plates supplemented with Cu (100 – 1000 ppm), and incubated
at 25oC for 7 days. Growth of the fungal isolates was observed and recorded after the 3rd
and 7th day.
3.2.3 Molecular Identification
Identification of endophytic fungi followed the procedures modified from Huhndorf et al.
(2004). In summary, a small amount of 3 days old mycelia was transferred into sterile 30
μl lysis solution (TE buffer – 10 mM Tris-HCL, 1 mM EDTA, at pH 8) in 1.5 ml
microcentrifuge tubes using a sterile toothpick, and incubated at -80°C overnight. On the

- 64 -
next day, the mixture was thawed at room temperature and 1 µl of the supernatant used
for Polymerase Chain Reaction. The rest of the crude extract was stored at -20°C until
further usage.
Twenty two (22) μl of the master mix (BIOLINE) were transferred into a sterile 0.3 ml
PCR tube together with 1 μl of the each forward and reversed primers (ITS4 {5’-
TCCTCCGCTTATTGATATGC-3’} and ITS5 {5’-
GGAAGTAAAAGTCGTAACAAGG-3’}), and 1 μl of the genomic DNA. The mixture
was then used for polymerase chain reaction.
The Polymerase Chain Reaction (PCR) consisted of an initial denaturing step of 5 minutes
at 94°C followed by 35 cycles (XY seconds at 94°C, 50 seconds at 54°C and 50 seconds
at 72°C), followed by a final extension step at 72°C for 10 minutes. The PCR products
were resolved by electrophoresis through 1% agarose gels in TAE and visualized by
staining with ethidium bromide for 10 minutes and destaining for 15 minutes. The PCR
products were then purified and sent for sequencing. The sequences obtained were
analyzed against the NCBI (USA) database (Zhang et al. 2000) and a phylogenetic tree
was constructed from genetic distance and bootstrap values calculated using MEGA 6
(see Figure 3.1; Tamura et al. 2013).
3.2.4 Biosorption of Copper by Living Fungal Biomass
Biosorption capacity was calculated following procedures outlined by (Zafar et al. 2007).
In summary, three single cylindrical blocks (agar plugs) of 5 day old fungal cultures were
inoculated into 80 ml of potato dextrose broth (PDB), with 500 ppm of heavy metal
copper added, and incubated for 2 months at 25oC, under static conditions.
After 2 months of incubation, the fungal biomass was filtered and dried at 70oC. The
weight of the dried fungal biomass were measured and recorded. The final concentration
of heavy metal was measured using Atomic Absorption Spectrometer (AAS; Xplor AA

- 65 -
(Serial No. A6945)). The biosorption capability of 1 gram living fungal biomass was
calculated using the formula:
Q [mg/g] = (Ci – Cf [mg/L] / M [g]) V [L]
where Q is mg of metal ion absorbed per gram of fungal biomass [mg/g], Ci and Cf are
the initial and final concentrations of the metal in the solution [mg/L]. M is the amount
of the added (bio)sorbent to the reaction mixture [g] and V is the volume reaction mixture
[L].
3.2.5 Biosorption of Copper by Dead Fungal Biomass
Biosorption capacity was calculated following procedures outlined by (Zafar et al. 2007).
A single cylindrical block (agar plug) of 5 days old fungal cultures was inoculated into
200 ml of potato dextrose broth (PDB) and incubated for 2 months at 25oC, under static
conditions. After 2 months of incubation, the fungal biomass was filtered, dried and killed
at 70oC. The dried biomass were then grinded into powder by using pestle and mortar.
The powered fungal biomass were stored into 1.5ml of centrifuge tube for further use.
An small amount of dried fungal biomass were pre-weighted and recorded, and added
into 10 ml of distilled water, with 500 ppm of heavy metal copper added, and incubated
for 2 months at 25oC.
After 2 months of incubation, the dead fungal biomass was filtered out and Cu
concentrations measured using Atomic Absorption Spectrometer (machine model). The
biosorption capability of 1 gram dead fungal biomass was calculated as follows:
Q [mg/g] = (Ci – Cf [mg/L] / M [g]) V [L]
where Q is mg of metal ion absorbed per gram of fungal biomass [mg/g], Ci and Cf are
the initial and final concentrations of the metal in the solution [mg/L]. M is the amount
of the added (bio)sorbent to the reaction mixture [g] and V is the volume reaction mixture
[L].

- 66 -
3.3 Results and Discussion
Endophyte Isolation and Purification
A Total of 147 fungal isolates were isolated from Nepenthes ampullaria. Ninety two (92)
isolates were from plants collected in Mentawai Jungle, while the other fifty five (55)
isolates were from plants collected at the roadside of Kota Samarahan, Kuching, Sarawak,
Malaysia.
Preliminary Screening of Copper Tolerance among Fungi
Of the 147 fungal isolates, Ninety two (92) isolates were from plants collected in
Mentawai Jungle, while the other fifty five (55) isolates were from plants collected at the
roadside of Kota Samarahan, Kuching, Sarawak, Malaysia. Only 11 managed to survive
in copper concentrations up to 1000 ppm (Table 3.1). Nine out of these 11 isolates (NA8,
NA25, NA27, NA28, NA31, NA40, NA41, S1 and S2) were isolated from a plant
collected from the roadside of Kota Samarahan, while only 2 isolates from Mentawai
Jungle (MNA3 and MNA27) were able to survive at 1000 ppm Cu. These 11 isolates
were chosen to carry out the Heavy Metal (copper) Biosorption Assay by using the living
and dead fungal biomass.
The results clearly showed that fungal isolates isolated from roadside plants display much
higher resistance towards Cu. This can be explained by increased exposure to Cu along
roadsides (and necessary adaptation to survive), as compared to the undisturbed jungle
environment. Zehetner et al. (2009) showed that roadside environments are often
contaminated by automobile traffic with a wide range of contaminants such as heavy
metals, which can be found in the wall of fuel tanks, engines, tires, brake pads and road
surface materials. Han et al. (2014) conducted a research on heavy metal concentrations
in road dust in Kuala Lumpur, and Cu ranked 2nd in concentration among the heavy metal
contaminants. These studies support our findings and indicate that the Nepenthes and their
endophytic fungi along the roadsides of Kota Samarahan have been exposed to heavy
metals released by the automobile traffic and have therefore developed stronger resistance
towards the metals.
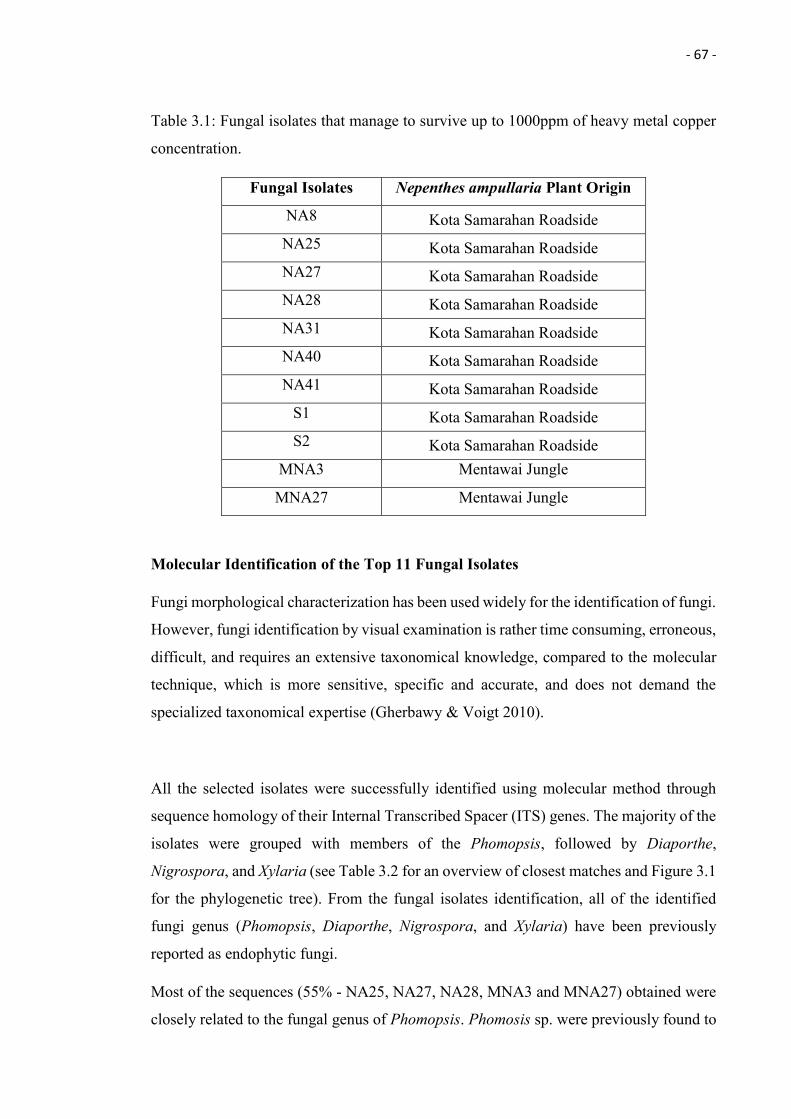
- 67 -
Table 3.1: Fungal isolates that manage to survive up to 1000ppm of heavy metal copper
concentration.
Fungal Isolates Nepenthes ampullaria Plant Origin
NA8 Kota Samarahan Roadside NA25 Kota Samarahan Roadside NA27 Kota Samarahan Roadside NA28 Kota Samarahan Roadside NA31 Kota Samarahan Roadside NA40 Kota Samarahan Roadside NA41 Kota Samarahan Roadside
S1 Kota Samarahan Roadside S2 Kota Samarahan Roadside
MNA3 Mentawai Jungle
MNA27 Mentawai Jungle
Molecular Identification of the Top 11 Fungal Isolates
Fungi morphological characterization has been used widely for the identification of fungi.
However, fungi identification by visual examination is rather time consuming, erroneous,
difficult, and requires an extensive taxonomical knowledge, compared to the molecular
technique, which is more sensitive, specific and accurate, and does not demand the
specialized taxonomical expertise (Gherbawy & Voigt 2010).
All the selected isolates were successfully identified using molecular method through
sequence homology of their Internal Transcribed Spacer (ITS) genes. The majority of the
isolates were grouped with members of the Phomopsis, followed by Diaporthe,
Nigrospora, and Xylaria (see Table 3.2 for an overview of closest matches and Figure 3.1
for the phylogenetic tree). From the fungal isolates identification, all of the identified
fungi genus (Phomopsis, Diaporthe, Nigrospora, and Xylaria) have been previously
reported as endophytic fungi.
Most of the sequences (55% - NA25, NA27, NA28, MNA3 and MNA27) obtained were
closely related to the fungal genus of Phomopsis. Phomosis sp. were previously found to

- 68 -
be plant pathogens that can cause foliar infections (Sharma & Florence 1997) and stem
cankers (Hadi & Nuhamara 1997) to some plants. However, our results show that
Pomopsis sp. appeared as Nepenthes ampullaria associated endophytic fungi. Similar
results were also obtained from an endophytic fungi study carried out by Lee and his team
(2014) which revealed that Phomopsis were isolated from Nepenthes plants. Besides that,
Phomopsis sp. also appeared as endophytes of the Tripterygium wilfordii plant (Kumar
& Hyde 2004), Spondias mombin plant (Rodrigues et al. 2000) and Garcinia plant
(Phongpaichit et al. 2007). Phomopsis have been previously recorded to be able to tolerate
heavy metals such as aluminium, lead, and chromium, with a copper tolerance up to 500
ppm (Sim et al. 2015). Our study highlights the higher tolerance of the Phomopsis sp.
against the heavy metal up to 1000 ppm, which is 2 times higher compared to the previous
study.
Three (27% - NA31, S1, S2) of the fungal isolates were grouped under Diaporthe sp..
Diaporthe is a teleomorph form of Phomopsis (Santos & Phillips 2009). Diaporthe
phaseolorum was previously recorded to be a fungal parasite in plants and also involved
in cutaneous infections in humans (Mattei et al. 2013). Some studies have shown that
Diaporthe phaseolorum can also appear as an endophyte in certain plants such as the
medicinal plant Baccharis trimera (Vieira et al. 2014), Laguncularia racemosa
(Sebastianes et al. 2012) and Taxus wallichiana var. mairei (Zaiyou et al. 2013).
Diaporthe are poorly understood in terms of their heavy metal tolerance, however,
Tomono and team (1982) have demonstrated that pycnospores of Diaporthe citri are able
to germinate with a germination rate of 22% under 10-5M (16 ppm) copper solution. Our
study shows the capability of Diaporthe to tolerate copper up to 1000 ppm.
Only 18% of the fungal isolates were grouped under the genus of Nigrospora (9%) and
Xylaria (9%). Both of the Nigrospora sphaerica and Xylaria sp. have been widely
reported as endophytes. For example, Senthilkumar and team (2014) reported endophytic
Nigrospora sphaerica isolation from tropical tree species (Tectona grandis L.) of India,
and Gallo and the team (2009) also reported isolation of endophytic Nigrospora sphaerica
from an Andean plant (Smallanthus sonchifolius). Xylaria species have a very wide
geographical distribution (Patil et al. 2012, Stadler et al. 2014). They have also been found
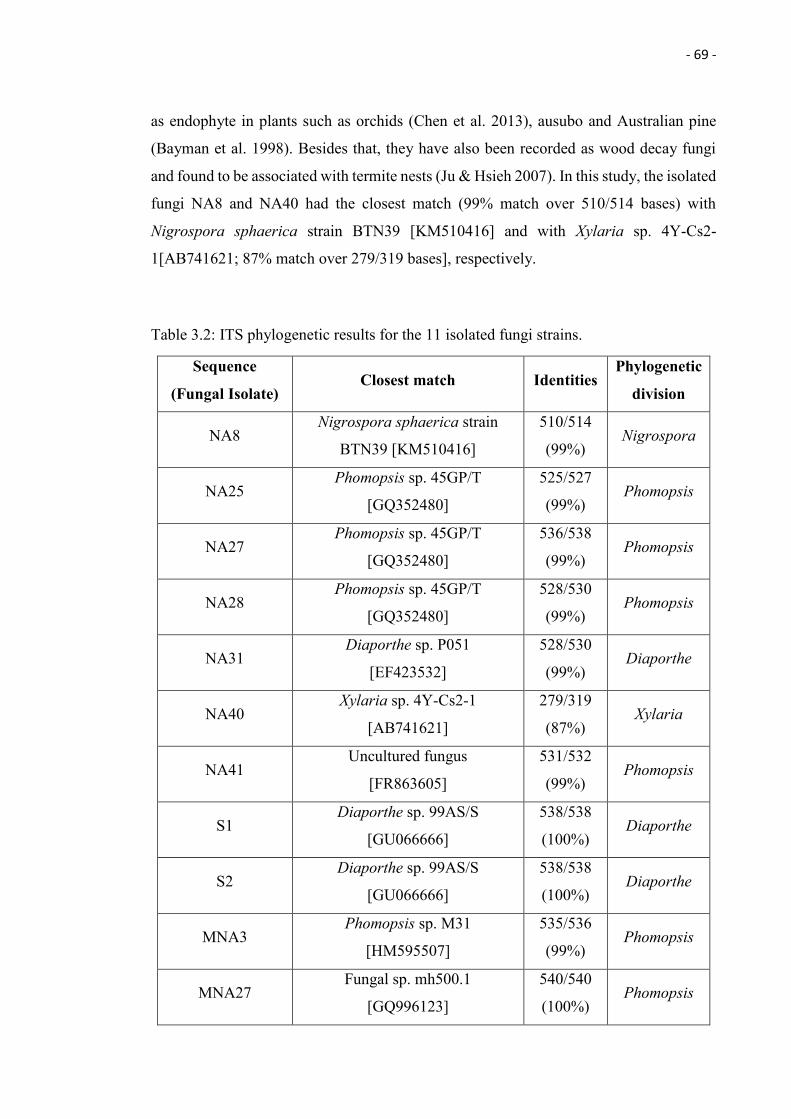
- 69 -
as endophyte in plants such as orchids (Chen et al. 2013), ausubo and Australian pine
(Bayman et al. 1998). Besides that, they have also been recorded as wood decay fungi
and found to be associated with termite nests (Ju & Hsieh 2007). In this study, the isolated
fungi NA8 and NA40 had the closest match (99% match over 510/514 bases) with
Nigrospora sphaerica strain BTN39 [KM510416] and with Xylaria sp. 4Y-Cs2-
1[AB741621; 87% match over 279/319 bases], respectively.
Table 3.2: ITS phylogenetic results for the 11 isolated fungi strains.
Sequence
(Fungal Isolate) Closest match Identities
Phylogenetic
division
NA8 Nigrospora sphaerica strain
BTN39 [KM510416]
510/514
(99%) Nigrospora
NA25 Phomopsis sp. 45GP/T
[GQ352480]
525/527
(99%) Phomopsis
NA27 Phomopsis sp. 45GP/T
[GQ352480]
536/538
(99%) Phomopsis
NA28 Phomopsis sp. 45GP/T
[GQ352480]
528/530
(99%) Phomopsis
NA31 Diaporthe sp. P051
[EF423532]
528/530
(99%) Diaporthe
NA40 Xylaria sp. 4Y-Cs2-1
[AB741621]
279/319
(87%) Xylaria
NA41 Uncultured fungus
[FR863605]
531/532
(99%) Phomopsis
S1 Diaporthe sp. 99AS/S
[GU066666]
538/538
(100%) Diaporthe
S2 Diaporthe sp. 99AS/S
[GU066666]
538/538
(100%) Diaporthe
MNA3 Phomopsis sp. M31
[HM595507]
535/536
(99%) Phomopsis
MNA27 Fungal sp. mh500.1
[GQ996123]
540/540
(100%) Phomopsis
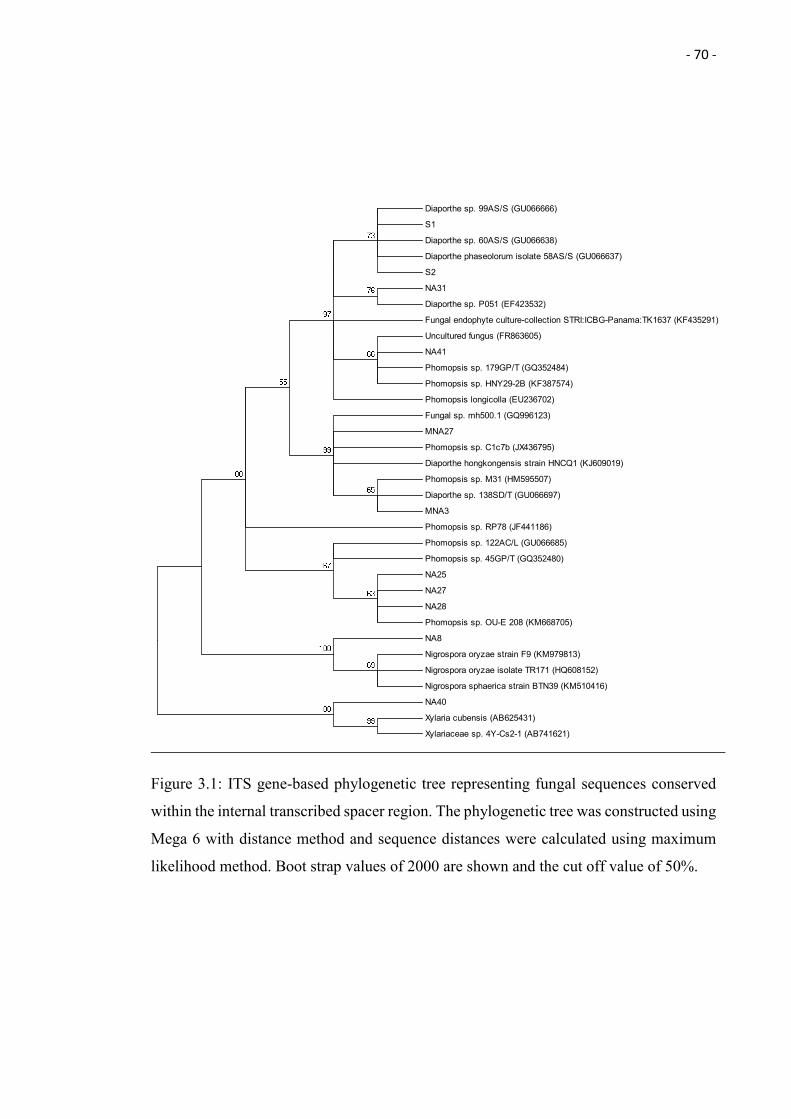
- 70 -
Figure 3.1: ITS gene-based phylogenetic tree representing fungal sequences conserved
within the internal transcribed spacer region. The phylogenetic tree was constructed using
Mega 6 with distance method and sequence distances were calculated using maximum
likelihood method. Boot strap values of 2000 are shown and the cut off value of 50%.
Diaporthe sp. 99AS/S (GU066666)
S1
Diaporthe sp. 60AS/S (GU066638)
Diaporthe phaseolorum isolate 58AS/S (GU066637)
S2
NA31
Diaporthe sp. P051 (EF423532)
Fungal endophyte culture-collection STRI:ICBG-Panama:TK1637 (KF435291)
Uncultured fungus (FR863605)
NA41
Phomopsis sp. 179GP/T (GQ352484)
Phomopsis sp. HNY29-2B (KF387574)
Phomopsis longicolla (EU236702)
Fungal sp. mh500.1 (GQ996123)
MNA27
Phomopsis sp. C1c7b (JX436795)
Diaporthe hongkongensis strain HNCQ1 (KJ609019)
Phomopsis sp. M31 (HM595507)
Diaporthe sp. 138SD/T (GU066697)
MNA3
Phomopsis sp. RP78 (JF441186)
Phomopsis sp. 122AC/L (GU066685)
Phomopsis sp. 45GP/T (GQ352480)
NA25
NA27
NA28
Phomopsis sp. OU-E 208 (KM668705)
NA8
Nigrospora oryzae strain F9 (KM979813)
Nigrospora oryzae isolate TR171 (HQ608152)
Nigrospora sphaerica strain BTN39 (KM510416)
NA40
Xylaria cubensis (AB625431)
Xylariaceae sp. 4Y-Cs2-1 (AB741621)

- 71 -
Biosorption of Copper by Fungal Live Biomass
As can be seen in Table 3.3, all the live biomass of the isolated fungi (Phomopsis,
Diaporthe, Nigrospora, and Xylaria) have been shown to biosorb metal ion. Fungal
isolate NA40 had the highest biosorption capacity of 73.26 mg/g and it was molecularly
identified to be closely related to Xylaria sp. 4Y-Cs2-1 [AB741621]. Xylaria sp. have
been previously studied for their capability of degrading lignin (Pointing et al. 2003) and
the pollutant polycyclicaromatic hydrocarbons (benzo(a)pyrene) (Chang et al. 2007). Our
research newly uncovered the ability of Xylaria sp. to tolerate the heavy metal copper up
to 1000 ppm, and to remove the heavy metal from the liquid solution by using live and
dead biomass. To the best of our knowledge, this is the 1st report of Xylaria sp. in regards
to copper tolerance and biosorption of copper.
The isolated fungus NA41 achieved the 2nd highest biosorption capacity of 71.34 mg/g,
followed by the other fungal isolates NA27, NA25, NA28 and MNA27, which had live
biomass copper biosorption capacities of 71.34, 59.81, 52.70, 52.31, 36.47 mg/g,
respectively (Table 3.3). Fungi MNA3 had the lowest biosorption capacity of 13.42mg/g.
All of the 6 fungal isolates mentioned above were grouped with Phomopsis. Phomopsis
have been previously reported to be able to degrade toxic recalcitrant N-heterocyclic
compounds (Indole and its derivatives) and remove heavy metal zinc from liquid solution
by its active biomass with a bioadsorption capacity of 0.055 mg/g ± 0.005 of dry weight
(Wang et al. 2014; Alozo et al. 2002). In our research, the isolates related to Phomopsis
sp. showed to be capable of removing heavy metal copper from the liquid solution at a
much higher rate.
Nigrospora sp. have been previously reported to have the ability of degrading hop bitter
acids (Huszcza et al. 2008), and toxic synozol red dye into non-toxic metabolites that are
safe for plant growth (Ilyas et al. 2013). A marine-derived Diaporthe sp. was found to be
able to decolorize and detoxify the raw textile effluent that contains toxic dyes such as
azo dye (Verma et al. 2010). As mentioned above, Phomopsis, an anamorph form of
Diaporthe, was found to be able to remove heavy metal zinc from liquid broth. Besides
that, the heavy metal copper ion absorption into the cytoplasm within the pychospore of
Diaporthe citri has been reported by Tomono et al. 1982, however, no specific amount of
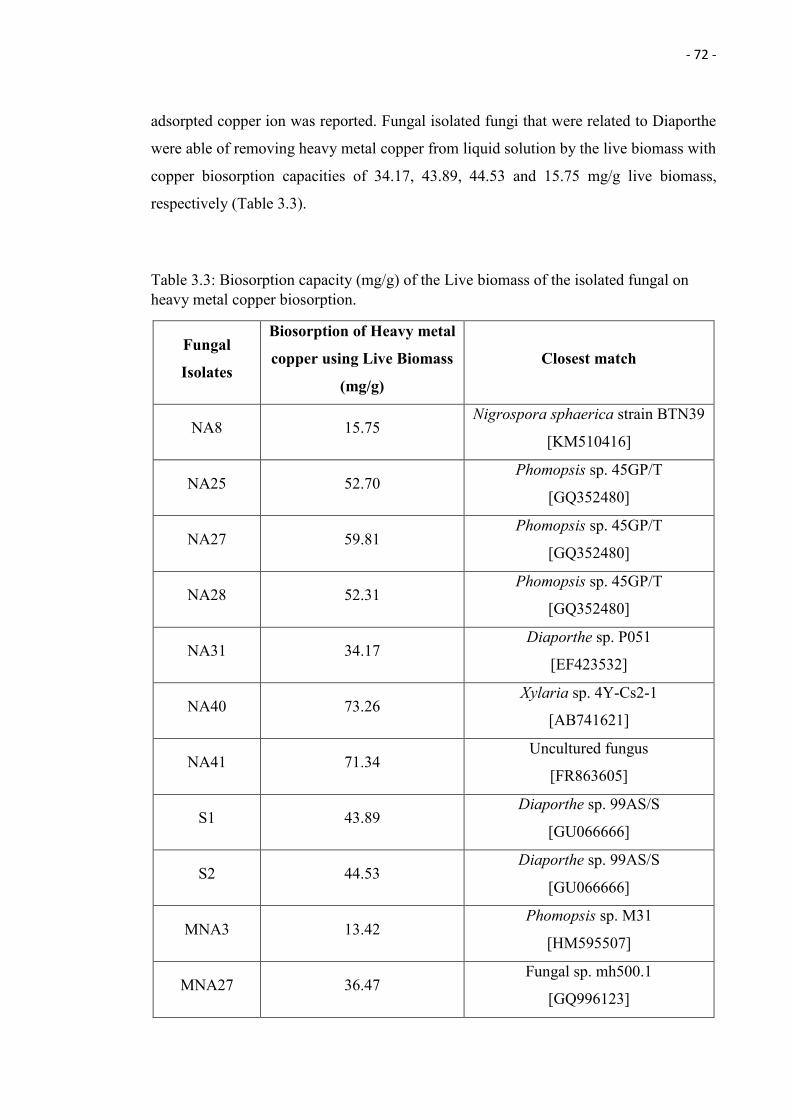
- 72 -
adsorpted copper ion was reported. Fungal isolated fungi that were related to Diaporthe
were able of removing heavy metal copper from liquid solution by the live biomass with
copper biosorption capacities of 34.17, 43.89, 44.53 and 15.75 mg/g live biomass,
respectively (Table 3.3).
Table 3.3: Biosorption capacity (mg/g) of the Live biomass of the isolated fungal on heavy metal copper biosorption.
Fungal
Isolates
Biosorption of Heavy metal
copper using Live Biomass
(mg/g)
Closest match
NA8 15.75 Nigrospora sphaerica strain BTN39
[KM510416]
NA25 52.70 Phomopsis sp. 45GP/T
[GQ352480]
NA27 59.81 Phomopsis sp. 45GP/T
[GQ352480]
NA28 52.31 Phomopsis sp. 45GP/T
[GQ352480]
NA31 34.17 Diaporthe sp. P051
[EF423532]
NA40 73.26 Xylaria sp. 4Y-Cs2-1
[AB741621]
NA41 71.34 Uncultured fungus
[FR863605]
S1 43.89 Diaporthe sp. 99AS/S
[GU066666]
S2 44.53 Diaporthe sp. 99AS/S
[GU066666]
MNA3 13.42 Phomopsis sp. M31
[HM595507]
MNA27 36.47 Fungal sp. mh500.1
[GQ996123]

- 73 -
Biosorption of Copper by Fungal Dead Biomass
The capability of the biosorption of heavy metal copper by using the dead biomass of the
isolated fungi (Phomopsis, Diaporthe, Nigrospora, and Xylaria) have been studied and
all of the isolates showed to be able to biosorb the heavy metal copper by their dead
biomass (Table 3.4). The fungal NA41 whichhad the closest match with an uncultured
fungus [FR863605] – phylogenetic division of Phomopsis (99% match over 531/532
bases), showed to have the highest biosorption capacity of 59.33 mg/g. The fungal isolate
NA25 that had the closest match with Phomopsis sp. 45GP/T [GQ352480; 99% match
over 525/527 bases], however, had the lowest biosorption capacity of copper (9.87 mg/g).
Apart from that, the other fungal isolates (NA27, NA28, MNA3, MNA27) that were all
grouped with the same genus, Phomopsis, showed to have copper biosorption capacities
of 11.05, 15.42, 23.07, and 39.73 mg/g, respectively (Table 3.4). Phomopsis sp. have been
previously shown to have the capability to bioadsorb 179 mg/g of lead, 26 mg/g of
cadmium, 6 mg/g of nickel, 10 mg/g of zinc and 25 mg/g of copper into the insoluble
polysaccharidic biomaterial, with the chitosan and glucans as the main components
(Saiano et al., 2005). Besides that, Sim et al. (2015) also demonstrated that the biosorption
of the heavy metal using the dead biomass of Phomopsis sp. had a biosorption capacity
of 19.6 of lead, 20.1 mg/g of cadmium, 18.1 mg/g of zinc, 16.9 mg/g of chromium and
17.4 mg/g of copper. Moreover, removal of pesticides oxadixyl from an aqueous solution
by using a very similar biomaterial that contains chitosan and glucans were also presented
by Saiano and Ciofalo (2007). Two of the isolates related to Phomopsis sp. (NA41 and
MNA27) showed to have a better biosorption capacity of 2.37 times higher for the isolate
NA41 and 1.58 times higher for the isolate MNA27, compared to the previously reported
study by Saiano et al. (2005). Three of the isolates related to Phomopsis sp. shows to have
better biosorption capacity of 3.41 times higher (isolate NA41), 2.28 times higher
(MNA27) and 1.33 times higher ( MNA3), compared to the previously reported study by
Sim et al. (2015).
Fungal isolate NA40 which was closely related to Xylaria sp. achieved the 2nd highest
biosorption capacity of 44.86 mg/g, while the isolates that were closely related to
Nigrospora sphaerica (NA8), Diaporthe sp. (NA31, S1 and S2), had the biosorption
capacities of 30.49, 30.93, 28.67, and 16.04 mg/g, respectively (Table 3.4). The dead
biomass of the anamorph of Diaporthe, Phomopsis, have been record to be able to biosorb
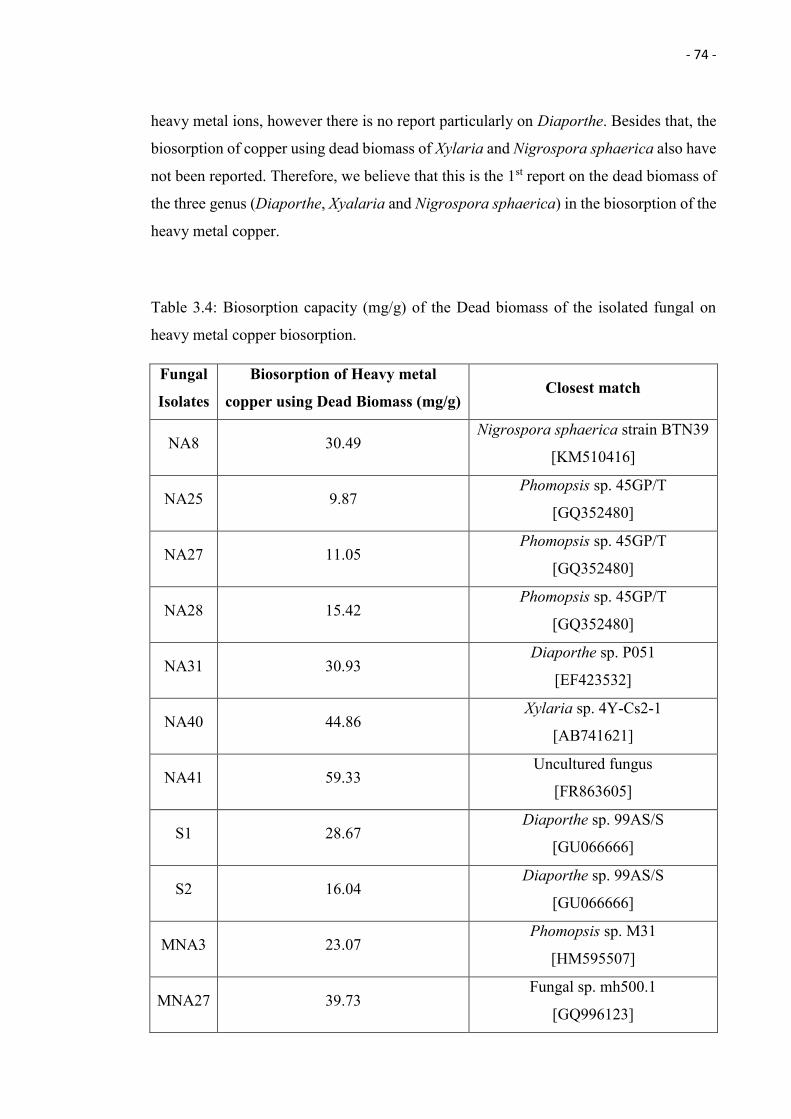
- 74 -
heavy metal ions, however there is no report particularly on Diaporthe. Besides that, the
biosorption of copper using dead biomass of Xylaria and Nigrospora sphaerica also have
not been reported. Therefore, we believe that this is the 1st report on the dead biomass of
the three genus (Diaporthe, Xyalaria and Nigrospora sphaerica) in the biosorption of the
heavy metal copper.
Table 3.4: Biosorption capacity (mg/g) of the Dead biomass of the isolated fungal on
heavy metal copper biosorption.
Fungal
Isolates
Biosorption of Heavy metal
copper using Dead Biomass (mg/g) Closest match
NA8 30.49 Nigrospora sphaerica strain BTN39
[KM510416]
NA25 9.87 Phomopsis sp. 45GP/T
[GQ352480]
NA27 11.05 Phomopsis sp. 45GP/T
[GQ352480]
NA28 15.42 Phomopsis sp. 45GP/T
[GQ352480]
NA31 30.93 Diaporthe sp. P051
[EF423532]
NA40 44.86 Xylaria sp. 4Y-Cs2-1
[AB741621]
NA41 59.33 Uncultured fungus
[FR863605]
S1 28.67 Diaporthe sp. 99AS/S
[GU066666]
S2 16.04 Diaporthe sp. 99AS/S
[GU066666]
MNA3 23.07 Phomopsis sp. M31
[HM595507]
MNA27 39.73 Fungal sp. mh500.1
[GQ996123]

- 75 -
Comparison between Biosorption of Copper by Fungal Live and Dead Biomass
As we can see in the Tables 3.3 and 3.4, some of the fungal isolates that belong to the
same genus, displayed very different biosorption capacities. For example, isolate NA25,
NA27 and NA28, all are closely related to Phomopsis sp., however their live and dead
biomass biosorption capacity of heavy metal copper are vary among each other
(biosorption capacity of 52.70 mg/g of live biomass and 9.87 mg/g of dead biomass for
NA25, 59.81 mg/g of live biomass and 11.05 of dead biomass for NA27, while 52.31 of
live biomass and 15.42 mg/g of dead biomass for NA28). Besides that, isolate S1 and S2
were closely related to Diaporthe sp. but recorded different biosorption capacities of
copper (Biosorption capacity of 43.89 mg/g of live biomass and 28.67 mg/g of dead
biomass for S1, while 44.53mg/g of live biomass and 16.04 mg/g of dead biomass of S2).
Similar results have been observed by Redman et al. (2011). Endophytic fungi of the same
species (Colletotrichum magna, path-1 and L2.5) were inoculated into pepper plant
(Capsicum annuum) and resulted in different drought tolerance. Biosorption capacity can
also be affected by different chemical compositions of the cell wall. Aspergillus fungus
and Mucor rouxii have been found to possess different percentages of chitin in the cell
wall, which has been linked to their differing biosorption capacities (Volesky 1990).
Interestingly, the 4 Phomopsis sp. isolated from the roadside (labelled with NA) have
higher copper biosorption capacity using live biomass compared to the other 2 Phomopsis
sp. isolated from the jungle plant (labelled with MNA). This might due to the reason
mentioned earlier, the pre-exposure and long-term adaptation of the endophytic fungi
along the roadside. Similar results also obtained from the research done by Helander
(1995) showing better tolerant of heavy metal nickel and copper in –vitro of the
endophytic fungal strain, Hormonema sp., isolated from the plant taken near the pollution
source in Harjavalta factory than those isolated from the plant taken 8 km from the factory,
where the pollution level is lower. Ge et al. (2011) suggested that the adaptation of the
fungi toward the heavy metal will also lead to the modification of the hyphae cell surface
which is believed to be involved in fungal intracellular detoxification of the heavy metal.
Based on our results, there is a clear distinction between absorption capacities if living or
dead biomass is used. Eighty percent of the roadside isolates displayed higher copper

- 76 -
biosorption capacity using living biomass, whereas none of the jungle isolates displayed
the same trend. However, when using dead biomass, no trend was observable between
roadside or jungle isolates. A reason might be that the biosorption of heavy metals using
dead biomass is metabolism independent, and therefore purely depends on physical
adsorption (Aksu et al. 1992, Kuyucak & Volesky 1988b), ion exchange (Muraleedharan
& Venkobachar 1990; Kuyucak & Volesky 1988) and complexation (Aksu et al. 1992).
Dead biomass is usually preferred for metal biosorption due to better biosorption
efficiency compared to living biomass. However, our research shows that living biomass
has a higher biosorption capacity compared to dead biomass. Similar results were
obtained by Abedin (2014), where the living biomass of Penicillium oxalicum JQ624873
had a higher Cu biosorption efficiency of 59%, compared to 48% using dead biomass
Another study by Kahraman et al. (2005) also found that living biomass of two white rot
fungi and Phanerochaete chrysosporium had higher Cu adsorption capacity compared to
the dried biomass. Kapoor et al. (1998) achieved significant improvements in Cu
biosorption when the live fungal biomass of Aspergillus niger was pre-treated with
formaldehyde, dimethyl sulphoxide and sodium hydroxide. However, such pre-treatment
has not been undertaken in this study, which underlines the capability of our isolates to
actively deal with high Cu concentrations and also opens possible routes of further
maximising their biosorption capacities.
3.4 Conclusion
In total, 147 fungal isolates were collected from Nepenthes ampullaria, ninety two (92)
isolates were from plants collected in Mentawai Jungle, while the other fifty five (55)
isolates were from plants collected at the roadside of Kota Samarahan, Kuching, Sarawak,
Malaysia. Only 7.5% of the total isolates managed to survive in copper concentrations up
to 1000 ppm. The highest Cu biosorption capacity of live biomass was achieved by fungal
isolate NA40 (related to Xylaria sp.; 73.26 mg/g), whereas NA41 (related to Phomopsis
sp.) exhibited the highest Cu biosorption capacity using dead biomass (73.26 mg/g).
This is the first time that the heavy metal copper tolerance of Xylaria, Diaporthe and
Nigrospora sphaerica and copper biosorption using live biomass of Xylaria and
Nigrospora sphaerica and dead biomass of Xylaria, Diaporthe and Nigrospora oryza

- 77 -
were studied. Our studies highlighted that fungal biosorption capacity is highly dependent
on the sampling area (roadside vs. jungle) and the fungal species. It also highlighted that
different biosorption mechanisms (live- metabolic dependent and dead biomass-
metabolic independent) result in different amounts of copper being removed from the
solutions.

- 78 -
Chapter 4
Proteomics analysis of the Nepenthes
ampullaria associated endophytic fungus,
Xylaria sp.
Changi Wong1*, Daniel Tan1, Samuel Lihan2, Aazani Mujahid2, Jameel R. Al-Obaidi3,
Norasfaliza Rahmad3, and Moritz Müller1
1 Faculty of Engineering, Computing and Science, Swinburne University of Technology
Sarawak, 93350 Kuching, Malaysia.
2 Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 93400
Kota Samarahan, Sarawak, Malaysia.
3 Agro-Biotechnology Malaysia Institutes, c/o MARDI Headquarters, Serdang 43400,
Selangor, Malaysia.
*Corresponding author
Email: [email protected]
Phone number: +60168716911
ABSTRACT
Proteomics study is one of the ways to study the differentially expressed proteins of the
endophytic fungus in response to heavy metal. In this study, the previously reported
Nepenthes ampullaria associated endophytic fungus, Xylaria sp., was used to perform
proteomics analysis in heavy metal copper exposure. In total, 11 proteins spots were
found to be up-regulated and 1 protein spots was found to be down-regulated in response
to heavy metal copper. The protein spots were identified to be related to the enzymes that
involve in heat shock protein, DNA repairing and antioxidant catalysation. To our
knownledge, this study is the first study on Xylaria and serves as a base line study for the
response of this particular fungus genus to the heavy metal copper.
Keywords: Endophytic fungi, Proteomics, Heavy metal copper, Xylaria, Nepenthes ampullaria

- 79 -
4.1 Introduction
Endophytes are microorganisms (bacteria (Hallmann et al. 1997) and fungi (Araújo et al.
2001; Hallmann et al. 1997) that live symbiotically and asymptomatically within their host
plants tissue. The host plant provides nutriments and restful habitation for them, while in
return they will protect their host plant from biotic and abiotic stress such as drought and
salinity resistance (Tadych & White 2009; Rodriguez et al. 2008), thermotolerance
(Redman et al. 2002), improved plant nutrients uptake and heavy metal tolerance (Tadych
& White 2009). Endophytic fungi are known to produce valuable bioactive compounds
and enzymes that exhibit anti-microbial (Li et al. 2000; Li et al. 2001; Miller et al. 1998),
anti-cancer (Li et al. 1996), immunosuppressant (Lee et al. 1995), pectinase, laccase, and
protease properties (Maria et al. 2005). Other than mutualism with the host plant,
endophytic fungi itself are known to be capable to resist certain toxic heavy metals such
as cadmium (Deng et al. 2014; El-Gendy et al. 2011), zinc (Deng et al. 2014) and copper
(El-Gendy et al. 2011), and bioaccumulate or biosorb them (Deng et al. 2014; El-Gendy
et al. 2011). For example, Phomopsis sp. NA41, an endophytic fungi isolated from
Nepenthes ampullaria, was found to be able to tolerate heavy metal copper concentrations
up to 1000 ppm and to remove the heavy metal from solution by using its live and dead
biomass (Wong et al. submitted). Besides that, fungi are also known to increase or
decrease the expression of certain proteins under heavy metal stress. For instance,
fourteen up-regulated and 21 down-regulated proteins were detected from Phanerochaete
chrysosporium under lead stress (Yıldırım et al. 2011), and when the particular fungal
strain underwent copper and cadmium stress, 74 copper upregulated proteins and 80
cadmium upregulated proteins spots were detected (Ozcan et al. 2007).
Heavy metal copper is one of the essential micronutrients required for the cell to carry
out biochemical processes. However, excess in copper may be toxic to the cell due to the
oxidative potential of the copper towards the proteins (Letelier et al. 2005), lipids (Zhao
et al. 2014) and nucleic acids (Linder 2012), leading to lethal microbial cell damage. In
order to survive the heavy metal induced oxidative stress, microorganisms such as yeast
(Jo et al. 2008) and fungi (Yıldırım et al. 2011), were found to increase the production of
the enzymes that involve in cell protecting such as antioxidant (Yıldırım et al. 2011, Rout
& Sahoo 2013), DNA repair (Jo et al. 2008; Yıldırım et al. 2011), and heat shock protein
(Yıldırım et al. 2011). According to González-Guerrero et al. (2010), GintABC1, which

- 80 -
encodes a putative ABC transporter of the MRP subfamily, was induced by copper and
cadmium stress in Glomus intraradices. Besides that, Pimt1 genes, which encode for the
metallothioneins that are involved in metal tolerance of most eukaryotes, were detected
in Paxillus involutus by Bellion et al. (2007). While it is possible to detect most of the
genes related to heavy metal resistance using genomic studies, the biological mechanisms
such as protein expression and protein-protein interaction, cannot be explored using
genomic studies, but by proteomics.
Proteomics, the study of the proteome, can provide information on the structure and
function of a protein which we cannot obtain using genomic studies. Proteomes are
dynamic, in contrast to the relatively static genomes. Proteomes often undergo changes
in response to extra- and intracellular signals and has been defined as “the proteins present
in one sample (tissue, organism, and cell culture) at a certain point in time” (Rastogi et al.
2006; Ravi et al. 2013).
Xylaria, under the family of Xylariaceae, is a fungal genus that has a wide geographical
distribution (Patil et al. 2012; Stadler et al. 2014). It has been found as wood decaying
fungi and associated with termite nests (Ju & Hsieh 2007). Besides that, Bayman et al.
(1998) and Ratnaweera et al. (2014) also showed that Xylaria exist as endophytic fungi
in certain plants such as asubo and Australian pine and orchid. Xylaria are previously
famous for their antimicrobial properties (Ratnaweera et al. 2014), cytotoxic properties
(Inthe et al. 2014) and wide enzymatic properties (Wei et al. 1992; Liers et al. 2007; Liers
et al. 2006). However, the study of its proteome expression towards copper has yet to be
explored and recorded.
In a previous study (Wong et al., submitted), an endophytic fungi isolate related to Xylaria
sp. exhibited the ability to survive copper concentrations up to 1000 ppm. It also displayed
the highest biosorption capability (using its living biomass) among all the others 11 fungal
isolates, including Phomopsis, Diaporthe, and Nigrospora. Therefore, it was chosen to
identify differentially expressed proteins in response to treatments with 3 different
concentration of copper (0, 300, and 500 ppm). This study is the first to our knowledge

- 81 -
on Xylaria and serves as a base line study for the response of this particular fungus genus
to the heavy metal copper.
4.2 Methodology
4.2.1 Culture Conditions
The fungus (NA40) was cultured in Potato Dextrose Broth (PDB) supplied with 2
different concentrations of copper (300 and 500 ppm) and another PDB without copper
as control for 3 weeks at 25oC. After 3 weeks incubation, the fungal biomass was
harvested and underwent protein extraction. All conditions were performed in triplicates.
4.2.2 Protein Extraction
The TCA-acetone extraction performed by Méchin et al. (2007) was used to extract the
protein from the fungal cells. The fungal biomass was grounded into fine powder using
mortar and pestle in the presence of liquid nitrogen. TCA-acetone extraction was
performed by mixing 1 g of the powdered fungal biomass with 1.8 ml of 10%
trichloroacetic acid in cold acetone containing 0.07% β-mercaptoethanol and vortexed at
4oC. After that, the mixture was incubated in -20oC overnight and on the following day,
the mixture was centrifuged at 10,000 g at 4oC for 15 minutes. The supernatant was
discarded and the pellet was re-suspended in rinsing solution (each of the 1 g with 1.8 ml
rinsing solution) containing 0.07% β-mercaptoethanol in cold acetone. The solution was
then incubated at -20°C for 1 h (mixed at 15 min intervals) and again centrifuged at
10,000g at 4oC for 15 minutes. The supernatant was discarded and this rinsing step
repeated twice. The pellet was then vacuum-dried and re-suspended with lysis buffer. The
supernatant was collected and stored at -80oC. The protein concentration was determined
using Biorad Bradford Reagent assay.
4.2.3 2-DE and Image Analysis of Protein Spots
2D gels of the control and each treatment were run in triplicates. Isoelectric focusing (IEF)
was performed using 13 cm Nonlinear IPG-strips (pH range 3-10). The IPG-strips were
initially rehydrated for 12 hours in the presence of 70 μg of protein samples. IEF was

- 82 -
performed using Biorad Protean i12 with standard protocol based on Biorad Handbook
(IEF Protocol), at 20 °C in a stepwise manner: 500 V (2 h), 1.0 kV (1 h), 8.0 kV (1 h),
8.0 kV (28000 VhS) and finally 750 V (hold). The strips were equilibrated in equilibration
buffer (based on GE Healthcare 2D SDS PAGE Handbook) containing 50 mM Tris–HCl
pH 8.8, 6 M urea, 30% (v/v) glycerol, 2% (w/v) SDS, 0.002% (w/v) bromophenol blue
and 1% (w/v) dithiothreitol (DTT) for 15 minutes, followed by equilibrated in the same
equilibration buffer containing 2.5% (w/v) iodoacetamide instead of DTT for another 15
minutes. The second dimension separation was performed in 12% polyacrylamide gels,
at 20oC, using SE 600 Ruby system (Hoefer SE 600 Ruby (Amersham Biosciences)), with
the running buffer contains 25 mM Tris–HCl, 192 mM glycine, 0.1% (w/v) SDS, at
10mA/gel (15min) and 20mA/ gel (3h 30min). The gels were stained with silver staining
(Shevchenko et al., 1996).
4.2.4 Protein Identification and Database Search
Stained gels were digitized by using image Scanner (GS800 Desitometer (Biorad)) and
the protein spots analysis were performed using Progenesis Samespots samespot software,
with the Max fold change ≥ 2, and Anova p-value ≤ 0.05. The chosen proteins spots
were then manually excised from the stained 2D gels and destained followed by in gel
digestion using trypsin overnight at 25oC (Shevchenko et al. 2007). The peptides were
extracted from the gel pieces by using 50% of acetonitrile and 100% acetonitrile for the
second time extraction. The solution is then vaccum dried and stored for further
identification.
Protein identification were done by Norasfaliza Rahmad which was accomplished by
mass spectrometry. Peptide Mass Fingerprinting (PMF) data search was performed using
Swiss-Prot database. The obtained protein ID is then further analysed and studied.

- 83 -
4.3 Results and Discussion
Proteome analyses of fungi in response to the heavy metal copper have been widely
studied using different types of fungi (Cherrad et al. 2012; Yıldırım et al. 2011). However,
no proteomic study of Xylaria sp. in response to copper has been published so far. The
endophytic fungi used in this study -NA40 (closely related to Xylaria sp. – identified
through molecular and morphological methods; see Wong et al., submitted)- was isolated
from Nepenthes ampullaria collected from the roadside of Kota Samarahan, Kuching,
Sarawak, Malaysia. In the following, we discuss its proteome response in the exposure
with varying levels of copper.
Effect on Copper treated on Fungal isolate NA40, Xylaria sp.
The fungus was exposed to two levels of copper (300 and 500 ppm) for three weeks before
protein extraction. After 3 weeks of incubation, the growth rate of the fungus was
significantly reduced in comparison to the copper-free control (data not shown). A total
of 12 protein spots were identified (see Table 4.1) with the 11 spots been up-regulated
and 1 spot been down-regulated (see Table 4.2 and 4.3). Some of the proteins have not
yet been recorded in heavy metal copper stress, while some of the proteins were known
to be heat shock protein, which are expressed under normal conditions and are highly
expressed under stress conditions such as oxidative stress (Becker & Craig 1994), viral
infections (Becker & Craig 1994; Valle et al. 2005) and elevated temperature (Becker &
Craig 1994).
Upregulated Protein spots
As can be seen in Figure 4.1, Formamidopyrimidine-DNA glycosylase is up-regulated by
the fungus in response to copper in order to repair DNA damage. Formamidopyrimidine-
DNA glycosylase is involved in repairing damaged DNA caused by oxidative stress
(UniProta). A study conducted by Boiteux et al. (1992) shows that,
Formamidopyrimidine-DNA glycosylase protein is able to specifically recognise 8-
hydroxypurines within the DNA and repair the damaged DNA in complement with
pyrimidine-specific enzymes, in vivo.

- 84 -
Ribosome recycling factor (RRF) was observed to be up-regulated by 7.39 fold in 300
ppm copper and 3.89 fold in 500 ppm copper (Table 4.2). RRF was previously recorded
in the bacterium Brucella melitensis as a heat shock protein (Teixeira-Gomes et al. 2000).
Besides that, RRF was also reported in plant (maize leaf) responses under chromium
stress (Wang et al. 2013). To the best of our knowledge, this is the 1st report of the
involvement of RRF in a fungus (Xyalaria sp.) in response to copper stress.
A newly described pantothenate kinase - Type III pantothenate kinase (CoaX), which
belongs to the acetate and sugar kinase/heat shock protein 70 /actin (ASKHA) protein
superfamily (Yang et al. 2006) was found to be up-regulated (see Table 4.1). So far,
pantothenate kinase have only been recorded in bacteria such as Helicobacter pylori,
Mycobacterium tuberculosis, Pseudomonas aeruginosa and Bordetella pertussis (Yang
et al. 2006; Awasthy 2010). To the best of our knowledge, there is so far no record of a
fungal strain producing this type of pantothenate kinase (CoaX). CoaX is known to be
involved in the catalyzation of the 1st step in the coenzyme A biosynthesis and is known
to be different from previously reported Type I and Type II pantothenate kinase proteins.
Unlike the Type I and Type II pantothenate kinase, CoaX is not inhibited by CoA and
thioesters. Nicely et al. (2007) suggested that CoaX is involved in maintaining higher
intracellular Coenzyme A levels in many bacteria in order to accommodate additional
functions for the coenzyme in thiol/disulphide redox homeostasis- a homeostasis that is
important for cellular defence against oxidative stress (Hansen et al. 2009). In order to
protect the fungus from copper oxidative stress, the CoaX was found to be up-regulated
in the fungal isolate NA40 (see Table 4.2/Figure 4.1).
Adenylosuccunate synthetase (purA) was previously reported by Winter et al. (2005) to
be up-regulated in Escherichia coli (ΔrpoH strains) when exposed to heat shock and
oxidative stress. The fungal isolate NA40 shows similar behaviour, purA was up-
regulated 6.31 fold when exposed to 300ppm copper and 1.93 fold when expose to
500ppm copper (Table 4.2/ Figure 4.1). purA is involved in the production of fumarate,
an important component of the tricarboxylic acid cycle (TCA) that yields high amounts
of NADH and ATP (Honzatko & Fromm 1999; Horecker & Stadtman 2014). The up-
regulation of the purA in the fungal isolate NA40 suggests that the protein is necessary
to provide energy for the fungal cell to reduce the oxidative damage by the copper, and
also for the metabolic changes that occur in the cell under oxidative stress.

- 85 -
Ketol-acid reductoisomerase has previously been recorded in response to heat stress
(Ferreira et al. 2006), cold shock (Graumann et al. 1996) and salt stress (Zhou et al. 2011),
and is known to be involved in the biosynthesis of amino acids (valine, leucine and
isoleucine) and catalysation of the following two processes (UniProtb):
1. (R)-2,3-dihydroxy-3-methylbutanoate + NADP+ = (S)-2-hydroxy-2-methyl-3-
oxobutanoate + NADPH
2. (2R,3R)-2,3-dihydroxy-3-methylpentanoate + NADP+ = (S)-2-hydroxy-2-ethyl-
3 oxobutanoate + NADPH.
Reduced Nicotinamide Adenine Dinucleotide (NADPH) is an important component in
the cellular antioxidation system and source for reductive synthesis of fatty acid, steroids
and DNA (Pollak et al. 2007). Besides that, it has been record to be involved in the
syntheses of flavonoid, NADPH oxidase and lignin in plant defense (Casati et al. 1999;
Torres 2006; Ying 2008). Ketol-acid reductoisomerase was up-regulated in the fungal
isolate NA40 when exposed to copper, likely leading to an increasing production of
NADPH, flavonoid and the amino acids in order to protect the cell from oxidative
damages and repair damaged DNA.
Cyanide is a toxic chemical that can be produced, metabolized and excreted by algae,
bacteria, insects, plants, and fungi (Cipollone et al. 2008). It can be catalysed by
thiosulfate sulfurtransferase to form the less toxic form of thiocyanate. Thiocyanate has
been previously reported to exhibit roles in the hosts’ defence (Chandler & Day 2012)
and also displayed antioxidant properties in humans (Xu et al. 2009; Chandler & Day 2012).
In order to protect the cell from copper oxidative stress, thiosulfate sulfurtransferase
concentration was up-regulated in the fungal cell at 300 ppm copper and even more at
500 ppm. Similar results (up-regulation of thiosulfate sulfurtransferase) were observed in
Mycobacterium tuberculosis in response to copper (Ward et al. 2008).
Two different types of ribosomal RNA methyltransferase were detected to be up-
regulated in response to the copper stress. One was ribosomal RNA large subunit
methyltarnseferase E (rlmE), which is similar to the heat shock proteins ftsJ and rrmJ

- 86 -
(UniProtc). In recent studies, Caldas et al. (2000) and Toh & Mankin (2008) have
demonstrated that a lack of rlmE decreases growth rate and makes the mutant strain more
sensitive to antibiotics (clindamycin, lincomycin, sparsomycin and hygromycin A)
compared to the wild-type strain. Besides that, Caldas and his research team (2000) also
suggested that the methylation of Um(2552) catalyzed by RrmJ in 23S RNA not only
improves the cell growth rates, it also increases protein synthesis activity, and strengthens
ribosomal subunit interactions even at non-heat shock temperatures. In our study, rlmE
was up-regulated to protect the fungal cell from copper oxidative stress. Another type of
ribosomal RNA methyltransferase detected was ribosomal RNA small subunit
methyltransferase G (rsmG), which was recorded to be involved in streptomycin
resistance and ribosomal functioning (Okamoto et al. 2007). It is known to methylate 16S
rRNA at the N7 position of the G527 nucleotide (Okamoto et al. 2007).
The up-regulation of the proteome rsmG that yields the production of S-adenosyl-L
homocysteine is believed to be involved in cysteine synthesis (UniProtd). This idea is
supported by the up-regulation of pyridoxamine 5' -phosphate oxidase homolog, which
catalyses the oxidation of either pyridoxamine 5' -phosphate (PMP) or pyridoxine 5' -
phosphate (PNP), to form pyridoxal 5' -phosphate (PLP) (Salvo et al. 2003)- an active
form of vitamin B6 that participates in the biosynthesis of the antioxidant compound
cysteine (Schnell et al. 2014). Besides that, vitamin B6 itself also serves as an important
cofactor of many enzymatic reactions such as racemization, transamination,
decarboxylation, trans-sulfuration and deamination (Mittenhube 2001). A few studies
have unravelled the antioxidant properties of vitamin B6 (Ehrenshaft et al. 1999, Osmani
et al. 1999). For instance, vitamin B6 acts as a potential antioxidant involved in the
resistance of Cercospora nicotianae (filamentous fungus) to its own product, cercosporin,
which is a strong photosensitizer of singlet molecular oxygen (1O2) (Bilski et al. 2000).
Besides that, vitamin B6 produced by the transcriptional regulation of GintPDX1 exhibits
antioxidant properties and acts as a modulator of the reactive oxygen species (ROS) of
Glomus intraradices (arbuscular mycorrhizal fungus) (Benabdellah et al. (2009).
Benabdellah et al. (2009) also demonstrated that the transcription of similar genes
(GintPDX1) -encoding for pyridoxal 5'-phosphate biosynthesis enzymes- was found up-
regulated in responses to copper-induced oxidative damage in arbuscular mycorrhizal
fungus, Glomus intraradices.

- 87 -
TtRNA modification GTPase (MnmE), also known as TrmE, can be found in both
prokaryotes and eukaryotes and has been labelled as MSS1 (orthologs of MnmE) in
Saccharomyces cerevisiae (Meyer et al. 2009) and Aspergillus clavatus (NCBIa). MnmE
(termed MSS1) catalyse the biosynthesis of the hyper-modified nucleoside 5-
methylaminomethyl-2-thiouridine (mnm5s2U34) (Elseviers et al. 1984), which is present
at the wobble position (position 34) of the specific tRNAs for lysine and glutamic acid
(Krüger & Sørensen 1998). It was found up-regulated by the fungal isolate NA40 in
response to heavy metal copper stress. MnmE (termed MSS1) have been previously
recorded to be involved in rapid growth at unfavourable conditions (Singh et al. 2009)
and stress responses such as low temperature in psychrophile Pseudomonas syringae
(Singh et al. 2009) and low pH resistance in Escherischia coli by regulating glutamate-
dependent acid resistance (Gong et al. 2004). It seems that they play a similar role for the
fungal isolate NA40.
Uncharacterized protein C24B10,16c is currently known to be an orphan gene (a gene
with no homologues of other organisms in genomes). The protein is upregulated when
exposed to 300 ppm of copper, however, down regulation of the protein was observed
when exposed to 500 ppm copper. The specific function of this protein remains poorly
understood, however, it is believed to be involved in meiotic cell cycle and proteasome
assembly (UniProte).
Down regulated protein spot
Down regulation of the protein Cell division protein SepF was observed in response to
copper (Table 4.3/ Figure 4.2). The protein SepF is involved in late cell division
(UniProtf). This can be an indication that the cell division rate of the fungal isolate NA40
was reduced when exposed to copper (observation supported based on visual inspection
of the fungal biomass; data not shown). Khan and Lee (2013) also demonstrated that
fungal growth rates will reduce in response to heavy metals such as copper and cadmium.
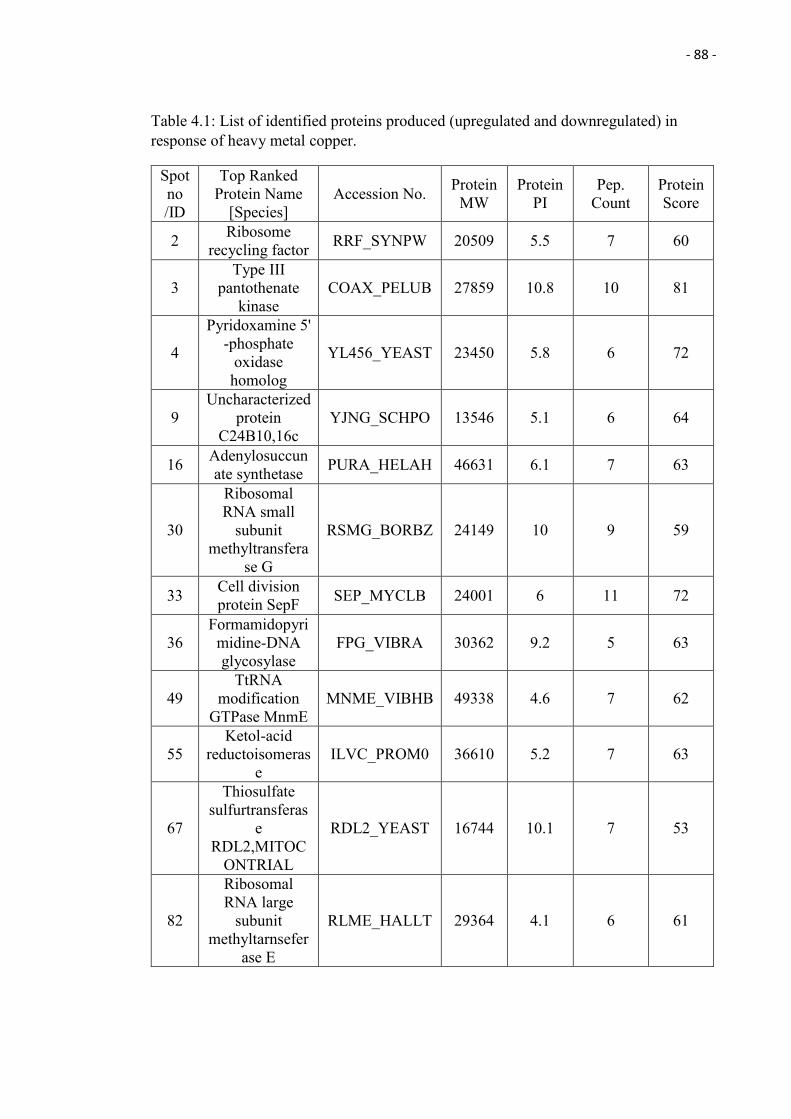
- 88 -
Table 4.1: List of identified proteins produced (upregulated and downregulated) in response of heavy metal copper.
Spot no /ID
Top Ranked Protein Name
[Species] Accession No. Protein
MW Protein
PI Pep.
Count Protein Score
2 Ribosome recycling factor RRF_SYNPW 20509 5.5 7 60
3 Type III
pantothenate kinase
COAX_PELUB 27859 10.8 10 81
4
Pyridoxamine 5' -phosphate
oxidase homolog
YL456_YEAST 23450 5.8 6 72
9 Uncharacterized
protein C24B10,16c
YJNG_SCHPO 13546 5.1 6 64
16 Adenylosuccunate synthetase PURA_HELAH 46631 6.1 7 63
30
Ribosomal RNA small
subunit methyltransfera
se G
RSMG_BORBZ 24149 10 9 59
33 Cell division protein SepF SEP_MYCLB 24001 6 11 72
36 Formamidopyri
midine-DNA glycosylase
FPG_VIBRA 30362 9.2 5 63
49 TtRNA
modification GTPase MnmE
MNME_VIBHB 49338 4.6 7 62
55 Ketol-acid
reductoisomerase
ILVC_PROM0 36610 5.2 7 63
67
Thiosulfate sulfurtransferas
e RDL2,MITOC
ONTRIAL
RDL2_YEAST 16744 10.1 7 53
82
Ribosomal RNA large
subunit methyltarnsefer
ase E
RLME_HALLT 29364 4.1 6 61
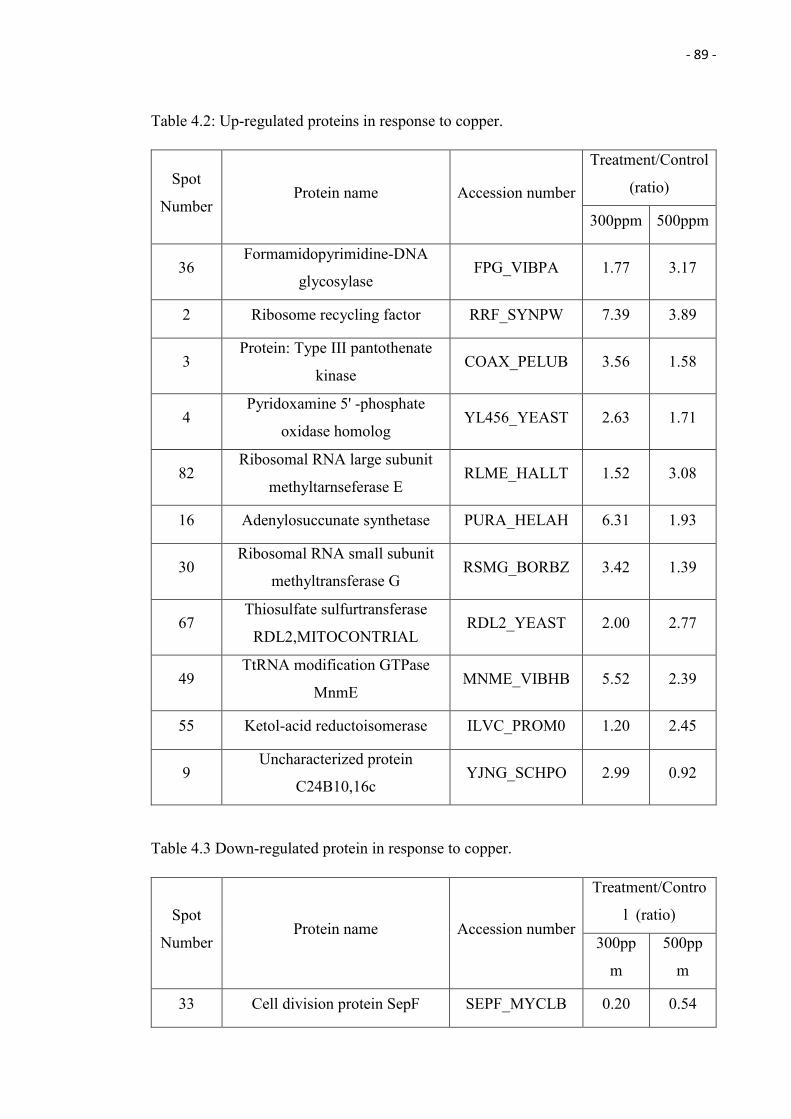
- 89 -
Table 4.2: Up-regulated proteins in response to copper.
Spot
Number Protein name Accession number
Treatment/Control
(ratio)
300ppm 500ppm
36 Formamidopyrimidine-DNA
glycosylase FPG_VIBPA 1.77 3.17
2 Ribosome recycling factor RRF_SYNPW 7.39 3.89
3 Protein: Type III pantothenate
kinase COAX_PELUB 3.56 1.58
4 Pyridoxamine 5' -phosphate
oxidase homolog YL456_YEAST 2.63 1.71
82 Ribosomal RNA large subunit
methyltarnseferase E RLME_HALLT 1.52 3.08
16 Adenylosuccunate synthetase PURA_HELAH 6.31 1.93
30 Ribosomal RNA small subunit
methyltransferase G RSMG_BORBZ 3.42 1.39
67 Thiosulfate sulfurtransferase
RDL2,MITOCONTRIAL RDL2_YEAST 2.00 2.77
49 TtRNA modification GTPase
MnmE MNME_VIBHB 5.52 2.39
55 Ketol-acid reductoisomerase ILVC_PROM0 1.20 2.45
9 Uncharacterized protein
C24B10,16c YJNG_SCHPO 2.99 0.92
Table 4.3 Down-regulated protein in response to copper.
Spot
Number Protein name Accession number
Treatment/Contro
l (ratio)
300pp
m
500pp
m
33 Cell division protein SepF SEPF_MYCLB 0.20 0.54
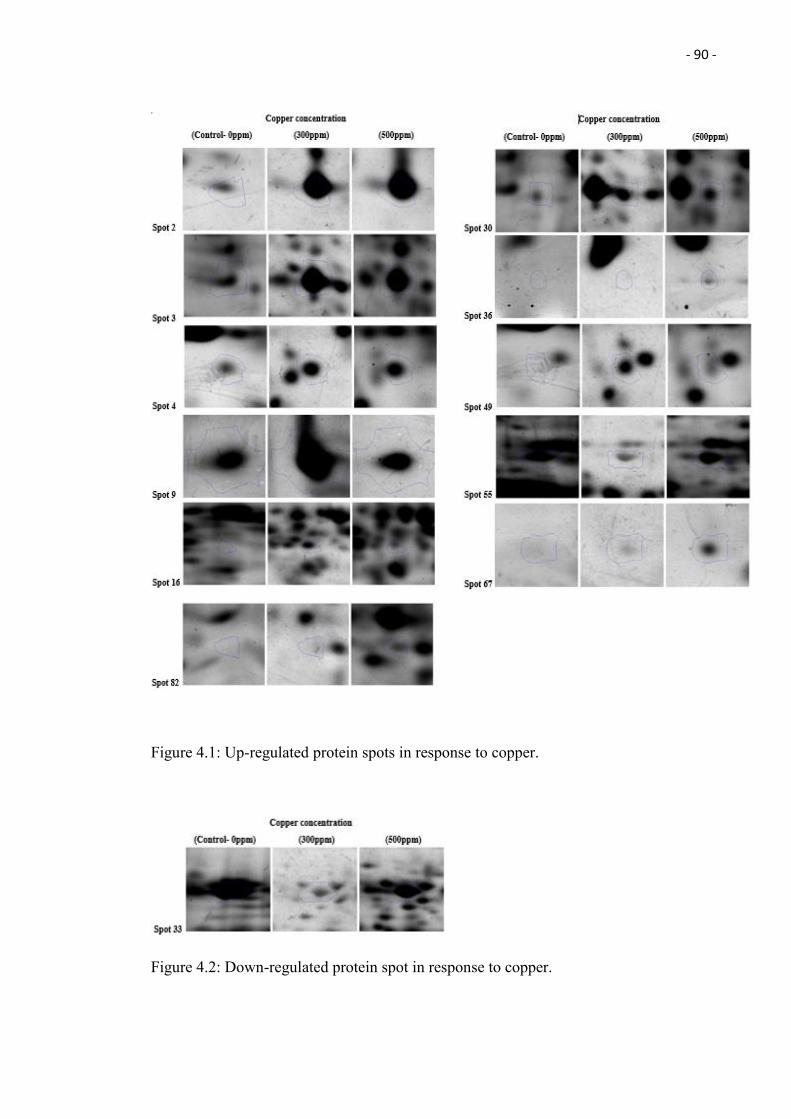
- 90 -
Figure 4.1: Up-regulated protein spots in response to copper.
Figure 4.2: Down-regulated protein spot in response to copper.

- 91 -
4.0 Conclusion
This study is the first to perform a proteomic analysis of the fungus Xylaria sp. in response
to heavy metal copper oxidative stress. The analysis shows that the particular fungus is
capable of producing a wide range of enzymes involved in repair of damaged DNA,
antioxidant catalysation, and heat shock proteins. The results can serve as a baseline study
for this particular fungus genus, Xylaria, on heavy metal copper proteome study.

- 92 -
Chapter 5
Summary, Conclusion and Future Work
5.1 Summary
This research study has presented (i) the capability of the isolated endophytic fungi from
Nepenthes ampullaria plants collected from undisturbed and anthropogenically affected
areas (Mentawai Jungle and Kota Samarahan roadside, Kuching) (a) to resist heavy metal
copper and (b) to biosorp copper from solution by using Live and Dead biomass, and (ii)
express different proteins (fungal isolate NA40) in response to copper stress.
In this study, ninety two (92) fungal isolates were isolated from the Nepenthes ampullaria
plants collected in Mentawai Jungle and fifty five (55) fungal isolates from the roadside
of Kota Samarahan Kuching, Sarawak, Malaysia. In total, there were 147 fungal isolates
collected from Nepenthes ampullaria plants, with the capability of the 7.5% fungal
isolates able to resist heavy metal copper concentration up to 1,000 ppm. The highest Cu
biosorption capacity of live biomass was achieved by fungal isolate NA40 (related to
Xylaria sp.; 73.26 mg/g), whereas NA41 (related to Phomopsis sp.) showed to have the
highest Cu biosorption capacity using its dead biomass (73.26 mg/g). To our knowledge,
this is the first reported study on the copper tolerance of Xylaria, Diaporthe and
Nigrospora sphaerica, and copper biosorption using live biomass of Xylaria and
Nigrospora sphaerica and dead biomass of Xylaria, Diaporthe and Nigrospora oryza.
This study highlights that fungal biosorption capacity is highly dependent on the sampling
area (roadside vs. jungle) and the fungal species. Moreover, the results also highlighted
that the different biosorption mechanisms (live- metabolic dependent and dead biomass-
metabolic independent) result in different amounts of copper being removed from the
solutions.
The proteomics analysis of the fungal isolates NA40 (related to Xylaria sp.) showed that
the particular fungus is able to produce a wide range of enzymes to protect itself from the
oxidative stress caused by copper. The proteins produced include enzymes that repair
damaged DNA, antioxidant and heat shock proteins. To our knowledge, this is the first
proteomic analysis study performed on the fungus Xylaria sp. in response to heavy metal

- 93 -
copper oxidative stress and the results obtained can serve as a baseline study for the
particular fungus genus, Xylaria, on heavy metal copper proteome study.
5.2 Future work
There is no doubt that the biosorption capabilities of the eleven fungi are highly promising,
and have the potential to be used as a new biosorbent materials in the near futures. We
have only just begun to ‘scratch the surface' of bioremediation using endophytes and the
biodiversity treasures of Borneo will surely yield many more surprises. However, future
work should expand our current knowledge and involve researches from chemistry,
biochemistry, genetics, and polymer sciences, in order to fully explore the potential of
endophytes in metal removal and (or) recovery.
Other than heavy metal resistant, biosorption and proteomics studies, endophytic fungi
are also known for their antimicrobial (Phongpaichit et al. 2006) and enzymatic properties
(Sculz et al. 2002). The world's first billion-dollar anticancer drug, Paclitaxel (taxol), was
found to be produced by a wide range of endophytic fungi (Strobel & Daisy 2003).
In the present study, a total number of 147 fungal isolates were collected from the
Nepenthes ampullaria plant and we would suggest to use the isolated fungal to carry out
further studies such as antimicrobial and enzymatic testing, to find out/ unleash/
understand more of their hidden abilities. Besides that, we would also suggest to carry
out the tolerance testing and biosorption on other heavy metals such as chromium, lead,
zinc, mercury and uranium. Moreover, pre-treatment of the fungal dead biomass that
might maximise the biosorption capacities, could also be carried out. Last but not least,
de novo (peptide) sequencing study can be used to confirm and expand upon the results
obtained from database searches (Cagney & Emili 2002).

- 94 -
References
Abedin, RMA 2014, “Optimization and Statistical Evaluation of Copper and Nickel Biosorption Capabilities by Dry Biomass of Penicillium oxalicum JQ624873”, Life Science Journal, vol. 11, no. 2, pp. 61-67.
Abrego, J 1997, “Comparative Evaluation of Collectors for Metal Removal by Precipitation/flotation”, International Journal of Environment and Pollution, vol. 8, no. 1/2, pp. 208–211.
Adam, JH 1997, “Prey Spectra of Bornean Nepenthes Species (Nepenthaceae) in Relation to Their Habitat”, Pertanika Journal of Tropical Agricultural Science, vol. 20, no. 2/3, pp. 121-134.
Adam, JH, Wilcock, CC & Swaine, MD 1992, “The Ecology and Distribution of Bornean Nepenthes”, Journal of Tropical Forest Science, vol. 5, no. 1, pp. 13-25.
Adlassnig, W, Peroutka, M, Lambers, H & Lichtscheidl, IK 2005, “The Roots of Carnivorous Plants”, Plant Soil, vol. 274, pp. 127-140.
Agency for Toxic Substances and Disease Registry (ATSDR) (2004) Public Health Statement for Copper, viewed 2nd January 2015, http://www.atsdr.cdc.gov/ToxProfiles/tp132-c1-b.pdf.
Ahalya, N, Ramachandra, TV & Kanamadi, RD 2003, “Review Paper: Biosorption of Heavy Metals”, Research Journal of Chemistry and Environment, vol. 7, no. 4.
Ahmad, AK, Mushrifah, I & Othman, MS 2009, “Water Quality and Heavy Metal Concentrations in Sediment of Sunagi Kelantan, Kelantan, Malaysia: A baseline study”, Sains Malaysiana, vol. 38, no. 4, pp. 435-442.
Ahmad, I, Ahmad, F, Pichtel, J 2011, Microbes and Microbial Technology: Agricultural and Environmental Applications, Springer Science & Business Media.
Aksu, Z, Sag, Y & Kutsal, T 1992, “The Biosorpnon of Copperod by C. vulgaris and Z. ramigera”, Environmental Technology, vol. 13, no. 6, pp. 579-586.
Angelova, MB, Pashova, SB, Spasova, BK, Vassilev, SV & Slokoska, LS 2005, “Oxidative Stress Response of Filamentous Fungi Induced by Hydrogen Peroxide and Paraquat”, Mycological Research, vol. 109, no. 2, pp. 150-8.
Araújo, WL, Maccheroni, WJr, Aguilar-Vildoso, CI, Barroso, PA, Saridakis, HO & Azevedo, JL 2001, “Variability and Interactions Between Endophytic Bacteria and Fungi Isolated From Leaf Tissues of Citrus Rootstocks”, Canadian Journal of Microbiology, vol. 47, no. 3, pp. 229-236.
Arnold, AE 2007, “Understanding the Diversity of Foliar Endophytic Fungi: Progress, Challenges, and Frontiers”, Fungal Biology Reviews, vol. 21, no. 2-3, pp. 51-66.
Ashish, B, Neeti, K & Himanshu, K 2013, “Copper Toxicity: A Comprehensive Study”, Research Journal of Recent Sciences, Vol. 2, pp. 58-67.

- 95 -
Awasthy, D, Ambady, A, Bhat, J, Sheikh, G, Ravishankar, S, Subbulakshmi, V, Mukherjee, K, Sambandamurthy, V & Sharma, U 2010, “Essentiality and Functional Analysis of Type I and Type III Pantothenate Kinases of Mycobacterium tuberculosis”, Microbiology, vol. 156, pp. 2691–2701.
Azevedo, MM, Carvalho, A, Pascoal, C, Rodrigues, F & Cássio, F 2007, “Responses of Antioxidant Defenses to Cu and Zn Stress in Two Aquatic Fungi”, Science of The Total Environment, vol. 377, no. 2–3, pp. 233–243.
Bacon, CW & Hill, NS 1996, “Symptomless Grass Endophytes: Products of Coevolutionary Symbioses and Their Role in the Ecological Adaptations of Grasses”, in SC Redkin & LM Carris (eds), Endophytic Fungi in Grasses and Woody Plants, St. Paul, APS Press, pp. 155–178.
Ban, Y, Tang, M, Chen, H, Xu, Z, Zhang, H & Yang, Y 2012, “The Response of Dark Septate Endophytes (DSE) to Heavy Metals in Pure Culture”, PLoS One, vol. 7, no. 10.
Barakat, MA 2011, “New Trends in Removing Heavy Metals From Industrial Wastewater Arabian”, Journal of Chemistry, vol. 4, no. 4, pp. 361–377.
Barakat, MA, Chen, YT & Huang, CP 2004, “Removal of Toxic Cyanide and Cu(II) Ions From Water by Illuminated TiO2 Catalyst,” Applied Catalysis B: Environmental, vol 53, no. 1, pp. 13–20.
Bayman, P, Angulo-sandoval, P, Baez-ortiz, Z & Lodge, DJ 1998, “Distribution and Dispersal of Xylaria endophytes in Two Tree Species in Puerto Rico”, Mycological Research, vol. 102, no. 8, pp. 944–948.
Baysal, A, Ozbek, N & Akman, S 2013, Determination of Trace Metals in Waste Water and Their Removal Processes, INTECH Open Access Publisher.
Becker, J & Craig, EA 1994, “Heat-shock Proteins as Molecular Chaperones”, European Journal of Biochemistry, vol. 219, no. 1-2, pp. 11–23.
Bellion, M, Courbot, M, Jacob, C, Guinet, F, Blaudez, D & Chalot, M. 2007, “Metal Induction of a Paxillus involutus metallothionein and its heterologous expression in Hebeloma cylindrosporum”, New Phytologist, vol. 174, no. 1, pp. 151-8.
Benabdellah, K, Azcón-Aguilar, C, Valderas, A, Speziga, D, Fitzpatrick, TB & Ferrol, N 2009, “GintPDX1 Encodes a Protein Involved in Vitamin B6 Biosynthesis That is Up-regulated by Oxidative Stress in the Arbuscular Mycorrhizal Fungus Glomus intraradices”, New Phytologist, vol. 184, no. 3, pp. 682-93.
Benito, Y & Ruíz, ML 2002, “Reverse Osmosis Applied to Metal Finishing Wastewater”, Desalination, vol. 142, no. 3, pp. 229–234.
Bennet, H 1986, Concise Chemical and Technical Dictionary, 4th enlarged ed., Edward Arnold, London.
Beveridge, TJ 1986, “The Immobilization of Soluble Metals by Bacterial Walls”, in HL Ehrlich & DS Holmes (eds), Biotechnology and Bioengineering Symposium No.

- 96 -
16: Biotechnology for the Mining, Metal-Refining, and Fossil Fuel Processing Industries, J. Wiley Interscience: New York, pp. 127- 140.
Bilski, P, Li, MY, Ehrenshaft, M, Daub, ME & Chignell, CF 2000, “Vitamin B6 (pyridoxine) and its Derivatives are Efficient Singlet Oxygen Quenchers and Potential Fungal Antioxidants”, Photochemistry and Photobiology, vol. 71, no. 2, pp. 129-34.
Bishnoi, NR & Garima, A 2005, “Fungus-an Alternative for Bioremediation of Heavy Metal Containing Wastewater: a review”, Journal of Science & Industrial Research, vol. 64, pp. 93-100.
Boiteux, S, Gajewski, E, Laval, J & Dizdaroglu, M 1992, “Substrate Specificity of the Escherichia coli Fpg Protein Formamidopyrimidine-DNA Glycosylase: Excision of Purine Lesions in DNA Produced by Ionizing Radiation or Photosensitization”, Biochemistry, vol. 31, no. 1, pp. 106–110.
Borges, JC & Ramos, CH 2005, “Protein Folding Assisted by Chaperones”, Protein and Peptide Letters, vol. 12, no. 3, pp. 257-61.
Bradford, MM 1976, “A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-dye Binding”, Analytical Biochemistry, vol. 72, no. 1–2, pp. 248–254.
Cagney, G & Emili, A 2002, “De novo Peptide Sequencing and Quantitative Profiling of Complex Protein Mixtures Using Mass-coded Abundance Tagging”, Nature biotechnology, vol. 20, no.2, pp. 163-170.
Caldas, T, Binet, E, Bouloc, P, Costa, A, Desgres, J & Richarme, G 2000, “The FtsJ/RrmJ Heat Shock Protein of Escherichia coli is a 23 S Ribosomal RNA Methyltransferase”, The Journal of Biological Chemistry, vol. 275, no. 22, pp. 16414-16419.
Carnivorous Plants / Insectivorous Plants in the Wilderness, Viewed 5th February 2015, http://www.honda-e.com/A02_World%20Maps/WorldMap_NepenDetail.htm.
Carrillo-González, R & González-Chávez, MC 2012, “Tolerance to and Accumulation of Cadmium by the Mycelium of the Fungi Scleroderma citrinum and Pisolithus tinctorius”, Biological Trace Element Research, vol. 146, no. 3, pp. 388-395.
Casati, P, Drincovich, MF, Edwards, GE & Andreo, CS 1999, “Malate Metabolism by NADP-malic Enzyme in Plant Defense”, Photosynthesis Research, vol. 61, no. 2, pp. 99-105.
Chandler, JD & Day, BJ 2012, “Thiocyanate: A Potentially Useful Therapeutic Agent With Host Defense and Antioxidant Properties”, Biochemical Pharmacology, vol. 84, no. 11, pp. 1381-7.
Chang, SC, Chen, MK, Wei, DL & Cheng, MK 2007, “Isolation and Characterization of A Polycyclic Aromatic Hydrocarbons-degrading Enzyme From Xylaria regalis”, Fungal Science, vol. 22, no. 1, 2, pp. 25–33.

- 97 -
Chen, J, Zhang, LC, Xing, YM, Wang, YQ, Xing, XK, Zhang, DW, Liang, HQ & Guo, SX 2013, “Diversity and Taxonomy of Endophytic Xylariaceous Fungi from Medicinal Plants of Dendrobium (Orchidaceae)”, PLoS One, vol. 8, no. 3.
Cho, DH, Kim, EY & Hung, YT 2010, “Heavy Metal Removal by Microbial Biosorbents”, Environmental Bioengineering, Handbook of Environmental Engineering, vol. 11, pp. 375-402.
Cipollone, R, Ascenzi, P, Tomao, P, Imperi, F & Visca, P 2008, “Enzymatic Detoxification of Cyanide: Clues From Pseudomonas aeruginosa Rhodanese”, Journal of Molecular Microbiology and Biotechnology, vol. 15, no. 2-3, pp. 199-211.
Clark, EM, White, JF & Patterson, RM 1983, “Improved Histochemical Techniques for the Detection of Acremonium coenophialum in Tall Fescue and Methods of in vitro Culture of the Fungus”, Journal of Microbiological Methods, vol. 1, no. 3, pp. 149-155.
Clarke, C 1997, Nepenthes of Borneo, Natural History Publications (Borneo) Sdn. Bhd., Kota Kinabalu.
Csermely, P & Yahara, I 2003, "Heat Shock Proteins." Molecular Pathomechanisms and New Trends in Drug Research, Taylor and Francis, London, pp. 67-75.
Cushnie, GC 1985, Electroplating wastewater pollution control technology, Noyes Publications, Park Ridge, N.J., U.S.A.
Da̧browski, A, Hubickia, Z, Podkościelnya, P & Robensb, E 2004, “Selective Removal of the Heavy Metal Ions From Waters and Industrial Wastewaters by Ion-exchange Method”, Chemosphere, vol. 56, no. 2, pp. 91–106.
Davies, BE & Jones, LHP 1988, “Micronutrients and TExic elements,” in A Wild (eds), Russell's Soil Conditions and Plant Growth, John Wiley & Sons; Interscience, New York, NY, USA, 11th edn., pp. 781–814.
Davis, RA, Welty, AT, Borrego, J, Morales, JA, Pendon, JG & Ryan, JG 2000, “Rio Tinto estuary (Spain): 5000 years of pollution Environmental”, Geology, Vol. 39, no. 10, pp. 1107-1116.
Deng, Z, Cao, L, Huang, H, Jiang, X, Wang, W, Shi, Y & Zhang, R 2011, “Characterization of Cd- and Pb-resistant Fungal Endophyte Mucor sp. CBRF59 Isolated From Rapes (Brassica chinensis) In a Metal-contaminated Soil”, Journal of Hazardous Materials, vol. 185, no. 2–3, pp. 717–724.
Deng, Z, Zhang, R, Shi, Y, Hu, L, Tan, H & Cao, L 2014, “Characterization of Cd-, Pb-, Zn-resistant Endophytic Lasiodiplodia sp. MXSF31 From Metal Accumulating Portulaca oleracea and Its Potential in Promoting the Growth of Rape in Metal-contaminated Soils”, Environmental Science and Pollution Research, vol. 21, no. 3, pp. 2346-57.

- 98 -
Dönmez, GC, Aksu, Z, Öztürk, A & Kutsal, T 1999, “A Comparative Study on Heavy Metal Biosorption Characteristics of Some Algae”, Process Biochemistry, vol. 34, no. 9, pp. 885–892.
Dyson, P, Ransing, R, Williams, PH & Williams, R 2008, Fluid Properties at Nano/Meso Scale: A Numerical Treatment, John Wiley & Sons.
Ehrenshaft, M, Bilski, P, Li, MY, Chignell, CF & Daub, ME 1999, “A Highly Conserved Sequence is A Novel Gene Involved in De novo Vitamin B6 Biosynthesis”, Proceedings of the National Academy of Sciences U S A. vol. 96, no. 16, pp. 9374-8.
Eife, R, Weiss, M, Barros, V, Sigmund, B, Goriup, U, Komb, D, Wolf, W, Kittel, J, Schramel, P & Reiter, K 1999, “Chronic Poisoning by Copper in Tap Water: I. Copper Intoxications With Predominantly Gastointestinal Symptoms”, European Journal of Medical Research, vol. 4, no. 6, pp. 219-23.
Elbersen, HW & West, CP 1996, “Growth and Water Relations of Field‐Grown Tall Fescue as Influenced by Drought and Endophyte”, Grass and Forage Science, vol. 51, no. 4, pp. 333-342.
El-Gendy, MMA, Hassanein, NM, El-Hay, Ibrahim, HA & El-Baky, DAH 2011, “Evaluation of Some Fungal Endophytes of Plant Potentiality as Low-cost Adsorbents for Heavy Metals Uptake from Aqueous Solution”, Australian Journal of Basic and Applied Sciences, vol. 5, no. 7, pp. 466-473.
Elifantz, H & Tel-Or, E 2002, “Heavy Metal Biosorption by Plant Biomass of the Macrophyte Ludwigia Stolonifera”, Water, Air, and Soil Pollution, vol. 141, no. 1-4, pp 207-218.
Elseviers, D, Petrullo, LA & Gallagher, PJ 1984, “Novel E. coli Mutants Deficient in Blosynthesis of 5-methylamlnomethyl-2-thiouridine”, Nucleic Acids Research, vol. 12, no. 8, pp. 3521-3534.
Erakhrumen & Agbontalor, A 2007, “Phytoremediation: An Environmentally Sound Technology for Pollution Prevention, Control and Remediation in Developing Countries”, Educational Research and Review, vol. 2, no. 7, pp. 151-156.
Ferguson, KD & Erickson, PM 1988, “Pre-Mine Prediction of Acid Mine Drainage”, in W Salomons & U Forstner (eds), Dredged Material and Mine Tailings, Springer-Verlag Berlin Heidelberg.
Ferreira, S, Hjernø, K, Larsen, M, Wingsle, G, Larsen, P, Fey, S, Roepstorff, P & Salomé, PM 2006, “Proteome Profiling of Populus euphratica Oliv. upon heat stress”, Annals of Botany, vol. 98, no. 2, pp. 361-377.
Fourest, E & Roux, J 1992, “Heavy Metal Biosorption by Fungal Mycelial By-Products: Mechanisms and Influence of pH”, Applied Microbiology and Biotechnology, vol. 37, no. 3, pp. 399-403.
Fulekar, MH 2012, Bioremediation Technology: Recent Advances, Springer Science & Business Media.

- 99 -
Gadd, GM 2004, “Microbial Influence on Metal Mobility and Application for Bioremediation”, Geoderma, vol. 122, no. 2–4, pp. 109–119.
Gallo, MB, Chagas, FO, Almeida, MO, Macedo, CC, Cavalcanti, BC, Barros, FW, de Moraes, MO, Costa-Lotufo, LV, Pessoa, C, Bastos, JK & Pupo, MT 2009, “Endophytic Fungi Found in Association With Smallanthus Sonchifolius (Asteraceae) as Resourceful Producers of Cytotoxic Bioactive Natural Products”, Journal of Basic Microbiology, vol. 49, no. 2, pp. 142-51.
Gamakaranage, CSSK, Rodrigo, C, Weerasinghe, S, Gnanathasan, A, Puvanaraj, V & Fernando, H 2011, “Complications and Management of Acute Copper Sulphate Poisoning; A Case Discussion”, Journal of Occupational Medicine and Toxicology, vol. 6.
Gaspard, S & Ncibi, MC 2013, Biomass for Sustainable Applications: Pollution Remediation and Energy, Royal Society of Chemistry.
Gaume, L, Gorb, S & Rowe, N 2002, “Function of Epidermal Surfaces in the Trapping Efficiency of Nepenthes alata Pitchers”, New Phytologist, vol. 156, pp. 479–489.
Gaur, A & Adholeya, A 2004, “Prospects of Arbuscular Mycorrhizal Fungi in Phytoremediation of Heavy Metal Contaminated Soils”, Current Science, vol. 86, no. 4, pp. 528-534.
Ge, W, Zamri, D, Mineyama, H & Valix, M 2011, “Bioaccumulation of Heavy Metals on Adapted Aspergillus foetidus”, Adsorption, vol. 17, no. 5, pp. 901-910.
Geen, A, Adkins, JF, Boyle, EA, Nelson CH & Palanques, A 1997, “A 120-yr Record of Widespread Contamination From Mining of the Iberian Pyrite Belt”, Geology, vol. 25, no. 4, pp. 291–294.
Gherbawy, Y & Voigt, K 2010, Molecular Identification of Fungi, Springer Science & Business Media.
Gold, H, Czupryna, G, Levy, RD, Calmon, C & Gross, RL 1987, “Purifying Plating Baths by Chelate Ion Exchange”, Metals Speciation, Separation, and Recovery, Lewis Publishers, Chelsea Michigan, pp. 619-642.
Gong, S, Ma, Z & Foster, JW 2004, “The Era-like GTPase TrmE conditionally activates gadE and glutamate-dependent acid resistance in Escherichia coli”, Molecular Microbiology, vol. 54, pp. 948-61.
González-Guerrero, M, Benabdellah, K, Valderas, A, Azcón-Aguilar, C & Ferrol, N 2010, “GintABC1 Encodes a Putative ABC Transporter of the MRP Subfamily Induced by Cu, Cd, and Oxidative Stress in Glomus intraradices”, Mycorrhiza, vol. 20, no. 2, pp. 137-46.
Gorb, E, Kastner, V, Peressadko, A, Arzt, E, Gaume, L, Rowe, N & Gorb, S 2004, “Structure and Properties of the Glandular Surfaces in the Digestive Zone of the Pitcher in the Carnivorous Plant Nepenthes ventrata and Its Role in Insect Trapping and Retention”, The Journal of Experimental Biology, vol. 207, pp. 2947–2963.

- 100 -
Graumann, P, Schröder, K, Schmid, R & Marahiel, MA 1996, “Cold Shock Stress-induced Proteins in Bacillus subtilis”, Journal of Bacteriology, vol. 178, no. 15, pp. 4611-4619.
Griffin, RC 2011, Water Policy in Texas: Responding to the Rise of Scarcity, RFF Press, Washington, DC.
Hadi, S & Nuhamara, ST 1997, “Diseases of Species and Provenances of Acacias in West and South Kalimantan, Indonesia”, in KM Old, SS Lee & JK Sharma (eds), Diseases of Tropical Acaclas. South Sumatra CIFOR Special Publication, Bogor, pp 23-26
Hallmann, J, Quadt-Hallmann, A, Mahaffee, WF & Kloepper, JW 1997, “Bacterial Endophytes in Agricultural Crops”, Canadian Journal of Microbiology, vol. 43, no.10, pp. 895-914.
Han, NMM, Latif, MT, Othman, M, Dominick, D, Mohamad, N, Juahir, H & Tahir, NM 2014, “Composition of Selected Heavy Metals in Road Dust From Kuala Lumpur City Centre”, Environmental Earth Sciences, vol. 72, no. 3, pp. 849-859.
Hansen, RE, Roth, D & Winther, JR. 2009, “Quantifying the Global Cellular Thiol-disulfide Status”, Proceedings of the National Academy of Sciences USA, vol. 106, no. 2, pp.422-7.
Harada, M 1995, “Minamata disease: Methylmercury Poisoning in Japan Caused by Environmental Pollution”, Critical Reviews in Toxicology, vol. 25, no. 1, pp. 1-24.
He, ZL, Yang, XE & Stoffella, PJ 2005, “Trace Elements in Agroecosystems and Impacts on the Environment”, Journal of Trace Elements in Medicine and Biology, vol. 19, no. 2-3, pp. 125-40.
Hegedűs, N, Emri, T, Szilágyi, J, Karányi, Z, Nagy, I, Penninckx, MJ & Pócsi I 2007, “Effect of Heavy Metals on the Glutathione Status in Different Ectomycorrhizal Paxillus involutus Strains”, World Journal of Microbiology and Biotechnology, vol. 23, no. 9, pp. 1339-1343.
Helander, MOL 1995, “Responses of Pine Needle Endophytes to Air Pollution”, New Phytologist, vol. 131, pp. 223-229.
Herrmann, JM 1999, “Heterogeneous Photocatalysis: Fundamentals and Applications to the Removal of Various Types of Aqueous Pollutants”, Catalysis Today, vol. 53, no. 1, pp. 115–129.
Hettler, J, Irion, G & Lehmann, B 1997, “Environmental Impact of Mining Waste Disposal on a Tropical Lowland River System: A Case Study on the Ok Tedi Mine, Papua New Guinea”, Mineralium Deposita, vol. 32, pp. 280-291.
Holan, ZR & Volesky, B 1994, “Biosorption of Lead and Nickel by Biomass of Marine Algae”, Biotechnology and Bioengineering, vol. 43, pp. 1001- 1009.
Holan, ZR & Volesky, B 1995, “Accumulation of Cadmium, Lead and Nickel by Fungal and Wood Biosorbents”, Applied Biochemistry and Biotechnology, vol. 53, no. 2, pp. 133-146.

- 101 -
Honzatko, RB & Fromm, HJ 1999, “Structure–Function Studies of Adenylosuccinate Synthetase from Escherichia coli”, Archives of Biochemistry and Biophysics, vol. 370, no. 1, pp. 1–8.
Horecker, BL & Stadtman, ER 2014, Current Topics in Cellular Regulation, Elsevier, vol. 22.
Hua, Y & Li, H 2005, “Food Web and Fluid in Pitchers of Nepenthes mirabilis in Zhuhai, China”, Acta Botanica Gallica, vol. 152, no. 2, pp. 165-175.
Huhndorf, SM, Miller, AN & Ferna´ndez, FA 2004, “Molecular Systematics of the Sordariales: the Order and the Family Lasiosphaeriaceae Redefined”, Mycologia, vol. 96, no. 2, pp. 368–387.
Huszcza, E, Bartmańska, A, Anioł, M, Mączka, W, Żołnierczyk, A & Wawrzeńczyk, C 2008, “Degradation of Hop Bitter Acids by Fungi”, Waste Management, vol. 28, no. 8, pp. 1406–1410.
Ilyas, S & Rehman, A 2013, “Decolorization and Detoxification of Synozol Red HF-6BN Azo Dye, by Aspergillus niger and Nigrospora sp.”, Iranian Journal of Environmental Health Sciences & Engineering, vol. 10.
International Institute for Environment and Development (IIED) 2002, “Ok Tedi Riverine. Disposal Case Study”, Mining, Minerals and Sustainable Development, viewed 3rd January, http://pubs.iied.org/pdfs/G00561.pdf.
Inthe, MG, Tarman, K & Safithri, M 2014, “Fractionation of Proteins of Marine Fungus Xylaria psidii KT30 and Their Cytotoxicity Against HeLa cells”, Jurnal Teknologi dan Industri Pangan, vol. 25, no. 1, pp. 39-46.
Iskandar, NL, Zainudin, NAIM & Tan, SG 2011, “Tolerance and Biosorption of Copper (Cu) and Lead (Pb) by Filamentous Fungi Isolated from a Freshwater Ecosystem”, Journal of Environmental Sciences, vol. 23, no.5, pp. 824-830.
Ismail, A & Safahieh, A 2005, “Copper and Zinc in Intertidal Surface Sediment and Telescopium telescopium from Lukut River, Malaysia”, Coastal marine science, vo. 29, no. 2, pp. 111-115.
Iyer, RR 1990, Hazardous waste treatment technologies. Gerald Rich and Kenneth Cherry, Pudvan Publishing Co. Inc., Northbrook, IL.
Jadia, CD & Fulekar, MH 2009, “Phytoremediation of Heavy Metals: Recent Techniques,” African Journal of Biotechnology, vol. 8, no. 6, pp. 921-928.
Javaid, A, Bajwa, R & Manzoor, T 2011, “Biosorption of Heavy Metals by Pretreated Biomass of Aspergillus niger”, Pakistan Journal of Botany, vol. 43, no. 1, pp. 419-425.
Jennings, GD & Sneed, RE 1996, “Nitrate in Drinking Water. North Carolina Cooperative Extension Service”, viewed 5th January, http://www.bae.ncsu.edu/extension/ext-publications/water/drinking/ag-473-4-nitrate-jennings.pdf.

- 102 -
Jensen, ON 2006, “Interpreting the Protein Language Using Proteomics”, Nature Reviews Molecular Cell Biology, vol. 7, no. 6, pp. 391-403.
Jo, WJ, Loguinov, A, Chang, M, Wintz, H, Nislow, C, Arkin, AP, Giaever, G & Vulpe, CD 2008, “Identification of Genes Involved in the Toxic Response of Saccharomyces cerevisiae Against Iron and Copper Overload by Parallel Analysis of Deletion Mutants”, Toxicological Science, vol. 101, no. 1, pp. 140-51.
Johnson, DB & Hallberg, KB 2005, “Acid Mine Drainage Remediation Options: A Review”, Science of the Total Environment, vol. 338, pp. 3-14.
Jones, EBG & Pang, KL 2012, Marine Fungi: and Fungal-like Organisms, Marine and Freshwater Botany, Walter de Gruyter.
Ju, YM & Hsieh, HM 2007, “Xylaria species Associated With Nests of Odontotermes formosanus in Taiwan”, Mycologia, vol. 99, no. 6, pp. 936–957.
Kahraman, S, Asma (Hamamci), D, Erdemoglu, S & Yesilada, O 2005, “Biosorption of Copper(II) by Live and Dried Biomass of the White Rot Fungi Phanerochaete chrysosporium and Funalia trogii”, Engineering in Life Sciences, vol. 5, pp. 72–77.
Kajitvichyanukula, P, Ananpattarachaia, J & Pongpomb, S 2005, “Sol–gel Preparation and Properties Study of TiO2 Thin Film for Photocatalytic Reduction of Chromium(VI) in Photocatalysis Process”, Science and Technology of Advanced Materials, vol. 6, no. 3–4, pp. 352–358.
Khan, AL & Lee, IJ 2013, “Endophytic Penicillium funiculosum LHL06 Secretes Gibberellin That Reprograms Glycine max L. Growth During Copper Stress”, BMC Plant Biology, vol. 13.
Khan, AL, Hamayun, M, Ahmad, N, Hussain, J, Kang, SM, Kim, YH, Adnan, M, Tang, DS, Waqas, M, Radhakrishnan, R, Hwang, YH & Lee, IJ. 2011, “Salinity Stress Resistance Offered by Endophytic Fungal Interaction between Penicillium minioluteum LHL09 and Glycine max. L.”, Journal of Microbiology and Biotechnology, vol. 21, no. 9, pp. 893-902.
Kim, AG, Heisey, BS, Kleinmann, RL & Deul M 1982, “Acid Mine Drainage: Control and Abatement Research”, Available from the National Technical Information Service, Springfield, VA.
Kirpichtchikova, TA, Manceau, A, Spadini, L, Panfili, F, Marcus, MA & Jacquet, T 2006 “Speciation and Solubility of Heavy Metals in Contaminated Soil Using X-ray Microfluorescence, EXAFS Spectroscopy, Chemical Extraction, and Thermodynamic Modeling”, Geochimica et Cosmochimica Acta, vol. 70, no. 9, pp. 2163–2190.
Kohanski, MA, Dwyer, DJ & Collins, JJ 2010, “How Antibiotics Kill Bacteria: From Targets to Networks”, Nature Reviews Microbiology, vol. 6, pp. 423-435.
Kratochvil, D & Volesky, B 1998, “Advances in Biosorption of Heavy Metals”, Trends in Biotechnology, vol. 16, pp. 291-300.

- 103 -
Kreisel, H, & Schauer, F 1987, Methoden Des Mykologischen Laboratoriums, Gustav Fischer Verlag.
Krings, M, Taylor, TN, Hass, H, Kerp, H, Dotzler, N & Hermsen, EJ 2007, “Fungal Endophytes in a 400-million-yr-old Land Plant: Infection Pathways, Spatial Distribution, and Host Responses”, New Phytologist, vol. 174, no. 3, pp. 648-57.
Krüger, MK & Sørensen, MA 1998, “Aminoacylation of Hypomodified tRNAGlu in vivo”, Journal of Molecular Biology, vol. 284, no. 3, pp. 609-20.
Kumar, DSS & Hyde, KD 2004, “Biodiversity and Tissue-recurrence of Endophytic Fungi in Tripterygium wilfordii”, Fungal Diversity, vol. 17, pp. 69-90.
Kurniawana, TA, Chana, GYS, Loa, WH & Babel, S 2006, “Physico–chemical Treatment Techniques for Wastewater Laden With Heavy Metals”, Chemical Engineering Journal, vol. 118, no. 1–2, pp. 83–98.
Kurup, R, Johnson, AJ, Sankar, S, Hussain, AA, Kumar CS & Sabulal, B 2013, “Fluorescent Prey Traps in Carnivorous Plants”, Plant Biology, vol. 15, no. 3, pp. 611–615.
Kuyucak, N & Volesky, B 1988a, “Biosorbents for Recovery of Metals from Industrial Solutions”, Biotechnology Letters, vol. 10, no. 2, pp. 137-142.
Kuyucak, N & Volesky, B 1988b, “New Algal Biosorbent for a Gold Recovery Process”, in PR Norris, DP Kelly, (eds), BioHydroMetallurgy: Proceedings of the International Symposium, Sci. Technol. Letters: Kew, Surrey, U.K., pp. 453-464.
Lambers, H, & Colmer, TD 2006, Root Physiology: from Gene to Function, volume 4 of Plant Ecophysiology, Springer Science & Business Media.
Lapakko, K 1993, “Mine Waste Drainage Quality Prediction: A Literature Review”, Draft Paper, Minnesota Department of Natural Resources, Division of Minerals, St. Paul, MN.
Lee, G 2014, Environment and Sustainability, WIT Press.
Lee, JC, Lobkovsky, E, Pliam, NB, Strobel, G & Clardy, J 1995, “Subglutinols A and B: Immunosuppressive Compounds from the Endophytic Fungus Fusarium subglutinans”, The Journal of Organic Chemistry, vol. 60, no. 22, pp. 7076–7077.
Lee, JM, Tan, WS & Ting, ASY 2014, “Revealing the Antimicrobial and Enzymatic Potentials of Culturable Fungal Endophytes from Tropical Pitcher Plants (Nepenthes spp.)”, Mycosphere, vol. 5, no. 2, pp. 364–377.
Lester, JN 1987, Heavy Metals in Wastewater and Sludge Treatment Processes, CRC Press Inc., Boca Raton, FL, United States.
Letelier, ME, Lepe, AM, Faúndez, M, Salazar, J, Marín, R, Aracena, P & Speisky, H 2005, “Possible Mechanisms Underlying Copper-induced Damage in Biological Membranes Leading to Cellular Toxicity”, Chemico-biological interactions, vol. 151, no. 2, pp. 71-82.

- 104 -
Li, JY, Harper, JK, Grant, DM, Tombe, BO, Bashyal, B, Hess, WM & Strobel, GA 2001, “Ambuic Acid, A Highly Functionalized Cyclohexenone With Antifungal Activity from Pestalotiopsis spp. and Monochaetia sp.”, Phytochemistry, vol. 56, no. 5, pp. 463-8.
Li, JY, Strobel, G, Harper, J, Lobkovsky, E & Clardy, J 2000, “Cryptocin, A Potent Tetramic Acid Antimycotic from the Endophytic Fungus Cryptosporiopsis cf. quercina”, Organic Letters, vol. 2, no. 6, pp. 767-70.
Li, JY, Strobel, G, Sidhu, R, Hess, WM & Ford, EJ 1996, “Endophytic Taxol-Producing Fungi from Bald Cypress, Taxodium distichum”, Microbiology, vol. 142, no. 8, pp. 2223-2226.
Liers, C, Ullrich, R, Pecyna, M, Schlosser, D & Hofrichter, M 2007, “Production, Purification and Partial Enzymatic and Molecular Characterization of A Laccase from the Wood-rotting Ascomycete Xylaria polymorpha”, Enzyme and Microbial Technology”, vol. 41, no. 6–7, pp. 785–793.
Liers, C, Ullrich, R, Steffen, KT, Hatakka, A & Hofrichter, M 2006, “Mineralization of 14C-labelled Synthetic Lignin and Extracellular Enzyme Activities of the Wood-colonizing Ascomycetes Xylaria hypoxylon and Xylaria polymorpha”, Applied Microbiology and Biotechnology, vol. 69, no. 5, pp. 573-579.
Lim, PE & Kiu, MY 1995, “Determination and Speciation of Heavy Metals in Sediments of the Juru River, Penang, Malaysia”, Environmental Monitoring and Assessment, vol. 35, no. 2, pp. 85-95.
Linder, MC 2012, “The Relationship of Copper to DNA Damage and Damage Prevention in Humans”, Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, vol. 733, no. 1–2, pp. 83–91.
Lloyd, FE 1942, The Carnivorous Plants. Chronica Botanica 9, Ronald Press Company, New York, U.S.A.
Malato, S, Maldonado, MI, Fernández, P, Oller, I & Polo, I 2014, “Decontamination of Water by Solar Irradiation”, Advanced Oxidation Technologies: Sustainable Solutions for Environmental Treatments, pp. 1–22.
Malinowski, D, Leuchtmann, A, Schmidt, D & Nsberger, J. 1997, “Growth and Water Status in Meadow Fescue (Festuca pratensis) is Differently Affected by Its Two Natural Endophytes”, Agronomy Journal, vol. 89, pp. 673-678.
Maria, GL, Sridhar, KR & Raviraja, NS 2005, “Antimicrobial and Enzyme Activity of Mangrove Endophytic Fungi of Southwest Coast of India”, Journal of Agricultural Technology, vol. 1, pp. 67-80.
Martínez-Juárez VM, Cárdenas-González JF, Torre-Bouscoulet ME & Acosta-Rodríguez I 2012, “Biosorption of Mercury (II) from Aqueous Solutions onto Fungal Biomass”, Bioinorganic Chemistry and Applications, vol. 2012.

- 105 -
Maslin, P & Maier, RM 2000, “Rhamnolipid-enhanced Mineralization of Phenanthrene in Organic-metal Co-contaminated Soils”, Bioremediation Journal, vol. 4, no. 4, pp. 295–308.
Mason, B & Moore, C 1982, Principles of Geochemistry. 4th edn. John Wiley & Sons, New York.
Mattei, AS, Severo, CB, Guazzelli, LS, Oliveira, FM, Gené, J, Guarro, J, Cano, J & Severo LC 2013, “Cutaneous Infection by Diaporthe phaseolorum in Brazil”, Medical Mycology Case Reports, vol. 2, pp. 85-7.
Mattuschka, B, Junghaus, K & Straube, G 1993 “Biosorption of Metals by Waste Biomass,” in AE Torma, ML Apel, & CLBrierley (eds), Biohydrometallurgical Technologies, The Minerals, Metals & Materials Society: Warrendale, PA, vol. 2, pp 125-132.
McCarthy, TS 2011, “The Impact of Acid Mine Drainage in South Africa”, South African Journal of Science, vol. 107, no. 5/6.
McPherson, S., 2009, Pitcher Plants of the Old World, Redfern Natural History Productions, Poole.
Méchin, V, Damerval, C & Zivy M 2007, “Total Protein Extraction with TCA-Acetone. Plant Proteomics”, Methods in Molecular Biology, vol. 355, pp. 1-8.
Merbach, MA, Zizka, G, Fiala, B, Maschwitz, U & Booth WE 2001, “Patterns of Nectar Secretion in Five Nepenthes species from Brunei Darussalam, Northwest Borneo, and Implications for Ant-plant Relationships”, Flora, vol. 196, pp. 153-160.
Meyer, S, Wittinghofer, A & Versées, W 2009, “G-Domain Dimerization Orchestrates the tRNA Wobble Modification Reaction in the MnmE/GidA Complex”, Journal of Molecular Biology, vol. 392, no. 4, pp. 910–922.
Miller, CM, Miller, RV, Garton-Kenny, D, Redgrave, B, Sears, J, Condron, MM, Teplow, DB & Strobel GA 1998, “Ecomycins, Unique Antimycotics from Pseudomonas viridiflava”, Journal of Applied Microbiology, vol. 84, no. 6, pp. 937-44.
Mirsal, I 2013, Soil Pollution: Origin, Monitoring & Remediation, Springer Science & Business Media.
Mithöfer, A 2011, “Carnivorous Plants: Insights in an Old Topic”, Phytochemistry, vol. 72, pp. 1678-1682.
Mittenhube, G 2001, “Phylogenetic Analyses and Comparative Genomics of Vitamin B6 (Pyridoxine) and Pyridoxal Phosphate Biosynthesis Pathways”, Journal of Molecular Microbiology and Biotechnology, vol. 3, no. 1, pp. 1-20.
Mohammadi, T, Moheb, A, Sadrzadeh, M & Razmi, A 2005, “Modeling of Metal Ion Removal from Wastewater by Electrodialysis”, Separation and Purification Technology, vol. 41, no. 1, pp. 73–82.

- 106 -
Mol, CD, Kuo, CF, Thayer, MM, Cunningham, RP & Tainer, JA 1995, “Structure and Function of the Multifunctional DNA-repair Enzyme Exonuclease III”, Nature, vol. 374, no. 6520, pp. 381-6.
Monnet, F, Vaillant, N, Hitmi, A, Coudret, A & Sallanon, H 2001, “Endophytic Neotyphodium lolii Induced Tolerance to Zn Stress in Lolium perenne”, Physiologia Plantarum, vol. 113, no. 4, pp. 557-563.
Muraleedharan, TR & Venkobachar, C 1990, “Mechanism of Biosorption of Copper(II) by Ganoderma iucidum”, Biotechnology and Bioengineering, vol. 35, no. 3, pp. 320–325.
Nakasone, KK, Peterson, SW & Jong, SC 2004, “Preservation and Distribution of Fungal Cultures”, Biodiversity of Fungi, pp. 37-47.
NCBIa ACLA_095590 mitochondrial GTPase (Mss1), putative [Aspergillus clavatus NRRL 1].
Ngai, KL 2011, Relaxation and Diffusion in Complex Systems, Springer Science & Business Media.
Nicely, NI, Parsonage, D, Paige, C, Newton, GL, Fahey, RC, Leonardi, R, Jackowski, S, Mallett, TC & Claiborne, A 2007, “Structure of the Type III Pantothenate Kinase from Bacillus anthracis at 2.0 Å Resolution: Implications for Coenzyme A-Dependent Redox Biology”, Biochemistry, vol. 46, no. 11, pp. 3234–3245.
Nriagu, JO & Pacynat, JM 1988, “Quantitative Assessment of Worldwide Contamination of Air, Water and Soils by Trace Metals”, Nature, vol. 333, pp. 134-139.
O'Farrell, PH 1975, “High Resolution Two-dimensional Electrophoresis of Proteins”, The Journal of Biological Chemistry, vol. 250, no. 10, pp.4007-21.
Okafor, CC & Onwuka, SU 2013, “Leachate Pollution of Soil of Enugu”, IOSR Journal of Environmental Science, Toxicology And Food Technology, vol. 5, no. 5, pp. 41-47.
Okamoto, S, Tamaru, A, Nakajima, C, Nishimura, K, Tanaka, Y, Tokuyama, S, Suzuki, Y & Ochi, K 2007, “Loss of A Conserved 7-methylguanosine modification in 16S rRNA Confers Low-level Streptomycin Resistance in Bacteria”, Molecular Microbiology, vol. 63, no. 4, pp. 1096-106.
Olade, MA 1987, “Heavy Metal Pollution and the Need for Monitoring: Illustrated for Developing Countries in West Africa”, in TC Hutchinson & KM Meema (eds), Lead, Mercury, Cadmium and Arsenic in the Environment, John Wiley & Sons Ltd., Chichester, UK, pp. 335-341.
Osmani, AH, May, GS, Osmani, SA 1999, “The Extremely Conserved pyroA Gene of Aspergillus nidulans is Required for Pyridoxine Synthesis and is Required Indirectly for Resistance to Photosensitizers”, The Journal of Biological Chemistry, vol. 274, no. 33, pp. 23565-9.

- 107 -
Ozcan, S, Yildirim, V, Kaya, L, Albrecht, D, Becher, D, Hecker, M & Ozcengiz, G 2007, “Phanerochaete chrysosporium Soluble Proteome as A Prelude for the Analysis of Heavy Metal Stress Response,” Proteomics, vol. 7, no. 8, pp. 1249-60.
Pandey, A & Mann, M 2000, “Proteomics to Study Genes and Genomes”, Nature, vol. 405, no. 6788, pp.837–846.
Patil, A, Patil, MS & Dangat, BT 2012, “Three Giant Ascomycetes (Pyrenomycetes) from Maharashtra, India”, Mycosphere, vol. 3, no. 3, pp. 353–356.
Pavoković, D, Križnik, B & Krsnik-Rasol, M 2012, “Evaluation of Protein Extraction Methods for Proteomic Analysis of Non-Model Recalcitrant Plant Tissues”, Croatica Chemica Acta, vol. 85, no. 2, pp. 177–183.
Pedersen, AJ 2003, “Characterization and Electrodialytic Treatment of Wood Combustion Fly Ash For the Removal of Cadmium”, Biomass and Bioenergy, vol. 25, no. 4, pp. 447–458.
Petrini, O & Dreyfuss, M 1981, “Endophytische Pilze in Epiphytischen Araceae, Bromeliaceae und Orchidaceae”, Sydowia, vol. 34, pp.135-148.
Phongpaichit, S, Nikom, J, Rungjindamai, N, Sakayaroj, J, Hutadilok-Towatana, N, Rukachaisirikul, V & Kirtikara, K. 2007, “Biological Activities of Extracts from Endophytic Fungi Isolated from Garcinia plants”, FEMS Immunology and Medical Microbiology, vol. 51, no. 3, pp. 517-25.
Pichat, P 2013, “Photocatalysis and Water Purification: From Fundamentals to Recent Applications”, New Materials for Sustainable Energy and Development, John Wiley & Sons.
Pivetz, BE 2001, “Phytoremediation of Contaminated Soil and Ground Water at Hazardous Waste Sites,” Ground Water Issue, Technical report, U.S. Environmental Protection Agency.
Pointing, SB, Parungao, MM & Hyde, KD 2003, “Production of Wood-decay Enzymes, Mass Loss and Lignin Solubilization in Wood by Tropical Xylariaceae”, Mycological Research, vol. 107, no. 2, pp. 231-235.
Pollak, N, Dölle, C & Ziegler, M 2007, “The Power to Reduce: Pyridine Nucleotides – Small Molecules With A Multitude of Functions”, Biochemical Journal, vol. 402, no. 2, pp. 205–218.
Prasad, MNV 2004, “Phytoremediation of Metals in the Environment for Sustainable Development,” Proceedings of the Indian National Science Academy, vol. 70, pp. 71-98.
Price, WA, & Errington, JC 1998, “Guidelines for Metal Leaching and Acid Rock Drainage at Minesites in British Columbia”, British Columbia Ministry of Energy, Mines and Petroleum Resources.
Pulford, ID & Watson, C 2003, “Phytoremediation of Heavy Metal-contaminated Land by Trees—A Review”, Environment International, vol. 29, no. 4, pp. 529–540.

- 108 -
Qdais, HA & Moussab, H 2004, “Removal of Heavy Metals from Wastewater by Membrane Processes: A Comparative Study”, Desalination, vol. 164, no. 2, pp. 105–110.
Rabilloud, T, Chevallet, M, Luche, S & Leize-Wagner, E 2005, “Oxidative Stress Response: A Proteomic View”, Expert Review of Proteomics, vol. 2, no. 6, pp. 949-956.
Rahim, BEA, Yusoff, I, Samsudin, AR, Yaacob, WZW & Rafek, AGM 2010, “Deterioration of Groundwater Quality in the Vicinity of an Active Open-tipping Site in West Malaysia”, Hydrogeology Journal, vol. 18, pp. 997-1006.
Rastogi, SC, Mendiratta, N & Rastogi, P 2006, “Bioinformatics Methods and Applications: Genomics, Proteomics and Drug Discovery”, Prentice-Hall of India (Private), Limited.
Ratnakumari, A & Sobha, K 2012, “Biosorption of Cu2+ Using Animal Polymers: Chick and Duck Feathers”, International Journal of Research in Pharmaceutical and Biomedical Sciences, vol. 3, no. 2, pp 664-669.
Ratnaweera, PB, Williams, DE, Silva, ED, Wijesundera, RLC, Dalisay, DS & Andersenc, RJ 2014, “Helvolic Acid, An Antibacterial Nortriterpenoid from A Fungal Endophyte, Xylaria sp. of Orchid Anoectochilus setaceus Endemic to Sri Lanka”, Mycology, vol. 5, no. 1, pp. 23–28.
Ravi, I, Baunthiyal, M & Saxena, J 2013, Advances in Biotechnology, Springer Science & Business Media.
Redman, RS, Dunigan, D, Rodriguez, RJ 2001, “Fungal Symbiosis from Mutualism to Parasitism: Who Controls the Outcome, Host or Invader?”, New Phytologist, vol. 151, pp. 705–716.
Redman, RS, Ranson, J, Rodriguez, RJ 1999, “Conversion of the Pathogenic Fungus Colletotrichum magna to A Non-pathogenic Endophytic Mutualist by Gene Disruption”, Molecular Plant-Microbe Interactions, vol. 12, pp. 969–975.
Redman, RS, Sheehan, KB, Stout, RG, Rodriguez, RJ, Henson, JM 2002, “Thermotolerance Generated by Plant/Fungal Symbiosis”, Science, vol. 298.
Ren, A, Li, C & Gao, Y 2011, “Endophytic Fungus Improves Growth and Metal Uptake of Lolium arundinaceum Darbyshire ex. Schreb.”, International Journal of Phytoremediation, vol. 13, no. 3, pp. 233-43.
Rich, G & Cherry, K 1987, Hazardous Waste Treatment Technologies, Pudvan Publishers, New York.
Rodrigues, KF, Hesse, M & Werner, C 2000, “Antimicrobial Activities of Secondary Metabolites Produced by Endophytic Fungi from Spondias mombin.”, Journal of Basic Microbiology, vol. 40, no. 4, pp. 261-7.
Rodriguez, R & Redman, R 2008, “More Than 400 million Years of Evolution and Some Plants still Can’t Make It On Their Own: Plant Stress Tolerance via Fungal Symbiosis”, Journal of Experimental Botany, vol. 59, pp. 1109-14.

- 109 -
Rodriguez, RJ, Henson, J, Volkenburgh, EV, Hoy, M, Wright, L, Beckwith, F, Kim, YO & Redman, RS 2008, “Stress Tolerance in Plants via Habitat-adapted Symbiosis”, The ISME Journal, vol. 2, pp. 404–416.
Rodriguez, RJ, Redman, RS & Henson, JM 2004, “The Role of Fungal Symbioses in the Adaptation of Plants to High Stress Environments”, Mitigation and adaptation strategies for global change, vol. 9, no. 3, pp. 261-272.
Rout, JR & Sahoo, SL 2013, “Antioxidant Enzyme Gene Expression in Response to Copper Stress in Withania somnifera L”, Plant Growth Regulation, vol. 71, no. 1, pp. 95-99.
Sablani, SS, Goosena, MFA, Al-Belushia, R & Wilfb, M 2001, “Concentration Polarization in Ultrafiltration and Reverse Osmosis: A Critical Review”, Desalination, vol. 141, no. 3, pp. 269–289.
Saiano, F & Ciofalo, M 2007, “Removal of Pesticide Oxadixyl from an Aqueous Solution” Bioremediation journal, vol. 11, no. 2, pp. 57-60.
Saiano, F, Ciofalo, M, Cacciola, SO & Ramirez, S 2005, “Metal Ion Adsorption by Phomopsis sp. Biomaterial in Laboratory Experiments and Real Wastewater Treatments”, Water Research, vol. 39, no. 11, pp. 2273-2280.
Salvo, ML, Safo, MK, Musayev, FN, Bossa, F & Schirch, V 2003, “Structure and Mechanism of Escherichia coli Pyridoxine 5'-phosphate Oxidase”, Biochimica et Biophysica Acta, vol. 1647, no. 1-2, pp. 76-82.
Santos, JM & Phillips, AJL 2009, “Resolving the Complex of Diaporthe (Phomopsis) species Occurring on Foeniculum vulgare in Portugal”, Fungal Diversity, vol. 34, pp. 111-125.
Schnell, R, Sriram, D & Schneider, G 2014, “Pyridoxal-phosphate Dependent Mycobacterial Cysteine Synthases: Structure, Mechanism and Potential as Drug Targets”, Biochim Biophys Acta.
Schulz, B, Boyle, C, Draeger, S, Römmert, AK & Krohn, K 2002, “Endophytic Fungi: A Source of Novel Biologically Active Secondary Metabolites”, Mycological Research, vol. 106, no. 9, pp. 996-1004.
Schulz, B, Wanke, U, Draeger, S & Aust, HJ 1993, “Endophytes from Herbaceous Plants and Shrubs: Effectiveness of Surface Sterilization Methods”, Mycological Research, vol. 97, no. 12, pp. 1447–1450.
Sebastianes, FL, Cabedo, N, El Aouad, N, Valente, AM, Lacava, PT, Azevedo, JL, Pizzirani-Kleiner, AA & Cortes, D 2012, “3-hydroxypropionic Acid as an Antibacterial Agent from Endophytic Fungi Diaporthe phaseolorum”, Current Microbiology, vol. 65, no. 5, pp. 622-32.
Seneviratne, M 2007, A Practical Approach to Water Conservation for Commercial and Industrial Facilities, 1st edn., Elsevier Science Australia, Marrickville, N.S.W.
Senthilkumar, N, Murugesan, S & Babu, DS 2014, “Metabolite Profiling of the Extracts of Endophytic Fungi of Entomopathogenic Significance, Aspergillus flavus and

- 110 -
Nigrospora sphaerica Isolated from Tropical Tree Species of India, Tectona grandis L.”, Journal of Agriculture and Life Sciences, vol. 1, no. 1, pp. 108-114.
Sharma, JK & Florence, EJM 1997, “Fungal Pathogens as a Potential Threat to Tropical Acacias: Case Study of India”, In: Proceedings of an International Workshop, Subanjeriji, South Sumatra, CIFOR Special Publication, pp 70-107.
Sherameti, I, Tripathi, S, Varma, A & Oelmüller, R 2008, “The Root-colonizing Endophyte Piriformospora indica Confers Drought Tolerance in Arabidopsis by Stimulating the Expression of Drought Stress-related Genes in Leaves,” Molecular Plant-Microbe Interaction, vol. 21, pp. 799-807.
Shevchenko, A, Tomas, H, Havli, J, Olsen, J V & Mann, M 2007, “In-gel Digestion for Mass Spectrometric Characterization of Proteins and Proteomes”, Nature protocols, vol. 1, no. 6, pp. 2856-2860.
Shevchenko, A, Wilm, M, Vorm, O & Mann, M 1996, “Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels”, Analytical Chemistry, vol. 68, pp. 850-858.
Sikora, RA, Pocasangre, L, Felde, AZ, Niere, B, Vu, TT & Dababat, AA 2008, “Mutualistic Endophytic Fungi and In-planta Suppressiveness to Plant Parasitic Nematodes”, Biological Control, vol. 46 no. 1, pp. 15–23.
Sim, CSF, Tan, WS, Ting, ASY 2015, “Endophytes from Phragmites for Metal Removal: Evaluating Their Metal Tolerance, Adaptive Tolerance Behaviour and Biosorption Efficacy”, Desalination and Water Treatment.
Singh, AK, Pindi, PK, Dube, S, Sundareswaran, VR & Shivaji, S 2009, “Importance of trmE for Growth of the Psychrophile Pseudomonas syringae at Low Temperatures”, Applied and Environmental Microbiology, vol. 75, no. 13, pp. 4419-26.
Singh, D, Tiwari, A & Gupta, R 2012, “Phytoremediation of Lead from Wastewater Using Aquatic Plants”, Journal of Agricultural Technology, Vol. 8, no. 1, pp. 1-11.
Singh, H 2006, Mycoremediation: Fungal Bioremediation, John Wiley & Sons.
Singh, LP, Gill, SS & Tuteja, N 2011, “Unraveling the Role of Fungal Symbionts in Plant Abiotic Stress Tolerance”, Plant Signaling & Behavior, vol. 6, no. 2, pp. 175–191.
Slack, A & Gate, J 2000, Carnivorous Plants, MIT Press.
Smith, AH, Lingas, EO & Rahman, M 2000, “Contamination of Drinking-water by Arsenic in Bangladesh: A Public Health Emergency”, Bulletin of the World Health Organization, vol. 78, no. 9, pp. 1903-1103.
Soleimani, M, Hajabbasi, MA, Afyuni, M, Mirlohi, A, Borggaard, O K & Holm, PE 2010, “Effect of Endophytic Fungi on Cadmium Tolerance and Bioaccumulation by Festuca arundinacea and Festuca pratensis”, International Journal of Phytoremediation, vol. 12, no. 6, pp. 535-549.

- 111 -
Stadler, M, Hawksworth, DL & Fournier J 2014, “The Application of the Name Xylaria hypoxylon, based on Clavaria hypoxylon of Linnaeus”, IMA Fungus, vol. 5, no 1, pp. 57–66.
Stern, BR, Solioz, M, Krewski, D, Aggett, P, Aw, TC, Baker, S, Crump, K, Dourson, M, Haber, L, Hertzberg, R, Keen, C, Meek, B, Rudenko, L, Schoeny, R, Slob, W & Starr, T 2007, “Copper and Human Health: Biochemistry, Genetics, and Strategies For Modeling Dose-Response Relationships”, Journal of toxicology and environmental health. Part B, Critical reviews, vol. 10, no. 3, pp. 157-222.
Stone, JK, Polishook, JA & White, JFJr 2011, “Endophytic Fungi”, in MS Foster & GF Bills (eds), Biodiversity of Fungi: Inventory and Monitoring Methods, Academic Press.
Strathmann, H 1992, “Introduction and Definitions”, Membrane Handbook, pp. 219-222.
Strobel, G & Daisy, B 2003, “Bioprospecting for Microbial Endophytes and Their Natural Products”, Microbiology and Molecular Biology Reviews, vol. 67, no. 4, pp. 491-502.
Tadych, M & White, JF 2009, “Endophytic microbes”, in M Schaechter (ed), Encyclopedia of Microbiology, 3rd edn. Elsevier, Dordrecht, pp. 431-442.
Tamura, K, Stecher, G, Peterson, D, Filipski, A & Kumar, S 2013, “MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0”, Molecular Biology and Evolution, vol. 30, no. 12, pp. 2725–2729.
Tangahu, BV, Abdullah, SRS, Basri, H, Idris, M, Anuar, N & Mukhlisin, M 2011, “A Review on Heavy Metals (As, Pb, and Hg) Uptake by Plants Through Phytoremediation,” International Journal of Chemical Engineering.
Tchounwou, PB, Yedjou, CG, Patlolla, AK & Sutton, DJ 2012, “Heavy Metals Toxicity and the Environment”, EXS, vol. 101, pp. 133–164.
Teixeira-Gomes, AP, Cloeckaert, A & Zygmunt, MS 2000, “Characterization of Heat, Oxidative, and Acid Stress Responses in Brucella melitensis”, Infection and Immunity., vol. 68, no. 5, pp. 2954-2961.
Toh, SM & Mankin, AS. 2008, “An Indigenous Posttranscriptional Modification in the Ribosomal Peptidyl Transferase Center Confers Resistance to an Array of Protein Synthesis Inhibitors”, Journal of Molecular Biology, vol. 380, no. 4, pp. 593-7.
Tomono, K, Kawai, M, Sasaki, H, Homma, Y & Misat, T 1982, “The Influence of Zinc, Magnesium and Calcium Ions on the Inhibitory Effect of Copper (II) Ion against Diaporthe citri.”, Journal of Pesticide Science, vol. 7, pp. 329-334.
Torres, MA, Jones, JDG & Dangl, JL 2006, “Reactive Oxygen Species Signaling in Response to Pathogens”, Plant physiology, vol. 141, no. 2, pp. 373-378.
Townsley, CC, Ross, IS & Atkins, AS 1986, “Biorecovery of Metallic Residues from Various Industrial Effluents Using Filamentous Fungi”, in RW Lawrence, RMR Branion & HG Ebner (eds), Fundamental and Applied Biohydrometallurgy, Elsevier: Amsterdam, pp 279-289.

- 112 -
U.S. Environmental Protection Agency 1994, “Technical Document Acid Mine Drainage Prediction”, Office of Solid Waste, Washington, DC.
U.S. Environmental Protection Agency 2013, Seminar Publication: Meeting Hazardous Waste Requirements for Metal Finishers, BiblioGov.
UniProta, Formamidopyrimidine-DNA glycosylase, viewed 15th Febuary 2015, http://www.uniprot.org/uniprot/Q87T81.
UniProtb, Ketol-acid reductoisomerase, viewed 15th Febuary 2015, http://www.uniprot.org/uniprot/A3PEE9.
UniProtc, Ribosomal RNA large subunit methyltransferase E, viewed 15th Febuary 2015, http://www.uniprot.org/uniprot/B9LSX2.
UniProtd, Ribosomal RNA small subunit methyltransferase G, viewed 15th Febuary 2015, http://www.uniprot.org/uniprot/B7J1B0.
UniProte, Uncharacterized protein C24B10.16c, viewed 15th Febuary 2015, http://www.uniprot.org/uniprot/Q9P7J0.
UniProtf, Cell division protein SepF, viewed 15th Febuary 2015, http://www.uniprot.org/uniprot/B8ZQQ6.
USDA Forest Service 1993, “Acid Mine Drainage from Mines on the National Forests”, A Management Challenge. Program Aid 1505, p.12.
Vaajasaari, K & Joutti, A 2006, “Field-Scale Assessment of Phytotreatment of Soil Contaminated with Weathered Hydrocarbons and Heavy Metals (9 pp)”, Journal of Soils and Sediments, vol. 6, no. 3, pp. 128-136.
Valle, JRD, Chávez-Salinas, S, Medina, F & Angel RMD 2005, “Heat Shock Protein 90 and Heat Shock Protein 70 are Components of Dengue Virus Receptor Complex in Human Cells”, Journal of Virology, vol. 79, no. 8, pp. 4557-67.
Varma, A, Verma, S, Sudha, Sahay, N, Bütehorn, B & Franken, P 1999, “Piriformospora indica, a Cultivable Plant-Growth-Promoting Root Endophyte”, Applied and Environmental Microbiology, vol. 65, no. 6, pp. 2741–2744.
Venugopal, V & Luckey, TD 1975, “Toxicology of Nonradio-Active Heavy Metals and Their Salts,” in TD Luckey, B Venugopal & D Hutcheson (eds), Heavy Metal Toxicity, Safety and Hormology, George Thieme, Stuttgart.
Verma, AK, Raghukumar, C, Verma, P, Shouche, YS & Naik, CG 2010, “Four Marine-derived Fungi for Bioremediation of Raw Textile Mill Effluents”, Biodegradation, vol. 21, no. 2 pp. 217-233.
Vieira, ML, Johann, S, Hughes, FM, Rosa, CA & Rosa, LH 2014, “The Diversity and Antimicrobial Activity of Endophytic Fungi Associated With Medicinal Plant Baccharis trimera (Asteraceae) from the Brazilian savannah”, Canadian Journal of Microbiology, vol. 60, no. 12, pp. 847-56.

- 113 -
Vigneswaran, S, Ngo, HH, Chaudhary, DS, & Hung, YT 2005, “Physicochemical Treatment Processes for Water Reuse”, Physicochemical treatment processes, Humana Press, pp. 635-676.
Volesky, B 1986, “Biosorbent Materials”, Biotechnol. Bioeng. Symp. Ser, vol. 16, pp. 121-126.
Volesky, B 1990, Biosorption of Heavy Metals, CRC press.
Wang, HW, Dai, CC, Zhu, H & Wang, XX 2014, “Survival of a Novel Endophytic Fungus Phomopsis liquidambari B3 in the Indole-contaminated Soil Detected by Real-time PCR and Its Effects on the Indigenous Microbial Community”, Microbiological Research, vol. 169, no. 12, pp. 881–887.
Wang, LK, Chen, JP, Hung, YT & Shammas, NK 2009, “Heavy Metals in the Environment,” Advances in Industrial and Hazardous Wastes Treatment, CRC Press.
Wang, LK, Shammas, NK & Hung, YT 2008, Waste Treatment in the Metal Manufacturing, Forming, Coating, and Finishing Industries (Advances in Industrial and Hazardous Wastes Treatment), CRC Press.
Wang, LK, Tay, JH, Tay, STL & Hun YT 2010, Environmental Bioengineering, Volume 11 of Handbook of Environmental Engineering, Springer Science & Business Media.
Wang, R, Gao, F, Guo, BQ, Huang, JC, Wang, L & Zhou, YJ 2013, “Short-Term Chromium-Stress-Induced Alterations in the Maize Leaf Proteome”, International Journal of Molecular Sciences, vol. 14, no. 6, pp. 11125–11144.
Ward, SK, Hoye, EA & Talaat, AM 2008, “The Global Responses of Mycobacterium tuberculosis to Physiological Levels of Copper”, Journal of Bacteriology, vol. 190, no. 8, p. 2939–2946.
Washburn, MP & Yates III, JR 2000, “Analysis of the Microbial Proteome”, Current Opinion in Microbiology, vol. 3, no. 3, pp. 292-297.
Webb, KM, Hill, AL, Laufman, J, Hanson, LE & Panella, L 2011, “Long-term Preservation of a Collection of Rhizoctonia solani Using Cryogenic Storage”, Annals of Applied Biology, vol. 158, pp. 297–304.
Webster 1976, 3rd New International Dictionary, Merriam, Chicago.
Wei, DL, Chang, SC, Wei, YH, Lin, YW, Chuang, CL & Jong, SC 1992, “Production of Cellulolytic Enzymes from the Xylaria and Hypoxylon species of Xylariaceae”, World Journal of Microbiology and Biotechnology, vol. 8, no. 2, pp. 141-146.
Weising, K, Nybom, H, Wolff, K & Meyer, W 1994, DNA fingerprinting in plants and fungi, CRC Press, London.
Weissenhorn, I, Leyval, C & Berthelin, J 1993, “Cd-tolerant Arbuscular Mycorrhizal (AM) Fungi from Heavy-Metal Polluted Soils”, Plant and Soil, vol. 157, no. 2, pp. 247-256.

- 114 -
Wenzel, WW 2009, “Rhizosphere Processes and Management in Plant-assisted Bioremediation (Phytoremediation) Oof Soils”, Plant and Soil, vol. 321, no. 1-2, pp. 385-408.
Wilkins, MR, Pasquali, C, Appel, RD, Ou, K, Golaz, O, Sanchez, JC, Yan, JX, Gooley, AA, Hughes, G, Humphery-Smith, I, Williams, KL & Hochstrasser, DF 1996, “From Proteins to Proteomes: Large Scale Protein Identification by Two-dimensional Electrophoresis and Amino Acid Analysis”, Biotechnology (N Y), vol. 14, pp. 61-5.
Winter, J, Linke, K, Jatzek, A & Jakob, U 2005, “Severe Oxidative Stress Causes Inactivation of DnaK and Activation of the Redox-Regulated Chaperone Hsp33”, Molecular Cell, vol. 17, no. 3, pp. 381–392.
World Health Organization 1996, Trace elements in human nutrition and health.
Xiao, X, Luo, S, Zeng, G, Wei, W, Wan, Y, Chen, L, Guo, H, Cao, Z, Yang, L, Chen, J & Xi, Q 2010, “Biosorption of Cadmium by Endophytic Fungus (EF) Microsphaeropsis sp. LSE10 Isolated from Cadmium Hyperaccumulator Solanum nigrum L.”, Bioresource Technology, vol. 101, no. 6, pp.1668-74.
Xu, Y, Szép, S & Lu, Z 2009, “The Antioxidant Role of Thiocyanate in the Pathogenesis of Cystic Fibrosis and Other Inflammation-Related Diseases”, Proceedings of the National Academy of Sciences USA, vol. 106, no. 48, pp. 20515-9.
Yang, K, Eyobo, Y, Brand, LA, Martynowski, D, Tomchick, D, Strauss, E & Zhang, H 2006, “Crystal Structure of a Type III Pantothenate Kinase: Insight into the Mechanism of an Essential Coenzyme A Biosynthetic Enzyme Universally Distributed in Bacteria”, Journal of Bacteriology, vol. 188, no. 15, pp. 5532–5540.
Yıldırım, V, Özcan, S, Becher, D, Büttner, K, Hecker, M & Özcengiz, G 2011, “Characterization of Proteome Alterations in Phanerochaete chrysosporium in Response to Lead Exposure”, Proteome Science, vol. 9.
Ying, W 2008, “NAD+/NADH and NADP+/NADPH in Cellular Functions and Cell Death: Regulation and Biological Consequences”, Antioxidants & Redox Signaling, vol. 10, no. 2, pp. 179-206.
Yunus, K, Yusuf, NM, Shazili, NAM, Chuan, OM, Saad, S, Chowdhury, AJK & Bidai, J 2011, “Heavy Metal Concentration in the Surface Sediment of Tanjung Lumpur,” Sains Malaysiana, vol. 40, pp. 89-92.
Zafar, S, Aqil, F & Ahmad, I 2007, “Metal Tolerance and Biosorption Potential of Filamentous Fungi Isolated from Metal Contaminated Agricultural Soil”, Bioresource Technology, vol. 98, no. 13, pp. 2557–2561.
Zaiyou, J, Li, M, Guifang, X & Xiuren, Z 2013, “Isolation of An Endophytic Fungus Producing Baccatin III from Taxus wallichiana var. mairei”, Journal of Industrial Microbiology & Biotechnology, vol. 40, no. 11, pp. 1297-302.

- 115 -
Zaurov, DE, Bonosa, S, Murphya, JA, Richardsonc, M & Belanger, FC 2001, “Endophyte Infection Can Contribute to Aluminum Tolerance in Fine Fescues”, Crop Science, vol. 41, no. 6, pp. 1981–1984.
Zehetner, F, Rosenfellner, U, Mentler, A & Gerzabek, MH 2009, “Distribution of Road Salt Residues, Heavy Metals and Polycyclic Aromatic Hydrocarbons Across a Highway-Forest Interface”, Water, Air, and Soil Pollution, vol. 198, no. 1-4, pp. 125-132.
Zhao, W, Han, J & Long, D 2014, “Effect of Copper−induced Oxidative Stress on Sclerotial Differentiation, Endogenous Antioxidant Contents, and Antioxidative Enzyme Activities of Penicillium thomii PT95”, Annals of Microbiology.
Zhou, S, Sauvé, RJ & Liu, Z 2011, “Sasikiran Reddy and Sarabjit Bhatti, Simon D. Hucko, Tara Fish and Theodore W. Thannhauser (2011) Identification of Salt-induced Changes in Leaf and Root Proteomes of the Wild Tomato, Solanum chilense”, JASHS, vol. 136, no. 4, pp. 288-302.

![Pomelo Peel: Agricultural Waste for Biosorption of …€¦ · · 2017-05-05controlling parameter in the biosorption process [19]. Most research conducted on heavy metal biosorption](https://img.pdfslide.net/doc/110x75/5b06dd247f8b9a79538d0e79/pomelo-peel-agricultural-waste-for-biosorption-of-parameter-in-the-biosorption.jpg)



























