Embed Size (px)
Citation preview

768
BIOTECHNOLOGY
Synthesis of bioplastics in plants
During the past decade, muchattention has been focused onthe development of new
biodegradable plastics. This interesthas been fueled by a greater aware-ness of the detrimental effect of syn-thetic plastics all our environment andof problems associated with their dis-posal. Among the various biodegrad-able plastics available, there is agrowing interest in the group of poly-hydroxyalkanoates (PH As). polyestersproduced by bacteria (1-4). Thesepolymers represent a valuable sourceof biodegradable and environmentallysafe biomnterials which can be used ina wide range of products such as uren-sits and dishes for the food industry.Films for packaging or hygienic prod-ucts, medical implants and gauze,agricultural mulches. etc. In contrastto many blodegrndeble plastics thatcan be partially broken down to small-er nondegradable constituents. eitherphotochemically or by nonenzymatichydrolysis. PHAs can be fullydegraded by microorganisms to waterand carbon dioxide. Thus, consumerproducts made of PHAs can bedegraded by including them in a com-post (Fig. I).
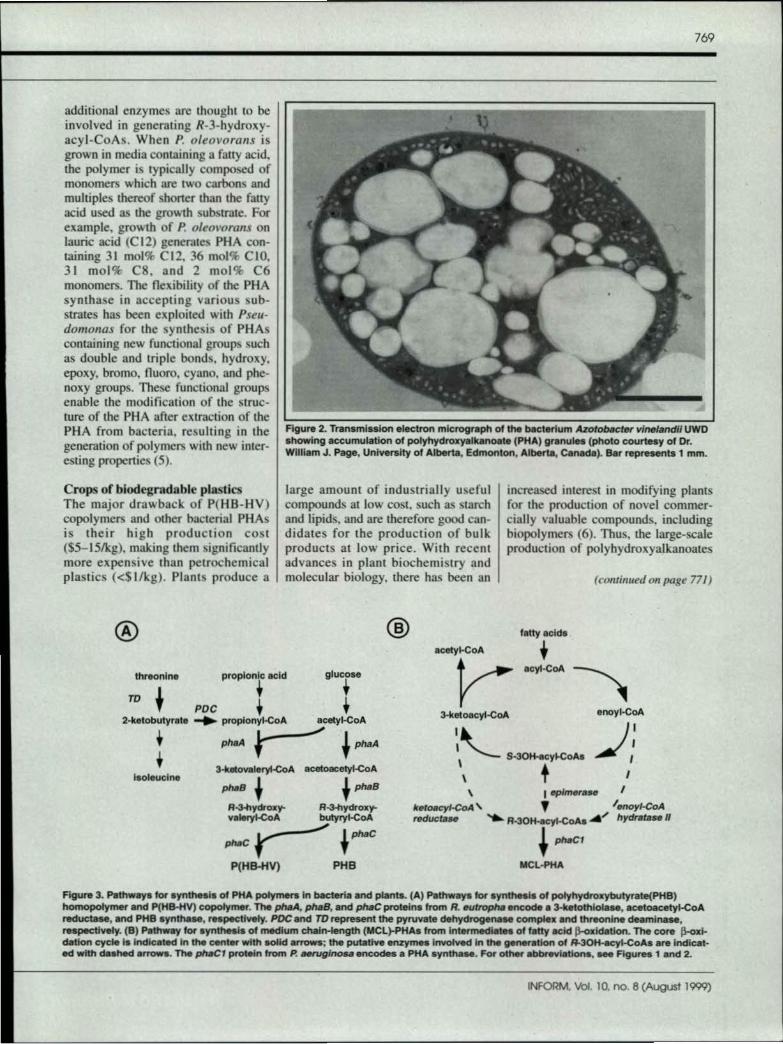
PH As represent a large family ofpolymers, composed primarily of 3-hydroxyacids. which are produced asintracellular granules by a wide vari-ety of bacteria (1-4) (Fig. 2). Thesepolymers act as a carbon reserve andelectron sink when bacteria are grownin media containing an excess of car-bon while one or more essential nutri-ents are limiting (e.g .. NO) or PO!l.When growth-limiting conditions arealleviated, the intracellular PHA is
This anicte is by \obnt!!.fD Bongcamand }Vt!s Poirier. In.rtilut d'Ecolo-gie-Biologie t!IPhydolog;~ Vigl-
tales, Uni~r:ri'ltk ~. CH-10ISLAwQIIM, SwiturlantL
INFORM. Vol. 10, ro. 8 (August 1999)
sizing SCL-PHAs. most of the knowl-edge has been obtained from Ralsto-nia eutropno (formerly Alcaligeneseutroplws). Synthesis of MCL-PHAsis mainly confined to the Pseu-domonas species, with most studiesdone on P. oleovorans.
In R, eusropba. polyhydroxybu-ryrare (PHB) is synthesized fromacetyl-Cox by the sequential action ofthree enzymes: 3-ketothiolase (plulA),acetoacetyl-Cox reductase (plIO B).and the PHA synthase (phaC) (Fig.3A). Copolymers of 3-hydroxybu-tyrate and 3-hydroxyvalerate [P(HB-HV)] can be produced by R. eutrophausing as substrates propionic or valer-ic acid mixed with glucose (Fig. 3A).P(HB-HV) is more flexible then PHB.which is relatively stiff and brittle,making P(HB-HV) a better polymerfor a wide variety of applications.Until recently. P(HB-HV) was pro-duced commercially under the tradename Biopel by Monsanto.
In Pseudomonas, the pathway ofMCL-PHAs synthesis is linked to thegeneration of 3-hydroxyacyl-CoAintermediates from the p-oxidationcycle of fatty acid degradation (Fig.38). MCL-PHA is made exclusivelyof the R isomer of 3-hydroxyacids.Since bacterial Ii-oxidation generatesS-3-hydroxyacyl-CoA intermediates.
Figure 1. Degradation In a compost of boHlel made of [P(HB·HVII cempl .. (photocourtesy of Dr. O. Seebach, ETH Zentrum, Zurich, Switzerland)
cleaved by a PHA depolymerase andthen catabolized to acetyl-CoA. Inaddition to the intracellular PHAdepolymerase. some bacteria andfungi possess extracellular PHAdepolymerases which enable theseorganisms to utilize external PHAs asa carbon source.
Bacteria that synthesize PHAs canbe subdivided into two generalgroups: the first one produces shortchain-length PH As (SCL-PHAs), hav-ing monomers three to five carbons inlength (C3-C5). while the secondgroup synthesizes medium chain-length PH As (MCL-PHAs). withmonomers ranging from C6-C 16(1-4). SCL-PHAs generally are morerigid thermoplastics while MCL-PHAs behave as elastomers. Despitethe large diversity of bacteria synthe-
"--"gum ---
769
propionic acid glucOl8, ,PDC, ,
2-katobutyr8te .... proplonyl-CoA acetyf.CoA
, phoA t--./,3-klltovaI8ry1~A acetoacetyl-CoA
pMB + + phs8R..:u.ydroxy- R-3-fIydJoxy..."leryI-CoA butyry1-CoA
pMC~ ~pMC
P{HB-HV) PHB
Figure 3. Pathway. for synthesll 01 PHA polymers in bacteria and p1anl$. (Al Pathway. for synthesis 01 ~trydroKYbutyrate(PHB)homopolymer and P(HB-HV) copotymer. The phsA. phaB, and phaC proleins from R. eutroplv encode I 3-kelothlolase, acetoaeetyJ.CoAreductase, and PHB syntha .. , respectively. PDC and TD represent the pynIvate dehydrogen ... comple. and threonine deaminase,respectlvfly. (Bl Pathway for synthes'l of medium chaJMength (MCL~PHAI from Intermediate. 01 fatty acid ji-olldatlon. The eeee ji-oxi-dation eycle.a Indic.ted In the cent ... whh solid arrows; the putative enzymes Involved In the generation 01 R-30H-aql-COAs are Indicat-ed with dashed arrow•. The plYCI protein from P. .enJglnosa enc:Qdes I PHAIynthne. For oth« abbreviations, _ Figures 1 and 2.
additional enzymes arc thought to beinvolved in generating R·3-hydroxy-acyl-CoAs. When P. oteovomns isgrown in media containing a fatty acid.the polymer is typically composed ofmonomers which are two carbons andmultiples thereof shorter thun the fauyacid used as the growth substrate. Forexample, growth of P. oleovorans onlauric acid (C 12) generates PHA con-taining 31 0101% C12. 36 0101% CIO,31 mol% C8. and 2 mol% C6monomers. The flexibility of the PHAsynthase in accepting various sub-strates has been exploited with Pseu-domonas for the synthesis of PH Ascontaining new functional groups suchas double and triple bonds. hydroxy.epoxy, bromo. flucro. cyano. and phe-noxy groups. These functional groupsenable the modification of the struc-ture of the PHA after extraction of thePHA from bacteria. resulting in thegeneration of polymers with new inter-esting properties (5).
Crops of biodegradable plasticsThe major drawback of P(HB-HV)copolymers and other bacterial PHAsis their high production cost($5-15Ikg). making them significantlymore expensive than petrochemicalplastics (<SlIkg). Plants produce a
®threonine
TD
Isoleuclr'lfll
FIgure 2. Transmission electron micrograph of the bacterium Azotobacter v/nelandll UWDahowlng accumulation of polyhydroxyallulnoate (PHA) granule, (photo courtesy of Dr.William J. Page, University of Atberts, Edmonton, Alberta, Caneda). Sar represents 1 mm.
large amount of industrially usefulcompounds at low cost, such as starchand lipids, and are therefore good can-didates for the production of bulkproducts at low price. With recentadvances in plant biochemistry andmolecular biology, there has been an
increased interest in modifying plantsfor the production of novel commer-cially valuable compounds, includingbiopolymers (6). Thus. the large-scaleproduction of polyhydroxyalkanoates
tcominued on page 771)
® fatty acids
acetyl-CoA +V ".-CoA~
aooyl--CoA
..J:II
, I ~rmI,a.e IketOllcyJ-CoA' , I enoy/-CoAreductaH ..... R-30H-ecyl-CoA. A.... hydrata$e II
~ pMC'
~ phaA
3-ket08Cyl-CoA
~\~ S-30H-tlcyl-CoAs
\ t
MCL-f'HA
tNFORM,Vol. 10,no. 8 (August 1999)

771
BIOTECHNOlOGY
[cominuedfrom page 769)
in crop plants became an anractivealternative. Production of PHAs on anagronomic scale could allow synthesisof biodegradable plastics in the rangeof IOL1()6 Ions per year, quantitiesdifficult to reach by bacterial fennen-ration alone. If PHA can be synthe-sized in plants to a level comparable\0 reserve lipids, i.e., approximately10--40% dry weight, PHA potentiallycould be produced at a cost compare-ble 10 vegetable oil ($O.5/kg) and thusbe competitive with petroleum-derived plastics (1,7).
PHB production in plantsSynthesis of PHA in plants was ini-tially explored by expression of thePHB biosynthetic genes of R.eutropha in Arabidopsis thaliana, aplant related 10 rapeseed. The firstexperiments aimed at assessing thefeasibility of synthesizing PHS inplants were done by expressing thePH S biosynthetic pathway in thecytoplasm (8). The R. eutropha genesencoding the acetoacetyl-CoA reduc-tase (phaB) and PHA synthase (plwC)were expressed constitutively intransgenic A. tnatiana. The highestamount of PHS obtained in theseplants was approximately 0.1 % of theshoot dry weight. Transmission elec-tron microscopy revealed thut PHSaccumulated in the form of granulesthat were indistinguishable from bac-terial PHA inclusions. Chemical anal-ysis of the plant polymer indicatedthat it was of high molecular weightand had properties similar to bacterialPHS (9). Although PHB synthesis inplants could be demonstrated success-fully, PHB accumulation was very lowand the high level of expression ofacetoacetyl-Cos, reductase led to areduction in growth of transgenicplants, with a corresponding decreasein seed yield. Both the low PHB pro-duction and the deleterious effect onplant growth were thought to be duemainly to the limited supply of cyto-plasmic acetyl-CoA available forPHB, isoprenoid, and flavonoidbiosynthesis (10). To solve this prob-lem, it was necessary to find anothercell compartment with a higher flux ofcarbon through ecetyl-Ccx.
Figure 4. TransmIssIon electron micrograph of transgenIc Arab/dope'e cells accumulatIngPHB granules (translucent areas Indicated by arrows) In the pla'tlds.Bar reprllentl 1 mm. For abbrevIation see FIgure 3.
In plants. fatty acid biosynthesisoccurs in the plastid using acetyl-CoAas a precursor. The plastid is thereforea site of high flux of carbon throughacetyl-CoA, particularly in developingseeds of oil-accumulating plants, suchas A. thaliana, where up to 40% of theseed dry weight is triacylglycerols.Expression of the PH B biosyntheticpathway in plastics was demonstratedin transgenic A. thaiiono (II). The R.eutropha plutA, pl/aB, and phaC geneswere modified for targeting the pro-teins to the plastids, and the geneswere constitutively expressed in A.thatiana. Transgenic plants accumu-lated PHS granules exclusively in theplastids (Figure 4). The maximalamount of PHS detected in pre-senescing leaves was 10-14% dryweight. In plants producing high lev-els of PHB, leaves showed chlorosis,indicating some defect in chloroplastfunction. although this phenotype wasnot accompanied by a reduction infresh weight. Thus, redirecting thePHB biosynthetic pathway from thecytoplasm to the plastid resulted in alOO-fold increase in PHB productionwith limited deleterious effects on theplants (I I).
Synthesis of PHS in crop plantswas initiated in the early 1990s byZeneca, which also produced PHBand P(HS-HV) by bacterial fermenta-tion. Rapeseed was chosen as the ini-tial target crop. Expression of thePHB pathway in the cytoplasm led 10low levels of PHB accumulation ill
leaves. similar to the level detected inArobidopsis (0.1% dry weight). In1995. Zeneea sold its PHA business toMonsanto. which also took up thechallenge to produce PHB in crops.Monsanto recently has shown thattransgenic rape expressing the PHBpathway in the leucoplast of develop-ing embryos accumulated PHB at lev-els up to 8% dry weight in matureseeds of heterozygous plants (12).
PHA copolymers in plantsPHB has relatively poor physicalproperties, being too stiff and brittlefor most applications. There is thus alarge interest in synthesizing PHAcopolymers with better physical prop-erties in plants. Two recent studieshave shown that it is possible to syn-thesize both SCL-PHA and MeL-PHA copolymers in plants.
In one study, accumulation ofMCL-PHA was obtained by expres-sion of the PHA synthase from thebacterium P. oeruginosa in plant per-oxlsomes (13). The peroxisome waschosen because it is the site of fattyacid p-oxidation in plants, which canprovide the 3-hydroxyacyl-CoA sub-strate to the PHA synthase for MCL-PHA synthesis. The P. aeruginosoPHA synthase was modified by theaddition of a peroxisomal targetingsignal from rapeseed isocitrate lyaseat the carboxy end and expressed in A.thaliana under the control of a consti-tutive promoter (13). Analysis oftransgenic plant material by gas cnro-
INFORM. Vol. 10. no. 8 (August 1m)

772
BIOTECHNOLOGY
enoyl-Coa hydratase II activity. or thehydration of 2-cis-cnoyl-CoA general-ed from the p-oxidation of fatty acidshaving cis double bonds at the evenposition (13).
In order to assess the factors tim-iring the synthesis of MCL·PHAs inperoxisomes, transgenic plants weregrown in liquid media supplementedwith Tween detergents as a source offatty acids. Addition of Tween-lO.containing primarily lauric acid.resulted in a IO-fold increase inMCL·PHA accumulation. Additionof unusual fatty acids to the mediaresulted in the incorporation. intothe polyester. of monomers derivedfrom the Bcoxldauon of these fattyacids. For example. addition oftridecanoic acid, tridecenoic acid(eI3: 1, .6.12), or 8-mcthyl-nonanoicacid resulted in the production of
PHA containing mainly saturatedodd-chain, unsaturated odd-chain,or branched-chain 3-hydroxyacidmonomers. respectively. These stud-ies demonstrated that the plant P-oxidation pathway was capable ofgenerating a large spectrum ofmonomers for PHA synthesis fromunusual fatty acids and that the sup-ply of fatty acids to the Bcoxidatlonpathway was a factor limiting theaccumulation of PHA in plants.
To overcome Ibis limitation in vivo.the endogenous nux of fatty acids top-oxidation was increased throughgenetic engineering by the expressionof a medium-chain acyl-ACP (acylcarrier protein) thioesterase fromCuohea tonceolata. Previous studiesdone with transgenic rapeseedexpressing the California bay Iauroyl-ACP uuoesterase had indicated that
matography and mass spectrometrydemonstrated the synthesis of a com-plex PHA containing over 15 differ-ent monomers, ranging from C6 toC 16 3-hydroxyacids. with somemonomers containing up to threeunsaturated bonds. Approximately0.4% dry weight of PHA was pro-duced in 7-day-old germinatingseedlings. Analysis of transgenic tis-sues by electron microscopy revealedthe presence of numerous PHA gran-ules in the peroxisomes. These resultsindicated that. like bacteria, plantsalso have enzymes capable of generat-ing the R-isomer of 3-hydroxyacyl-CoA from p-oxidation intermediates.It is hypothesized that a broad spec-trum of R-3-hydroxyacyl-CoAs couldbe generated either by the epimeriza-lion of the $-3- hydroxyacyl-CoA, thehydration of 2-lrans-enoyl-CoA by a
REINARTZ screw presses give priorityto an efficient mechanical cold pressingfrom small scale up to a high capacityrange.
The decisive advantage of a cold press-ing with REINARTZ screw presses isthat no trans fatty acids are produced inthe oil and the press cake. Trans fattyacids, which are most unwanted inhuman and animal nutrition, occur whenpreheating the oilseed prior to pressing.
Furthermore, due to its low phosphorlipids content, the cold-pressed oil fromREINARTZ screw presses has an excel-lent quality for the substitution of dieseloil in combustion engines.
GegrOndet• Founded1853
Manufacturers of screw pressesfor more than 50 years.
PRODUCTION OF VEGETABLE Oil
Head Office:Maschinenfabrik ReinartzGmbH & Co. KGP.O. Box 10 09 500-41409 NeusslGermanyPhone: (02131) 97 61·0Fax: (02131) 97 61 - 12
For inJon'no1i04'1 circle .110 on form on loll page
INFORM Vol. 10. no. 8 (August 1999)

773
medium-chain fatty acids synthesizedin plastids from leaves expressing thethioesterase were degraded by p-oxi-dation before any accumulation couldbe detected in vegetative tissues (14).It was thus hypothesized that expres-sion of a medium-chain thioesrerase inconjunction with a PHA synthasewould create a higher nux of carbontoward Fatty acid p-oxidation andPHA synthesis. Transgenic plantsexpressing the peroxisomal PHA syn-thase and the plastidial medium-chainacyl-ACP unoesterase from C. tonce-otato produced IO-fold more PHA inmature shoots compared to plantsexpressing only the PHA synthase.PHA produced in double-transgenicplants contained mainly saturatedmonomers ranging from 6 to 10 car-bons. consistent with the specificity ofthe C. lanceolato rhioesterase forcaproyl-ACP. and in contrast to MCL-PHA produced from plants expressingonly the PHA synthase. which wasmainly composed of unsaturated long-chain monomers.
These results clearly showed thatthe quality and quantity of MCL-PHAs in plants can be modulated bythe co-expression of enzymes respon-sible for the synthesis of unusual fattyacids together with a peroxisomalPHA synthase. Furthermore, thesestudies indicate that analysis of thenature and proportion of monomersfound in peroxisomal MCL-PHA canbe used in basic plant biochemistry toelucidate the pathways involved in thedegradation of unsaturated and unusu-al funy acids. while the quantity ofPHAs can be used to study nux offatty acids toward peroxlscmat ~-oxi-dation.
In an alternative approach aimed atthe synthesis of P(HB-HV) copoly-mers in plants. scientists at Monsantoco-expressed in the plastids a threo-nine deaminase from Escherichia colialong with the PHB biosynthetic path-way (12) (Figure 3A). Threoninedeamina.se catalyzes the conversion ofthreonine 10 u-kerobutyrate. whichitself can be convened. albeit at lowefficiency. to propionyl-CoA by theaction of the pyruvate dehydrogenasecomplex. The propionyl-CoA can becondensed with acetyl-Cox to formvaleryl-CoA. which can then be used
for P(HB-HV) synthesis. Expressionof the threonine deaminase togetherwith the PHB biosynthetic pathway inleucoplasts of rapeseed embryosresulted in P(HB-HV) accumulation to2.3% dry weight in seeds. with a
type of polymer from seedlings. Alter-natively, one also can envisage pro-ducing PHA in the leaves or stems ofa variety of crops. thus adding valueto the plant biomass that is oftenregarded as a waste material. It has
Important Jactors affectingthe commercial viability
oj PHA production ill cropsare the extraction and
the purification oj the polymer.
hydroxyvalerare content up to 6.5mol% (12). Demonstration of P(HB-HV) synthesis in plants represents asignificant advance for commercial-ization of PHA from crop plants sincethe equivalent bacterial copolymerBiopol has been well-studied and isalready in commercial use.
also been suggested that carbohydrnte-accumulating crops. such as potato orsugar beet. could be used to producePHA. In this last example, synthesisof PHA would be dependent on indi-reedy diverting carbon away from car-bohydrate synthesis and toward PHA.
Important factors affecting thecommercial viability of PHA pro-duction in crops are the extractionand the purification of the polymer.Although PHA synthesized in bacte-ria can account for up to 85% dryweight and can be relatively easilypurified by protease and detergenttreatment, extraction of 10-20%PHA from plant material is likely tobe more difficult. adding to the pro-duction cost of the polymer. Finally.one must keep in mind that high-level synthesis of PHA in any cropis likely to require the coordinatedand stable expression of numeroustransgenes affecting severalmetabolic pathways. Synthesis ofP(HB-HV) requires at least fourtrausgenes affecting fany acid andamino acid biosynthetic pathways.Thus, in comparison to the first-gen-eration transgenic crops actually inthe field. which typically expressonly one or two transgenes, the suc-cessful development of a PHA-pro-ducing crop represents a very com-plex and challenging genetic engi-neering project.
Bringing PHA synthesis to the fieldIt has now been shown that it is pos-sible for plants to synthesize a vari-ety of PH As having properties rang-ing from stiff plastic (PH B). softerplastic [P(HB-HV)], and elastomers(MCL-PHAs). The challenge for thefuture is to reach commercial levelsof the right PHA in an appropriatecrop. It is estimated that the break-even point for commercial viabilityof PHA production in plants wouldbe approximately 15% dry weightPHA. assuming a relatively lowdecrease in biomass yield. From thisperspective. the accumulation ofPHB in rapeseed embryos of up 108% dry weight in heterozygousplants appears promising.
Several strategies for synthesis ofPHA in crops can be envisaged. Syn-thesis of PH As in seeds of oil cropshas several advantages. PHD andP(HB-HV) synthesis is dependent onan abundant source of acetyl-CoA.which can be provided in the leu-cop last of developing seeds of oilcrops. These PHAs could thus be har-vested from dry mature seeds. ForMCL-PHA synthesis, the large tria-cylglycerol reserve which is degradedthrough ~-oxidalion during germina-tion could enable the harvest of this
References
I. Poirier, Y.. C. Nawrath, and C.Somerville. Production of poly-hydroxyalkanoates. a family of
INFORM. Vol. 10. no. 8 (August 1999)

774
BIOTECHNOLOGY
High Vacuum Systems and Jet Ejectors for Fat and Oil Industry200 equipments installed in 20 countries of the world
HI. Pedroni y Asociados S.l_. CI•... _" •• w-c-.IyIt.I- ..... 77 I • -11* ......
biodegradable plastics and elas-tomers, in bacteria and plants.Biotechnology 13: 142-150(1995).
2. Steinbilchel A., Polyhydroxyalka-noic acids, in Novel Biomaterialsfrom Biological Sources, editedby D. Byrom. MacMillan. NewYork, 1991. pp. 123-213.
3. Anderson. A.J., and E.A. Dawes,Occurrence. metabolism. meta-bolic role, and industrial uses orbacterial polyhydroxyalkanoatcs,Microbiol. Rev. 54:450-472(1990).
4. Srelnbucbet. A., and B. Fuchten-busch. Bacterial and other biolog-ical systems for polyester produc-tion, Trends Biatechnol,16,419-427 (1998).
5. De Koning. G.J.M.• H.M.M. VanBilesen, P.J. Lemstra, W. Hazen-berg. B. Withold, H. Preusting,J.G. Van der Galien, A. Schirmer.and D. Jendrossck, A biodegrad-able rubber by crosslinkingpoly(hydroxyalkanoate) fromPseudomonas oieovorons, Poly-mer 35:2090--2097 (1994).
6. Nawrath. C .. Y. Poirier, and C.R.Somerville, Plant polymers forbiodegradable plastics: cellulose,starch and polyhydroxyalka-
noares. Mol. Breeding l :I05-122(1995).
7. Poirier, Y., and C. Nawrath,Transgenic plants for the produc-tion of polyhydroxyalkanoates. afamily of biodegradable thermo-plastics and elastomers. in Trans-genic Plant Research, edited byK. Lindsey, Harwood AcademicPublishers, Amsterdam, TheNetherlands, 1998, pp. 201-208.
8. Poirier, Y., D.E. Dennis. K.Klomparens, and C. Somerville.Polyhydroxybutyrate. a biode-gradable thermoplastic, producedin transgenic plants, Science256,520-523 (1992).
9. Poirier, Y., L.A. Schechtman,M.M. Satkowski. L Noda, and C.Somerville, Synthesis of highmolecular weight poly([Rl-(-)-3-hydroxybutyrate) in transgenicArabidopsis thaliana plant cells,Int. J. BioI. Macromol. 17:7-12(1995).
10. Poirier, Y., D. Dennis, K. Klom-parens, C. Nawrath. and C.Somerville, Perspectives on theproduction of polyhydrcxyalka-noetes in plants, FEMS Microbi-al. Rev. 103:237-246 (1992).
II. Nawrath, c., Y. Poirier, and C.R.Somerville, Targeting of the poly-
hydrcx ybutyrate biosyntheticpathway to the plastids of Ara-bidopsis thaliana results in highlevels of polymer accumulation,Proc. Natl. Acad. Sci. USA91,12760-12764 (1994).
12. Valentin. H.E., D.L. Broyles, L.A.Casagrande, S.M. Colburn, W.L.Creely, P.A. DeLaquil, H.M. Fel-ton, K.A. Gonzalez. K.L.Houmiel. K. Lutke, D.A.Mahadeo, T.A. Mitsky, S.R. Pad-gette, S.E. Reiser, S. Slater, D.M.Stark, R.T. Stock, D.A. Stone,N.B. Taylor, G.M. Thorne, M.Tran, and K. Gruys, PHA produc-tion, from bacteria to plants. Int.J. Bioi. Macromol. (in press)(1999)
13. Minendorf, V., E.J. Robertson,R.M. Leech, N. Kruger, A. Stein-bucbel. and Y. Poirier, Synthesisof medium-chain-length polyhy-droxyalkanoates in Arabidopsisthaliana using intermediates ofperoxisomal fauy acid ~-ollida-tion, Proc. Natl. Acad. Sci. USA95,13397-13402 (1998).
14. Eccleston, V.S .. A.M. Cranmer.T.A. Voelker, and J.B. Ohlrogge.Medium-chain fatty acid biosyn-thesis and utilization in Brasstcanapus plants expressing lauroyl-acyl carrier protein tbioesterase.Plama 198:46-53 (1996). •
CNI.~14n1140 ................... Aa... AI. ..
TIt (M-11)4-712 ... • Fa: (14-11)4-712""'· E...t:J •..... - .....
For i IIoImaIkIn circle "08 on torm on lost poge
INFORM. VOl. 10, no. 8 (August 1999)





























