Embed Size (px)
Citation preview

BÖLÜM 2
BİTKİLERDE OKSİDATİF STRES

8
2. 1. Aktif Oksijen Türlerinin Oluşumu ve Antioksidant Sistem
Atomik oksijen yer kabuğunda en yaygın olarak bulunan elementtir. Atmosfer ve suyun
yapısında bulunan moleküler oksijen ise tüm aerobik yaşam formlarının varlığı için gereklidir.
Yeryüzünün mevcut oksijen rezervi, oksijenin suyun yapısından serbest hale geçmesini
sağlayan fotosentez olayı ile sağlanmıştır. Atmosferdeki oksijen miktarı fotosentez ve
oksijenli solunum olayları sayesinde hemen hemen sabit tutulur. Bunun dışında oksijen
atomları enzimatik olmayan olaylar dışında, oksigenazlar gibi enzimler yardımıyla çeşitli
organik bileşiklerin yapısına girebilir. Oksijenin ortamdaki konsantrasyonunun belirli bir
seviyenin üzerine çıkması, aerobik organizmalar için bile tehlikeli olabilir. Atmosferik oksijen
miktarının üzerindeki değerler, canlı sistemlerdeki bazı enzimlerin inaktif hale gelmesine
veya inhibisyonuna yol açabilir. Bunun dışında oksijen miktarının artması, fotosentetik
verimliliğinin azalmasına neden olur. Çünkü bu koşullarda ribuloz -1,5- bisfosfat
karboksilaz/oksigenaz enzimi C3 yolu yerine fotorespirasyon olayının hızlanmasını sağlar.
Diğer aerobik organizmalar gibi bitkiler de etkili bir şekilde enerji üretebilmek için oksijene
ihtiyaç duyarlar. Ancak oksijenin varlığı hücresel yapıların ve reaksiyonların sürekli oksidatif
bir tehdit altında olmasına da yol açmaktadır (Alscher et al., 1997). Bitki hücrelerinde
oksijenin suya tam olarak indirgenmesi, gerekli enerjinin açığa çıkmasını sağlarken; oksijenin
tam olarak indirgenememesi ise oldukça reaktif olan ve DNA, proteinler ve lipidler gibi
birçok makromoleküle zarar veren aktif oksijen türlerinin (AOT) oluşumuna neden
olmaktadır (Dat et al., 2000). Kloroplast ve mitokondrilerdeki redoks reaksiyonları sırasında,
bazı stres faktörleri nedeniyle elektronların asıl alıcı molekül yerine moleküler oksijene (O2)
verilmesi sonucu AOT’ler oluşmaya baslar. Ağır metaller, kuraklık, yüksek ve düşük sıcaklık,
mekanik yaralanma, UV ışık, fotoinhibisyona yol açan yüksek ışık yoğunluğu, patojen
infeksiyonu ve hava kirliliği gibi stres faktörlerinin de AOT oluşumuna neden olduğu
bilinmektedir (Bray et al., 2000; Öncel ve ark., 2000).
Oksijenin aktivasyonu kimyasal ve fiziksel mekanizmalarla gerçekleşebilir. Kimyasal
aktivasyon mekanizmasında oksijenin dışarıdan elektron alması söz konusudur. Oksijen temel
durumunda iken (O2, moleküler oksijen, atmosferik oksijen), sahip olduğu paylaşılmamış iki
elektronunun paralel spinli olmasından dolayı çok fazla reaktif değildir. Bu nedenle
paylaşılmamış elektronları ters spinli olan (antiparalel) bir molekülden, bir elektron çifti
alamaz. Çünkü Pauli prensibine göre, ancak ters spinli olan elektronlar bir elektron çifti
oluşturabilirler. Bir elektronun spinini ters çevirmek için gereken zaman fazla olduğu için

9
oksijen, başka moleküllere elektron vermek yerine kendisi elektron alma eğilimindedir. Bu
durumda moleküler oksijenin indirgenmesi, birbirini takip eden, her basamakta tek bir
elektronun oksijen tarafından alınmasını ve farklı AOT’lerin oluşumunu sağlayan dört
aşamadan ibarettir. Bu olaya “monovalent redüksiyon” adı verilir (Salin, 1987; Dat et al.,
2000; Edrewa, 2005). Monovalent oksijen redüksiyonu yoluyla oluşan çeşitli AOT’ler şekil 2.
1’de görülmektedir. Bu reaksiyon zincirinin ilk basamağı enerji gerektirmektedir. Ancak diğer
basamakları ekzotermik özelliğe sahiptir ve bu nedenle kendiliğinden veya kataliz yoluyla
gerçekleşebilmektedir (Vranova et al., 2003). Süperoksit radikali orta derecede reaktif bir
moleküldür. Dokulardaki yarı ömrü yaklaşık olara 2-4 μs’dir. Biyolojik membranları geçemez
ve hızla hidrojen perokside dönüştürülür (dismutasyon). Hem oksidasyon hem de redüksiyon
yeteneğine sahip olan süperoksit radikali bazen kendiliğinden de hidrojen perokside
dönüştürülebilir. Bir serbest radikal olmayan hidrojen peroksit de süperoksit radikali gibi orta
derecede reaktiviteye sahiptir. Ancak farklı olarak hem dokulardaki yarı ömrü daha uzundur
(yaklasık 1 ms) hem de oluştuğu yerden belli bir mesafeyi difüzyonla kat edebilmektedir
(Vranova et al., 2003). Hidrojen peroksit demir, bakır ve mangan gibi geçiş metallerinin
katalizörlüğünde “Haber-Weiss” veya “Fenton” reaksiyonu yoluyla oldukça reaktif olan
hidroksil radikalini oluşturabilir. Hem süperoksit radikalinin hem de hidrojen peroksidin canlı
hücreler için oluşturduğu asıl tehdit budur. Canlı sistemlerde hidroksil radikalini etkisiz hale
getirebilen herhangi bir enzimatik mekanizma bulunmadığı için bu molekülün dokulardaki
aşırı birikimi hücre ölümüne neden olmaktadır (Vranova et al., 2003).
Şekil 2. 1. Monovalent oksijen redüksiyonu ve oluşan AOT’ler (Edrewa, 2005).
Yukarıda bahsedilen farklı AOT’ler bitki hücrelerine farkı yollarla zarar verebilirler. Bu
mekanizmalar arasında bazı duyarlı enzimlerin inaktivasyonu, klorofil pigmentinin
parçalaması ve lipid peroksidasyonu sayılabilir. Lipid peroksidasyonu olayında bazı AOT’ler
ile hücresel membranların yapısında bulunan doymamış yağ asitleri arasında meydana gelen
etkileşimler sonucu lipid hidroperoksitleri oluşur. Ayrıca oramda bazı metal iyonlarının

10
bulunması alkoksil ve peroksil radikallerinin oluşumuna da yol açabilir. Sonuçta lipid
yapısında meydana gelen değişimler, membranların organizasyonunu, bütünlüğünü ve
geçirgenliğini de bozabilir. Ayrıca özelikle hidroksil radikali ile DNA arasında meydana
gelen reaksiyonlar, DNA molekülündeki baz sırasının değişmesine, zincir kırılmalarına ve bu
hasarların onarılma olasılığının azalmasına da neden olur. Proteinlerle süperoksid radikali
arasındaki etkileşimler de proteinlerin amino asit kompozisyonunda değişimlere, polipeptid
fragmentasyonuna, protein denatürasyonuna ve proteinlerin proteolizis olayına
duyarlılıklarının artmasına yol açar.
Oksijenin fiziksel aktivasyonu, klorofil molekülünün uyarılması (eksitasyonu) aracılığıyla
ışığa bağımlı reaksiyonlarla da sağlanabilmektedir (Hippeli et al., 1999). Bu reaksiyonda ışık
enerjisinin absorblanması ile klorofil molekülündeki bir elektron temel durumundan (Klo,
So), yüksek enerjili bir duruma (Klo*, S2) fırlatılmaktadır. Klorofil molekülü çok kısa bir süre
içerisinde bu elektronunun enerjisinin bir kısmını kaybederek daha düşük enerjili bir duruma
(S1) ve daha sonra yine enerji kaybı ile triplet (T*) duruma geçmektedir. S1 durumundan
triplet durumuna dönüşüm sırasında, klorofil molekülünün uyarılmış elektronunun spini de
değişmektedir. Enerjinin triplet klorofil molekülünden temel durumdaki oksijene transferi ile
klorofil molekülü temel durumuna dönerken; bu kez de temel durumdaki oksijenin
elektronlarından birinin spini değişmekte ve sonuçta 1O2 (singlet oksijen) oluşmaktadır
(Niyogi, 1999). Şekil 2. 2’de singlet oksijenin oluşum mekanizması görülmektedir.
2. 2. Bitki Hücrelerindeki AOT Kaynakları
2. 2. 1. Kloroplastlar
Bitki hücrelerinde AOT oluşturma yeteneğine sahip birçok bölge bulunmaktadır. Ancak
kloroplastların bitki hücrelerindeki AOT’lerin en büyük kaynağı olduğu kabul edilmektedir.
Kloroplastlardaki fotosistem I ve II’nin ışık toplayıcı sistemleri tarafından absorblanan ışık
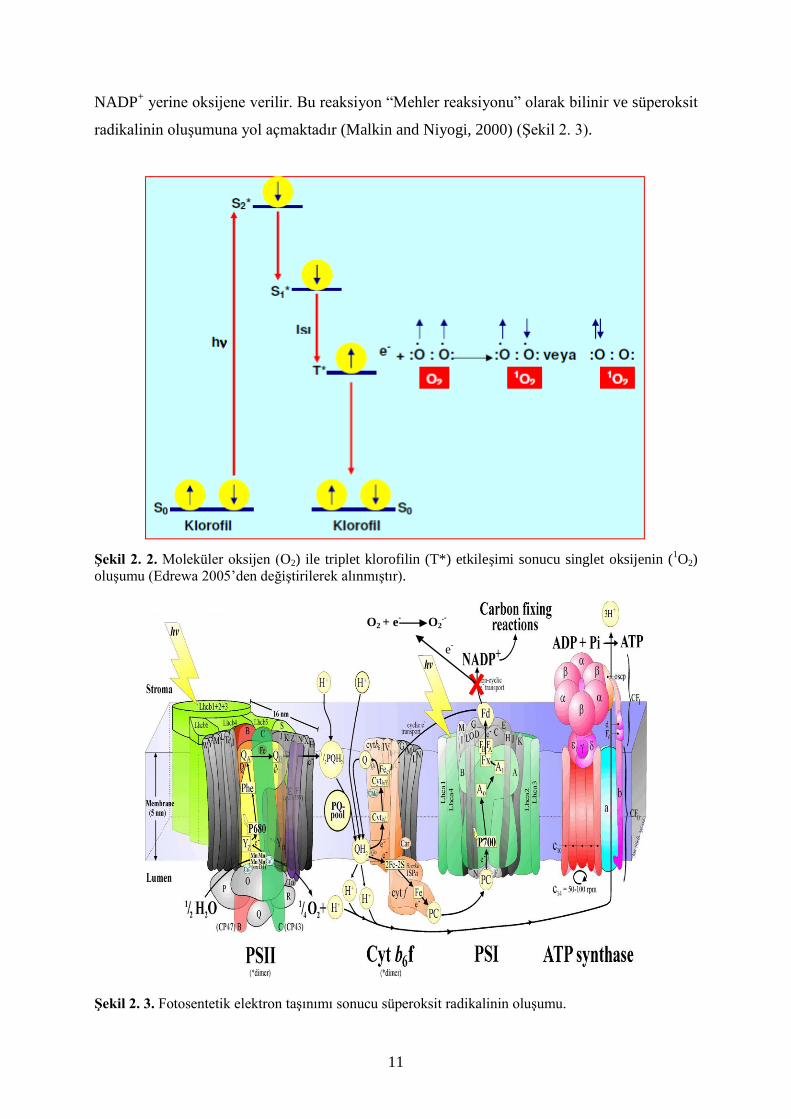
enerjisi, elektron taşınım reaksiyonları vasıtasıyla NADP+ molekülüne ulaştırılmakta ve
böylece oluşturulan NADPH karbon fiksasyon reaksiyonlarında kullanılmaktadır (Malkin and
Niyogi, 2000). Tuz stresi, kuraklık, ozon, yüksek ve düşük sıcaklıklar gibi karbon fiksasyon
reaksiyonlarını kısıtlayan koşullarda NADP+ molekülünün rejenerasyonu da kısıtlanır ve
fotosentetik elektron taşınım reaksiyonları sonunda elektronlar ferrodoksin aracılığı ile
11
NADP+ yerine oksijene verilir. Bu reaksiyon “Mehler reaksiyonu” olarak bilinir ve süperoksit
radikalinin oluşumuna yol açmaktadır (Malkin and Niyogi, 2000) (Şekil 2. 3).
Şekil 2. 2. Moleküler oksijen (O2) ile triplet klorofilin (T*) etkileşimi sonucu singlet oksijenin (1O2)
oluşumu (Edrewa 2005’den değiştirilerek alınmıştır).
Şekil 2. 3. Fotosentetik elektron taşınımı sonucu süperoksit radikalinin oluşumu.
e-
O2 + e- O2
.-
12
2. 2. 2. Peroksizomlar
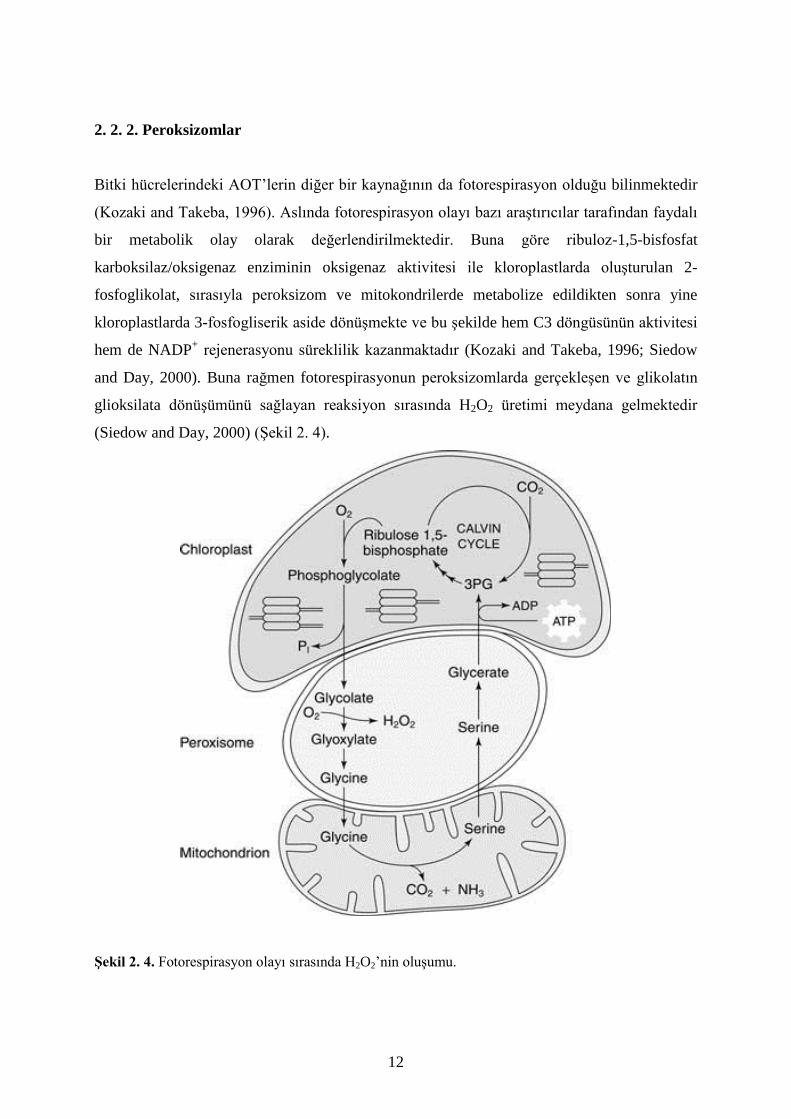
Bitki hücrelerindeki AOT’lerin diğer bir kaynağının da fotorespirasyon olduğu bilinmektedir
(Kozaki and Takeba, 1996). Aslında fotorespirasyon olayı bazı araştırıcılar tarafından faydalı
bir metabolik olay olarak değerlendirilmektedir. Buna göre ribuloz-1,5-bisfosfat
karboksilaz/oksigenaz enziminin oksigenaz aktivitesi ile kloroplastlarda oluşturulan 2-
fosfoglikolat, sırasıyla peroksizom ve mitokondrilerde metabolize edildikten sonra yine
kloroplastlarda 3-fosfogliserik aside dönüşmekte ve bu şekilde hem C3 döngüsünün aktivitesi
hem de NADP+ rejenerasyonu süreklilik kazanmaktadır (Kozaki and Takeba, 1996; Siedow
and Day, 2000). Buna rağmen fotorespirasyonun peroksizomlarda gerçekleşen ve glikolatın
glioksilata dönüşümünü sağlayan reaksiyon sırasında H2O2 üretimi meydana gelmektedir
(Siedow and Day, 2000) (Şekil 2. 4).
Şekil 2. 4. Fotorespirasyon olayı sırasında H2O2’nin oluşumu.

13
Ayrıca yine peroksizomlarda yağ asitlerinin β- oksidasyonu sırasında da H2O2 oluşumu söz
konusudur (Somerville et al., 2000). Peroksizomlardaki pürin metabolizması sırasında ise,
ksantinin ürik aside dönüştüğü reaksiyonda süperoksit radikali oluşmaktadır (del Rio et al.,
1998).
2. 2. 3. Mitokondriler
Bunun dışında hücrelerde büyüme ve enerji gerektiren diğer olayların stres etkisiyle
yavaşlaması veya durması halinde de, mitokondrilerdeki elektron taşınım sisteminin aşırı
indirgenmesi sonucu süperoksit radikali oluşabilmektedir (Purvis and Shewfelt, 1993).
Mitokondrilerde AOT oluşumunda rol oynayan asıl mekanizma siyanide duyarlı olmayan
alternatif oksidaz sistemidir.
2. 2. 4. Endoplazmik retikulum ve apoplast
Yine bitki hücrelerinin sitosol, endoplazmik retikulum, plazma membranı ve apoplast gibi
bölgelerinde bazı AOT’lerin oluştuğu bilinmektedir (Vranova et al., 2003). Endoplazmik
retikulumda bulunan ve mevalonik asit yolundaki hidroksilasyon rekasiyonlarını katalizleyen
sitokrom P450 gibi karışık fonksiyonlu oksigenazlar, oksijen atomlarının substratlara ilave
edilmesini sağlarlar. Bu reaksiyonlarda elektron vericisi olarak rol oynayan NAD(P)H
molekülleri süper oksid radikallerinin oluşumuna neden olabilir. Apoplastlar da bazı
AOT’lerin oluştuğu bölgeler olarak tanımlanmaktadır. Apoplast, patojen saldırıları sırasında
ilk hedef bölgedir. Bu nedenle patojenlere karşı savunma mekanizmalarının da ilk olarak aktif
hale geldiği alanlardır. Apoplastlardaki en yaygın biyosentetik mekanizma lignin
biyosentezidir. Bu reaksiyonlar sırasında fenilpropanoid öncüleri arasında hidrojen peroksit
vasıtasıyla ve peroksidaz grubu enzimlerin katalizörlüğünde çapraz bağlanmalar meydana
gelir. Bu reaksiyonlarda ihtiyaç duyulan NADH molekülleri hücre çeperindeki malat
dehidrogenaz enzimi ile oluşturulur ve muhtemelen daha sonra hidrojen peroksid oluşumunda
kullanılır. Bitki hücrelerinde AOT oluşumunun görüldüğü bölgeler ve ilgili reaksiyonlar şekil
2. 5’de verilmiştir.

14

15
2. 3. Bitkilerde antioksidant sistem
Bitkilerde, AOT’lerin toksik etkilerinden korunmak için enzimatik ve enzimatik olmayan
bileşenlerden oluşan etkili bir antioksidant savunma sistemi bulunmaktadır. Antioksidant
sistemin enzimatik bileşenleri arasında süperoksit dismutaz (SOD), askorbat peroksidaz
(APX), monodehidro askorbat redüktaz (MDHAR), dehidroaskorbat redüktaz (DHAR),
glutatyon redüktaz (GR), katalaz (CAT) ve peroksidaz grubundan olan diğer enzimler
sayılabilir. Enzimatik olmayan bileşenler ise askorbik asit, glutatyon, α-tokoferol ve
karotenoidlerdir (Dat et al., 2000; Foyer et al., 1994; Edrewa, 2005).
SOD, süperoksit radikalinin moleküler oksijen ve hidrojen perokside parçalanmasını sağlayan
reaksiyonu katalizleyen bir enzimdir (Bowler et al., 1992). SOD enzimi, süperoksidin fazla
miktarda üretildiği kloroplast, mitokondri ve sitosolde bulunur. Bunun dışında SOD’nin
peroksizomal, nuklear ve hücre dışı izoformları da belirlenmiştir (Ogawa et al., 1996). Oluşan
hidrojen peroksit, askorbat-glutatyon döngüsüne girer ve sonuçta su ve oksijene kadar
parçalanır (Sekil 2. 6). Askorbat-glutatyon döngüsünde bulunan enzimler APX, MDHAR,
DHAR ve GR’dir. Bu enzimlerden APX, indirgeyici molekül olarak askorbatı kullanarak
hidrojen peroksidi parçalarken; MDHAR ve DHAR enzimleri askorbatın rejenerasyonunu
sağlar. Askorbatın rejenerasyonunun sağlanması için görev yapan enzimlerden MDHAR
indirgeyici molekül olarak NADPH’yi, DHAR ise glutatyonu kullanmaktadır. Elektronunu
vererek oksitlenen glutatyon ise GR enziminin katalizlediği bir reaksiyonla yeniden
indirgenir. Askorbat-glutatyon döngüsü, kloroplastlarda, sitosolde, mitokondrilerde ve
peroksizomlarda bulunmaktadır (Bowler et al., 1992; Jimenez et al., 1997; Noctor and Foyer,
1998; Vranova, 2003). CAT’nin hücrelerin hidrojen peroksitten korunması konusundaki
etkinliği sınırlıdır. Feierabend and Enger (1986) ve Feierabend et al. (1992), bunun nedenini
CAT’nin büyük ölçüde peroksizomlarda bulunması, hidrojen perokside olan afinitesinin zayıf
olması ve ışık etkisiyle aktivitesini kaybetmesi gibi faktörlere bağlamışlardır (Foyer et al.,
1994). Dat et al., (2000) ise çeşitli stres faktörlerinin etkisiyle CAT proteini miktarındaki
azalmanın, translasyonun inhibisyonundan veya parçalanma hızındaki artıştan
kaynaklandığını rapor etmiştir. Ayrıca CAT aktivitesinin düşük sıcaklık stresi ve yüksek
sıcaklık şoku etkisiyle inhibe olduğu da belirlenmiştir (Dat et al., 1998; Lopez-Dalgado et al.,
1998). Bundan dolayı hidrojen peroksitin kloroplast ve diğer hücresel yapılarda
oluşturabileceği oksidatif zararlar, çok büyük ölçüde APX enzimi ile önlenmektedir (Foyer et
al., 1994).

16
Sekil 2. 6. Askorbat-glutatyon döngüsü (O2.-, süperoksit radikali; SOD, süperoksit dismutaz; H2O2,
hidrojen peroksit; APX, askorbat peroksidaz; AsA, askorbik asit; MDA, monodehidroaskorbik asit;
MDHAR, monodehidro askorbat redüktaz; DHA, dehidroaskorbik asit; DHAR, dehidroaskorbat
redüktaz; GSSG, okside glutatyon; GSH, indirgenmis glutatyon; GR, glutatyon redüktaz) (Mittler
2002’den degistirilerek alınmıstır)
AOT’lerin detoksifikasyonunu sağlayan diğer bir mekanizma da su-su döngüsüdür (Sekil 2.
7). Bu olayda iki molekül suyun fotolizi sonucu ortaya çıkan dört elektron, fotosistem II
aracılığı ile fotosistem I’e ulaştırılır. Bu elektronlardan iki tanesi, iki molekül O2’yi
indirgeyerek, iki molekül süperoksit radikalini oluşturur. Oluşan süperoksit radikalleri, SOD
ile hidrojen perokside indirgenir. Hidrojen peroksit ise APX ile suya indirgenir. Bu reaksiyon
sırasında oksitlenen askorbik asit de diğer iki elektronla yeniden indirgenir. Yani iki molekül
suyun fotolizi ile oluşan dört tane elektron, yine iki molekül suyun oluşturulmasında
kullanılmaktadır (Asada, 1999).

17
Sekil 2. 7. Su-su döngüsü (FS II, fotosistem II; FS I, fotosistem I; SOD, süperoksit dismutaz; H2O2,
hidrojen peroksit; APX, askorbat peroksidaz; MDA, monodehidro askorbik asit; AsA, askorbik asit)
(Asada, 1999).
2. 4. Bitkilerde Oksidatif Stresle Diğer Stres Tipleri Arasındaki İlişkiler
Antioksidant enzim aktivitelerinde meydana gelen değişimler, farklı bitki türlerinin biyotik ve
abiyotik stres faktörlerine karsı duyarlılık ve dayanıklılık dereceleri konusunda fikir
vermektedir (Foyer et al., 1994). Yapılan çeşitli çalışmalarda yüksek ışık yoğunluğu, kuraklık
stresi, tuz stresi, düşük ve yüksek sıcaklık stresi, ağır metal toksisitesi, UV ışık, herbisitler ve
hava kirleticileri gibi faktörlerin antioksidant enzimlerin aktivitelerinde değişikliklere yol
açtığı belirlenmiştir (Çakmak and Horst, 1991; Runeckles and Vaartnou, 1997; Giardi et al.,
1997; Dat et al., 1998; Smirnoff, 1998; Surplus et al., 1998; Savoure et al., 1999; Kingston-
Smith et al., 1999; Keleş ve Öncel, 2002; Ekmekçi ve Terzioglu, 2005). Bu yüzden stres
altındaki bitkilerde antioksidant sistemde meydana gelen değişimlerin incelenmesi önemlidir.
Birçok herbisitin bitkilerde AOT’lerin oluşumuna neden olduğu bilinmektedir. Örneğin
paraquatın elektronları PSI’den alarak ışıklı koşullarda süperoksit radikalinin oluşumuna
neden olduğu belirlenmiştir. Fungal bir toksin olan serkosporin adlı bileşik de ışıkla aktive
olarak singlet oksijeni oluşturmakta ve bu molekül de ciddi boyutlarda lipid
peroksidasyonuna neden olmaktadır. Asiflurfen gibi bazı herbisitler de klorofil
metabolizmasında rol oynayan bazı ara ürünlerin meydana gelmesine yol açar. Tetrapirol

18
yapısındaki bu ara ürünler de ışık altında singlet oksijeni meydana getirerek bitki hücrelerinin
ölümüne yol açar.
Endüstriyel faaliyetler veya tarımsal uygulamalar sonucu toprakta ve atmosferde birikim
gösteren metaller de bitkiler için zararlı olur. Bu tip metaller bitkilerde ya büyümeyi ve
biyosentetik reaksiyonları doğrudan doğruya inhibe ederek ya da AOT oluşumuna neden
olarak hasar oluştururlar. Örneğin yüksek konsantrasyonlarda bakır iyonlarına maruz kalan
bitkilerde lipid peroksidasyonu ve klorofil parçalanması gözlenmiştir. Bakır ve demir gibi
metaller aynı zamanda redoks bakımından aktif metaller olduğu için, Fenton reaksiyonunun
meydana gelmesine yol açarlar. Lipid peroksitleri de bakır elementinin lipoksigenaz enzimini
indüklemesi sonucu ortaya çıkar. Kadmiyuma maruz kalan bitkilerde de yine klorofil
miktarında azalma ve antioksidant enzimlerin inhibisyonu görülmüştür.
Ozon ve kükürt dioksit gibi atmosferik kirleticiler de bitki dokularında AOT’lerin oluşumuna
neden olmaktadır. Yapılan çalışmalar ozonun kükürt diokside göre bitkiler için daha büyük
tehlike olduğunu göstermiştir. Ozonun yüksek oksitleme potansiyeli ve bunun sonucunda
AOT’lerin oluşması bunun en büyük nedenidir. Ozona maruz kalan bitkilerde lipid
peroksidasyonu, klorofil parçalanması, PSII’deki D1 proteininin parçalanması ve RuBisCo
miktarında ve aktivitesinde azalmalar görülmüştür.
Güneşten çıkan UV-B ışınları (290-320 nm), stratosferdeki ozon tabakasının azalması sonucu
yer yüzüne kadar ulaşır. UV-B ışınlarının PSII’de neden olduğu hasarların sebepleri arasında
elektron taşınımının yavaşlaması ve özellikle D1 proteinindeki yapısal hasarlar sayılabilir.
UV-B ışınlarına maruz kalan bazı bitkilerde SOD gibi antioksidant enzimlerin
akümülasyonunun indüklendiği belirlenmiştir. İzole edilmiş tilakoid membranların UV-B ile
ışıklandırılması sonucu özellikle hidroksil radikalinin oluştuğu gözlenmiştir. UV-B ayrıca
bitkisel dokularda AOT’lerin oluşmasına neden olan bazı reaksiyonları başlatabilme
yeteneğine de sahiptir.
Tuz stresi de kloroplast ve mitokondrilerdeki önemli metabolik olayları etkilemektedir.
Yaprak mitokondri ve peroksizomları ile yapılan çalışmalar tuz stresinin hem süperoksit
radikali oluşumunu hızlandırdığını hem de MnSOD miktarını azalttığını göstermiştir. Tuz
stresine dayanıklı olan bitki genotiplerinde APX ve Cu/ZnSOD enzimlerinin aktiviteleri ile

19
askorbat miktarının arttığı, duyarlı bitkilerde ise antioksidant enzim aktivitelerinde herhangi
bir değişim olmaksızın H2O2 ve lipid peroksidasyonunun arttığı belirlenmiştir.
Kuraklık stresine maruz kalan bitkilerde absisik asit sinyali ile stomalar kapanır ve aşırı
derecede indirgenen fotosentetik aygıt oksidatif strese maruz kalabilir. Yapılan araştırmalar
kuraklığa maruz kalan bitkilerde protein sentezinin inhibe edildiği, kloroplastta bulunan
birçok enzimin inaktif hale geldiği, elektron taşınımının yavaşladığı, membran geçirgenliğinin
azaldığı ve özellikle H2O2’yi parçalayan enzimlerin aktivitesinin arttığı belirlenmiştir.
Düşük sıcaklıklarla yüksek ışık yoğunluğu bitkilerde fotooksidatif strese neden olur. Bu
olayın temelinde fotoinhibisyon olarak bilinen olay yatmaktadır. Fotoinhibisyon, fotosentezin
ışığa bağımlı reaksiyonlarının yine ışık etkisiyle inhibe olmasıdır. Fotoinhibisyon doğal bir
olaydır ve absorblanan ışık enerjisi miktarının, bu enerjinin fotosentetik elektron taşınım
reaksiyonlarındaki kullanım hızından daha fazla olması durumunda ortaya çıkmaktadır
(Savitch et al., 2000). Bu durumda fotosistem II’nin aşırı derecede indirgenmesi söz
konusudur (Aro et al., 1993; Long et al., 1994; Vasilikiotis and Melis, 1994). Fotoinhibisyon
olayı çok farklı çevresel stres faktörlerinin etkisiyle veya ortamın ışık şiddetinin aniden
artmasıyla meydana gelebilmektedir (Demmig-Adams et al., 2004). Soğuk ve ılıman bölge
bitkileri açık havalarda sıklıkla düşük sıcaklıklara maruz kalırlar ve bu koşullar
fotoinhibisyonun şiddetini artırır (Krause, 1994). Düşük sıcaklıkların fotosentez olayının
fotoinhibisyona karsı duyarlılığını artırdığı ve sonuçta düşük ışık yoğunluklarında bile
fotoinhibisyonun indüklenebileceği rapor edilmiştir (Xu et al., 1999). Birçok bitkide
fotosistem II’nin fotoinhibisyona, fotosistem I’e göre daha duyarlı olduğu bildirilmiştir
(Sonoike, 1996; Terashima et al., 1998; Ivanov et al., 1998). Fotosistem II reaksiyon
merkezlerinin ışık etkisiyle zarara uğrama hızı onarım hızını aştığı zaman geri dönüşümü
olmayan veya kronik fotoinhibisyon söz konusudur (Baroli and Melis, 1996; Melis, 1999). Bu
onarım mekanizması, zarar gören D1 polipeptidlerinin proteolitik olarak fotosistem II’nin
yapısından uzaklastırılması ve yeni sentezlenen D1 polipeptidlerinin fotosistem II yapısına
ilavesinden ibarettir (Savitch et al., 2000). Geri dönüşümlü fotoinhibisyonda ise, enerjinin
tüketim kapasitesi ile fotosistem II aktivitesi arasındaki dengenin korunması amacıyla,
fotosistem II’nin aktivitesinin azalması söz konusudur. Bu yüzden geri dönüşümlü
fotoinhibisyon, koruyucu bir mekanizma olarak kabul edilmektedir (Öqüist et al., 1992;
Osmond, 1994; Adams et al., 1995a, b).

20
Singlet oksijen, hidrojen peroksit, süperoksit ve hidroksil radikalleri gibi AOT’lerin
oluşumunun, metabolik olaylar üzerinde zararlı etkilere sahip olduğu bilinmektedir. Ancak
Asada (1993), bu moleküllerin normal hücresel reaksiyonlar sonucunda da oluştuğunu rapor
etmiştir (Foyer et al., 1994). Edrewa (2005) ise AOT’lerin düşük konsantrasyonlarda hücresel
fonksiyonların yerine getirilebilmesi için oldukça önemli olduğunu belirtmiştir. Bunun
dışında AOT’lerin bitkilerde patojen infeksiyonlarından korunma ve sinyal iletiminde de
önemli roller oynadığı belirlenmiştir (Grant and Loake, 2000; Van Breusegem et al., 2001;
Neill et al., 2002). Normal koşullardaki bir bitki hücresinde metabolik reaksiyonlar sırasında
AOT’lerin oluşumu ile parçalanması olayları kontrol altında tutulmaktadır. Oksijen toksisitesi
ise kontrol edilemeyen aşırı üretim veya savunma mekanizmalarının yetersizliği sonucu
oluşmaktadır (Edrewa, 2005).









![Everglades City School - District School Board of Collier County · 2015-11-03 · Everglades City School 415 SCHOOL DR, Everglades City, FL 34139 [ no web address on file ] School](https://img.pdfslide.net/doc/110x75/5f3397183d00ce1f415a65d1/everglades-city-school-district-school-board-of-collier-county-2015-11-03-everglades.jpg)

















