Embed Size (px)
Citation preview

Breeding Seasons of North Scandinavian Starlings (Sturnus vulgaris): Constrained by Food orTime?Author(s): Per LundbergSource: Journal of Animal Ecology, Vol. 56, No. 3 (Oct., 1987), pp. 847-855Published by: British Ecological SocietyStable URL: http://www.jstor.org/stable/4952 .
Accessed: 02/05/2014 06:41
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
British Ecological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal ofAnimal Ecology.
http://www.jstor.org
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

Journal of Animal Ecology (1987), 56, 847-855
BREEDING SEASONS OF NORTH SCANDINAVIAN STARLINGS (STURNUS VULGARIS):
CONSTRAINED BY FOOD OR TIME?
BY PER LUNDBERG
Department of Animal Ecology, University of Umea, S-901 87 Umea, Sweden
SUMMARY
(1) The breeding seasons of two North Scandinavian starling (Sturnus vulgaris L.) populations with considerably different annual cycles were investigated in relation to their food supply. Starlings of Andoya, northern Norway, regularly reared two broods each year and were sedentary, whereas starlings in Umea, northern Sweden, reared a single brood and were migratory.
(2) The potential prey fauna extracted from soil core samples revealed a parallel seasonal variation at both locations, both with respect to prey abundance and biomass. Moreover, the spatial distribution of potential prey did not show any consistent variation over time at either location.
(3) Starlings in Ume'a terminated their breeding season when the food supply began to decrease, but starlings at Andoya reared an additional brood, albeit smaller than the first one.
(4) It is suggested that food supply alone cannot account for differences in the breeding patterns. In addition, the inter-relation with other annual processes, such as moult and migration, might well increase the costs of an additional brood, thus ultimately terminating the breeding season.
INTRODUCTION
Theories on the timing of breeding seasons of birds have generally emphasized that food supply is the major ultimate factor regulating the onset and termination of breeding (e.g. Lack 1968; Perrins 1970; Immelmann 1971; Murton & Westwood 1977). The empirical evidence supporting this concept is now overwhelming (see Murton & Westwood 1977 for a review).
Many bird species in the temperate zone rear only a single brood each year, but a variety of species produce two or more broods (Lack 1950, 1968; Immelmann 1971; Perrins & Birkhead 1983). Several factors are involved in the regulation of the number of broods reared in a season; these include food availability, the duration of parental care and the costs to the parents. If the breeding season is prolonged by commitment to a second, or perhaps, a third, brood, the parents (or in some cases the juveniles) will have less time to complete their post-nuptial or post-juvenile moult and/or migratory preparations. Lewis (1975) pointed out that it is not only environmental conditions per se (such as the food supply), but also the interrelation between breeding and other processes
Present address: Department of Wildlife Ecology, Swedish University of Agricultural Sciences, S-901 83 Umea, Sweden.
847
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

Breeding seasons of starlings
in the birds' annual cycle, such as moult, that might influence the timing and length of the breeding season. This idea was discussed further by Gwinner (1981), Lundberg & Eriksson (1984) and Lundberg & Silverin (1986). Greenberg (1980) has also analysed the relationship between time and migration in more detail.
In a previous paper (Lundberg & Silverin 1986) we suggested that time in the context of all other annual processes might be a factor limiting the starlings' possibilities of rearing two broods in northern Scandinavia. In this paper, empirical data supporting this hypothesis are presented and it is shown that the seasonal variations in food supply probably cannot fully account for the different breeding systems of Scandinavian starling populations.
MATERIALS AND METHODS
Study area and the starling populations
Starlings at two northern Scandinavian locations were studied; Andoya, northern Norway (69?16'N, 16?05'E) and Umea, northern Sweden (63?50'N, 20?16'E). At Andoya starlings breed in nest-boxes close to farms and villages, particularly in the vicinity of Andenes. Here, two broods are reared regularly by about 70% of the pairs (Lundberg & Silverin 1986). The total breeding season lasts from early May to mid-August. After the completion of the second brood, starlings at Andoya go through a complete moult which lasts till the end of November or the beginning of December. Starlings at Andoya are sedentary.
In the study area in Umea, starlings breed in nest-boxes and occasionally in natural nest-holes in an old park close to the river Umealven. Their single brood is completed by the end of June, and subsequently the birds go through the post-nuptial (or post-juvenile) moult before they migrate by the end of September, mainly to British wintering grounds.
Lundberg & Silverin (1986) give further details on the two populations for the years 1981-84 inclusive. Karlsson (1983) should also be consulted for a thorough description of Scandinavian starlings' breeding habits.
Methods
In 1983-84 the food supply for breeding starlings was estimated at both locations. Unfortunately, I have no detailed information on the prey choice of the two starling populations at breeding time. However, Feare (1984) has summarized available data on food selection in the species from a variety of locations in Europe. The diet of breeding starlings varies considerably between locations depending on the dominant prey species in the soil surface. I therefore estimated the total abundances and biomasses of the soil surface fauna. Observations on feeding starlings at both locations revealed that at least leatherjackets (Tipulidae larvae), smaller dipteran larvae, caterpillars and lumbricids constitute a significant part of the items brought to the young and consumed by starlings at that time of the year.
Breeding starlings (as well as juveniles when they had fledged) at both locations changed their feeding sites somewhat during the course of the breeding season (Table 1). The changes of feeding sites were in all cases the same for both adults and juveniles and they did not differ in feeding sites at any time of the year (%2-test, P = 0-46). At Andoya the wrack belt on the sea shore was visited frequently at the beginning of the breeding season. From May onwards, however, and throughout the breeding season, short-grass meadows
848
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

P. LUNDBERG
TABLE 1. The distribution of feeding visits by breeding and juvenile starlings to the different feeding sites, the mean distance to cover between nests and feeding areas, and the dispersion of prey of Andoya and Ume'a. The frequency of visits to different sites is indicated by mean monthly percentages. The dispersion of prey is expressed as X2-values according to Elliott (1979) (in parentheses). N indicates the number of
soil samples taken. Means are given with their standard deviations
Andoya
Feeding visits (%) Shore
Wrack belt (m) meadows (m)
62+ 13 (281)t 35+ 18 (101)t 4?+ 15 8+15
11+9 7+3
27 +7 (74)t 38 + 21 (91)t 34+ 12 (66)t 20? 6 (127)t 44?+17 (50)t 53 + 7 (68)t
Feeding visits (%)
Meadows (m)
17+5 29+ 12 (133)t 12 +?4 (80)t 31 + 17 (59)t 28 +8 52 + 11 (185)t
Grazed No. of meadows (m) visits
11+3 103 27 + 7 (41)t 144 62+ 16 (32)* 261 72+ 24 (48)t 83 45 + 12 (22)* 176 40?+ 19 (104)t 101
Umea
Grazed No. of fields (m) visits
65+ 13 (73)t 78 59+ 10 (10)* 189 81 +28 (19)* 206 66 ? 12(144)t 111 72 + 26 (72)t 95 48?24(116)t 164
* Random distribution. t Clumped distribution.
Umea EIZZi Andoya :
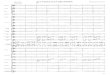
A M J J A S Month
FIG. 1. Seasonal changes in prey abundance (means + S.E.) within the upper 3 cm soil surface at And0ya (0) and in Umea (O). Bars above the graphs indicate incubation (shaded) and nestling
(black) periods at the two locations.
849
April May June July August September
April May June July August September
River slope
18 +5 12+8 7+2 3+1 0 0
N
57 180 60 58 60 60
N
30 59 60 60 30 60
Mean distance to cover
245 ? 32 168 +40 89+ 11
122 + 29 111 +44 162 +23
Mean distance to cover
270 + 54 84 + 27
133+ 13 104 + 10 183 + 35 120 + 26
30-
E a
a) Q- a) 20-
-a
-a -
a)
a -a 10 r-
a
L-
0-
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions
Breeding seasons of starlings
Ume8 L1 _ And0yo sL -
(a) I
300-
200 -
-~ 00- -)
EZ 0 -
0~
CP' 0 E
0
20
10 -
0-
(b)
A M J J A S
Month
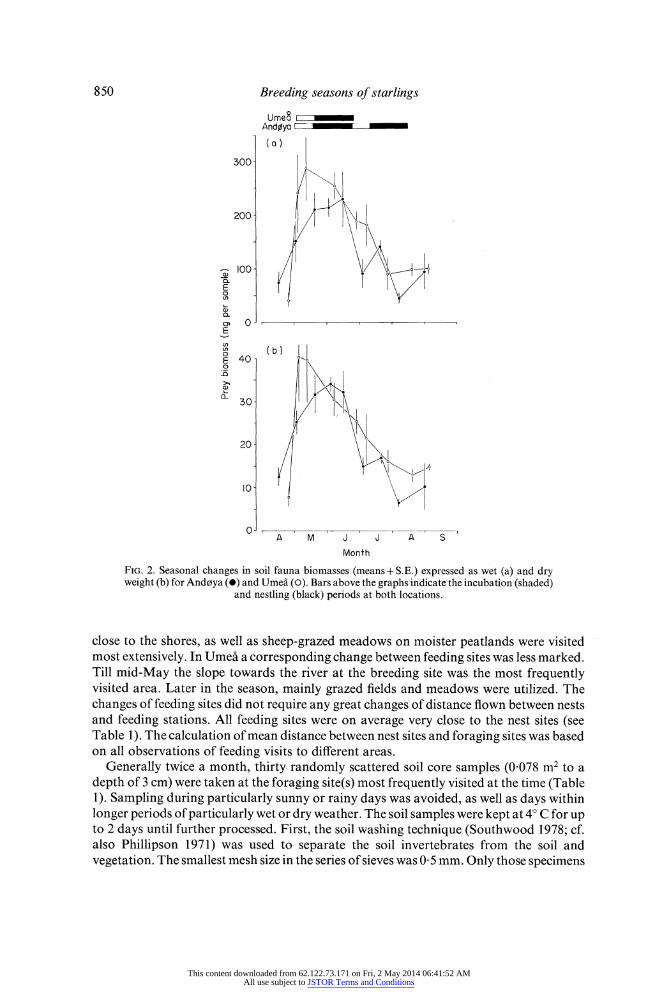
FIG. 2. Seasonal changes in soil fauna biomasses (means + S.E.) expressed as wet (a) and dry weight (b) for Andoya (0) and Umea (0). Bars above the graphs indicate the incubation (shaded)
and nestling (black) periods at both locations.
close to the shores, as well as sheep-grazed meadows on moister peatlands were visited most extensively. In Ume'a a corresponding change between feeding sites was less marked. Till mid-May the slope towards the river at the breeding site was the most frequently visited area. Later in the season, mainly grazed fields and meadows were utilized. The changes of feeding sites did not require any great changes of distance flown between nests and feeding stations. All feeding sites were on average very close to the nest sites (see Table 1). The calculation of mean distance between nest sites and foraging sites was based on all observations of feeding visits to different areas.
Generally twice a month, thirty randomly scattered soil core samples (0-078 m2 to a depth of 3 cm) were taken at the foraging site(s) most frequently visited at the time (Table 1). Sampling during particularly sunny or rainy days was avoided, as well as days within longer periods of particularly wet or dry weather. The soil samples were kept at 4? C for up to 2 days until further processed. First, the soil washing technique (Southwood 1978; cf. also Phillipson 1971) was used to separate the soil invertebrates from the soil and vegetation. The smallest mesh size in the series of sieves was 0-5 mm. Only those specimens
850
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

(a) 100- ^- Others
_ ::
M Lumbricidae
-; ~ ' 3Lepidoptera larvae
all monthy smDiptera larvae (others)
/ _Tipulidae larvae 0
= (b) o 100- e> _ Others
0-
50 : ; ::: E
-
- - .. Lumbricidoe
50 0 i 5 Lepidoptera larvae
l bDiptera larvae (others) 0O t - -...... .Tipulidae larvae
FIG. 3. The seasonal changes in soil fauna composition based on number of specimens caught in all monthly samples from And0ya (a) and Umea (b).
exceeding 1 mm in length were retained since smaller items were probably not exploited by the starlings (cf. Tinbergen 1976). Specimens found during this procedure were preserved in 70% ethanol. The rest of the soil core was kept in 80% ethanol for at the most 2 weeks until further analysis. The remainder from the sieving was then hand-sorted. Gastropods were ignored since they apparently play only a minor role in the diet of breeding starlings (Feare 1984).
Prey abundanc es were expressed as the number of specimens per sample. Biomass was expressed both in wet weight (the specimens dried on a filter paper 30 s before weighing) and dry weight (specimens dried at 60 ?C for 24 h before weighing). The dispersion of the potential prey items in the soil was calculated by a %2-test according to Elliott (1979).
RESULTS
Starlings at both locations started egg-laying at the beginning of May and the juveniles from the first brood fledged by the end of June (Figs 1 and 2). However, more than 70% of the pairs at Andoya laid a second clutch soon after the first one was completed, i.e. the total breeding time for the Norwegian birds was about 1-5 months longer than for the starlings in Umea. The size of the second brood at Andoya was significantly smaller than the first one (Lundberg & Silverin 1986).
The prey abundances showed clear seasonal variations at both locations (Fig. 1). This was particularly pronounced during the beginning of the breeding season in Umei where
851 P. LUNDBERG
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

Breeding seasons of starlings (a )
100 ~ Others
Lepidoptera larvae
50-^ g ~~ ~Coleaptera larvae
a - _ Diptera larvae (others)
,, ~~~ 0_~~ ~Tipulidae larvae
?. (b)__ > 00- Others
i;:-';''~i... ;',:,I 2Lumbricidae
0-
-E m'::;p?::u , . I . I - L
I I1,1
Lepidoptera larvae
Coleoptera larvae
Diptera larvae (others)
Tipulidae larvae
A M J J A Month
FIG. 4. The seasonal changes in soil fauna composition based on dry weight of all monthly samples from Andoya (a) and Umea (b).
the prey abundance increased rapidly. After reaching a peak in the middle of June, prey abundances decreased throughout the rest of the breeding season. At Andoya the change in prey abundances at the beginning of the breeding season was not as marked as in Umea. Instead, there was a marked decrease in prey abundance during July to very low values in August. The seasonal variations in soil fauna biomasses were similar to those in prey abundances (Fig. 2), the difference in biomass between Andoya and Umea, however, being smaller particularly at the beginning and the end of the season. At both locations the soil fauna biomass reached a peak between early May and the middle of June. I
The composition of the soil fauna varied throughout the breeding season at both locations. At Andoya, dipteran larvae (including Tipulidae) were the most abundant groups during all months, whereas coleopteran larvae (mainly Elateridae, Curculionidae and Staphylinidae) increased in abundance towards the end of the season (Fig. 3a). In Ume'a the abundance of Tipulidae larvae showed a marked decrease through the season. Lumbricids were the second most abundant group, dominating in August (Fig. 3b).
At both locations, lumbricids and leatherjackets were the major components of the soil fauna biomass throughout the season. At Andoya there was a marked decrease in leatherjacket biomass from mid-June, but this was not the case at Ume'a, where the biomass of leatherjackets did not change very much during the course of the breeding season (Fig. 4).
The dispersion of potential prey changed somewhat between months as well as between feeding sites. Most of the time, the soil fauna was strongly clumped (X2-test), or in some cases randomly distributed. This was true both at Andoya and in Ume'a. The degree of
50-
852
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

patchiness did change with time, but without any consistent seasonal pattern (Table 1). Again, no consistent differences between the two locations were found.
DISCUSSION
The two starling populations studied live under considerably different environmental conditions, and differ also in the organization of their annual cycles (Lundberg & Eriksson 1984; Lundberg 1985). The food supply during the possible breeding season from April to August was, however, surprisingly similar. The peaks in prey abundances and biomasses coincided well with the time when starlings at both locations had young of the first brood in the nests. The decline in food supply after June might then explain the termination of breeding. However, despite the fact that the food supply decreased towards the end of the summer, starlings at Andoya laid another clutch, albeit smaller than the first one. Moreover, the distance flown by the adults between nest-sites and feeding areas, as well as the dispersion of prey they collected, did not change significantly during the season.
Thus, these findings give circumstantial evidence for the hypothesis that the food supply alone cannot fully account for the differences in breeding system between the populations. Possibly, starlings at Umea, which cannot survive the winter at the breeding grounds, cannot rear a second brood because of limitations of time available rather than restrictions imposed by the food supply. Adult and juvenile starlings in Ume'a have to complete a post-nuptial or post-juvenile moult and prepare for the migration before the end of September. Starlings at Andoya on the other hand, can continue the post-nuptial or post-juvenile moult well into November and December after the completion of the second clutch (Lundberg & Eriksson 1984). One could imagine that starlings in Umea might have been able to overcome the problem of the short breeding and moulting season by postponing moult until after migration. Some northern populations of tropical migrants, e.g. the whitethroat (Sylvia communis Latham), differ from their southern conspecifics by either suspending moult or excluding it at the breeding grounds (Swann & Baillie 1979; Svensson 1984). It is not clear why migrant starlings in northern Scandinavia have not adopted this strategy, which might allow for an additional brood.
The occurrence of second clutches in the starling in relation to the food supply, was investigated by Dunnet (1955) and Anderson (1961). Dunnet suggested that the decline of the chief prey species (leatherjackets) by the end of the summer would allow only early breeders to rear a second brood, and then only in years with comparatively high prey densities. Thus, he argued that the food supply solely determined the occurrence of second clutches. Later, Anderson (1961) pointed out that Dunnet's interpretation of the data was questionable. The food supply is certainly important for additional broods (e.g. Perrins 1965), but the point here is that the costs of prolonging the breeding season involve neglecting other annual processes vital for survival and future reproduction.
The data from Andoya show that a lowered food supply does not necessarily mean that a second brood is impossible, especially if the brood size is adjusted accordingly. Apparently, the extra costs in terms of decreased adult (and juvenile) survival are relatively small. This is in line with the time allocation hypothesis suggested by Greenberg (1980). He concluded that migrants gain in survival at the expense of annual productivity, whereas residents maximize productivity at the expense of annual survival. Thus, for migrants, any increase in the length of the breeding season (by additional broods) will
P. LUNDBERG 853
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

Breeding seasons of starlings
decrease the amount of time spent in the wintering areas resulting in lowered survival chances.
The data on food supply presented here should of course be interpreted with caution. The success of a predator in utilizing its prey depends not only on the prey density (or biomass), but also on the prey availability. The total food supply was similar throughout the summer at Andoya and in Umea, but there could have been differences for instance in the vegetation cover or soil structure. However, at both locations starlings foraged in similar habitats, i.e. short-grass meadows or cultivated fields where the vegetation never became particularly high or dense. The soil samples were also taken from the surface layer which is likely to be readily accessible to foraging starlings.
Moreover, one could argue that the shifts in preferred feeding sites which occurred at both locations during the course of the season might have increased the costs in terms of energy expenditure significantly but differently at the two sites. Since neither average distance flown between nests and feeding sites, nor the dispersion of prey, changed significantly, it seems doubtful that the differences in the number of broods could be explained by differences in food supply only. Moreover, the populations in this study fit well to the general pattern of number of clutches in Scandinavian starlings (Karlsson 1983). If small differences in travel costs and prey dispersion patterns have any significant affect on the possibility of rearing additional broods, one would expect to find great local variation in the number of clutches laid, both in northern Norway and in northern Sweden. Since this is not the case (Karlsson 1983), I find it difficult to reject the time-limit hypothesis for that reason.
On the basis of the time-limit hypothesis, one would expect second broods to occur less frequently (or even be absent) in most migratory starling populations. This seems to be true for most Scandinavian populations. Practically no second broods occur in Sweden and Finland where almost all populations are migratory (Karlsson 1983; Fliege 1984). Along the Norwegian coast on the other hand, several populations are sedentary and also regularly rear two broods each year (Karlsson 1983). On the other hand, in mid-Europe second broods are rather common, albeit most populations are migratory, at least for short distances (Karlsson 1983; Feare 1984). However, the data are incomplete and the occurrence of second broods varies considerably between populations as well as between years (Karlsson 1983). Possibly, early breeders do have the time for another clutch, whereas later ones manage to rear only a single one. This option would not generally be open to starlings farther north (as in Scandinavia) where the summers are too short anyway. There are also indications of partial migration among west European starling populations (Feare 1984), which could mean that sedentary individuals would have time for another brood, whereas migrants would be able to rear only a single one. This could also explain why only a proportion of the pairs rear two broods, as well as the year-to-year variations. These ideas, however, remain to be tested.
ACKNOWLEDGMENTS
K. Danell, L.-O. Eriksson, K. Leonardsson, C. Otto and two anonymous referees improved the manuscript with their comments. J. Lundberg, E.-B. Schildt, J. Linner and M.-L. L6fgren gave technical assistance. Financial support was received from the Swedish Natural Science Research Council (grant to L.-O. Eriksson).
854
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions

REFERENCES
Anderson, A. (1961). The breeding of the starling in Aberdeenshire. The Scottish Naturalist, 70, 60-74. Dunnet, G. M. (1955). The breeding of the starling Sturnus vulgaris in relation to its food supply. Ibis, 97, 619-
662. Elliott, J. M. (1979). Some methods for the statistical analysis of samples of benthic invertebrates. Freshwater
Biological Association, Scientific Publication No. 25. Titus Wilson & Sons Ltd., Kendal. Feare, C. (1984). The Starling. Oxford University Press, Oxford. Fliege, G. (1984). Das Zugverhalten des Stars (Sturnus vulgaris) in Europa: Eine Analyse der Ringfunde. Journal
fur Ornithologie, 125, 393-446. Greenberg, R. (1980). Demographic aspects of long-distance migration. Migrant Birds in the Neotropics:
Ecology, Behavior, Distribution, and Conservation (Ed. by A. Keast & E. S. Morton), pp. 493-504. Smithsonian Institution Press, Washington, D.C.
Gwinner, E. (1981). Annual rhythms: perspective. Handbook of Behavioral Neurobiology, Vol. 4 (Ed. by J. Aschoff), pp. 381-389. Plenum Publishing Corporation.
Immelmann, K. (1971). Ecological aspects of periodic reproduction. Avian Biology, Vol. 1, (Ed. by D. S. Farner & J. R. King), pp. 341-389. Academic Press, New York.
Karlsson, J. (1983). Breeding of the starling (Sturnus vulgaris). Unpublished Ph. D. thesis, University of Lund, Lund, Sweden.
Lack, D. (1950). The breeding seasons of European birds. Ibis, 92, 64-78. Lack, D. (1968). Ecological Adaptations for Breeding in Birds. Methuen & Co. Ltd. Lewis, R. A. (1975). Reproductive biology of the white-crowned sparrow (Zonotrichia leucophrys pugetensis
Grinnell). I. Temporal organization of reproductive and associated cycles. Condor, 77, 46-59. Lundberg, P. (1985). The annual cycle organization in birds-a case study of two North Scandinavian starling
Sturnus vulgaris populations. Unpublished Ph. D. thesis, University of Umea, Umea, Sweden. Lundberg, P. & Eriksson, L.-O. (1984). Postjuvenile moult in two North Scandinavian starling Sturnus vulgaris
populations-evidence for difference in the circannual time-program. Ornis Scandinavica, 15, 105-109. Lundberg, P. & Silverin, B. (1986). Breeding cycles of two North Scandinavian starling Sturnus vulgaris
populations and the circannual testicular and gonadotrophin cycles. Ornis Scandinavica, 17, 18-23. Murton, R. K. & Westwood, N. J. (1977). Avian Breeding Cycles. Clarendon Press, Oxford. Perrins, C. M. (1965). Population fluctuations and clutch-size in the great tit Parus major L. Journal of Animal
Ecology, 34, 601-647. Perrins, C. M. (1970). The timing of birds' breeding seasons. Ibis, 112, 242-255. Perrins, C. M. & Birkhead, T. R. (1983). Avian Ecology. Blackie & Son Ltd, Glasgow. Phillipson, J. (1971). Methods of study in quantitative soil ecology: population, production and energy flow. IBP
Handbook No. 18. Blackwell Scientific Publications, Oxford. Southwood, T. R. E. (1978). Ecological Methods. 2nd edn. Chapman and Hall, London. Svensson, L. (1984). Identification Guide to European Passerines. 3rd edn. Stockholm. Swann, R. L. & Baillie, S. R. (1979). The suspension of moult by trans-Saharan migrants in Crete. Bird Study, 26,
55-58. Tinbergen, J. M. (1976). How starlings (Sturnus vulgaris L.) apportion their foraging time in a virtually single-
prey situation on a meadow. Ardea, 64, 155-170.
(Received 3 March 1986)
P. LUNDBERG 855
This content downloaded from 62.122.73.171 on Fri, 2 May 2014 06:41:52 AMAll use subject to JSTOR Terms and Conditions