Embed Size (px)
Citation preview

1
BrpA is Involved in Regulation of Cell Envelope Stress 1
Responses in Streptococcus mutans 2
3
4
J. P. Bitoun1, S. Liao1, X. Yao1, S.-J. Ahn2, R. Isoda2, A. H. Nguyen1, L. J. 5
Brady2, R. A. Burne2, J. Abranches3, and Z. T. Wen1, 4* 6
7
1Department of Oral and Craniofacial Biology, School of Dentistry, Louisiana State 8
University Health Sciences Center, New Orleans, LA 70119, USA; 2Department of Oral 9
Biology, College of Dentistry, University of Florida, Gainesville, FL 32610, USA; 10
3Center for Oral Biology, University of Rochester School of Medicine and Dentistry, 11
Rochester, NY 14610; and 4Department of Microbiology, Immunology, and Parasitology, 12
School of Medicine, Louisiana State University Health Sciences Center, New Orleans, 13
LA 70112. 14
15
16
Running title: Cell envelope stress response in Streptococcus mutans 17
18
Contact Information: 19
Email: [email protected] 20
Phone: 5049418465 21
Fax: 5049418319 22
23
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.07823-11 AEM Accepts, published online ahead of print on 10 February 2012
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

2
Abstract 24
25
Previous studies have shown that BrpA plays a major role in acid and oxidative stress 26
tolerance and biofilm formation by S. mutans. Mutant strains lacking BrpA also display 27
increased autolysis and decreased viability, suggesting a role for BrpA in cell envelope 28
integrity. In this study, we examined the impact of BrpA-deficiency on cell envelope 29
stresses induced by envelope-active antimicrobials. As compared to the wild-type strain 30
UA159, the BrpA-deficient mutant (TW14D) was significantly more susceptible to 31
antimicrobial agents, especially lipid II inhibitors. Several genes involved in 32
peptidoglycan synthesis were identified by DNA microarray analysis as down-regulated 33
in TW14D. Luciferase reporter gene fusion assays also revealed that expression of brpA 34
is regulated in response to environmental conditions and stresses induced by exposure to 35
sub-inhibitory concentrations of cell envelope antimicrobials. In a Galleria mellonella 36
(wax worm) model, BrpA-deficiency was shown to diminish the virulence of S. mutans 37
OMZ175, which unlike S. mutans UA159, efficiently kills the worms. Collectively, these 38
results suggest that BrpA plays a role in the regulation of cell envelope integrity and that 39
deficiency of BrpA adversely affects the fitness and diminishes the virulence of 40
OMZ175, a highly invasive strain of S. mutans. 41
42
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

3
Introduction 43
The oral cavity is a dynamic environment in which frequent and often rapid 44
fluctuations in pH and the concentrations of antimicrobial agents and other stressors 45
occur. Dental care products, such as toothpastes and mouth rinses, contain a variety of 46
antibacterial compounds, including hydrogen peroxide, sodium lauryl sulfate and 47
chlorohexidine. Many bacteria in the highly complex oral flora can produce hydrogen 48
peroxide and antibacterial peptides, better known as bacteriocins, allowing the producers 49
to ensure their presence in the community by killing competing organisms (24). To 50
survive in the relatively hostile environment of oral biofilms, bacteria must be able to 51
sense, respond to, and cope with these insults. The cell envelope plays a vital role during 52
these processes as it protects the cell from the environment, maintains cell shape, acts as 53
a molecular sieve, and provides a platform for components of the cell involved sensing 54
and transmission of environmental signals. Ensuring envelope integrity is therefore 55
crucial for bacterial cells to survive. 56
Streptococcus mutans, a primary causative agent of human dental caries, lives almost 57
exclusively in biofilms on the tooth surface. This bacterium is known for its ability to 58
survive and adapt to environmental insults, including mounting protective responses in 59
reaction to various stimuli (10, 28). Multiple pathways are utilized by S. mutans to 60
modulate its capacity to cope with stresses, but certain two-component signal 61
transduction systems (TCS), including CiaHR, VicRK and LiaSR, play integral roles in 62
survival and adaptation to low pH, reactive oxygen species (ROS) and cell envelope 63
stress induced by antimicrobial agents (5, 6, 11, 38, 40). For example, mutants lacking 64
LiaSR in S. mutans displayed increased susceptibility to Lipid II cycle interfering 65
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

4
antibiotics and chemicals that perturb cell membrane integrity (40). In addition, BrpA, 66
for biofilm regulatory protein A, is also involved in acid- and oxidative stress tolerance 67
response and biofilm development by S. mutans (42, 43). Relative to the parent strain, S. 68
mutans strains lacking BrpA had a limited ability to grow and accumulate on a surface 69
and displayed enhanced sensitivity to low pH and hydrogen peroxide. 70
A predicted surface-associated protein, BrpA contains a region homologous to the 71
LytR-CpsA-Psr (LCP) domain of the LCP family of proteins. The LCP family of proteins 72
is widely distributed among Gram-positive bacteria and its members are generally 73
annotated as cell wall associated transcriptional regulators (17). Originally, the LytR 74
protein of Bacillus subtilis was identified as an autogenous transcriptional attenuator that 75
also regulated the promoter of the divergently transcribed lytABC operon, which encodes 76
a lipoprotein (LytA), an N-acetylmuramoyl-L-alanine amidase (autolysin, LytC), and a 77
modifier protein of LytC (LytB) (26). The LytR paralogue CpsA of Streptococcus 78
agalactiae was subsequently shown to function as a transcriptional activator of the 79
capsule operon (14, 16). Recently, LytR in Streptococcus pneumoniae was reported to be 80
essential for normal septum formation (20), with the mutant displaying variability in size 81
and shape. The lytR mutants were also found to form multiple asymmetrical septa. 82
Similar functions were also observed with MsrR, a Psr-like protein in Staphylococcus 83
aureus (36). A mutant lacking MsrR was reported to have a 4-fold decrease in minimal 84
inhibitory concentration (MIC) against oxacillin and a 2-fold reduction against 85
teicoplanin, when compared to the parental strain. 86
Previously, we showed that BrpA-deficiency in S. mutans causes major defects in 87
biofilm formation and acid- and oxidative stress responses (42, 43). Relative to the parent 88
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

5
strain, the deficient mutant also had an increased rate of autolysis and a decreased 89
viability, suggesting compromise in cell envelope biogenesis /homeostasis. In this study, 90
we used reporter gene fusion and antibacterial susceptibility assays to further characterize 91
S. mutans strains deficient of BrpA. Results showed that S. mutans strains lacking BrpA 92
were more susceptible to cell envelope-targeting antimicrobials and that cell envelope 93
and environmental stresses enhanced the expression of BrpA. In addition, we show that 94
BrpA is required for optimal binding to salivary agglutinin. These results extend previous 95
studies showing that BrpA plays a critical role in cell envelope biogenesis and cell 96
envelope stress responses in S. mutans. 97
98
Materials and Methods 99
Plasmids, bacterial strains, cell lines, and growth conditions. Bacterial strains and 100
plasmids used in this study are listed in Table 1. S. mutans strains were maintained on 101
brain heart infusion (BHI) medium. For biofilm formation, S. mutans was grown in 102
modified biofilm medium (BM) with glucose (18 mM) and sucrose (2 mM) as 103
supplemental carbon and energy sources (BMGS) (29, 42, 43). All solid media were 104
prepared similarly with inclusion of Bacto agar (Difco Laboratories, Franklin Lakes, NJ) 105
at the level of 1.5% (w/v). When needed, erythromycin (10 µg/ml), kanamycin (1 106
mg/ml), and/or spectinomycin (1 mg/ml) were added. Unless otherwise stated, cells were 107
grown at 37oC in an aerobic environment with 5% CO2. All E. coli strains were grown in 108
Luria Bertani medium at 37oC aerobically, with or without inclusion of kanamycin (40 109
µg/ml), ampicillin (100 µg/ml), spectinomycin (100 µg/ml), and/or erythromycin (300 110
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

6
µg/ml). Human coronary artery endothelial cells (HCAEC) were grown and maintained 111
in endothelial cell basal medium-2 (EBM-2, Lonza) (2, 31). 112
113
DNA manipulation, transcriptional initiation site mapping, and construction of 114
reporter fusions. Standard recombinant DNA procedures were used (12, 37). All 115
restriction and modifying enzymes were purchased from Invitrogen (Carlsbad, CA) or 116
New England Biolabs (Ipswich, MA) and used as recommended by the suppliers. All 117
primers (Table 1) were synthesized by Integrated DNA Technologies, Inc. (Iowa City, 118
IA). RNA Ligase Mediated Rapid Amplification of cDNA Ends (RLM-RACE) 119
(Ambion, Inc., Foster City, CA) was used to map the transcription initiation site (TIS) of 120
brpA. Briefly, total RNA was prepared from early- (OD600nm = 0.2) and late- (OD600nm = 121
0.8) exponential phase cultures grown in BHI using hot phenol (1, 42). The preparations 122
were then treated with RNase-free DNase I (Ambion, Inc.), and RNA was retrieved using 123
the Qiagen RNeasy purification kit (Qiagen, Inc., Valencia, CA). For cDNA synthesis, 124
total RNA was treated with calf intestinal phosphatase and with Tobacco Acid 125
Pyrophosphatase by following the supplier’s recommendations, and then ligated to the 126
supplied 5’ RACE adapter. cDNA was synthesized using iScript reverse transcriptase III 127
(Invitrogen) and followed by a nested PCR using either the 5’ RACE outer primer or the 128
5’ RACE inner primer (Ambion, Inc.) and a brpA-specific reverse primer. The TIS was 129
determined by sequencing of the resulting PCR amplicon. 130
To analyze the regulation of brpA expression, a promoterless luciferase gene (luc) 131
was used as a reporter (23, 35). Briefly, the cognate brpA promoter region was amplified 132
by PCR with primers pbrpA5’NheI and pbrpA3’ XhoI. Following proper restriction 133
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

7
digestions, the amplicon was cloned directly in front of the promoterless luc gene in 134
integration vector pFW11-luc (22), which also contains a Shine-Dalgarno sequence 135
optimized for group A streptococci (35). Following confirmation of the correct sequence 136
of the cloned element, the resulting construct, pFW11::PbrpA::luc, was introduced into S. 137
mutans UA159 and TW14D and maintained on BHI agar containing 1 mg/mL 138
spectinomycin. The expression of BrpA under different environmental conditions and 139
cell envelope stressors were analyzed using luciferase assay by following the protocol of 140
Podbielski (22, 35). 141
142
DNA microarray and RealTime-PCR analysis. For DNA microarray analysis, 143
total RNAs were extracted from early-exponential phase (OD600nm≅0.3) cultures, treated 144
with DNaseI (Ambion, Inc.) to remove all DNA, and then retrieved with the RNeasy 145
purification kit (QIAGEN, Inc.) (42). Array analysis was performed by using the whole-146
genome S. mutans microarrays (version 2) that were obtained from The J. Craig Venter 147
Institute (JCVI, http://pfgrc.jcvi.org) by following the protocols recommended by JCVI 148
as described elsewhere (1, 42). Expression levels of selected genes identified by DNA 149
microarray analysis were confirmed by Real-time PCR procedures detailed elsewhere 150
(Table 1) (5, 42). Microarray data have been deposited in NCBI (accession #GSE35349). 151
152
Cell envelope antimicrobial susceptibility assays. The susceptibility of S. mutans 153
strains to antimicrobial agents was analyzed using microtiter plate-based assays as 154
described previously (30, 40). Cell envelope antimicrobial agents tested included the 155
antibiotics vancomycin (Sigma, St. Louis, MO), bacitracin (Sigma), and β-lactam 156
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

8
antibiotic pencillin G (Sigma); the bacteriocin nisin (Sigma); and the cell envelope active 157
compounds sodium dodecyl sulfate (SDS) and chlorhexidine (Sigma). Briefly, 100 µl of 158
properly diluted mid-exponential phase cultures were added to 96-well plates containing 159
BHI medium supplemented with two-fold serial dilutions of cell envelope antimicrobial 160
agents. After 48 hours, bacterial growth was measured spectrophotometrically using a 161
Synergy 2 plate reader (BioTek, Inc.), and relative cell density percentages (OD490nm of 162
cultures with antimicrobial agents divided by the OD490nm of the untreated cultures X 163
100) were calculated. Minimal inhibitory concentration (MIC) was defined as the lowest 164
concentration at which the cultures did not grow to over 10% of the relative cell density. 165
Minimal bactericidal concentration (MBC) assays were carried out using the MIC test 166
plates. MBC was determined as the lowest concentration where fewer than 5 colony-167
forming-units (CFU) were observed after 48 hours when 20µl of the cultures was plated 168
on non-selective medium. 169
170
Biofilm formation and BIAcore assays. Biofilm formation on 96-well plates pre-171
coated with salivary agglutinin was carried out as previously described (4, 42, 43). 172
Interactions of S. mutans whole cells with salivary agglutinin were analyzed using 173
BIAcore assays in which the receptor was immobilized on Pioneer F1 sensor chips (32). 174
175
Preparation of protein fractions and Western blot analysis. Various fractions of 176
proteins were prepared from BHI-grown early-exponential phase (OD600nm = 0.3) cultures 177
of S. mutans (3, 41, 45). Briefly, whole cell lysates were obtained by glass bead-beating 178
in SDS-boiling buffer (60 mM Tris, pH 6.8, 10% glycerol, and 5% SDS). For surface-179
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

9
associated fractions, cells from 500 ml cultures were suspended in 25 ml of 0.2% N-180
dodecyl-N,N-dimethyl-3-ammonio-1-propanesulfonate (Zwittergent, Sigma) and 181
incubated at 28oC with shaking at 80 rpm for 1 hour. Following centrifugation, the 182
supernatants were further concentrated using Amicon Ultra centrifugal filters (Millipore, 183
Billerica, MA). In other cases, bacterial cells were suspended in 4% SDS and incubated 184
at room temperature for 30 minutes. For cell-free fractions, cultural supernatants were 185
precipitated by ammonium sulfate. For Western blot analysis, proteins (10 µg total) were 186
separated using 7.5% SDS-PAGE, blotted onto Immobilon-FL membranes, and then 187
probed with anti-P1 monoclonal antibodies (8, 41). 188
189
Bacterial invasion assay. The impact of BrpA-deficiency on S. mutans ability to 190
invade host tissues were analyzed using primary human coronary artery endothelial cells 191
(HCAEC) as described elsewhere (2, 31). Briefly, overnight cultures were harvested by 192
centrifugation at 14,000xg for 5 minutes, and pellets were washed twice with phosphate-193
buffered saline (pH 7.2) and resuspended in endothelial cell basal medium-2 (EBM-2, 194
Lonza). Aliquots (1 ml) of bacterial cells (with ~5x107 cfu/ml) were mixed with HCAEC 195
monolayers in 24-well plates for 2-hours. Following proper washes and additional 196
incubation with gentamicin and penicillin G to eliminate extracellular bacterial cells, 197
HECAE cells were lysed by osmotic shock, and serial dilutions of the lysates and 198
bacterial cells released were plated on BHI agar in triplicate. The percentage of 199
intracellular bacteria relative to the initial inoculum was calculated. 200
201
Wax worm infection model. Galleria mellonella killing assays were performed by 202
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

10
following the procedures described previously (21). Briefly, groups of 20 larvae, ranging 203
from 200 to 300 mg in weight and with no signs of melanization, were randomly 204
assigned. A 5 µl aliquot of properly diluted mid-exponential phase (OD600nm = 0.5) 205
cultures of S. mutans wild-type or the BrpA-deficient mutant were injected into the 206
hemocoel using a 10 µl Hamilton syringe (Hamilton Co., Reno, NV). Groups receiving 207
heat-inactivated (10 min at 75°C) S. mutans wild-type or saline were used as controls. 208
After injection, larvae were incubated at 37°C, and appearance (signs of melanization) 209
and survival were recorded at selected intervals. Kaplan-Meier killing curves were 210
plotted and estimates of differences in survival were compared using a log rank test. A P 211
value of ≤0.05 was considered significant. All data were analyzed with GraphPad Prism, 212
version 4.0. 213
214
Results 215
BrpA-deficiency affects binding to immobilized salivary agglutinin. Binding to 216
salivary agglutinin and other glycoproteins, primarily through the multi-functional 217
adhesin P1 (also called Antigen I/II, PAc, or SpaP), is considered to be a major 218
mechanism used by S. mutans to colonize the tooth surface (15, 19, 25, 27). On 96-well 219
plates pre-coated with whole saliva and purified salivary agglutinin (4), S. mutans wild-220
type UA159 formed robust biofilms after 24 hours, consistent with previous findings (4). 221
Relative to UA159, however, biofilm accumulation by the BrpA-deficient mutant, 222
TW14D was significantly lower (P<0.05) (Figure 1A). We also used BIAcore assays to 223
analyze the impact of BrpA-deficiency on P1-mediated adherence and biofilm formation. 224
Affinity-purified, high molecular weight salivary glycoprotein agglutinin was 225
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

11
immobilized on a Pioneer F1 sensor chip and interaction of S. mutans with immobilized 226
agglutinin was measured by BIAcore, a proven technique for assessment of salivary 227
agglutinin mediated adherence (32). When compared to the wild-type, the capacity of 228
salivary agglutinin-mediated whole cell-receptor interaction in the mutant deficient of 229
BrpA was decreased by more than 57%, with the average resonance signal for the wild-230
type, UA159 at 938.95(±102.45) versus 413.6(±186.7) (P<0.01) for the mutant, TW14D 231
(Figure 1B). 232
Western blot analysis was then carried out to further examine the levels of P1 in 233
whole cell lysates, cell-free fractions and surface-associated fractions from UA159 and 234
TW14D using monoclonal antibodies (mcAb) against P1 as probes (3, 8, 41, 45). When 235
probed with mcAb 6-8C, which reacts the C-terminus of P1 (8), a single band with 236
molecular weight (MW) around 200 kDa was apparent in the surface-associated fractions 237
of both UA159 and TW14D (Figure 2A). In comparison, however, the density of this 238
reactive band in TW14D was over 2-fold of the one in UA159. A similar band was also 239
detected in whole cell lysate of TW14D, but was not detectable in UA159. When probed 240
with mcAb 4-10A that recognizes the A-P stalk, one major band of MW around 200 kDa 241
was apparent in both TW14 and UA159 (Figure 2B), with the density of the band in 242
TW14D being more than 4-fold higher than that in UA159. Besides, multiple bands with 243
MW around 150 kDa were apparent in the whole cell lysates, but again these bands in 244
TW14D were more than 9-fold denser than those in UA159. With mcAb 3-8D as a 245
probe, which recognizes the A-region of the P1, multiple bands with similar MW around 246
100 kDa were identified in the whole cell lysates of both UA159 and TW14D (Figure 247
2C). In comparison, the densities of these bands in the mutant were about 6-fold higher 248
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

12
than those the wild-type. Multiple bands reactive to mcAb 3-8D were also seen in the 249
surface-associated fractions, but different from the whole cell lysates, these bands were 250
mostly around 75 kDa. 251
252
BrpA-deficient mutants are more sensitive to cell envelope-active antimicrobials. 253
Previously, it was shown that BrpA-deficiency in S. mutans caused elevations in autolysis 254
and reductions in viability, with more dead cells and cell debris in biofilms relative to the 255
wild-type strain (13, 42). To analyze whether BrpA in S. mutans affects cell envelope 256
integrity, MIC and MBC against several cell envelope antibacterial agents were measured 257
using microtiter plate-based assays. As shown in Table 2, the BrpA-deficient mutant had 258
a decreased ability to survive the treatment of several different antimicrobial agents, 259
when compared to the wild-type strain under the same conditions. TW14D had a 1.8- and 260
2-fold reduction in MIC to nisin and bacitracin, respectively. Similar trends were also 261
detected with vancomycin, penicillin-G, D-cycloserine, SDS and triclosan, although the 262
differences between these two strains were not statistically significant. When MBC’s 263
were analyzed, the deficient mutant had ≥1.5-fold reductions in sensitivity to nisin, 264
chlorhexidine and SDS. Slight, but not statistically significant decreases were also seen 265
with bacitracin, vancomycin and triclosan. 266
267
BrpA-deficiency affects virulence in a Galleria mellonella model. Recent studies 268
have shown that certain strains of S. mutans, such as OMZ175 (serotype f), are highly 269
invasive, and consequently may play a significant role in development of certain systemic 270
diseases, such as infective endocarditis (2, 31). In contrast, UA159, a commonly used 271
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

13
laboratory strain, possesses only limited capacity to invade endothelial cell lines (2). To 272
create a BrpA-deficient mutant of OMZ175, PCR reactions were conducted with genomic 273
DNA from TW14D as the template (Table 1) (42). The resulting amplicon containing 274
DNA fragments flanking brpA and an erythromycin resistance element (ermr) was used 275
to replace the brpA-coding sequence in S. mutans OMZ175, and mutants were selected 276
on BHI with erythromycin and further confirmed by DNA sequencing. When analyzed 277
by invasion assays using human coronary artery endothelial cells (HCAEC) (2), the 278
BrpA-deficient mutant, TW230 had a slightly reduced invasion efficiency, when 279
compared to OMZ175, with an average invasion rate of 0.18% for TW230 vs 0.49% for 280
OMZ175 (P=0.089). When tested in the G. mellonella (wax worm) virulence model (21), 281
the survival rate of worms receiving the BrpA-deficient mutants was significantly 282
(P<0.01) higher than those receiving strain OMZ175 (Figure 3). Not surprisingly, 283
considering the poor virulence of strain UA159 in this model, no major differences 284
(P>0.05) were observed when TW14D and UA159 were compared in the wax worms 285
(data not shown). 286
287
brpA is cotranscribed with SMU.409. The brpA gene (SMU.410) in S. mutans is 288
flanked by downstream SMU.411 and by upstream SMU.409 (Figure 4), which encodes a 289
streptococcus-specific, hypothetical protein and a putative bacterial ATPase/GTPase 290
(www.oralgen.lanl.gov), respectively. The open reading frames in SMU.409 and brpA 291
are arranged in the same orientation, while SMU.411 and brpA are transcribed in 292
opposite directions. To map the promoter region of brpA, TIS was examined using 293
5’RACE with total RNA extracted from BHI-grown cultures. Results of 5’ RACE PCR 294
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

14
showed that multiple brpA transcripts existed (data not shown). Sequence analysis of the 295
major complementary DNA product revealed that the major TIS of brpA was 774 bp 296
upstream of the translational start codon ATG (Figure 4), suggesting that SMU.409 and 297
brpA are co-transcribed under the conditions studied. Reverse transcription-PCR with 298
total RNA extracted from BHI-grown planktonic cultures and 3-day biofilms grown on 299
BMGS confirmed that brpA was co-transcribed with the upstream gene SMU.409 (Figure 300
5A) under both of the conditions tested. Insertion of polar kanamycin resistance element, 301
ΩKm (34) at SMU.409 also caused a reduction of more than 25-fold in brpA 302
transcription, as shown by RealTime-PCR with total RNA preps of early-exponential 303
(OD600nm = 0.25) cultures of the insertional mutant and its parent strain UA159 (data not 304
shown). Similar results were also obtained with Western blot analysis (data not shown). 305
306
Expression of BrpA is regulated in response to environmental conditions. In 307
cultures grown in BHI broth, luciferase expression from the full brpA promoter (a 1119 308
bp fragment) was measured at its maximum during early-exponential phase (OD600nm = 309
0.3; Figure 5B), consistent with our earlier study by Northern blotting (44). Considering 310
the fact that mutants lacking BrpA had significant defects in their abilities to survive low 311
pH and hydrogen peroxide challenge (42), cells of early-exponential phase cultures 312
carrying the reporter fusion were treated with hydrogen peroxide and methyl viologen 313
(paraquat, Sigma) in the growth medium for 90 min. Results showed that, relative to the 314
un-treated controls, the level of luciferase activity in cells treated with hydrogen peroxide 315
and methyl viologen was increased significantly (Table 3). Such increases appeared to be 316
concentration dependent when the amount used was within a certain threshold (Figure 317
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

15
S1). However, beyond the threshold, luciferase activity was decreased with further 318
increases of hydrogen peroxide and methyl viologen used. Similar results were also 319
obtained with cells treated with various cell envelope antimicrobials at sub-inhibitory 320
concentrations (Tables 2&3), with the most significant impact measured with 321
chlorhexidine, a chemical commonly used in dental care products and in prevention of 322
tooth decay. Efforts were also made to evaluate the impact of pH on BrpA expression by 323
incubating the cells carrying the reporter fusion in BHI broth adjusted to different pH 324
values, but the results were inconclusive, probably due to the impact of low pH on the 325
luciferase enzyme (data not shown). To circumvent this problem, study is underway 326
using chloramphenicol acetyltransferase as a reporter under controlled conditions in a 327
chemostat. 328
329
BrpA-deficiency causes substantial alterations in the transcriptional profiles of 330
the deficient mutant. In consideration of the fact that BrpA expression is at its 331
maximum during early-exponential phase as shown by Northern blotting (44) and 332
reporter gene fusion assays, we carried out another DNA microarray analysis using total 333
RNA from early exponential phase cultures (OD600nm =0.3). It was found that 92 genes 334
were up-regulated and 90 down-regulated in TW14D by a factor of at least 1.5-fold (p ≤ 335
0.001, Tables S1&S2). At a level of P<0.01, 176 additional genes were found to be 336
differentially expressed in TW14D, with 77 up- and 90 down-regulated (data not shown). 337
Based on the description and putative functions of the genes identified at the significance 338
level of P<0.001, BrpA-deficiency affects almost every aspect of the cellular physiology 339
as well as virulence properties, including amino acid biosynthesis (10), carbohydrates and 340
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

16
energy metabolism (18), nucleic acids and DNA metabolism (10), transcriptional 341
regulation (9), ABC transporters (29), molecular chaperones and other cellular processes 342
(13), and hypothetical and conserved hypothetical proteins (50). The breadth of impact 343
of BrpA-deficiency on the transcriptional profile of the deficient mutant is similar to what 344
was observed previously with mid-exponential phase cells (42). However, comparison of 345
the two transcriptional profiles revealed that only a small number of genes were 346
consistently up- or down-regulated in both early- and mid-exponential phase cultures, 347
which include recA (for recombinant protein RecA), gtfD (for glucosyltransferase D), 348
wapA (for surface-associated protein WapA), groEL (for molecular chaperone GroEL) 349
and sod (for Mn-dependent superoxide dismutase SOD) (Tables S1&S2) (42). In 350
addition, the magnitude of alterations in gene expression was also more dramatic in cells 351
of the early-exponential phase, when compared to that in the mid-exponential phase 352
cultures. 353
354
Discussions 355
The cell envelope is of essential importance for growth, cell division, interaction with 356
the environment, and antimicrobial resistance. Previous studies have shown that BrpA, a 357
paralogue of the LCP family of cell wall-associated transcriptional attenuators, strongly 358
influences S. mutans biofilm formation and survival against low pH and reactive oxygen 359
species (42, 43). Strains deficient of BrpA also displayed increased autolysis rates and 360
decreased viability, suggesting a role for BrpA in regulation of cell envelope biogenesis 361
or homeostasis (13, 42, 43). In this study, BrpA-deficiency was shown to significantly 362
weaken the ability of S. mutans to survive cell envelope stresses induced by cell 363
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

17
envelope-targeting antimicrobials (Table 2). Among the antimicrobial agents tested, the 364
most significant influences in MIC were measured with nisin and bacitracin, two 365
antibiotics that interfere with Lipid II cycling, blocking peptidoglycan and cell wall 366
biosynthesis (9). The most significant impact in MBC was seen with chlorhexidine and 367
SDS, two chemicals commonly used in oral health care products that compromise 368
membrane integrity. These results provide further support for a role for BrpA in 369
regulation of cell envelope biogenesis or maintenance by S. mutans, consistent with the 370
roles of certain LCP paralogues in other bacterial species (18, 20, 33, 36, 39). 371
The bacterial cell wall is a repeating, three-dimensional polymer, known as 372
peptidoglycan or murein consisting of a linear, alternating N-acetylmuramic acid 373
(MurNAc) and N-acetylglucosamine (GlcNAc) motif, cross-linked via peptides appended 374
to MurNAc. Of the genes altered as a result of BrpA-deficiency in TW14D, several were 375
found to encode proteins with potential roles in peptidoglycan biosynthesis (Table 4). 376
Among them are SMU.246 for a phospho-MurNAc-pentapeptide-transferase (RgpG), 377
SMU.549 for an undecaprenyl-PP-MurNAc-pentapeptide-UDP-GlcNAc transferase 378
(MurG), SMU.599 for a D-alanine-D-alanine ligase (DdlA), and SMU.1677 for a UDP-379
MurNAc-tripeptide synthetase (MurE). While the exact role of these gene products in S. 380
mutans cellular physiology awaits further investigation, down-regulation of genes 381
involved in peptidoglycan synthesis would have an impact on cell envelope biogenesis, 382
likely leading to defects in wall integrity. Such a defect would be consistent with the 383
weakened resistance to cell envelope antimicrobials, reduced viability and increased 384
autolysis observed for BrpA-deficient mutants (13, 43). In support of a role in cell 385
envelope biogenesis, the expression of a luciferase reporter fusion under the direction of 386
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

18
brpA promoter was also up-regulated in response to cell envelope stresses induced by 387
exposure to sub-inhibitory concentrations of antimicrobial agents that target the cell 388
envelope (Table 3). Defects in cell envelope integrity would likely result in vulnerability 389
of the bacterial cells to environmental insults, and therefore can partly explain the 390
weakened acid- and oxidative stress response of the BrpA-deficient mutants (42). 391
P1, a cell wall anchored adhesin, is considered a key contributor to S. mutans 392
colonization of the tooth surface (7, 15). P1 mediates the adherence through interactions 393
with high molecular weight salivary agglutinin in the enamel pellicle. Both biofilm 394
formation assays and BIAcore analysis showed that BrpA affects the ability of S. mutans 395
to interact with salivary agglutinin (Figure 1). As shown by Western blotting, however, 396
the level of P1 expression was increased by more than 2-fold as a result of BrpA-397
deficiency (Figure 2A). When analyzed by DNA microarrays, several genes encoding 398
components of the Sec translocase were also found altered in TW14D. These included 399
secA, secE and secY encoding the ATP-dependent motor of the translocation machinery, 400
SecA, and the translocon pore components, SecE and SecY, respectively. Both secA and 401
secE were down-regulated by more than 2-fold, while secY was up-regulated by more 402
than 2-fold (Table 4). The Sec secretion system participates in translocation of 403
polypeptides across, or integration into, the cytoplasmic membrane (46). Alteration in 404
expression of individual members of the Sec translocon complex, as well as global 405
defects in cell envelope integrity, will likely influence the function of the translocation 406
/secretion machinery. As a result of altered Sec function, the P1 adhesin may be 407
compromised in conformation, stability and /or distribution on the surface. Therefore, 408
the increased expression could be a compensatory response to such a compromise, while 409
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

19
the underlying mechanism awaits further investigation. In addition, the disproportional 410
increases in density of the lower MW bands in TW14D that were reactive to mcAb 3-8D 411
and 4-10A, which are shown to recognize truncated peptides (8), also suggest the stability 412
of P1 may be reduced in the brpA mutant (Figure 2B&C). Therefore, decreased stability 413
and /or mis-folding of P1 may underlie the reduced binding to salivary agglutinin by 414
strain, TW14D. These in vitro results also suggest that BrpA-deficiency may affect 415
bacterial adherence and biofilm initiation by S. mutans in the oral cavity as well. 416
Previously, Northern blotting showed that transcription of brpA was maximal during 417
early-exponential phase (OD600nm ≈0.3) and that deficiency of LuxS dramatically 418
decreased brpA transcription, indicating that expression of brpA is regulated in response 419
to environmental conditions and by LuxS-mediated quorum sensing (44). In this study, 420
we used luciferase reporter gene fusion assays to show that the expression of BrpA is 421
strongly dependent on growth phase, with maximal activity measured during early-422
exponential phase (Figure 5B). These results again suggest that environmental conditions 423
and cell density play an important role in regulation of BrpA expression. Differences in 424
environmental conditions, such as pH and concentration of ROS, and cell density could 425
in part account for some of the discrepancies observed between the two transcriptional 426
profiles between the early- and mid-exponential phase cultures (Tables S1&S2) (42). 427
However, it awaits further investigation whether BrpA affects different group of genes in 428
response to environmental stimuli. 429
Both 5’RACE and RT-PCR showed that under the conditions studied the major 430
transcript was a product of co-transcription of brpA with its upstream locus SMU.409 431
(Figures 4&5). Consistently, both RealTime-PCR and Western blot analysis (data not 432
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

20
shown) showed that polar insertion at SMU.409 resulted in a dramatic reduction of BrpA 433
expression. However, we have previously showed that possession in trans of the brpA-434
coding sequence plus a 344 bp fragment upstream of its start codon was able to partially 435
complement the deficient mutant TW14 in acid tolerance response (42). Luciferase 436
reporter fusion with a fragment of 683 bp upstream of brpA also showed promoter 437
activity in this intergenic region, although it is much weaker than the full-length (1119 438
bp) promoter (data not shown). Computer-based analysis of this intergenic region using 439
BPROM, a bacterial sigma70 promoter recognition program, and Virtual Footprint, a 440
program especially designed to analyze transcription factor binding sites, also revealed 441
putative -10 (TATAAc) and -35 (TTGAgA) sites and regions with high similarity to 442
binding sites for several putative transcriptional regulators (Figure 4). These results 443
further suggest that transcription of brpA may be initiated at different sites under different 444
environmental conditions. Study is currently under way to dissect the underlying 445
mechanisms, including the cis-and trans-acting elements involved in regulation of brpA 446
expression. SMU.409 encodes a putative ATPase/GTPase. While the exact role of 447
SMU.409 in regulation of S. mutans cellular physiology and brpA expression is still 448
under investigation, the close association of this gene with brpA also suggests its likely 449
involvement in BrpA-regulated cell envelope biogenesis /homeostasis. 450
In summary, results presented here further support that S. mutans BrpA is involved in 451
the regulation of cell envelope biogenesis /maintenance and that deficiency of BrpA 452
affects the fitness of the deficient mutants and decreases the virulence of OMZ175, a 453
highly invasive strain in a wax worm model. Current effort is directed to further 454
investigation of the underlying mechanisms. 455
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

21
456
Acknowledgement 457
This work is supported by NIDCR grant DE19452 to Z.T.W. and in part by the South 458
Louisiana Institute of Infectious Disease Research. We would like to thank Dr. Fengxia 459
(Felicia) Qi at the University of Oklahoma Health Sciences Center for her kind gift of the 460
integration vector pFW11-luc, and Mr. James H. Miller for his assistance with invasion 461
and wax worm infection assays. 462
463
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

22
Figure legends 464
Figure 1. Biofilm formation (A) and BIAcore assays (B). A: S. mutans UA159 and 465
TW14D were grown on 96-well plates that were pre-coated with unstimulated whole 466
saliva (WS) or affinity-purified salivary agglutinin (AG). Data shows the average density 467
(±standard deviation in error bars) of 24-hour biofilms from more than three independent 468
sets of experiments, with a * indicating significant difference between UA159 and 469
TW14D under the conditions specified (P<0.05). B: P1-mediated S. mutans whole cell 470
interaction with salivary agglutinin was measured using BIAcore assays. Salivary 471
agglutinin was immobilized on an F1 chip surface. S. mutans UA159 and the BrpA-472
deficient mutant, TW14D were injected for 60 seconds. S. mutans UA159 yielded an 473
average resonance signal of 938.95, while TD14D had 413.6. Results indicate that BrpA-474
deficiency affects P1-mediated whole cell adhesin-receptor interactions. Panel shows 475
representatives of two independent experiments. 476
477
Figure 2. Western blot (A-C) and SDS-PAGE (D) analysis of P1 in S. mutans wild-type 478
UA159 (lanes 1&3) and the BrpA-deficient mutant TW14D (lanes 2&4). Proteins (10 µg 479
total) of whole cell lysates (lanes 1&2) and surface-associated fractions (lanes 3&4) were 480
separated using 7.5% SDS-PAGE (D), blotted onto a PVDF nitrocellulose membrane, 481
and then probed with anti-P1 monoclonal antibodies (mcAb). Panel A shows results 482
when probed with mcAb 6-8C, which recognizes the C-terminus of the P1. A single 483
band with molecular weight (MW) around 200 kDa was apparent and the density of this 484
band in TW14D was more than 2-fold of the one in UA159. Panels B and C show results 485
when probed with mcAb 4-10A and mcAB 3-8D, which recognizes the A-P stalk and the 486
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

23
alanine-rich region of P1, respectively, but both are shown to react to truncated peptides 487
(8). Multiple bands with MW around 150 (B) and 100 (C) kDa were identified in both 488
UA159 and TW14D, but the densities of these bands in TW14D were significantly higher 489
than those in UA159. M, molecular weight markers. 490
491
Figure 3. Killing of G. mellonella larvae infected with S. mutans wild-type and the 492
corresponding BrpA-deficient mutants at 37°C. Figure shows survival (Kaplan-Meier) 493
plots of S. mutans OMZ175 (solid squares) and the BrpA-deficient mutant, TW230 (open 494
cycles) injected at 1x107 CFU/larva. There was no killing of larvae injected with saline 495
and minimum killing of larva injected with heat-killed S. mutans UA159 cells (data not 496
shown). The experiments were repeated three times, and the data presented here are 497
results representative of a typical experiment. Compared to the wild-type parent strain, 498
OMZ175, the BrpA-deficient mutant, TW230 showed attenuated virulence (P ≤0.01). 499
No significant differences were measured between UA159 and TW14D (data not shown). 500
501
Figure 4. Schematic diagram of the brpA-flanking region (A) and analysis of the brpA 502
promoter region (B). Panel A shows the genetic organization of the regions flanking 503
brpA, with locus name and size labeled under and above the arrows, respectively. 504
Approximate positions of the primers used for RT-PCR were shown with a solid arrow 505
above SMU.409 and under brpA, respectively. Panel B highlights the promoter region of 506
brpA, which includes the coding sequence of SMU.409, with the translation initiation site 507
indicated by an arrow above the start codon of brpA and SMU.409, respectively. As 508
determined by 5’-RACE, the transcription initiation site (TIS, underlined) of brpA was 509
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

24
774 bp upstream of its translation start site, which overlaps the translation start site of 510
SMU.409 as annotated by NCBI (www.oralgen.lanl.gov). Computer-based analysis 511
using BPROM, a bacterial sigma70 promoter recognition program, revealed two sets of 512
putative -35 and -10 sites (boxed) and binding sites for putative transcriptional regulators 513
(underlined). Further analysis using Virtual Footprint, a program especially designed to 514
analyze transcription factor binding sites, also identified sequences with high similarity to 515
binding sites for transcriptional factors GltC, TnrA, and Fnr (underlined). 516
517
Figure 5. RT-PCR analysis (A) and Luciferase activity assays (B). A. RT-PCR analysis 518
was carried out using total RNA isolated from mid-exponential phase (OD600nm=0.5) 519
cultures. Lanes M, 1, 2, and 3 represent DNA markers, RT-PCR products, control with 520
no reverse transcriptase, and PCR products with genomic DNA as the template, 521
respectively. Similar results were obtained with total RNA extracted from 3-day biofilms 522
(data not shown). B. For reporter gene fusion study, the full-length brpA promoter region 523
was cloned in front of a promoterless firefly luciferase gene, and the resulting promoter-524
reporter fusion was integrated into the S. mutans genome. Cells carrying the reporter 525
fusion were collected at different times during growth and assayed for luciferase activity 526
by mixing 200 µL of whole cells with 50 µL of D-luciferin, 1 mM, pH 6.0. Data shows 527
reporter luciferase activity (open squares) in relative light units (RLU) calibrated with the 528
optical density (OD600nm) of the cultures (solid circles) at each time point. 529
530
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

25
References 531
1. Abranches, J., M. Candella, T. Z. Wen, H. V. Baker, and R. A. Burne. 2006. 532 Different roles of EIIABMan and EIIGlc in the regulation of energy metabolism, 533 biofilm development and competence in Streptococcus mutans. J. Bacteriol. 534 188:3748-3756. 535
2. Abranches, J., L. Zeng, M. Belanger, P. H. Rodrigues, P. J. Simpson-536 Haidaris, D. Akin, W. A. Dunn, Jr., A. Progulske-Fox, and R. A. Burne. 537 2009. Invasion of human coronary artery endothelial cells by Streptococcus 538 mutans OMZ175. Oral Microbiol. Immunol. 24:141-5. 539
3. Ahn, S. J., and R. A. Burne. 2006. The atlA operon of Streptococcus mutans: 540 role in autolysin maturation and cell surface biogenesis. J. Bacteriol. 188:6877-541 88. 542
4. Ahn, S. J., Z. T. Wen, L. J. Brady, and R. A. Burne. 2008. Characteristics of 543 biofilm formation by Streptococcus mutans in the presence of saliva. Infect. 544 Immun. 76:4259-68. 545
5. Ahn, S. J., Z. T. Wen, and R. A. Burne. 2006. Multilevel control of competence 546 development and stress tolerance in Streptococcus mutans UA159. Infect. Immun. 547 74:1631-42. 548
6. Biswas, I., L. Drake, D. Erkina, and S. Biswas. 2008. Involvement of sensor 549 kinases in the stress tolerance response of Streptococcus mutans. J. Bacteriol. 550 190:68-77. 551
7. Brady, L. J., S. E. Maddocks, M. R. Larson, N. Forsgren, K. Persson, C. C. 552 Deivanayagam, and H. F. Jenkinson. 2011. The changing faces of 553 Streptococcus antigen I/II polypeptide family adhesins. Mol. Microbiol. 77:276-554 86. 555
8. Brady, L. J., D. A. Piacentini, P. J. Crowley, and A. S. Bleiweis. 1991. 556 Identification of monoclonal antibody-binding domains within antigen P1 of 557 Streptococcus mutans and cross-reactivity with related surface antigens of oral 558 streptococci. Infect. Immun. 59:4425-35. 559
9. Breukink, E., and B. de Kruijff. 2006. Lipid II as a target for antibiotics. Nat. 560 Rev. Drug Discov. 5:321-32. 561
10. Burne, R. A. 1998. Oral streptococci...products of their environment. J. Dent. 562 Res. 77:445-452. 563
11. Burne, R. A., J. Abranches, S. J. Ahn, J. A. Lemos, Z. T. Wen, and L. Zeng. 564 2011. Functional Genomics of Streptococcus mutans., p. 185-204. In P. E. 565 Kolenbrander (ed.), Oral Microbial Communities: Genomic Inquires and 566 Interspecies Communication. ASM Press. 567
12. Burne, R. A., Z. T. Wen, Y. M. Chen, and J. E. C. Penders. 1999. Regulation 568 of expression of the fructan hydrolase gene of Streptococcus mutans GS-5 by 569 induction and carbon catabolite repression. J. Bacteriol. 181:2863-2871. 570
13. Chatfield, C. H., H. Koo, and R. G. Quivey, Jr. 2005. The putative autolysin 571 regulator LytR in Streptococcus mutans plays a role in cell division and is 572 growth-phase regulated. Microbiol. 151:625-31. 573
14. Cieslewicz, M. J., D. L. Kasper, Y. Wang, and M. R. Wessels. 2001. 574 Functional analysis in type Ia group B Streptococcus of a cluster of genes 575
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

26
involved in extracellular polysaccharide production by diverse species of 576 streptococci. J. Biol. Chem. 276:139-46. 577
15. Crowley, P. J., L. J. Brady, S. M. Michalek, and A. S. Bleiweis. 1999. 578 Virulence of a spaP mutant of Streptococcus mutans in a gnotobiotic rat model. 579 Infect. Immun. 67:1201-6. 580
16. Hanson, B. R., B. A. Lowe, and M. N. Neely. Membrane topology and DNA-581 binding ability of the Streptococcal CpsA protein. J. Bacteriol. 193:411-20. 582
17. Hubscher, J., L. Luthy, B. Berger-Bachi, and P. Stutzmann Meier. 2008. 583 Phylogenetic distribution and membrane topology of the LytR-CpsA-Psr protein 584 family. BMC Genomics 9:617. 585
18. Hubscher, J., N. McCallum, C. D. Sifri, P. A. Majcherczyk, J. M. Entenza, R. 586 Heusser, B. Berger-Bachi, and P. Stutzmann Meier. 2009. MsrR contributes to 587 cell surface characteristics and virulence in Staphylococcus aureus. FEMS 588 Microbiol. Lett. 295:251-60. 589
19. Jenkinson, H. F., and R. J. Lamont. 2005. Oral microbial communities in 590 sickness and in health. Trends Microbiol. 13:589-95. 591
20. Johnsborg, O., and L. S. Havarstein. 2009. Pneumococcal LytR, a protein from 592 the LytR-CpsA-Psr family, is essential for normal septum formation in 593 Streptococcus pneumoniae. J Bacteriol 191:5859-64. 594
21. Kajfasz, J. K., I. Rivera-Ramos, J. Abranches, A. R. Martinez, P. L. Rosalen, 595 A. M. Derr, R. G. Quivey, and J. A. Lemos. 2010. Two Spx proteins modulate 596 stress tolerance, survival, and virulence in Streptococcus mutans. J. Bacteriol. 597 192:2546-56. 598
22. Kreth, J., J. Merritt, W. Shi, and F. Qi. 2005. Co-ordinated bacteriocin 599 production and competence development: a possible mechanism for taking up 600 DNA from neighbouring species. Mol. Microbiol. 57:392-404. 601
23. Kreth, J., J. Merritt, W. Shi, and F. Qi. 2005. Competition and coexistence 602 between Streptococcus mutans and Streptococcus sanguinis in the dental biofilm. 603 J. Bacteriol. 187:7193-203. 604
24. Kuramitsu, H. K., X. He, R. Lux, M. H. Anderson, and W. Shi. 2007. 605 Interspecies interactions within oral microbial communities. Microbiol. Mol. Biol. 606 Rev. 71:653-70. 607
25. Lamont, R. J., D. R. Demuth, C. A. Davis, D. Malamud, and B. Rosan. 1991. 608 Salivary-agglutinin-mediated adherence of Streptococcus mutans to early plaque 609 bacteria. Infect. Immun. 59:3446-50. 610
26. Lazarevic, V., P. Margot, B. Soldo, and D. Karamata. 1992. Sequencing and 611 analysis of the Bacillus subtilis lytRABC divergon: a regulatory unit 612 encompassing the structural genes of the N-acetylmuramoyl-L-alanine amidase 613 and its modifier. J. Gen. Microbiol. 138:1949-61. 614
27. Lee, S. F., A. Progulske-Fox, and A. S. Bleiweis. 1988. Molecular cloning and 615 expression of a Streptococcus mutans major surface protein antigen, P1 (I/II), in 616 Escherichia coli. Infect. Immun. 56:2114-9. 617
28. Lemos, J. A., and R. A. Burne. 2008. A model of efficiency: stress tolerance by 618 Streptococcus mutans. Microbiol. 154:3247-55. 619
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

27
29. Loo, C. Y., D. A. Corliss, and N. Ganeshkumar. 2000. Streptococcus gordonii 620 biofilm formation: identification of genes that code for biofilm phenotypes. J. 621 Bacteriol. 182:1374-82. 622
30. McBain, A. J., R. G. Ledder, P. Sreenivasan, and P. Gilbert. 2004. Selection 623 for high-level resistance by chronic triclosan exposure is not universal. J. 624 Antimicrob. Chemother. 53:772-7. 625
31. Nakano, K., K. Fujita, K. Nishimura, R. Nomura, and T. Ooshima. 2005. 626 Contribution of biofilm regulatory protein A of Streptococcus mutans, to systemic 627 virulence. Microbes Infect. 7:1246-55. 628
32. Oli, M. W., W. P. McArthur, and L. J. Brady. 2006. A whole cell BIAcore 629 assay to evaluate P1-mediated adherence of Streptococcus mutans to human 630 salivary agglutinin and inhibition by specific antibodies. J. Microbiol. Methods 631 65:503-11. 632
33. Over, B., R. Heusser, N. McCallum, B. Schulthess, P. Kupferschmied, J. M. 633 Gaiani, C. D. Sifri, B. Berger-Bachi, and P. Stutzmann Meier. 2011. LytR-634 CpsA-Psr proteins in Staphylococcus aureus display partial functional redundancy 635 and the deletion of all three severely impairs septum placement and cell 636 separation. FEMS Microbiol. Lett. 320:142-51. 637
34. Perez-Casal, J., M. G. Caparon, and J. R. Scott. 1991. Mry, a trans-acting 638 positive regulator of the M protein gene of Streptococcus pyogenes with similarity 639 to the receptor proteins of two-component regulatory systems. J. Bacteriol. 640 173:2617-24. 641
35. Podbielski, A., M. Woischnik, B. A. Leonard, and K. H. Schmidt. 1999. 642 Characterization of nra, a global negative regulator gene in group A streptococci. 643 Mol. Microbiol. 31:1051-64. 644
36. Rossi, J., M. Bischoff, A. Wada, and B. Berger-Bachi. 2003. MsrR, a putative 645 cell envelope-associated element involved in Staphylococcus aureus sarA 646 attenuation. Antimicrob. Agents Chemother. 47:2558-64. 647
37. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a 648 laboratory manual, 2nd ed, vol., Cold Spring Harbor laboratory. Cold Spring 649 Harbor N.Y. 650
38. Senadheera, D., K. Krastel, R. Mair, A. Persadmehr, J. Abranches, R. A. 651 Burne, and D. G. Cvitkovitch. 2009. Inactivation of VicK affects acid 652 production and acid survival of Streptococcus mutans. J. Bacteriol. 191:6415-24. 653
39. Steidl, R., S. Pearson, R. E. Stephenson, N. Ledala, S. Sitthisak, B. J. 654 Wilkinson, and R. K. Jayaswal. 2008. Staphylococcus aureus cell wall stress 655 stimulon gene-lacZ fusion strains: potential for use in screening for cell wall-656 active antimicrobials. Antimicrob. Agents Chemother. 52:2923-5. 657
40. Suntharalingam, P., M. D. Senadheera, R. W. Mair, C. M. Levesque, and D. 658 G. Cvitkovitch. 2009. The LiaFSR system regulates the cell envelope stress 659 response in Streptococcus mutans. J. Bacteriol. 191:2973-84. 660
41. Wen, T. Z., P. Suntharaligham, D. G. Cvitkovitch, and R. A. Burne. 2005. 661 Trigger factor in Streptococcus mutans is involved in stress tolerance, competence 662 development, and biofilm formation. Infect. Immun. 73:219-25. 663
42. Wen, Z. T., H. V. Baker, and R. A. Burne. 2006. Influence of BrpA on critical 664 virulence attributes of Streptococcus mutans. J. Bacteriol. 188:2983-92. 665
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

28
43. Wen, Z. T., and R. A. Burne. 2002. Functional genomics approach to identifying 666 genes required for biofilm development by Streptococcus mutans. Appl. Environ. 667 Microbiol. 68:1196-203. 668
44. Wen, Z. T., and R. A. Burne. 2004. LuxS-mediated signaling in Streptococcus 669 mutans is involved in regulation of acid and oxidative stress tolerance and biofilm 670 formation. J. Bacteriol. 186:2682-91. 671
45. Wilkins, J. C., D. Beighton, and K. A. Homer. 2003. Effect of acidic pH on 672 expression of surface-associated proteins of Streptococcus oralis. Appl. Environ. 673 Microbiol. 69:5290-6. 674
46. Zimmer, J., Y. Nam, and T. A. Rapoport. 2008. Structure of a complex of the 675 ATPase SecA and the protein-translocation channel. Nature 455:936-43. 676
677 678
679 680
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

Table 1. Bacterial strains, plasmids and primers used in the study
Strains /plasmids Major properties References S. mutans UA159 Wild-type, serotype c
S. mutans OMZ175 Wild-type, serotype f, (2)
S. mutans TW14 UA159 derivative, brpA, erythromycin resistant (43)
S. mutans TW14D UA159 derivative, brpA, erythromycin resistant (42)
S. mutans TW14K UA159 derivative, brpA, kanamycin resistant This study
S. mutans TW230 OMZ175 derivative, brpA, erythromycin resistant This study
E. coli DH10B Cloning host, mcrA, mcrBC, mrr, and hsd Invitrogen, Inc.
pFW5-luc
Integration vector containing promoterless luciferase gene and a spectinomycin resistance marker
(22)
Primers for RACE
Name DNA sequence (5’ to 3’) Applications
5` RACE Adapter gcugauggcgaugaaugaacacugcguuugcuggcuuugaugaaa 5’ RACE
5` RACE Outer gctgatggcgatgaatgaacactg 5’ RACE
5` RACE Inner cgcggatccgaacactgcgtttgctggctttgatg 5’ RACE
RACE brpA Rev gtcaactcccattaagaggatac 5’ RACE
brpA check Fw gtttaccttaggaggaaactga 5’ RACE
brpA SP1 acctttggcaattccttttgtca sequencing
brpA SP2 aactgacttgacagataaaaact sequencing Primers for reporter fusion, RT-PCR and qPCR
Name Forward (5’ to 3’) Reverse (5’ to 3’) Applications
PbrpA ttatgatgctagcaagtctcaaagaca tcagtttcctcctcgagtaaacatc promoter-reporter fusion
SMU.409-brpA aaggctgccactttatcatttggatg aatcttaatatcaagcatatcctgaa RT-PCR
brpA:erm agctcagataaggctgagctccta aaaccgtctttcatgcccatgtgcat ∆brpA:erm amplification
SMU.409P5 tacagctaactcttctgcaacaccatc attcgatagggatccaaatgataaagtg 5’ fragment for polar insertion
SMU.409P3 cactttatcatttggatccctatcgaat acgatacttgctgacactgtctaaagct 3’ fragment for polar insertion
SMU.246 tccttcttatgattggtgtt ctactacttcttgacggtaat SMU.246 fragment, 135 bp
SMU.549 gcagtctcttacgattatgg gctacaacaggaggaact SMU.549 fragment, 84 bp
SMU.599 gtgcgactactattcctcaa tcttcaacttctgccaact SMU.599 fragment, 82 bp
SMU.1677 ctcattatggaagtctcaa aagtaggatgttcaatcg SMU.1677 fragment, 121 bp
SecA gtgcttccattacctatca attcctcttcttctgtcttc secA fragment, 87 bp
SecY caggaagtgtggttgtaa gcttgaacggatattgac secY fragment, 155 bp
BrpA cgtgaggtcatcagcaaggtc cgctgtaccccaaaagtttagg brpA fragment, 148 bp Note: Nucleotides underlined are restriction site engineered for cloning.
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from
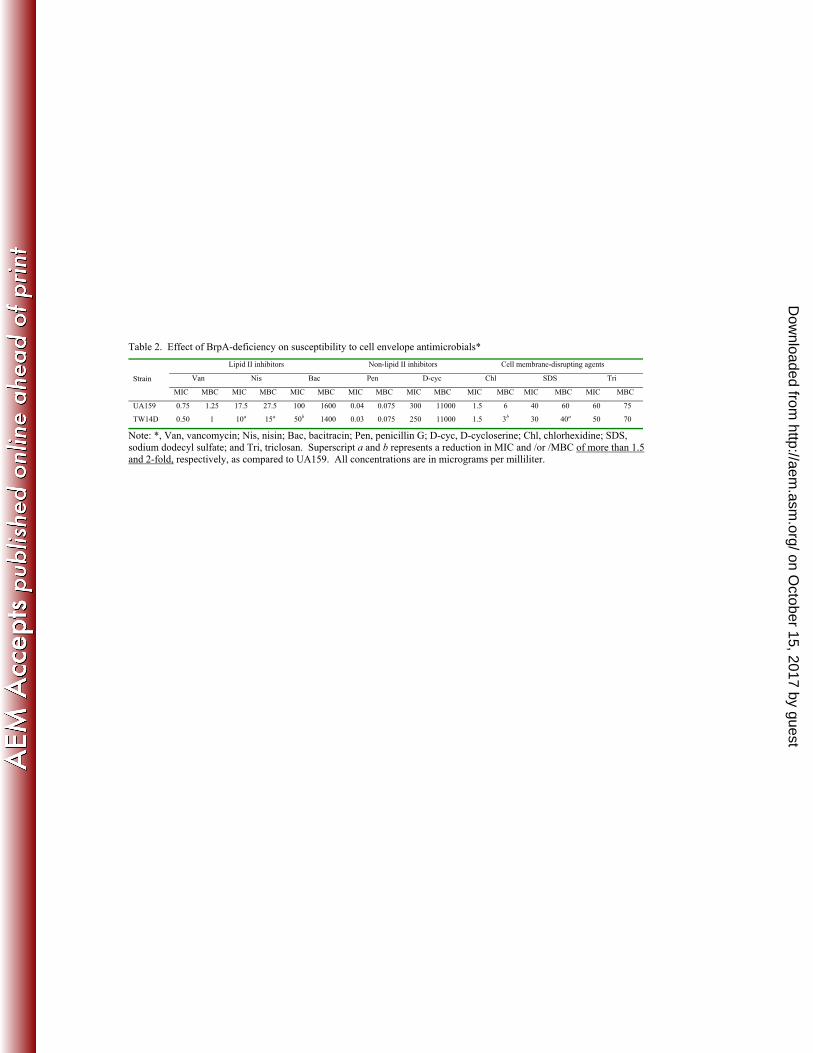
Table 2. Effect of BrpA-deficiency on susceptibility to cell envelope antimicrobials*
Strain
Lipid II inhibitors Non-lipid II inhibitors Cell membrane-disrupting agents
Van Nis Bac Pen D-cyc Chl SDS Tri
MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC MIC MBC
UA159 0.75 1.25 17.5 27.5 100 1600 0.04 0.075 300 11000 1.5 6 40 60 60 75
TW14D 0.50 1 10a 15a 50b 1400 0.03 0.075 250 11000 1.5 3b 30 40a 50 70
Note: *, Van, vancomycin; Nis, nisin; Bac, bacitracin; Pen, penicillin G; D-cyc, D-cycloserine; Chl, chlorhexidine; SDS, sodium dodecyl sulfate; and Tri, triclosan. Superscript a and b represents a reduction in MIC and /or /MBC of more than 1.5 and 2-fold, respectively, as compared to UA159. All concentrations are in micrograms per milliliter.
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

Table 3. Luciferase expression in response to oxidative stress and cell envelope active antimicrobial agents*
Stimuli and stressor (concentration) RLU
hydrogen peroxide (0.4 mM) 2.09 ± 0.33c
methyl viologen (10 mM) 2.07 ± 0.55b
SDS (8 µg/mL) 2.51 ± 0.50c
chlorhexidine (1.5 µg/mL) 3.03 ± 0.42c
vancomycin (0.75 µg/mL) 1.41 ± 0.03a
bacitracin (20 µg/mL) 2.24 ± 0.13a
nisin (10 µg/mL) 2.25 ± 0.47b
D-cycloserine (20 µg/mL) 1.62 ± 0.31b
penicillin G (0.04 µg/mL) 1.66 ± 0.35a
Note: *, Data were expressed as the ratio (average ±standard deviation) of the luciferase activity in RLU at the conditions specified over controls that received equal amount of solvent instead of the stressor indicated, with a superscript of a, b, and c indicating a difference between the groups at the significance level of P<0.05, 0.01 and 0.001, respectively.
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

Table 4. Selected genes identified by DNA microarray analysis Unique ID
Description /putative functions*
Array Ratio#
qPCR Ratio@
P value
SMU.246 glycosyl transferase N-acetylglucosaminyltransferase, RgpG -2.6 -4.0 5.9E-3 SMU.549 undecaprenyl-PP-MurNAc-pentapeptide-UDPGlcNAc GlcNAc transferase, MurG -2.7 -2.0 3.6E-3 SMU.599 D-alanine-D-alanine ligase, DdlA -1.8 -3.2 7.1E-3 SMU.1677
UDP-N-acetylmuramoylananine-D-glutamate-2,6-diaminopimelate ligase, UDP-MurNac-tripeptide synthetase, MurE
-13.1
-2.7
3.6E-3
SMU.1689 D-alanyl carrier protein, DltC, 3.4 nd 6.9E-3 SMU.1691 D-alanine-D-alanyl carrier protein ligase, DltA, 3.4 nd 3.7E-5 SMU.1838 preprotein translocase subunit SecA -1.9 -2.0 6.6E-3 SMU.1948 preprotein translocase subunit SecE -2.8 nd 2.1E-3 SMU.2006 preprotein translocase SecY protein 3.9 1.9 1.6E-5
Note: *, description and putative function of the identified genes are based upon the published S. mutans database. # and @, defined as levels of expression in the BrpA-deficient mutant relative to those of the wild-type shown by DNA microarray analysis and RealTime-PCR, respectively, with “-“ representing down-regulation. nd, not done.
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

0 9
A B
0.5
0.6
0.7
0.8
0.9UA159
TW14D
dens
ity,O
D57
5nm
**
0
0.1
0.2
0.3
0.4
Opt
ical
d
WS AG
Coating conditions
Figure 1g
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

A B
1 2 3 4
A B
41 2 3250150
M
250150
M
150
100
75100
75150
C41 2 3M M
D41 2 341 2 3
250150
100
M
25015010075
M 41 2 3
Figure 2100
75
75
50
37
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

100OMZ175
40
60
80 TW230
rcen
t sur
viva
l
0 10 20 30 40 500
20Per
hours
Figure 3
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

441 bp 1218 bp 293 bp 419 bpA.
SMU.409 brpA (SMU.410) SMU.411 SMU.412
TCATTTTTTGAAAATAGTTGGAAGTAATAAAAGATTTTGATATAATACTTTTATGTTTTATAGCCAAAATGAAAATCAGCTAATGGCTC
B. SMU.409
PhoB
TTGGACAGAGAATTGGTCAAAAACTACAAGCTCAAGACGTTCTTGTTCTGACAGGCGATTTAGGATCTGGGAAAACCACTCTGACAAAAG
-35 -10 TIS
5’ primer for RT-PCR
GltC
GAATTGCCAAAGGTTTGGGGATTAAGCAGATGATTAAGAGTCCAACCTATACGATTGTTCGTGAATATGAGGGAAGGCTGCCACTTTATC
ATTTGGATGTCTATCGAATTGGTGATGATCCTGATTCTATTGATCTGGATGATTTTCTTTTTGGAGATGGCGTAACAGTGATTGAGTGGG Stop codon GltCGGAGAGCTCTTAGACGATAGCCTTTTGAGCGATTATTTGACAGTCCTTTTAGATAAAACAGAGGGTGGCAGACAAATTACTTTATTACCTCATGGCCTGCGCTCTCAGCAATTGGTTGCGGAACTTGAGCATGACTGAGCAGGAAGTTTTTATCTGTCAAGTCAGTTTTGAAGATACAGAA -35’ -10’OxyR-like
Stop codo
TnrA-like
AAAAAGTTTATCAGAAATCTCGAAAAAACCGATTAAGAGATTAATTTTAATTTGAGATGAAGAGTGCCCTCATATAACTGCTCAAGAAA
TGGCTGCTTTTATTGAAAAAAGCTCAATCAGTCAATGAAATTTGTGTGATTGTAAGCTAGTCGGCCAAGTGATAGAGCTTGTGATGATTTTAGCACATAGCGAAATGATACGCTGTTTGGAATGAACGGCTCAAAAACAAAAGATGGAGTATTTCTTGATGTTTACCTTAGGAGGAAACT
Fnr-likebrpA
CT
GATAGATTAGAAAAACAAATGFigure 4
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

A. B.
M 1 2 3
0 8
1
400
500
0.6
0.8
300
400
OD
600n
m
D60
0nm
1 kb
0.2
0.4
100
200
Cul
tura
l
RLU
/OD
001 2 3 4 5 6 7 8
Time, hours
Figure 5Figure 5
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from














![3-BrPA eliminates human bladder cancer cells with highly ......cessfully evolve and promptly pass the proof-of-principle tests in preclinical models and clinical trials [1, 3, 6]](https://img.pdfslide.net/doc/110x75/60aaa3c19d8e4b6fe7142ca4/3-brpa-eliminates-human-bladder-cancer-cells-with-highly-cessfully-evolve.jpg)




![3a Brpa>KAaHe Pa3Mep: 86 X 50 CM 3a u.]Kacþ CBeTbJ1 50 CM KopL,1To: 40 X 40 CM rnaAKa Abn601-lVlHa 20 CM C OTBOP 3a CMeCVlTen VI aBTOMaTV1qeH KnanaH KOM"IJIEKT TEKAWAY STYLO IE 3a](https://img.pdfslide.net/doc/110x75/5c8754ea09d3f2bc6b8b9066/3a-brpakaahe-pa3mep-86-x-50-cm-3a-ukacb-cbetbj1-50-cm-kopl1to-40-x-40-cm.jpg)







